Embed Size (px)
Citation preview

GLd
JGa
Gb
2c
B
a
ARRA
KLLIGNS
1
ta
mFJ
0h
Veterinary Parasitology 193 (2013) 193– 199
Contents lists available at SciVerse ScienceDirect
Veterinary Parasitology
jo u rn al hom epa ge : www.elsev ier .com/ locate /vetpar
enetic characterization of the blood-sucking nematodesibyostrongylus dentatus and Libyostrongylus douglassii supports theirifferent evolutionary history
osiana G. Andradea, Alena M. Iniguezb,∗, Adriane N. Souzac, Valéria C.L. Marquesc,onc alo A. de Souza Filhoc, Clóvis P. Santosa, Renato A. DaMattaa
Laboratório de Biologia Celular e Tecidual, Centro de Biociências e Biotecnologia, Universidade Estadual do Norte Fluminense, 28013-602 Campos dosoytacazes, RJ, BrazilLaboratório de Genética Molecular de Microorganismos, Laboratório de Biologia de Tripanosomatídeos, Instituto Oswaldo Cruz, Fundac ão Oswaldo Cruz,1045-900 Rio de Janeiro, RJ, BrazilLaboratório de Biotecnologia, Centro de Biociências e Biotecnologia, Universidade Estadual do Norte Fluminense, 28013-602 Campos dos Goytacazes, RJ,razil
r t i c l e i n f o
rticle history:eceived 6 May 2012eceived in revised form 29 October 2012ccepted 23 November 2012
eywords:ibyostrongylus douglassiiibyostrongylus dentatusnternal Transcribed Spacerenetic diagnosisematodestruthio camelus
a b s t r a c t
Libyostrongylus sp. are nematodes that infect ostriches. Libyostrongylus douglassii was firstdescribed in ostriches from several countries in the world. Later Libyostrongylus dentatuswas morphologically identified in ostriches in the USA and Brazil, and mixed infection iscommon in the latter country. The internal transcribed spacer (ITS) region of the ribosomalDNA gene is used for genetic variability assessment and phylogenetic reconstruction formany organisms. Through genetic analysis the status of different species morphologicallydefined was confirmed and a molecular method was developed to differentiate both species.ITS1, 5.8S, ITS2 regions of L. douglassii and L. dentatus were characterized. Regarding com-plete ITS region, the K2-p genetic distance between the species was 0.060 (SE 0.008) and theintra-specific distance was 0.002 (SE 0.001) for L. dentatus and 0.006 (SE 0.002) for L. dou-glassii. NJ and MP phylogenetic analysis of ITS1 and ITS2 regions indicated that both species
belong to the Trichostrongylidae family, and are evolutionarily different, suported by highbootstrap value. Based on ITS DNA polymorphisms, a molecular approach was designed todetect both species. These results are the first molecular characterization of L. douglassiiand L. dentatus, and provide new tools for the identification of these parasites of veterinaryimportance.. Introduction
L. douglassii and L. dentatus are ostrich-specific nema-odes found on the mucosal lining of the proventriculusnd ventriculus (Hoberg et al., 1995; Craig and Diamond,
∗ Corresponding author at: Laboratório de Biologia de Tripanoso-atídeos, Pavilhão Rocha Lima, Sala 518, Instituto Oswaldo Cruz,
undac ão Oswaldo Cruz, Av. Brasil 4365, Manguinhos, 21045-900 Rio deaneiro, RJ, Brazil. Tel.: +55 21 2562 1416.
E-mail address: [email protected] (A.M. Iniguez).
304 4017- © 2012 Elsevier B V . . ttp://dx.doi.org/10.1016/j.vetpar.2012.11.024
Open access under the Elsevier OA license.
© 2012 Elsevier B.V .
1996; Ederli et al., 2008a; Andrade et al., 2011a). L. dou-glassii, first described by Cobbold (1882) in ostriches inAfrica, is associated to clinical signals as anorexia, weightloss, anemia, proventriculitis and mortality in chicks andadults (Reinecke, 1983; Bastianello et al., 2005; Santoset al., 2010). However, clinical signals have not beenassociated to L. dentatus probably because this species
Open access under the Elsevier OA license.
was only described in 1995 as morphologically simi-lar to L. douglassii. L. dentatus was first morphologicallydescribed in ostriches in the United States of America(Hoberg et al., 1995), and later it was found in Brazil when

ry Paras
194 J.G. Andrade et al. / Veterinanew morphological characters were added to the originaldescription (Ederli et al., 2008a). By using a morphologicaldiagnosis method based on the examination of infectivelarvae obtained after coproculture of feces (Ederli et al.,2008b), all Brazilian farms examined that were positive forLibyostrongylus sp., so far, have mixed infections (Andradeet al., 2011b). The presence of L. dentatus in ostriches pos-itive for L. douglassii has been probably ignored in othercountries because ostriches on Brazilian farms are orig-inated from countries without notification of the formerparasite. Thus, mixed infections may occur in ostriches allover the world. A molecular method to differentiate bothspecies is needed to allow rapid identification which isessential to a better understanding of libyostrongylosis inthe world.
The genus Libyostrongylus had been classified as a mem-ber of the Trichostrongylidae family, based on analysisof 22 morphological characters (Hoberg and Lichtenfels,1994). However, Durette-Desset et al. (1999) based on 48morphological characters classified Libyostrongylus in theCooperiidae family. Phylogenetic relationships betweenhelminthes species based only on morphological datahave been repeatedly redefined, but continue to providethe foundation for nematode, identification, taxonomicclassification and evolution comprehension. Moreover,molecular data have contributed to nematodes phylo-genetic classification (Hoste et al., 1998; Bellocq et al.,2001), and includes L. douglassii (Chilton et al., 2001).However, L. dentatus has not been genetically analyzedor its molecular phylogenetic position determined, untilnow.
The ribosomal DNA (rDNA) internal transcribed spacer(ITS) is a genetic marker largely used to distinguish speciesof nematodes (Lichtenfels et al., 1997; Chilton et al., 2001;Cutillas et al., 2007). This part of the nuclear rDNA, whichcontains the first internal transcribed spacer (ITS1), the5.8S rRNA gene and second internal transcribed spacer(ITS2), is subject to significant variation both in size andnucleotide sequence that correlate with the divergence ofnematode species (Chilton, 2004). The only one phyloge-netic analysis of L. douglassii, is based only on the ITS2sequence of one individual and situates this species on theTrichostrongylidae family (Chilton et al., 2001), corrobo-rating with the morphological classification proposed byHoberg and Lichtenfels (1994). The present work aimeda molecular characterization of ribosomal ITS region ofL. dentatus and L. douglassii, infer a molecular phylogeny,and develop a molecular approach to differentiate bothspecies.
2. Materials and methods
2.1. Parasites
Adults of L. dentatus and L. douglassii were collectedfrom the proventriculus during necropsy of an ostrich.These nematodes were washed repeatedly with bi-distilled
water. Female species were identified by morpholog-ical characteristics (Hoberg et al., 1995; Ederli et al.,2008a). Samples were preserved at −70 ◦C until DNAextraction.itology 193 (2013) 193– 199
2.2. DNA extraction
Genomic DNA extractions were performed on 12 indi-vidual females of each species (deposition number ofrepresentative species to L. dentatus and L. douglassiiare 35413 and 35414, respectively, in the Helmintholog-ical Collection of the Oswaldo Cruz Institute, FundacãoOswaldo Cruz, Rio de Janeiro, Brazil). The specimens ofL. dentatus and L. douglassii were identified as Lde andLdo, respectively, and numbered. Each individual nema-tode was macerated in 0.3 ml of the Plant DNAzol® reagentwith 30 �g of RNAse. A volume of 0.3 ml of chloroform wasadded, mixed vigorously, further incubated at room tem-perature for 5 min and centrifuged (12,000 × g, 10 min). Theaqueous phase was transferred to a fresh tube, mixed with0.225 ml of 100% ethanol, incubated at room temperaturefor 5 min and centrifuged to precipitate DNA. PrecipitatedDNA was mixed with 0.3 ml of 75% Plant DNAzol reagentin ethanol, incubated at room temperature for 5 min, cen-trifuged and the supernatant was discarded. The DNA pelletwas washed with 0.3 ml of 75% ethanol, centrifuged, andthe supernatant discarded. The DNA extraction protocolused was according to Plant DNAzol® manufacturer with afew modifications. The DNA pellet was air-dried and elutedon 25 �L of water.
2.3. DNA amplification and sequencing
Complete ITS region was amplified by PCR using the for-ward primer NC5 (5′-GTAGGTGAACCTGCGGAAGGATCATT-3′) and reverse primer NC2 (5′- TTAGTTTCTTTTCCTCCGCT-3′) (Hillis and Dixon, 1991). The PCR reaction volumewas 50 �L containing 1× PCR buffer, 200 �M of dNTP,3 mM of MgCl2, 40 �mol of each primer, 2 units ofPlatinum® Taq DNA polymerase (Invitrogen) and 2 �Lof extracted DNA template. The PCR program usedwas: 94 ◦C/3 min, (94 ◦C/40 s, 60 ◦C/40 s, 72 ◦C/1 min) × 40cycles, 72 ◦C/7 min. The amplified products were visualizedin 1.8% agarose gel stained with ethidium bromide. PCRproducts were purified using GFX PCR DNA and Gel BandPurification Kit (GE) according to manufacturer’s protocol.PCR products were sequenced using Big Dye Termina-tor v 3.1 Cycle Sequencing Ready Reaction kit (AppliedBiosystems) in a 3100 Automated DNA Sequencer (AppliedBiosystems) as recommended by the supplier.
2.4. Sequence analysis
Amplicons were sequenced in each direction usingthe NC5 and NC2 primers, in separate reactions. Bio Editv7.0.4.1 and DNASTAR Lasergene SeqMan v7.0.0 programswere used for analyzing and editing the DNA sequences.Sequences were aligned using Clustal W (Thompson et al.,1994). Genetic signatures of L. douglassii that represent thespecific nucleotides of the species were determined.
The genetic distances and phylogenetic analyseswere conducted using MEGA v4 (Tamura et al., 2007).
Intra-specific and inter-species genetic distances werecalculated using Kimura 2-parameter (K2-p) model(Kimura, 1980), complete deletion treatment and standarderror estimated (SE) by a bootstrap procedure (2000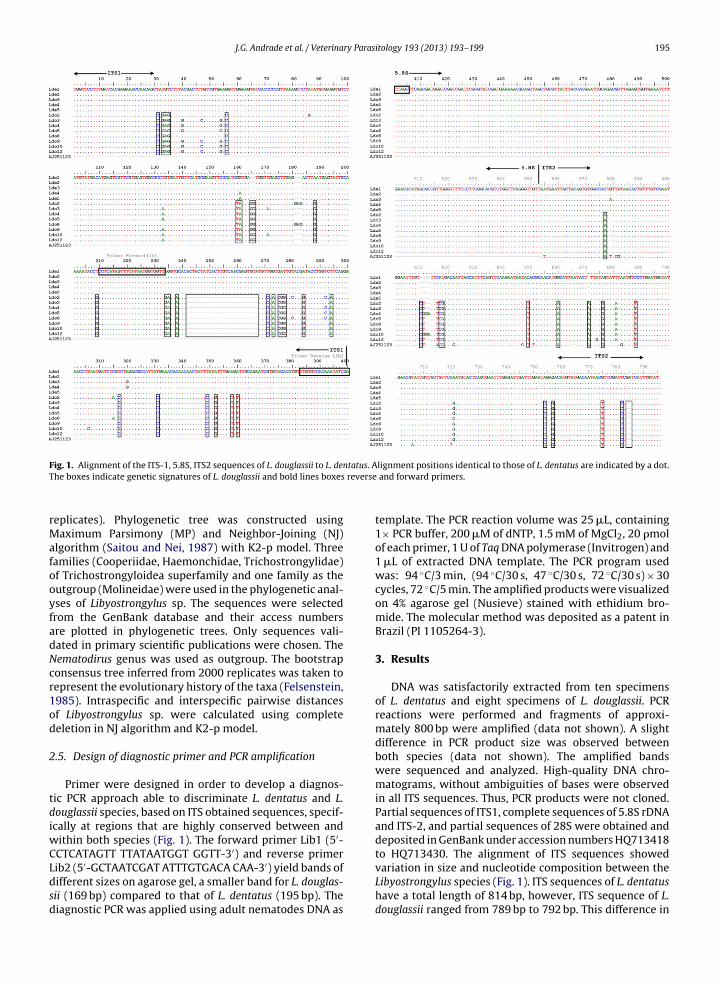
J.G. Andrade et al. / Veterinary Parasitology 193 (2013) 193– 199 195
F tatus. AT reverse
rMafooyfadNcr1od
2
tdiwCLdsd
ig. 1. Alignment of the ITS-1, 5.8S, ITS2 sequences of L. douglassii to L. denhe boxes indicate genetic signatures of L. douglassii and bold lines boxes
eplicates). Phylogenetic tree was constructed usingaximum Parsimony (MP) and Neighbor-Joining (NJ)
lgorithm (Saitou and Nei, 1987) with K2-p model. Threeamilies (Cooperiidae, Haemonchidae, Trichostrongylidae)f Trichostrongyloidea superfamily and one family as theutgroup (Molineidae) were used in the phylogenetic anal-ses of Libyostrongylus sp. The sequences were selectedrom the GenBank database and their access numbersre plotted in phylogenetic trees. Only sequences vali-ated in primary scientific publications were chosen. Theematodirus genus was used as outgroup. The bootstraponsensus tree inferred from 2000 replicates was taken toepresent the evolutionary history of the taxa (Felsenstein,985). Intraspecific and interspecific pairwise distancesf Libyostrongylus sp. were calculated using completeeletion in NJ algorithm and K2-p model.
.5. Design of diagnostic primer and PCR amplification
Primer were designed in order to develop a diagnos-ic PCR approach able to discriminate L. dentatus and L.ouglassii species, based on ITS obtained sequences, specif-cally at regions that are highly conserved between and
ithin both species (Fig. 1). The forward primer Lib1 (5′-CTCATAGTT TTATAATGGT GGTT-3′) and reverse primer
ib2 (5′-GCTAATCGAT ATTTGTGACA CAA-3′) yield bands ofifferent sizes on agarose gel, a smaller band for L. douglas-ii (169 bp) compared to that of L. dentatus (195 bp). Theiagnostic PCR was applied using adult nematodes DNA aslignment positions identical to those of L. dentatus are indicated by a dot. and forward primers.
template. The PCR reaction volume was 25 �L, containing1× PCR buffer, 200 �M of dNTP, 1.5 mM of MgCl2, 20 �molof each primer, 1 U of Taq DNA polymerase (Invitrogen) and1 �L of extracted DNA template. The PCR program usedwas: 94 ◦C/3 min, (94 ◦C/30 s, 47 ◦C/30 s, 72 ◦C/30 s) × 30cycles, 72 ◦C/5 min. The amplified products were visualizedon 4% agarose gel (Nusieve) stained with ethidium bro-mide. The molecular method was deposited as a patent inBrazil (PI 1105264-3).
3. Results
DNA was satisfactorily extracted from ten specimensof L. dentatus and eight specimens of L. douglassii. PCRreactions were performed and fragments of approxi-mately 800 bp were amplified (data not shown). A slightdifference in PCR product size was observed betweenboth species (data not shown). The amplified bandswere sequenced and analyzed. High-quality DNA chro-matograms, without ambiguities of bases were observedin all ITS sequences. Thus, PCR products were not cloned.Partial sequences of ITS1, complete sequences of 5.8S rDNAand ITS-2, and partial sequences of 28S were obtained anddeposited in GenBank under accession numbers HQ713418to HQ713430. The alignment of ITS sequences showed
variation in size and nucleotide composition between theLibyostrongylus species (Fig. 1). ITS sequences of L. dentatushave a total length of 814 bp, however, ITS sequence of L.douglassii ranged from 789 bp to 792 bp. This difference in
196 J.G. Andrade et al. / Veterinary Parasitology 193 (2013) 193– 199
Ldo 3
Ldo1 0
Ldo 9
Ldo 4
Ldo 2
Ldo 8
L.doug lass ii
Lde 5
Lde 1
Lde 3
Lde 4
L.den tatus
Libyostrong ylus sp
Y15880 T.tenu is
Y15875 T.axei
Y15876 T.colub riformis
Y15879 T.ruga tus
Y15877 T.probo lurus
Y15878 T.retortae formis
Y15881 T.vitrinu s
Tricho strong ylus sp
EU084691 H.con tortus
AF044933 O.ostertag i
AF044931 O.lep tospicularis
AJ250659 O.arctica
AF044932 O.mossi
Ostertag ia sp
AF194138 N.batt us
89
97
56
58
100
66
79
92
90
85
99
95
78
76
84
100
54
55
0.05
99
99
99
93
63
52
84
Fig. 2. Phylogenetic tree of the ITS-1 sequences. The evolutionary history was inferred using the MP and NJ algorithm with K2-p model. The percentageootstra
of replicate trees in which the associated taxa clustered together in the bitalic and bolded.
the ITS size between both species is mainly due to the pres-ence of a gap of 26 bp at nucleotide position 242 in all the L.douglassii sequences (Fig. 1). The genetic analysis revealedthat ITS haplotype sequence of Ldo2 and Ldo8 sampleshave an insertion of three nucleotides, GAG, at position 180when compared to the Ldo haplotypes (Fig. 1). An insertionof three nucleotides, GGA and a deletion at nucleotide posi-tion T611 was observed in the ITS2 haplotype of Ldo 4 andLdo10 samples when compared to other Ldo haplotypes(Fig. 1). A set of 21 genetic signatures was identified in L.douglassii when comparing to L. dentatus in the ITS1 region:C31, GAG33, C56, TA160, GG165, G188, G209, GA234, A238,(ACTACTATCACTTGTCAACGAGTGTA) 242 del, C271, A273,CGG275, G284, A294, C317, C331, C349, G352, T358, T360;13 in the ITS2 region: A577, C610, CG617, T649, A660, A671,G676, T688, C754, G757, T775, C782, (GA) 784del; and nonein the 5.8S rDNA gene (Fig. 1).
In the genetic analysis, identical ITS haplotypessequences were omitted and only ITS haplotypes wereanalyzed. L. douglassii ITS sequences (∼765 bp) revealed8 ITS haplotypes with a mean genetic distance of 0.006(K2-p = 0.001–0.013; SE = 0.002). L. dentatus ITS sequences(786 bp) revealed 6 ITS haplotypes with a lower mean
genetic distance of 0.002 (K2-p = 0.001–0.003; SE = 0.001).Between the two species the genetic distance is 0.060(SE = 0.008), demonstrating the evolutionary status ofp test (2000 replicates) is shown next to the branches with MP values in
distinct species. When only ITS1 was analyzed, a meangenetic distance between the species was of 0.085(K2-p = 0.077–0.093; SE = 0.014). Within the 4 ITS hap-lotypes of L. dentatus the genetic distance was 0.004(K2-p = 0.003–0.005; SE = 0.002), and more elevated in the6 ITS haplotypes of L. douglassii, 0.014 (K2-p = 0.003–0.028;SE = 0.004). ITS2 dataset, 2 and 3 ITS haplotypes of L. denta-tus and L. douglassii, respectively, revealed an inter-speciesgenetic distance of 0.056 (K2-p = 0.054–0.059; SE = 0.015).Curiously, the same value of 0.056 (K2-p = 0.054–0.059;SE = 0.016) was observed between the unique L. douglassiisequence available and the L. douglassii from this study.The interspecies genetic distance between L. douglassiiGenBank AJ251123 and L. dentatus was visible higher andvalue 0.094 (SE = 0.015).
Phylogenetic trees based on ITS1 and ITS2 regionswere obtained and evolutionary distances were designed(Figs. 2 and 3). The NJ and MP trees were inferred based onITS1 and ITS2 sequences using 12 and 17 Trichostrongy-loidea sequences, respectively, including one referencesequence of all available species of Trichostrongylus,Haemonchus, Cooperia, and Ostertagia (GenBank; 11/2011).The ITS1 tree revealed that all L. douglassii sequences
grouped in a monophyletic clade with a high bootstrapvalue (NJ = 99%, MP = 99%), and the same topology wasobserved for all L. dentatus haplotypes (NJ bootstrap = 100%,
J.G. Andrade et al. / Veterinary Parasitology 193 (2013) 193– 199 197
X78065 T.axei
X78066 T.retortae formis
Y14817 T.probo lurus
Y14818 T.ruga tus
X78064 T.vitrinu s
X78063 T.colub riformis
X78067 T.tenu is
Tricho strong ylus sp
Lde 1
Lde 3L.den tatus
AJ251123L .doug lassii
Ldo1 2
Ldo 2
Ldo 4
L.doug lassii
Libyostrong ylus sp
X78803 H.con tortus
X78812 H.placeiHae mon chu s sp
AJ000033 C.curticei
X83561 C.on copho ra
X83560 C.pun ctna ta
Coope ria sp
X86026 Te.circumcinta
X86027 O.ostertag i
AJ250657 O.arctica
DQ354334 O.lep tospicularis
Ostertag ia sp
AF194138 N.batt us
100
95
100
70
97
81
99
92
55
100
66
70
97
65
85
51
0.05
87
74
9099
68
99
95
75
F y was ino ootstrapi
MtCpbsLMoclmwsLLswtfOObtdta
ig. 3. Phylogenetic trees of the ITS-2 sequences. The evolutionary historf replicate trees in which the associated taxa clustered together in the btalic and bolded.
P = 99%) (Fig. 2). Both Libyostrongylus species clus-ered in a unique clade (NJ bootstrap = 95%, MP = 99%).lusters genus-specific with strong and moderate sup-ort were detected from genus Trichostrongylus sp. (NJootstrap = 100%, MP = 99%), and Ostertagia sp (NJ boot-trap = 66%, MP = 84%). The closer taxonomic group toibyostrongylus was Trichostrongylus sp. (bootstrap = 76%,P = 63%). The ITS2 NJ tree (Fig. 3) showed a similar topol-
gy as described for ITS1, but with another well-supportedluster (NJ = 100%, MP = 90%) formed only by Libyostrongy-us species All L. dentatus grouped in a strongly supported
onophyletic clade (NJ bootstrap = 100%, MP = 99%), asell as a clade with L. douglassii sequences from this
tudy by NJ method (92%) but moderate with MP (68%).. douglassii GenBank AJ251123 sequence clustered with. douglassii sequences from this study with low boot-trap value (55%) when the NJ method was applied but notith the MP method. Both NJ and MP trees revealed clus-
ers genus-specific supported by strong bootstrap valuesor: Trichostrongylus sp., Haemonchus sp., Cooperia sp., andstertagia sp. (Fig. 3). Teladorsagia circumcincta grouped onstertagia sp. cluster with moderate bootstrap value (NJootstrap = 81%, MP = 85%).The polymorphisms between
he ITS sequences of L. dentatus and L. douglassii allowed theevelopment of a diagnosis approach that discriminatedhese species (Fig. 4A). The potential diagnosis PCR waspplied to a mixture of L. douglassii and L. dentatus DNA,ferred using the MP and NJ algorithm with K2-p model. The percentage test (2000 replicates) is shown next to the branches with MP values in
and two different bands of expected size were observed(Fig. 4B).
4. Discussion
Libyostrongylus is a nematode that infects ostriches(Reinecke, 1983; Bastianello et al., 2005; Andrade et al.,2011a). Although adults (Hoberg et al., 1995; Ederli et al.,2008a) and infective larvae (Ederli et al., 2008b; Andradeet al., 2011b) of L. douglassii and L. dentatus were definedmorphologically as two species, no molecular datum con-firm this statement at the moment. Furthermore, thereis no agreement if this genus should be included in theTrichostrongylidae (Hoberg and Lichtenfels, 1994) or inthe Cooperiidae (Durette-Desset et al., 1999) family. Inaddition, a fast identification method is needed to betterunderstand this infection. In this study, a characteriza-tion of both species based on complete ITS region wasperformed and it was determined that they are genet-ically different. Moreover, both species belong to theTrichostrongylidae family and can be differentiated by amolecular approach.
The ITS region is known to be useful as a target for
species identification, and for inferring phylogenetic rela-tionships between nematodes (Hoste et al., 1998; Audebertet al., 2000; Nadler et al., 2000; Chilton et al., 2001).Here, new complete ITS sequences were obtained for L.
198 J.G. Andrade et al. / Veterinary Parasitology 193 (2013) 193– 199
Fig. 4. A. Schematic representation of complete ITS region with the diagnostic approach proposed in this study to discriminate L. douglassii to L. dentatus.The location and size of the expected PCR products are shown. The white block represents the gap fragment on L. douglassii species and arrows represent thediagnostic primers. B. PCR amplification using diagnostic primers resulted in products with different sizes. Lane 1 and 5: 100 bp DNA ladder (Invitrogen);
tus DNA
lane 2: PCR products of L. douglassii DNA; lane 3: PCR products of L. denta6: negative control. 4% agarose gel (Nusieve) ethidium bromide stained.douglassii and for the first time for L. dentatus species infect-ing ostriches from Brazil. Despite the ITS2 L. douglassiisequence available (Chilton et al., 2001), no molecular char-acterization of the Libyostrongylus genus was conducted.
In this study, the genetic characterization based on thetwo genetic regions supports the evidence of two differentspecies. The interspecies genetic distance based on totalITS and both ITS1 and ITS2 regions, established that thesespecies are each unique genetic entities. The phylogenetictree topologies based on two methods, were also in sig-nificant agreement, demonstrating that L. douglassii andL. dentatus represents a taxonomic unit genetically sep-arated from each other and grouped in a Libyostrongylusgenus clade. In this regard, the analysis of both geneticregions placed Libyostrongylus genus on Trichostrongyli-dae that confirms the taxonomic position proposed beforeby Hoberg and Lichtenfels (1994) based on a morpholog-ical analysis. Moreover, the proximity of Libyostrongylusto Trichostrongylus genus is in agreement with the ITS2phylogeny data of Chilton et al. (2001), using L. douglas-sii AJ251123 and other members of Trichostrongyloideasuperfamily.
Concerning the genetic relationships between L. dou-glassii AJ251123 and Libyostrongylus ITS2 sequences
generated in this study, it is interesting to note that thegenetic distance that discriminates L. douglassii and L. den-tatus is the same between L. douglassii AJ251123 and L.; lane 4: PCR products of mixed DNA of L. douglassii and L. dentatus; lane
douglassii. ITS2 L. douglassii AJ251123 sequence was iso-lated in 2001 from ostriches raised in Australia.
The genetic distance between L. douglassii lineagescould be explained by the process of sub-speciation ofostrich hosts. During the ostriches evolution at leastfour sub-species occurred in distinct regions of theAfrican continent. This sub-species are: Struthio cameluscamelus, S. c. molybdophanes, S. c. massaicus e S. c. aus-tralis (Shanawany and Dingle, 1999). L. douglassii of theseostriches sub-species could co evolved within its host pro-ducing a genetic heterogeneity. With the domesticationof ostriches, the host sub-species interbred generatingostrich sub-species populations with L. douglassii sub-species populations. Thus, with the exportation of ostrichto different continents different subspecies of L. dou-glassii might have been shipped to Brazil and Australia,explaining the heterogeneity found between L. douglas-sii sequences. On the other hand, a very well supportedclade is formed with only L. douglassii from Brazilianostriches. More L. douglassii database from Australia andother worldwide regions is necessary to confirm thishypothesis.
The in silico ITS interspecies polymorphism analysisallowed us to detect a set of genetic signatures that char-
acterized L. douglassii and L. dentatus and also to designa diagnosis PCR for differentiation of both species. Theuse of ITS in this study corroborated the value of this
ry Paras
gc
idsadseuittd
ntmbosd
C
F
cTds(
A
Bam
R
A
A
A
B
J.G. Andrade et al. / Veterina
enetic marker on molecular taxonomy as potential bar-odes allowing genetic species identification.
Ribosomal DNA is a variable multicopy gene familyn organisms which facilitates PCR amplification proce-ures and their use for diagnostic identification. The ITSequences of this work showed marked interspecies vari-bility between both Libyostrongylus species allowing theevelopment of identification method to discriminate thepecies. The possibility of rapid identification of the pres-nce of L. douglassii and/or L. dentatus will allow a betternderstanding of the mixed infection, as well as an increase
n the biology information of these parasites. This will openhe possibility to better understand the epidemiology ofhis mixed infection and the correct use of anthelminthic,ecreasing the ostrich production cost.
L. dentatus and L. douglassii are distinct species phyloge-etically supported based on ITS1 and ITS2 sequences andhey belong to the Trichostrongylidae family, confirming
orphological characterizations. Genetic polymorphismetween L. dentatus and L. douglassii permitted the devel-pment of a molecular approach able to discriminate bothpecies. This is the first genetic characterization of the L.entatus and L. douglassii.
onflict of interest
There are no conflicts of interest.
inancial support
This study was funded by the Brazilian fostering agen-ies Conselho Nacional de Desenvolvimento Científico eecnológico (CNPq), Fundac ão Carlos Chagas Filho do Rioe Janeiro (FAPERJ), Fundac ão de Coordenac ão de Pes-oal de Nível Superior (CAPES) and Instituto Oswaldo CruzIOC/Fiocruz). AMI and RAD are CNPq research fellows.
cknowledgments
The authors would like to thank Thais Wolino da Silvaoa Morte and Lauren Hubert Jaeger for technical helpnd thank Andrèa Carvalho César for proof reading theanuscript.
eferences
ndrade, J.G., Carvalho, E.C.Q., Santos, C.P., DaMatta, R.A., 2011a. Mixedinfection with Libyostrongylus dentatus and Libyostrongylus douglassiiinduces a heterophilic inflammatory infiltrate in the proventriculusof ostriches. Avian Pathol. 40, 367–370.
ndrade, J.G., Lelis, R.T., DaMatta, R.A., Santos, C.P., 2011b. Occurrenceof nematodes and anthelmintic management of ostrich farms fromdifferent Brazilian states: Libyostrongylus douglassii dominates mixedinfections. Vet. Parasitol. 178, 367–370.
udebert, F., Durette-Desset, M.-C., Chilton, N.B., 2000. Internal tran-scribed spacer rDNA can used to infer the phylogenetic relationships
of species within the genus Nematodirus (Nematoda: Molineoidea).Int. J. Parasitol. 30, 187–191.astianello, S., McKenna, P.B., Hunter, J., Julian, A.F., 2005. Clinical andpathological aspects of Libyostrongylus infection in ostriches. Surveil-lance 32, 3–6.
itology 193 (2013) 193– 199 199
Bellocq, J.G., Ferté, H., Depaquit, J., Justine, J.-L., Tillier, A., Durette-Desset, M.-C., 2001. Phylogeny of the Trichostrongylina (Nematoda)inferred from 28S rDNA sequences. Mol. Phylogenet. Evol. 19,430–442.
Chilton, N.B., 2004. The use of nuclear ribosomal DNA markers for theidentification of bursate nematodes (order Strongylida) and for thediagnosis of infections. Anim. Health Res. Rev. 5, 173–187.
Chilton, N.B., Newton, L.A., Beveridge, I., Gasser, R.B., 2001. Evolu-tionary relationships of trichostrongyloid nematodes (Strongylida)inferred from ribosomal DNA sequence data. Mol. Phylogenet. Evol.19, 367–386.
Cobbold, T.S., 1882. New entozoon from the ostrich. J. Linnean Soc. 16,184–188.
Craig, T.M., Diamond, P.L., 1996. Parasites of ratites. In: Tully, T.N., Shane,S.M. (Eds.), Ratites Management, Medicine, and Surgery. Kreiger Pub-lishing Company, Malabar, pp. 115–126.
Cutillas, C., de Rojas, M., Ariza, C., Ubeda, J.M., Guevara, D.C., 2007. Molec-ular identification of Trichuris vulpis and Trichuris suis isolated fromdifferent hosts. Parasitol. Res. 100, 383–389.
Durette-Desset, M.C., Hugot, J.P., Darlu, P., Chabaud, A.G., 1999. A cladisticanalysis of the Trichostrongyloidea (Nematoda). Int. J. Parasitol. 29,1065–1086.
Ederli, N.B., Bonadiman, S.F., Moraes Neto, A.H.A., DaMatta, R.A., San-tos, C.P., 2008a. Mixed infection by Libyostrongylus douglassii and L.dentatus (Nematoda: Trichostrongylidae) in Struthio camelus (Ratites:Struthioniformes) from Brazil with further morphological character-ization of adults. Vet. Parasitol. 151, 227–232.
Ederli, N.B., Oliveira, F.C.R., Lopes, C.W.G., DaMatta, R.A., Santos, C.P.,Rodrigues, M.L.A., 2008b. Morphological diagnosis of infective larvaeof Libyostrongylus douglassii (Cobbold, 1882) Lane, 1923 and L. denta-tus Hoberg, Lloyd and Omar, 1995 (Nematoda: Trichostrongylidae) ofostriches. Vet. Parasitol. 155, 323–327.
Felsenstein, J., 1985. Confidence limits on phylogenies: an approachingusing the bootstrap. Evolution 29, 783–791.
Hillis, D.M., Dixon, M.T., 1991. Ribosomal DNA: molecular evolution andphylogenetic inference. Q. Rev. Biol. 66, 441–453.
Hoberg, E.P., Lichtenfels, J.R., 1994. Phylogenetic systematic analysis of theTrichostrongylidae (Nematoda), with an initial assessment of coevo-lution and biogeography. Parasitology 80, 976–996.
Hoberg, E.P., Lloyd, S., Omar, H., 1995. Libyostrongylus dentatus n. sp.(Nematoda: Trichostrongylidae) from ostriches in North America,with comments on the genera Libyostrongylus and Paralibyostrongylus.J. Parasitol. 81, 85–93.
Hoste, H., Chilton, N.B., Beveridge, I., Gasser, R.B., 1998. A comparison of thefirst internal transcribed spacer of ribosomal DNA in seven species ofTrichostrongylus (Nematoda: Trichostrongylidae). Int. J. Parasitol. 28,1251–1260.
Kimura, M., 1980. A simple method for estimating evolutionary rateof base substitutions through comparative studies of nucleotidesequences. J. Mol. Evol. 16, 111–116.
Lichtenfels, J.R., Hoberg, E.P., Zarlenga, D.S., 1997. Systematics of gas-trointestinal nematodes of domestic ruminants: advances between1992 and 1995 and proposals for future research. Vet. Parasitol. 72,225–245.
Nadler, S.A., Hoberg, E.P., Hudspeth, D.S.S., Rickard, L.G., 2000. Rela-tionships of Nematodirus species and Nematodirus battus isolates(Nematoda: Trichostrongyloidea) based on nuclear ribosomal DNAsequences. J. Parasitol. 86, 588–601.
Reinecke, R.K., 1983. Veterinary Helminthology. Butterworth, Durban, 295pp.
Saitou, N., Nei, M., 1987. The neighbor-joining method: a new method forreconstructing phylogenetic tree. Mol. Biol. Evol. 4, 406–425.
Santos, C.P., Andrade, J.G., DaMatta, R.A., 2010. An update onLibyostrongylus, a gastro-intestinal nematode of ostriches. In: Lamann,G.V. (Ed.), Veterinary Parasitology. Nova Science Publishers, USA,pp. 179–192.
Shanawany, M.M., Dingle, J., 1999. Ostrich Production Systems. Food andAgriculture Organizations of the United Nations, 256 pp.
Tamura, K., Dudley, J., Nei, M., Kumar, S., 2007. MEGA4: Molecular Evo-lutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol.
Evol. 24, 1596–1599.Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W: improvingthe sensitivity of progressive multiple sequence alignment throughsequence weighting, position specific gap penalties and weight matrixchoice. Nucleic Acids Res. 22, 4673–4680.





























