Embed Size (px)
Citation preview

Invest. agr.: Prod. Sanid. anim., 3 (1), 1988 109
GENETICA DE LA PROLIFICIDAD EN EL CERDO IBERICO: REVISION DE METODOLOGIAS Y RESULTADOS
M.T. DOBAO J. RODRIGAÑEZ
L. SILIO M. A. TORO
Departamento de Genética y Mejora Animal, INIA Apartado 811 1, 28080-Madrid
E. DE PEDRO
Departamento de Producciones Animales, ETSIA Apartado 3048, 14080-Córdoba
RESUMEN El análisis del cruce dialélico completo de cuatro antiguas estirpes de cerdo Ibérico evidencia una
heterosis individual de 0,40 lechones /camada, lo que sugiere que el cruce de líneas de Ibérico es el método más sencillo de mejora de prolificidad para aquellos ganaderos interesados en la producción de animales Ibéricos puros.
La utilización de cruces con razas del tipo Taihu ofrece superiores expectativas. El tamaño medio de camada en los tres primeros partos de cerdas Ibéricas (IB) e híbridas Jiaxingxlbérico (JXxIB) es de 8,5 ± 0,3 y de 11,4 ± 0,4 lechones respectivamente. Pero el cruce de estas cerdas híbridas con verracos Ibéricos resulta insatisfactorio por el escaso crecimiento y rendimiento en piezas nobles de los productos terminales (l/4JX,3/41B), por lo que deben ensayarse esquemas alternativos de cruza-miento.
La selección con empleo de índices familiares posibilita, dadas las estimas disponibles en poblacio-nes de cerdo Ibérico (h2=0,10; 2
pσ = 4,00; t=0,15), respuestas teóricas por generación de 0,20 a 0,36 lechones/camada. Aunque estudios de simulación evidencian diversos problemas en el uso de estos índices en poblaciones de tamaño moderado: diferenciales de selección inferiores a los esperados y elevadas tasas de consanguinidad.
Una disminución de 0,20 lechones por cada 10 p. 100 de aumento de la consanguinidad de la
madre o de la camada se ha estimado sobre datos registrados en tres poblaciones cerradas de pequeño tamaño (Ne= 11,96; 15,13 y 36,14). Diversos métodos de minimización de la tasa de consanguinidad se analizan a partir de dos programas de conservación de estirpes. El mantenimiento de una estructura
Recibido: 10-1 -1987. Aceptado para su publicación: 24-1-88. Redactor Asociado: R. Alenda.

M. T. DOBAO et al.
110
familiar equilibrada y el apareamiento de la combinación de reproductores de mínimo parentesco en cada generación se proponen como normas básicas en programas de este tipo.
PALABRAS CLAVE: Prolificidad, cerdo Ibérico, consanguinidad, cruzamiento, selección.
INTRODUCCION
Los progresos registrados en las últimas décadas en Europa y América en la productivi-dad numérica en el cerdo son fundamentalmente atribuibles a mejoras en manejo y sani-dad, y en lo genético, se deben a la sustitución de razas y líneas y al empleo generalizado de hembras híbridas, métodos ambos que agotan su potencial en pocas generaciones. Por contra, la proporción de magro en canal, objetivo de la práctica totalidad de los programas de selección, se encuentra próxima al óptimo en las poblaciones europeas de explotación intensiva. Ello ha renovado el interés por los caracteres reproductivos acompañado de un reexamen teórico de nuevas estrategias para la mejora genética de los mismos, en particular la prolificidad (Hill y Webb, 1982).
En este sentido están siendo experimentados en diversos países métodos como la crea-ción de líneas hiperprolíficas, mediante la aplicación de fuertes intensidades de selección en grandes bancos de datos, la selección indirecta para variables fisiológicas correlacionadas con el tamaño de camada o el uso de índices familiares a partir de datos de prolificidad registrados en hembras adultas emparentadas con los candidatos (Haley et al., 1986). Otro enfoque alternativo es la utilización en esquemas de cruzamiento de un grupo de razas chinas, del tipo denominado Taihu, caracterizado por sus excepcionales cualidades repro-ductoras (Legault y Sellier, 1986).
La aplicación de cualquiera de estos métodos al cerdo Ibérico, en el que dada su baja productividad numérica, la mejora de la prolificidad reviste gran interés, plantea numerosos problemas específicos, a comenzar por la práctica inexistencia en esta raza de estimas de parámetros genéticos y fenotípicos de los principales caracteres reproductivos.
El presente trabajo sintetiza diversos estudios, realizados en los tres últimos años, orien-tados a la obtención de estimas de algunos de estos parámetros y a la evaluación de las posibilidades de mejora genética de la prolificidad en el cerdo Ibérico. A partir de resultados experimentales se analizan las expectativas y problemas de los tres enfoques que, por el momento, consideramos más adecuados: a) cruces entre líneas de Ibérico, b) cruces con la raza china Jiaxing, perteneciente al tipo Taihu y e) selección mediante el empleo de índices familiares.
El conocimiento de la genética de los caracteres reproductivos presenta en el cerdo Ibérico un interés adicional dada la precaria conservación de la población, y, en particular, de algunos tipos tradicionales en riesgo de extinción. Por ello, es igualmente objeto de este trabajo el análisis de los efectos negativos de la consanguinidad sobre caracteres reproducti-vos en cerdos Ibéricos y sus implicaciones en el diseño de programas de conservación de estirpes en poblaciones cerradas.
Los resultados presentados se refieren, en su mayor parte, al tamaño de camada al nacimiento, que constituye el componente de la productividad numérica preferentemente estudiado desde el punto de vista genético. Los efectos ambientales sobre este carácter, aunque serán incluidos en algunos casos, en los modelos de análisis empleados, no constitu-yen el objetivo central de este trabajo. Sobre este tema, particularmente los aspectos de influencia estacional sobre caracteres reproductivos, así como sobre la genética de otros

GENETICA DE PROLIFICIDAD EN EL CERDO IBERICO
Invest. agr.: Prod. Sanid. anim., 3 (1), 1988 111
caracteres de cría como el número de mamas en el cerdo Ibérico, hemos realizado con anterioridad diversos trabajos (Dobao et al., 1983 y 1986; Toro et al., 1986a).
DIFERENCIAS ENTRE POBLACIONES DE IBERICO Y MEJORA POR CRUZAMIENTO ENTRE LINEAS
DIFERENCIAS ENTRE POBLACIONES DE CERDO IBERICO
Si bien la práctica totalidad de las razas ganaderas comparten como rasgo común una considerable heterogeneidad interna, posiblemente sea el cerdo Ibérico entre nuestras razas autóctonas la que históricamente haya presentado mayor diversidad de tipos morfológicos dentro de las características comunes al conjunto de la población: capa oscura, hocico alargado, apetito y metabolismo reducidos, madurez temprana, tamaño pequeño y formas compactas. La población de cerdos Ibéricos mantuvo hasta el inicio de los años 60, junto a un censo de efectivos muy considerable (567.424 cerdas de vientre censadas en 1955), una estructura tradicional de carácter reticulado con coexistencia de distintos tipos de difusión comarcal y escaso flujo genético entre los mismos.
El último cuarto de siglo, que ha conocido una espectacular reducción del censo, ha supuesto un cambio drástico de la estructura populacional con la irrupción generalizada del cruzamiento industrial, aunque, afortunadamente, todavía se conserven en nuestro país algunos tipos de Ibérico de características morfológicas y productivas bien diferenciadas: estirpes Torbiscal y Guadyerbas, línea Valdesequera y otros varios en manos de esforzados ganaderos privados.
Ciertamente, en 1945 no hubo demasiados problemas para elegir, tras una detenida exploración de las ganaderías más acreditadas de la península, cuatro de ellas representati-vas de los principales tipos, con vistas a la puesta en marcha del Centro de Cría de Cerdos Ibéricos en «El Dehesón del Encinar» (Oropesa, Toledo). Su denominación y procedencia eran como sigue:
A) Ervideira, procedente del Alentejo Central, de capa colorada dorada similar a la del rubio andaluz y tipo rústico y relativamente tardío.
B) Campanario, originario de La Serena, cerdos robustos y negro lampiños. C) Caldeira, de la región de Elvas, de capa retinta oscura. D) Puebla, del tipo pelón guadianés, negro lampiño, muy precoz y menos rústico que los
anteriores. Estos cuatro grupos fundacionales procedían de antiguas ganaderías de reconocido aisla-
miento genético y fueron mantenidos como estirpes puras en el Centro de «El Dehesón» hasta su fusión en 1963, aunque se ensayasen regularmente los cruzamientos recíprocos entre todas ellas. Los datos registrados en este período inicial de la piara experimental tienen por ello un singular valor en cuanto al conocimiento de las características productivas de los diferentes tipos de Ibérico. Aunque los datos relativos a los principales caracteres de cría, registrados en los períodos 1945/63 y 1964/73 de la citada piara experimental, fueron bri-llantemente analizados por su creador y responsable genético (Odriozola, 1976), los mode-los estadísticos empleados no se plantearon la comparación entre los grupos fundacionales ni consideraron los posibles efectos entre los mismos. Un análisis de los datos de prolifici-dad se llevó a cabo por nuestro grupo con esta orientación (Rodrigáñez et al., 1 985a). Su metodología y resultados se resumen seguidamente.
.

M. T. DOBAO et al.
112
Se dispuso de datos registrados en 1.109 camadas nacidas en las 11 parideras habidas entre 1948 y 1954, en las que se efectuaron simultáneamente los 16 tipos de cruzamiento posibles (cruce dialélico completo) entre las 4 estirpes fundacionales, oscilando el número de camadas representativo de cada tipo de apareamiento entre 44 y 107, siendo 65,7 su media armónica. Estos datos fueron analizados mediante el procedimiento descrito por Bulmer (1980), pp. 116-119, con una estimación previa de las medias de los tamaños de camada de los 16 tipos de apareamiento a partir de la cual se realizó el análisis de los efectos aditivos y heteróticos en las 4 estirpes estudiadas.
En la Tabla 1 se presentan los valores medios del número de nacidos y nacidos vivos por camada en cada uno de los 16 cruces, estimados por mínimos cuadrados mediante un modelo que incluye como efecto fijo el Tipo de Cruce (1 a 16), como efecto aleatorio la Paridera (1 a 11), la interacción de ambos Cruce x Paridera y como covariables, la edad y el coeficiente de consanguinidad de la madre.
La medida de la heterosis de un cruzamiento se obtiene fácilmente por diferencia respec-to a la media de los parentales. Los valores correspondientes de heterosis entre las estirpes (a) Ervideira, (b) Campanario, (e) Caldeira y (d) Puebla para las dos variables estudiadas (Nacidos Vivos) Hab=0,33 /0,41, Hac=0,42 /0,41 Had =0,58 /0,50 Hbc =0,76 /0,86, Hbd =0,53 /0,68 y Hcd =0,33 /0,44. Las heterosis medias (H) son de 0,49 nacidos y 0,55 nacidos vivos, equivalentes al 6,2 y 7,1 p. 100 de las correspondientes medias del carácter.
El análisis con un modelo de efectos genéticos de las medias estimadas de tamaños de camada se presenta en le Tabla 2. Los resultados indican la ausencia de efectos recíprocos significativos, que podrían ser atribuibles a efectos de ambiente materno. Con excepción de la heterosis media (H) ninguno de los otros efectos genéticos considerados en el modelo: aditivos, heterosis de estirpe y específica para cada cruzamiento, presentan significación es-tadística.
TABLA 1 MEDIAS ESTIMADAS DEL TAMAÑO DE CAMADA EN EL CRUCE DIALELICO COMPLETO
ENTRE 4 ESTIRPES DE CERDO IBERICO
Least-squares means of litter size in a complete diallel cross on four strains of Iberian pig.
Estirpe paterna Estirpe materna Ervideira Campanario Caldeira Puebla
Nacidos
(A) Ervideira ................................................................ 7,9 7,8 8,3 8,5 (B) Campanario ............................................................ 7,9 7,2 8,2 7,8 (C) Caldeira .................................................................. 8,2 8,2 7,7 8,3 (D) Puebla .................................................................... 8,2 8,0 7,6 7,6
(error típico = 0,28 nacidos)
Nacidos vivos
(A) Ervideira ................................................................ 7,8 7,7 8,2 8,2 (B) Campanario ............................................................ 7,7 6,8 7,8 7,6 (C) Caldeira .................................................................. 7,7 8,0 7,3 8,0 (D) Puebla .................................................................... 7,8 7,9 7,4 7,3
(error típico = 0,28 nacidos vivos)

GENETICA DE PROLIFICIDAD EN EL CERDO IBERICO
Invest. agr.: Prod. Sanid. anim., 3 (1), 1988 113
TABLA 2 ANÁLISIS DE VARIANZA DE LOS DATOS DE LA TABLA 1 (Rodrigáñez et al., 1985a)
Analysis of variance of data in Table I (Rodrigañez et al., 1985 a)
Total nacidos Nacidos vivos Fuente de variación g.l.
C.M. F C.M. F
Estirpes ..................................................... 3 0,095 1,88 NS 0,166 2,66 NS Heterosis mnedia ...................................... l 0,717 14,27 ** 0,897 4,36 ** Heterosis estirpes...................................... 3 0,008 0,15 NS 0,034 0,55 NS Heterosis específica .................................. 2 0,118 2,35 NS 0,066 1,05 NS Efectos recíprocos .................................... 6 0,050 0,063
La heterosis general observada debe atribuirse a diferencias entre las estirpes en las frecuencias génicas de loci que presentan dominancia para el carácter prolificidad. Por contra, los resultados no evidencian diferencias significativas en la prolificidad de las estir-pes comparadas atribuibles a genes de efecto aditivo sobre el tamaño de camada. El análisis de datos relativos a otros caracteres procedentes de este diseño dialélico ha confirmado asimismo la existencia de heterosis entre estas estirpes de cerdo Ibérico para características de crecimiento, y canal, así como para el número de mamas (Odriozola, 1956; Dobao et al., 1986).
HETEROSIS Y MEJORA POR CRUCE ENTRE LINEAS DE IBERICO
El interés de los resultados anteriores no se limita al conocimiento retrospectivo de tipos de 1bérico, algunos prácticamente desaparecidos, sino que tiene implicaciones útiles en cuanto a la obtención de moderadas mejoras de prolificidad por cruzamiento entre estirpes y líneas de Ibérico.
Los valores observados de heterosis se refieren a lo que se conoce como heterosis indivi-dual, debida al efecto directo del genotipo sobre el tamaño de la camada y son considerable-mente altos: 6-7 p. l00 de la media del carácter, cuando los valores comparables descritos en la literatura son únicamente del orden del 2 p. 100 (Sellier, 1976). Ello puede explicarse por la consanguinidad acumulada a lo largo del prolongado aislamiento genético de estas estirpes en manos de sus antiguos propietarios privados.
Es muy verosímil que la consanguinidad de algunas de las poblaciones diferenciadas de Ibérico que todavía se conservan en pureza sea bastante más elevada que la de las 4 estirpes aquí estudiadas. En Portugal, Antunes-Correia y Oliveira (1985) han señalado la elevada consanguinidad de los núcleos existentes de cerdo Alemtejano. Los reducidos censos de reproductores mantenidos, en ocasiones con esporádicos cuellos de botella, la proporción de machos y hembras generalmente inadecuada y la ausencia de control de genealogías y de diseño de los apareamientos son factores que, como veremos en un capitulo posterior, incrementan sensiblemente las tasas de consanguinidad por generación.
Parece pues razonable esperar del cruzamiento entre líneas actuales de Ibérico, y como resultado de heterosis directa, unos incrementos de prolificidad superiores a los 0,50 lechones por camada observados en los cruces analizados entre las estimes fundacionales de la piara de «El Dehesón».

M. T. DOBAO et al.
114
Posibilidades aun mayores en el incremento de la prolificidad ofrece el uso como madres de cerdas híbridas obtenidas por cruce entre líneas, aprovechando la heterosis ma-ternal, efecto del genotipo de la madre sobre el tamaño de camada, que en la literatura se estima del orden del 6 p. l00 de la media del carácter, y que por las razones anteriores expuestas, puede suponerse superior en el caso de cerdas híbridas de diferentes líneas de Ibérico, en especial cuanto más acreditado esté el anterior aislamiento reproductivo entre las mismas.
DIFERENCIAS ENTRE RAZAS Y MEJORA POR CRUZAMIENTO ENTRE RAZAS
DIFERENCIAS ENTRE RAZAS
El moderado progreso genético que ha conocido en las últimas décadas el carácter proli-ficidad en el cerdo, se ha basado principalmente en la utilización de las diferencias entre razas, bien mediante esquemas de cruzamiento que incluyen como líneas maternales cerdas obtenidas por hibridación entre razas o por el mero proceso de sustitución de razas que ha favorecido la expansión internacional de las más productivas.
El número de razas de cerdos en el mundo se considera próximo a 350, la mayor parte de las cuales serian razas locales. El conocimiento de su posible utilidad como líneas de madres requiere la evaluación conjunta tanto de sus características reproductivas como de las de crecimiento y composición corporal. La dificultad de esta tarea ha sido puesta de manifiesto en un reciente trabajo bibliográfico de Sutherland et al. (1984). Las poblaciones denominadas como razas son frecuentemente muy heterogéneas y mal definidas, sufriendo cambios considerables a lo largo del tiempo debidos a selección, deriva genética o incorpo-ración de genes de otras razas y aunque ciertamente algunas grandes razas disponen de libros genealógicos, los más antiguos no superan apenas el siglo de antigüedad, siendo la mayoría mucho más recientes. Las comparaciones objetivas aun entre estas razas de gran difusión vienen dificultadas porque las exportaciones de animales se realizan de forma sesgada y a pequeña escala, lo que unido a que los métodos de selección del país importador pueden diferir del originado hace que las características genéticas de razas que comparten un mismo nombre pueden ser gradualmente divergentes en diferentes países (King y Ava-los, 1986).
A pesar de las dificultades que plantea la comparación rigurosa entre razas, es claro que éstas, en conjunto, presentan una considerable variación en prolificidad, por lo que algunos autores como Legault (1985) distinguen cuatro categorías generales de razas de cerdos, desde el punto de vista de su potencial reproductivo y productivo:
Razas de doble aptitud, cuya característica común es un rendimiento satisfactorio tanto en prolificidad como en crecimiento y características de canal: Large White. Landrace y Duroc en Europa y Chester White y Yorkshire en Estados Unidos. El tamaño medio de camada en estas razas oscila en un intervalo de 10-11,5 lechones al parto y 8-9,5 destetados en buenas condiciones de manejo (Hill y Webb, 1982).
Razas de buena conformación, especializadas en la obtención de verracos para cruza-mientos terminales: Pietrain y Landrace Belga en Europa y Hampshire, Duroc y Poland China en Estados Unidos. Caracterizadas por una elevada proporción de magro en la canal y por una prolificidad moderada (8-10 lechones nacidos y 6-8 destetados por camada).

GENETICA DE PROLIFICIDAD EN EL CERDO IBERICO
Invest. agr.: Prod. Sanid. anim., 3 (1), 1988 115
Razas hiperprolíficas, grupo circunscrito aun pequeño número de razas originarias de la República Popular China, país donde se cría un tercio de la población mundial de cerdos, con más de 40 razas de importancia económica, de las que un número inferior a 20, y muy en particular las denominadas del tipo Taihu, presentan una excepcional capacidad repro-ductiva entre otras características que se resumen seguidamente:
1. Extrema precocidad sexual (primer celo a los 3-4 meses de edad), elevada prolificidad y excelente aptitud maternal como muestran los datos recogidos en la Tabla 3 correspon-dientes a 4 razas del tipo Taihu, 2 de las cuales (Meishan y Jiaxing) son estudiadas en Francia desde 1980 (Legault y Sellier, 1986). La base fisiológica de la superior prolificidad de estas razas, cuyas tasas de ovulación son similares a las razas europeas, parece radicar en sus menores tasas de mortalidad embrionaria (Bolet et al., 1986).
2. Reducido crecimiento y tamaño adulto: 80 kg de peso corporal se alcanzan con 8 meses de edad en China y 6 meses en Francia. Canales grasas y de mala conformación: La proporción de magro en canales Meishan y Jiaxing es l6-18 p. l00 inferior a la de Large White o Landrace, aunque la carne es de muy buena calidad organoléptica (Legault et al., 1985).
3. Rusticidad y excelente aptitud para el aprovechamiento de forrajes y subproductos domésticos e industriales como resultado de los hábitos de racionamiento chinos que histó-ricamente se han basado en estos productos, y en concreto en la región del lago Taihu, en plantas acuáticas, minimizando el uso de concentrados,
TABLA 3 PROLIFICIDAD Y NUMERO DE MAMÁS EN CUATRO RAZAS CHINAS DEL TIPO TAIHU
Prolificacv and teat number in four Taihu breeds
Tamaño de camada
Raza Partos Nº de
Camadas Nacidos Totales
Nacidos Vivos Destetadosa
Número de
mamas Referenciab
Meishan ........................ 1-2 386 14,0 12,9 11,3 16,8(664) (1) >3 511 17,0 14,8 2,9 - (1) 2-8 159 16,7 14,2 11,3 - (2) 1-5 305 15,6 14,5 13,0 16,1(1293) (3)
Jiaxing........................... 1-2 248 12,8 11,9 9,9 16,4(829) (1) >3 207 16,9 14,4 12,1 - (1) 2-8 537 13,2 11,9 - - (2) 1-5 173 11,9 113 10,2 18,8(472) (3)
Er Hua Lian .................. 1-2 1116 12,4 11,5 10,0 - (1) >3 1278 15,3 13,6 11,5 - (1) 2-8 421 16,1 14,6 - 18,1(414) (2)
Fen Ping........................ 1-2 346 14,2 12,8 11,7 17,4(4717) (1) >3 426 17,0 14,8 12,1 - (1) 2-8 523 16,7 14,5 - - (2)
a El destete en los datos chinos referencias 1 y 2) se produce a los 60 días y a 28 días en los obtenidos en Francia referencia 3).
b Zhang et al. (1983): (2) Cheng (1984) y 3) Bidanel y Legault (1986).

M. T. DOBAO et al.
116
Por último, razas locales, en número próximo a las 300 en todo el mundo, cuyo rasgo común es su buena adaptación a desfavorables condiciones ambientales de clima, alimenta-ción y manejo, Aunque sus rendimientos en cuanto a características reproductivas y de crecimiento son, usualmente, poco destacados, estas raras son muy apreciadas como com-ponentes maternales de esquemas de cruzamiento adaptados a sistemas de manejo extensi-vos. En el caso de nuestro cerdo Ibérico, desde hace un cuarto de siglo, ésta es la práctica seguida por una abrumadora mayoría de ganaderos, que tras ensayar el cruzamiento con la práctica totalidad de las razas europeas y americanas de capa oscura (Large Black, Berkshi-re, Tamworth. Wessex-Saddleback y Duroc-Jersey), han optado en general por los cruces con Duroc. Se acepta comúnmente que el empleo de madres híbridas Durocxlbérico posibi-lita aumentos del tamaño de camada del orden de 1-1,5 lechones, respecto al uso de hem-bras Ibéricas puras, aunque existe un vacío de estudios experimentales sobre este tema que acrediten de forma rigurosa las ventajas de estos u otros cruces posibles en cuanto a la mejora del carácter prolificidad.
MEJORA POR CRUZAMIENTO ENTRE RAZAS: EVALUACION DEL CRUCE CON LA RAZA CHINA JIAXING
Las características de las razas del tipo Taihu dan un gran interés potencial a su cruza-miento con el Ibérico como método drástico de disponer de hembras de prolificidad muy superior. Otras peculiaridades de estas razas chinas como su gran rusticidad, reducidas exigencias alimenticias y buena calidad de carne resultan asimismo idóneas, mientras que los aspectos más problemáticos evidenciados en los estudios realizados en Francia, relativos al inferior crecimiento y al engrasamiento y mala conformación de las canales, resultaban, a priori, inconvenientes menores en la comparación con cerdos Ibéricos, que en buena medi-da comparten estos rasgos.
A finales de 1984 se inició un proyecto de investigación, en el marco de la colaboración INIA-INRA y financiado por la CAICYT orientado a la realización de un programa de cruzamiento entre animales de razas Ibérica y Jiaxing que, mediante el registro de un conjunto de controles y medidas permitiera evaluar experimentalmente la posible aplica-ción de los cruces con esta raza china a los sistemas de producción extensivos típicos del cerdo Ibérico.
La raza Jiaxing pareció inicialmente más adecuada que la Meishan para este ensayo concreto, dado su mayor número de mamas, su superior aptitud combinatoria para el carácter prolificidad (Legault y Caritez, 1983) y el hecho, no anecdótico dadas las caracterís-ticas singulares del mercado de productos ibéricos, del color completamente negro de sus extremidades, mientras que los cerdos Meishan tienen cabos y uñas blancas.
Para la obtención del material biológico necesario, se importaron de Francia dosis de semen refrigerado de verracos Jiaxing (JX), proporcionadas por el INRA. En marzo de 1985 nacieron en el Centro de Investigación Agropecuario «Dehesón del Encinar» las 9 primeras camadas procedentes de la inseminación artificial, con las dosis citadas, de cerdas Ibéricas de la estirpe Torbiscal IB (E), con un total de 29 machos y 19 hembras Fl (JXxIB), que constituyen el material biológico de partida del presente estudio. Posteriormente, una nueva serie de inseminaciones ha permitido obtener otras 8 camadas, con un total de 28 machos y 19 hembras, de las que l0 se han reservado para la realización de estudios de fisiología

CRECIMIENTO Y CARACTERÍSITCAS DE CANAL DE CERDOS
Invest. agr.: Prod. Sanid. anim., 3 (1), 1988 117
reproductiva en el Departamento de Reproducción Animal del INIA, que asimismo se encargó de la realización del programa de inseminaciones.
El sistema de parideras agrupadas que se sigue en la piara experimental de «El Dehesón» ha permitido disponer como controles de animales Ibéricos coetáneos de los de origen Jiaxing, tanto en lo referente al registro de caracteres reproductivos (edad a la pubertad, prolificidad, etc.) como en los estudios de crecimiento y características de canal realizados en el propio Centro así como en cooperación con diversos ganaderos e industriales colabo-radores.
Evaluación de cruces con Jiaxing: caracteres reproductivos
La edad de aparición del primer celo fue registrada en el grupo de 19 hembras JXxIB(E) obtenido inicialmente así como en un grupo control de 18 cerdas Torbiscal IB(E). Un pequeño número de machos se introdujo diariamente de forma controlada en los aloja-mientos de ambos grupos de hembras, con vistas a la detección de celos. Los resultados mostraron diferencias significativas en la edad a la pubertad, siendo las medias respectivas de 130 ± 7 días en las hembras JXxIB y 207 ± 9 en las Ibéricas. Las determinaciones de niveles de progesterona en el primer celo mostraron valores similares en ambos grupos (Martín-Rillo et al., comunicación personal).
Los resultados de prolificidad en los 3 primeros partos de cerdas cruzadas JXxIB y controles Ibéricas se presentan en la Tabla 4. La cubrición de ambos tipos genéticos se ha realizado con el mismo contingente de verracos Ibéricos de la estirpe Torbiscal. Las cubri-ciones se realizaron mediante monta natural con control de genealogía, permaneciendo cada verraco encerrado durante 24 días con 5 a 8 cerdas de ambos tipos.
Los datos resumidos en la Tabla 4 proceden de camadas nacidas en 7 sucesivas parideras o agrupaciones discontinuas de partos que tienen lugar trimestralmente en «El Dehesón del Encina,», Un análisis de mayor rigor estadístico se ha realizado con el empleo de un modelo mixto de análisis de varianza, que incluye como efectos fijos el Tipo Genético (JXxIB e IB)
TABLA 4 TAMAÑOS DE CAMADA EN CERDAS DE TIPOS GENETICOS JIAXINC x IBERICO
E IBERICO Litter size comparison between Jiaxing x Iberian and Iberian control sows
Tipo Genético de cerda Parto Tamaño de camada
Jiaxing x Ibérico lberico
Significación de
diferencias
(N = 28) (N = 132) 1 Nacidos ................................................ 10,2 ± 0,5 7,5 ± 0,2 *** Nacidos vivos 10,0 ± 0,5 7,2 ± 0,2 *** (N = 9) (N = 53) 2 Nacidos ................................................ 11,4 ± 0,6 8,2 ± 0,3 *** Nacidos vívos 110 ± 0,7 7,7 ± 0,3 *** (N = 19) (N = 17) 3 Nacidos . ............................................... 12,6 ± 0,6 9,8 ± 0,3 * Nacidos vivos 120 ± 0,7 9,5 ± 0,3 *
M. T. DOBAO et al.
118
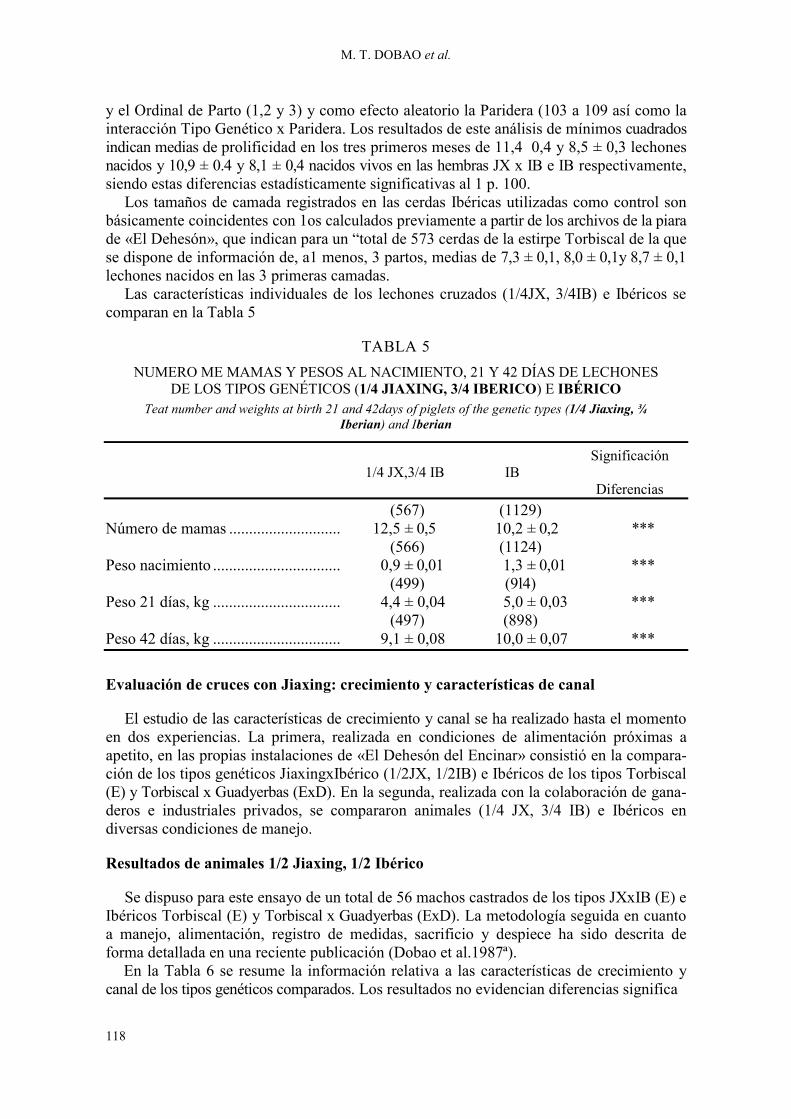
y el Ordinal de Parto (1,2 y 3) y como efecto aleatorio la Paridera (103 a 109 así como la interacción Tipo Genético x Paridera. Los resultados de este análisis de mínimos cuadrados indican medias de prolificidad en los tres primeros meses de 11,4 0,4 y 8,5 ± 0,3 lechones nacidos y 10,9 ± 0.4 y 8,1 ± 0,4 nacidos vivos en las hembras JX x IB e IB respectivamente, siendo estas diferencias estadísticamente significativas al 1 p. 100.
Los tamaños de camada registrados en las cerdas Ibéricas utilizadas como control son básicamente coincidentes con 1os calculados previamente a partir de los archivos de la piara de «El Dehesón», que indican para un “total de 573 cerdas de la estirpe Torbiscal de la que se dispone de información de, a1 menos, 3 partos, medias de 7,3 ± 0,1, 8,0 ± 0,1y 8,7 ± 0,1 lechones nacidos en las 3 primeras camadas.
Las características individuales de los lechones cruzados (1/4JX, 3/4IB) e Ibéricos se comparan en la Tabla 5
TABLA 5 NUMERO ME MAMAS Y PESOS AL NACIMIENTO, 21 Y 42 DÍAS DE LECHONES
DE LOS TIPOS GENÉTICOS (1/4 JIAXING, 3/4 IBERICO) E IBÉRICO Teat number and weights at birth 21 and 42days of piglets of the genetic types (1/4 Jiaxing, ¾
Iberian) and Iberian
1/4 JX,3/4 IB IB
Significación
Diferencias (567) (1129) Número de mamas ............................ 12,5 ± 0,5 10,2 ± 0,2 *** (566) (1124) Peso nacimiento ................................ 0,9 ± 0,01 1,3 ± 0,01 *** (499) (9l4) Peso 21 días, kg ................................ 4,4 ± 0,04 5,0 ± 0,03 *** (497) (898) Peso 42 días, kg ................................ 9,1 ± 0,08 10,0 ± 0,07 ***
Evaluación de cruces con Jiaxing: crecimiento y características de canal
El estudio de las características de crecimiento y canal se ha realizado hasta el momento en dos experiencias. La primera, realizada en condiciones de alimentación próximas a apetito, en las propias instalaciones de «El Dehesón del Encinar» consistió en la compara-ción de los tipos genéticos JiaxingxIbérico (1/2JX, 1/2IB) e Ibéricos de los tipos Torbiscal (E) y Torbiscal x Guadyerbas (ExD). En la segunda, realizada con la colaboración de gana-deros e industriales privados, se compararon animales (1/4 JX, 3/4 IB) e Ibéricos en diversas condiciones de manejo.
Resultados de animales 1/2 Jiaxing, 1/2 Ibérico
Se dispuso para este ensayo de un total de 56 machos castrados de los tipos JXxIB (E) e Ibéricos Torbiscal (E) y Torbiscal x Guadyerbas (ExD). La metodología seguida en cuanto a manejo, alimentación, registro de medidas, sacrificio y despiece ha sido descrita de forma detallada en una reciente publicación (Dobao et al.1987ª).
En la Tabla 6 se resume la información relativa a las características de crecimiento y canal de los tipos genéticos comparados. Los resultados no evidencian diferencias significa

CRECIMIENTO Y CARACTERÍSITCAS DE CANAL DE CERDOS
Invest. agr.: Prod. Sanid. anim., 3 (1), 1988 119
TABLA 6 CRECIMIENTO Y CARACTERÍSICAS DE CANAL DE CERDOS CASTRADOS IBÉRICOS
Y JIAXING x IBÉRICO (Dobao et al., 1987a) Growth and carcass performance of Iberian x Jiaxing, and Iberian barrows
Ibérico
Torbiscal Torbiscal
x Guadyerbas
Jiaxing x
Ibérico (Torbiscal)
s.e. medias
(N) (19) (19) (18)
Crecimiento
Peso inicial (1), kg ......................... 14,4 a 14,8 a 18,0 b 0,86 ** Peso ceba (2), kg............................ 90,2 88,9 90,1 2,44 G M D (1-2), kg/día....................... 0,53 0,52 0,50 0,01 E T M (90 kg), mm,....................... 32,4 32,8 30,9 0,73 Peso sacrificio (3) kg ..................... 138,9 a 139,9 a 129,2 b 2,02 *** G M D (2-3), kg/día....................... 0,70 a 0,73 a 0,56 b 0,04 ** Canal
Peso canal, kg ................................ 114,6 a 116,2 a 107,2 b 1,52 *** Peso lomos, kg .............................. 2,8 a 2,8 a 2,5 b 0,06 *** Peso jamones + kg ......................... 19,9 a 19,7 a 18,2 b 0,26 *** Peso paletas + kg ........................... 13,2 a 12,8 a 12,1 b 0,16 ***
+ Jamones y paletas recortadas en redondo.
tivas en el crecimiento en la recría, pero si en el periodo de ceba, en el que los animales JXxE registraron una ganancia media inferior a la de ambos tipos Ibéricos en 0,15kg, diferencia sólo atribuible en parte al menor apetito de aquellos animales. Mientras los índices de conversión en la recría fueron de 3,77, 3,83 y 3,95 en los lotes E. ExD y JXxE, en el periodo de ceba, aun descontando el pienso rechazado en el último lote, aproximadamen-te el 11 p. 100 del suministrado, los índices de conversión correspondientes fueron 4,73, 4,52 y 5,35. En cuanto a características productivas de las canales, tanto las medias del peso canal como de lomos, paletas y jamones son significativamente menores en los cerdos JXxE que en los dos tipos de Ibérico,
Un análisis más completo de las diferencias en composición corporal muestra, para un mismo peso canal, diferencias significativas tanto entre el tipo JXxE y los Ibéricos E y ExD como, en algunos casos, entre estos mismos. Como aspectos más notables, en los animales JXxE es menor el peso de ciertas partidas grasas con la excepción de la manteca en rama y mayor el peso de la cabeza y de los huesos de jamón y paleta. Las canales ExD son las que muestran un mayor engrasamiento así como un esqueleto más fino y las canales E son las de superior rendimiento en magro y piezas nobles (Dobao et al, 1987a).
Resultados de cerdos 1/4 Jiaxing, 3/4 Ibéricos
Desde el punto de vista práctico, los rendimientos de crecimiento y canal que presentan mayor interés, entre los diversos tipos de animales que intervienen en un esquema de cruzamiento son los de los productos terminales, que en definitiva constituyen la mayoría

M. T. DOBAO et al.
120
de los cerdos enviados a matadero. En el esquema de utilización de la raza Jiaxing, valorado inicialmente por nosotros, el tipo genético de los productos terminales es 1/4 JX, 3/4 IB, al obtenerse por el cruzamiento de hembras hibridas JiaxingxIbérico con verracos Ibéricos. Como controles de esta experiencia se dispuso de animales Ibéricos nacidos simultáneamen-te de hembras Ibéricas cubiertas por el mismo elenco de verracos, mediante el sistema de monta natural con genealogía controlada ya descrito en un apartado anterior.
A lo largo del último año, lotes de animales castrados de ambos tipos genéticos han sido proporcionados hasta un número total próximo a los 600, a 8 ganaderos colaboradores de diversas regiones españolas y con sistema de producción dispares, que incluyen la práctica totalidad de los empleados en cerdos del tipo Ibérico. Los animales, marcados individual-mente, fueron pesados a su salida del Centro «El Dehesón del Encinar», y a lo largo de su permanencia en cada una de las fincas colaboradoras, sometidos al régimen de manejo y alimentación habituales en las mismas, se registraron el peso y el espesor de tocino dorsal en el momento, próximo a los ocho meses de edad, de inicio de la ceba, y finalizada la misma el peso de envío al sacrificio. En matadero se registró el peso de la canal, los lomos, jamones y paletas recortados de cada uno de los animales sacrificados, de los que se prepararon asimismo muestras de lomo y grasa subcutánea para la realización posterior de análisis quí-micos.
Los resultados que se muestran en la Tabla 7 corresponden a un total de 199 animales pertenecientes a los lotes experimentales suministrados a un primer grupo de 5 ganaderos colaboradores, y para los que ya se dispone de la mayoría de los datos previstos. Los valores presentados son las medias estimadas por mínimos cuadrados con un modelo de análisis de varianza que incluye como efectos fijos: Sexo (machos o hembras castrados) y Tipo Genético (1/4 JX o IB), y como efecto aleatorio el Colaborador (1 a 5) que combina los efectos no controlables de cada explotación y matadero, así como las interacciones Sexo x
TABLA 7 CRECIMIENTO Y CARACTERISTICAS DE CANAL DE MACHOS Y HEMBRAS CASTRADOS
DE LOS TIPOS GENETICOS (1/4 JIAXING, 3/4 IBERICO) E IBERICO Growth and carcass performance of 1/4 Jiaxing, 3/4 Iberian x Jiaxing, and Iberian barrows and
castrated females
1/4 JX, 3/4
IB IB
Machos Hembras Machos Hembras
Significacióndiferencias
tipos genéticos
(N) (60) (61) (40) (38)
Peso inicial (1), kg .................... 21,2 20,6 21,5 21,3 NS Peso ceba (2), kg, ..................... 85,4 78,0 92,0 88,7 ** G MD (1-2), kg/día................... 0,41 0,37 0,46 0,43 * E T M, mm ............................... 32,7 30,9 34,6 33,6 ** Peso sacrificio (3), kg ............... 141,0 129,1 159,5 150,0 G M D (2-3), kg/día.................. 0,47 0,45 0,52 0,51 NS
Peso canal, kg ........................... 112,7 103,7 128,6 121,6 ** Jamones, kg .............................. 16,7 15,8 19,1 18,2 ** Paletas, kg................................. 11,1 10,7 12,9 12,1 ** Lomos, kg ................................. 2,8 2,8 3,2 3,2 ** Suma piezas noble .................... 31,029,3 35,2 33,5 **

CRECIMIENTO Y CARACTERÍSITCAS DE CANAL DE CERDOS
Invest. agr.: Prod. Sanid. anim., 3 (1), 1988 121
Tipo Genético y Tipo Genético x Colaborador, cuyo correspondiente cuadrado medio se utilizó como estima del error en el contraste de la significación estadística del efecto Tipo Genético. Estos resultados evidencian fuertes diferencias desfavorables a los animales cruza-dos 1/4 JX, 3/4 IB en la práctica totalidad de las características analizadas de crecimiento y canal.
Anteriormente se ha mencionado el menor apetito observado en los animales 1/2 JX, que rechazaban alrededor del 11 p. 100 del pienso suministrado durante la coba. Un proble-ma análogo se presenta en los animales 1/4 JX, que de acuerdo con las observaciones realizadas por los ganaderos colaboradores, consumen una menor cantidad de pienso mos-trando gran apetencia por el consumo de hierba y, en menor medida, bellotas. En el origen de esta menor apetencia por los concentrados, manifestada igualmente en la lactación de las hembras Fl(JXxIB) y evidenciada asimismo por los trabajos franceses, se hallan las caracte-rísticas singulares del tracto digestivo de los cerdos del tipo Taihu, de mayor longitud que el de las razas europeas, resultante de su adaptación a lo largo de los siglos al consumo de forrajes, como componente fundamental de su alimentación.
En lo relativo a la calidad de las canales, tanto la valoración visual de las mismas por parte de los responsables de los mataderos colaboradores como los parámetros químicos registrados permiten afirmar la inexistencia de diferencias considerables en calidad de carne entre animales Ibéricos y cruzados con Jiaxing. Los inconvenientes de las canales l/4JX, 3/41B proceden, lógicamente de su menor tamaño, 16-18kg inferior a los Ibéricos, que unido a la inferior conformación de paletas y jamones, se traduce en diferencias del orden de 4,2 kg en la suma de piezas nobles entre animales sacrificados con la misma edad. Asimismo la comparación de canales 1/2 JX e Ibéricos (Tabla 7), así como los resultados previamente obtenidos en China y Francia, sugieren como inconveniente adicional de las canales de origen Jiaxing la importancia relativa de componentes de canal de escaso aprove-chamiento como vísceras, piel y hueso.
SELECCION PARA PROLIFICIDAD
NUEVOS MÉTODOS DE SELECCION PARA PROLIFICIDAD EN CERDOS
Los métodos de cruzamiento entre razas o entre líneas, previamente discutidos, son sencillos y ofrecen rápidas posibilidades de mejora, pero agotan su máximo potencial en unas pocas generaciones. Los métodos de selección, basados en las diferencias genéticas entre individuos dentro de las poblaciones, tienen una mayor complejidad, aunque presen-tan el atractivo, al menos teórico, de incrementos del carácter seleccionado continuados a lo largo de cierto número de generaciones.
La tasa anual de respuesta teórica (AG) en un carácter seleccionado viene expresada por la siguiente fórmula:
∆G = ihσpρ/t
siendo i la intensidad de selección, h2 y 2pσ la heredabilidad y la varianza fenotípica del
carácter, ρ, la eficiencia de la selección de los candidatos o correlación entre el índice utiliza-do y el valor mejorante de los mismos y t, el intervalo generacional.
En el caso del tamaño de camada en cerdos, se aceptan como parámetros usuales en razas de difusión universal: h2 = 0,10 y σp = 2,8. Asumiendo una selección de los candidatos

M. T. DOBAO et al.
122
basada en la media de los 2 primeros partos de sus madres (ρ = 0,21), y con una proporción seleccionada del l0 p. l00 en los machos y del 33 p. l00 en las hembras (i = 1,43), la res-puesta teórica predecible por generación es 0,26 lechones/camada (Bichard y David, 1985).
Pese a la baja heredabilidad del tamaño de camada (h2 = 0,l0), la considerable varianza fenotípica del carácter (σp = 8,1) posibilita una apreciable respuesta esperada, aun con un método de selección relativamente simple. Lamentablemente, estas optimistas perspectivas teóricas no se han confirmado hasta el momento experimentalmente (Vangen, 1981; Olli-vier y Bolet, 1981), ya que tanto los diferenciales de selección como los parámetros genéti-cos realizados resultan ser menores a los inicialmente previstos, por lo que las respuestas son muy inferiores a las esperadas.
A la vista del renovado interés existente por el carácter prolificidad y contando con las nuevas técnicas ahora disponibles, se han propuesto en los últimos años nuevas estrategias selectivas para la mejora del tamaño de camada en el cerdo (Haley et al., 1986):
1. Los grandes bancos de datos informatizados de países como Francia, Gran Bretaña, Noruega o Australia permiten la formación de líneas hiperprolíficas mediante fuertes inten-sidades de selección (i > 2,4) y evaluación con índices basados en el registro de hasta 4 partos por hembra (Legault y Gruand, 1976: Skjervold, 1983).
2. En los Estados Unidos (Oklahoma y Nebraska) se ha ensayado la selección indirecta para variables fisiológicas correlacionadas con el tamaño de camada, como tasas de ovula-ción y de supervivencia embrionaria, método que se orienta al aumento de la eficacia de la selección (ρ) y a la reducción del intervalo generacional (t) (Young et al., 1983; Johnson et al., 1984).
3. El uso dé índices familiares se ha propuesto como método idóneo para un carácter de baja heredabilidad y expresión limitada a un sexo, condiciones que favorecen la selección a partir de los datos de prolificidad de las hembras emparentadas con los candidatos, lo que aumenta la eficiencia de la selección hasta ρ = 0,36 en los índices más complejos, Con intensidades de selección similares a las previamente mencionadas, las tasas teóricas de respuesta anual son de hasta 0,50 lechones/camada (Avalos y Smith, [987).
La aplicación de estos métodos a poblaciones españolas de cerdos Ibéricos parece, por el momento, problemática, dadas las altas exigencias de infraestructura y manejo que plan-tean, prácticamente inalcanzables actualmente, Baste decir como referencia, que el banco de datos en que se apoya el programa francés de selección de una línea hiperprolifica controla alrededor del 39 p. l00 del censo nacional de cerdas madres: 800.000 camadas en 1984 (Legault. 1985). La selección para variables fisiológicas, por ahora en fase experimen-tal, plantea dificultades importantes en cuanto al registro de medidas como tasas ovulatorias o de mortalidad embrionaria, Otras medidas, aparentemente más sencillas, como el tamaño de testículos, parecen asimismo presentar correlaciones genéticas positivas con el tamaño de camada en cerdos Duroc y Yorkshire (Toelle y Robison, 1985; Young et al., 1986) por lo que podría resultar de interés su estudio experimental en el cerdo Ibérico.
La selección con índices familiares, que ofrece las mayores expectativas teóricas, en términos de tasas anuales de respuesta, plantea problemas organizativos complejos aunque no insuperables en el caso del cerdo Ibérico. Al examen detallado de este método dedicare-mos los siguientes apartados.

CRECIMIENTO Y CARACTERÍSITCAS DE CANAL DE CERDOS
Inv. Agrar.: Prod. Sanid. Anim., 2(1), 1987 123
ESTIMAS DE HEREDARILIDAD DEL TAMAÑO DE CAMADA EN CERDOS IBERICOS
Heredabilidad y correlaciones genéticas son los parámetros genéticos centrales en cual-quier programa de selección. La inexistencia de estimas de los valores de estos parámetros para caracteres reproductivos en el cerdo Ibérico, hace que la obtención de las mismas constituya tarea obligada a la hora de analizar un programa de selección para estos caracte-res.
En la Tabla 8 se presentan las estimas de heredabilidad obtenidas en un análisis prelimi-nar de datos de tamaño de camada procedentes de los 3 primeros partos de hembras lbéri-cas de la estirpe Torbiscal. Las estimas se obtuvieron mediante el empleo de los 2 métodos más usuales: regresión de madres e hijas y correlación entre medias hermanas de padre. Asimismo se calcularon por regresión, para las parejas madre/hija en que se dispone de datos de partos sucesivos, las heredabilidades de las medias de los partos (1-2), (2-3) y (1-2-3).
Dado el número de datos disponible, de 491 a 1.000 cerdas, y lo primario de los análisis realizados, en los que los datos no han sido corregidos para efectos ambientales, las estimas obtenidas, particularmente las calculadas por correlación, vienen afectadas por considera-bles errores típicos, que deberán ser reducidos en posteriores análisis con un mayor volu-men de datos y modelos más sofisticados. En cualquier caso, estos resultados preliminares son consistentes con las estimas obtenidas en poblaciones europeas, que cifran en 0,10 el valor de la heredabilidad del tamaño de camada en el cerdo (Hill y Webb, 1982; Bichard y David, 985; Vangen, 1986; Haley et al., 1987).
Asumiendo una correlación genética + 1,0 en la prolificidad de partos sucesivos, las heredabilidades esperadas en las medias de 2 o 3 partos serian de 0,17 y 0,23. Nuestros resultados indican valores algo inferiores, especialmente la media de los partos 1 y 2, lo que resulta coincidente con lo sugerido por algunos autores que consideran que el control genéti-co del tamaño de camada en el primer parto corresponde a genes parcialmente distintos a los que controlan la prolificidad de los partos posteriores (Tartar y Bolet, 1984). Los errores típicos muy elevados de las correlaciones genéticas estimadas en este análisis preliminar no permiten contrastar por el momento esta hipótesis, muy sugestiva en el caso del Ibérico, en
TABLA 8 ESTIMAS DE HEREDABILIDAD (h 2) DEL TAMAÑO DE CAMADA EN CERDO IBERICO
OBTENIDAS POR REGRESION MADRES/HIJAS Y POR CORRELACION ENTRE HERMANAS DE PADRE
Estimates of heritahility (h2) of litter size in Iberian breed from daughter/dam regression and paternal half-sib analyses
Regresión madres/hijas Correlación hermanas de padre
Partos (n) h2 ± s.e. (n) h2 ± s.e.
(1000) 0,05± 0,07 (1000) 0,19 ± 0,21 2 (709) 0,10± 0,08 (745) 0,06 ± 0,26 3 (491) 0,06± 0,09 (573) 0,06 ± 0,33 -2 (709) 0,03± 0,08 — — 2-3 (491) 0,11± 0,09 — — 1-2-3 (491) 0,16± 0.09 — —

M. T. DOBAO et al.
124
el que son muy acentuadas las diferencias de tamaño de camada entre los primeros partos: 7,20 ± 0,07; 7,92 ± 0,09 y 8,75 ± 0,09 lechones por camada, respectivamente, en cerdas de la estirpe Torbiscal.
INDICES FAMILIARES: RESPUESTAS TEORICAS EN CERDOS IBÉRICOS
La elevada tasa reproductiva de cerdos y conejos está en el origen del particular interés que presenta en estas especies el uso de índices de selección que combinan información familiar, El apareamiento de un cerdo puede realizarse a los 8 meses de edad, por lo que en el momento de su valoración como candidato a reproductor puede disponerse de informa-ción relativa a los 2 primeros partos de su madre, parida con 12 y 18 meses de edad, así como de otras hembras emparentadas, hermanas o medias hermanas, coetáneas de la misma.
Si suponemos que se dispone del registro de datos reproductivos de 3 hembras por camada y una proporción de reproductores de 1 verraco por l0 hembras, podemos conside-rar 5 índices familiares distintos, combinando la información correspondiente a los 2 pri-meros partos de la madre, índice Básico (II), y, adicionalmente, los datos de las 2 hermanas (12) y 27 medias hermanas (13) de la madre, así como de las 3 hermanas (14) y 27 medias hermanas (15) del padre.
En cuanto a intensidad de selección, se han considerado 2 proporciones de machos seleccionados: 1/10 (Caso A) y 1/30 (Caso B), así como el supuesto de selección dentro de familias de verracos con proporción 1/10 de machos seleccionados (Caso C). En los 3 casos se ha considerado una proporción 1/3 de hembras seleccionadas.
Las respuestas teóricas correspondientes a las combinaciones de los 5 índices (11 a 15) y 3 casos (A, B y C) descritos se presentan en la Tabla 9. Los valores esperados en lechones/ camada, son algo menores a las respuestas teóricamente esperadas en otras razas (Large White y Landrace), en las que es mayor la varianza fenotípica del carácter ( 2
pσ = 8,1),
TABLA 9 RESPUESTAS ESPERADAS POR GENERACION (LECHONES/CAMADA) CON EL EMPLEO
DE INDICES FAMILIARES DE SELECCION EN CERDO IBERICO Selection responses in piglets per litter per generation expected using different family selection indices in
Iberian pig
CasoA Caso B Indices
pm = 1 / 10 pm = 1 / 30 Caso C
Intrafamiliar (pm = 1 / 10)
Registros familia materna: Madre (I. Básico)...................... 11 0,18 0,22 0,17 + 2 hermanas madre ................. 12 0,21 0,25 0,20 + 27 medias hermanas .............. 13 0,24 0,28 0,22
Registros familia paterna:
+ 3 hermanas padre................... 14 0,29 0,33 0,24 + 27 medias hermanas .............. 15 0,32 0,36 0,25
Parámetros asumidas: h2 = 0,10; 2pσ =4,00; t 0.15.

CRECIMIENTO Y CARACTERÍSITCAS DE CANAL DE CERDOS
Inv. Agrar.: Prod. Sanid. Anim., 2(1), 1987 125
frente al valor 2pσ = 4,0 observado en el Ibérico (Avalos y Smith, 1987). Sin embargo, las
tasas de respuesta expresadas en porcentaje de la media del carácter son similares, en torno al 3-4 p. l00 anual, pues el coeficiente de variación del tamaño de camada en el cerdo Ibérico (CV = 0,27) es sólo ligeramente inferior al valor (CV = 0,28) comúnmente citado en trabajos con otras razas europeas. Estas tasas anuales de respuesta exceden, de hecho, a las esperadas en caracteres como crecimiento (2-3 p. l00) o proporción de magro (1-2 p. l00) (Smith, 1984).
El empleo práctico de este tipo de índices plantea algunos problemas, que pudieran resultar aún más acusados en su aplicación al cerdo Ibérico, dado el tamaño pequeño o moderado de las piaras.
Los problemas del uso de índices familiares en la selección para tamaño de camada en poblaciones finitas han sido estudiados recientemente por nuestro grupo mediante simula-clon en ordenador, suponiendo una población seleccionada de lo machos y 100 hembras, de estructura familiar y de datos similar a lo anteriormente indicado (Toro et al., 1988). Las respuestas observadas en la simulación de l0 generaciones de selección son inferiores a las teóricas, como resultado de dos efectos: a) los diferenciales de selección observados son menores que los esperados y b) el incremento de consanguinidad por generación (∆F) es hasta 3 veces superior al esperado sin selección, como se aprecia en los datos presentados en la Tabla l0. Como discutiremos en el siguiente apartado, la consanguinidad tiene un efecto detrimental sobre los caracteres reproductivos, por lo que de considerarse en la simulación un modelo con dominancia, las respuestas observadas llegan a ser, en los casos analizados, hasta un 60 p. l00 menores que las esperadas.
Existen igualmente considerables problemas operacionales relacionados con la dificultad para disponer en el momento de la evaluación de los candidatos del conjunto de la informa-ción requerida, pues cabe esperar pérdidas de información debidas a bajas, camadas falli-das, etc., lo que resultará en variación en el volumen y calidad de información disponible para evaluar cada candidato. Por último el registro de datos plantea ciertas necesidades de personal y de instalaciones cuya justificación desde el punto de vista práctico sólo puede proceder del valor de la mejora conseguida.
TABLA 10 TASAS DE CONSANGUINIDAD POR GENERACION (∆F, %) CALCULADAS POR
SIMULACION EMPLEANDO DIFERENTES INDICES FAMILIARES DE SELECCION EN UNA POBLACION SELECCIONADA DE 10 MACHOS Y 100 HEMBRAS
Rates of inbreeding per generation (∆F, %) obtained by simulation using different family selection indices in a selected herd of 10 males and 100 females
CasoA Caso B Indices
pm = 1 / 10 pm = 1 / 30 Caso C
Intrafamiliar (pm = 1 / 10)
11 1,63 2,47 1,30 12 1,93 2,78 1,53 13 2,25 3,05 1,92 14 2,97 3,88 2,01 15 3,37 3,95 2,17
∆F en ausencia de selección: 1,375 p.10

M. T. DOBAO et al.
126
CONSANGUINIDAD, CARACTERES REPRODUCTIVOS Y PROGRAMAS DE CONSERVACION
CONSANGUINDAD Y CARACTERES REPRODUCTIVOS EN EL CERDO IBERICO
La consanguinidad es una consecuencia inevitable de la cría de animales en poblaciones cerradas, particularmente, como ya hemos visto, cuando se realiza selección en las mismas. Existe un acuerdo general sobre la influencia negativa de la consanguinidad sobre los carac-teres más relacionados con la eficacia biológica, atribuible a dominancia direccional en los loci que controlan estos caracteres, Así, el tamaño de camada en mamíferos puede ser afectado tanto por la consanguinidad de las madres (Fd) a través de una menor tasa de ovulación o cambios desfavorables en el medio intrauterino, como por la consanguinidad de la camada (FI), resultante en una mayor mortalidad prenatal.
En el cerdo, los efectos negativos de la consanguinidad sobre la prolificidad son bien conocidos (Bereskin et al., 1970; Mikami et al., 1977). En el caso del cerdo Ibérico, el conocimiento preciso de los efectos de la consanguinidad sobre caracteres reproductivos tiene un interés singular, dada la necesidad de programas de conservación de líneas amenazadas de riesgo de extinción (Dobao et al., 1 987b).
Los archivos de la piara experimental de cerdo Ibérico de «El Dehesón del Encinar» ofrecen información muy valiosa para estos análisis. Explicada anteriormente su formación en 1945, cabe añadir que a partir de 1963, la piara quedó estructurada en 3 estirpes: Torhis-cal, resultado de la fusión de los cuatro grupos fundacionales, Guadyerbas, nueva denomi-nación del grupo Puebla fundacional, pelón guadianés y Gamito, estirpe lampiña configurada a partir de la anterior pero de capa colorada. Estas estirpes se mantuvieron con generaciones solapadas, tamaños efectivos respectivos de 36,14; 11,96 y 15,13 e intervalos generacionales de 2,24; 2,32 y 2,08 años. Los datos registrados hasta 1963 fueron analizados por Odriozola (1976) y los presentados aquí proceden de un análisis posterior realizado sobre 4.389 camadas de las estirpes nacidas entre 1974 y 1984, que incluyen algunas proce-dentes de apareamientos de consanguinidad forzada (Rodrigañez et al., 1985b).
Los efectos de a consanguinidad de la madre y de la camada fueron estudiados en cada estirpe mediante análisis de regresión múltiple, utilizándose los mismos modelos empleados por Odriozola (1976). Las variables dependientes consideradas son el número de lechones nacidos vivos por camada <Ny) y el peso en kilogramos de la camada a los 2 l días (W21). Ambos análisis incluyen las siguientes variables regresoras: P, Ordinal de la Paridera, Ad, Edad de la madre en trimestres y los Coeficientes de Consanguinidad de la madre (Fd) y de la camada (FI). expresados en porcentajes y calculados a partir de las genealogías mediante un programa de ordenador, atribuyéndose un valor F=0 a los animales fundadores. El modelo empleado en el análisis del peso de camada (W21) incluye asimismo la variable regresora N21, número de lechones vivos a 2 1 días, utilizándose en estos análisis únicamente los datos registrados en 1.604 camadas sin lechones adoptados y cedidos.
La Tabla 11 muestra, para las 3 estirpes, las medias e intervalos de variación de las variables consideradas en ambos análisis, y en la Tabla 12, se presentan los valores de los coeficientes de regresión parcial (b) de Nv y W21 sobre la consanguinidad de la madre (Fd) y de la camada (F1) sus errores típicos y niveles de significación estadística. El empleo de la técnica de probabilidades combinadas permite contrastar la hipótesis nula (Ho: b = 0) utili-zando conjuntamente los resultados de los análisis realizados independientemente en las 3

CRECIMIENTO Y CARACTERÍSITCAS DE CANAL DE CERDOS
Inv. Agrar.: Prod. Sanid. Anim., 2(1), 1987 127
TABLA 11 CONSANGUINIDAD Y CARACTERES REPRODUCTIVOS EN TRES ESTIRPES DE CERDO
IBERICO: MEDIAS E INTERVALOS DE VARIACION DE LAS VARIABLES CONSIDERADAS EN EL ANALISIS DEL NUMERO DE NACIDOS VIVOS (NV) Y EL PESO DE LA CAMADA A 21
DIAS (W21) (Rodrigáñez et al., 1985b)
Inbreeding and litter performance in three strains of Iberian pig: Means and ranges of variation of variables included in the analysis of litter size at birth (Nv) and litter weight at 21 days (W21)
(Rodrigañez et al., 1985b)
Torbiscal Guadyerbas Gamito
Variables Media Intervalo Media Intervalo Media Intervalo
a) Nacidos vivos (Nv) (N = 3029) (N = 720) (N = 640)
P................................... 78,6 58-96 74,3 58-96 73,2 58-96 Ad(3m) ........................ 10,7 4-35 9,9 4-30 9,9 4-27 Fd(%)........................... 10,8 5,3-42,1 25,4 15,8-44,1 33,1 22,7-56,6 Fl(%)............................ 12,0 6,9-53,6 28,0 20,2-52,0 35,2 27,6-60,5 ........................................ N 8,2 1-15 7,2 1-14 7,5 1-13
b) Peso camada 21d (W21): (N = 1090) (N = 293) (N = 221)
P................................... 79,3 58-96 74,8 58-96 73,1 58-96 Ad(3rn) ........................ 10,8 4-31 10,2 4-30 9,9 4-25 Fd(%)........................... 10,9 5,9-35,2 25,3 15,8-44,1 32,5 27,7-47,9 Fl(%)............................ 11,9 6,9-42,4 27,8 20,2-52,0 34,8 27,6-55,5 Nv ................................. 7,2 1-11 6,5 2-9 6,6 2-10 W21(kg) ........................ 30,9 3,9-47,5 27,6 3,9-45,4 28,6 5,1-43,4
estirpes. En la última columna de la Tabla 12 se presentan los resultados de este test conjun-to en los casos en que su auo resulta legitimo.
Hay un consistente efecto negativo sobre el tamaño de camada tanto de la consanguini-dad de la madre (Fd) como de la camada (FI). En el análisis previamente realizado por
TABLA 12 EFECTOS DE l0 p.100 DE AUMENTO DE LA CONSANGUINIDAD DE LA MADRE (Fd} Y DE LA CAMADA (F1) SOBRE EL NUMERO DE NACIDOS VIVOS (NV) Y EL PESO DE LA CAMADA
A 21 DÍAS (W21) EN TRES ESTIRPES DE CERDO IBERICO (Rodrigáñez et al. 1985b)
Effects of a .10 p 100 increase in dam (Fd) and litter (F1) inbreeding on litter size at birth and litter weight at 21 days in three strains of Iberian .pig (Rodrigañez et al., 1985b)
Torbiscal Guadyerbas Gamito Variables
b ± s.e. (P) (x10) b ± s.e. (P) (x10) b ± s.e. (P) (x10) Prob.
Combinada
Nacidos vivos: Fd -0,14 ± 0,08 (0,107) -0,24 ± 0,16 (0,133) -0,32 ± 017 (0,052) 0,025 FI -0,30 ± 0,09 (0,001) -0,36 ± 0,15 (0,020) -0,24 ± 0,16 (0,131) 0,0003
Peso a 21 d (kg): Fd -0,54 ± 0,27 (0,042) -1.12 ± 0,45 (0,013) -0,27 ± 0,70(0,707) 0,015 F1 -0,56 ± 0,29 (0,054) -1,00 ± 0,45 (0,029) -0,16 ± 0,63(0,802) -

M. T.DOBAO et al.
128
Odriozola (1976) sólo se encontraron efectos negativos significativos sobre el tamaño de camada de Fd y F1 en Guadyerbas, y de la consanguinidad de la madre (Fd) en Gamito. La mayor consistencia de los resultados del presente análisis puede atribuirse al más amplio intervalo de variación de los coeficientes de consanguinidad además del mayor número de datos disponibles.
Los efectos estimados sobre a variable W21, peso de camada a 21 días, son asimismo negativos y significativos, con la excepción del efecto de la consanguinidad de la camada en Gamito.
Las estimas obtenidas de los efectos de consanguinidad, indican que a un 10 p. 100 de incremento de la consanguinidad de la madre o de la camada corresponde a una reducción de -0,2 lechones nacidos vivos por camada, valor similar a los recogidos en la literatura (Hill y Webb, 1982).
MINIMIZACIÓN DE CONSANGUINIDAD EN PROGRAMAS DE CONSERVACION
Como hemos explicado en diversas ocasiones, la drástica reducción del censo de la población de cerdo Ibérico unido al establecimiento de una estructura poblacional basada en el cruzamiento industrial con razas extranjeras, ha llevado a la extinción de tipos de Ibérico como el rubio andaluz y a una situación de precariedad extrema de otros como el negro lampiño o el manchado de Jabugo. Ello haría necesaria la puesta en marcha de un Plan de Gestión de Recursos Genéticos de la Población Ibérica, que, entre otras cosas permitiera conservar los tipos más amenazados en poblaciones sometidas a un riguroso control genético, basado en la minimización del incremento de consanguinidad en las mis-mas (Dobao et al., 1 987b).
Los aspectos teóricos de la evolución de la consanguinidad y de los sistemas de aparea-miento que la minimizan han sido tratados en profundidad por la genética de poblaciones clásica. Si el objetivo de una población control es mantener el máximo número de alelos, esto es, minimizar la deriva genética, la mejor estrategia es subdividir la población en el máximo número posible de sublíneas permanentes, las cuales contendrían animales alta-mente consanguíneos (Robertson, 1964). En situaciones prácticas, sin embargo, el objetivo es más bien minimizar la disminución de heterocigosidad para evitar el deterioro de la eficacia biológica de las poblaciones conservadas. En este último caso es bien conocido que para minimizar el avance de la consanguinidad el factor más importante a tener en cuenta es la varianza del número de gametos contribuido por cada padre. El censo efectivo de población (Ne9 es, de acuerdo con la fórmula de Hill (1972):
∆F = l/2Ne = l/32M{2+ 2mmσ +2M/F cov (mm, mf) + (M/F)2 + 2
mfσ } +
+l/32F{2+(F/M)2 2fmσ + 2F/M cov(fm,ff) + ffσ }
En esta fórmula, válida estrictamente para censos altos, 2mmσ es la varianza del número
de hijos por padre, 2mfσ es la varianza del número de hijas por padre y cov(mm,mf) es la
covarianza de estos valores. Las cantidades correspondientes si consideramos la descenden-cia de las madres son 2
fmσ ffσ y cov(fm,ff). Aunque el mantenimiento de una estructura poblacional equilibrada es el criterio funda-
mental para la conservación de los recursos genéticos de una población, hay otras sugeren-cias como el apareamiento no aleatorio que pueden contribuir también a minimizar la

CRECIMIENTO Y CARACTERÍSITCAS DE CANAL DE CERDOS
Inv. Agrar.: Prod. Sanid. Anim., 2(1), 1987 129
consanguinidad. Wright (1921) determiné el avance del coeficiente de consanguinidad en aquellos sistemas en los que los individuos que se aparean tienen el menor parentesco posible y que denominó de máxima evitación de consanguinidad (y que aquí denominare-mos de mínimo parentesco) ya que pensé que dicho sistema minimizaba el incremento de la consanguinidad en todas las generaciones. Sin embargo, Kimura y Crow (1963) descu-brieron que los sistemas de apareamiento circular y circular en parejas producen un menor incremento de consanguinidad a largo plazo aunque no en las generaciones iniciales,
El apareamiento de mínimo parentesco, tal y como fue diseñado por Wright, sólo puede aplicarse a poblaciones de censo real que sea potencia de 2. En otras situaciones, para censos reducidos (menos de l0 machos y l0 hembras) y siempre que las facilidades computaciona-les lo permitan, hay que elaborar un programa de ordenador que elija, entre todas las combinaciones posibles, aquellas en las que las parejas están mínimamente emparentadas. Para censos mayores el cómputo puede facilitarse mediante el uso de técnicas de programa-ción lineal.
Los programas de conservación de las estirpes Torbiscal y Guadyerbas pueden constituir una referencia práctica útil.
En 1984 se decidió desglosar la estirpe Torhiscal en dos núcleos, uno de conservación y otro de selección. En el núcleo de conservación se consideró que debía mantenerse un censo efectivo que asegurara que, como mínimo, no se incrementase la tasa de endogamia a la que ha estado sometida dicha estirpe desde su fundación y que cálculos previos indican que es del orden del 1 p. 100 por generación. Por ello, se adoptó como objetivo un tamaño efectivo de Ne > 50, puesto que en la práctica el tamaño efectivo se verá disminuido por problemas prácticos inevitables: baja de algunos reproductores, fallos en las cubriciones, camadas con todos los individuos del mismo sexo, etc.
Hay muchas combinaciones de número de machos y hembras, tamaños familiares, etc, que producen un mismo tamaño efectivo. Pero como se ha señalado anteriormente lo más importante es conseguir que cada macho sea reemplazado por un hijo suyo y lo mismo ocurra con cada hembra. En nuestras circunstancias se consideró que censos reales de 15 machos y 45 hembras (a los que corresponde un censo efectivo de 72) presentaba varias ventajas: a) la tasa de endogamia esperada es algo menor que el 1 p. l00 por generación; b) la proporción de sexos 1:3 no resulta excesivamente gravosa en cuanto a su mantenimiento, posibilitando el suministro a las ganaderías o entidades interesadas de hasta 600 lechones/ año; c) al ser múltiplos de 3, las cifras de reproductores se ajustan cómodamente a una tasa de reposición de 1/3 anual, correspondiente a un intervalo generacional de 3 años que se considera idóneo y d) estos valores están comprendidos entre los recomendados internacio-nalmente (Maijala, 1974).
Parece, por tanto, de interés considerar el efecto de diferentes sistemas de apareamiento en la evolución del coeficiente de consanguinidad de poblaciones equilibradas. Para ello se ha preparado un programa de ordenador que calcula el avance de la consanguinidad me-diante las bien conocidas relaciones de recurrencia de los parentescos explicadas en Falco-ner (1960). En la Tabla 13 se presenta la evolución del coeficiente de consanguinidad en condiciones de apareamiento al azar (basado en 200 repeticiones), apareamiento circular en parejas (CP) y apareamiento de mínimo parentesco (MP) evaluadas para poblaciones de relaciones de parentesco y efectivo de reproductores análogos a los de las estirpes Guadyer-bas (Nm = 7; Nf = 21) y Torbiscal (Nm = 16; Nfr = 48).
A corto plazo el sistema de apareamiento MP puede suponer una contribución no

M. T.DOBAO et al.
130
TABLA 13 EVOLUCION SIMULADA DE LA CONSANGUINIDAD EN LAS ESTIRPES GUADYERBAS Y TORBISCAL. CON TRES MÉTODOS DE APAREAMIENTO: AL AZAR (AZ), CIRCULAR EN
PAREJAS (CP) Y MINIMO PARENTESCO (MP)
Simulated evolution of inbreeding in the Guadyerbas and Torbiscal strains ,with three different mating systems: random (ÁZ). circular (CF) and maximum avoidance of inbreeding (MP)
Tipo de Generaciones
Estirpe Apareamiento 0 5 15 20 100
Guadyerbas (Nm =7; Nf = 21) Azar (AZ) 0,29 0,38 0,47 0,5! 0,26 Circular (CP) 0,29 0,42 0,52 0,56 0,89 Min. parentesco (MP) 0,29 0,35 0,43 0,47 0,82
Torbiscal (Nm =16; Nf = 48) Azar (AZ) 0,12 0,15 0,21 0,23 0,55 Circular (CP) 0,12 0,25 0,38 0,42 0,67 Mm. parentesco (MP) 0,12 0,13 0,18 0,20 0,50
desdeñable al mantenimiento de heterocigosidad respecto al sistema de apareamientos al azar. A largo plazo, este último es superior aunque la diferencia respecto al apareamiento al azar es escasa, siendo la generación a la que esta superioridad empieza a manifestarse mayor conforme el censo aumenta. Aunque el apareamiento circular en parejas produce un menor incremento de a consanguinidad a muy largo plazo, los incrementos de la misma en las generaciones iniciales son bastante altos, por lo que su utilidad en programas de conserva-ción es dudosa a pesar de ser frecuentemente recomendado.
Existen modificaciones de estos 3 tipos básicos de apareamiento como los apareamien-tos cíclicos que se utilizan en poblaciones control de ratones que pueden ser adecuadas en algunas circunstancias por su simplicidad pero que no superan al sistema MP mientras que presentan la desventaja de manifestar ciclos de aumento y disminución de la consanguini-dad que no son deseables en un programa de conservación.
DISCUSION
Las expectativas de los distintos métodos analizados en cuanto a la mejora genética de la prolificidad del cerdo Ibérico, pueden resumirse como sigue:
i) el cruce de líneas de Ibérico posibilita aumentos de tamaño de camada superiores a los 0,50 lechones, equivalentes al 6-7 p. l00 de la media del carácter, atribuibles a heterosis individual o directa.
ii) la superior prolificidad de las hembras híbridas Jiaxing por Ibérico respecto a cerdas Ibéricas, se cifra en un incremento de 3 lechones por camada equivalente al 35-40 p, l00 de la media.
iii) entre los diversos métodos de selección para tamaño de camada, la selección me-diante índices familiares ofrece las mayores posibilidades, con respuestas teóricas por gene-ración comprendidas entre 0,20 y 0,36 lechones por camada, equivalentes al 2,5-4,5 p. l00 de la media.
El interés práctico del cruzamiento entre líneas de Ibérico para aquellos ganaderos inte

CRECIMIENTO Y CARACTERÍSITCAS DE CANAL DE CERDOS
Inv. Agrar.: Prod. Sanid. Anim., 2(1), 1987 131
resados en disponer como productos terminales de animales Ibéricos puros viene reforzado por el efecto positivo adicional de la heterosis individual sobre los caracteres de crecimiento y canal, de acuerdo con los resultados obtenidos por Odriozo!a (1956).
Por el contrarío, el cruzamiento inicialmente ensayado de hembras híbridas JXxIB con verracos Ibéricos resulta no satisfactorio al valorarse el conjunto de ciclo productivo. La mayor prolificidad de estas hembras se traduce en una reducción sustancial del coste unita-no por lechón, que puede valorarse en 500-1.200 ptas/lechón, para costes anuales de mante-nimiento por cerda madre comprendidos entre 20.000 y 50.000 pesetas. Sin embargo, las diferencias registradas en el peso de sacrificio entre animales l/4JX,3/4IB e Ibéricos de la misma edad, equivalen a una reducción del margen bruto de beneficio por cerdo sacrificado entre 3.800 y 5.700 pesetas, para el amplio intervalo de precios de la última campaña (200-310 ptas/kg). Esta reducción del margen de beneficio supera ampliamente la disminu-ción del coste unitario por lechón.
El aprovechamiento del superior potencial reproductivo de las hembras de origen Jia-xing, así como de las características positivas de estos animales en cuanto a rusticidad y buena calidad de carne, requiere por lo tanto el ensayo de otros esquemas de cruza-miento.
En primer lugar, la utilización como reproductoras de hembras l/4JX, 3/4IB para la obtención como productos terminales de animales l/8JX en los que cabe esperar menos dificultades para alcanzar el elevado peso de sacrificio (150-170kg) requerido por el merca-do de productos de tipo Ibérico, Una segunda alternativa sería el cruzamiento de las hem-bras 1/2JX, l/2IB por verracos Duroc-Jersey, lo que permitiría disponer como productos terminales de animales de superior crecimiento. La aplicación práctica de este esquema de cruzamiento a 3 vías, posiblemente el de mayores ventajas productivas, estaría condiciona-da, más allá de su evaluación experimental, a la deseable clarificación del mercado de productos ibéricos. En nuestra opinión, sólo en un contexto de mayor transparencia de este mercado y de protección efectiva de la población de cerdos Ibéricos, estaría justificada la introducción de esquemas de cruzamiento con nuevas razas.
Las tasas de respuesta esperadas con el uso de índices de selección que combinan infor-mación familiar presentan un gran atractivo, en panicular si se considera su carácter acu-mulativo, Sin embargo, los resultados obtenidos por simulación, indican que las respuestas observadas pueden ser sensiblemente inferiores a las teóricas, siendo mayores las discrepan-cias para los índices más complejos, que deberían ser utilizados cautelosamente, en particu-lar con objeto de reducir los incrementos de consanguinidad (Toro et al., 1988). En este sentido, cuando existen restricciones al aumento del número de reproductores, puede resul-tar aconsejable el empleo de tácticas como la selección intrafamilias de verracos (Caso C), el apareamiento de reproductores de mínimo parentesco o una superior proporción (1/5) de machos y hembras,
Existen igualmente considerables problemas operacionales relacionados con la dificultad para disponer en el momento de la evaluación de los candidatos del conjunto de la informa-ción requerida, pues cabe esperar pérdidas de información, debidas a bajas, camadas fallidas, etcétera, lo que resultará en variación en el volumen y calidad de información disponible para evaluar cada candidato. Por último, dada la limitada infraestructura de las piaras de cerdo Ibérico, el registro de datos plantea ciertas necesidades de personal y de instalaciones cuya justificación desde el punto de vista práctico sólo puede proceder del valor de la mejora conseguida.

M. T.DOBAO et al.
132
Los efectos detrimentales de la consanguinidad que han sido evidenciados sobre caracte-res como la prolificidad o el peso de la carnada son de relevancia en cuanto a la conserva-ción de estirpes o tipos diferenciados de cerdo Ibérico, generalmente en forma de poblacio-nes de tamaño reducido. La descripción de los programas de conservación de las estirpes Torbiscal y Guadyerbas indica las reglas básicas que, a nuestro juicio, deben orientar este tipo de programas: a) maximización del censo efectivo con mantenimiento de una estructu-ra familiar equilibrada y anulación de la varianza de los tamaños familiares mediante la reposición de cada reproductor por un hijo de su mismo sexo y b) apareamiento, en cada generación, de la combinación de reproductores de mínimo coeficiente de parentesco.
AGRADECIMIENTOS Los autores expresan su gratitud a la colaboración del personal del Centro de Investigación Agrope-
cuaria «Dehesón del Encinar», así como a La Consejería de Agricultura de la Junta de Comunidades de Castilla La Mancha, por el apoyo prestado a la continuidad de la piara experimental.
La obtención de hembras F1(JXxIB) ha sido posible con la ayuda de C. Legault, J.C. Caritez y J. L. Vrillon del INRA, así como de nuestros compañeros S. Martin-Rillo y E. Alias del Departamento de Reproducción Animal del INIA.
Debemos asimismo agradecer su colaboración a Paula Vázquez y a Carmen García y a la Sección de Proceso de Datos del INIA.
El trabajo experimental ha estado subvencionado por el INIA y la CAICYT (Proyectos núms. 5972, 4082/0229/Sl y 5205/2958/83.
SUMMARY
Genetics of prolificacy in the Iberian pig: A revision of methodologies and results
The analysis of a complete diallelic cross among four old Iberian pig strains evidences an individual heterosis of 0,40 piglets per litter which suggests that crossing among Iberian lines is the simplest method to improve prolificacy for those breeders interested on pure Iberian pigs production.
The utilization of crosses with Taihu breeds offers better perspectives, the average litter size for the three first parities of Iberian (IB) and hybrid Jiaxing x Iberian (JXxlB) sows are 8,5 ± 0,3 and 11,4±0,4 piglets respectively. But the backcross of these hybrid sows with, Iberian boars is not satisfactory because of the scarce growth, rate and poor carcass conformation of the terminal products (l/4JX, 3/41B). Alternative crossing schemes must be essayed.
The use of family selection indices allows theoretical responses per generation from 0,20 to 0,36 piglets per litter, using available parameter estimates from Iberian pig populations (h2 = 0,10 2
pσ = 4,00; t= 0,15). Although simulation studies evidence some problems in the use of these indices in populations of low or moderate size: selection differentials lower and higher inbreeding rates than expected ones.
A disminution of.20 piglets per 10 p. 100 increase of dam or litter inbreeding has been estimated from data recorded in three closed small size populations (Ne~ 1.96; 15.13 and 36.14). Different methods of minimization of the inbreeding rate are analyzed in two programs of breed conservation. The maintenance of an equilibrated family structure and minimum co-ancestry matings in each gene-ration are proposed as basic norms for this type of programs.
KEY WORDS: Prolificacy, Iberian pig. inbreeding, crossbreeding, selection.
REFERENCIAS BIBLIOGRAFICAS
ANTUNES-CORREIA J.C., OLIVEIRA A. R., 1985. Evoluçao da consanguinidade en nucleos fecha-dos de porcos alentejanos. XXI Jornadas Luso-Españolas de Genética. Vila-Real, 24-26 septiem-bre.

CRECIMIENTO Y CARACTERÍSITCAS DE CANAL DE CERDOS
Inv. Agrar.: Prod. Sanid. Anim., 2(1), 1987 133
AVALOS E., SMITH C., 1987. Genetic improvement of litter size in pigs. Animal Production, 44,153-64. BERESKIN B., CHELBY C. E., ROWE K. E., REMPEL W, E., DETTMERS A. E., NORTON H. W.,
1970. Inbreeding and swine production in Minnesota experimental herds. Journal of Animal Science, 31, 278-288.
BICHARD M., DAVID P. J., 1985. Effectiveness of genetic selection for prolificacy in pigs. Journal of Reproduction and Fertility, Suppl., 33, 127-138.
BIDANEL J, P., LEGAULT C., 1986. Experimental and prospective aspects of the utilization ofproli-ftc Chinese pig breeds a Europe. 37th Annual Meeting of the EAAP, Budapest, 1-4 septiembre.
BOLET G., MARTINAT-BOTTE F., LOCATELLI P., CORNAND J., TERQUI M., BERTHCLOT F., 1986. Components of prolificacy in hyperprolific Large White sows compared with the Meishan and Large White breeds. Génétique, Séléction, Evolution. 18(3), 333-342.
BULMER M. G., 1980. The Mathematical Theory of Quantitative Genetics. Clarendon Press. Oxford. CHENG P., ‘983. A highly prolific breed of China. The Taihu pig. Parts I and II. Pig News and
Information, 4. 407-416. DOBAO M. T., RODRIGAÑEZ J., SILIO L., 983. Seasonal influence on fecundity and litter perfor-
mance characteristics in Iberian pigs. Livestock Production Science, 10, 601-610. DOBAO M. T., RODRIGAÑEZ J., SILIO L., TORO M. A., 986. Heterosis para número de mamas
en 4 estirpes de cerdo Ibérico, XXII Jornadas Luso-Españolas de Genética, Oviedo, 30 septiembre-20 octubre.
DOMO M. T., RODRIGAÑEZ 3., SILIO L., TORO M. A., de PEDRO E., GARCIA DE SILES J. L., 1987a, Crecimiento y características de canal en cerdos Ibéricos, Duroc-Jersey x Ibérico y Jiaxing x Ibérico. Investigación Agraria, Serie Producción y Sanidad Animal, 2(l), 9-23.
DOBAO M. T., RODRIGAÑEZ J., SILIO L. TORO M. A., l987b. Implicaciones del cambio de estructura de población en La conservación del cerdo Ibérico. En Conservación y desarrollo de las dehesas portuguesa y española. (Eds. P. Campos Palacin y M. Martin Bellido): 181-188. Ministerio de Agricultura, Madrid.
FALCONER D, 5., 960. Introduction to Quantitative Genetics. Oliver and Boyd, Edimburgo. HALEY C. S., AVALOS E., SMITH C., 1986. A review of selection for reproductive performance
in the pig. 37th Annual Meeting of EAAP, Budapest, -4 septiembre. HILL W. G., 1972. Estimation of genetic change. 1. General theory and design of control populations.
Animal Breeding Abstracts, 40, 1-l5. HILL W. G., WEEB A, J., 1982. Genetics of reproduction in the pig. En Control of Pig Reproduction.
(Eds. D. J. A, Cole y G. R. Foxcroft): 541-564. Butterworth, Londres. JOHNSON R. K., ZIMMERMAN D, R., KJTTOK R. J., 1984. Selection for components of reproduc-
tion in swine. Livestock Production Science, 11, 54-558. KIMURA M., CROW J. F., 1963. On the maximum avoidance of inbreeding. Genetical Research, 4,
399 -4 15. KING J. W. B., AVALOS E., 1986, Breed resources: use of breeds as dam lines. 3rd World Congress on
Genetics Applied to Livestock Production, Lincoln, Nebraska, X, 26-34. LEGAULT C., 1985. Selection of breeds, strains and individual pigs for prolificacy. Journal of Repro-
duction and Fertility, Suppl. 33: 151-166, LEGAULT C., CARITEZ J.C., 1983. L' expérimentation sur e port chinois en France. 1. Performan-
ces de reproduction en race pure et en croisement. Génétique, Séléction, Evolution, 15, 225-240. LECAULT C., GRUAND J., 1976. Amélioration de la prolificité des truies par la création d'une
lignée «hyperprolifique» et l'usage de l’insémination artificielle: principe et résultats expérimentaux préliminaires. Journées Recherche Porcine en France, 8, 201-212.
LEGAULT C., SEILIER P., CAR]TEZ J.C., DANDO P., GRUAND J., 1985. L'expérimentation sur le porc chinois en France. II. Performances de production en croisement avec les races européen-nes. Génétique Séléction, Evolution, 17, 133-152.
LEGAUI,T C., SELLIER P., 1986. The Chinese prolific breeds of pigs: examples of extreme genetic stocks, En Exploiting New Technologies in Animal Breeding. (Eds. 5. C. Smith, J, W. 3. King y J.C. McKay» 153-162. Oxford University Press, Oxford.
MAIJALA K., 1974, Conservation of animal breeds in general. l’~ World Congress on Genetics Ap-plied to Livestock Production, II, 37-46.
MIKAMI H., FREDEEN H.T., SATHER A, P., 1977. Mass selection in a pig population. 2. The

M. T.DOBAO et al.
134
effects of inbreeding within the selected populations. Canadian Journal of Animal Science. 57, 627-634,
ODRIOZOLA M., 1956, The significance of the phenomenon of heterosis and of environmental condi-tions, particularly that of climate, on the quality of pig carcasses. Publicaciones del VII Congreso Internacional de Zootecnia, Madrid.
ODRIOZOLA M., 1976. Investigación sobre los datos acumulados en dos piaras experimentales. IRY-DA, Madrid.
OLLIVIER L., BOLET G., 1981. La sélection sur la prolificité chez le porc: résultats d’une expérience de sélection sur dix générations. Journées Recherche Porcine en France, 3, 261-268.
ROBERTSON A., 1964, The effect of non-random mating within inbred lines on the rate of inbree-ding. Genetical Research, 5, 164-167,
RODRIGAÑEZ J., DOBAO M. T., SILIO L, TORO M. A., 1985a. Heterosis para prolificidad en cuatro antiguas estirpes de cerdo Ibérico. ITEA, 5, 104-106.
RODRIGANEZ J., DOBAO M. T., SILIO L., TORO M. A., 1985b. Consanguinidad y caracteres reproductivos en el cerdo Ibérico. ITEA. 5.107-109.
SELLIER P,. 1976. The basis of crossbreeding in pigs: a review. Livestock Production Science, 3,203-226,
SKJERVOLD H., 1983. A breeding scheme to improve prolificacy in pigs by intense sire and dam selection, Pig News Information, 4 (4). 427-430.
SMITH C., 1984. Rates of genetic improvement in farm livestock, Research and Development in Agriculture, 1, 79-85,
SUTHERLAND R, A., WEBB A, J., KING J. W. 8., 985. A survey of world pig breeds and compan-sons. Animal Breeding Abstracts, 53, 1-22.
TARTAR M., BOLET G., 1984. Application de la théorie des indices de séléction à des caractères répétés. Génétique, Séléction. Evolution, 16, 319-334.
TOELLE V. D., ROBISON O, W., 198$, Estimates of genetic relationships between testes measure-ments and female reproductive traits in swine. Journal of Animal Breeding and Genetics, 102, 125-130,
TORO M. A., DOBAO M. T., RODRIGAÑEZ J., SILIO L., 1986, Heritability of a canalized trait: teat number in Iberian pigs. Génétique, Séléction, Evolution, 18, 173-184.
TORO M, A., SILIO L., RODRIGAÑEZ J., DOBAO M, T., 988, Inbreeding and family index selection for prolificacy in pigs. Animal Production, 46(l), (en prensa).
VANGEN O., 1981. Problems and possibilities for selection for fecundity in multiparous species. Pig News and Information, 2, 257-263.
VANGEN O., 1986. Genetic control of reproduction iii pigs from parturition to puberty. 3rd World Congress on Genetics Applied to Livestock Production, XI. 168-179,
WRIGHT 5., 1921, Systems of mating, Genetics, 6, 111-178. YOUNG L. D., PUMFREV R. A., CLJNNINGHAM P. J., ZIMMERMAN D, R., 1978. Heritabilities
and genetic and phenotypic correlations for prebreeding traits, reproductive traits and principal components. Journal of Animal Science, 46, 937-949.
YOUNG L. D., LEYMASTER K. A., LUNSTRA D. D., 1986, Genetic variation in testicular develop-ment and its relationship to female reproductive traits in swine. Journal of Animal Science, 63, 17-26.
ZHANG W. C., WU J, S, REMPEL W. E., 1983. Some performance characteristics of prolific breeds of pigs in China, Livestock Production Science. l0, 59-68.





























