Embed Size (px)
Citation preview

Genetic and Environmental Influences on
Personality: A Study of Twins Reared
Together Using the Self- and Peer Report
NEO-FFI Scales
Rainer Rlemann Alois AngleitnerUniversity of Bieleield, Germany
Jan StrelauUniversity of Warsaw, Poland
ABSTRACT Previous behavior-genetic research on personality has beenalniosi exclusively based tin self-report questionnaire measures. The purposeof this research was to measure personahty constructs via self- and peerreports on tbe items of tbe NEO Five-Factor Inventory (Costa & McCrae.1989). The sample included 660 monozygotic and 200 same sex and 104opposite sex dizygoiic twin pairs. We collected self- and two independent peerreports for each of the twins. Our analysis of self-report data replicates earlierfmdings of a substantial genetic influence on the Big Five (/r = .42 to .56).We also found this influence for peer reports. Our results validate tindingsbased solely on self-reports. However, estimates of genetic contributions tophenotypic variance were substantially higher when based on peer reports (/i' =.57 to .81) or self- and peer reports (ir = .66 to .79) because these data allowed
Preliminarj' analyses of the data were presented at the seventh meeting of the Interna-tional ScKiety for the Study of Individual Differences, Warsaw, Poland. July 15-19,1995, and in Angleitner, Riemann, Spinath. Hempel. Thiel. and Strelau (1995). Thisresearch was supported in part hy funds from the Max Planck Forschungspreis(awarded by the Alexaiider von Huinboldt Stiftung and the M;ix-Planck Gesellschaft)10 Alois Angleitner and Jan Strelau. We thank Peter BorkLMiau. Rainer Reisen/ein. andFrank M. Spinath for helpful comments, and Wolfgang Thiel for his assistance in ana-lyzing the data. CorrespKmdence conceming this article should he addressed to RainerRiemann. Department of Psychology. University of Bielefeld. Postfach 100131, 33501Bielefeld, Germany. E-mail may he sent to [email protected].
Joumal of PersonaUty 65:3, September 1997.Copyright © 1997 by Duke University Press.

Riemann et al.
us to separate error variance from variance due to nonshared environmental in-fluences. Correlations between self- and peer reports reflected the same geneticinfluences to a much higher extent than identical environmental effects.
Humati behavior-genelic research has provided important itisights intothe etiology of individual differences in personality traits (Loehlin,1992; Plotnin & McClearn, 1993; Rose, 1995; Rowe, 1994). This re-search has focused most strongly on Extraversion and Neuroticism asthe two major dimensions of personality. In recent years, however, thefive-factor model has gained a prominent position within personalityresearch (for reviews see Digman, 1990; John, 1990; John, Angleitner,& Ostendorf, 1988: McCrae, 1989). Ntimerous studies have shownthat five broad factors—Extraversion or Surgency, Agreeableness,Conscientiousness, Emotional Stability or Neuroticism, and Culture/Intellect or Openness to Experience—account for much of the com-mon variance in personality rating and questionnaire data. They aremeaningful dimensions for the description of individual differences inpersonality and robustly found in various sources of data. The NEOPersonality Inventory-Revised (NEO-PI-R; Costa & McCrae, 1992)and the short version of the NEO-PI, the NEO Eive-Factor Inventory(NEO-FFI; Costa & McCrae, 1989) are now standard questionnairemeasures of the five-factor model.
Four major research designs that are used in human behavior-genetic studies can be distinguished: (a) studies of twins reared to-gether in the same family, (h) studies of twins who have been reared indifferent families, (r) studies of adoptive families (ideally includingmeasures of the adoptees' biological parents), and (d) family studiesanalyzing the data of participants who are genetically related to varyingdegrees. All of these designs have their particular strengths and weak-nesses, and firm conclusions regarding the etiology of individual dif-ferences are based on an integrative analysis of data based on all fourdesigns (e.g., Bouchard & McGue, 1981; Loehlin, 1992). The mostpopular design, the comparison of monozygotic (MZ) and dizygotic(DZ) twins reared together, requires several assumptions to estimategenetic and environmental effects on traits: (a) There are only twotypes of twins (MZ and DZ) and DZ twins share on average 50% oftheir genes (i.e., there are no effects of assortative mating),' (b) twins
1. There is some evidence of assortative mating for Openness and Conscientiousness(see McCrae, 1996). A.ssortative mating increases the correlation of additive and

TWin Study of NEO-FFI Scales 451
are a representative sample of the population to which one wishes togeneralize, (c) the effects of environmental influences shared by twins isno greater for MZ than for DZ twins ("equal environments" assump-tion), and (d) gene-environment correlation and interaction have a neg-ligible effect on the trait under study. Finally, studies of twins rearedtogether do not allow simultaneous estimation of environmental eflectsshared by the twins and nonadditive genetic influences (e.g., due togenetic dominance). The validity of these assumptions is thoroughly dis-cussed in the behavior-genetic literature (see, e.g.. Eaves, Eysenck, &Martin. 1989; Loehlin, 1992). From this discussion it can be concludedthat studies of twins reared together provide a valuable tool for behavior-genetic research. The strength of this design is that large samples areavailable representing the whole range of genotypes and environments.
Combining data from twin, adoption, and family studies into anextremely large data set, Loehlin (1992) thoroughly reviewed the thenavailable behavior-genetic research on the five-factor model of person-ality. All of the studies summarized by Loehlin used self-report question-naires that, with one exception (Bergeman et al., 1993), had not beenconstructed with the goal of measuring the Big Five. Instead, Extra-version and Neuroticism were frequently measured by the EysenckPersonality Inventory (EPI; Eysenck & Eysenck, 1964), the EysenckPersonality Questionnaire (EPQ; Eysenck & Eysenck, 1975), the EPQ-Revised (Eysenck & Eysenck, 1991), or short versions of one of thesequestionnaires. Measures related to Agreeableness comprised scalesdeveloped to measure masculinity versus femininity, aggression, altru-ism, empathy, and related constructs. Conscientiousness was indexedby measures of, for example, socialization, task orientation, and con-formity. Finally, Culture/Openness was assessed, among others, byscales for flexibility, artistic interests, science interests, and achieve-ment via independence.
Two behavior-genetic models could reasonably be fitted to this exten-sive data collection. One of these models involved effects of additivegenes (A), genetic epistasis (I), shared twin or sibling environments (C),
nonadditive genetic effects for DZ twitis. Thus, the failure to take assortative matinginto account results in an overestimation of effects due to etivironmental influencesshiired hy sihiings and an underestimation of nonadditive genetic effects. Because weneither have appropriate estimates of assortative mating for otjr sample nor a thoroughtinderstanding of its etiology (see Nealc & Cardon, 1992). we did not correct the cor-relations of genetic effects for DZ twins in our models.

Kiemannetal.
and a combination of specific (not shared by siblings) environmental,genotype-environment interaction, and measurement error (E). Theother model included effects of A, C, and E, and of a special MZ twinenvironment (contributing to the similarity of MZ twins rearedtogether). Given the evidence on the "equal environments" assumption,we focus on the data for the first model here. Several conclusions canbe drawn from Loehlin's (1992) meta-analysis. For all five factors,additive genetic effects were the strongest source of the phenotypicvariance, explaining 43% of the variance in Culture/Openness, 32% ofExtraversion, 27% of Neuroticism, 24% of Agreeableness, and 22% ofConscientiousness. Conversely, shared sibling environment effectscontributed little to phenotypic variance. They were negligible forExtraversion (2%) and small for Culture/Openness (6%), Conscien-tiousness (7%). Neuroticism (7%), and Agreeableness (11%). Nonaddi-tive genetic effects had a noticeable influence on all dimensions (11%to 17%) except Culture/Openness (2%).
Highly similar results are reported by Bouchard (1994) for the Min-nesota study of twins reared apart and twins reared together usingscales or factors of the Multidimensional Personality Questionnaire(MPQ; Tellegen et al., 1988) to measure the five factors, although forthese data a model involving A and E effects provide the most appro-priate description of the data. Earlier studies of twins reared together,however, yielded heritability estimates that were somewhat higher andindicated less influence of nonadditive genetic factors.
Two studies of twins reared together (Beer & Rose, 1995; Jang.Livesley, & Vemon, 1996) that used direct measures of the five-factormodel, the NEO-PI and the NEO-FFI (Costa & McCrae, 1985)yielded results similar to those reported by Loehlin (1992). The esti-mates of genetic effects on the phenotypic variance of the NEO-PIscales reported by Jang et al. (1996) were 61% for Openness, 53% forExtraversion, 44% for Conscientiousness, 41% for Neuroticism, and41% for Agreeableness. However, the results of Bergeman et al.'s(1993) study of twins reared together and twins reared apart weremarkedly different. Bergeman et al. used a Swedish translation ofshort preliminary NEO-PI scales for Openness (25 items). Conscien-tiousness (10 items), and Agreeableness (10 items). They found smallgenetic effects for Conscientiousness (29%) and Agreeabieness (12%).Shared environmental influences were substantial for Agreeableness(21%) and noticeable for Conscientiousness (11%). The resuhs forOpenness were in line with previous research. For Agreeableness and

Twin Study of NEO-FFI Scales 4S3
Conscientiousness, twins reared apart were substantially less similarthan twins reared together. This results, however, was not replicated inBouchard's (1994) analysis and may, as the authors concede, be attrib-uted to psychometric limitations of the measures or to peculiarities ofthe Swedish sample.
In general, the data summarized here are in good agreement withmany behavior-genetic studies of personality traits. They indicate sub-stantial heritability of all dimensions of the five-factor model. Geneticfactors explain about 40% to 50% of the variance in self-reported per-sonality traits; environmental influences shared by siblings or twinsare absent or marginal (Bouchard, 1994). Nonadditive genetic effectsor, alternatively, a special MZ environment resemblance are importantfor Neuroticism, Extraversion, and Conscientiousness.
These conclusions for adult personality measures rely almost exclu-sively on self-report measures, which are used because they allowresearchers to collect reliable and valid personality measures for sev-eral hundred or even several thousand participants. Since its begin-ning, however, the systematic study of personality has used multiplemethods (e.g., Allport, 1937; Cattell, 1950; Fiske, 1971; Heymans &Wiersma, 1906), primarily because they allow examination of method-specific effects. This is important for personality constructs like traits,which summarize divergent modes of responding. In addition, byincluding multiple sources of data in behavior-genetic research, theinfluence of environmental effects that are not shared by twins can beseparated from effects due to measurement error.
Although it may be argued that people generally know their ownbehaviors, thoughts, and feelings better than observers, there are reasonsto believe that self-reports of twins reared together are a problematicsource of data. Specifically, several authors (e.g., Borkenau, 1993; Carey,1986; Heath, Neale, Kessler, Eaves, & Kendler, 1992; Loehlin, 1986;Rose, 1995; Saudino & Eaton, 1991) have suggested that self-reportsmay be distorted by "contrast effects," which result in an underestimationof the similarity between DZ twins and may obscure real shared environ-mental influences or inflate the estimates of nonadditive genetic influences.
We can conceive of several processes that would result in a distor-tion of self-reports provided by twins reared together. For example,real behavioral differences may be emphasized by DZ twins becausethey compare their own behavior to that of their co-twin instead of thepopulation average. Alternatively, MZ twins may discount behavioraldifferences because of their generally high similarity.

454 Riemann et al.
A second shortcoming of self-report data in behavior-genetic researchis that no personality judgment directly reflects behavioral differences(except the answering of questionnaire items). To varying extentsresponses lo questionnaire items or rating scales require inferencesand interpretations. Thus, it remains unclear to what extent heritabilitycoefficients and estimates of environmental influences reflect contentor stylistic (response-set) variance. This argument is not limited tostudies of twins reared together.
In the present study of twins reared together, we use peer reports inaddition to self-reports on the items of the NEO-FFI to measure theBig Five. Peer reports are an established source of data in personalityresearch (Funder, 1987; Kenny, 1994). Depending on the number ofpeers in the study and additional moderator variables (Chaplin, 1991;Paunonen, 1989) they correlate consistently but far from perfectlyamong each other and with self-report data. Several factors have beenidentified that mediate consensus among peers (Kenny. 1991). Assimi-lation effects in peer report data can be reduced to the degree thatpeers know one of the twins very well but have little or no contactwith the other. More important, peers judging only one of a pair of DZtwins should be less susceptible to contrast effects.
Our study has five goals. First, we will estimate environmental andgenetic influences using an established measure of the five-factor model.Estimates will be compared between the methods. If the estimates forself-report data deviate consistently from those for peer report data, thiswould be an indicator of contrast or assimilation effects. Second, peerreport data provided by more than one peer allow the separation ofeffects due to environmental influences on personality traits that are notshared by twins from measurement error. Third, we will provide heri-tability estimates for personality constructs that are measured by morethan one method. Fourth, and closely related to the third point, we willestimate genetic and environmental correlation.s between self- and peerreport measures, which are important if we are to understand thegenetic and environmental basis of the correlation between self- andpeer reports. For example, even if we find high heritability estimatesfor both sources of data, they may reflect different genetic effects.Thus, a joined analysis of self- and peer reports may obscure the under-lying processes. If this is the case, more detailed analyses should becarried out for both measures separately. On the other hand, the identi-fication of the components that make up the correlation between self-and peer reports provides information on the basis of this agreement.

Twin Study ol NEO-FFI Scales
Finally, we will test for age differences in estimates of genetic andenvironmental influences on personality measures (differential heri-tability). Since our study has a cross-sectional design, age effects areconfounded with cohort effects. Recent meta-analyses of differentialheritability (McCartney, Harris, & Bemieri. 1990; Pedersen, 1993)found that heritability is somewhat lower in older age groups than inyounger groups, but due to methodological problems and inconsistentfindings, the size of this effect is still undetermined. Bergeman et al.(1993) found no significant age effects for Openness, Agreeableness,and Conscientiousness.
METHOD
Participants
Participants were 509 female and 151 male MZ twin pairs and 149 female. 51male, and 104 opposite sex twin pairs diagnosed as DZ. All twin pairs volun-teered for this study. Because there is no population-based twin register inGermany, al! participants were approached through announcements in news-papers, magazines, radio and TV stations, or through twin elubs and twinmeetings. Participants were between 14 and 80 years old (M = 32.98, SD =13.40 years). There were no marked differences between MZ and DZ twinswith respect to age, education, or occupational status.
Zygosity Dete rmina t ion
Zygosity of twins was determined using physical similarity criteria. Partici-pants completed a zygosity questionnaire (Oniszczenko. Angleitner, Strelau,& Angeri. 1993) in which they were asked to describe and compare them-selves with their co-twin on a numher of physical characteristics (e.g., height,hair color, texture of hair, eye color) and to judge the extent of twin confusionby parents, relatives, peers, and .strangers. From a subsample of 110 (84 MZ,38 DZ) same sex twin pairs, blood samples were ohtained during a separatemedical examination. For these participants, zygcsity was determined by theanalysis of single-gene markers with a probability of less than .O(X)I to falselyclassify a DZ pair as MZ. Discriminant function scores and classificationfunctions were calculated in this subsample to predict the zygosity of theremaining 750 same sex twin pairs. Among the 15 variables entered in theanalysis, extent of confusion by strangers, differences in eye color, height, haircolor, and suffering from illness yielded the highest standardized discriminantfunction coefficients. The predicted zygosity was correct 93.64% of the timein the subsample, a result which is in the range of hit rates typically achieved

456 Riemann et al.
hy zygosity questionnaires (Lykken, 1978; Nichols & Bilbro, 1966; Plomin.DeFries, & McCleam, 1990). Misclassifications occurred more frequently forDZ twins than for MZ twins: Six DZ twins were classified as MZ, while onlyone MZ twin was classified as DZ.
Materials
A number of temperament and personality questionnaires and the inventoryfor zygosity diagnosis were mailed to participants. Among these measureswas the German version of the NEO-FFI (Costa & McCrae, 1989; Borkenau& Ostendorf, 1993), an established measure of the five-factor model of per-sonality. The peer report version of the NEO-FFI was identical to the self-report version, except that the first-person form had been changed to thethird-person form.
Procedure
Participants were instructed to complete the questionnaires independently. Inaddition, each twin gave the peer report versions of the questionnaires to twopeers who knew one twin hut (preferably) not the co-twin very well. Partici-pants returned self- and peer reports by mail; the latter were sealed hy thepeers in separate envelopes.
Peers were mostly friends (62%). relatives (16%), spouses (10%), and col-leagues (9%) who had known the participants for 11.06 {SD = 10.46) years onaverage and who, on the whole, judged their acquaintance with the target per-son as "very good" or "good" (82%). Very few (\%) indicated that they hadlittle or very little knowledge about the target. The majority of peers werefemale (62%).
Analyses
Personality scores on the NEO-FFI change significantly with age and are cor-related with the respondents' sex (Borkenau & Ostendorf, 1993). Because theages of twin pairs were obviously perfectly correlated, age was expected tocontribute to the correlation between twins. Covariance between twins due tomaturation may thus he misrepresented as caused by environmental influ-ences shared by twins (Eaves et ai,, 1989; McGue & Bouchard. 1984). In thesame manner sex differences resulted in spuriously higher correlationsbecause most of our participants were same sex twins.
To control for artificial sex and linear age effects on the correlationsamong our MZ and DZ twin samples, age- and sex-corrected scores werecomputed. We regressed the twins' NEO-FFI scores on their age and sex andcomputed standardized residuals (i.e., the standardized difference between

Twin Study of NEO-FFI Scales 457
the scores predicted from knowing participants' age and sex and their actualscore). This was done separately for first- and second-bom twins.
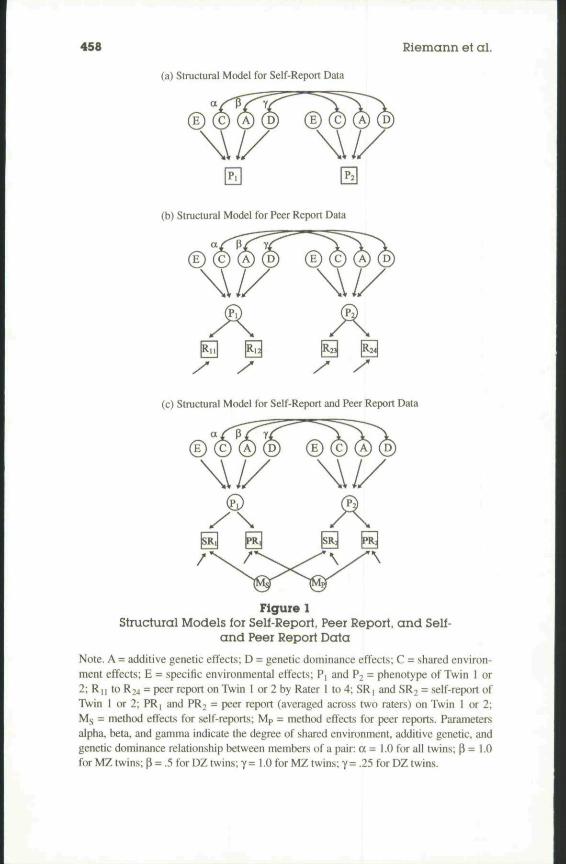
We used maximum-likelihood model fitting to estimate the influence ofgenetic and environmental sources of variance on phenotypic individual dif-ferences. Goodness-of-fit indexes and parameter estimates were obtainedusing LISREL 8.03 (Joreskog & Sorbom. 1993). These analyses were limited toage- and sex-corrected scores of same sex twin pairs using variance-covariancematrices. Due to the small number of male DZ twins, more detailed analyses ofgenetic and environmental effects on male and female phenotypes would addlittle, if any, additional information. We considered four sources of variance inour analyses: (a) additive genetic effects at multiple loci (A), (b) genetic domi-nance effects at multiple loci (D), (c) environmental effects shared by twins(C), and (d) nonshared environmental influences (E). When the analyseswere based exclusively on self-report data (Model I). E confounded variancedue to nonshared environmental effects and error variance. The availability oftwo peer reports per twin (Model 2) and of self- and peer reports (Model 3)allowed us to separate error variance from nonshared environmental variance.The models fitted to our data are depicted in Figure I (a-c).
Model 1 is a standard model for the analysis of twin data. The twins' phe-notypic personality traits were measured by a single indicator (i.e., NEO-mscale). We tixed the paths from phenotypic trait expressions (Pi and P2) totheir respective indicators to unity. Iti Model 2 there were two indicators ofeach phenotypic trait, and the paths from traits to their measures and the mea-sures' error variances were estimated. Model 3 was the rater bias modeldescribed by Neale and Cardon (1992). It differed from Model 2 insofar astwo method factors were introduced. Paths from the method factors to themeasures were estimated for each method (self- vs. peer report) separately.They were constrained to equality within methods and between MZ and DZtwin groups. Although self- and peer reports were provided by differentjudges for each twin, we decided to include method factors in the modelbecause the similarity among self- and among peer ratings, even if they stemfrom different persons, may be greater than the similarity between self- andpeer ratings. For example, self-raters who were reared together and sharedgenetic influences may have employed more similar meaning systems thanpeer raters, who showed no family resemblance. In tum, peer ratings mayhave reflected shared stereotypes on the relation between physical and psy-chological characteristics or rare but striking behaviors of twins to a higherdegree than self-report measures.
In a study of twins reared together, D and C effects cannot be estimatedsimultaneously. Following the procedure described in detail hy Neale andCardon (1992) we tested the following hypotheses: (a) The data do not indi-cate any family resemblance (E model), (b) family resemblance is caused byadditive genetic effects (AE model), (c) family resemblance is caused by
458
(a) Structural Model for Self-Report Data
Riemann et al.
fb) Structural Model for Peer Report Data
W// W//
(c) Structural Model for Self-Reporl and Peer Report Data
W//
Figure 1Structural Models tor Sell-Report, Peer Report, and Self-
and Peer Report Data
Note. A = additive genetic effects; D = genetic dominance effects; C = shared environ-ment effects; E = specific environmental effects; P, and P, = phenotype of Twin 1 or2; R[| to R24 = peer report on Twin I or 2 by Rater I to 4; SR] and SR2 = self-report ofTwin I or 2; PRj and PR2 = peer report (averaged across two raters) on Twin 1 or 2;Ms = method effects for self-reports; Mp = method effects for peer reports. Parametersalpha, beta, and gamma indicate the degree of shared environment, additive genetic, andgenetic dominance relationship between members of a pair: a = 1.0 for all twins; p = 1.0for MZ twins; p = .5 for DZ twins; 7= 1.0 for MZ twins; y= .25 for DZ twins.

Twin Study of NEO-FFI Scales
environmental influences shared by twins (CE model), (d) family resem-blance is due to additive genetic and dominance genetic effects (ADE model),and ( / ) family resemblance is caused by additive genetic plus shared envi-ronmental influences (ACE model). Thus, we identified the best-fitting modelby starting from tbe model with the fewest parameters (usually the AEmodel) that was not rejected significantly. We then tested whether modelswith more parameters (ADE, ACE) fitted the data significantly better than thereduced model using X' difference tests. However, we did not use the DEmodel because it is highly unlikely that there are genetic dominance effectswithout additive genetic effects (Roberts, 1967). If either of the two environ-mental models could not be rejected, this result is also reported.
RESULTS
Psychometric Quality of the Data
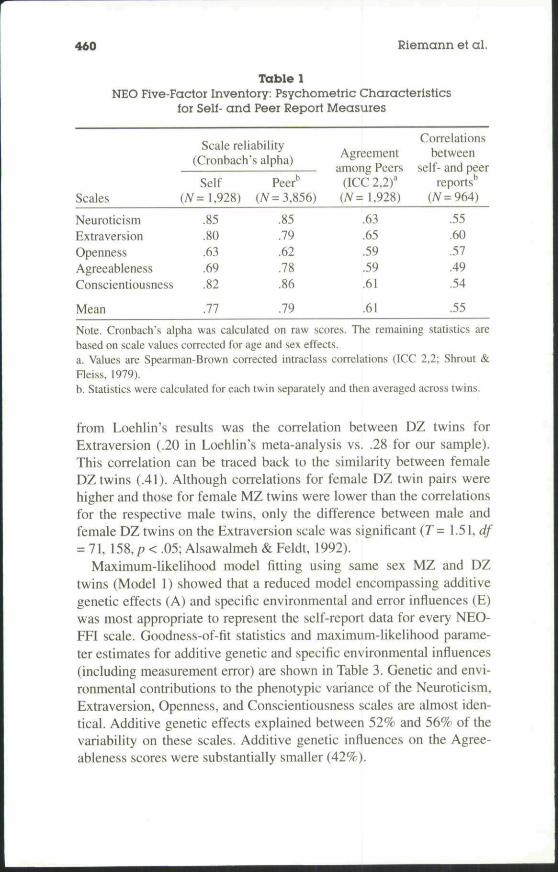
The intemal consistency of the self- and peer report scales of the NEO-FFI for Neuroticism, Extraversion, Agreeableness, and Conscientious-ness was acceptable. The intemal consistency for Openness toExperience can be judged as satisfactory. On average, the intemal con-sistency was marginally higher for the peer reports than for self-reports(see Table 1), and it was lower for Openness than for the other traits.
Altogether, the agreement between the two raters was good. Onaverage, Spearman-Brown corrected intraclass correlations betweenthe peer ratings were higher than .60 (see Table 1). Rater agreementshowed no substantial differences between MZ and DZ twins butmoderate differences between male and female targets. Averagedacross scales, rater agreement was .66 for male and .59 for female MZtwins and .60 for both female and male DZ twins.
Correlations between self-reports and averaged peer reports were inthe range of .46 to .60, indicating high validity for the self- and peerreport measures. They were only slightly inflated by age and sex dif-ferences. Table 1 lists correlations of age- and sex-corrected scores.
Sell-Report Data
We calculated twin similarities (intraclass correlations) for raw scoresand for age- and sex-corrected scores. Both analyses yielded nearlyidentical results. Therefore we limit our presentation of results to thecorrected data.
Intraclass correlations for the self-report data (Table 2) correspondedclosely to the data summarized by Loehlin (1992). The greatest deviation
460 Riemann et al.
Table INEO Five-Factor Inventory: Psychometric Characteristics
for Self- and Peer Report Measures
Scales
NeuroticismExtraversionOpennessAgreeablenessConscientiousness
Mean
Scale reliability(Cronbach's alpha)
Self(TV =1,928)
.85
.80
.63
.69
.82
.77
Peer^(A = 3,856)
.85
.79
.62
.78
.86
.79
Agreementamong Peers
(ICC 2,2)^(A'=1,928)
.63
.65
.5959.61
.61
Correlationsbetween
self- and peerreports
(TV =964)
.55
.60
.57
.49
.54
.55
Note. Cronbach's alpha was calculated on raw scores. The remaining statistics arebased on scale values corrected for age and sex effects.a. Values are Spearman-Brown corrected intraclass correlations (ICC 2,2; Shrout &Fleiss, 1979).b. Statistics were calculated for each twin separately and then averaged across twins.
from Loehlin's results was the correlation between DZ twins forExtraversion (.20 in Loehlin's meta-analysis vs. .28 for our sample).This correlation can be traced back to the similarity between femaleDZ twins (.41). Although correlations for female DZ twin pairs werehigher and those for female MZ twins were lower than the correlationsfor the respective male twins, only the difference between male andfemale DZ twins on the Extraversion scale was significant {T= 1.51. df= 71, 158,/7<.05;Alsawalmeh&Feldt, 1992).
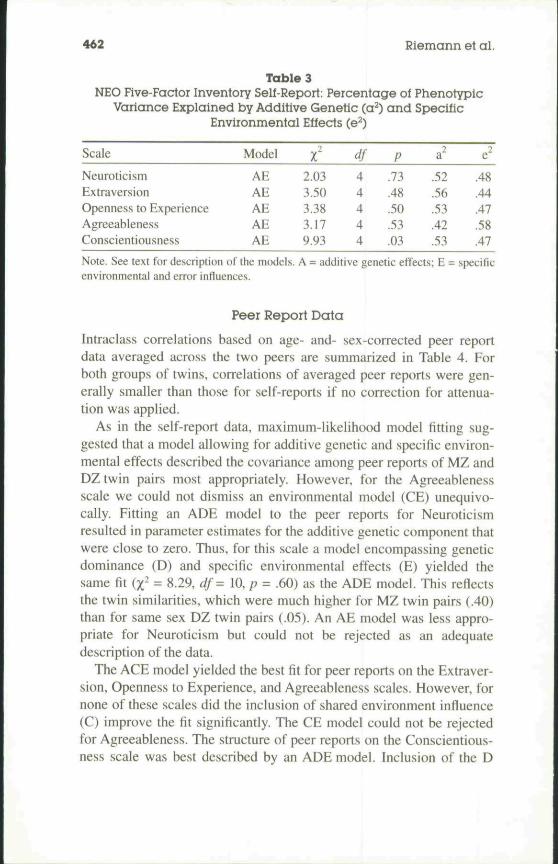
Maximum-likelihood model fitting using same sex MZ and DZtwins (Model 1) showed that a reduced model encompassing additivegenetic effects (A) and specific environmental and error influences (E)was most appropriate to represent the self-report data for every NEO-FFI scale. Goodness-of-fit statistics and maximum-likelihood parame-ter estimates for additive genetic and specific environmental influences(including measurement error) are shown in Table 3. Genetic and envi-ronmental contributions to the phenotypic variance of the Neuroticism,Extraversion, Openness, and Conscientiousness scales are almost iden-tical. Additive genetic effects explained between 52% and 56% of thevariability on these scales. Additive genetic influences on the Agree-ableness scores were substantially smaller (42%).

NQ
0) ^-v
(U o
:/) O
E ii
3 ^.O II
aO —
r-i 00— O
— (N
(N
^ II .1 = -2o > = S 'u
z w o < u
462 Riemann et al.
Table 3NEO Five-Factor Inventory Self-Report: Percentage of Phenotypic
Variance Explained by Additive Genetic (a^) and SpecificEnvironmental Effects (e^)
Scale
NeuroticismExtraversionOpenness to ExperienceAgreeablenessConscientiousness
Model
AEAEAEAEAE
2.033.503.383.179.93
df44444
P
.73
.48
.50
.53
.03
a"
.52
.56
.53
.42
.53
e"
.48
.44
.47
.58
.47
Note. See text for description of the models. A = additive genetic effects; E = specificenvironmental and error influences.
Peer Report Data
Intraclass correlations based on age- and- sex-corrected peer reportdata averaged across the two peers are summarized in Table 4. Forboth groups of twins, correlations of averaged peer reports were gen-erally smaller than those for self-reports if no correction for attenua-tion was applied.
As in the self-report data, maximum-likelihood model fitting sug-gested that a model allowing for additive genetic and specific environ-mental effects described the covariance among peer reports of MZ andDZ twin pairs most appropriately. However, for the Agreeablenessscale we could not dismiss an environmental model (CE) unequivo-cally. Fitting an ADE model to the peer reports for Neuroticismresulted in parameter estimates for the additive genetic component thatwere close to zero. Thus, for this scale a model encompassing geneticdominance (D) and specific environmental effects (E) yielded thesame fit (x^ = 8.29, df ^ 10, p = .60) as the ADE model. This reflectsthe twin similarities, which were much higher for MZ twin pairs (.40)than for same sex DZ twin pairs (.05). An AE model was less appro-priate for Neuroticism but could not be rejected as an adequatedescription of the data.
The ACE model yielded the best fit for peer report.s on the Extraver-sion, Openness to Experience, and Agreeableness scales. However, fornone of these scales did the inclusion of shared environment influence(C) improve the fit significantly. The CE model could not be rejectedfor Agreeableness. The structure of peer report.s on the Conscientious-ness scale was best described by an ADE model. Inclusion of the D

0H
•coad)CAhi
T3
0)d5
u
ou
N
6
o"
t l .
•:= O
u. >
o
OO fO O^ " ^
o — o —in Tf r~- r*— (N — (N
r - i r o i n o O N O O i n ' T i - r ' iO O C N ' ^ f S - ^ f N - ^ t N
O(N
OO tN -^ m rn(N m tN m O
(N r--
(N m
in CO CT^ ON '—I inO O fN ^ fN "n fN
— o
— r (N ^o o tN m 00 O
— oo O — (N00 tN m Tt r~-
.- — ^ ra i>
o '* " '
tN tN m00 m in
ou .
>
XU
ViVI<acc<L>a.O
ess
a_u
aj
3J
cVi3
_oc.—
coU

4164 Riemann et al.
parameter, however, did not improve the fit significantly. The modelthat allowed solely for specific environmental infiuences was rejectedfor each scale at the .001 significance level.
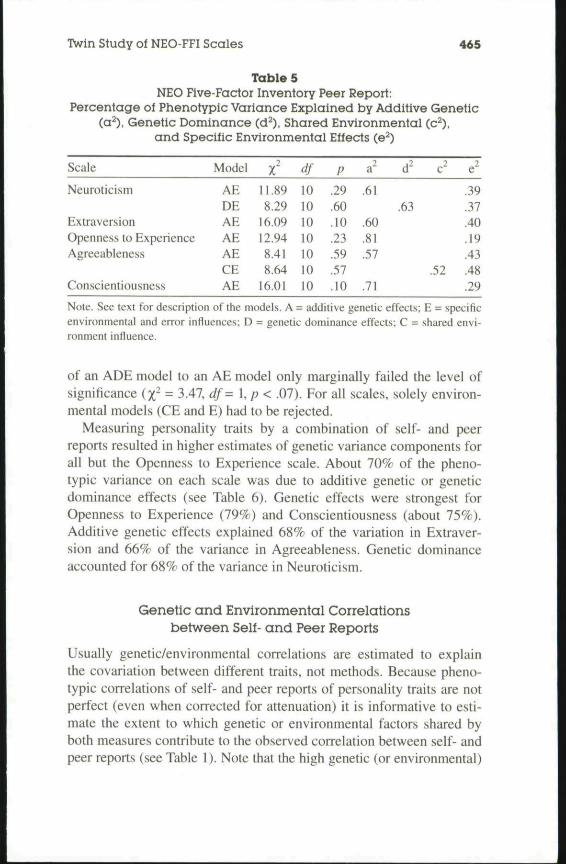
Maximum-likelihood parameter estimates for all models that couldnot be rejected are summarized in Table 5. Because our model for peerreport data allows for a separation of variance due to specific environ-mental influences and measurement error, we yielded substantiallyhigher estimates for genetic infiuences on personality variables com-pared to the self-report data. Genetic influences were highest forOpenness (81%) and Conscientiousness (71%). About 60% of the vari-ance on the Neuroticism, Extraversion, and Agreeableness scales wasexplained by genetic infiuences.
Self- a n d Peer Report Data
To obtain the most reliable parameter estimates we analyzed self- andpeer report data in a joint model. First, we tested whether a modelwithout method factors (Model 2) fits the variance-covariance matricesfor self- and peer reports. For all scales except Agreeableness, thismodel had to be rejected. However, the model including method fac-tors (Model 3) yielded a substantially better fit also for Agreeableness.Thus, we restrict our presentation to the results for Model 3. In someof our analyses the loadings on one of the method factors approachedzero and could not be identified. If this was the case, we fixed theloadings on the method factor for this and all nested analyses to zero.
The reduced AE model was most appropriate for Openness to Expe-rience and Agreeableness. For Extraversion the ACE model yieldedthe best fit, but this model was not significantly superior to the AEmodel {y} = 1.11, df= 1, ns). For Openness to Experience, both theACE and the AE models were rejected at the .05 level. Inclusion ofshared environmental infiuences for this scale did not improve the fitsignificantly (X" = 2.05. df= I, ns). Allowing for genetic dominancevariance improved the fit for Agreeableness only marginally (x^ =0.05. df= Ins).
For Neuroticism and Conscientiousness, genetic dominance was animportant source of variance. Inclusion of D improved the fit for Neu-roticism significantly (X' = 4.60, £//= \, p < .05) over the AE model.Fitting an ADE model for the Neuroticism data resulted in a parameterestimate for additive genetic variance close to zero. Thus a reducedDE model yielded the same fit. For Conscientiousness the comparison
Twin Study of NEO-FFI Scales 465
Table 5NEO Five-Factor Inventory Peer Report:
Percentage of Phenotypic Variance Explained by Additive Genetic(a^), Genetic Dominance (d^), Shared Environmental (c^),
and Specific Environmental Effects (e^)
Scale
Neuroticism
ExtraversionOpenness to ExperienceAgreeableness
Conscientiousness
Model
AEDEAEAEAECEAE
X11.898.29
16.0912.948.418.64
16.01
df
10101010101010
p
.29
.60
.10
.23
.59,57.10
a"
.61
.60
.81
.57
.71
d-
.63
c
.52
e"
.39
.37
.40
.19
.43
.48
.29
Note. See text for description of the models. A = additive genetic effects; E = specificenvironmental and error influences; D = genetic dominance effects; C = shared envi-ronment influence.
of an ADE model to an AE model only marginally failed the level ofsignificance {y} - 3.47, df - U p < .07). Eor all scales, solely environ-mental models (CE and E) had to be rejected.
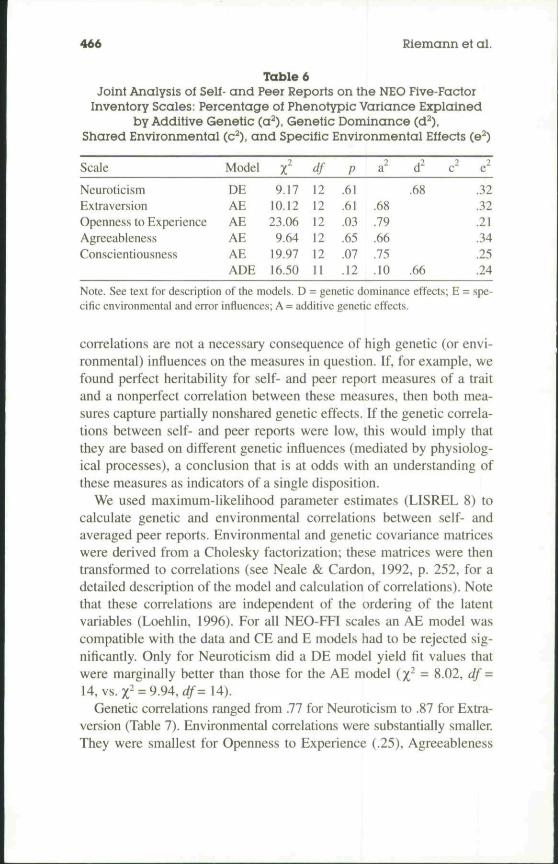
Measuring personality traits by a combination of self- and peerreports resulted in higher estimates of genetic variance components forall but the Openness to Experience scale. About 70% of the pheno-typic variance on each scale was due to additive genetic or geneticdominance effects (see Table 6). Genetic effects were strongest forOpenness to Experience (79%) and Conscientiousness (about 75%).Additive genetic effects explained 68% of the variation in Extraver-sion and 66% of the variance in Agreeableness. Genetic dominanceaccounted for 68% of the variance in Neuroticism.
Genetic and Environmental Correlationsbetween Self- and Peer Reports
Usually genetic/environmental correlations are estimated to explainthe covariation between different traits, not methods. Because pheno-typic correlations of self- and peer reports of personality traits are notperfect (even when corrected for attenuation) it is informative to esti-mate the extent to which genetic or environmental factors shared byboth measures contribute to the observed correlation between self- andpeer reports (see Table I). Note that the high genetic (or environmental)
466 Riemann et al.
Table 6Joint Analysis of Self- and Peer Reports on the NEO Five-Factor
Inventory Scales: Percentage of Phenotypic Variance Explainedby Additive Genetic (a^), Genetic Dominance (d^),
Shared Environmental (c^), and Specific Environmental Effects (e^)
Scale
NeuroticismExtraversionOpenness to ExperienceAgreeablenessConscientiousness
Model
DEAEAEAEAEADE
r9.17
10.1223.069.64
19.9716.50
df121212121211
P
.61
.61
.03
.65
.07
.12
a'
.68
.79
.66
.75
.10
d' (
.68
.66
•} e'.32.32.21.34.25.24
Note. See text for description of the models. D = genetic dominance effects; E = spe-cific environmental and error influences; A = additive genetic effects.
correlations are not a necessary consequence of high genetic (or envi-ronmental) influences on the measures in question. If, for example, wefound perfect heritability for self- and peer report measures of a traitand a nonperfect correlation between these measures, then both mea-sures capture partially nonshared genetic effects. If the genetic correla-tions between self- and peer reports were low, this would imply thatthey are based on different genetic influences (mediated by physiolog-ical processes), a conclusion that is at odds with an understanding ofthese measures as indicators of a single disposition.
We used maximum-likelihood parameter estimates (LISREL 8) tocalculate genetic and environmental correlations between self- andaveraged peer reports. Environmental and genetic covariance matriceswere derived from a Cholesky factorization; these matrices were thentransformed to correlations (see Neale & Cardon, 1992, p. 252, for adetailed description of the model and calculation of correlations). Notethat these correlations are independent of the ordering of the latentvariables (Loehlin, 1996). Eor all NEO-EEI scales an AE model wascompatible with the data and CE and E models had to be rejected sig-nificantly. Only for Neuroticism did a DE model yield fit values thatwere marginally better than those for the AE model (X^ = 8.02, df =14, vs. x = 9.94, 4/"= 14).
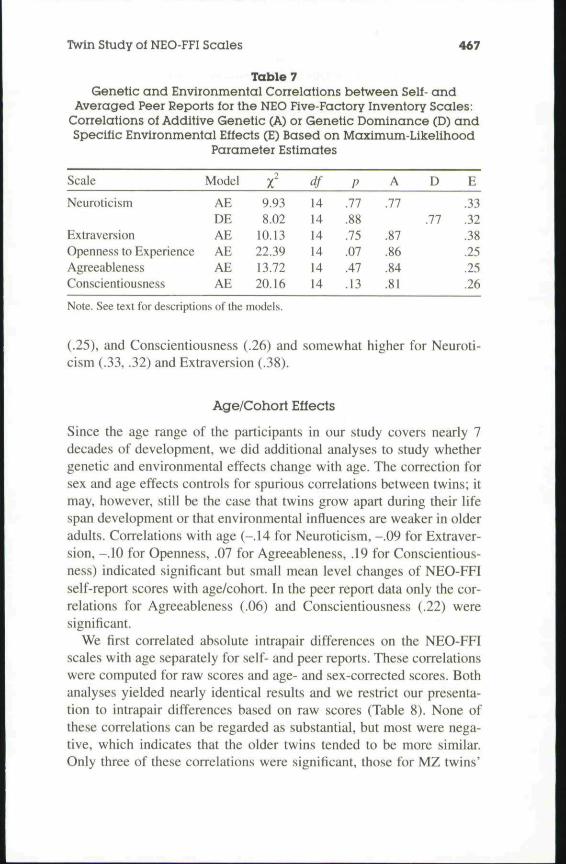
Genetic correlations ranged from .77 for Neuroticism to .87 for Extra-version (Table 7). Environmental correlations were substantially smaller.They were smallest for Openness to Experience (.25), Agreeableness
Twin Study of NEO-FFI Scales 467
Table 7Genetic and Environmental Correlations between Self- and
Averaged Peer Reports for the NEO Five-Factory Inventory Scales:Correlations of Additive Genetic (A) or Genetic Dominance (D) andSpecific Environmental Effects (E) Based on Maximum-Likelihood
Parameter Estimates
Scale
Neuroticism
ExtraversionOpenness to ExperienceAgreeablenessConscientiousness
Model
AEDEAEAEAEAE
9.938.02
10.1322.3913.7220.16
df141414141414
P
.11
.88
.75
.07
.47
.13
A
.77
.87
.86
.84
.81
D
.77
E
.33
.32
.38
.25
.25
.26
Note. See text for descriptions of the models.
(.25), and Conscientiousness (.26) and somewhat higher for Neuroti-cism (.33, .32) and Extraversion (.38).
Age/Cohort Effects
Since the age range of the participants in our study covers nearly 7decades of development, we did additional analyses to study whethergenetic and environmental effects change with age. T\\c correction forsex and age effects controls for spurious correlations between twins; itmay, however, still be the case that twins grow apart during their lifespan development or that environmental influences are weaker in olderadults. Correlations with age (-.14 for Neuroticism, -.09 for Extraver-sion, -.10 for Openness, .07 for Agreeableness, .19 for Conscientious-ness) indicated significant but small mean level changes of NEO-FFIself-report scores with age/cohort. In the peer report data only the cor-relations for Agreeableness (.06) and Conscientiousness (.22) weresignificant.
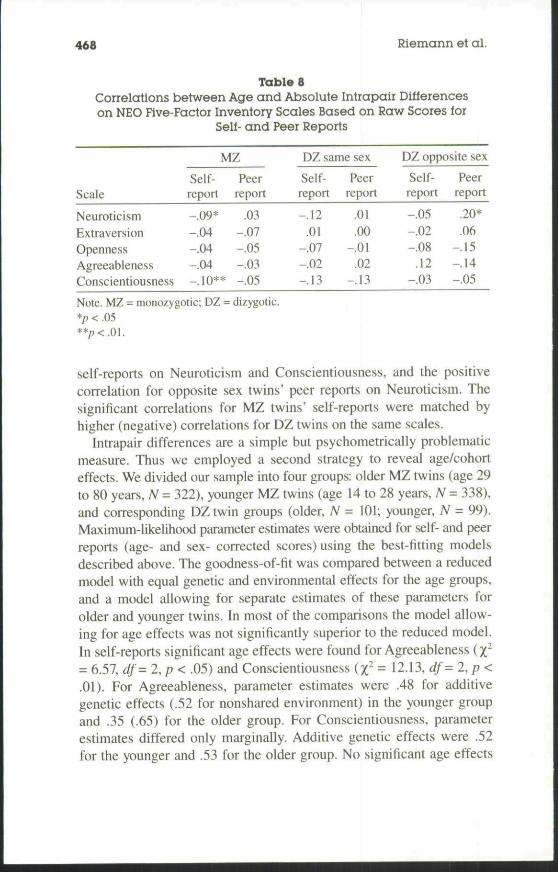
We first correlated absolute intrapair differences on the NEO-FFIscales with age separately for self- and peer reports. These correlationswere computed for raw scores and age- and sex-corrected scores. Bothanalyses yielded nearly identical results and we restrict our presenta-tion to intrapair differences based on raw scores (Table 8). None ofthese correlations can be regarded as substantial, but most were nega-tive, which indicates that the older twins tended to be more similar.Only three of these correlations were significant, those for MZ twins'
468 Giemann et al.
TablesCorrelations between Age and Absolute Intrapair Differenceson NEO Five-Factor Inventory Scales Based on Raw Scores for
Self- and Peer Reports
Scale
NeuroticismExtraversionOpennessAgreeablenessConscientiousness
MZ
Self-report
-.09*-.04-.04-.04-.10**
Peerreport
.03-.07-.05-.03-.05
DZ same sex
Self-report
-.12.01
-.07-.02-.13
Peerreport
.01
.00-.01
.02-.13
DZ opposite sex
Self-report
-.05-.02-.08
.12-.03
Peerreport
.20*
.06-.15-.14-.05
Note. MZ = monozygotic; DZ = dizygotic.*/; < .05
self-reports on Neuroticism and Conscientiousness, and the positivecorrelation for opposite sex twins' peer reports on Neuroticism. Thesignificant correlations for MZ twins' self-reports were matched byhigher (negative) correlations for DZ twins on the same scales.
Intrapair differences are a simple but psychometrically problematicmeasure. Thus we employed a second strategy to reveal age/cohorteffects. We divided our sample into four groups: older MZ twins (age 29to 80 years, A = 322), younger MZ twins (age 14 to 28 years, N = 338),and corresponding DZtwin groups (older, N = 101; younger. A' = 99).Maximum-likelihtXKl parameter estimates were obtained for self- and peerreports (age- and sex- corrected scores) using the best-fitting modelsdescribed above. The goodness-of-fit was compared between a reducedmodel with equal genetic and environmental effects for the age groups,and a model allowing for separate estimates of these parameters forolder and younger twins. In most of the comparisons the model allow-ing for age effects was not significantly superior to the reduced model.In self-reports significant age effects were found for Agreeableness ( / -= 6.57, df= 2, p < .05) and Conscientiousness (x^ = \2A3, df == 2, p <.01). For Agreeableness, parameter estimates were .48 for additivegenefic effects (.52 for nonshared environment) in the younger groupand .35 (.65) for the older group. For Conscientiousness, parameterestimates differed only marginally. Additive genetic effects were .52for the younger and .53 for the older group. No significant age effects

Twin Study of NEO-FFI Scales 469
were found in peer reports, although the effect for Conscientiousnesswas nearly replicated (x^ = 5.40, df= 2.p< .07; additive genetic effectfor younger group, .69, vs. .78 for older group).
To summarize, the results of these two analyses do not support gen-eral claims about the association between age and genetic or environ-mental effects on personality. We found stronger age effects inself-reports than in peer reports. These were most consistent for Con-scientiousness but, contrary to expectations, on this scale older twinswere more similar than younger twins.
DISCUSSION
The results of our study support six major conclusions. First, ouranalyses of self-report data replicate earlier findings of a substantialgenetic influence on all broad personality dimensions measured by theNEO-FFI. Second, this influence is not only demonstrated for self-report data but also for peer reports. Third, the pattern of genetic andenvironmental influences is similar for self- and peer report data; thusour analyses validate findings solely based on self-reports and invali-date the hypothesis that self- report data of DZ twins are substantiallydistorted by contrast effects. Fourth, self-report studies of single per-sonality measures tend to underestimate genetic influences becausethey do not allow the separation of error variance from variance due tospecific environmental influences. Fifth, correlations between self- andpeer reports reflect the same genetic influences to a much higherextent than identical environmental influence. Finally, there is littleevidence of differential heritability across the age cohorts. We shallbriefly discuss each of these conclusions in tum.
Self-Reports
If we compare the correlations of self-reports between MZ and DZtwins that were obtained in our study to those of the twin studies sum-marized by Loehlin (1992), Bergeman et al. (1993), and Jang et al.(1996), striking similarities are obvious. Consistent differences betweenthe five scales are hardly noticeable across the studies, although MZtwin correlations tend to be highest for Extraversion and lowest forAgreeableness. DZ twins tend to correlate highest on Openness. Ourheritability estimates are highly similar to those reported by Jang et al.(1996) for the NEO-PI-R and Bouchard's (1994) results for twins

470 Riemann etal.
reared together. They are higher than those reported by Loehlin (1992)and Bergeman et al. (1993), and genetic dominance effects (or theinfluence of more similar MZ twins' environments) are absent in ourdata. This difference may be caused by the differences in the question-naire measures, divergent procedures regarding the recruitment oftwins, and the absence of twins reared apart in our study.
Peer Reports
The analysis of peer report data clearly demonstrates family resem-blance for the five dimensions under study. Comparing the results forself-report and peer report data across the NEO-FFI scales, we found asubstantial similarity. A model that explains family resemblance viaadditive genetic and specific environmental effects is most appropriatefor each self-report scale and cannot be rejected for any peer reportmeasure. However, tendencies that are observed for the self-reportsseem to be stronger in the peer report data. For self-reports we foundthe highest difference between MZ twin and DZ twin correlations forNeuroticism and the smallest difference for Agreeableness. For thesescales, models allowing for genetic dominance and shared environ-mental effects, respectively, could not be rejected in the domain ofpeer report data.
It is important to note that the differences for these two scales are inopposite directions. For Neuroticism the difference between MZ and DZtwins is more pronounced (due to low DZ twin correlations) in peerreports, whereas for Agreeableness this difference becomes smaller (dueto low MZ twin correlations). This pattern of results contradicts sug-gestions that self-reports of MZ and DZ twins are generally distortedby assimilation or contrast effects and validates self-report studies.
Sell- and Peer Reports
We regard the joint analysis of self- and peer reports as providing themost reliable and valid parameter estimates in our study because it uti-lizes all available measures and derives trait scores from independentsources. It emphasizes the importance of additive genetic and nonsharedenvironmental influences on Bxtraversion, Openness to Experience, andAgreeableness, and of genetic dominance effects for Neuroticism andConscientiousness. Despite the similarities between .self- and peerreport data, both sources of data result in substantially different esti-

Twin Study of NEO-FFI Scales 471
mates of environmental and genetic influences on the five personalityvariables under study. The increase in the averaged estimates ofgenetic influences from .51 for self-reports to .66 for peer report mea-sures is a consequence of separating error variance from nonsharedenvironmental influences in the latter analyses. This is comparable to acorrection for attenuation in classical test theory. Subtracting errorvariance from nonshared environmental effects (taking the averageinternal consistency of the NEO-FFI scales as a crude estimate of truescore variance) simulates this effect for the self-report data and yieldsheritability estimates of about .70.
The finding that heritability estimates for the joint analysis of self-and peer reports are somewhat higher (.71 on average) than those forpeer reports is not merely a trivial result of aggregating self- and peerreports into a more reliable measure because error variance is con-trolled for in both analyses. The joint analysis demonstrates that thebroad personality traits (constructs) measured by independent judg-ments are strongly determined by genetic influences. This result sug-gests that the covariance between self- and peer reports is largely dueto shared genetic effects, which is confirmed by our analysis of geneticand environmental correlations. The correlations of additive geneticeffects were on average higher than .80. Nonshared environmentaleffects are reflected commonly by self- and peer reports to a substan-tially smaller extent (.32). This result implies that behavior-geneticresearch can profit from the joint analyses of self- and peer report data.
None of our analyses shows marked differences between the dimen-sions of the five-factor model regarding genetic versus environmentalinfluences. Although heritability estimates were lowest for Agreeable-ness in all of our analyses, the difference between this scale and Neu-roticism and Extraversion is only marginal in the analyses involvingpeer reports. The latter data result in estimates of genetic influences onOpenness to Experience and Conscientiousness somewhat higher thanthose for the remaining scales. Although previous studies have shownthe importance of nonadditive genetic effects for Neuroticism, Consci-entiousness, and Extraversion, these effects are particularly high in ourdata for Neuroticism and Conscientiousness. The robustness of thesefindings has to be tested in independent samples.
The present analyses are based on only one instrument to measurethe five-factor model, the NEO-FFI. Thus it remains to be seen how theinclusion of multiple measures of the five-factor model, in combinationwith multiple perspectives (self- vs. peer), will aftect our findings. An

472 Siem.ann et al.
extension of the multitrait multi-observer approach presented in thisstudy may help to clarify the genetic and environmental structure ofpersonality traits. A thorough understanding of this structure is alsonecessary for the interpretation of molecular-genetic studies linkingpersonality traits to quantitative gene loci (Cloninger. Adolfsson, &Svrakic, 1996; Ebstein et al., 1996; Plomin, 1995).
Age Eftects
The absence of substantial differential heritability of the five factorsacross age/cohort groups replicates earlier findings of Bergeman et al.(1993). It is, however, at odds with recent meta-analyses. This discrep-ancy is difficult to explain. It would be informative to examinewhether differential heritability is systematically associated with spe-cific facets of the five-factor model. Likewise, differential heritabilitymay be stronger in comparisons including children and adolescents,who are underrepresented in Bergeman et al.'s and the present study.
CONCLUSION
ln conclusion, the present study has shown that the inclusion of peerreports into behavior-genetic studies of personality variables providesa valuable tool. Peer report data provide the opportunity to validate theresults of self-report studies and, more important, they shift the focusfrom personality measures to personality constructs. Further researchshould include additional sources of data (e.g., objective personalitymeasures or physiological indicators) or multiple questionnaires toconduct systematic multitrait multimethod behavior-genetic studies ofpersonality.
REFERENCES
Allport, G. W. (1937). Personality: A psychological interpretation. New York: Holt.Alsawalmeh, Y. M., & Feldt, L. S. (1992). Test of the hypothesis that the intraclass
reliability coefficient is the same for two measurement procedures. Applied Psy-chological Measurement, 16,195-205.
Angleitner. A.. Riemann, R.. Spinath. F. M.. Hempel. S . Thiel, W, & Strelau, J. (1995,July). Fir.sl report on the Bielefeld samples. Paper presented at the Workshop onGenetic Studies on Temperament and Personality, Warsaw-Pultusk, Poland.
Beer, J. M., & Rose, R. J. (1995). The five factors of personality: A study of twins.Behavior Genetics, 25, 254.

Twin Study of NEO-FFI Scales
Bergeman. C. S., Chipuer, H. M., Plomin, R., Pedersen, N. L., McCleam, G. E.. Nes-selroade, J. R.. Costa. P. T. Jr.. & McCrae, R. R. (199.^). Genetic and environmen-tal effects on Openness to Experience, Agreeableness, and Conscientiousness: Anadoption/twin study. Journal of Personality. 61, 159-179.
Borkenau, P (1993). Aniage and Umwelt [Nature and nurture]. Gottingen. Germany:
Hogrefe.Borkenau, P.. & Ostendorf. F. (1993). NeoFUnf-Faktoren-lnventar (NEO-FFI) [NEO
Five Factor Inventory], Gottingen, Germany: Hogrefe.Bouchard, T. J, (1994, June 17). Genes, environment, and personality. Science, 264,
1700-1701.Bouchard. T J.. & McGue. M. (1981, May 29). Familial studies of intelligence: A
review. Science. 212. 1055-1059.Carey, G. (1986). Sibling imitation and contrast effects. Behavior Genetics, 16,
319-343.Cattell, R. B. (1950). Personality: A systetnutic theoretical ar^d factual study. New
York: McGraw-Hill.Chaplin, W. F. (1991). The next generation of moderator research in personality psy-
chology. Journal of Personality, 59. 141-178.Cloninger, C. R., Adolfsson, R., & Svrakic, N. M. (1996). Mapping genes for human
personality. Nature Genetics. 12, 3-4.Costa, P. T, Jr., & McRae, R. R. (1985). NEO Personality Inventory: Manual Form S
and Form R. Odessa. FL: Psychological Assessment Resources.Costa, P. T. Jr.. & McCrae, R. R. (1989). NEO-Pl/FFI manual supplement. Odessa,
FL: Psychological Assessment Resources.Costa, P T., Jr., & McCrae, R. R. (1992). Revised NEO Personality Inventory (NEO
PJ-R) and NEO Five-Factor Inventory: Professional manual. Odessa, FL: Psycho-logical Assessment Resources.
Digman. J. M. (1990). Personality structure: Emergence of the five-factor model.Amiual Review of Psychology. 41. 417-440.
Eaves. L. J.. Eysenck, H. J., & Martin, N, G. (1989). Genes, culture and personality:An empirical approach. London: Academic Press.
Ebstein, R. P., Novick, O., Umansky, R., Priel, B., Osher. Y., Blaine. D., Bennett,E. R.. Nemanov. L.. Katz, M.. & Belmaker. R. H. (1996). Dopamine D4 receptor(D4DR) polymorphism associated with the human personality trait of novelty seek-ing. Nature Genetics, 12. 78-80.
Eysenck, H. J.. & Eysenck. S. B. G. (1964). Manual of the Eysenck Personality Inven-tory. London: University of London Press.
Eysenck. H. J., & Eysenck. S. B. G. (1975). Manual of the Eysenck Personality Ques-tionnaire. London: Hodder& Stroughton.
Eysenck. H. J., & Eysenck. S. B. G. (1991). Manual of the Eysenck Personality Scales.
London: Hodder & Stroughton.Fiske. D. W. (1971). Measuring the concepts of personality. Chicago: Aldine.Funder, D, C. (1987). Errors and mistakes: Evaluating the accuracy of social judg-
ment. Psychological Bulletin, 101, 75-90.Heath, A. C . Neale, M.C., Kes.sler, R. C , Eaves, L. J.. & Kendler. K. S. (1992). Evi-
dence for genetic influences from self-reports and informant ratings. Journal ofPersonality and Social Psychology, 63, 85-96.

474 Riemann etal.
Heymans, G., & Wiersma, E. (1906). Beitrage zur speziellen Psychologie auf Grundeiner Massenuntersuchung [Contributions to differential psychology based on astudy of a large sample]. Zeitschrift fUr Psychologie, 42, 8-127.
Jang, K. L.. Livesley. W. J., & Vemon, P. A. (1996). Heritability of the Big Fivedimensions and their facets: A twin study. Journal of Personality. 64. 577-591.
John, O. P. (1990). The "Big Five" factor taxonomy: Dimensions of personality in thenatural languages and in questionnaires. In L. A. Pervin (Ed.), Handbook of per-sonality: Theory and research (pp. 66-100). New York: Guilford.
John, O. P., Angleitner, A., & Ostendorf, F. (1988). The lexical approach topersonality: A historical review of trait taxonomic research. European Journal ofPersonality, 2, 171-203.
Joreskog. K.. & Sorbom, D. (1993). LISREL 8: Structural equation modeling with theSIMPLIS command language. Chicago: Scientific Software International.
Kenny. D. A. (1991). A general model of consensus and accuracy in interpersonal per-ception. P.sychological Review, 98, 155-163.
Kenny. D. A. (1994). Interpersonal perception: A social relations analysis. New York:Guilford.
Loehlin, J. C. (1986). Heredity, environment, and the Thurstone Temperament Sched-ule. Behavior Genetics, 16. 61-73.
Loehlin. J. C. (1992). Genes and environment in personality development. NewburyPark, CA: Sage.
Loehlin, J. C. (1996). The Cholesky approach: A cautionary note. Behavior Genetics,26, 65-69.
Lykken. D. T. (1978). The diagnosis of zygosity in twins. Behavior Genetics. 8.437-473.
McCartney. K., Harris, M., & Bemieri, F. (1990). Growing up and growing apart: Adevelopmental meta-analysis of twin studies. Psychological Bulletin, 107. 226-237.
McCrae. R. R. (1989). Why I advocate the five-factor model: Joint factor analyses ofthe NEO-PI with other instruments. In D. M. Buss & N. Cantor (Eds). Personalitypsychology: Recent trends and emerging directions (pp. 237-245). New York:Springer.
McCrae. R. R. (1996). Social consequences of experiential openness. PsychologicalBulletin, 120, 323-337.
McGue. M., & Bouchard. T. J. (1984). Adjustment of twin data for the effects of ageand sex. Behavior Genetics, 14, 325-343.
Neale, M. C , & Cardon. L. R. (1992). Methodology for genetic studies of twins andfamilies. Dordrecht, The Netherlands: Kluwer Academic.
Nichols, R. C , & Bilbro. W. C. (1996). The diagnosis of twin zygosity. Acta Genelica,16, 265-275.
Oniszczenko, W., Angleitner, A., Strelau, J., & Angert, T. (1993). The questionnaire oftHins'physical resemblance. Unpublished questionnaire. Department of Psychol-ogy, University of Warsaw and University of Bielefeld.
Paunonen. S.V. (1989). Consensus in personality judgments: Moderating effects oftarget-rater acquaintanceship and behavior observability. Journal of Personalityand Social Psychology, 56. 823-833.

Twin Study of NEO-FFI Scales 475
Pedersen, N. L. (1993). Genetic and environmental continuity and change in personal-ity. In T. J. Bouchard & P. Propping (Eds.). Twins as a too! of behavior genetics(pp. 147-162). London: Wiley.
Plomin. R. (1995). Molecular genetics and psychology. Current Directions in Psycho-logical Science, 4, 114—117.
Plomin, R.. DeFries, J. E., & McCleam, G. E. (1990). Behavioral genetics: A primer(2nd ed.). New York: Academic Press.
Plomin, R.. & McCleam. G. E. (Eds.). (1993). Nature, nurture and psychology. Wash-ington. DC: American Psychological A.ssociation.
Roberts. R. C. (1967). Some concepts and methods in quantitative genetics. In J. Hirsch(Ed.), Behavior-genetic analysis (pp. 214-257). New York: McGraw-Hill.
Rose. R. (1995). Genes and human behavior. Annual Review of P.sychology. 46,625-654.
Rowe. D. C. (1994). The limits of family influence. New York: Guilford.Saudino, K. J., & Eaton. W. O. (1991). Infant temperament and genetics: An objective
study of motor activity level. Child Development, 62. 885-898.Shrout. P. E.. & Fleiss. J. L. (1979). Intraclass correlation: Use in assessing rater relia-
bility. Psychological Bulletin, 86. 420-428.Tellegen, A., Lykken. D. T., Bouchard. T. J.. Wilcox, K. M.. Segal, N. L., & Rich. S.
(1988). Personality similarity in twins reared apart and together. Journal of Person-aHty and Social Psychology, 54, 1031 -1039.






























