Embed Size (px)
Citation preview

Genetic Influences on Cellular Reactionsto CNS Injury: The Reactive Responseof Astrocytes in Denervated NeuropilRegions in Mice Carrying a Mutation
(WldS) That Causes DelayedWallerian Degeneration
OSWALD STEWARD1,2* AND PATRICIA A. TRIMMER1,3
1Department of Neuroscience, University of Virginia Health Sciences Center,Charlottesville, Virginia 22908
2Department of Neurosurgery, University of Virginia Health Sciences Center,Charlottesville, Virginia 22908
3Department of Neurology, University of Virginia Health Sciences Center,Charlottesville, Virginia 22908
ABSTRACTThis study compares the reactive changes in astrocytes in denervated neuropil regions in
normal mice and in mice carrying the WldS mutation which leads to delayed Walleriandegeneration. In situ hybridization and immunocytochemical techniques were used to definethe time course of changes in the levels of glial fibrillary acidic protein (GFAP) and GFAPmRNAin the denervated neuropil of the hippocampus after unilateral aspiration lesions of theentorhinal cortex. In control mice, GFAP mRNA levels increased rapidly in the denervatedneuropil to a peak that was about tenfold higher than control at 2–4 days, decreased between 6and 8 days postlesion, and then increased again to a second peak at 10 days postlesion. In-creases in immunostaining for GFAP were evident by 2 days, remained elevated until 12 dayspostlesion and then decreased slowly. In mice carrying theWldSmutation, the upregulation ofGFAP mRNA levels in the denervated laminae was substantially delayed. Strikingly absentwas the dramatic increase in labeling at 2–4 days postlesion which was such a prominentfeature of the response in control animals. Peak labeling in the denervated laminae was notseen until 10–12 days postlesion. The development of a well-defined band of intenselyimmunostained and hypertrophied astrocytes in the denervated zone was also delayed in theWldS animals, although there were modest increases in immunostaining as early as 2 dayspostlesion that were seen throughout the hippocampus ipsilateral to the lesion. These resultssuggest that degenerative changes in axons and synaptic terminals are the principal triggerfor upregulating GFAP expression in the denervated neuropil, although other signals alsoplay a role in the early postlesion response. J. Comp. Neurol. 380:70–81, 1997.r 1997 Wiley-Liss, Inc.
Indexing terms: mutant mice; GFAP; immunocytochemistry; hippocampus; regeneration
This paper and its companion (Schauwecker and Stew-ard, 1997) take advantage of a naturally occurring muta-tion (termed WldS) to begin to unravel the signals thattrigger reactive responses of non-neuronal cells in dener-vated neuropil zones. These responses are of interestbecause they are thought to play an important role in theprocess of synapse reorganization that occurs after injury.Mice carrying the WldS mutation exhibit a remarkable
delay in the onset of Wallerian degeneration of bothperipheral and central axons (Perry et al., 1990a,b, 1991;
Steward, 1992). The evidence that is available to dateindicates that the mutation somehow affects intracellularsignalling cascades within damaged axons that trigger the
Contract grant sponsor: NIH; Contract grant number: NS29875.*Correspondence to: Oswald Steward, Department of Neuroscience and
Neurosurgery, University of Virginia Health Sciences Center, Charlottes-ville, VA 22908. E-mail: [email protected] 9 November 1994; Revised 11 October 1996; Accepted 15
November 1996
THE JOURNAL OF COMPARATIVE NEUROLOGY 380:70–81 (1997)
r 1997 WILEY-LISS, INC.

processes of self-destruction that lead eventually to axonaldisintegration (Buckmaster et al., 1995). Because theprocess of degeneration is delayed, there are delays in therecruitment of polymorphonuclear cells and macrophagesto sites of damage in peripheral nerves or the centralnervous system (CNS; Fujuki et al., 1996; Schauweckerand Steward, 1997, companion paper), as well as delays inthe activation of microglia in areas that contain degenerat-ing axons or synaptic terminals (Fujuki et al., 1996;Schauwecker and Steward, 1997, companion paper).The present experiments focused on one model system
that has been used extensively for studies of lesion-induced synaptic plasticity, the hippocampal dentate gyrusfollowing unilateral lesions of the entorhinal cortex. Thishas been an important system for studies of postlesionsynaptic reorganization because of the extent of the post-lesion growth. Following lesions of the entorhinal cortex,about 90% of the synaptic terminals in the outer portion ofthe molecular layer of the dentate gyrus degenerate. Overtime, these synapses are replaced as a result of thesprouting of several afferent systems, which results in anearly complete reinnervation of the dentate granule cells(for a review, see Steward, 1989).A number of recent studies have characterized the
responses of non-neuronal cells in the denervated dentategyrus during lesion-induced synapse turnover; these re-sults have provided evidence that microglia and astrocytesmay play an important role in the reinnervation process.Within 1–2 days after the injury, microglia in the dener-vated neuropil become activated and upregulate theirexpression of complement receptors and various cytokines(for recent references, see Schauwecker and Steward,1997). During the same time period, astrocytes exhibit‘‘reactive’’ responses which involve a dramatic upregula-tion of gene expression followed by hypertrophy (Faganand Gage, 1990, 1994; Steward et al., 1990). In the reactivestate, astrocytes play the key role in removing degenerat-ing synaptic terminals from postsynaptic cells (Matthewset al., 1976a,b; Raisman, 1969). Reactive astrocytes mayalso produce neuronotrophic factors (Furukawa et al.,1986; Houlgatte et al., 1989; Ip et al., 1993) and/orextracellular matrix molecules that play a role in inducingor promoting the postlesion sprouting response (McKeonet al., 1991).An important issue is defining the relationship between
the different reactive processes. Of particular interest iswhether the responses are in sequence or in parallel. Inthis regard, the timing of the responses of microglia andastrocytes, together with the fact that activated microgliarelease a variety of cytokines that are capable of inducingreactive changes in astrocytes, has led to the hypothesisthat the response of astrocytes is triggered via signals fromactivated microglia (Fagan and Gage, 1990). However, analternative possibility is that the responses of microgliaand astrocytes are triggered independently. For example,intense physiological activity (seizures or spreading depres-sion) can induce changes in astroglial gene expression thatmimic the initial stages of the reactive response. Specifi-cally, even a single seizure or a few episodes of spreadingdepression lead to dramatic increases in glial fibrillaryacidic protein (GFAP) expression (Bonthius and Steward,1993; Kraig et al., 1991; Steward, 1994; Steward et al.,1991). The induction of GFAP expression that occurs afterelectroconvulsive seizures is not associated with detect-able neuronal damage or microglial activation (Schau-
wecker and Steward, unpublished observations). More-over, recent studies have revealed that increases in GFAPmRNA levels in nondenervated zones of the hippocampusfollowing electrolytic entorhinal cortex lesions are due inpart to recurrent limbic seizures that occur during the first24 hours after the lesion (Kelley and Steward, 1996a,b).These results have led to the hypothesis that the reactiveresponse of astrocytes may be a composite response trig-gered by different signals operating in parallel, includingsignals released from degenerating synaptic terminals oraxons, factors released by activated microglia; and physi-ological events that occur as a consequence of the lesion.To define the relative role of different potential signals, it
is important to design experiments that allow one todissociate the different candidate signals. TheWldS muta-tion provides an interesting opportunity in this regard;indeed, the mutation provides a setting in which to testspecific hypotheses. In previous studies, we demonstratedthat the onset of terminal degeneration and the activationof microglia within denervated neuropil zones is substan-tially delayed in animals carrying the WldS mutation(Schauwecker and Steward, 1997; Steward, 1992). Theseresults lead to specific predictions regarding the activationof astrocytes. In particular, components of the reactiveresponse of astrocytes that are triggered by signals fromactivated microglia should not occur in WldS mutantanimals until after microglial activation. Components ofthe reactive response that are directly triggered by degen-erative changes in axons or synaptic terminals should alsobe substantially delayed in the mutant animals, althoughnot necessarily delayed to the same extent as the delay inmicroglial activation. Finally, components of the reactiveresponse that are triggered in other ways (for example, asa result of episodes of spreading depression induced by thelesion or postlesion seizures) should occur with the sametime course in normal andWldSmutant animals.The present study evaluates these predictions by compar-
ing the reactive response of astrocytes in the denervatedneuropil of the hippocampus of normal and mutant micefollowing unilateral lesions of the entorhinal cortex. Theresults indicate that multiple signals regulate the reactiveresponse of astrocytes following injury, and that the re-sponse that occurs in areas containing degenerating axonsand synaptic terminals is substantially delayed in animalscarrying theWldSmutation.
MATERIALS AND METHODS
Experimental animals
Experimental animals were of two types: (1) C57BL/6-WldS mice obtained from Olac (Bicester, Oxfordshire,England). These animals carry theWallerian degenerationdelayed (WldS) mutation; and (2) C57BL/6 mice obtainedfrom Harlan Sprague-Dawley Inc. (Indianapolis, IN). Thelatter are control mice in which degeneration occurs with atime course that is comparable to other species. Most of theWldS animals used for these studies were the first genera-tion of a breeding colony established at our facility.
Surgery
The entorhinal cortex (EC) and portions of the surround-ing retrohippocampal cortex were removed unilaterally byaspiration as described in Schauwecker and Steward(1997).
GENETIC INFLUENCES ON REACTIVE ASTROCYTOSIS 71

For the in situ hybridization analyses of GFAP mRNAlevels, animals were allowed to survive for 2, 4, 6, 8, 10, 12,14, and 21 days postlesion (two to four animals per timepoint; for details, see Results). The total number of ani-mals per group varied depending on the number of ani-mals that were eliminated due to lesions that were eitherincomplete or that extended into the hippocampus. Threeunoperated (control) mice of each substrain were alsoprepared. Separate groups of mutant and control animalsreceived EC lesions and were prepared for GFAP immuno-cytochemistry at 2, 4, 8, 12, and 20 days postlesion (n 5 3per time point). At the appropriate postlesion interval,animals were euthanized with an overdose of Avertin andperfused with 4% paraformaldehyde in 0.1 M phosphatebuffer (pH 7.3).The protocols for probe preparation, in situ hybridiza-
tion, and immunocytochemistry are the same as in ourprevious studies in rats (Steward et al., 1990, 1993). Theseprotocols are summarized briefly below.
In situ hybridization
For in situ hybridization, brains were removed andplaced in 4% paraformaldehyde in 0.1 M phosphate bufferovernight. They were then transferred to 30% sucrose for24 hours. Brains were sectioned on a cryostat and sectionswere thaw-mounted onto poly-lysine-coated RNAse freemicroscope slides. Sections were stored at 280°C prior toin situ hybridization.In situ hybridization was carried out using a cRNA
probe for GFAP mRNA that has previously been described(Steward et al., 1990). The cRNAprobe was derived from a2.5 kilobase cDNA clone for mouse GFAP (Lewis et al.,1984). A 1.26 kilobase fragment from the 58 portion of thisclone was recloned into the HindIII site of a BluescriptM13-vector by D. Chikaraishi (Tufts University), and theplasmid was provided to us as a gift. 35S-labeled antisensecRNA probes of about 1,000 bases in length were synthe-sized from linearized plasmid using the T3 promoter aspreviously described (Steward et al., 1990). Control experi-ments documenting the specificity of the probe have beendescribed previously (Steward et al., 1990).Slides were postfixed in 4% paraformaldehyde, treated
with proteinase K (1 µg/ml in RNAse buffer for 30 min-utes), washed in 0.53 standard saline citrate (SSC) anddried. The slides were placed flat in humidified petridishes, and the sections were covered with 100 µl ofhybridization buffer and prehybridized for 2 hours. Then20 µl of additional hybridization buffer containing 1 µl ofprobe (0.5–1 3 106 cpm) and 2 µg of tRNAwas added and acoverslip was placed over the section. Sections were hybrid-ized overnight at 55°C in humidified Petri dishes. Theslides were then washed in 0.53 SSC, treated with RNAsefor 30 minutes, washed for 2 hours at 55°C in a stringencybuffer (0.13 SSC, 10 mM b mercaptoethanol (BME) 1 mMEDTA), dehydrated through ethanols, and dried. Sectionswere exposed to photographic film to obtain film autoradio-graphs and then dipped in Kodak NTB-2 photographicemusion. The slides were exposed for 1 week, developed inD19, and stained with cresyl violet.For quantitative analyses, grain counts were carried out
on the emulsion-coated slides using a Gould IP8500 imageanalysis system and software developed by the BiomedicalImage Processing Center at the University of Virginia.Slides were selected for quantitative analysis that did notexhibit obvious histological artifacts or other technical
artifacts. Grain density was evaluated over the outer halfof the molecular layer of the dentate gyrus using a squaremeasuring window that was 43 3 43 µm. Three indepen-dent measurements were made on each slide, and fiveslides were evaluated for each animal. Counts were alsocarried out in the thalamus and rostral cerebral cortex toprovide a measure of grain density in control regions. Inorder to control for differences in the overall level ofhybridization between slides and cases, data were normal-ized by expressing the counts as an ipsi/contralateral (I/C)ratio of grain density and the average I/C ratio for eachanimal was then calculated. In this way, each slide servesas its own control. Data were analyzed by Kruskal-Wallisone-wayANOVA (on ranks).It is important to note that the contralateral side could
be used as an intra-animal control because the mice didnot exhibit the bilateral increases in GFAP expressionwhich occur to a variable degree in rats at 2 days postle-sion, and which appear to be a result of postlesion seizures(Kelley and Steward, 1996b). As noted further in theDiscussion, the difference in responses between mice andrats may be due to differences in the method of lesionproduction (aspiration lesions in mice vs. electrolytic le-sions in rats); the latter have been shown to result inpostlesion seizures due to the deposition of iron at thelesion site.
Measurement of GFA protein byimmunocytochemistry
For GFAP immunostaining, animals were perfused with4% paraformaldehyde as described above and the brainswere placed in fixative overnight. The brains were thensectioned in the horizontal plane using an Oxford Vibra-tomet (section thickness was 40 µm) and sections werecollected in 0.1 M phosphate buffer (pH 7.4). The sectionswere washed thoroughly in 0.1 M phosphate buffer prior toimmunostaining.Sections were immunostained by the avidin-biotin per-
oxidase complex, the ‘‘ABC method,’’ by using a commer-cial anti-GFAP antibody (Dako), and the Vectastain R kit.Sections were incubated in 0.5% Triton X-100, 0.25% lCarrageenan in 0.1 M phosphate buffer (pH 7.4) for 2hours at room temperature to block nonspecific staining,and then reacted for 18 hours at 5°C in the same solutionwith the primary antibody (at a dilution of 1:500). Thesections were rinsed 43 in 0.05 M Tris buffer (pH 7.6), andthen incubated in the 0.5% Triton X-100, 0.25% l Carra-geenan solution with a goat antirabbit secondary antibody(at a dilution of 1:100), together with 5% normal goatserum. The sections were reacted in the avidin-biotinsolution for 30 minutes according to the protocol providedby the manufacturer, washed in Tris buffer, and thenreacted for 10 minutes in 0.1 M phosphate buffer (pH 7.4)containing diaminobenzidine (0.5mg/ml), ammonium chlo-ride (0.4 mg/ml), glucose oxidase (40 µg/ml), and 0.2%beta-D-glucose. The sections were mounted on microscopeslides, dehydrated through xylene, and coverslipped withDPX mounting medium. Immunocytochemical controlsinvolved omission of the primary antibody, in which caseimmunostaining was abolished.Quantitative analyses of the level of immunostaining
were carried out using an MCID system using the sameapproach as described in the companion paper. The sec-tions were viewed using a 163 objective and a rectangularmeasuring window ranging from 50 µm 3 300 µm to
72 O. STEWARD AND P.A. TRIMMER

70 µm 3 300 µm. The image was digitized and thethreshold was adjusted manually so that immunostainedcells and processes were above threshold and unstainedareas were below threshold. The threshold was set for thecontrol side on each section, and then the same thresholdsetting was used for the side ipsilateral to the lesion. Thepercentage area of the tissue that was occupied by immu-nostained cells and processes was determined taking fivemeasurements per section and analyzing five to ten sec-tions per animal. We then calculated the ipsilateral/contralateral (I/C) ratio of the measure, which provides aninternal control for section-to-section variability ofimmunostaining.
RESULTS
To interpret the patterns of increased labelling forGFAP, it is important to recall the organization of theprojections from the entorhinal cortex to the hippocampalformation. The basic organization of the projections inmice is similar to what has been described in other species(Steward, 1992). The projections to the dentate gyrusterminate in the outer two-thirds of the stratum molecu-lare. The projections to CA1 and CA3 terminate in thestratum lacunosm-moleculare and stratum moleculare,respectively. The projections to the dentate gyrus and CA3are almost exclusively ipsilateral, whereas the projectionto CA1 is bilateral. Thus, the only substantial crossedprojection to the hippocampus is the one that terminatesin the stratum lacunosum-moleculare of the CA1 region.There is, however, a small crossed projection from theentorhinal area that terminates in a triangular-shapedpatch in the molecular layer of the subiculum. Thisprojection from the entorhinal area to the subiculum hasnot been previously described to the best of our knowledgebut can be seen in illustrations from previous tract tracingstudies in rats (see for example Steward, 1980). Lesions ofthe entorhinal cortex lead to the appearance of terminaldegeneration in all of these areas.
Time course of increases in GFAP expressionas revealed by in situ hybridization
Figure 1 illustrates the pattern of labeling in the hippo-campus following hybridization with the probe for GFAPmRNA in C57BL/6 control mice 2, 4, 10, and 20 days fol-lowing unilateral EC lesions. The pattern of the increasedGFAP expression was comparable to what has been previ-ously described in rats (Steward et al., 1990). At 2 dayspostlesion, GFAP mRNA levels were dramatically in-creased in the dentate gyrus and hippocampus ipsilateralto the lesion, with the greatest increases occurring in thezones that are innervated by the entorhinal cortex (Fig.1A). However, there were also general increases in labelingthroughout the hippocampus on the ipsilateral side, espe-cially in the CA1 region. In contrast to what is seen afterelectrolytic lesions of the EC in rats, none of the miceexhibited the transient increases in labeling in the dentategyrus on the side contralateral to the lesion (Fig. 1B).At 4 days postlesion, the levels of labeling were higher,
and the labeling was primarily localized in the areas thatwould contain degeneration debris, specifically, the por-tions of the dentate gyrus and hippocampus that receiveprojections from the EC and the areas surrounding thelesion site (Fig. 1C). There was also a patch of intenselabeling in the hippocampus contralateral to the lesion in
the area of termination of the crossed temporoammonicpathway in the stratum lacunosum-moleculare of CA1 (seeFig. 1D, arrow) and another patch of increased labeling inthe molecular layer of the subiculum corresponding to theterminal field of the crossed projection that terminates inthis site.The pattern of labeling was essentially similar at 4, 6, 8,
10, and 12 days postlesion (see, for example, Fig. 1E whichillustrates the pattern at 10 days). Throughout this period,there were high levels of labeling over the denervatedzones in the dentate gyrus, CA1, and CA3 regions. At 20days postlesion, GFAP mRNA levels were still elevated,but not to the same extent as at earlier intervals (Fig. 1F).Quantitative analyses of grain density in the outer
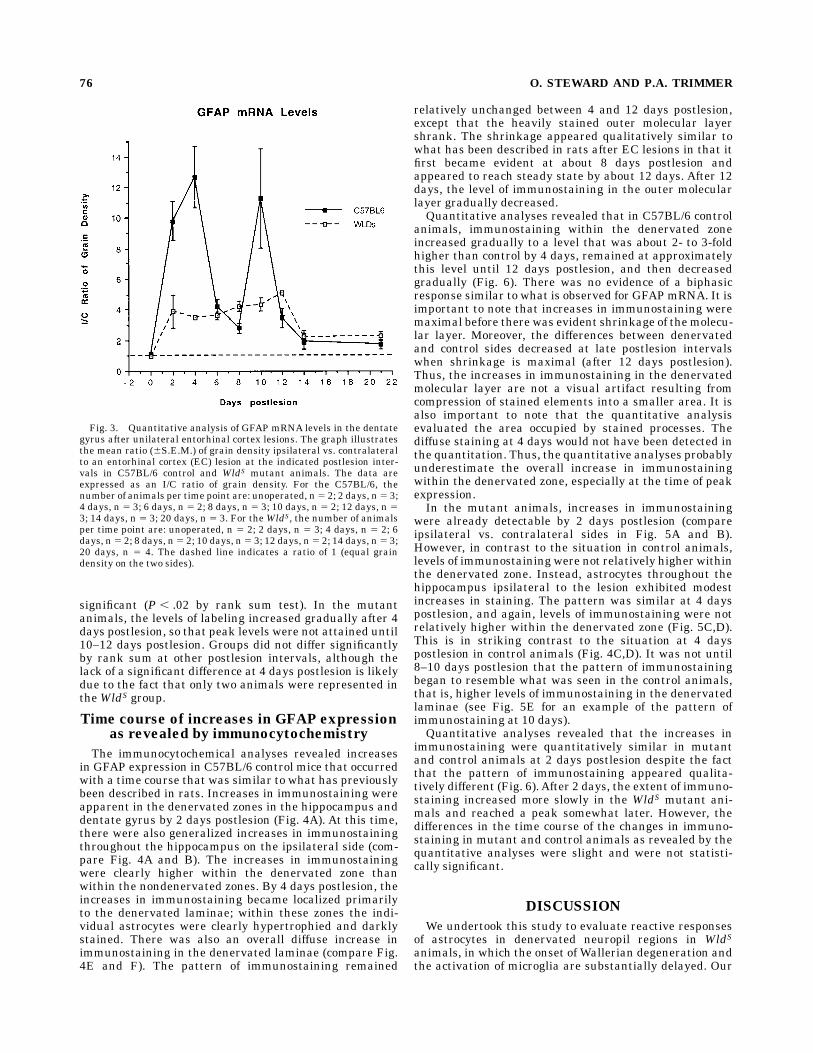
molecular layer of the dentate gyrus revealed that peaklevels of labeling occurred at 2–4 days postlesion (Fig. 3).After the prominent peak at 2–4 days postlesion, levels oflabeling declined to a plateau that was about 2- to 3-foldhigher than control levels and then increased again to asecond peak at 10 days postlesion. This biphasic change inGFAPmRNA levels after EC lesions is similar to what hasbeen described in rats (Steward et al., 1990). Despitedifferences in the levels of labeling, the pattern of labelingat 6 and 8 days was virtually indistinguishable from thepattern at 10 days, which is illustrated in Figure 2E.In the WldS mutant mice, there were detectable in-
creases in GFAP mRNA levels as early as 2 days postle-sion. Labeling was especially high near the lesion site, butthere was also increased labeling within the hippocampus(Fig. 2A). Strikingly absent, however, was the relativelyhigher labeling within the denervated laminae. Instead,there was a generalized increase in labeling throughoutthe hippocampus and dentate gyrus. Thus, the overallpattern of labeling in the denervated hippocampus wasquite different than what was seen in the control animals.The pattern of labeling was essentially the same at day 4
postlesion (Fig. 2C,D), except that in the case illustrated,the overall level of labeling was somewhat lower, so thatthe increased labeling adjacent to the lesion site is lessapparent. Despite the fact that the overall level of labelingin this case was relatively less, the pattern of labeling withrespect to the denervated laminae was clearly apparent.Most importantly, it was clear that there was no indicationof relatively increased labeling within the denervated zone.The pattern in which there was relatively higher levels
of labeling within the denervated laminae began to appearat 6 days postlesion and was more evident by 8 days.However, at these time points, the labeling had a patchydistribution (not shown). By 10 days postlesion, the pat-tern of labeling resembled what was seen at 4 dayspostlesion in control animals in that levels of labeling wereclearly elevated in all of the areas that are innervated bythe entorhinal cortex (Fig. 3E). Levels of labeling weresomewhat higher at 12 days postlesion, and remainedelevated at 20 days postlesion (Fig. 3F).The quantitative analyses revealed clear differences in
the overall time course of the changes in GFAP mRNAlevels in C57BL/6 controls vs.WldS mutant animals (over-all comparison of groups across days 5 P , .003 byKruskal-Wallis ANOVA). The most obvious difference wasthe absence of the large peak in GFAPmRNA levels at 2–4days postlesion in the mutant animals. At 2 days postle-sion, levels of labeling in the denervated zone were about4-fold higher in the mutant animals vs. 10-fold in theC57BL/6 control mice. This difference was statistically
GENETIC INFLUENCES ON REACTIVE ASTROCYTOSIS 73
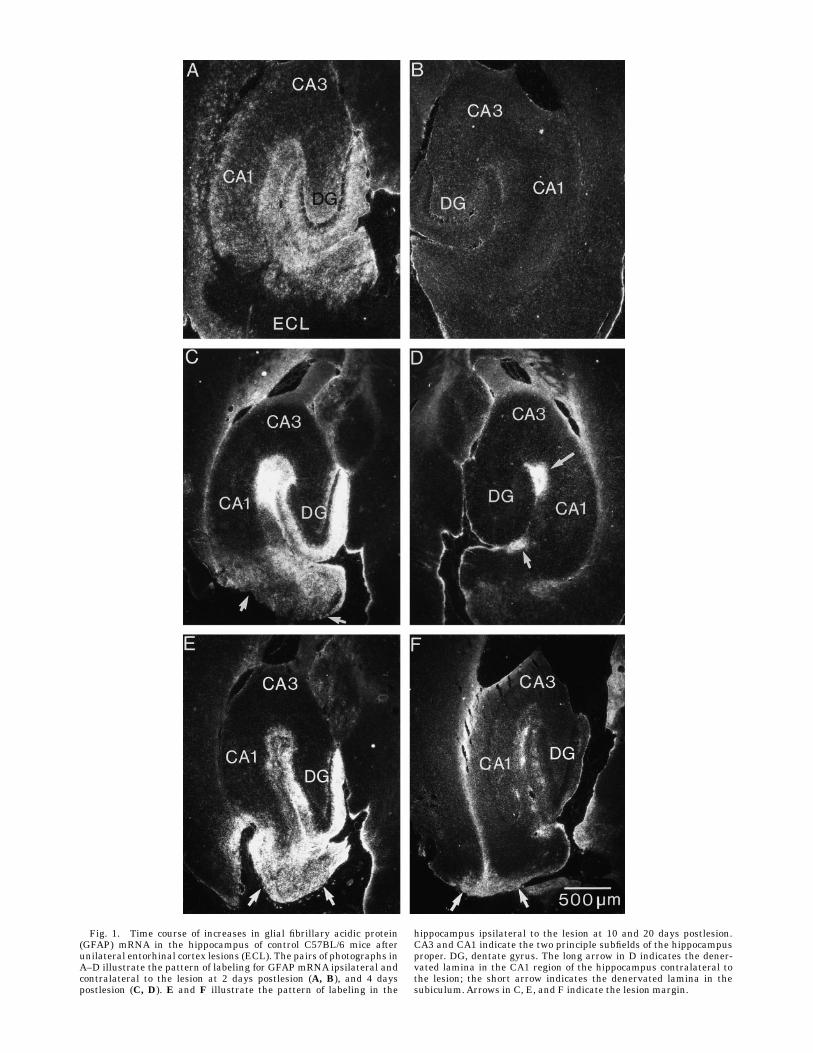
Fig. 1. Time course of increases in glial fibrillary acidic protein(GFAP) mRNA in the hippocampus of control C57BL/6 mice afterunilateral entorhinal cortex lesions (ECL). The pairs of photographs inA–D illustrate the pattern of labeling for GFAP mRNA ipsilateral andcontralateral to the lesion at 2 days postlesion (A, B), and 4 dayspostlesion (C, D). E and F illustrate the pattern of labeling in the
hippocampus ipsilateral to the lesion at 10 and 20 days postlesion.CA3 and CA1 indicate the two principle subfields of the hippocampusproper. DG, dentate gyrus. The long arrow in D indicates the dener-vated lamina in the CA1 region of the hippocampus contralateral tothe lesion; the short arrow indicates the denervated lamina in thesubiculum.Arrows in C, E, and F indicate the lesion margin.
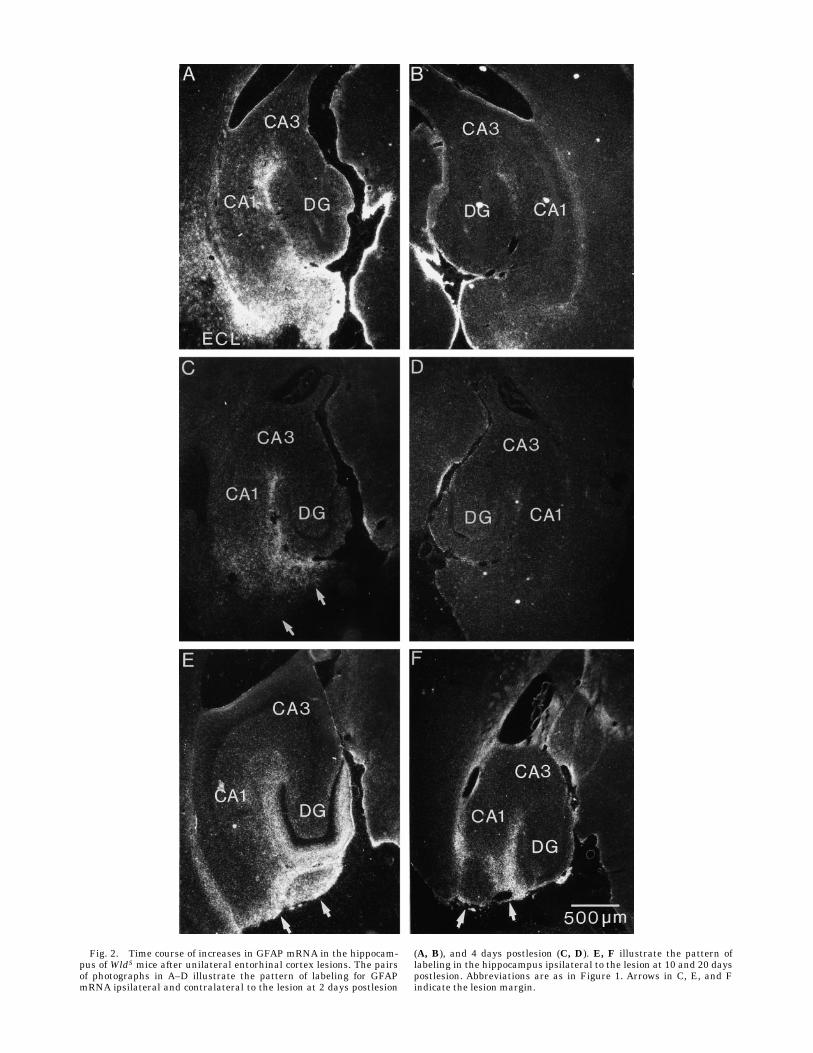
Fig. 2. Time course of increases in GFAP mRNA in the hippocam-pus of WldS mice after unilateral entorhinal cortex lesions. The pairsof photographs in A–D illustrate the pattern of labeling for GFAPmRNA ipsilateral and contralateral to the lesion at 2 days postlesion
(A, B), and 4 days postlesion (C, D). E, F illustrate the pattern oflabeling in the hippocampus ipsilateral to the lesion at 10 and 20 dayspostlesion. Abbreviations are as in Figure 1. Arrows in C, E, and Findicate the lesion margin.
significant (P , .02 by rank sum test). In the mutantanimals, the levels of labeling increased gradually after 4days postlesion, so that peak levels were not attained until10–12 days postlesion. Groups did not differ significantlyby rank sum at other postlesion intervals, although thelack of a significant difference at 4 days postlesion is likelydue to the fact that only two animals were represented intheWldS group.
Time course of increases in GFAP expressionas revealed by immunocytochemistry
The immunocytochemical analyses revealed increasesin GFAP expression in C57BL/6 control mice that occurredwith a time course that was similar to what has previouslybeen described in rats. Increases in immunostaining wereapparent in the denervated zones in the hippocampus anddentate gyrus by 2 days postlesion (Fig. 4A). At this time,there were also generalized increases in immunostainingthroughout the hippocampus on the ipsilateral side (com-pare Fig. 4A and B). The increases in immunostainingwere clearly higher within the denervated zone thanwithin the nondenervated zones. By 4 days postlesion, theincreases in immunostaining became localized primarilyto the denervated laminae; within these zones the indi-vidual astrocytes were clearly hypertrophied and darklystained. There was also an overall diffuse increase inimmunostaining in the denervated laminae (compare Fig.4E and F). The pattern of immunostaining remained
relatively unchanged between 4 and 12 days postlesion,except that the heavily stained outer molecular layershrank. The shrinkage appeared qualitatively similar towhat has been described in rats after EC lesions in that itfirst became evident at about 8 days postlesion andappeared to reach steady state by about 12 days. After 12days, the level of immunostaining in the outer molecularlayer gradually decreased.Quantitative analyses revealed that in C57BL/6 control
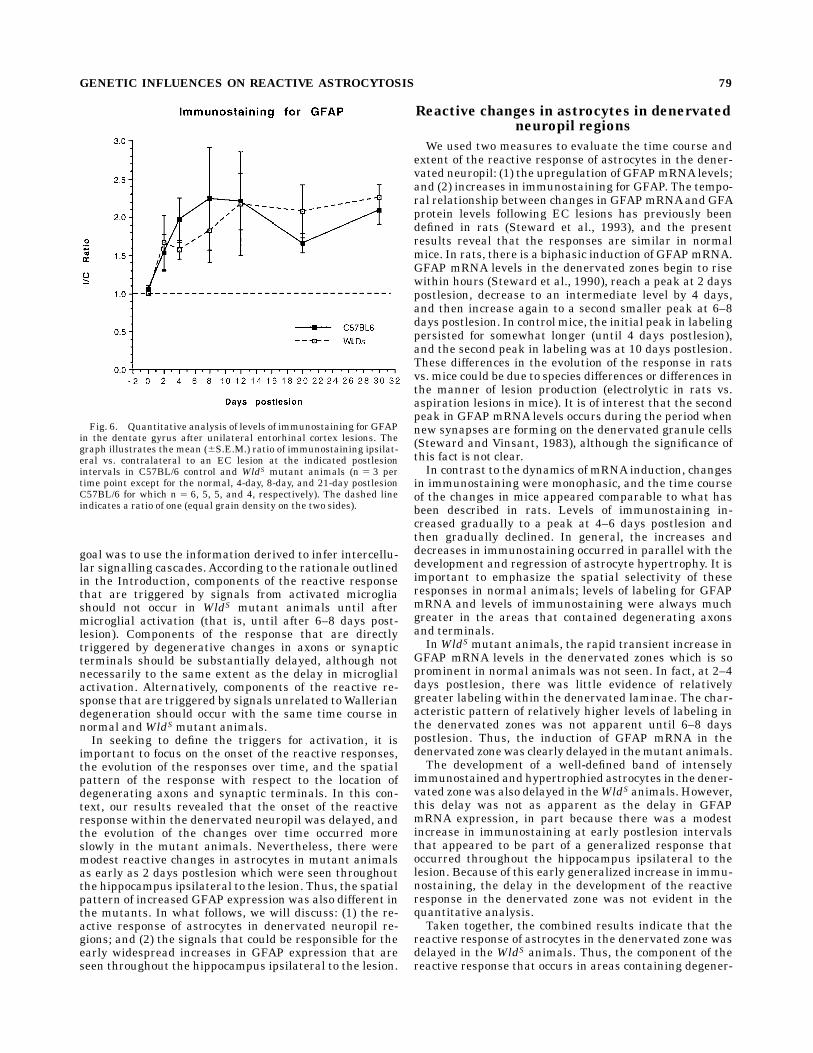
animals, immunostaining within the denervated zoneincreased gradually to a level that was about 2- to 3-foldhigher than control by 4 days, remained at approximatelythis level until 12 days postlesion, and then decreasedgradually (Fig. 6). There was no evidence of a biphasicresponse similar to what is observed for GFAPmRNA. It isimportant to note that increases in immunostaining weremaximal before there was evident shrinkage of themolecu-lar layer. Moreover, the differences between denervatedand control sides decreased at late postlesion intervalswhen shrinkage is maximal (after 12 days postlesion).Thus, the increases in immunostaining in the denervatedmolecular layer are not a visual artifact resulting fromcompression of stained elements into a smaller area. It isalso important to note that the quantitative analysisevaluated the area occupied by stained processes. Thediffuse staining at 4 days would not have been detected inthe quantitation. Thus, the quantitative analyses probablyunderestimate the overall increase in immunostainingwithin the denervated zone, especially at the time of peakexpression.In the mutant animals, increases in immunostaining
were already detectable by 2 days postlesion (compareipsilateral vs. contralateral sides in Fig. 5A and B).However, in contrast to the situation in control animals,levels of immunostaining were not relatively higher withinthe denervated zone. Instead, astrocytes throughout thehippocampus ipsilateral to the lesion exhibited modestincreases in staining. The pattern was similar at 4 dayspostlesion, and again, levels of immunostaining were notrelatively higher within the denervated zone (Fig. 5C,D).This is in striking contrast to the situation at 4 dayspostlesion in control animals (Fig. 4C,D). It was not until8–10 days postlesion that the pattern of immunostainingbegan to resemble what was seen in the control animals,that is, higher levels of immunostaining in the denervatedlaminae (see Fig. 5E for an example of the pattern ofimmunostaining at 10 days).Quantitative analyses revealed that the increases in
immunostaining were quantitatively similar in mutantand control animals at 2 days postlesion despite the factthat the pattern of immunostaining appeared qualita-tively different (Fig. 6).After 2 days, the extent of immuno-staining increased more slowly in the WldS mutant ani-mals and reached a peak somewhat later. However, thedifferences in the time course of the changes in immuno-staining in mutant and control animals as revealed by thequantitative analyses were slight and were not statisti-cally significant.
DISCUSSION
We undertook this study to evaluate reactive responsesof astrocytes in denervated neuropil regions in WldSanimals, in which the onset of Wallerian degeneration andthe activation of microglia are substantially delayed. Our
Fig. 3. Quantitative analysis of GFAP mRNA levels in the dentategyrus after unilateral entorhinal cortex lesions. The graph illustratesthe mean ratio (6S.E.M.) of grain density ipsilateral vs. contralateralto an entorhinal cortex (EC) lesion at the indicated postlesion inter-vals in C57BL/6 control and WldS mutant animals. The data areexpressed as an I/C ratio of grain density. For the C57BL/6, thenumber of animals per time point are: unoperated, n5 2; 2 days, n5 3;4 days, n 5 3; 6 days, n 5 2; 8 days, n 5 3; 10 days, n 5 2; 12 days, n 53; 14 days, n 5 3; 20 days, n 5 3. For theWldS, the number of animalsper time point are: unoperated, n 5 2; 2 days, n 5 3; 4 days, n 5 2; 6days, n5 2; 8 days, n5 2; 10 days, n5 3; 12 days, n5 2; 14 days, n5 3;20 days, n 5 4. The dashed line indicates a ratio of 1 (equal graindensity on the two sides).
76 O. STEWARD AND P.A. TRIMMER
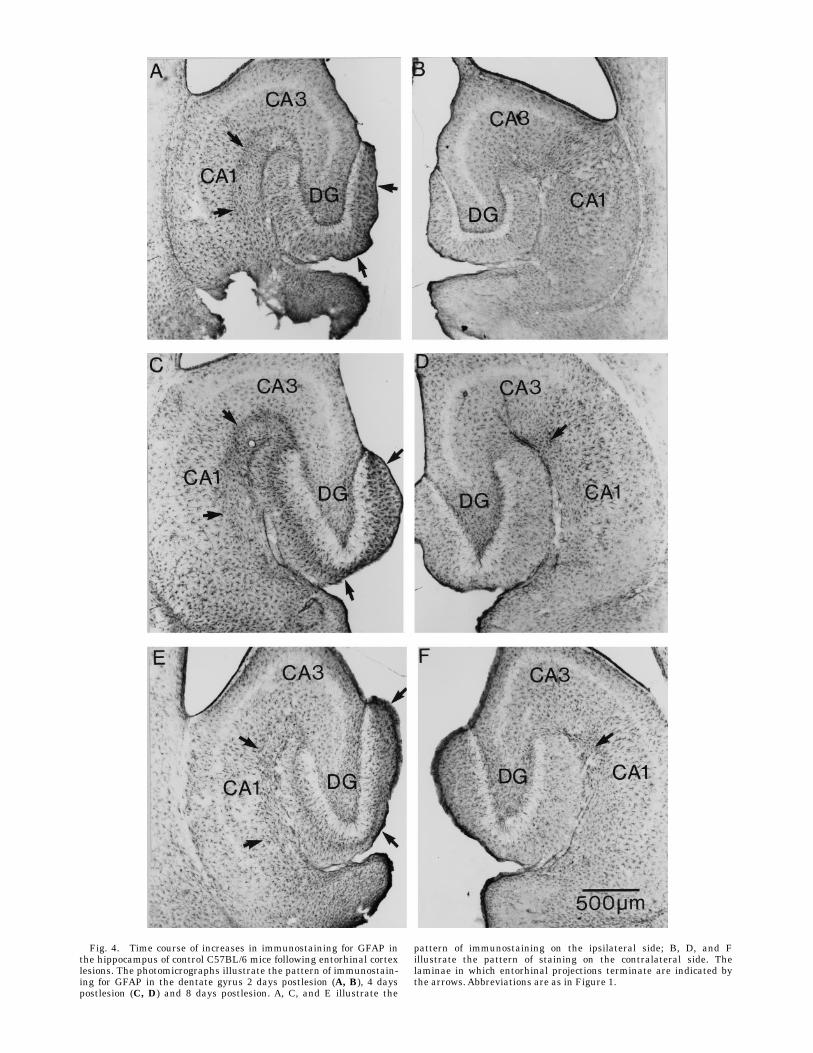
Fig. 4. Time course of increases in immunostaining for GFAP inthe hippocampus of control C57BL/6 mice following entorhinal cortexlesions. The photomicrographs illustrate the pattern of immunostain-ing for GFAP in the dentate gyrus 2 days postlesion (A, B), 4 dayspostlesion (C, D) and 8 days postlesion. A, C, and E illustrate the
pattern of immunostaining on the ipsilateral side; B, D, and Fillustrate the pattern of staining on the contralateral side. Thelaminae in which entorhinal projections terminate are indicated bythe arrows. Abbreviations are as in Figure 1.
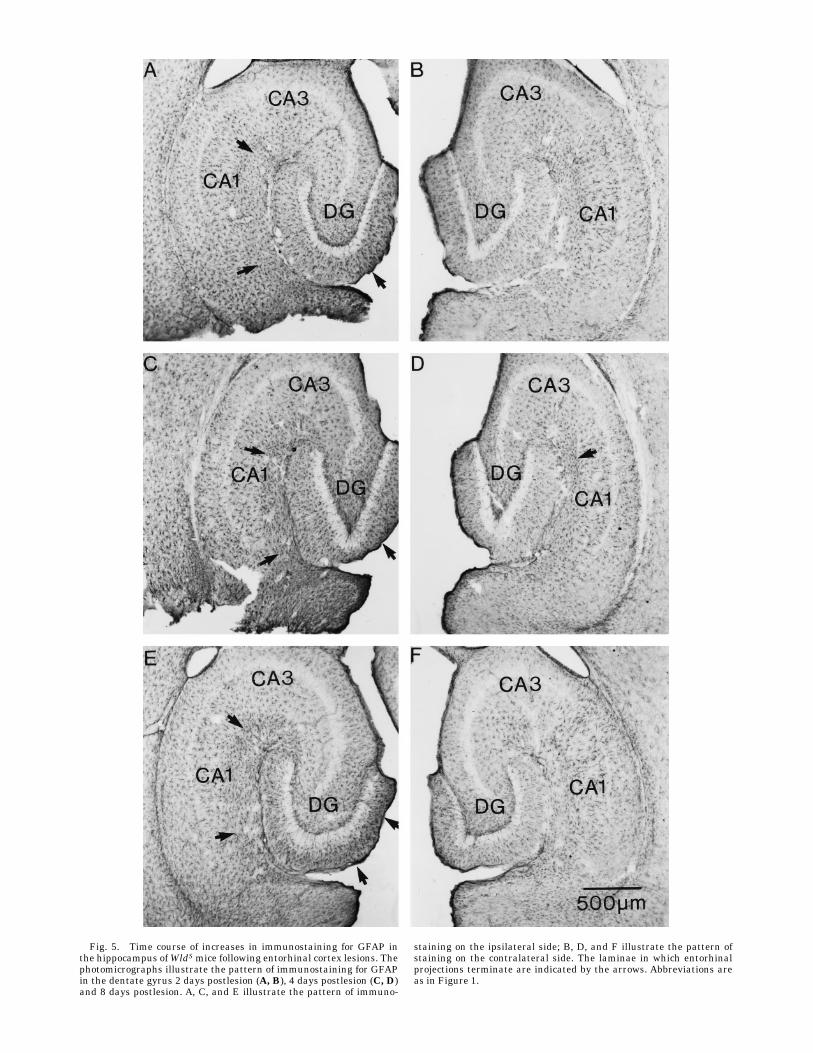
Fig. 5. Time course of increases in immunostaining for GFAP inthe hippocampus ofWldSmice following entorhinal cortex lesions. Thephotomicrographs illustrate the pattern of immunostaining for GFAPin the dentate gyrus 2 days postlesion (A, B), 4 days postlesion (C, D)and 8 days postlesion. A, C, and E illustrate the pattern of immuno-
staining on the ipsilateral side; B, D, and F illustrate the pattern ofstaining on the contralateral side. The laminae in which entorhinalprojections terminate are indicated by the arrows. Abbreviations areas in Figure 1.
goal was to use the information derived to infer intercellu-lar signalling cascades. According to the rationale outlinedin the Introduction, components of the reactive responsethat are triggered by signals from activated microgliashould not occur in WldS mutant animals until aftermicroglial activation (that is, until after 6–8 days post-lesion). Components of the response that are directlytriggered by degenerative changes in axons or synapticterminals should be substantially delayed, although notnecessarily to the same extent as the delay in microglialactivation. Alternatively, components of the reactive re-sponse that are triggered by signals unrelated toWalleriandegeneration should occur with the same time course innormal andWldSmutant animals.In seeking to define the triggers for activation, it is
important to focus on the onset of the reactive responses,the evolution of the responses over time, and the spatialpattern of the response with respect to the location ofdegenerating axons and synaptic terminals. In this con-text, our results revealed that the onset of the reactiveresponse within the denervated neuropil was delayed, andthe evolution of the changes over time occurred moreslowly in the mutant animals. Nevertheless, there weremodest reactive changes in astrocytes in mutant animalsas early as 2 days postlesion which were seen throughoutthe hippocampus ipsilateral to the lesion. Thus, the spatialpattern of increased GFAP expression was also different inthe mutants. In what follows, we will discuss: (1) the re-active response of astrocytes in denervated neuropil re-gions; and (2) the signals that could be responsible for theearly widespread increases in GFAP expression that areseen throughout the hippocampus ipsilateral to the lesion.
Reactive changes in astrocytes in denervatedneuropil regions
We used two measures to evaluate the time course andextent of the reactive response of astrocytes in the dener-vated neuropil: (1) the upregulation of GFAPmRNAlevels;and (2) increases in immunostaining for GFAP. The tempo-ral relationship between changes in GFAPmRNAand GFAprotein levels following EC lesions has previously beendefined in rats (Steward et al., 1993), and the presentresults reveal that the responses are similar in normalmice. In rats, there is a biphasic induction of GFAPmRNA.GFAP mRNA levels in the denervated zones begin to risewithin hours (Steward et al., 1990), reach a peak at 2 dayspostlesion, decrease to an intermediate level by 4 days,and then increase again to a second smaller peak at 6–8days postlesion. In control mice, the initial peak in labelingpersisted for somewhat longer (until 4 days postlesion),and the second peak in labeling was at 10 days postlesion.These differences in the evolution of the response in ratsvs. mice could be due to species differences or differences inthe manner of lesion production (electrolytic in rats vs.aspiration lesions in mice). It is of interest that the secondpeak in GFAPmRNA levels occurs during the period whennew synapses are forming on the denervated granule cells(Steward and Vinsant, 1983), although the significance ofthis fact is not clear.In contrast to the dynamics of mRNA induction, changes
in immunostaining were monophasic, and the time courseof the changes in mice appeared comparable to what hasbeen described in rats. Levels of immunostaining in-creased gradually to a peak at 4–6 days postlesion andthen gradually declined. In general, the increases anddecreases in immunostaining occurred in parallel with thedevelopment and regression of astrocyte hypertrophy. It isimportant to emphasize the spatial selectivity of theseresponses in normal animals; levels of labeling for GFAPmRNA and levels of immunostaining were always muchgreater in the areas that contained degenerating axonsand terminals.InWldSmutant animals, the rapid transient increase in
GFAP mRNA levels in the denervated zones which is soprominent in normal animals was not seen. In fact, at 2–4days postlesion, there was little evidence of relativelygreater labeling within the denervated laminae. The char-acteristic pattern of relatively higher levels of labeling inthe denervated zones was not apparent until 6–8 dayspostlesion. Thus, the induction of GFAP mRNA in thedenervated zonewas clearly delayed in themutant animals.The development of a well-defined band of intensely
immunostained and hypertrophied astrocytes in the dener-vated zone was also delayed in theWldS animals. However,this delay was not as apparent as the delay in GFAPmRNA expression, in part because there was a modestincrease in immunostaining at early postlesion intervalsthat appeared to be part of a generalized response thatoccurred throughout the hippocampus ipsilateral to thelesion. Because of this early generalized increase in immu-nostaining, the delay in the development of the reactiveresponse in the denervated zone was not evident in thequantitative analysis.Taken together, the combined results indicate that the
reactive response of astrocytes in the denervated zone wasdelayed in the WldS animals. Thus, the component of thereactive response that occurs in areas containing degener-
Fig. 6. Quantitative analysis of levels of immunostaining for GFAPin the dentate gyrus after unilateral entorhinal cortex lesions. Thegraph illustrates the mean (6S.E.M.) ratio of immunostaining ipsilat-eral vs. contralateral to an EC lesion at the indicated postlesionintervals in C57BL/6 control and WldS mutant animals (n 5 3 pertime point except for the normal, 4-day, 8-day, and 21-day postlesionC57BL/6 for which n 5 6, 5, 5, and 4, respectively). The dashed lineindicates a ratio of one (equal grain density on the two sides).
GENETIC INFLUENCES ON REACTIVE ASTROCYTOSIS 79

ating axons and terminals is most likely triggered directlyor indirectly by degeneration debris. Unfortunately, adetermination of whether the linkage is direct or indirectis problematic, because the widespread transient increasein GFAP expression complicates the quantitative analysisof the evolution of the changes in the denervated zone. Forthis reason, it is not possible to determine whether thedelay in the denervated is proportionate to the delay inmicroglial activation, as would be expected if this compo-nent of the reactive response was directly triggered viamicroglia.
The reactive changes in astrocytes that occurbefore the onset of degeneration or the
activation of microglia
In considering the signals that might induce reactivechanges prior to the onset of degeneration, it is importantto again recall the spatial distribution of the reactivechanges at the early postlesion intervals (2–4 days). Incontrol C57BL/6 mice, increases in GFAP mRNA andincreases in immunostaining were relatively higher in thedenervated zones, but there were also generalized in-creases in GFAP expression throughout the hippocampusipsilateral to the lesion. In contrast, in mutant animals,there were generalized increases in GFAP expressionthroughout the hippocampus ipsilateral to the lesion, butno indication of a relatively greater reactive response inthe denervated zone. These findings are consistent withearlier studies indicating that there are two components ofthe reactive response after EC lesions, one that occurs atearly postlesion intervals throughout the hippocampusand dentate gyrus, and one that occurs selectively in thedenervated laminae (Steward et al., 1990, 1993). Only theformer response is seen in WldS mutant animals at 2–4days postlesion.In rats, the increases in GFAP mRNA in nondenervated
zones in the hippocampus following EC lesions have beenshown to be due in part to physiological events. The role ofphysiological events in inducing this response was demon-strated in studies in which physiological recordings weremade during the production of electrolytic lesions andduring the first 24 hours following the lesion (Kelley andSteward, 1996a) and then the pattern of GFAP upregula-tion was evaluated in the same animals (Kelley andSteward, 1996b). These studies focused especially on theenigmatic increases in GFAP mRNA that were seen in thehippocampus contralateral to the lesion in some animals.The physiological recordings revealed that a variablenumber of episodes of spreading depression were triggeredduring lesion production and that all animals exhibitedspontaneous seizures during the 24-hour period after thelesion. There was no relationship between the occurrenceof episodes of spreading depression and the transientincreases in GFAP expression in the contralateral hippo-campus. Instead, increases in GFAP mRNA levels in thecontralateral hippocampuswere entirely predictable basedon the timing, number, and spatial distribution of seizuresthat occurred during the early postlesion interval.The occurrence of seizures also predicted increases in
GFAPmRNA in the nondenervated zones in the hippocam-pus ipsilateral to the lesion. That is, increases in GFAPexpression were greater in the animals that exhibited alarger number of seizures. At the same time, however,some of the increases in GFAP mRNA in nondenervatedlaminae ispilateral to the lesion could also have been due
to the episodes of spreading depression that occurredduring lesion production.The findings in rats provide a basis for understanding
the pattern of increases in GFAP mRNA levels in mice. Inthe first place, it is important to note that mice probably donot experience the same pattern of postlesion seizures.The seizures that have been documented in rats were afterelectrolytic lesions (Dasheiff and McNamara, 1981; Kelleyand Steward, 1996a). It is thought that an importantfactor in these seizures is the deposition of iron whichoccurs at the tip of the electrode used to produce theelectrolytic lesion (Dasheiff and McNamara, 1981). If thisis so, the lesions in mice produced by aspiration would notbe expected to produce the same type of postlesion sei-zures. This probably accounts for the lack of increases inGFAP expression in the hippocampus contralateral to thelesion in the mice.The widespread transient increases in GFAP expression
in the hippocampus ipsilateral to the lesion are probablynot related to postlesion seizures, but may be related tophysiological events that occur at the time of the lesion. Inparticular, aspiration lesions could produce bouts of spread-ing depression in the ipsilateral hippocampus which arecapable of strongly upregulating GFAP expression (Bon-thius and Steward, 1993). This could account for thewidespread upregulation of GFAP mRNA levels in thehippocampus ipsilateral to the lesion in the mice. It isimportant to emphasize, however, that until the physiologi-cal events that occur as a result of aspiration lesions inmice are defined, the above hypothesis remains specula-tive.
CONCLUSION
We began our studies of the WldS mutation with theexpectation that the entire cascade of lesion-induced cellu-lar events would be quite different than in control animals.The present results, together with the results of previousstudies (Fujiki et al., 1996; Schauwecker and Steward,1997; Steward, 1992; Zhang et al., 1996) confirm thisexpectation. If the reactive responses of non-neuronal cellsplay a role in the synaptic reorganization that occurs afterinjury, one would expect this reorganization to be quitedifferent in the mutant animals. Indeed, there is alreadyevidence that this is the case for at least one form ofpostlesion growth (the sprouting of the cholinergic affer-ents; see Steward, 1992). It will be of considerable interestto define exactly how different the neuronal growth re-sponses are in a tissue environment in which the timingand nature of the responses of non-neuronal cells aresubstantially modified as a result of the mutation.
LITERATURE CITED
Bonthius, D., and O. Steward (1993) Induction of cortical spreadingdepression with potassium chloride upregulates levels of messengerRNA for glial fibrillary acidic protein in cortex and hippocampus:Inhibition by MK-801. Brain Res. 618:83–94.
Buckmaster, E.A., V.H. Perry, and M.C. Brown (1995) The rate of Walleriandegeneration in cultured neurons from wild type and C57BL/WidS micedepends on time in culture and may be extended in the presence ofelevated K1 levels. Eur. J. Neurosci. 7:1596–1602.
Dasheiff, R.M., and J.O. McNamara (1981) Electrolytic entorhinal lesionscause seizures. Brain Res. 231:444–450.
Fagan, A.M., and F.H. Gage (1990) Cholinergic sprouting in the hippocam-pus: a proposed role for IL-1. Exp. Neurol. 110:105–120.
80 O. STEWARD AND P.A. TRIMMER

Fagan, A.M., and F.H. Gage (1994) Mechanisms of sprouting in the adultcentral nervous system: Cellular responses in areas of terminal degen-eration and reinnervation in the rat hippocampus. Neuroscience 58:705–725.
Fujiki, M., Z. Zhang, L. Guth, and O. Steward (1996) Genetic influences oncellular reactions to spinal cord injury: Activation of macrophages/microglia and astrocytes is delayed in mice carrying a mutation (WldS)that causes delayedWallerian degenerarion. J. Comp. Neurol. 371:469–484.
Furukawa, S., Y. Furukawa, E. Satoyoshi, and K. Hayashi (1986) Synthesisand secretion of nerve growth factor bymouse astroglial cells in culture.Biochem. Biophys. Res. Commun. 136:57–73.
Houlgatte, R., M. Mallet, P. Brachet, and A. Prochiantz (1989) Secretion ofnerve growth factor in cultures of glial cells and neurons derived fromdifferent regions of the mouse brain. J. Neurosci. Res. 24:143–152.
Ip, N.Y., S.J. Wiegand, J. Morse, and J.S. Rudge (1993) Injury-inducedregulation of ciliary neurotrophic factor mRNA in the adult rat brain.Eur. J. Neurosci. 5:25–33.
Kelley, M.S., and O. Steward (1996a) The process of reinnervation in thedentate gyrus of adult rats: Physiological events at the time of lesionand during the early postlesion period. Exp. Neurol. 139:73–82.
Kelley, M.S., and O. Steward (1996b) The role of postlesion seizures andspreading depression in the upregulation of glial fibrillary acidicprotein mRNA after entorhinal cortex lesions. Exp. Neurol. 139:83–94.
Kraig, R.P., L. Dong, R. Thisted, and C.B. Jaeger (1991) Spreadingdepression increases immunohistochemical staining of glial fibrillaryacidic protein. J. Neurosci. 11:2187–2198.
Lewis, S.A., J.M. Balcarek, V. Krek, M. Shelanski, and N.J. Cowan (1984)Sequence of a cDNA clone encoding mouse glial fibrillary acidic protein:Structural conservation of intermediate filaments. Proc. Nat.Acad. Sci.,U.S.A. 81:2743–2746.
Matthews, D.A., C. Cotman, and G. Lynch (1976a) An electron microscopicstudy of lesion-induced synaptogenesis in the dentate gyrus of the adultrat. I Magnitude and time course of degeneration. Brain Res. 115:1–21.
Matthews, D.A., C. Cotman, and G. Lynch (1976b) An electron microscopicstudy of lesion-induced synaptogenesis in the dentate gyrus of the adultrat. II Reappearance of morphologically normal synaptic contacts.Brain Res. 115:23–41.
McKeon, R.J., R.C. Schreiber, J.S. Rudge, and J. Silver (1991) Reduction ofneurite outgrowth in a model of glial scarring following CNS injury is
correlated with the expression of inhibitory molecules on reactiveastrocytes. J. Neurosci. 11:3398–3411.
Perry, V.H., M.C. Brown, E.R. Lunn, P. Tree, and S. Gordon (1990a) Evi-dence that very slow Wallerian degeneration in C57BL/Ola mice is anintrinsic property of the peripheral nerve. Eur. J. Neurosci. 2:802–808.
Perry, V.H., E.R. Lunn, M.C. Brown, S. Cahusac, and S. Gordon (1990b)Evidence that the rate of Wallerian degeneration is controlled by asingle autosomal dominant gene. Eur. J. Neurosci. 2:408–413.
Perry, V.H., M.C. Brown, and E.R. Lunn (1991) Very slow retrograde andWallerian degeneration in the CNS of C57BL/Ola mice. Eur. J. Neuro-sci. 3:102–105.
Raisman, G. (1969) Neuronal plasticity in the septal nuclei of the adult rat.Brain Res. 14:25–48.
Schauwecher, P.E., and O. Steward (1997) Genetic influences on cellularreactions to brain injury: Activation of microglia in denervated neuropilin mice carrying a mutation (WldS) that causes delayed Walleriandegeneration. J. Comp. Neurol. 380:82–94.
Steward, O. (1980) Trajectory of contralateral entorhinal axons whichreinnervate the fascia dentata of the rat following ipsilateral entorhinallesions. Brain Res. 183:277–289.
Steward, O. (1989) Reorganization of neuronal connections followingtrauma: Principles and experimental paradigmns. J. Neurotrauma6:99–152.
Steward, O. (1992) Signals that induce sprouting in the central nervoussystem: Sprouting is delayed in a strain of mouse exhibiting delayedaxonal degeneration. Exp. Neurol. 118:340–351.
Steward, O. (1994) Electroconvulsive seizures upregulate astroglial geneexpression selectively in the dentate gyrus. Mol. Brain Res. 25:217–224.
Steward, O., E.R. Torre, L.L. Phillips, and P.A. Trimmer (1990) The processof reinnervation in the dentate gyrus of adult rats: Time course ofincreases in mRNA for glial fibrillary acidic protein. J. Neurosci.10:2373–2384.
Steward, O., E.R. Torre, R. Tomasulo, and E. Lothman (1991) Neuronalactivity up-regulates astroglial gene expression. Proc. Nat. Acad. Sci.,U.S.A. 88:6819–6823.
Steward, O., M. Kelley, and E. Torre (1993) The process of reinnervation inthe dentate gyrus of adult rats: Temporal relationship between changesin the levels of glial fibrillary acidic protein (GFAP) and GFAPmRNA inreactive astrocytes. Exp. Neurol. 124:167–183.
GENETIC INFLUENCES ON REACTIVE ASTROCYTOSIS 81







![Original Article Glioneuronal tumor with neuropil-like ... tumor with neuropil-like islands: a histological, immunohistochemical, ... located in the cerebrum [2-8]. ... cular proliferation](https://img.pdfslide.net/doc/110x75/5ab547337f8b9a0f058c9d40/original-article-glioneuronal-tumor-with-neuropil-like-tumor-with-neuropil-like.jpg)





















