Embed Size (px)
Citation preview

Generalist versus specialist strategies of plasticity: snailresponses to predators with different foraging modes
JASON T. HOVERMAN*, RICKEY D. COTHRAN† AND RICK A. RELYEA†
*Department of Forestry and Natural Resources, Purdue University, West Lafayette, IN, U.S.A.†Department of Biological Sciences, University of Pittsburgh, Pittsburgh, PA, U.S.A.
SUMMARY
1. Phenotypic plasticity is a common adaptation to environmental heterogeneity, and theory predicts
that the evolution of constitutive versus plastic strategies should depend on the frequency of alterna-
tive environments, the magnitude of constraints and the costs of plasticity per se. However, it is
unclear how species should evolve when they experience more than two environments that favour
divergent phenotypes, particularly when they have absolute constraints on their morphology.
2. We examined the plasticity of three freshwater snail species (Helisoma anceps, H. campanulata and
H. trivolvis) in response to three environments: (i) no predator; (ii) shell-invading water bugs (Belos-
toma flumineum) and (iii) shell-crushing crayfish (Orconectes rusticus). We found distinct responses by
each snail species to the predator treatments. Helisoma anceps starts with a relatively low, narrow and
thick shell that becomes lower and thicker in response to crayfish but is unresponsive to water bugs.
In contrast, H. campanulata starts with a relatively high, wide and thin shell that becomes lower and
wider in response to water bugs but is unresponsive to crayfish. Helisoma trivolvis starts with a shell
of intermediate height and width while the predators induce defences in different directions.
3. These results suggest that H. trivolvis has a generalist plastic strategy while H. anceps and H. cam-
panulata have specialised plastic strategies orientated against a single type of predator at the potential
cost of being unable to respond to others.
4. We then performed predation trials to determine predator preferences using a mixture of the three
species. After 2 weeks of exposure to crayfish cues, H. anceps had higher survival than both H. trivol-
vis and H. campanulata with uncaged crayfish. After 2 weeks of exposure to water bug cues, both
H. trivolvis and H. campanulata had higher survival than H. anceps with uncaged water bugs. When
predation trials were conducted after 5 weeks of exposure to predator cues, H. trivolvis and H. cam-
panulata reached a size refuge from both predators and this shifted predation pressure to H. anceps.
5. Collectively, these results suggest that closely related prey species with different absolute con-
straints in their morphology had different defences that are either specialised or generalised to alter-
native environments.
Keywords: functional tradeoff, gastropod, inducible defence, phylogeny, selection
Introduction
Natural selection in heterogeneous environments may
lead to the evolution of phenotypic plasticity, defined as
the ability of a single genotype to produce different
phenotypes in response to different environments
(Schlichting & Pigliucci, 1998; Pigliucci, 2001).
Phenotypic plasticity exists in many species and in
response to a wide range of environmental conditions,
and substantial variation in the expression of phenotypic
plasticity can exist among closely related species
(Harvell, 1991; Kusch, 1993; Colbourne, Hebert & Taylor,
1997; Van Buskirk, 2002; Berendonk, Barraclough &
Barraclough, 2003). However, the evolution of plasticity
may be constrained by several mechanisms. For
example, absolute constraints (sensu Brakefield, 2006)
arise because the basic body plans of species are quite
‘difficult’ to change by natural selection (i.e. new traits
Correspondence: Jason T. Hoverman, Department of Forestry and Natural Resources, Purdue University, West Lafayette, IN 47907, U.S.A.
E-mail: [email protected]
© 2014 John Wiley & Sons Ltd 1
Freshwater Biology (2014) doi:10.1111/fwb.12332

that break the constraint may be deleterious). Addition-
ally, allocation tradeoffs can occur when resource limita-
tion constrains simultaneous investment in multiple
traits (Dewitt, 1998; Auld, Agrawal & Relyea, 2010).
These constraints, combined with the amount of envi-
ronmental heterogeneity experienced by a species over
time or space, can drive the evolution of different mean
phenotypes in closely related species (i.e. averaged
across all environments), as well as different directions
and magnitudes of phenotypic plasticity (Van Tienderen,
1991; Dewitt, Sih & Wilson, 1998; Schlichting & Pigliucci,
1998; Pigliucci, 2001; Van Kleunen & Fischer, 2007; Auld
et al., 2010). Because closely related species are likely to
be similar in biochemical, physiological and structural
constraints, comparative studies have the potential to
identify a core set of constraints that may limit the
expression of environmentally induced traits, which
may bias evolution towards ‘fixed’ solutions to environ-
mental heterogeneity (Pfennig et al., 2010).
The inducible defences of freshwater snails represent
an ideal system to assess how absolute constraints and
allocation tradeoffs may influence the divergent evolu-
tion of plasticity across species. For example, shell geom-
etry creates constraints on shell shape. The shells of
most gastropods can be described using three parame-
ters: expansion rate (W), translation (T) and distance (D)
of the generating curve from the axis of coiling (Raup,
1962; Rice, 1998). Physical relationships among these
shell parameters limit shell shape (i.e. not all regions of
morphospace can be achieved) and therefore affect the
range of options for morphological defences against var-
ious predators. For example, species with rapid shell
expansion rates (i.e. a coiling tube that rapidly increases
in diameter) produce shells with relatively large aper-
tures, which are more vulnerable to predators that enter
the shell. However, these species can invest more in
shell thickness, a defence against shell-crushing preda-
tors, because fewer coils around the central axis are
needed to increase overall body size (Raup, 1962). Alter-
natively, snails with slow expansion rates generate shells
with relatively small apertures that are difficult to enter,
although such shells are typically thinner because more
coils around the central coiling axis are required to grow
to a particular body size (Raup, 1962). In addition to
absolute constraints on shell geometry, snails also
face allocation tradeoffs (Dewitt & Langerhans, 2003;
Hoverman & Relyea, 2007a, 2009); for instance, there can
be a tradeoff between investing a limited amount of
shell material to thickness or coiling (Russell-Hunter,
1978; Kemp & Bertness, 1984; Brodersen & Madsen,
2003). As a result, snails commonly face tradeoffs in
how they respond to shell-invading versus shell-crush-
ing predators (Dewitt, 1998; Hoverman & Relyea, 2008,
2009; Bourdeau, 2009). Thus, interspecific differences in
expansion rate, coupled with constraints on shell thick-
ness, could influence patterns of phenotypic plasticity
and phenotypic diversification of snail species (Edgell &
Miyashita, 2009).
We examined the inducible defences of three closely
related planorbid snails (Helisoma trivolvis, H. anceps and
H. campanulata; Fig. 1) to determine how these snails
respond to predators with different foraging modes,
given the absolute constraints and allocation tradeoffs
that limit shell morphology. The three snail species
occur together in semipermanent to permanent water-
bodies, where they encounter a diversity of predators
including water bugs, crayfish and fish (Hoverman et al.,
2011). In a series of studies, we have explored the
responses of H. trivolvis to different predators (Hover-
man, Auld & Relyea, 2005; Hoverman & Relyea, 2007a,b,
2008, 2009). In the presence of the water bug Belostoma
flumineum, H. trivolvis invests in shell coiling but the
aperture remains relatively small because of their
moderate expansion rate; this reduces the ability of the
bug to reach the snail’s soft tissues when withdrawn
inside the shell. In contrast, H. trivolvis forms thicker
shells in the presence of the crayfish Orconectes rusticus,
which reduces the predator’s ability to crack or crush
the shell.
In contrast to the extensive research on H. trivolvis,
there appear to be no studies on the predator-induced
morphology of H. campanulata and H. anceps. Among the
three species, the main difference in shell shape is the
rate of shell expansion (Raup, 1962). Visual inspection
suggests that the expansion rate is low for H. campanula-
Fig. 1 Left side (i.e. spire) view of Helisoma anceps, H. campanulata
and H. trivolvis. These three species differ in the rate at which the
diameter of the shell increases with each rotation around the
coiling axis (termed w; Raup, 1962); w is low for H. campanulata,
intermediate for H. trivolvis and high for H. anceps, respectively. As
a consequence of variation in w, H. anceps exhibits a relatively large
aperture for a given body mass, whereas H. campanulata exhibits a
relatively small aperture.
© 2014 John Wiley & Sons Ltd, Freshwater Biology, doi: 10.1111/fwb.12332
2 J. T. Hoverman et al.

ta, intermediate for H. trivolvis and high for H. anceps.
Consequently, H. campanulata has a relatively small shell
aperture relative to its overall size, whereas H. trivolvis
has an intermediate aperture and H. anceps has a rela-
tively large aperture. Additionally, shell thickness also
appears to differ among species; shells are relatively
thick for H. anceps, intermediate for H. trivolvis, and rela-
tively thin for H. campanulata (Osenberg & Mittelbach,
1989; Brown, 1998).
These differences in basic shell geometry and thick-
ness might constrain the predator-induced defences of
each species, resulting in differences in predation risk by
shell-invading versus shell-crushing predators. For
example, because H. anceps has a thick shell and a high
shell expansion rate, it should be well defended against
shell-crushing predators but vulnerable to shell-invading
predators. Because H. campanulata has a thin shell with a
low shell expansion rate, it should be well defended
against shell-invading predators but vulnerable to crush-
ing predators. To test these hypotheses experimentally,
we assessed the phenotypic responses of the snails to
water bugs or crayfish and then examined the relative
susceptibility of each snail species to the predators.
Methods
Induction experiment
The goal of the induction experiment was to assess
predator-induced morphology in the three Helisoma spe-
cies. We collected ~100 adults of H. trivolvis, H. anceps
and H. campanulata from ponds near the University of
Pittsburgh’s Pymatuning Laboratory of Ecology (PLE) in
Linesville, PA. These ponds contain both water bugs
(Belostoma flumineum) and crayfish (Orconectes rusticus).
For each snail species, 10 individuals were placed into
each of 10 culture pools filled with 100 L of well water.
Egg deposition began immediately and continued until
the adults were removed after 2 weeks. Upon hatching,
snails were fed rabbit food ad libitum until the experi-
ment began.
In a mesocosm experiment, we examined the effects of
caged predators on the growth and morphology of each
species. We designed a completely randomised, factorial
experiment composed of three predator treatments (no
predator, caged water bug [B. flumineum], or caged cray-
fish [O. rusticus]) crossed with the three snail species.
These nine treatments were replicated eight times for a
total of 72 experimental units.
The experimental units were 90-L pools filled with
well water. We added 10 g of rabbit food and an aliquot
of pond water, containing periphyton, phytoplankton
and zooplankton, to sustain food for snails and maintain
water quality. Each pool received 100 juvenile snails of
the appropriate species. Initial mean mass � 1 SD of
H. trivolvis, H. anceps or H. campanulata was 2.0 �1.4 mg, 1.2 � 0.6 mg and 1.5 � 0.9 mg, respectively. For
each snail species, 20 snails were set aside to assess mor-
tality due to handling; 24-h survival was 100%.
After adding the snails, we placed a single predator
cage into each pool. The cages were made from corru-
gated pipes (10 cm long 9 10 cm diameter) capped with
shade cloth. For caged predator treatments, we added
one water bug or crayfish to each cage. Caged predators
emit water-borne chemical cues, which provide the
opportunity for prey to detect and respond to predators
without reducing prey density (Chivers & Smith, 1998).
The caged predators were fed 300 mg of snail biomass
(total wetmass including shells, two to five snails) of the
appropriate snail species three times per week. Based on
previous research, this amount of consumed snail bio-
mass by the predators is sufficient to elicit phenotypic
responses in H. trivolvis (Hoverman & Relyea, 2007b,
2008, 2009). The predators consumed all the snails
between feedings. To equalise disturbance, we briefly
lifted the cages in the no-predator treatment from the
water and then returned them. We placed a shade cloth
lid over each pool to prevent colonisation by insects and
amphibians.
During the experiment, we observed that the spe-
cies varied greatly in growth rates; growth was fast in
H. trivolvis, intermediate in H. campanulata and slow in
H. anceps. Because analyses of morphological plasticity
are sensitive to differences in mass, we decided to take
down the experimental units for each species at different
times so that they were similar in mass at the end of the
experiment. Although this approach resulted in differ-
ences among the species in duration of predator expo-
sure, our previous work with H. trivolvis found that the
magnitude of predator-induced morphological change is
relatively constant over ontogeny (Hoverman & Relyea,
2007a, 2009). For H. trivolvis, H. campanulata and
H. anceps, the experiment was ended after 14, 21 and
39 days, respectively. On each date, all surviving snails
were removed and preserved in 10% formalin. In one
experimental unit, all the H. campanulata died and this
was excluded from analyses. For the remaining experi-
mental units, survival was high (>95%) and did not
differ among caged predator treatments or snail species
(predator, F2,63 = 0.7, P = 0.499; species, F2,63 = 0.3,
P = 0.706; interaction, F4,63 = 1.3, P = 0.274). For each
experimental unit, 10 individuals were randomly
© 2014 John Wiley & Sons Ltd, Freshwater Biology, doi: 10.1111/fwb.12332
Defensive strategies of Helisoma snails 3

selected and dried at 60 °C for 24 h. Each individual
was then weighed to the nearest mg (total dry mass
included shell and tissue) and measured for shell width
and height, and aperture width and height using digital
imaging software (Optimas Co., Bothell, WA, U.S.A.).
We also measured the shell thickness of each snail at the
leading edge of the aperture using digital calipers.
To examine the effects of our caged predator treat-
ments on snail morphology, we began by assessing the
allometric relationship between each shell dimension
and log10-transformed mass. While there was no rela-
tionship between shell thickness and mass for each spe-
cies, the remaining shell dimensions showed positive
relationships with mass. To account for these allometric
relationships, we used analysis of covariance (ANCO-
VA) with mass as a covariate and snail species and
predator treatment as main effects. A critical assumption
in the ANCOVA procedure is that the treatments share
a common regression slope and our data met this
assumption (tests of interactions with mass all
P ≥ 0.171). From the ANCOVA, we used the estimated
marginal means and residuals from within-treatment
regressions to calculate a mass-adjusted value for each
individual. Using all the measured individuals in the
ANCOVA, we had ample power to capture the allomet-
ric relationship between shell dimensions and mass. For
each shell dimension, we then calculated the mean size-
adjusted shell dimensions for each experimental unit
and used these means as the morphological response
variables. Because shell thickness did not covary with
mass, we calculated the mean shell thickness for the
snails from each experimental unit and this served as
the response variable.
We used a multivariate analysis of variance (MANO-
VA) to analyse the effect of caged predators and snail
species on final mass, mass-adjusted shell dimensions
(shell width and height, and aperture width and height)
and shell thickness. The data were normally distributed,
and variances were equal across treatments. Significant
multivariate effects were followed by univariate tests.
When univariate tests were significant, we conducted
mean comparisons using Tukey’s HSD test. We also con-
ducted correlation analyses to examine the relationships
between the morphological traits and assess whether
allocation tradeoffs were evident across snail species
and predator treatments. For each pairwise combination
of the five morphological traits, we calculated the Pear-
son product–moment correlation coefficient across the
nine experimental treatments. We used the mean trait
value for each experimental unit in the analysis
(N = 71). We used a Bonferroni-corrected a = 0.005 to
assess the significance of the 10 correlations that were
conducted.
Predation trials
For the predation trials, the objective was to test preda-
tor preference for the different snail species after
defences in response to the cues of water bugs or cray-
fish had been induced. Pilot experiments indicated that
the two species of predators were capable of consuming
all three species of snails when they were small and of
similar mass (i.e. ~50 mg dry mass; R.D. Cothran et al.
unpublished data), but the key issue is the predation
risk when predators are given a choice of all three spe-
cies that have been exposed to predator cues for the
same amount of time. Because the species differ not
only in relative shape but also in size (i.e. they grow at
different rates), the predation trials tested how the
entire suite of traits affects relative predation risk when
a predator is given a choice among the three snail spe-
cies. In short, the predation trials were designed to
assess differences in predation risk using the typical
variation in size and shape of snails that would occur
in nature.
For each snail species, we collected 100 adult snails
from the E.S. George Reserve in Livingston County, MI,
U.S.A. The three species were collected from a single
pond (Crane pond) that contained both predators (Hov-
erman et al., 2011). For each snail species, we divided
individuals equally among five 100-L wading pools (i.e.
20 per pool) and allowed them to deposit egg masses
for 2 weeks. After hatching, the juvenile snails were fed
rabbit food ad libitum until the start of the experiment.
Water bugs and crayfish were collected from local ponds
and housed in the laboratory, where they were fed
snails weekly until used in the predation trials.
For each snail species, we created water bug- and
crayfish-induced snails by raising them in 90-L wading
pools (n = 6 for each snail species and predator combi-
nation, for a total of 36 pools). These pools contained
either a caged crayfish or a caged water bug and were
set up identically to the pools used in the induction
experiment. We added 60, 1-week-old snails of the
appropriate species to each pool.
We tested the snails for their relative susceptibility to
a given predator after being exposed to predator cues
for 2 weeks (i.e. juvenile snails) and again after 5 weeks
(i.e. adult snails). For each snail species and predator
combination, we collected 20 snails from each of the six
induction pools and combined them for a total of 120
snails that could be randomly assigned to predation
© 2014 John Wiley & Sons Ltd, Freshwater Biology, doi: 10.1111/fwb.12332
4 J. T. Hoverman et al.
trials. We randomly selected a subsample of snails from
each treatment combination to test for species differ-
ences in mass (mg oven-dried mass including shell).
Using these induced animals, we randomly selected
10 individuals from each of the three snail species and
added the combined 30 animals to each experimental
unit. Experimental units were 10-L plastic tubs contain-
ing 7 L of well water and a predator refuge. For cray-
fish, the predator refuge was a half-piece of PVC pipe
(10.5 cm long, 4 cm high). For water bugs, the predator
refuges were two plastic ‘macrophytes’ composed of 10
pieces of polypropylene rope (10 cm long, 0.5 cm diame-
ter) bound together with a small cable tie and anchored
by a small rock. For water bugs, the size of the preda-
tors (length from tip of head to tip of abdomen) used in
the trials was 21.5 � 1.1 mm (mean � SD) and
20.8 � 0.9 mm for the 2 and 5 week trials, respectively.
For crayfish, the size of the predators (length from tip of
rostrum to tip of telson) used in the trials was
63.5 � 7.5 mm (mean � SD) and 73.4 � 4.6 mm for the
2 and 5 week trials, respectively.
The 30 snails in each tub were given 24 h to acclimate
before we added uncaged predators that had been
starved for 1 day. We set up 10 replicates containing the
mixture of snail species for each of the two predator
species. For the predation trial on snails that had been
exposed to predator cues for 2 weeks, we ended each
replicate after a predator had consumed 40–60% of the
snails. For the predation trial on snails that had been
exposed to predator cues for 5 weeks, we had to relax
this criterion because predators were not able to con-
sume many of the adult H. trivolvis and H. campanulata
(i.e. some tubs had up to 73% survival). We also
included two no-predator controls to monitor snail mor-
tality without predators and confirmed that survival
without a predator was 100%.
We used a general linear model (GLM) to compare
snail species survival after 2 and 5 weeks of exposure to
predator cues. ‘Species of snail’ was used as the within-
subject variable and ‘predator type’ (crayfish or water
bug) was used as the between-subject variable. We also
used a GLM to test for differences in size among the
snail species.
Results
Induction experiment
In the induction experiment, we found that responses to
the caged predators varied across the three snail species
(Table 1). Because of the interaction between snail
species and caged predators, we explored the effects of
caged predators within each snail species.
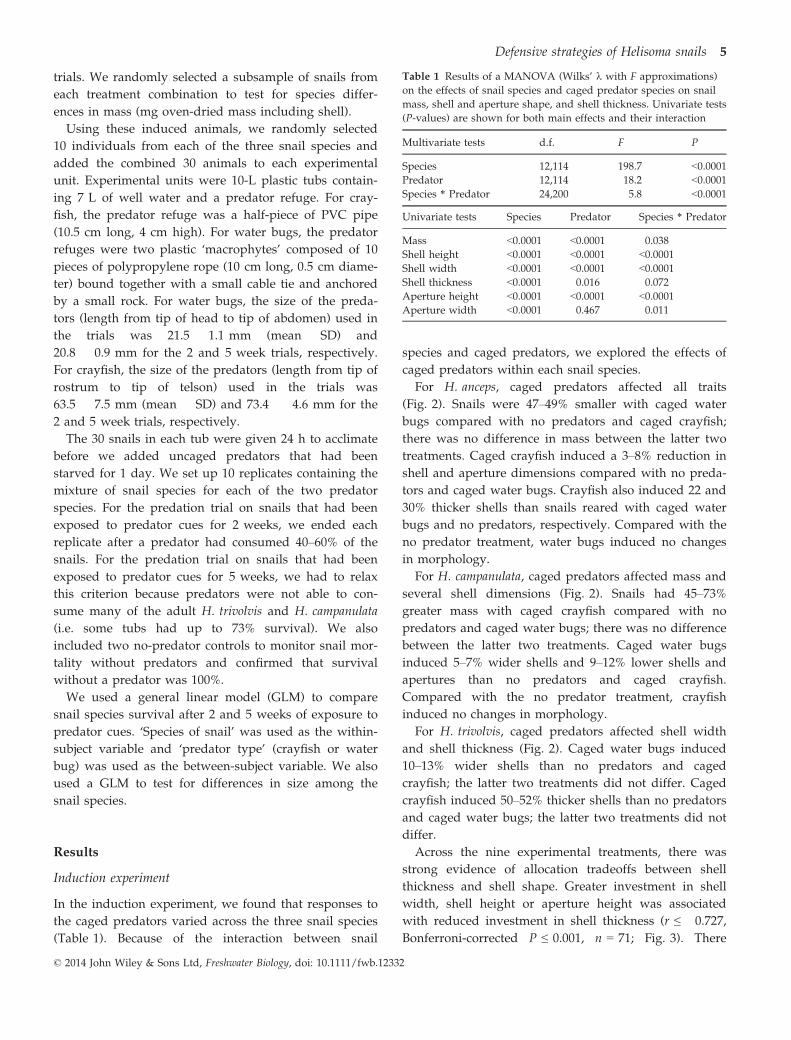
For H. anceps, caged predators affected all traits
(Fig. 2). Snails were 47–49% smaller with caged water
bugs compared with no predators and caged crayfish;
there was no difference in mass between the latter two
treatments. Caged crayfish induced a 3–8% reduction in
shell and aperture dimensions compared with no preda-
tors and caged water bugs. Crayfish also induced 22 and
30% thicker shells than snails reared with caged water
bugs and no predators, respectively. Compared with the
no predator treatment, water bugs induced no changes
in morphology.
For H. campanulata, caged predators affected mass and
several shell dimensions (Fig. 2). Snails had 45–73%
greater mass with caged crayfish compared with no
predators and caged water bugs; there was no difference
between the latter two treatments. Caged water bugs
induced 5–7% wider shells and 9–12% lower shells and
apertures than no predators and caged crayfish.
Compared with the no predator treatment, crayfish
induced no changes in morphology.
For H. trivolvis, caged predators affected shell width
and shell thickness (Fig. 2). Caged water bugs induced
10–13% wider shells than no predators and caged
crayfish; the latter two treatments did not differ. Caged
crayfish induced 50–52% thicker shells than no predators
and caged water bugs; the latter two treatments did not
differ.
Across the nine experimental treatments, there was
strong evidence of allocation tradeoffs between shell
thickness and shell shape. Greater investment in shell
width, shell height or aperture height was associated
with reduced investment in shell thickness (r ≤ �0.727,
Bonferroni-corrected P ≤ 0.001, n = 71; Fig. 3). There
Table 1 Results of a MANOVA (Wilks’ k with F approximations)
on the effects of snail species and caged predator species on snail
mass, shell and aperture shape, and shell thickness. Univariate tests
(P-values) are shown for both main effects and their interaction
Multivariate tests d.f. F P
Species 12,114 198.7 <0.0001Predator 12,114 18.2 <0.0001Species * Predator 24,200 5.8 <0.0001
Univariate tests Species Predator Species * Predator
Mass <0.0001 <0.0001 0.038
Shell height <0.0001 <0.0001 <0.0001Shell width <0.0001 <0.0001 <0.0001Shell thickness <0.0001 0.016 0.072
Aperture height <0.0001 <0.0001 <0.0001Aperture width <0.0001 0.467 0.011
© 2014 John Wiley & Sons Ltd, Freshwater Biology, doi: 10.1111/fwb.12332
Defensive strategies of Helisoma snails 5
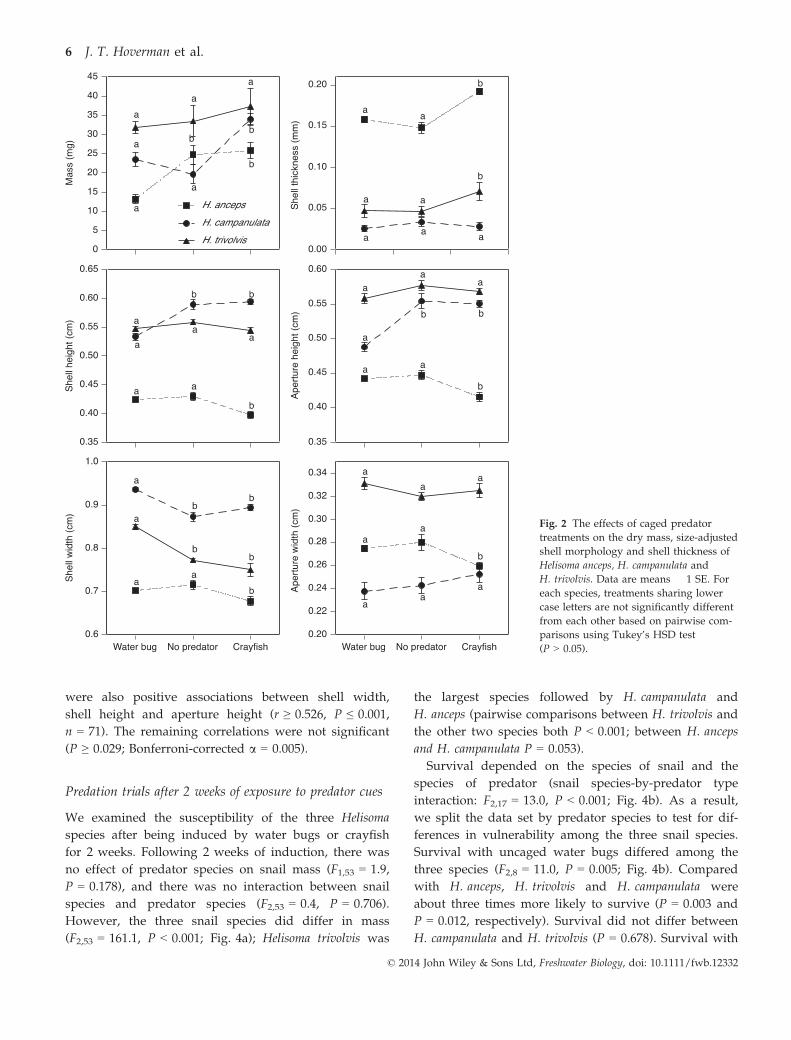
were also positive associations between shell width,
shell height and aperture height (r ≥ 0.526, P ≤ 0.001,
n = 71). The remaining correlations were not significant
(P ≥ 0.029; Bonferroni-corrected a = 0.005).
Predation trials after 2 weeks of exposure to predator cues
We examined the susceptibility of the three Helisoma
species after being induced by water bugs or crayfish
for 2 weeks. Following 2 weeks of induction, there was
no effect of predator species on snail mass (F1,53 = 1.9,
P = 0.178), and there was no interaction between snail
species and predator species (F2,53 = 0.4, P = 0.706).
However, the three snail species did differ in mass
(F2,53 = 161.1, P < 0.001; Fig. 4a); Helisoma trivolvis was
the largest species followed by H. campanulata and
H. anceps (pairwise comparisons between H. trivolvis and
the other two species both P < 0.001; between H. anceps
and H. campanulata P = 0.053).
Survival depended on the species of snail and the
species of predator (snail species-by-predator type
interaction: F2,17 = 13.0, P < 0.001; Fig. 4b). As a result,
we split the data set by predator species to test for dif-
ferences in vulnerability among the three snail species.
Survival with uncaged water bugs differed among the
three species (F2,8 = 11.0, P = 0.005; Fig. 4b). Compared
with H. anceps, H. trivolvis and H. campanulata were
about three times more likely to survive (P = 0.003 and
P = 0.012, respectively). Survival did not differ between
H. campanulata and H. trivolvis (P = 0.678). Survival with
Fig. 2 The effects of caged predator
treatments on the dry mass, size-adjusted
shell morphology and shell thickness of
Helisoma anceps, H. campanulata and
H. trivolvis. Data are means � 1 SE. For
each species, treatments sharing lower
case letters are not significantly different
from each other based on pairwise com-
parisons using Tukey’s HSD test
(P > 0.05).
© 2014 John Wiley & Sons Ltd, Freshwater Biology, doi: 10.1111/fwb.12332
6 J. T. Hoverman et al.
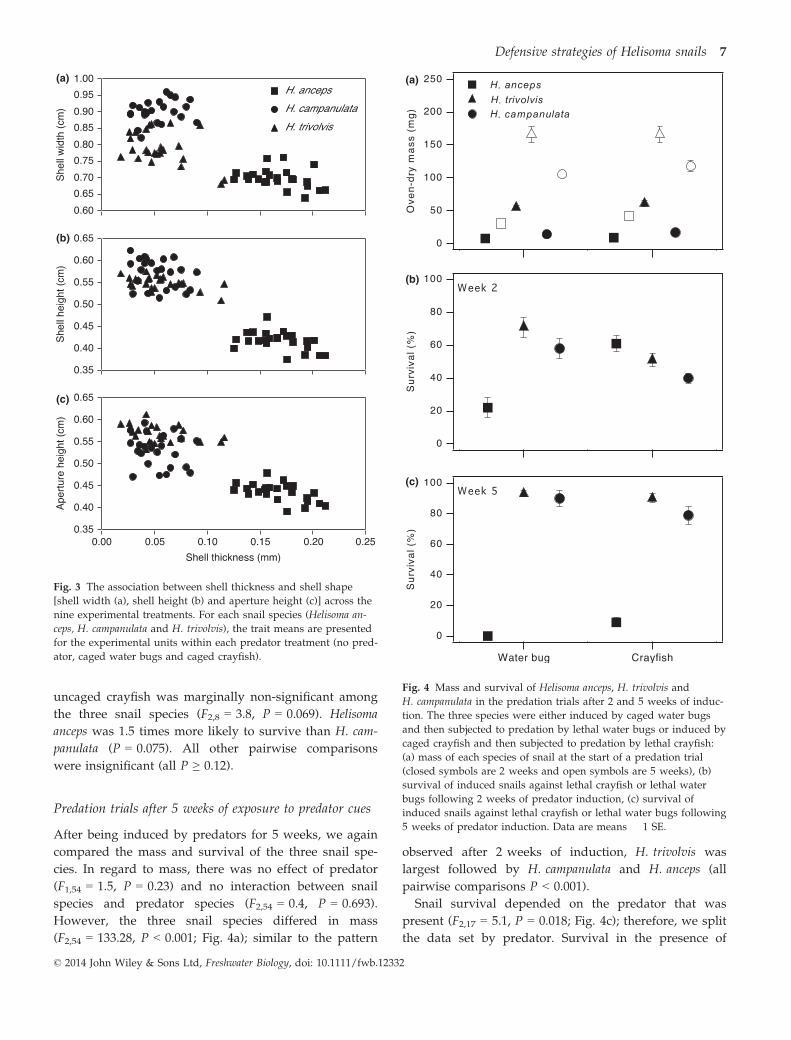
uncaged crayfish was marginally non-significant among
the three snail species (F2,8 = 3.8, P = 0.069). Helisoma
anceps was 1.5 times more likely to survive than H. cam-
panulata (P = 0.075). All other pairwise comparisons
were insignificant (all P ≥ 0.12).
Predation trials after 5 weeks of exposure to predator cues
After being induced by predators for 5 weeks, we again
compared the mass and survival of the three snail spe-
cies. In regard to mass, there was no effect of predator
(F1,54 = 1.5, P = 0.23) and no interaction between snail
species and predator species (F2,54 = 0.4, P = 0.693).
However, the three snail species differed in mass
(F2,54 = 133.28, P < 0.001; Fig. 4a); similar to the pattern
observed after 2 weeks of induction, H. trivolvis was
largest followed by H. campanulata and H. anceps (all
pairwise comparisons P < 0.001).
Snail survival depended on the predator that was
present (F2,17 = 5.1, P = 0.018; Fig. 4c); therefore, we split
the data set by predator. Survival in the presence of
(a)
(b)
(c)
Fig. 3 The association between shell thickness and shell shape
[shell width (a), shell height (b) and aperture height (c)] across the
nine experimental treatments. For each snail species (Helisoma an-
ceps, H. campanulata and H. trivolvis), the trait means are presented
for the experimental units within each predator treatment (no pred-
ator, caged water bugs and caged crayfish).
(a)
(b)
(c)
Fig. 4 Mass and survival of Helisoma anceps, H. trivolvis and
H. campanulata in the predation trials after 2 and 5 weeks of induc-
tion. The three species were either induced by caged water bugs
and then subjected to predation by lethal water bugs or induced by
caged crayfish and then subjected to predation by lethal crayfish:
(a) mass of each species of snail at the start of a predation trial
(closed symbols are 2 weeks and open symbols are 5 weeks), (b)
survival of induced snails against lethal crayfish or lethal water
bugs following 2 weeks of predator induction, (c) survival of
induced snails against lethal crayfish or lethal water bugs following
5 weeks of predator induction. Data are means � 1 SE.
© 2014 John Wiley & Sons Ltd, Freshwater Biology, doi: 10.1111/fwb.12332
Defensive strategies of Helisoma snails 7

uncaged water bugs differed among the three snail
species (F2,8 = 168, P < 0001). None of the H. anceps
survived, whereas survival was high for H. campanulata
and H. trivolvis (comparisons against H. anceps: both
P < 0.001; H. campanulata versus H. trivolvis: P = 1.0).
Survival with uncaged crayfish also differed among the
three snail species (F2,8 = 158, P < 0.001). Few H. anceps
survived, whereas most of the H. campanulata and H. tri-
volvis did (comparisons with H. anceps: both P < 0.001;
H. campanulata versus H. trivolvis: P = 0.486).
Discussion
We found that Helisoma trivolvis, H. anceps and H. cam-
panulata all expressed predator-induced plasticity, yet
they exhibited individual responses to the predators.
The responses of H. trivolvis to caged predators were
consistent with previous research on this species
(Hoverman et al., 2005; Hoverman & Relyea, 2007b).
When exposed to water bug cues, they formed wider
shells (i.e. coiled more) but aperture size did not change.
These responses enable H. trivolvis to withdraw into its
shell, which reduces the water bug’s ability to access the
snail’s soft tissues (Hoverman & Relyea, 2009). They
formed thicker shells when exposed to crayfish cues,
which reduces shell-crushing or chipping by crayfish
(Hoverman & Relyea, 2009). Given the adaptive value of
these responses for H. trivolvis and the co-occurrence of
H. anceps and H. campanulata with the same predators,
we predicted similar phenotypic responses across the
snail species. However, the morphology of H. anceps and
H. campanulata changed in response to one or the other
predator but not to both. Helisoma anceps responded to
crayfish by forming thicker shells, narrower shells and
apertures, and lower shells and apertures; however,
there was no response to water bugs. Helisoma campanu-
lata responded to water bugs by forming wider and
lower shells and low apertures, although there was no
response to crayfish. Although there were no
morphological responses in H. anceps and H. campanulata
when exposed to water bugs and crayfish, respectively,
mass did change. Specifically, H. anceps was smaller
with caged water bugs while H. campanulata was larger
with caged crayfish. Thus, both species did detect these
predators, but morphology did not respond. Interest-
ingly, the predator-specific morphological responses
observed in H. anceps and H. campanulata to crayfish and
water bugs, respectively, are similar to many of the
adaptive responses observed in H. trivolvis. Thus, the
responses of H. anceps to crayfish, and the responses of
H. campanulata to water bugs, might reduce risk to each
inducing predator, but the lack of response to the alter-
native predator might increase risk in the presence of
that predator (a hypothesis later tested in the predation
trials).
The differences in morphological responses to water
bugs between the species appear to reflect the geometric
constraints of shell shape. While all three species have
planospiral coiling, H. campanulata and H. trivolvis have
low expansion rates, so the diameter of their coiled tube
increases only gradually as whorls are added. In contrast,
H. anceps has a high expansion rate, so the diameter of its
coiled tube increases substantially as more whorls are
added. Consequently, H. campanulata and H. trivolvis can
respond to water bugs by increasing shell width (i.e. rap-
idly growing a longer coiled tube) because this places the
aperture of the shell further away from the soft tissue of
the snail, without substantially increasing the size of the
aperture; this makes it harder for a water bug to reach
and pierce the soft tissues. If H. anceps was to respond to
water bugs in this manner, the soft tissue would be fur-
ther from the aperture, but the aperture would also
become much larger and easier for a water bug to enter
and pierce the soft tissues. This geometric constraint of
H. anceps may explain its lack of morphological response
to water bugs.
The existence of allocation tradeoffs is an underlying
assumption of phenotypic plasticity (Schlichting & Pig-
liucci, 1998; Tollrian & Harvell, 1999; Pigliucci, 2001).
Using correlation analyses, we found strong evidence for
allocation tradeoffs across the experimental treatments;
investment in shell thickness was negatively correlated
with investment in shell shape. Thus, in addition to geo-
metric constraints on shell shape, snails face allocation
tradeoffs that constrain their ability to invest simulta-
neously in shell thickness and shape. These results are
consistent with previous work in marine snails. For
instance, Trussell & Nicklin (2002) found a negative asso-
ciation between shell length and thickness in Littorina
obtusata. This allocation tradeoff appears to be driven by
competing demands for calcium, such that thicker shells
can only be produced when investment in shell length is
reduced (Palmer, 1981; Kemp & Bertness, 1984).
As a consequence of allocation tradeoffs, snails are
faced with a fundamental tradeoff between responses to
shell-entering and shell-crushing predators. In H. trivol-
vis, the induction of wider shells, by extending the length
of the coiled tube, makes it more difficult for water bugs
to enter the aperture and pierce the soft tissue (Hoverman
& Relyea, 2009). However, this comes at the cost of
reduced investment in shell thickness, which makes the
shell more vulnerable to crushing predators such as
© 2014 John Wiley & Sons Ltd, Freshwater Biology, doi: 10.1111/fwb.12332
8 J. T. Hoverman et al.

crayfish. Similarly, the body plan of H. campanulata is well
suited to defence against water bugs, because it can rap-
idly grow a long, thin tube that makes it difficult for water
bugs to reach the soft tissue. However, this strategy leaves
few resources for thickening the shell making H. campanu-
lata vulnerable to crushing predators such as crayfish. In
other freshwater and marine snails, research has also doc-
umented contrasting responses to predators with diver-
gent foraging modes that appear to be driven by
allocation tradeoffs (Dewitt, Robinson & Wilson, 2000;
Bourdeau, 2009). For instance, Bourdeau (2009) showed
that the marine snail, Nucella lamellosa, forms thick, round
shells when exposed to shell-crushing crabs but elongate
shells with shell-entry seastars. Thus, morphological
defences in snails appear to be driven by a fundamental
tradeoff between shell thickness and shape.
Our study focussed on morphological responses to pre-
dators, yet behavioural responses have also been
observed in H. trivolvis and other snails (Dewitt, Sih &
Hucko, 1999; Turner, Bernot & Boes, 2000; Hoverman
et al., 2005; Hoverman & Relyea, 2007b). Behavioural
responses often involve spatial avoidance, such that snails
move into areas that provide shelter (e.g. rock refuges) or
are difficult for the predator to reach (e.g. the water sur-
face). In our previous work, we did not detect behaviour-
al responses to water bugs because this predator is
capable of foraging for snails throughout the water col-
umn and under ‘refuges’ (Hoverman et al., 2005; Hover-
man & Relyea, 2007b). However, snails often move
towards the water surface or above the water line in the
presence of crayfish (Hoverman et al., 2005), although
such responses are not always observed (Hoverman &
Relyea, 2007b). As crayfish are largely benthic, this behav-
iour can reduce the probability of encounter. While
behaviour was not quantified in our experiments, it is
possible that it could play an important role in the
defences of these snails. For instance, several studies have
demonstrated trait compensation, where individuals with
poorly formed morphological defences display strong
behavioural responses to predators (Dewitt et al., 1999;
Rundle & Br€onmark, 2001; Cotton, Rundle & Smith, 2004;
Rundle et al., 2004). Thus, it is possible that H. campanula-
ta and H. anceps could display anti-predator behaviours
in the presence of crayfish and water bugs, respectively,
compensating for their vulnerable shell morphology.
Additional research is necessary to explore whether
behavioural responses are employed by these species.
The results of the predation trials conducted after
2 weeks of exposure to the cues of water bugs or
crayfish demonstrated how the phenotypes of the snails,
when all three species were combined, affected their
relative predation risk. In this trial, all three species were
within a mass range that could be consumed by either
predator (R.D. Cothran et al., unpublished data), but the
survival of each snail species with uncaged crayfish or
water bugs was strongly associated with the observed
changes in mass and morphology. When crayfish-
induced snails were exposed to crayfish, survival was
high for H. anceps, intermediate for H. trivolvis and low
for H. campanulata. This was particularly interesting
because H. anceps was the smallest of the three species,
yet it was the best defended against crayfish. When
water bug-induced snails were exposed to water bugs,
survival was low for H. anceps and high for H. trivolvis
and H. campanulata. Comparing the mass of H. trivolvis
and H. campanulata, the latter was considerably smaller,
yet its morphological defences against water bugs were
stronger. In contrast, H. campanulata and H. anceps were
more similar in mass, yet H. campanulata had better
survival, further confirming that the morphological
responses to water bug cues provided H. campanulata
with effective defences against water bugs. These results
are consistent with the argument that snails with
relatively wider shells are better defended against water
bugs and snails with thicker shells are better defended
against crayfish.
After being exposed to predator cues for 5 weeks, we
observed that H. trivolvis and H. campanulata had
very high survival with both uncaged predators while
H. anceps had very low survival. The main driver of
these contrasting outcomes of predation with time
appears to be the occurrence of size refuges from preda-
tion. We have previously shown that adult H. trivolvis
can reach a size refuge against both water bugs and
crayfish, regardless of their shell phenotype (Hoverman
& Relyea, 2009). Thus, it appears that H. trivolvis and
H. campanulata exceeded the size required to reach a ref-
uge from both predators, whereas H. anceps was still
vulnerable. As a result, it appears that the inability of
either predator to consume H. trivolvis and H. campanu-
lata shifted all of the predation pressure to H. anceps,
thereby causing much lower survival of this species. In
short, when the snails of any species are within a vul-
nerable size range, morphological defences are impor-
tant for reducing predation risk, but when a species
achieves a size refuge, the morphology of the large spe-
cies becomes unimportant to predation risk and preda-
tion can become more intense on the small species in
the community; even when the small species has a
defended phenotype. One of the limitations of our
experimental design was that we used a narrow size
range for each predator species. Although there is little
© 2014 John Wiley & Sons Ltd, Freshwater Biology, doi: 10.1111/fwb.12332
Defensive strategies of Helisoma snails 9

variation in the size of adult water bugs, crayfish can
attain a larger size than represented in our study. As
adults, crayfish tend to shift to more herbivorous diets,
but their greater size does allow them to feed on larger
macroinvertebrates (Lodge & Lorman, 1987; Lodge et al.,
1994). Because larger crayfish can consume larger prey,
it is possible that the results of the predation trials after
5 weeks of induction would be different if larger cray-
fish were selected for the experiment. However, if snails
were not in a size refuge from predation, we would
expect the results to be similar to the outcome observed
in the predation trials after 2 weeks of induction.
While predation is a common threat for freshwater
snails that has been hypothesised to affect patterns in
species richness, species abundance and community
structure (Lodge et al., 1987; Dillon, 2000), our
understanding of how inducible defences contribute to
patterns in species distribution and abundance remains
limited. Theory suggests that species with inducible
defences against a number of predators (i.e. generalists)
should be more broadly distributed than species that
specialise for a particular predator (Agosta & Klemens,
2008). We find support for this prediction for snail meta-
communities in Michigan, U.S.A. (Hoverman et al.,
2011). Helisoma trivolvis is the most broadly distributed
of the three species with a range that spans temporary
ponds to permanent lakes, H. campanulata is found in
semi-permanent ponds to lakes, and H. anceps is only
found in a subset of lakes. Across this habitat gradient,
there is also a gradient in predator composition; water
bugs are more frequently found in temporary and
semipermanent ponds, while crushing predators such as
crayfish and fish are more common in permanent lakes
(Hoverman et al., 2011). In addition to predation,
however, factors such as habitat size, hydroperiod and
competition can structure snail assemblages (Lodge
et al., 1987; Dillon, 2000; Hoverman et al., 2011).
Freshwater habitats also vary in abiotic conditions that
could influence the interaction between snails and their
predators. For instance, ponds with low calcium concen-
tration or low pH could interfere with shell deposition
in snails and, consequently, constrain the expression of
inducible defences (Thomas et al., 1974; Madsen, 1987;
Rundle et al., 2004). While additional research examining
the interactions between predators and different snail
species in natural populations is needed, our results
suggest that differing defences against predators may be
an important driver of species distributions in fresh-
water snails.
Our study demonstrates that three congeners have
phenotypically plastic strategies that span the full range
of possibilities: specialised defences against a shell-crush-
ing predator, specialised defences against a shell-invading
predator and generalised, flexible defences against both
predators. Theory predicts that a population should
evolve either fixed or plastic phenotypes depending on
the costs and benefits of alternative phenotypes and the
frequency of alternative environments (Pfennig et al.,
2010). However, our work suggests that species can
become somewhat specialised to a given environment, yet
still maintain some plasticity. This may reflect a condition
in which the plasticity has not yet achieved fixation for a
constitutive defence. Alternatively, it may simply be that
each specialist most commonly experiences the presence
or absence of one type of predator over space and time
(e.g. environments containing either no predators or a
shell-crushing predator) but rarely experiences the other
type of predator (e.g. a shell-invading predator). In such a
scenario, there would be little selective pressure to evolve
responses to the other type of predator. While more
research needs to be conducted on this system to under-
stand how selection shapes the evolution of phenotypic
plasticity, it is clear that an understanding of absolute
constraints on body shape, combined with knowledge of
allocation tradeoffs, can provide critical insights into the
mechanisms that produce interspecific variation in the
expression of phenotypic plasticity and patterns of
phenotypic diversification.
Acknowledgments
We thank N. Diecks, B. French, D. Jones, P. Monahan, P.
Noyes and A. Stoler for their assistance with the
experiment and S. Bagnull for measuring the snails. This
work was supported by a National Science Foundation
grant to RAR, and grants from the Conchologists of
America, the Pennsylvania Academy of Science, and
Sigma Xi to JTH.
References
Agosta S.J. & Klemens J.A. (2008) Ecological fitting by phe-
notypically flexible genotypes: implications for species
associations, community assembly and evolution. Ecology
Letters, 11, 1123–1134.
Auld J.R., Agrawal A.A. & Relyea R.A. (2010) Re-evaluating
the costs and limits of adaptive phenotypic plasticity.
Proceedings of the Royal Society B: Biological Sciences, 277,
503–511.
Berendonk T.U., Barraclough T.G. & Barraclough J.C. (2003)
Phylogenetics of pond and lake lifestyles in Chaoborus
midge larvae. Evolution, 57, 2173–2178.
© 2014 John Wiley & Sons Ltd, Freshwater Biology, doi: 10.1111/fwb.12332
10 J. T. Hoverman et al.

Bourdeau P.E. (2009) Prioritized phenotypic responses to
combined predators in a marine snail. Ecology, 90, 1659–
1669.
Brakefield P.M. (2006) Evo-devo and constraints on selec-
tion. Trends in Ecology & Evolution, 21, 362–368.
Brodersen J. & Madsen H. (2003) The effect of calcium con-
centration on the crushing resistance, weight and size of
Biomphalaria sudanica (Gastropoda: Planorbidae). Hydrobio-
logia, 490, 181–186.
Brown K.M. (1998) The role of shell strength in selective
foraging by crayfish for gastropod prey. Freshwater Biol-
ogy, 40, 255–260.
Chivers D.P. & Smith R.J.F. (1998) Chemical alarm signaling
in aquatic predator-prey systems: a review and prospec-
tus. Ecoscience, 5, 338–352.
Colbourne J.K., Hebert P.D.N. & Taylor D.J. (1997) Evolu-
tionary origins of phenotypic diversity in Daphnia. In:
Molecular Evolution and Adaptive Radiation (Eds T.J.
Givnish & K.J. Sytsma), pp. 163–188. Cambridge Univer-
sity Press, Cambridge, UK.
Cotton P.A., Rundle S.D. & Smith K.E. (2004) Trait compen-
sation in marine gastropods: shell shape, avoidance
behavior, and susceptibility to predation. Ecology, 85,
1581–1584.
Dewitt T.J. (1998) Costs and limits of phenotypic plasticity:
tests with predator-induced morphology and life history
in a freshwater snail. Journal of Evolutionary Biology, 11,
465–480.
Dewitt T.J. & Langerhans R.B. (2003) Multiple prey traits,
multiple predators: keys to understanding complex com-
munity dynamics. Journal of Sea Research, 49, 143–155.
Dewitt T.J., Robinson B.W. & Wilson D.S. (2000) Functional
diversity among predators of a freshwater snail imposes
an adaptive trade-off for shell morphology. Evolutionary
Ecology Research, 2, 129–148.
Dewitt T.J., Sih A. & Hucko J.A. (1999) Trait compensation
and cospecialization in a freshwater snail: size, shape and
antipredator behaviour. Animal Behaviour, 58, 397–407.
Dewitt T.J., Sih A. & Wilson D.S. (1998) Costs and limits of
phenotypic plasticity. Trends in Ecology & Evolution, 13,
77–81.
Dillon R.T. (2000) The Ecology of Freshwater Molluscs. Cam-
bridge University Press, Cambridge, UK.
Edgell T.C. & Miyashita T. (2009) Shell shape and tissue
withdrawal depth in 14 species of temperate intertidal
snail. Journal of Molluscan Studies, 75, 235–240.
Harvell C.D. (1991) Coloniality and inducible polymor-
phism. American Naturalist, 138, 1–14.
Hoverman J.T., Auld J.R. & Relyea R.A. (2005) Putting prey
back together again: integrating predator-induced behav-
ior, morphology, and life history. Oecologia, 144, 481–491.
Hoverman J.T., Davis C.J., Werner E.E., Skelly D.K., Relyea
R.A. & Yurewicz K.L. (2011) Environmental gradients
and the structure of freshwater snail communities. Ecog-
raphy, 34, 1049–1058.
Hoverman J.T. & Relyea R.A. (2007a) How flexible is phe-
notypic plasticity? Developmental windows for trait
induction and reversal. Ecology, 88, 693–705.
Hoverman J.T. & Relyea R.A. (2007b) The rules of engage-
ment: how to defend against combinations of predators.
Oecologia, 154, 551–560.
Hoverman J.T. & Relyea R.A. (2008) Temporal environmen-
tal variation and phenotypic plasticity: a mechanism
underlying priority effects. Oikos, 117, 23–32.
Hoverman J.T. & Relyea R.A. (2009) Survival trade-offs
associated with inducible defences in snails: the roles of
multiple predators and developmental plasticity. Func-
tional Ecology, 23, 1179–1188.
Kemp P. & Bertness M.D. (1984) Snail shape and growth
rates: evidence for plastic shell allometry in Littorina litto-
rea. Proceedings of the National Academy of Sciences of the
United States of America, 81, 811–813.
Kusch J. (1993) Behavioral and morphological changes in
Ciliates induced by the predator Ameba proteus. Oecologia,
96, 354–359.
Lodge D., Brown K.M., Klosiewski S., Stein R., Covich A.,
Leathers B. et al. (1987) Distribution of freshwater snails:
spatial scale and relative important of physicochemical
and biotic factors. American Malacological Bulletin, 5, 73–
84.
Lodge D.M., Kershner M.W., Aloi J.E. & Covich A.P. (1994)
Effects of an omnivorous crayfish (Orconectes Rusticus) on
a freshwater littoral food-web. Ecology, 75, 1265–1281.
Lodge D.M. & Lorman J.G. (1987) Reductions in submersed
macrophyte biomass and species richness by the crayfish
Orconectes rusticus. Canadian Journal of Fisheries and Aquatic
Sciences, 44, 591–597.
Madsen H. (1987) Effect of calcium concentration on growth
and egg laying of Helisoma duryi, Biomphalaria alexandrina,
Biomphalaria camerunensis and Bulinus truncatus (Gastro-
poda: Planorbidae). Journal of Applied Ecology, 24, 823–836.
Osenberg C.W. & Mittelbach G.G. (1989) Effects of body
size on the predator-prey interaction between pumpkin-
seed sunfish and gastropods. Ecological Monographs, 59,
405–432.
Palmer A.R. (1981) Do carbonate skeletons limit the rate of
body growth? Nature, 292, 150–152.
Pfennig D.W., Wund M.A., Snell-Rood E.C., Cruickshank
T., Schlichting C.D. & Moczek A.P. (2010) Phenotypic
plasticity’s impacts on diversification and speciation.
Trends in Ecology & Evolution, 25, 459–467.
Pigliucci M. (2001) Phenotypic Plasticity: Beyond Nature and
Nurture. Johns Hopkins University Press, Baltimore, MD,
U.S.A.
Raup D.M. (1962) Computer as aid in describing form in
gastropod shells. Science, 138, 150–152.
Rice S.H. (1998) The bio-geometry of mollusc shells. Paleobi-
ology, 24, 133–149.
Rundle S.D. & Br€onmark C. (2001) Inter- and intraspecific
trait compensation of defence mechanisms in freshwater
© 2014 John Wiley & Sons Ltd, Freshwater Biology, doi: 10.1111/fwb.12332
Defensive strategies of Helisoma snails 11

snails. Proceedings of the Royal Society of London Series
B-Biological Sciences, 268, 1463–1468.
Rundle S.D., Spicer J.I., Coleman R.A., Vosper J. & Soane J.
(2004) Environmental calcium modifies induced defences
in snails. Proceedings of the Royal Society of London Series
B-Biological Sciences, 271, S67–S70.
Russell-Hunter W.D. (1978) Ecology of freshwater pulmo-
nates. In: The Pulmonates. Vol. 2A: Systematics, Evolution,
and Ecology (Eds V. Fretter & J. Peake ), pp. 335–383. Aca-
demic Press, Orlando, FL, U.S.A.
Schlichting C.D. & Pigliucci M. (1998) Phenotypic Evolution:
A Reaction Norm Perspective. Sinauer, Sunderland, MA,
U.S.A.
Thomas J.D., Benjamin M., Lough A. & Aram R.H. (1974)
The effects of calcium in the external environment on the
growth and natality rates of Biomphalaria glabrata. Journal
of Animal Ecology, 43, 839–860.
Tollrian R. & Harvell D. (1999) The Ecology and Evolution of
Inducible Defenses. Princeton University Press, Princeton,
NJ, U.S.A.
Trussell G.C. & Nicklin M.O. (2002) Cue sensitivity, induc-
ible defense, and trade-offs in a marine snail. Ecology, 83,
1635–1647.
Turner A.M., Bernot R.J. & Boes C.M. (2000) Chemical cues
modify species interactions: the ecological consequences
of predator avoidance by freshwater snails. Oikos, 88,
148–158.
Van Buskirk J. (2002) A comparative test of the adaptive
plasticity hypothesis: relationships between habitat and
phenotype in anuran larvae. American Naturalist, 160,
87–102.
Van Kleunen M. & Fischer M. (2007) Progress in the detec-
tion of costs of phenotypic plasticity in plants. New Phy-
tologist, 176, 727–730.
Van Tienderen P.H. (1991) Evolution of generalists and spe-
cialists in spatially heterogeneous environments. Evolu-
tion, 45, 1317–1331.
(Manuscript accepted 13 January 2014)
© 2014 John Wiley & Sons Ltd, Freshwater Biology, doi: 10.1111/fwb.12332
12 J. T. Hoverman et al.