Embed Size (px)
Citation preview

Genetic Analysis of the Role of yfiR in the Ability of Escherichia coliCFT073 To Control Cellular Cyclic Dimeric GMP Levels and ToPersist in the Urinary Tract
Erica L. Raterman, Daniel D. Shapiro, Daniel J. Stevens, Kevin J. Schwartz, Rodney A. Welch
Department of Medical Microbiology and Immunology, University of Wisconsin—Madison, Madison, Wisconsin, USA
During urinary tract infections (UTIs), uropathogenic Escherichia coli must maintain a delicate balance between sessility andmotility to achieve successful infection of both the bladder and kidneys. Previous studies showed that cyclic dimeric GMP (c-di-GMP) levels aid in the control of the transition between motile and nonmotile states in E. coli. The yfiRNB locus in E. coliCFT073 contains genes for YfiN, a diguanylate cyclase, and its activity regulators, YfiR and YfiB. Deletion of yfiR yielded a mu-tant that was attenuated in both the bladder and the kidneys when tested in competition with the wild-type strain in the murinemodel of UTI. A double yfiRN mutant was not attenuated in the mouse model, suggesting that unregulated YfiN activity andlikely increased cytoplasmic c-di-GMP levels cause a survival defect. Curli fimbriae and cellulose production were increased inthe yfiR mutant. Expression of yhjH, a gene encoding a proven phosphodiesterase, in CFT073 �yfiR suppressed the overproduc-tion of curli fimbriae and cellulose and further verified that deletion of yfiR results in c-di-GMP accumulation. Additional dele-tion of csgD and bcsA, genes necessary for curli fimbriae and cellulose production, respectively, returned colonization levels ofthe yfiR deletion mutant to wild-type levels. Peroxide sensitivity assays and iron acquisition assays displayed no significant dif-ferences between the yfiR mutant and the wild-type strain. These results indicate that dysregulation of c-di-GMP productionresults in pleiotropic effects that disable E. coli in the urinary tract and implicate the c-di-GMP regulatory system as an impor-tant factor in the persistence of uropathogenic E. coli in vivo.
Uropathogenic Escherichia coli (UPEC) organisms are the mostprevalent isolates of patients with urinary tract infections
(UTIs). Unlike many other bacterial infections, UTIs are commonin young healthy women and are among the most frequently di-agnosed infections in the United States (1–5). The contributionsof colonization and virulence factors such as type I fimbriae, ironacquisition systems, and hemolysin to successful infection of theurinary tract by UPEC are well documented (6–14). However, thecontribution of second messengers such as bis-(3=-5=)-cyclic di-meric GMP (c-di-GMP) to virulence have thus far not been welldocumented for UPEC. Given the recent advances in the under-standing of the downstream effectors of c-di-GMP, interest in therole of c-di-GMP signaling in the virulence of bacterial pathogenshas increased.
c-di-GMP is created by diguanylate cyclases (DGCs) from twoGTP molecules and is broken down to 5=-phosphoguanylyl-(3=-5=)-guanosine (pGpG) by specific phosphodiesterases (PDEs).Other enzymes are responsible for further breaking down pGpGto two GMP molecules. All DGCs possess the GGDEF (Gly-Gly-Asp-Glu-Phe) active-site domain, and PDEs are associated withthe EAL (Glu-Ala-Leu) domain (15). Various external or internalstimuli are sensed by the DGCs and translated to the various c-di-GMP effector molecules to alter the behavior of the bacteria. Be-cause c-di-GMP effectors are present at all levels of regulation,including transcriptional, translational, and posttranslationalfunctions, c-di-GMP signaling can produce rapid responses toquickly changing environmental conditions. The most commonalterations made by c-di-GMP signaling in E. coli are changes inmotility and production of adhesins and exopolysaccharides. Ingeneral, increased levels of c-di-GMP decrease motility and in-crease multicellular community behaviors, while the reverse istrue when c-di-GMP levels are low (16–27).
Groups working with nonpathogenic K-12 strains of E. colihave established a link between curli fimbria production and c-di-GMP levels (20, 28, 29). Curli fimbriae are coiled surface struc-tures that promote both adhesion to surfaces as well as cell-cellinteractions. The curli locus is organized in two divergent operons(csgDEFG and csgBA), and the transcriptional activator CsgDstimulates expression of csgBA as well as its own operon. CsgA isthe major subunit of the curli fiber, and CsgB is needed for thenucleation of the CsgA oligomer. Increased levels of c-di-GMPlead to increased activity of CsgD, although this effect likely in-volves other intermediary proteins (20, 29–32). Various studieshave demonstrated the ability of curli fimbriae to induce an im-mune response through Toll-like receptor 2 (TLR2) activationand generation of proinflammatory fibrinopeptides and by acti-vating inducible nitric oxide synthase, a defense mechanism thatproduces large amounts of nitric oxide (33–35). Increased bacte-rial resistance to the antimicrobial peptide LL-37 in curli-fimbri-ated E. coli has also been demonstrated (36).
Increased activity of CsgD also indirectly affects the produc-tion of cellulose. Expression of adrA and yedQ, two genes that
Received 10 December 2012 Returned for modification 14 January 2013Accepted 28 May 2013
Published ahead of print 17 June 2013
Editor: J. B. Bliska
Address correspondence to Rodney A. Welch, [email protected].
Supplemental material for this article may be found at http://dx.doi.org/10.1128/IAI.01396-12.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/IAI.01396-12
September 2013 Volume 81 Number 9 Infection and Immunity p. 3089–3098 iai.asm.org 3089
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

encode DGCs, is activated by CsgD. The subsequent productionof c-di-GMP by either or both of the proteins AdrA and YedQ,depending on the strain of E. coli, stimulates increased activity ofBcsA, the catalytic subunit of the cellulose biosynthetic machin-ery. Transcription levels of the cellulose operons (bcsEFG andyhjRQbcsABZC) do not seem to be affected by c-di-GMP levels(16, 20, 31, 37–39). Production of cellulose in concert with curlifimbriae seems to dampen the immune response produced by thecurli fibers (36). The coproduction of these factors also produces adistinctive in vitro plate phenotype termed the rdar morphotype,for the red, dry and rough colonies observed on agar plates thatcontain Congo red dye (29). The rdar phenotype is similar to thesmall-colony variant (SCV) phenotype seen in Pseudomonasaeruginosa isolates.
This study investigated the links between the DGC YfiN and itsactivity regulator YfiR, first identified in a screen for loss of motil-ity in E. coli K-12 (40), with changes observed in curli fimbriaeproduction, cellulose production, and motility. The implicationsof the expression and activity of these systems for the virulence ofUPEC strain CFT073 in the urinary tract were also assessed usingthe mouse model of UTI. Deletion of yfiR and subsequent in-creased activity of YfiN lead to increased production of curli fim-briae and cellulose and a near total loss of motility. We furthershow that these phenotypes lead to attenuation in the competitivemouse model of UTI. This is the first report to demonstrate thatproper control of c-di-GMP production is essential for UPEC tomount successful infections in the urinary tract.
MATERIALS AND METHODSStrains and media. E. coli strain CFT073 was isolated from a patient withpyelonephritis at the University of Maryland Medical System. Strains weregrown in Luria-Bertani broth (LB), morpholinepropanesulfonic acid(MOPS) glycerol broth, tryptone broth (2 g tryptone and 1 g NaCl in 200ml double-distilled water [ddH2O]), or filter-sterilized human urine col-lected from a female volunteer. Curli fimbriae and cellulose expressionwas determined by growth on modified LB agar plates (10 g tryptone, 5 gyeast extract, and 15 g agar per liter ddH2O with or without 40 �g/mlCongo red and 20 �g/ml Coomassie blue added after autoclaving) orurine agar plates (750 ml sterile human urine mixed with an autoclavedmixture of 15 g agar in 250 ml ddH2O with or without 40 �g/ml Congored and 20 �g/ml Coomassie blue). Swimming motility was assessed in0.3% tryptone agar plates (1 g tryptone, 0.5 g NaCl, and 0.3 g agar per 100ml ddH2O), and swarming motility was assessed on 0.45% swarm agar(0.3 g meat extract, 1 g Bacto peptone, and 0.45 g agar per 100 ml ddH2O,with 0.5 g glucose added after autoclaving). Antibiotics were added in thefollowing concentrations as appropriate: 50 �g/ml kanamycin, 20 �g/mlchloramphenicol, and 200 �g/ml carbenicillin. A 10 mM concentration ofarabinose was added to the media to induce expression from the pBADpromoter when necessary.
Construction of mutant strains, allelic repair, and complementa-tion systems. All primers used for the generation of deletion mutants andcloning are listed in Table S1 in the supplemental material. All nonpolargene deletions were performed using the �-Red recombination systemdeveloped by Datsenko and Wanner (41). The PCR products used fortransformations in CFT073 were generated off pKD3 or pKD4 templatesusing primers specific for the targeted gene. The kanamycin- or chloram-phenicol-marked gene deletions were then transduced into a freshCFT073 background using the EB49 transducing phage (42). The antibi-otic resistance cassette used to replace the target gene was removed usinga Flp recombinase encoded on pCP20 (41). All gene deletions were veri-fied by PCR and loss of antibiotic resistance on LB agar containing theappropriate antibiotic. The yfiR locus was repaired in the yfiR deletionmutant by insertion of the kanamycin cassette via �-Red near, but outside,
the yfiLRNB locus, being careful not to disrupt any other loci. The EB49transducing phage was used to transduce the wild-type yfiLRNB locus intothe yfiR deletion mutant background. Restoration of the locus was con-firmed by PCR and sequencing.
Complementation of the yfiR deletion strain was obtained by cloningPCR-amplified yfiR into the NcoI-PstI restriction sites of pBAD24d. Se-quencing of the resulting plasmid confirmed the insertion of yfiR.pBAD30-yhjH and pBAD30-yhjHE136A were obtained from Ute Römling(15). The lacZ transcriptional fusion strains used for the �-galactosidaseassays were obtained using the pFUSE system (43). Briefly, approximated500 bp of the 5= end of the targeted gene was cloned into the XbaI/SmaIsites of pFUSE to link the gene to the plasmid-encoded lacZYA locus. Theresulting plasmids were used in a tetraparental mating with two otherstrains carrying the transposase on a suicide plasmid for homologousrecombination into a CFT073 �lacZ::Kan background strain. Thismethod produces a transcriptional fusion of the lacZYA operon with thetargeted gene in the CFT073 genome. Kanamycin and chloramphenicolwere used to select positive strains, and inserts were verified by PCR andsequencing.
Murine model of UTI. Colonization of the urinary tract was deter-mined using the competitive murine model of urinary tract infection asdescribed previously (44). CFT073�lacZYA was used as the wild-typestrain, and the various mutants had an intact lacZYA locus. To select forpiliated bacteria, all bacterial strains were grown statically in 3 ml LB at37°C for 2 days. The pellicle formed on the rim of the test tube was thentransferred to fresh LB, incubated for 2 more days, and, finally, passagedagain to 40 ml LB for a final, 2-day incubation. The broths were adjustedto an optical density at 600 nm (OD600) �0.4 with 1� phosphate-buff-ered saline (PBS), and the wild-type strain and the mutant strain weremixed equally. The mixed broth was then pelleted and washed once in 1�PBS and resuspended in 500 �l 1� PBS. Isoflurane-anesthetized 6- to7-week-old female Swiss Webster mice (Harlan, USA) were inoculated viaurethral catheterization with 50 �l (�108 CFU) mixed bacterial suspen-sion. Mice were euthanized via CO2 asphyxiation, and the bladder andkidneys were aseptically harvested in 1� PBS. The organs were homoge-nized, serially diluted in 1� PBS, and plated on MacConkey agar medium(Fisher, USA). Ratios of surviving strains were determined by countingwhite (wild-type) and red (mutant) colonies. Colonization levels weregraphed and analyzed using a paired Wilcoxon signed-ranked test andPrism (GraphPad). Competitive indices were calculated by dividing mu-tant CFU by wild-type CFU from the mice and then dividing this ratio bythe ratio of the mutant to the wild type in the original inoculum. Allanimal experiments were approved by the UW-Madison Animal Care andUse Committee.
�-Galactosidase assay. All �-galactosidase assays were performed aspreviously described (20). Briefly, bacterial strains were grown on LB agarplates without salt and containing 200 �g/ml carbenicillin and 10 mMarabinose if necessary. Plates were incubated for 16 to 20 h at 30°C. Thebacteria were then collected with cotton swabs and suspended in 3 mltryptone broth without salt to an OD600 between 0.4 and 0.8. Five hun-dred microliters each broth was mixed with 500 �l Z buffer (60 mMNa2HPO4, 60 mM NaH2PO4, 10 mM KCl, 1 mM MgSO4, �-mercapto-ethanol [pH 7.0]), 20 �l chloroform, and 10 �l 0.1% SDS and vortexed for10 s. Samples were then incubated at 28°C for 5 min, and 200 �l 4-mg/mlo-nitrophenyl-�-D-galactopyranoside (ONPG) was then added. The re-action was stopped with 500 �l Na2CO3. The OD420 and OD550 were thenrecorded, and the Miller units were calculated according to the followingequation: Miller units � 1,000 � {[OD420 � (1.75 � OD550)]/(t � V �OD600)}, where t is the reaction time in min and V is the volume ofbacterial broth used in milliliters.
Congo red and calcofluor binding assays. Congo red binding assayswere performed as previously described (45). Briefly, strains wereswabbed on LB agar plates without salt containing 200 �g/ml carben-icillin and 10 mM arabinose if necessary. Plates were incubated for 16to 20 h at 30°C. The bacteria were collected with cotton swabs and
Raterman et al.
3090 iai.asm.org Infection and Immunity
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

suspended in 5 ml tryptone broth with 40 �g/ml Congo red (Sigma-Aldrich) and without NaCl. Samples were taken for dilution plating onLB plates, and broths were separated into two tubes and incubatedwith shaking at 37°C for 2 h. The bacteria were pelleted at 8,000 rpm,and the OD490 was determined for the supernatants. The amount ofCongo red dye left in the supernatant was determined using a standardcurve of known concentrations of the dye.
Congo red plates were used to determine the rdar morphotype of eachstrain as previously described (45). Strains were grown statically in 3 mlLB at 37°C for 2 days. The pellicle formed on the rim of the test tube wasthen transferred to fresh LB, incubated for 2 more days, and, finally, pas-saged again to 3 ml LB for a final 2-day incubation. Broths were dilutionplated onto Congo red plates and incubated for 5 to 7 days at 30°C. Colonymorphology was then recorded. To determine cellulose production,strains were streaked on LB plates without salt containing 200 �g/mlcalcofluor (Sigma-Aldrich) and 10 mM arabinose if necessary. Plates weregrown at 30°C overnight before the results were recorded.
Western blot analysis of CsgA expression. Strains were grown over-night at 30°C on LB plates without salt with carbenicillin and arabinose forinduction of pBAD-yfiR strains. Bacteria were collected using cottonswabs and resuspended in 1 ml 1� PBS. A sample of each broth was takenfor plate counts, and the remaining sample was prepared according torecommendations from the laboratory of Matt Chapman at the Univer-sity of Michigan, Ann Arbor, MI. The bacteria were pelleted at 8,000 rpm,resuspended in 100 �l 99% formic acid, and incubated for 10 min on ice.Samples were then evaporated to dryness using a Speedvac. Pellets wereresuspended in 200 �l 2� CRACK buffer (4% [wt/vol] sodium dodecylsulfate, 0.2% [wt/vol] bromophenol blue, 20% [vol/vol] glycerol, 200 mMdithiothreitol). The Precision Plus Protein dual standard (Bio-Rad, USA)and the samples were then loaded on 15% SDS-PAGE gels and run at 250V for 3 h. Amounts loaded were normalized using the CFU obtained fromthe plate counts. Gels were either Coomassie stained or transferred tonitrocellulose at 40 V for 1 h. Nitrocellulose membranes were thenblocked with 5% milk in 1� Tris-buffered saline with Tween 20 (TBST)for 1 h at room temperature. Anti-CsgA antibody (1:7,000) (obtainedcourtesy of the Chapman laboratory) was added, and blots were incubatedovernight at 4°C. Blots were then washed 3 times with 1� TBST andprobed with anti-rabbit horseradish peroxidase (HRP) (1:20,000) in 5%milk in 1� TBST for 30 min at room temperature. Blots were washed with1� TBST 3 more times and developed using an enhanced chemilumines-cence (ECL) kit (GE Healthcare, USA).
Motility assays. Swimming motility was assessed in 0.3% agar tryp-tone plates. Strains were grown overnight at 37°C in tryptone broth.Broths were adjusted to an OD600 of �0.1 with fresh tryptone broth before2 �l was injected in the center of the agar swim plates. Ten-microlitervolumes of the same broths were placed on the surface of swarm agarplates to assess swarming motility. Swim plates were incubated for 12 h at
30°C, and swarm plates were incubated for 2 days at 30°C. Assays wereindependently repeated at least three times.
RESULTSPredicted functions of the YfiLRNB system of E. coli CFT073 aresimilar to those of the YfiBNR system of P. aeruginosa. Recentreports on P. aeruginosa identified the YfiBNR system as a regula-tory complex that plays a role in the development of the SCVphenotype important for persistence in a subcutaneous cathetermodel. YfiN was identified as an active membrane-associatedDGC, and YfiR was identified as a periplasmic regulator of YfiNactivity (46, 47). The authors proposed that YfiB sequesters YfiR tothe outer membrane, which, in turn, relieves repression of YfiNactivity (47). In the analogous operon, E. coli contains an addi-tional gene, yfiL, that has no predicted function (Fig. 1A). Liketheir P. aeruginosa counterparts, E. coli YfiN has a predicted ac-tive-site domain with the conserved GGDEF sequence, while YfiRhas a predicted signal sequence for export to the periplasm with apredicted cleavage site between amino acids 22 and 23 (Fig. 1B).YfiN also has a predicted allosteric inhibition site for feedbackinhibition by c-di-GMP (Fig. 1B). However, the sequence identityat both the nucleotide and protein levels between the P. aeruginosaand E. coli proteins is low, with only 30% and 39% amino acididentity between the two predicted YfiR proteins and YfiN pro-teins, respectively (Fig. 1B).
A yfiR mutant is attenuated in vivo. A yfiR mutant of P. aerugi-nosa was attenuated in the acute phase of a murine skin infectionmodel (46). To test this phenotype in E. coli CFT073, a nonpolaryfiR deletion mutant was constructed and tested in competitionwith the wild-type strain in the murine model of urinary tractinfection. As shown in Fig. 2, the yfiR mutant was attenuated in
FIG 1 Organization of the yfiLRNB locus and predicted protein functions. (A)The yfiLRNB locus consists of four genes that are expressed off a single pre-dicted promoter. (B) YfiR is a predicted periplasmic protein of unknown func-tion with a signal sequence (S.S.), and YfiN is a predicted membrane-bounddiguanylate cyclase with an intact GGDEF active site and an allosteric inhibi-tion site (I site). YfiL and YfiB are predicted lipoproteins of unknown function.
FIG 2 Deletion of yfiR causes attenuation in the mouse model of competitiveUTI. Shown are results for coinfection of female Swiss Webster mice withCFT073 �yfiR (n � 21) (A), CFT073 �yfiN (n � 26) (B), CFT073� yfiRN (n �16) (C), or the CFT073 �yfiR phage restoration mutant (n � 18) (D) andCFT073 �lacZYA using the mouse model of UTI. The competitive indices werecalculated by the following equation: (yfi mutant CFU recovered/wild-typeCFU recovered)/(mutant inoculum CFU/wild-type inoculum CFU). The solidline indicates the median value.
c-di-GMP Level Control in Escherichia coli CFT073
September 2013 Volume 81 Number 9 iai.asm.org 3091
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

both the bladder (P � 0.0008) and kidneys (P � 0.004) at averagesof 7.4-fold and 24.8-fold, respectively, at 48 h postinfection (Fig.2A). Single deletions of yfiN (Fig. 2B), yfiL (see Fig. S1A in thesupplemental material), and yfiB (see Fig. S1B) were not attenu-ated in vivo. Similarly, deletion of yfiN from the yfiR mutant res-cued the competitive defect of the yfiR single mutant (Fig. 2C),suggesting that unrestricted activity of YfiN c-di-GMP productionleads to attenuation in the urinary tract. Allelic restoration of theyfiR mutation to its wild-type form by transduction also resultedin loss of attenuation in vivo (Fig. 2D). Single infections with theyfiR mutant and wild-type CFT073 also showed that the yfiR mu-tant is carried at lower loads in the bladder and kidneys than thewild-type strain at 48 h postinfection (see Fig. S1C). Growthcurves performed in LB, minimal media, and urine showed nodifference in the growth rates between wild-type CFT073 and theyfiR mutant (see Fig. S2A). The yfiR mutant also competed like thewild-type strain in competitive LB broth and urine cultures (seeFig. S2B). These results indicate that a simple growth defect isunlikely to account for the attenuation of the yfiR mutant in theurinary tract.
Growth on agar plates induces expression of the yfiRNB lo-cus. To determine the optimal conditions for expression of yfiRand yfiN, transcriptional fusions linking yfiR and yfiN to lacZYAwere made using the pFUSE system (43). Strains were grown toexponential or late log phase in tryptone broth cultures or on LBagar plates overnight. �-Galactosidase assays were then per-formed on the strains. No expression was detected in the brothcultures (data not shown), but expression was detected when thestrains were grown on the LB agar plates (see Fig. S3). This indi-cates that expression of the yfiRNB locus may be surface induced.
Deletion of yfiR causes a motility defect. The detrimental ef-fects of increased levels of c-di-GMP on motility are well docu-mented (15, 22). To determine whether the mutant strains used inthis study also displayed motility defects, the strains were grown in0.3% tryptone swim plates and compared to the wild-type strain.A representative image of a swim plate and the average diameter ofeach strain’s swim ring are shown in Fig. 3A and B. As expected,the yfiR mutant showed a drastic decline in its ability to swim outin the agar plates. The yfiN, as previously shown (48), and yfiRNmutants behaved like the wild-type strain. Complementation ofthe yfiR mutant with arabinose-induced pBADyfiR restored mo-tility to the wild-type level, as did restoration of the yfi locus witha CFT073 transducing phage. The swimming defect observed withthe yfiR deletion strain does not seem to be due to an inability toassemble the flagella. Intact flagellar filaments can be observed inelectron micrographs of the cells, and motility is seen in live bac-teria observed under a light microscope (Fig. 3C). This suggeststhat the motility defect observed in the agar swim plates for theyfiR deletion mutant may be caused by the bacteria becomingstuck in the agar rather than the complete lack of functional fla-gella.
Deletion of yfiR causes increased production of cellulose andcurli fimbriae. In other strains of E. coli, as well as other species ofbacteria, an increase in c-di-GMP levels usually results in in-creased fimbriation and exopolysaccharide production. E. colistrains specifically upregulate production of curli fimbriae andcellulose (15, 20, 22, 29, 49). A phenotypic assay to test whethercurli fimbriae and cellulose are being produced involves platingthe bacteria on LB plates that contain Congo red dye but lack salt.After 5 to 8 days of growth, colonies that express both products
will have a deep red, rugose phentotype that is referred to as therdar morphotype (for red, dry and rough). Colonies that produceonly curli fimbriae will lack the rugose characteristic but will stillbe deep red and slightly rough. Expression of cellulose only resultsin a rugose colony that is pink instead of deep red, and loss ofexpression of both products results in a colony that is white andsmooth (29, 50). Wild-type CFT073 and the yfiRNB locus mutantswere grown for 6 days in LB at 37°C without shaking before serialdilution and plating on LB Congo red plates. The colony pheno-types obtained for each strain under these conditions are shown inFig. 4A. All colonies obtained for the yfiR deletion mutant dis-played the rdar morphotype, while the wild-type strain and theyfiN and yfiRN deletion mutants produced a mix of rdar andsmooth and white colonies. Both the rdar and the smooth andwhite colony types of the wild-type strain would give a mixture ofsmooth and white and rdar colonies when replated on fresh LBCongo red plates (data not shown). The two distinct colony types
FIG 3 Motility phenotypes of mutants with alterations in the yfiLRNB locus.(A) Swim phenotype in 0.3% tryptone agar plates with 10 mM arabinosegrown at 30°C. (B) Average diameters of swim rings of strains grown in 0.3%tryptone agar plates at 30°C. (C) Transmission electron micrographs of wild-type CFT073 and CFT073 �yfiR grown in 0.3% tryptone agar plates at 30°Cand 37°C.
Raterman et al.
3092 iai.asm.org Infection and Immunity
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

obtained for the wild-type strain and their ability to revert to theopposite morphology suggest that at some point during theirgrowth, the bacteria commit to a regulated state that results ineither the production or suppression of curli and cellulose. It isnot clear at this time what the signal(s) may be that promoteseither state or how those signals are propagated through the prog-eny to ultimately produce a colony that is either all rdar or allsmooth and white. Restoration of the yfiR mutation via transduc-
tion returned the phenotype of the yfiR deletion mutant to a mix-ture of both rdar and smooth and white colonies (Fig. 4A and B).Phenotypes on plates containing calcofluor, a fluorescent dye thatbinds cellulose, confirmed that yfiR deletion strains produce cel-lulose (see Fig. S4 in the supplemental material). OverexpressionyfiR carried on pBAD in the CFT073� yfiR background producedall smooth colonies, although most were slightly rough in the cen-ter, presumably from loss of arabinose induction as a result of
%rd
ar
∆yfiRpBADyfiR
∆yfiRpBAD∆yfiRWild-type
∆yfiR∆csgA
FIG 4 Curli fimbriae and cellulose production are increased in CFT073 �yfiR. (A) Phenotypes of yfiLRNB locus mutants grown on LB Congo red plates at 30°C.All colony phenotypes that were observed are pictured. (B) Percentage of mutant colonies from the LB Congo red plates displaying the rdar phenotype. (C)Congo red binding assay in liquid broth. The amount of Congo red dye bound is presented as pg of bound dye per CFU. P values were calculated using Student’st test. (D) �-Galactosidase assays performed on wild-type CFT073 and CFT073�yfiR strains with the csgD::lacZYA transcriptional fusion. P values were calculatedusing Student’s t test. (E) Western blot of acid-treated, whole-cell preparations grown on LB no-salt plates at 30°C. A rabbit anti-CsgA antibody was used to detectCsgA monomers.
c-di-GMP Level Control in Escherichia coli CFT073
September 2013 Volume 81 Number 9 iai.asm.org 3093
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
metabolism (see Fig. S5A). Congo red binding assays were alsoperformed on lawns of bacteria grown overnight at 30°C on LBagar plates without NaCl. As expected, the yfiR mutant boundmore Congo red dye per CFU than the other strains, and thisincrease in bound Congo red was mitigated either by restorationof the mutated yfi locus to the wild type or by plasmid-expressedyfiR (Fig. 4C).
To determine whether deletion of yfiR altered expression ofcsgD, the promoterless lacZYA locus was inserted directly down-stream of csgD to create a transcriptional fusion using the pFUSEsystem (43). Expression levels of csgD could then be monitored by�-galactosidase assays. Deletion of yfiR resulted in a drastic in-crease in the Miller units expressed off the csgD operon (Fig. 4D).Deletion of both yfiR and yfiN in the csgD::lacZYA strain returnedcsgD expression to wild-type levels, as did complementation withpBADyfiR in the yfiR deletion strain (Fig. 4D). Increased produc-tion of the curli fibrils was verified by Western blotting using anantibody specific for CsgA, the major curlin subunit (Fig. 4E).Electron micrographs of the wild-type and yfiR mutant strains alsoshowed an increase in what appear to be curli fibrils in the yfiRmutant (see Fig. S6). Plate phenotypes and expression of the adrAoperon suggested that expression of adrA, but not yedQ (data notshown), is affected by CsgD activity (see Fig. S5A and B). Expres-sion off the bcs operon was also monitored using the same systemto place lacZYA downstream of bcsA. However, bcsA and bcsEexpression levels in the mutant strains remained unchanged fromthe wild-type strain (see Fig. S7). This is not surprising given thatc-di-GMP is thought to bind directly to BcsA to increase its activ-ity levels and, thus, elevate cellulose production.
As CFT073 did not form a robust biofilm in plastic plates inour hands, pellicle formation on sterile glass test tubes wasmonitored as the closest equivalent to the biofilm plate assay.Deletion of yfiR dramatically increased pellicle formation. As inthe Congo red binding assay, additional deletion of yfiN orcomplementation with a plasmid-borne copy of yfiR in the yfiRdeletion mutant returned pellicle formation to the wild-typelevel (see Fig. S8A and B).
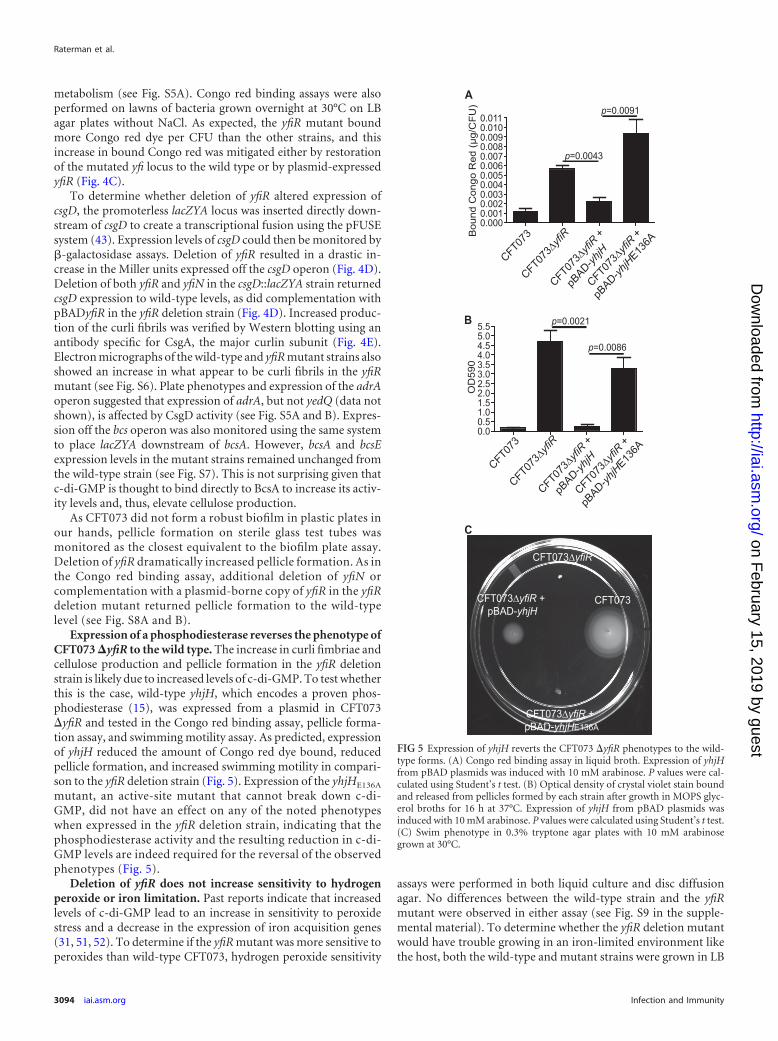
Expression of a phosphodiesterase reverses the phenotype ofCFT073 �yfiR to the wild type. The increase in curli fimbriae andcellulose production and pellicle formation in the yfiR deletionstrain is likely due to increased levels of c-di-GMP. To test whetherthis is the case, wild-type yhjH, which encodes a proven phos-phodiesterase (15), was expressed from a plasmid in CFT073�yfiR and tested in the Congo red binding assay, pellicle forma-tion assay, and swimming motility assay. As predicted, expressionof yhjH reduced the amount of Congo red dye bound, reducedpellicle formation, and increased swimming motility in compari-son to the yfiR deletion strain (Fig. 5). Expression of the yhjHE136A
mutant, an active-site mutant that cannot break down c-di-GMP, did not have an effect on any of the noted phenotypeswhen expressed in the yfiR deletion strain, indicating that thephosphodiesterase activity and the resulting reduction in c-di-GMP levels are indeed required for the reversal of the observedphenotypes (Fig. 5).
Deletion of yfiR does not increase sensitivity to hydrogenperoxide or iron limitation. Past reports indicate that increasedlevels of c-di-GMP lead to an increase in sensitivity to peroxidestress and a decrease in the expression of iron acquisition genes(31, 51, 52). To determine if the yfiR mutant was more sensitive toperoxides than wild-type CFT073, hydrogen peroxide sensitivity
assays were performed in both liquid culture and disc diffusionagar. No differences between the wild-type strain and the yfiRmutant were observed in either assay (see Fig. S9 in the supple-mental material). To determine whether the yfiR deletion mutantwould have trouble growing in an iron-limited environment likethe host, both the wild-type and mutant strains were grown in LB
FIG 5 Expression of yhjH reverts the CFT073 �yfiR phenotypes to the wild-type forms. (A) Congo red binding assay in liquid broth. Expression of yhjHfrom pBAD plasmids was induced with 10 mM arabinose. P values were cal-culated using Student’s t test. (B) Optical density of crystal violet stain boundand released from pellicles formed by each strain after growth in MOPS glyc-erol broths for 16 h at 37°C. Expression of yhjH from pBAD plasmids wasinduced with 10 mM arabinose. P values were calculated using Student’s t test.(C) Swim phenotype in 0.3% tryptone agar plates with 10 mM arabinosegrown at 30°C.
Raterman et al.
3094 iai.asm.org Infection and Immunity
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

with 2,2-dipyridyl, an iron chelator. Again, no differences ingrowth were observed between the wild-type and yfiR mutantstrains (see Fig. S10).
Deletion of ycgR in the yfiR mutant partially restores coloni-zation levels in the mouse model. Motility is a known fitness traitof uropathogenic E. coli strains in the urinary tract and is neededfor ascension of the ureters to reach the kidneys (13, 53, 54).Recent studies indicate that YcgR can bind c-di-GMP and theninteract with the flagellar motor. This interaction acts to eitherinhibit rotation or keep the flagella turning counterclockwise,causing smooth-swimming cells that may become stuck in theagar matrix (55, 56). To determine whether the swimming de-fect seen in the yfiR mutant causes the attenuated phenotype invivo, ycgR was deleted from the yfiR mutant and tested in thecompetitive mouse model of UTI. Swim plates of this mutantconfirmed that motility was partially restored (see Fig. S11 inthe supplemental material). As shown in Fig. 6A, this partialrestoration of motility did not result in recovery of the yfiRdeletion mutant to wild-type colonization levels in vivo. Byitself, a single ycgR deletion mutant was not attenuated in thecompetitive mouse model against wild-type CFT073 (see Fig.S12A). Given that the colonization levels were not restored, fullmotility may be needed for full virulence, or another factormay also be partially responsible for the observed attenuationof the yfiR deletion mutant.
Deletion of both bcsA and csgD in the yfiR mutant returnscolonization to wild-type levels. Previous studies have shownthat curli fibers produce an immune response by activating thehost coagulation system and thus promoting an influx of whiteblood cells (34). However, curli fimbriae can also promote resis-tance to the host antimicrobial peptide LL-37 (36). Additionalmutations in the yfiR deletion strain were therefore made to de-termine if the increased expression of curli fimbriae, or possiblyeven the increase in cellulose production, could be responsible forthe attenuation of the yfiR deletion strain in vivo. Deletion of csgDor bcsA from the yfiR mutant did not return colonization to wild-type levels in the competitive UTI model, although perhaps aslight improvement in the mutant fitness is seen in the kidneys forboth double-deletion strains (Fig. 6B and C). Strains with singledeletions of csgD or bcsA competed like the wild-type strain in vivo(see Fig. S12B and C). Additional deletion of csgD from the �yfiR�ycgR double-deletion strain also did not relieve the attenuatedphenotype in the mouse model (see Fig. S12D). However, whenboth csgD and bcsA were deleted from the yfiR mutant, coloniza-tion was restored to wild-type levels (Fig. 6D). The motility of thedouble and triple mutants was determined using tryptone swimplates, on which they remained poorly motile like the yfiR singlemutant (see Fig. S11). All mutant strains were also tested on cal-cofluor plates, which showed that only deletion of bcsA from a yfiRdeletion strain eliminated cellulose production (see Fig. S13).
FIG 6 Deletion of both bcsA and csgD from the yfiR deletion mutant restores fitness to wild-type CFT073 levels in the mouse model of competitive UTI. Shownare results for coinfection of female Swiss Webster mice with CFT073 �yfiR �ycgR (n � 14) (A), CFT073 �yfiR �csgD (n � 11) (B), CFT073 �yfiR �bcsA (n �7) (C), or CFT073 �yfiR �csgD �bcsA (n � 24) (D) and CFT073 �lacZYA using the mouse model of UTI. The insets show the phenotypes of the mutants onCongo red plates grown at 30°C. The competitive indices were calculated by the following equation: (yfi mutant CFU recovered/wild-type CFU recovered)/(mutant inoculum CFU/wild-type inoculum CFU). The solid line indicates the median value.
c-di-GMP Level Control in Escherichia coli CFT073
September 2013 Volume 81 Number 9 iai.asm.org 3095
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Urine inhibits the rdar morphotype and csgD expression. Inaddition to growing the mutant strains on LB Congo red plates, weinoculated them on urine agar Congo red plates to see whetherexposure to urine caused a change in the rdar phenotype. All of thewild-type CFT073 colonies grown on the urine Congo red platesdisplayed a smooth and white phenotype, indicating that urinerepresses the expression or activity of the proteins needed for pro-duction of cellulose and curli fimbriae (see Fig. S14A in the sup-plemental material). Similar results were obtained when the wild-type strain was grown in liquid urine and then plated on LB Congored plates (see Fig. S14B), suggesting that the effect urine has onthe regulatory controls for curli fimbriae and cellulose are long-lasting. The change was not permanent, though, because a returnof the rdar phenotype was observed after growth in LB liquidculture followed by growth on LB Congo red plates (data notshown). Congo red plates that combined urine and LB were alsotested with the wild-type strain as a control to ensure that the lossof the rdar phenotype was not due to growth inhibition on the urine-only plates (see Fig. S14A). The presence of urine also affected thephenotype of the yfiR mutant on the urine Congo red plates, but allcolonies were still deep red and somewhat rugose (see Fig. S14A),which indicated that curli fimbriae and cellulose were both still beingproduced. Growth in liquid urine followed by growth on LB Congored plates produced no differences in the rdar phenotype for the yfiRmutant (see Fig. S14B). Salt (NaCl) is a known inhibitor of the rdarphenotype (29, 57). To determine which salts found in urine might beresponsible for the all-smooth-and-white phenotype seen in wild-type CFT073, individual salt components of urine [NaCl, Ca3(PO4)2,K2SO4, KCl, KHCO3, MgCO3, and MgSO4] at their physiologicalconcentrations were added to LB Congo red plates for testing with thewild-type strain. Of the salts tested, only NaCl produced a phenotypesimilar to that found on the urine Congo red plates (see Fig. S14A).
To determine whether urine and NaCl affected the expressionof csgD, the wild-type and yfiR mutant csgD::lacZYA strains wereemployed. �-Galactosidase assays performed on cells grown onurine plates showed a decrease in expression from the csg pro-moter in the presence of urine compared to LB, although thischange was not yfiR dependent. However, expression levels in theyfiR mutant remained elevated compared to those in the wild-typestrain under all conditions tested (see Fig. S14C in the supplemen-tal material). Growth on LB-NaCl plates caused no change in ex-pression compared to growth on LB-only plates (see Fig. S14C).
DISCUSSION
We set out to study the effect of altering the levels of the signalingmolecule c-di-GMP on the pathogenesis of the model uropatho-genic E. coli strain CFT073. We suggest in this report that loss ofperiplasmic YfiR relieves inhibition of the activity of the mem-brane-bound DGC YfiN, and this leads to an elevation in cellularc-di-GMP levels (Fig. 7). Loss of yfiR and the presumed increase inc-di-GMP levels then lead to an increase in production of bothcurli fimbriae and cellulose, a reduction in swimming motility,and, ultimately, attenuation in the mouse model of UTI. Spurbecket al. found that deletion of yfiN decreased adherence of CFT073to cultured human bladder epithelial cells (48). However, singledeletions of yfiL, yfiB, or yfiN in this study did not attenuate E. coliCFT073 in vivo. This discrepancy may be explained by the expres-sion of other fimbriae by CFT073 that may compensate for anyloss of curli-mediated binding in vivo. It is also possible that thehuman bladder cells used by Spurbeck et al. and the bladder epi-
thelium of the Swiss Webster mice express different receptors thatallow for binding by different sets of fimbriae; thus, fimbriae thatare important for establishing infection in the human bladder maydiffer from those necessary for the colonization of the mouse blad-der. Additional deletion of yfiN from the yfiR deletion strain orallelic restoration of the yfiR deletion strain brought urinary tractcolonization back to wild-type CFT073 levels. This indicates thatYfiR exerts an inhibitory effect on the activity of YfiN that is im-portant for maintaining control of c-di-GMP levels in vivo. With-out this control of c-di-GMP signaling, the bacteria become im-paired in their ability to colonize the urinary tract.
A study of the homologous P. aeruginosa system postulated thatYfiR prevents the dimerization of YfiN, a state that is necessary foractivity of most DGCs. The follow-up report further indicated thatYfiB sequesters YfiR, which would inhibit its ability to disrupt YfiNdimerization (47). Overall, though, the function of YfiB and themechanism by which the YfiRNB system operates are still largelyunknown. Further studies would be necessary to determine the hier-archy of the YfiRNB system and the set of external stimuli that mayactivate or repress YfiN activity. Our work also indicates that at leasttranscription of the yfi locus may be surface induced in CFT073. Ifattachment to the host epithelium also induces expression of thislocus, the regulation of both yfiLRNB transcription and YfiN activitycould give insights into the series of events necessary for successfulcolonization of and persistence in the urinary tract.
The downstream targets of increased c-di-GMP levels in E. coli
Curli Fimbriae
Cellulose
YcgR
YfiRYfiB
YfiN
CsgD
BcsA
AdrA
csgDEFG csgBA
yedQ
adrA
bcsABZC yhjQ yhjR bcsEFG
OM
OM
IM
IMYfiR
2 GTP
2 GTP
←
←
←
┬
←
←
←
←
←
←
Signal?
←
X
FlagellarMotor
c-di-GMP
c-di-GMP
FIG 7 Model of the putative YfiLRNB system and its downstream effects oncurli and cellulose production. YfiR is a predicted periplasmic inhibitor ofYfiN diguanylate cyclase activity. In the absence of YfiR, YfiN produces c-di-GMP, which, in turn, increases the production of curli fimbriae and celluloseand inhibits motility. CsgD, AdrA, BcsA, and YcgR are all predicted to interactdirectly or indirectly with c-di-GMP to produce the indicated phenotypes.
Raterman et al.
3096 iai.asm.org Infection and Immunity
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

CFT073 are similar to those that were previously described forother E. coli strains (22). The loss of yfiR resulted in decreasedmotility, increased expression of the csgD promoter, and increasedproduction of both curli fimbriae and cellulose. Expression ofyhjH, which encodes a phosphodiestrase, in the CFT073 �yfiRbackground returned these phenotypes to the wild type. Thephosphodiesterase activity of YhjH was required for this reversal,suggesting that an increased c-di-GMP level is indeed the factordriving the increased expression of curli fimbriae and celluloseand the observed decrease in motility in the yfiR deletion strain.The motility defect of the yfiR deletion strain was partially rescuedby additional deletion of ycgR. YcgR is capable of interacting withthe flagellar motor to either promote smooth swimming or act asa braking mechanism when it is bound to c-di-GMP (55, 56).Deletion of ycgR therefore prevented alteration of flagellar rota-tion by this mechanism. Although the additional deletion of ycgRpartially relieved the attenuation of the motility phenotype, theCFT073 �yfiR �ycgR strain was still attenuated in the urinarytract. This observation indicates that the motility defect did notcontribute to the in vivo attenuation. However, motility may bejust one factor involved in the attenuation of the yfiR deletionmutant, and restoration of motility alone may not have beenenough to rescue colonization to wild-type levels in vivo.
Included in these possible contributing factors, the elevatedproduction of curli fimbriae and cellulose also appears to have adetrimental effect on the colonization ability of CFT073. The linkbetween the expression of csgD and the activity of the cellulosebiosynthetic machinery was established previously in E. coli K-12strains. CsgD not only activates expression of the curli operon butalso activates expression of adrA and yedQ, genes that encodeDGCs. The c-di-GMP produced by either AdrA or YedQ wouldthen stimulate increased activity of the cellulose synthetic machin-ery by binding to components of its core catalytic module (29, 39,50). Deletion of adrA in CFT073, but not yedQ, resulted in a loss ofcellulose production on LB Congo red plates. Therefore, we pos-tulate that AdrA expression stimulated by CsgD activity is thelikely link involved in CsgD-dependent increases of cellulose pro-duction in E. coli CFT073.
Curli fibers are known activators of the host coagulase systemthat leads to increased influx of immune cells and inflammation(34). Another report also implicated the expression of curli fim-briae in the absence of cellulose production as an immune stimu-lant that results in increased clearance of uropathogenic E. colistrain UTI89 from the urinary tract (36). In contrast to these re-ports, a CFT073 �yfiR �csgD mutant was still attenuated in thecompetitive mouse model of UTI even though it was confirmedthat curli fimbriae were no longer being produced. Only addi-tional deletion of bcsA from the �yfiR �csgD mutant rescued theattenuated phenotype. Although the yfiR single-deletion mutantdid not display any growth defects in vitro in various media whengrown singly or in competition with the wild-type strain, it ispossible that the increased stress of the urinary tract environmentresulted in the attenuation of the metabolically overburdened mu-tant. The significant amount of cellular resources being consumedfor the production of large and unnecessary amounts of curli fim-briae and cellulose may divert resources away from the produc-tion of other virulence or colonization factors vital for infection ofthe urinary tract. Further studies into alterations in the globalproduction of bacterial proteins in the yfiR deletion mutant could
give insight into whether the expression of any other virulencefactors was altered.
Given that single deletions of csgD or bcsA did not attenuateCFT073 in vivo, biofilm formation associated with curli or celluloseis likely not a persistence factor involved in colonization in the urinarytract. Additionally, growth in urine decreases expression of csgD andinhibits the rdar morphology. The salt content of urine appears to bethe main suppressor leading to this outcome. Growth at 37°C alsoinhibits the production of these biofilm components (data notshown). Although curli and cellulose may be produced during UTIs,these data suggest that robust curli and cellulose-dependent biofilmsare not formed by E. coli CFT073 in our mouse model of UTI andmay instead be more important for adaptation and persistence inenvironments outside the urinary tract.
This study not only emphasizes the importance of regulation ofcurli fimbriae, cellulose production, and motility in the virulenceof UPEC in the urinary tract but also highlights c-di-GMP pro-duction as a key regulator of these products. Although the bacteriaseem to survive well in the short term in the absence of curlifimbriae and cellulose, these molecules could still play a role in thepersistence of the bacteria in the urinary tract during chronic in-fections. Given the abundance of DGCs (�19) and PDEs (�17) inE. coli, c-di-GMP most likely also plays an important role in thesurvival of the bacteria in vivo. Further studies into the distinct orperhaps redundant roles played by the various DGCs and PDEsshould illuminate the steps in the fine-tuned control that the bac-teria have over the expression of their colonization and virulencefactors to mount successful infections in the host.
ACKNOWLEDGMENTS
This research was funded by the National Institutes of Health grant R01DK063250-06 and by the National Institutes of Health, National ResearchService Award AI55397.
REFERENCES1. Bacheller CD, Bernstein JM. 1997. Urinary tract infections. Med. Clin.
North Am. 81:719 –730.2. Faro S, Fenner DE. 1998. Urinary tract infections. Clin. Obstet. Gynecol.
41:744 –754.3. Hooton TM. 2000. Pathogenesis of urinary tract infections: an update. J.
Antimicrob. Chemother. 46(Suppl 1):1–7; discussion, 63– 65.4. Kunin CM. 1994 Urinary tract infections in females. Clin. Infect. Dis.
18:1–10; quiz, 11–12.5. Orenstein R, Wong ES. 1999. Urinary tract infections in adults. Am. Fam.
Physician 59:1225–1234, 1237.6. Bahrani-Mougeot FK, Buckles EL, Lockatell CV, Hebel JR, Johnson DE,
Tang CM, Donnenberg MS. 2002. Type 1 fimbriae and extracellularpolysaccharides are preeminent uropathogenic Escherichia coli virulencedeterminants in the murine urinary tract. Mol. Microbiol. 45:1079 –1093.
7. Donnenberg MS, Welch RA. 1996. Virulence determinants of uropatho-genic Escherichia coli, p 135–174. In Mobley HLT, Warren JW (ed), Uri-nary tract infections: molecular pathogenesis and clinical management.ASM Press, Washington, DC.
8. Garcia EC, Brumbaugh AR, Mobley HL. 2011. Redundancy and speci-ficity of Escherichia coli iron acquisition systems during urinary tract in-fection. Infect. Immun. 79:1225–1235.
9. Kau AL, Hunstad DA, Hultgren SJ. 2005. Interaction of uropathogenicEscherichia coli with host uroepithelium. Curr. Opin. Microbiol. 8:54 –59.
10. Nielubowicz GR, Mobley HL. 2010. Host-pathogen interactions in uri-nary tract infection. Nat. Rev. Urol. 7:430 – 441.
11. Smith YC, Rasmussen SB, Grande KK, Conran RM, O’Brien AD. 2008.Hemolysin of uropathogenic Escherichia coli evokes extensive shedding of theuroepithelium and hemorrhage in bladder tissue within the first 24 hours afterintraurethral inoculation of mice. Infect. Immun. 76:2978–2990.
12. Wiles TJ, Kulesus RR, Mulvey MA. 2008. Origins and virulence mech-anisms of uropathogenic Escherichia coli. Exp. Mol. Pathol. 85:11–19.
c-di-GMP Level Control in Escherichia coli CFT073
September 2013 Volume 81 Number 9 iai.asm.org 3097
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

13. Wright KJ, Seed PC, Hultgren SJ. 2005. Uropathogenic Escherichia coliflagella aid in efficient urinary tract colonization. Infect. Immun. 73:7657–7668.
14. Yamamoto S, Nakata K, Yuri K, Katae H, Terai A, Kurazono H, TakedaY, Yoshida O. 1996. Assessment of the significance of virulence factors ofuropathogenic Escherichia coli in experimental urinary tract infection inmice. Microbiol. Immunol. 40:607– 610.
15. Simm R, Morr M, Kader A, Nimtz M, Romling U. 2004. GGDEF andEAL domains inversely regulate cyclic di-GMP levels and transition fromsessility to motility. Mol. Microbiol. 53:1123–1134.
16. Amikam D, Galperin MY. 2006. PilZ domain is part of the bacterialc-di-GMP binding protein. Bioinformatics 22:3– 6.
17. D’Argenio DA, Miller SI. 2004. Cyclic di-GMP as a bacterial secondmessenger. Microbiology 150:2497–2502.
18. Hengge R. 2009. Principles of c-di-GMP signalling in bacteria. Nat. Rev.Microbiol. 7:263–273.
19. Jenal U, Malone J. 2006. Mechanisms of cyclic-di-GMP signaling inbacteria. Annu. Rev. Genet. 40:385– 407.
20. Kader A, Simm R, Gerstel U, Morr M, Romling U. 2006. Hierarchicalinvolvement of various GGDEF domain proteins in rdar morphotype de-velopment of Salmonella enterica serovar Typhimurium. Mol. Microbiol.60:602– 616.
21. Mills E, Pultz IS, Kulasekara HD, Miller SI. 2011. The bacterial secondmessenger c-di-GMP: mechanisms of signalling. Cell. Microbiol. 13:1122–1129.
22. Povolotsky TL, Hengge R. 2012. ‘Life-style’ control networks in Esche-richia coli: signaling by the second messenger c-di-GMP. J. Biotechnol.160:10 –16.
23. Römling U, Gomelsky M, Galperin MY. 2005. c-di-GMP: the dawning ofa novel bacterial signalling system. Mol. Microbiol. 57:629 – 639.
24. Ryjenkov DA, Simm R, Romling U, Gomelsky M. 2006. The PilZdomain is a receptor for the second messenger c-di-GMP. The PilZ do-main protein controls motility in enterobacteria. J. Biol. Chem. 281:30310 –30314.
25. Ryjenkov DA, Tarutina M, Moskvin OV, Gomelsky M. 2005. Cyclicdiguanylate is a ubiquitous signaling molecule in bacteria: insights intobiochemistry of the GGDEF protein domain. J. Bacteriol. 187:1792–1798.
26. Wolfe AJ, Visick KL. 2008. Get the message out: cyclic-Di-GMP regulatesmultiple levels of flagellum-based motility. J. Bacteriol. 190:463– 475.
27. Yan H, Chen W. 2010. 3=,5=-Cyclic diguanylic acid: a small nucleotidethat makes big impacts. Chem. Soc. Rev. 39:2914 –2924.
28. Bokranz W, Wang X, Tschape H, Romling U. 2005. Expression ofcellulose and curli fimbriae by Escherichia coli isolated from the gastroin-testinal tract. J. Med. Microbiol. 54:1171–1182.
29. Römling U. 2005. Characterization of the rdar morphotype, a multicel-lular behaviour in Enterobacteriaceae. Cell. Mol. Life Sci. 62:1234 –1246.
30. Hammar M, Arnqvist A, Bian Z, Olsen A, Normark S. 1995. Expressionof two csg operons is required for production of fibronectin- and congored-binding curli polymers in Escherichia coli K-12. Mol. Microbiol. 18:661– 670.
31. Brombacher E, Baratto A, Dorel C, Landini P. 2006. Gene expressionregulation by the curli activator CsgD protein: modulation of cellulosebiosynthesis and control of negative determinants for microbial adhesion.J. Bacteriol. 188:2027–2037.
32. Römling U, Bian Z, Hammar M, Sierralta WD, Normark S. 1998. Curlifibers are highly conserved between Salmonella typhimurium and Esche-richia coli with respect to operon structure and regulation. J. Bacteriol.180:722–731.
33. Olsén A, Jonsson A, Normark S. 1989. Fibronectin binding mediated bya novel class of surface organelles on Escherichia coli. Nature 338:652– 655.
34. Persson K, Russell W, Morgelin M, Herwald H. 2003. The conversion offibrinogen to fibrin at the surface of curliated Escherichia coli bacteria leadsto the generation of proinflammatory fibrinopeptides. J. Biol. Chem. 278:31884 –31890.
35. Tükel C, Nishimori JH, Wilson RP, Winter MG, Keestra AM, vanPutten JP, Bäumler AJ.. 2010. Toll-like receptors 1 and 2 cooperativelymediate immune responses to curli, a common amyloid from enterobac-terial biofilms. Cell. Microbiol. 12:1495–1505.
36. Kai-Larsen Y, Luthje P, Chromek M, Peters V, Wang X, Holm A, KadasL, Hedlund KO, Johansson J, Chapman MR, Jacobson SH, Romling U,Agerberth B, Brauner A. 2010. Uropathogenic Escherichia coli modulatesimmune responses and its curli fimbriae interact with the antimicrobial
peptide LL-37. PLoS Pathog. 6:e1001010. doi:10.1371/journal.ppat.1001010.
37. Römling U. 2002. Molecular biology of cellulose production in bacteria.Res. Microbiol. 153:205–212.
38. Ross P, Mayer R, Benziman M. 1991. Cellulose biosynthesis and functionin bacteria. Microbiol. Rev. 55:35–58.
39. Zogaj X, Nimtz M, Rohde M, Bokranz W, Romling U. 2001. Themulticellular morphotypes of Salmonella typhimurium and Escherichia coliproduce cellulose as the second component of the extracellular matrix.Mol. Microbiol. 39:1452–1463.
40. Girgis HS, Liu Y, Ryu WS, Tavazoie S. 2007. A comprehensive geneticcharacterization of bacterial motility. PLoS Genet. 3:e154. doi:10.1371/journal.pgen.0030154.
41. Datsenko KA, Wanner BL. 2000. One-step inactivation of chromosomalgenes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci.U. S. A. 97:6640 – 6645.
42. Battaglioli EJ, Baisa G, Weeks AE, Schroll R, Hryckowian AJ, WelchRA. 2011. Isolation of generalized transducing bacteriophages for uro-pathogenic strains of Escherichia coli. Appl. Environ. Microbiol. 77:6630 –6635.
43. Bäumler AJ, Tsolis RM, van der Velden AWM, Stojiljkovic I, Anic S,Heffron F. 1996. Identification of a new iron regulated locus of Salmonellatyphi. Gene 183:207–213.
44. Redford P, Welch RA. 2006. Role of sigma E-regulated genes in Esche-richia coli uropathogenesis. Infect. Immun. 74:4030 – 4038.
45. Ma Q, Wood TK. 2009. OmpA influences Escherichia coli biofilm forma-tion by repressing cellulose production through the CpxRA two-component system. Environ. Microbiol. 11:2735–2746.
46. Malone JG, Jaeger T, Spangler C, Ritz D, Spang A, Arrieumerlou C,Kaever V, Landmann R, Jenal U. 2010. YfiBNR mediates cyclic di-GMPdependent small colony variant formation and persistence in Pseudomo-nas aeruginosa. PLoS Pathog. 6:e1000804. doi:10.1371/journal.ppat.1000804.
47. Malone JG, Jaeger T, Manfredi P, Dötsch A, Blanka A, Bos R, CornelisGR, Häussler S, Jenal U. 2012. The YfiBNR signal transduction mecha-nism reveals novel targets for the evolution of persistent Pseudomonasaeruginosa in cystic fibrosis airways. PLoS Pathog. 8:e1002760. doi:10.1371/journal.ppat.1002760.
48. Spurbeck RR, Tarrien RJ, Mobley HL. 2012. Enzymatically active andinactive phosphodiesterases and diguanylate cyclases are involved in reg-ulation of motility or sessility in Escherichia coli CFT073. mBio 3:e00307–12. doi:10.1128/mBio.00307-12.
49. Pesavento C, Becker G, Sommerfeldt N, Possling A, Tschowri N, MehlisA, Hengge R. 2008. Inverse regulatory coordination of motility and curli-mediated adhesion in Escherichia coli. Genes Dev. 22:2434 –2446.
50. Monteiro C, Saxena I, Wang X, Kader A, Bokranz W, Simm R, NoblesD, Chromek M, Brauner A, Brown RM, Jr, Romling U. 2009. Charac-terization of cellulose production in Escherichia coli Nissle 1917 and itsbiological consequences. Environ. Microbiol. 11:1105–1116.
51. Méndez-Ortiz MM, Hyodo M, Hayakawa Y, Membrillo-Hernandez J.2006. Genome-wide transcriptional profile of Escherichia coli in responseto high levels of the second messenger 3=,5=-cyclic diguanylic acid. J. Biol.Chem. 281:8090 – 8099.
52. Lacey MM, Partridge JD, Green J. 2010. Escherichia coli K-12 YfgF is ananaerobic cyclic di-GMP phosphodiesterase with roles in cell surface re-modelling and the oxidative stress response. Microbiology 156:2873–2886.
53. Lane MC, Alteri CJ, Smith SN, Mobley HL. 2007. Expression of flagellais coincident with uropathogenic Escherichia coli ascension to the upperurinary tract. Proc. Natl. Acad. Sci. U. S. A. 104:16669 –16674.
54. Schwan WR. 2008. Flagella allow uropathogenic Escherichia coli ascen-sion into murine kidneys. Int. J. Med. Microbiol. 298:441– 447.
55. Boehm A, Kaiser M, Li H, Spangler C, Kasper CA, Ackermann M,Kaever V, Sourjik V, Roth V, Jenal U. 2010. Second messenger-mediatedadjustment of bacterial swimming velocity. Cell 141:107–116.
56. Paul K, Nieto V, Carlquist WC, Blair DF, Harshey RM. 2010. Thec-di-GMP binding protein YcgR controls flagellar motor direction andspeed to affect chemotaxis by a “backstop brake” mechanism. Mol. Cell38:128 –139.
57. Prigent-Combaret C, Brombacher E, Vidal O, Ambert A, Lejeune P,Landini P, Dorel C. 2001. Complex regulatory network controls initialadhesion and biofilm formation in Escherichia coli via regulation of thecsgD gene. J. Bacteriol. 183:7213–7223.
Raterman et al.
3098 iai.asm.org Infection and Immunity
on February 15, 2019 by guest
http://iai.asm.org/
Dow
nloaded from





























