Embed Size (px)
Citation preview

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 1/82
serie de biologia monografia no. 15
Programa Regional de Desarrollo Cientrfico y Tecnol6gico
Departamento de Asuntos Cientrficos
Secretarfa General de la
Organizaci6n de los Estados Americanos

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 2/82
G E N E T I C A
D E P O B L A C I O N E S H U M A N A S
por
Francisco Rothhammer
Departamento de Biologia Celular y GemHica
Facultad de Medicina
Universidad de Chile
e
Instituto de Ecologia y Evolucion
Universidad Austral de Chile
Santiago, CHILE
Programa Regional de Desarrollo Cientifico y Tecnoloqico
Departamento de Asuntos Cientificos
Secretaria General de la
Organizacion de los Estados Americanos
Washington, D.C. - 1977

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 3/82
CC'Copyright1977 by
The General Secretariat of the
Organization of American States
Washington, D.C.
Derechos Reservados, 1977Secreta ria General de la
Organizaci6n de los Estados Americanos
Washington, D.C.
Esta monografia ha sido preparada para su publicaci6n en el
Departamento de Asuntos Cientfficos de laSecretaria General
de la Organizaci6n de los Estados Americanos
Editora: EvaV. Chesneau
Asesor Tacnico: Prof. Danko Brncic
Departamento de Genetica
Universidade Federal do Rio Grande do Sui
Porto Alegre, Rio Grande do Sui, Brasil

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 4/82
El programa de rrron og r-affa s ci e ntffi ca s e s una faceta de la va s ta la-
bor de Ia Or-ga ni zac idn de los Estados Americanos, a cargo del Depa r-
tamento de Asuntos Cie ntff'i c o s de la Sec r eta rfa General de di cha
Organi~aci6n, a cuyo financiarniento contribuye en forma importante el
Prograrna Regional de Desarrollo Cientffico y Tecno16gico.
Goncebido pur los Jef"s de Estado Arne r t ce.no s en su Reuni6n cele-
brada en Punta del Este, Uruguay, en 1967, y c ris ta Ii zad o en las deli-
beraciones y rria nda to s de la Quinta Reuni6n del Consej 0 Interamericano
Cultural, llevada a cabo en Ma r a c ay , Ve ne aue Ia , en 1968, e l Programa
Regional de De sa rrolIo Ci entff'i c0 y Tecno16gico e s Ia expresi6n de las
aspiraciones pr e c orri za da s por los Jefes de Estaclo Americanos en el
sentido de pone r Ia ciencia y la t ecnolog Ia al servicio de los pueblos
Ia tinoame rica no s ,
Demostrando gran visi6n, di c ho s clignatarios reconocieron que la
ciencia y Ia te crio l og Ia e atan transformanclo Ia estructura econ6mica y
s oc ia l de muchas naciones y que, en e s ta hora, par ser instrumento in-
dispensable de progreso en America Latina, neeesitan un 'irnpu l s o sin
preeedentes.
IIIEI Programa Regional de Desarrollo Ci entffi co y Tecno16gico es un
complemento de los e sfuerzos na ciona.Ie s de los parses latinoame ricanos
y se orienta hacia la adopci6n de rrie di.da s que permitan e l fo:rnento de la
investigaci6n, la ense1'\anza y la difusi6n de la ciencia y Ia tecnologfa; la
formaci6n y pe rfeccionamiento de personal cierit if'i c0; el intercambio de
informacione s, y la transfe reneia y adaptaci6n a los parses latinoa:rne-
ricanos del eonocirniento y las rec nol ogfa s generadas en otras regiones.
En el cumplimiento de estas premisas furrdarrie nta le s , el programa
de rnonog r affa e repre s e nta una contribuci 6n di recta a Ia ens eilanza de la s
ciencia 5 en niveles educati vos que aba rca n inp or ta nt fs i.rrros sec to r e s de
la poblaci6n y, al rriis rrio tierrrp o , p ropugna la difusi6n del sabe r cicntrfico.
La colecci6n de rnoriog r affa s cientffica s cons ta de cuatro serie s, en
espa1'\ol y po r tugue s , sobre terna s de frsica, qufrrri ca , biologla y :rnate-
matiea. De sde sus corni enz os , e stas obra s se destinaro s a profe sore s
y alumnos de ciencia s de eneena n ea secunda ria y de los primeros a1'\os
de la univer sitaria; de e st os se tiene ya te s t. i rn orrio de 8U buena acogida.
Esta introducei6n brinda al Pr og r arna Regional de Desarrollo Cientr-
fi c o y Tecno16gico de la Secretarla General de la Organizaci6n de 108
Estados Americanos l.a ocasi6n de agradeeer al doctor Francisco
Rothhammer, autor de esta monog r-arta , y a quienes tengan el Inre r e s ybuena voluntad de contribuir a su divulgaci6n.
rna r zo de 1977

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 5/82
INDICE
A los Lectores ..
CAPITULO PRIMERO. INTRODUCCION .
CAPITULO SEGUNDO . ANTECEDENTESH ISTOR ICOS ...
Las Contribuciones de Darwin y Mendel _ .
Mendelistas y Biometristas .
La Genetica de Poblac::iones .Emerge corrio Ciencia
lndependiente _. _.... _. . . . . .. .. _ _.. _..
Se Afianza 121.Nueva Cienda . . . .. . .
Hacia Una Srntesis _Clasicistas y Balancistas .
Seleccionistas y Neutra Iista s
CAPITULO TERCERO. POBLACIONES MENDELIANAS
Y LA LEY DE HARDY-WEINBERG _ '.' .
Algunas Definiciones Basicas , , .
Sobre Frecuencias Fenotlpicas, Genotlpicas y
Genicas _ _.
Un Pritner Intento de Hacer Fredicciones , .
Las Suposiciones Irnpli"citas en el Teorerna de
Hardy-Weinberg... . . , .
Generalizad6n deI Teorema de Hardy- Weinbe rg
La Distribuci6n de Hardy-Weinberg Corresponde a
Ia Hi.potesis de Inercia Evolutiva _ _ .
CAPITULO CUARTO. MUTACIONES Y SELECCION
NATURAL .
Clasificaci6n de los Factore s Evolutivos .
Evolud6n por Mutaciones . _ , _.. _ .
Condiciones pa.r'a el Es tablecirniento de Un Equilibria
Entre Mutaciones y Selecci6n Natural. _ , .La Estimaci6n de Tasas Mutacionales _.. , ..
La Estimaci6n de Coeficientes de Selecci6n - ..
CAPITULO QUINTO. POLIMORFISMOS GENETIGOS
La Definicion de Polirnorfistno Genetico - .
Algunos Mecanisrnos de Mantenci6n de Polimorfismos
Estables _ ,. . .
Condiciones para Ia Estabilidad de Un PolimorfisIno ..
EI Coeficiente de Retorno 21.1Equilibrio - .Niveles de Variabilidad Polirn6rfica .
Polimorfismos Fosiblernente Mantenidos por
Ventaja Hete rocigota , _ -
Otros F'act or e s que Originan Po lirno rHsmo s .
La Relajaci6n de la Selecci6n Natural .
Pagina
iii
3
3
4
5
5
66
7
9
9
9
10
V12
13
is
17
17
17
1821
22
25
25
25
27
29
30
33
34
35

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 6/82
CAPITULO SEXTO. CONSANGUINIDAD.................. 37
Matrimonios Con s arigufne o s . . . . . . . . . . . . . . . . . . . . . . . . . . 37
E1 Coeficiente de Endogamia 37
E1 Me to do de Kudo para Ca1cu1ar e1 Coeficiente
de Endogamia 40
E1 Efecto de la Endogamia a Nive1 Poblacional 43
El Coeficiente de Endogamia de la Poblacion . . . . . . . . . . . 44
Cargas Gen et.i c a s 44
. . . . . . . . . , .CAPITULO SEPTIMO. CONSTITUCION GENETICA Y
MEDIO AMBIENTE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
Consideraciones Generales. . . . . . . . . . . . . . . . . . . . . . . . . . . 49
El Modelo Clasico de Herencia P'o Iig entc a 50
Algunos Me tod os para Estimar la Heredabilidad y
la Determinacion Ge ne tica . . . . . . . . . . . . . . . . . . . . . . . . . . 52
UtiIi za ci on de Mellizos Mono cigotic os y Di ci.go t.i c os
para Estimar la De te r rrrina.ci.on Ge n etic a 53
Estimacion de la Determinacion Ge ne ti ca a Partir de
Estudios con Hijos Adoptivos . . . . . . . . . . . . . . . . . . . . . . . . 53
La Obte n cion de Estirnadores de la Determinacion
Gene ti ca a Partir de Correlaciones Familiares 53
CAPITULO ,OCTAVO. FACTORES EVOLUTIVOS
ESTOCASTICOS " . . . .. . . . . .. . 57
VI El Concepto de Deriva Ge ne tica .
Un Pequeno Experimento que !lustra la Deriva .
P'r ed icci cn de los Efectos de la Deriva .
La Probabilidad de F'Lja cion de Un Gen Mutante .
Deriva Ge ne ti ca en Poblaciones Subdivididas .
Modelos de Estructura Poblacional .
'I'a.rnafio de Pob la ci Sn Efectivo .
57
57
58
59
60
62
63
CAPITULO NOVENO. ESTIMACI6N DE FRECUENCIAS
GENICAS 65
Estadlstica y Gerie tica de Poblaciones . . . . . . . .. . . . . . 65
Propiedades de Un Estimador 66
Estimacion de Frecuencias Gerii ca.s de Loci Di.ale Ii co sCodominantes 66
Es tima cton de Frecuencias Ge nica s de Loci Di aIe Ii co s
Dominantes 67
Estimaci6n de Frecuencias Geni cas de Loci
Codominantes Ligados al Sexo . . . . . . . . . . . . . . . . . . . . . . . 67
Estimaci6n de Frecuencias Ge ni ca s de Loci
e exo .
Estimaci6n de Frecuencias Genicas de Loci
Mu lt.i.a l.eIi co s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68
Bibliografi"a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
Agradecimientos ..... 74

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 7/82
1INTRODUCCI6N
lA que se debe que los seres hurna no s sean individualITlente consi-
derados rini co s y sin ernba r g o se puedan agrupar en poblacione s dis tin-
tas? ~C6ITlo se origin6 esta variabilidad? lEs estable 0 ca.rnbia con
el tiempo? lQue mecanismos la mantienen? lQue consecuencias ha
tenido la evoluci6n cultural sobre la evoluci6n biol6gica hurri arra ? Estas
son algunas de la preguntas a las que pretende dar respuesta la gene-
tica de poblaciones. M. Ki rnu r-a y T. Ohta(l} definen Ia g en etic a de po-
blaciones como "aquella r arna de la g en et ic a cuyo objeto es investigar
las leyes que gobiernan l a estructura g en eti ca de poblaciones naturales
y mediante ellas clarificar los mecanismos de la evoluci6n".
Si se acepta con Dobzhansky(2} que "evoluci6n es el carnb io de la
composici6n genet ica de las poblaciones" se podrla deIinir en forma
mas gene ra l la ge net ica de poblaciones como la rama de la bio log Ia evo-
lutiva que investiga las leyes de t.raneformaci.on que rigen el proceso
evolutivo. 5i bien es cierto que la gerret ica de poblaciones e s una sub-
disciplina importante de la biologla evolutiva, no conviene exa ge r a r su
alcance. Lewontin(3} destaca que la genetica de poblaciones ha contri-
buido poco a I conocimiento de los procesos de es pe c ia ci on y nada al co-
nocimiento de la ext ln c i.on de las razas yespecies. No se puede poner
en duda , sin embargo, que la e spe c ia cion y la ext in ci on son aspectos
fundamentales del proceso evolutivo. El progreso de la gerie tica de po-
blaciones de pe nd e r a en gran parte de la claridad con que se especifi-
quen sus objetivos. Lewontin (3) esquematiza la estructura de la teorla
geneticopoblacional como
G 1
1 ;
Fig. 1. Esquema de la estructura de la teorla geneticopoblacional.
donde 0 y F representan descripciones genotlpicas y fe not fpica s de di-
ferentes estados del sistema evolutivo, y T las leyes de transformaci6n
a que se ha aludido con anterioridad. En este esquema 01 representa
la composici6n g enet l ca de una poblaci6n en un instante determinado.
En primer lugar, es necesario conocer un conjunto de leyes epigeneti-
a 01 en diferentes ambientes. 1'2 indica las leyes de apareamiento, mi-
graci6n y selecci6n natural que regulan las transformaciones del con-
junto de fenotipos en el lapso de una generaci6n. Por otra parte, 1'3
representa un conjunto imponente de leyes que permiten establecer una
relaci6n entre la distribuci6n de fenotipos F ' ? , y la composici6n gen ett ca
subyacente, O2, Finalmente, las leyes 1 '4 son las leyes de Mendel y
Morgan mediante las cuales se puede en principio predecir la distribu-
ciori de genotipos O~, a partir de los genotipos de la gene r a ci on anterior,

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 8/82
G2• Estas ultimas son las mejor conocidas. En estamonografi"ase in-
t.errtara revisar algunas leyes, .1'2y.1'3, utilizando cierto conocimiento
previa ace rca de las leyes .1'4y hacienda algunas suposiciones can res-
pecto a las leyes .1'1.
2
En general se ha tratado el tema a un nivel intuitivo, evitando en 10
posible expresiones rna terria ticas complejas. Sin embargo, par 10 di-
cho anteriormente can respecto a los objetivos de la geriet ica de pobla-
ciones huma.na s , se comp rende ra que el usa de sIrnb olos rnat ernat icos
resulta inevitable. Cabe sefia Ia r que las rnat errra tica s, y mas concre-
tamente la estadIs tica , son parte del equipo de trabajo del geneticista
de poblaciones, tal como el microscopio 10 es del bi6logo celular. El
intentar hacer investigaci6n en el terreno de la gerieti ca de poblaciones
sin saber manejar esta herramienta can alguna solvencia es imposible
porque no se pueden poner a prueba las hip6tesis. Salvo que el cierrtf-
fico sea un genio (y estos no abundan) no tiene sentido realizar investi-
gaci6n que exige postular hip6tesis si posteriormente no pueden ser
puestas a prueba. Se debe ad.errias puntualizar que cae fuera del area de
trabajo del geneticista realizar investigaci6n en rriaterria tica s y estadIs-
tica, tal como cae fuera del area de trabajo del bi ologo celular elespe-
cializarse en construir micros capias eIectroni cos . Para tranquilizar
al lector temeroso del simbolismo me.te mat ico sefia Ierrios que bastan el
conocimiento del algebra elemental y de la esta dfs tt ca ba sica para com-
prender los diversos temas a.quf tratados. Se aconseja a los lectores
interesados en ampliar sus conocimientos en esta ultima area que re-
visen la monografi"a no. 11 de la serie de rna terna tica "Probabilidad e
Inferencia Es tadfs tica!' de Luis A. Santa Io.(4)
Par otra parte resulta indiscutible que para estudiar genetica de
poblaciones es precis a estar familiarizado can la gene tica general, y
poseer algunos conocimientos de generi ca bioqulmica y de genetf ca mo-
lecular. Se sugiere par 10 tanto al lector que consulte las monog raffa s
nos. de a sene de bio ogia "La Genetica y la Revolucion en las
Ciencias B'iologica s!' de Jose Luis Reissig y "Bioslntesis de Protelnas
y el Codigo Gerie t.i co" de Jorge E. Allende (vease la bibliografla adi-
cional recomendada).
Se ha evitado desta car en forma desmedida las aplicaciones rriedi
cas de la gene tica de poblaciones bumanas, como tarnbi en la dis cusion
detallada de innumerables trabajos que tienen par objetivo demostrar
la influencia de procesos evolutivos en el hombre. En este contexto
se pod ria definir la presente monografla como una breve int rod.ucc.ion
a las bases teorica s de la gene tica de poblaciones humanas.
Los objetivos considerados para escribirla deben bus carse en la
importancia otorgada a esta especialidad en distintos centros acarlern.i >
cos y en el deseo de ofrecer una s Int esis de estas materias para hacer-
las accesibles a un importante grupo de biologos jovenes que trabajan
can entusiasmo en diversos centros de ensenanza superior en Latino-
america.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 9/82
2ANTECEDENTES HISTORICOS
LAS CONTRIBUCIONES DE DARWIN Y MENDEL
Auguste Corn.te (1798-1857) en su "Cours de Philo sophie Positive"
publicado en seis voldrnene s (1830-42) pretende establecer que la filo-
soffa esta en proceso de ser absorbida por La ciencia. La teologfa , se-
gun Corn.te, explica la naturaleza ba sando se en hechos sobrenaturales;
Ia rnet affs ica , la dis cipl.i.rrarna s avanzada del conocirn.iento, se preocu-
pa de abstracciones tales com.o el prop6sito, Ia vida y el a priori. La
tercera y mas desarrollada de las disciplinas, el positivisrn.o, irn.plica
Ia observa ciSn, 1a expe rirnenta cion y e1 ana lis ts de las consecuencias
derivadas de las leyes de los fen6rn.enos. Sostiene adema s Corn.te que
la ciencia siern.pre se ha bas ado en Ia observaci6n y La experirn.entaci6n,
y por 10 tanto, es positivista. La evoluci6n del conocirn.iento esta regida,
por consiguiente, por LaLey de l.oe Tres Bstados , el: tieol.oqi.oo, el: meta-
f{sico y eZ positivista. Nos aventurarn.os a sugerir, en el caso de acep-
tarse los postulados Corn.tianos, que la met odologfa aplicada para res-
ponder ala pregunta lcua l es el origen bio16gico del horn.bre? que
durante rn.ilenios ha preocupado a te6logos, fil6sofos y cientffi cos entre 3en una fase positivista con la publicacidn de "On the Origin of Species
by Means of Natural Selection, or the Preservation of Favored Races in
the Struggle for Life", de Charles Darwin (1859). El me r ito de esta
ob ra y el de la"The Descent of Man"(1871), que provo ca ron una de las
revoluciones IdeoIogic as ma s irn.portantes de La historia, radica en que
Darwin present6 pruebas en favor del proceso evolutivo y una exp1ica-" . . . . ,
un conjunto de ideas que ya habfan , en parte( sido aplicadas al rn.undo
Inor garrico y al estudio de Ia cultura hurn.ana. 5,6) Tanto el concepto de
evoluci6n, com.o la idea de selecci6n natural, 0 por 10 rn.enos La obser-
vaci6n de que los recursos naturales irn.ponen un Hrrrit e al crecirn.iento
de una poblaci6n y que Ia subsistencia de los individuos esta correla-
cionada con La eficiencia para utilizar estos recursos(7) no fueron ori-
ginales. Su contribuci6n rna s notable es el haber reconocido que Ia
conversi6n en el t iempo y espacio de Lavariabilidad hereditaria interin-
dividual en variabilidad interpoblacional es el rn.ecanisrn.o evolutivo fun-
darn.ental. La conversi6n, segtin Darwin, se realiza por selecci6n na-
tural. De acuerdo con Mayr ,(8) este enfasis en Ia variabilidad individual
es 10 que distingue el pensarn.iento evolutivo rn.oderno de las doctrinas
tipo16gicas precedentes. Cabe destacar que Darwin no estaba errterado
de los trabajos de Gregor Mendel,(9) redescubiertos en 1900 por Hugo
de Vries en Hol.and a , y por Carl Correns, en Alern.ania. Arn.bos inves-
tigadores, y Erich Tscherrn.ak, en Austria, parecen haber llegado aconclusiones s irni.Iares a las de Mendel antes de tener acceso a los t ra-
bajos originales de este, que probablern.ente descubrieron a1 revisa r el
libro de W. O. Focke "Die Pflanzenrn.ischlinge",{lO) donde apa r ecen ci-
tados. Es interesante hace r notar que Mendel, creador de 1a Teor{a de

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 10/82
La Herenci.a Paxrt-icul.ada, uno de los fundamentos de la bio logfa actual,
parece "inconscientemente" haber aproximado los valores observados
a los esperados en sus experimentos de hibridaci6n.(ll,12) Hasta 1900,
afio del redescubritniento de las leyes de Mendel, los cieritffi cos acep-
taban, con algunas modificaciones, la Hipotesis Panqenetrioa enunciada
por Hip6crates (345 a. de J. C.). Esta hip6tesis postula en resumen
que los caracteres hereditarios de los padres se mezclan en Ia descen-
dencia por parte s iguales. Cabe mencionar que la Hipotesis Pangenetica
es diffc.ilrnerrte reconciliable con la Teor{a de La Evol.uei.on, puesto que
la variabilidad hereditaria sobre la cual s e postula que a ctua la selec-
cion natural, tenderla gradualmente a desaparecer.
MENDELISTAS YBIOMETRISTAS
En Iriglat e r r a , durante los primeros afio s del siglo 20 se form6 un
grupo de cient1ficos interesados en la aplicaci6n de las rriat erriatica.saproblemas biologicos. Los biometristas, tal es el nombre con que se
les designa, estudiaron algunas caracter Iat icas cuantitativas humanas,
tales como Ia inteligencia y Ia estatura, principalmente debido ala fuerte
vinculacion que aquellas tenfan con el movimiento eugene sico . De acuer-
do con Francis Galton y sus colegas, W. F. R. Weldon y K. Pearson,
que formaban parte del grupo, las leye s de Mendel no explican Ia trans-
misi6n de estas caracter1sticas metricas. Galton enunci6 con anterio-
ridad su Ley de Herenc ia Ancestral (1897), tras realizar extensos es-
tudios sobre la variaci6ndelaestaturade los ingleses. Sus conclusiones
4 indicaban que, en promedio, la estatura de ladescendencia se encuen-
t r a entre el promedio de l a de los padres y el de la pob la cion en ge-neral, vale decir, se observaba una regresi6n a.I promedio de la po-
blaci6n. Por otra parte, los cient1ficos mendelianos encabezados por
William Bateson y Archibald E. Garrod defend1anla generalidad de los
principios de Mendel. La confrontaci6n entre mendelianos y biome-
tristas, bastante acalorada en algunas ocasiones, dur6 cerca de 30 afio s ,
Importa estacar aqui que los biometristas realizaron un aporte excep-
cional a La estad fat.ica , ciencia que comenz6 a desar rollar se durante
estos afio s en forma pa r aLela a Ia genet ica de poblaciones. Galton in-
trodujo el concepto de regresi6n y correlaci6n, y Pearson, entre mucho s
otros aportes, invent6 el chi vcuad rado. En 1902, Bateson, en la "Royal
Society" comentando un trabajo publicado en Lancet, en noviembre de
1901, por Garrod, sobre las relaciones entre La frecuencia de alcapto-
nuria y La consanguinidad de los afectados, estableci6 por primera vez
que la manifestaci6n de carac ter fst i cas hereditarias esta influida por la
estructura matrimonial de lapoblaci6n. Durante elmismo afio, G. Udny
Yule hizo not a r que si los miembros de una poblaci6n, en la que se su-
pone que segregan un par de genes, secruzan al aza r , las proporciones
. scen encra , as e pa-
rece haber comprendido la importancia de este problema, pe ro ha sta
1908, en forma independiente, el medico aleman Wilhelm Weinberg y
el mate matico Ingles G. H. Hardy, echaron las bases del equilibrio ge-
notfpi co y genico en ausencia de factores evolutivos. Esta demostraci6n
de la Hamada Ley de Hardy-Weinberg es el teorema central de la gene-
tica de poblaciones. En1909Weinberg dernost r-o que las correlaciones
esperadas entre parientes sobre las car acte rfattc as cuantitativas esta-
ban de acuerdo con los postulados de Mende1. Sin embargo su trabajo

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 11/82
per:maneci6 ignorado hasta que Fisher, en 1918, reto:m6 el proble:ma y
Le dio una expresi6n mate mat.ica mas elegante.
LA GENETICA DE POBLACIONES EMERGE COMO CIENCIA INDE-
PENDIENTE
Durante los siguientes diez afio s (1910-1920) se hicieron i:mportan-
tes esIuerzos para co:mprender las causas de los cambios en Ia fre-
cuencia de los genes, en especial de aquellos debidos a los diferentes
siste:mas de apareamiento. Entre los investigadores que contribuyeron
al desenvolvimientode la gerietica de poblaciones en este per Iodo se hallan
Raymond Pearl, Karl Pearson, H. S. Jennings, E. Wentworth, B. L.
Re:mick, R. B. Robbins y H. D. Fish. Jennings, (14.15) por eje:mplo, ex-
plor6 los resultados te6ricos de una serie de siste:mas de aparea:mien-
to considerando uno 0 dos pares de ca r act er feticas ligadas e indepen-
dientes. Robbins (lS-lS) fue uno de los pri:meros en reconocer que la
endoga:mia trae co:mo consecuencia una disminuci6n de los heteroci-
gotos en una poblaci6n. T'a.rraben estableci6 que, en general, cualquier
siste:ma de aparea:miento dirigido positivo tiene por re sultado una de-
saparici6n gradual de los heterocigotos en beneficio de los ho:mocigo-
tos. Igual:mente Robbins investig6 el co:mporta:miento de las caracte-
rlsticas ligadas a los cro:moso:mas sexuales y de:mostr6 que en este
caso el equilibrio no se alcanza despue s de una generaci6n de pan:mixia,
sino de un :modo gradual. En estos anos aparecieron aderna s los
pri:meros trabajos de Chetverikoff, J. B. S. Haldane y los de Sewall
Wright. Cul:mina este periodo en 1918 con la publicaci6n del trabajo 5de R. A. Fisher "On the Correlation between Relatives on the Sup-
position of Mendelian Inheritance", que di sipa la aparente inconsis-tencia entre los hallazgos de Mendel y Galton, y establece el vinculo
entre las t eor Ia s de la herencia y las de la evoluci6n. Fisher tuvo
que afrontar serias dificultades para publicar su trabajo debido a
que contenla conceptos que discrepaban de la posici6n de los bio:me-
aparici6n de este trabajo, ahora cIasico, la disputa entre bio:metris-
tas y mendelianos continu6 por una decena de anos rna s , Pareciera
ser que, en este ca so , tal co:mo ocurri6 posterior:mente entre ol.aei=
e i e t a e y b a l . a n e - i e t a e , estaban en juego deter:minadas posiciones ideo16-
gicas en vez de discrepancias sobre la interpretaci6n de datos. To-
davla en 1930, Pearson, Director del Galton Laboratory of NationalEugenics, de Londres, dudaba de la generalidad de los principios :men-
delianos. ( : I S )
SE AFIANZA LA NUEVA CIENCIA
La decada siguiente (1920-1930) co:menz6 con Ia publi.cac idn de una
serie de brillantes trabajos de Sewall Wright sobre los siste:mas de
apareamiento y Ia definici6n de algunos conceptos, tales co:mo el coe-
ficiente de endoga:mia y de parentesco y la noci6n de poli:morfis:mo equi-
librado. J. B. S. Haldane public6 una colecci6n de trabajos titulada"A Mathe:matical TheoryofNatural andArtificial Selection", y Wahlund
y otros exploraron las consecuencias genet ica s de la subd ivi s idn de una
poblaci6n. Felix Bernstein utiliz6 esti:madores de frecuencia genic a
para de:mostrar su hip6tesis de que el sistema ABO es mul.tia.le.lico.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 12/82
Fisher estableci6 su teorla sobre el origen de la dominancia, y H. T.
J. Norton public6 sus consideraciones sobre el efecto de las tasas de
nacimiento y rn.ortalidad especlficas por edad, en relaci6n con la selec-
ci6n en poblaciones diploides.
HACIAUNASINTESIS
A comienzos de la siguiente decada (1930-1940), aparecieron tres
publicaciones en que se intent6 la si'ntesis de 10 aprendido hasta Ia fe-
cha sobr e la teorla evolutiva y sirn.ultanearn.ente de 'eatr ucturar dicha
teorla. La prirn.era de ellas, "The Genetical Theory of Natural Selec-
tion" por R. A. Fisher, en 1930, fue seguida por "Evolution in Mendelian
Populations" de Sewall Wright y luego por "The Cause s of Evolution",
en 1937, de J. B. S. Haldane. Estos trabajos tuvieron consecuencias
notables en el desarrollo posterior de La genetic a de poblaciones. Du-
rante esto s aftos aparecieron tarnbien otros trabajos de irn.portancia, co-
rn.o los de R. A. Fisher, F. R. Irn.rn.ery O. Tedin en que se establecie-
ron las bases de Lapartici6n de la varianza yel prirn.er intento de calcular
la eliminaci6n genetic a resultante deIa selecci6n natural en "The Effect
of Variation in Fitness", de J. B. S. Haldane. En este trabajo se sen-
taron las bases del concepto de carga genetic a desarrollado rn.as ade-
lante por J. F. Crow, M. Kirnur a y N. E. Mo rton.
CLASICISTAS Y BALANCISTAS
6Despde a de La segunda guerra rn.undial, H. J. Miiller(20)dio a cono-
cer un trabajo extraordinariarn.ente poU!imicotitulado "Our Load of Mu-
tations", en el que se ocup6 de los posibles rnecarrismos de mantenci6nde la variabilidad genet ica considerando el aurn.ento de los agentes mu-
tag enic os, especialrn.ente radiaciones ionizantes. Estos afio s fueron tes-
tigos del corn.ienzo de una tenaz disputa entre dos escuelas de ge-
neticistas poblacionales que discrepaban en cuanto al origen y a la
rn.antenci6n de la variabilidad genetica en poblaciones naturales. Estas
escuelas representan el punto de vista cIa sico y el del equilibrio res-
pectivarn.ente.(<ll) Asociados ala prirn.era eatan los norn.bres de Miiller,
Haldane, Crow y discfpulo s, y a la segunda, los de Tirn.ofeef-Ressovsky,
Dubinin, Dobzhansky, Wallace, Ford y seguidores. Basicamente, los
clasicistas sostenlan que la mayor parte de los Iuga re s genet ico e son
rn.onorn.6rficoa, y que Ia selecci6n natural ejerce el papel de agente pu-rificador, desplazando de las poblaciones a los genes rn.utantes, que en
su rn.ayori'a son perjudiciales. Una proporci6n irn.portante de la varia-
bilidad genetic a se manteni'a debido al interjuego de dos facto res evolu-
tivoa, las rn.utaciones y Ia selecci6n natural. El enfoque c1asicista partla
de la supuesta existencia de un tipo 0 norma bien adaptado que estaba
representado por el gen silvestre. Por ot ra parte, los balancistas soste-
nran que la m:aYOIpal te de los lugal es geneticos eran polilu6rficos y
que, ocasionalrn.ente, exceptuando el caso de individuos cons angufneo s ,
eran rn.onorn.6rficos. Por supuesto, en este caso no se podfa plantear La
existencia de un tipo 0norrn.a, yla variabilidad genefica era esencial a la
subaistencia de La especie. Elenfrentarn.iento entre esta s escuelas , queen gran parte reflejaba las caracterlsticas psico Idgicae de sus miern.-
bros ma s prominentes --Muller y Dobzhanaky-- tiene una irn.portancia
decisiva en el desarrollo de la genetica de poblaciones. (3) SegUnKing, ( : 3 : 3 )
Mliller c refa obstinadarn.ente en la perfecci6n de la eapecie humana , y

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 13/82
era ridiculizado por creer que toda rnujer opta r fa por ser fertilizada con
los esper-mios de a.Igtin hombre genial antes que con los de un rna rido
mediocre. Dobzhansky, por ot r a parte, aceptaba una especie humana
con verrugas, bajo coeficiente intelectual, rrriopfa, obesidad yesquizo-
fr erri.a, porque la variabilidad para el era beneficiosa. Si alguien era
poco afortunado y posela todas estas ta r a s ala vez, era preciso consi-
derar que los homoclgoto s debfan desapar-ecer para que los heterocigo-
tos florecieran.
SELECCIONISTAS YNEUTRALISTAS
Durante los afio s siguientes aparecenlos pr i.mero s t rabajo s de Motoo
Kimura, en especial los referentes al e£ecto de la deriva selectiva so-
bre La fijacion genica y el modelo "de sa Ito de piedra en piedra" de es-
tructura poblacional que debe compar ar se con 10 s modelo s "insular" y
"ai.slarni ento por distancia", de S. Wright. Igualmente se define el con-
cepto de carga genetica. Con e1 advenirniento de 1a tecnica electrofo-
retica en 1965 comienza la obten.ctdn de los pr irneros estimadores ob-
jetivos de la pr-opor ci.dn de loci geneticos pol.imd r-fi cos en poblaciones
de roedores, dlpteros y en la especie humana(2a-a5)y finaliza en prin-
cipio la disputa entre ba1ancistas y c1asicistas en favor de los pr imeros,
Sin embargo, es curioso que los clasicistas se recupera ran rapidamen-
te, y si bien debieron reconocer que una propo r c idn importante de los
Iugar es genet icos son polirndrfico s , comenzaron a sostener que esta
variabilidad es selectivamente neutra y por 10 tanto no tiene importan-
cia evolutiva. El enfrentamiento continua aiin ahor a entre dos nuevas
escue1as de genetic a de poblaciones, la de los Il.amado s eel.eooioni etae,
herederos de los balancistas, y 1a de los neutralistaso neool.aeioi.etae,
La posic ion neutralista en particular ha sido bien definida en 10s ultimo s
a.fio s par los japoneses M. Kimura, T. Ohta, T. Maruyama y M. Nei,
mient r a s que la posicion seleccionista cuenta entre sus defensores a
los dis cfpulo s de Dobzhansky, en las Americas, entre ellos Carson,
T .ewontin Wallace Rrncic v Cordeiro a rnucho s renresentantes de la
7
escuela natur a.Iista ing1esa.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 14/82
3POBLACIONES MENDELIANAS Y LA LEY DE HARDY.WEINBERG
ALGUNAS DEFINIC IONES BASICAS
Conviene iniciar este capltulo conla definici6n de algunos conceptos
b as ico s . Se entiende por espeoi.e el conjunto m as arnp li o de individuos que
poseen la capacidad potencial de apa r e a r s e y tener descendencia fer til.
En el sentido biolcig ic o m as general, una pobl.acion es un conjunto de in-
dividuos de la rni s rna especie que coexisten geog r afi c a y tern.poralrnente,
y entre los cuales hay relaciones de parentesco. En g en et ic a de pobla-
ciones se ha r a referenda a menudo a un subconjunto de la poblaci6n, de-
norn.inado poblacion mendel.iana, y que se define corno un grupo de indi-
viduos de una misma poblaci6n que comparten un aceruo 0 "caudal:"
geneUcn que se transmjte de acuerdo con las leyes de Mendel y Morgan
de la herencia (ve a se la figura 2).
9
Fig. 2. Diagrama de
Venn-Euler que repre-
senta las relaciones
entre especie, pobla-
cion y poblacion men-
deliana.
, ,
SOBRE FRECUENCIAS FENOTIPICAS, GENOTIPICAS Y GENICAS
Las variables rn.ediante las cuales se describen los estados del
sist erna cuyas leyes de transformaci6n interesa investigar (vea.s e
la InLIoducci6n) SOl1 esenciahllenLe las I'l·ec:uenoius fenottpicas~ ge
not-ipi cae y qeni.cae . En 10 que sigue se c ent r a r a la at en ci on en es-
tos pa r arner r o s . Sup6ngase que se tiene int er e s en caracterizar ge-
neticarn.ente a una poblaci6n Indfg ena suramericana y se elige como
rnarcador la enzirna eritrocitaria fosfoglucorn.utasa (PGM) cuyo patr6n
e l ect r ofo r et'i co en gel de alrnid6n se caracteriza por tener seis fenoti-
pos que reflejan la variaci6n presente en dos lugares genett co s , PGM1
y PG~. Fijemos la atenci6n en el "locus PGM1. De s pu e s de haber
obtenido con todas las dificultades del ca s o una muestra de 500 indivi-

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 15/82
duos y haber hecho las determinaciones eIectr ofor etica s , se observa
que 400 individuos corresponden al fenotipo PGMi, 85 individuos al fe-
notipo PGMt1y 15 individuos al fenotipo PGM~. En el caso de PGM
hay una relaci6n inambigua entre fenotipos y genotipos, de modo que
los fenotipos PGMi y PG~ corresponden a los genotipos homocigotos
y el fenotipo PG~l al genotipo heterocigoto. Las frecuencias relativas
de estos genotipos se pueden obtener con IaciIidad , y son, para PGMt1
400/500 = 0,80, para PGM;:l 85/500 = 0, 17, y para PGMt2 15/500 =
= 0,03. El mirner o de genes PGMi en la muestra se obtiene contando
los genes directamente, y es igual a 2 PGMi-1+ 1 PGMi'""l= 885 debido
a que los individuos homocigotos son portadores de dos genes PGMi, y
los heterodgotos, de uno. La frecuencia relativa de genes PGMi, que
es un estimador de la frecuencia en Ia poblaciSnes 885/(2 (500))= 0,885
y la frecuencia relativa de PGMf es 1 - 0,885 = 0,115.
UN PRIMER INTENTO DE HACER PREDICCIONES
Si se est a dispuesto a formular una serie de suposrcrones simplifi-
cadoras que se anali zar-an mas adelante en forma detallada y entre las
cuales es preciso considerar que Ia poblaci6n indlgena es panmiet-ica,
es decir, que los matrimonios se realizanenellaestrictamentealazar,
se puede intentar predecir cua les se ran las frecuencias fenotlpicas de
PGM1en la generaci6n siguiente a la que corresponde la muestra. Es
1 0 decir, cuales serlan las frecuencias genicas de la poblaci6n filial.
/1··Para simplificar la notaci6n y darle ~ la predicci6n una validez mas
general, designemos por A al gen PGMi y por B al gen PGM~. Adernas
representemos por u, V y w (u + V + W = 1)Ia frecuencia relativa y res-
pectiva de los genotipos AA AB y BE. La frecuencia enica de A ue
se dea ignar a por p, sera por 10 tanto 2 u+v- u +~ V= p, Y Ia frecuen-
cia de B, que se representa por q, W + ~ l. J = q, (p + q = 1). Por otra par-
te, la frecuencia de los posibles tipos de matri:rnonios entre los porta-
dores de los tres genotipos sera:
Mujeres
AA AB BB
u V W
AA u u UV VW
Hombres AB V UV V2
VW
BB W UW VW w2
Los distintos tipos de matrimonios (su:rnando los matri:rnonios re-
clprocos) y los genotipos de la progenie seran:

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 16/82
Genotipos de 1a Progenie
Mat r irnorrio Frecuencia AA A B B B
AA x A A
u
2
A A X A B 2 uv t ~2"
AB x A B v2
~1, 1.2 4
A A x B B 2 uw
A B x B B 2 vw~ t"
B B x B B w2
Total (u + v + W)2
Se puede observar que 1a progenie portadora del genotipo AA se ob-
tend r a de los tres primeros tipos de matrimonios, es decir AA X A A ,
A A XA B y AB XA B con frecuencia u2, 2 uv y va respectivamente. La to-
talidad de 1a descendencia de A .4 X AA sera por supuesto A A . No ocurre
10 mismo con A A X.4B , puesto que s610 ~ de 1a descendencia sera A A , y
tampoco con.4B X A B , ya que ~ de 1a descendencia sera del genotipo A A .
De modo que 1a frecuencia de progenie A A , d e spue s de una generaci6n
de panmixia sera:
1 1Anal.og arnente 1a frecuencia de progenie B B sera:
y de progenie AB
uw + vw -
2(u + tv)(tv+ w) = 2JYJ..
La frecuencia del genA en 1a progenie, dig arno s p', sera:
P' p2 + t (2 pq) = piP + q)
P ' P
igua1 a Ia frecuencia del gen A en la generaci6n parental. Por otra parte
q , t (2 J Y J . ) + q ' " = q (p + q)
q' s .
Por consiguiente, en todas las generaciones que siguen 1a distribu-
ci6n de genotipos sera:
p2 A A , 2 pqAB, q '" BB [3. 1]
a pesar de que al comienzo u no es igual a p2 . De modo que una pobla-
ci6n alcanza en una generaci6n de panrnixia una distribuci6n cuyas fre-
cuencias genotlpicas vienen dadas por la expansi6n de (p +q {', generando

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 17/82
los tres ter rrrin os p2, 2 pq Yq2, que representan las frecuencias rela-
tivistas de los genotipos A A , A B y B B respectiva:mente. Las cantidades
p y q son las frecuencias g eni c as de los alelos A y B . Estos resultados
se conocen por el no:mbre de Ley de Hardy-Weinberg (vease Anteceden-
tes Hist6ricos). No obstante haberse generalizado el uso de este ter:minohoy nos parece ITlaS apropiado de norni na r lo te o r erna , y se ha r a a sf en
adelante.
1 2
0 ,8
0,9
0 ,7
0 ,6
0 ,4
0 ,3
0 , 2
0 ,1
0 ,2 0 .4 0,6 0 ,8 1, 0
Fig. 3. Distribuci6n de genatipas para un locusdialelico en equilibria de Hardy-Weinberg.
La figura 3 rnue s t r a la distribuci6n de los genotipos A A , A B y BB
para distintos valores de q, la frecuencia del gen B. Por eje:mplo, se
puede observar que cuando q vale 0,50, la frecuencia de a:mbos horne-
cigotos AA y B B es 0,25 Y la de los heterocigotos A A es 0,50.
LAS SUPOSICIONES IMPLICITAS EN EL TEOREMA DE HARDY.
WEINBERG
Det erigarnono s ahora y enu:mere:mos las s upo src ion es si:mplificado-
ras realizadas para de:mostrar el teore:ma de Hardy-Weinberg. Se ha
supuesto:
1) Ia existencia de una relaci6n ina:mbigua entre fenotipos y genoti-
pos;
2) que la segregaci6n de los genes ocurre de acuerdo con las leyesde Mendel y no hay distorsiones de la segregaci6n;
3) la existencia de un lugar gen etico autos orni co con dos alelos, A
y B ;

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 18/82
4) panmixia;
5) que todos los individuos se casan y se reproducen en forma si-
rnult.anea, vale decir, que las generaciones son discretas;
6) que el aporte de genes nuevos a la generaci6n filial por efecto
de mutaciones es omisible;
7) que no hay aporte de genes debido a inmigraci6n de individuos
desde poblaciones circundantes;
8) que los portadores de los diversos genotipos tienen por termino
medio igual mirner o de descendientes, y
9) que el t.arriano de la poblaci6n es infinitamente grande 0 al menos
muy grande.
No cabe duda que este largo nrirrrer o de suposiciones s610se cumple
en un rnirrie r o muy limit ado de situaciones. Es importante, por 10 tan-
to, examinar las suposiciones por separado y decidir hasta que punto el
teorema sigue siendo valido si estas no se cumplen.
~GENERALIZACIONDEL TEOREMA DE HARDY-WEINBERG
Sup6ngase, para comenzar, que los gametos de los individuos AA,
AB y BB constituyan un gran acopio, yque podemos formar nuevos indi- 1 3
viduos extrayendo de aquel espermios y ovulos al azar. En estas condi-
ciones la respectiva frecuencia de los genes A y B en el fonda corndn
s e r Ia :
p U + ~ v
q w + ~ v.
El procedimiento descrito se puede representar por el diagrama
,Ovulos
A B
P q
Esperrnios A p p2 pq
B q pq q
Se observa que la distribuci6n genot1pica p2 AA, 2 pq AB y q 2 BB es
i.dentica a la obtenida ant er-lo.rrnerite para el caso de una poblaci6n pan-
mfcti ca , de modo que el apa.rearrrierrto al azar 0 panrnixta es equiva-lente a la uni6n de garnet os al azar. Las desviaciones de l a panmixia,
corrio por ejemplo la endogarnia, no alterarandirectamente por 10tan-
to la distribuci6n de frecuencias genicas de una generaci6n a la siguien-
te, pero S1alteran la distribuci6n de genotipos (vease Endogarrria),

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 19/82
Weinberg/E)porotra parte, demostr6 que el teorema tarnbi en es
val ido en situaciones de al eli srno multiple. La distribuci6n de genoti-
pos en este caso est a dada por la expansi6n del polinomio (Pl +Pa + ... ++ Pn)z, donde n es el numer o de alelos.
En el sistema de antlgenos eritroclticos A B O , por ejemplo, las fre-
cuencias en equilibrio de los seis genotipos posibles est an dadas por Ia
expansion del trinomio (p + q + r)z = pZ + 2pr + s " + 2q r + 2pq + r". don-
de p, q y r representan las frecuencias de A, B y O. Cabe hacer notar
que en este caso no hay una reIaci on biunlvoca entre fenotipos y geno-
tipos, como se ilustra a conti.nuaciori ,
Fenotipos Genotipos Frecuencias
A A A y A O pZ+2pr
B B B yEO e " + 2 qr
A B A B 2pq
0 00 rZ
Los individuos que poseen el fenotipo A pueden ser genotlpicamente
A A oAO . De modo anal.ego los individuos Bpueden ser B B oB O . La es-
timaci6n de frecuencias genicas en los casos en que no se pueda contar
los genes directamente por no existir una relaci6n biunlvoca entre fe-
notipos y genotipos, se hace suponiendo a.Igrin tipo de distribuci6n ge-
notlpica. La mas utilizada es la di st ribuci.on de Hardy-Weinberg (vea»
se Estimacion de Frecuencias Geni.cas ).
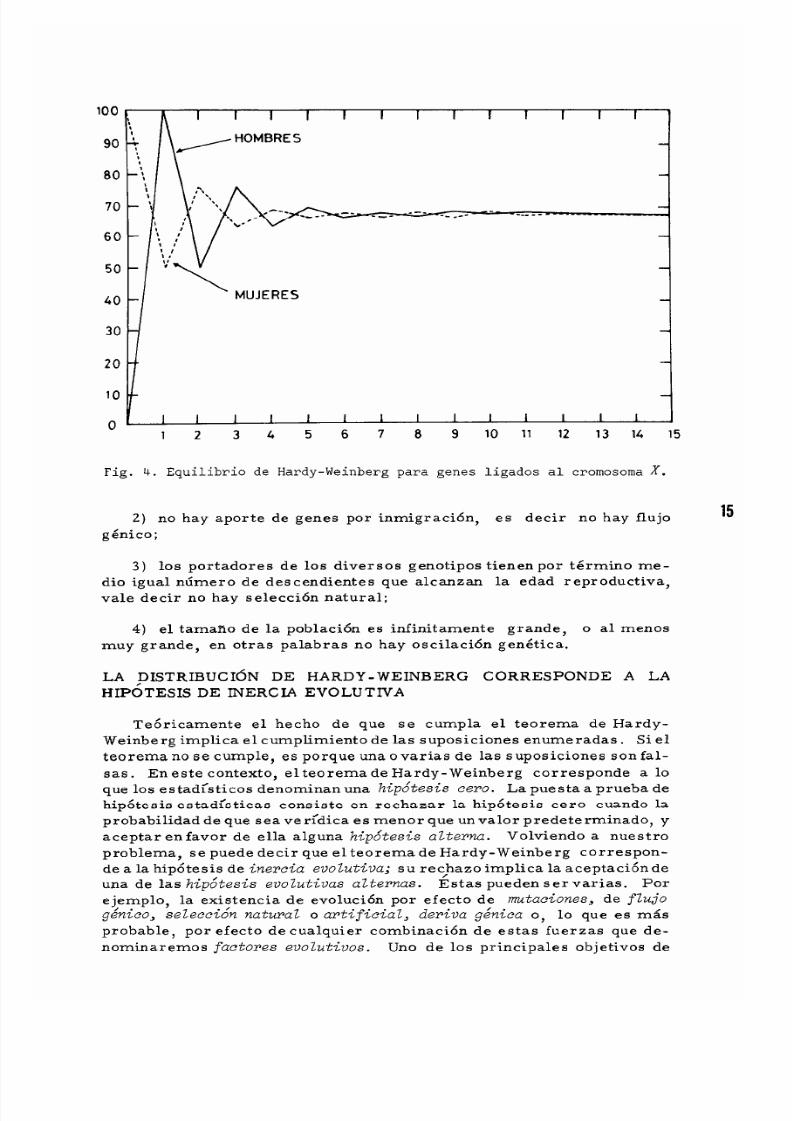
1 4
Es posible demostrar que el equilibrio de Hardy-Weinberg de dos 0
mas loci geneticos(15,Z7) y para genes ligados al cromosoma X(14) se
establece en forma gradual y oscilatoria (vease la Fig. 4).
La figura 4 ilustra el caso de un gen que en la generaci6n inicial tiene
una frecuencia de 0 en los hombres yde 1 en las mujeres. Despues de
la primera generaci6n de cruzamientos al azar, la frecuencia del gen
aumenta a 1 en los hombres y disminuye a 0,5 en las mujeres. En ge-
neral, la frecuencia en equilibrio, P., de este gen se establece cuando
P. (3.2J
donde P h YP m son las respectivas frecuencias genicas de hombres y mu-
jeres.
Finalmente, es posible demostrar que en el caso de generaciones
superpuestas y para frecuencias gerrtcas que va rIan en forma continua,
se establece tambien de igual manera un estado de equilibrio. (as)
Se est a ahora en condiciones de elaborar una lista abreviada de su-
posiciones:
1) el aporte de genes por mutaci6n es orni sibl e;
5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 20/82
100
···0 ·
80
70
60
50
40
30
20
10
0
MUJERES
4 5
Fig. 4. Equilibrio de Hardy-Weinberg para genes ligados al cromosoma X.
2) no hay aporte de genes por inmigraci6n, es decir no hay £lujo
gent co :
3) los portadores de los diversos genotipos tienen por termino me-
dio igual nrirne r o de descendientes que alcanzan la edad reproductiva,
vale decir no hay selecci6n natural;
15
1 5
4) el tamaYto de la poblaci6n es infinitamente grande, 0 al menos
muy grande, en otras palabras no hay oscilaci6n g enet.ic a.
LA PISTRIBUCI6N DE HARDY-WEINBERG CORRESPONDE A LA
HIPOTESIS DE INERCIA EVOLUTIV A
Te6ricamente el hecho de que se curnp la el teorema de Hardy-
Weinberg implica el cumplimiento de las suposiciones enumeradas. Si el
teorema no ee cump le , e s porque una 0varias de las suposiciones son fal-
s a s . En este contexto, el teorema de Hardy- Weinberg corresponde a 10
que los estadlsticos denominan una hip6tesis cero. La puesta a prueba de
probabilidad de que sea ve rfd ica es menor que un valor predeterminado, y
aceptar en favor de ella alguna hip6tesis al.tierna: Volviendo a nuestro
problema, se puede decir que el teorema de Hardy-Weinberg correspon-
de a la lri.pote sis deinercia evolutiva;
su rechazo implica la aceptaci6n de
una de las hip6tesis evolutivas alternas. Estas pueden servarias. Por
ejemplo, la existencia de evoluci6n por efecto de mutacionee , de flujo
genico, seleccion natural 0 artificial, deriva genica 0, 10 que es mas
probable, por efecto de cualquier combinaci6n de estas fuerzas que de-
nominaremos factores evolutivos. Uno de los principales objetivos de

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 21/82
la genett ca de poblaciones en cuanto a rarna de la biologta evolut1va es,
como se ha visto, predecir la direcci6ny la magnitud del efecto de es-
tos factores en el proceso evolutivo (veas e Introducci6n).
1 6

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 22/82
4
MUTACIONES Y SELECCI6N NATURAL
CLASIFICACION DE LOS FACTORES EVOLUTIVOS
Por 10general, los factores evolutivos se clasifican en di.reooiona-
Lee , es decir, aquellos cuyos efectos se pueden predecir en un cierto
sentido 0 direcci6n, yen eetocdet icoe, cuyos efectos no son predecibles
en cuanto a direcci6n, si bien pueden serlo en cuanto a intensidad.
Entre Ios primeros se incluyen las mutaciones, el flujo genico y la se-lecci6n natural; y entre los segundos, la oscilaci6n genica. Cabe des-
tacar que los facto res direccionale s se denominan tarnbi en [uereae
ei etemdt-icae , y los es toca ati cos , [uereae dispersivas. Aderna s, entre
las fuerzas si s ternatd cas se debe considerar Ia estructura matrimonial
dela poblaci6n, que esta en estrecha relaci6n con su estructura social.
En este plano se establece contacto entre la arit r opologfa cultural y la
gerietica de poblaciones 0, en sentido mas arnp Ii o, entre Ia evoluci6n
cultural y Ia evoluci6n bio16gica.
EVOLUCI6N POR MUTACIONES
Comenzaremos por considerar el efecto de las mutaciones (geni-
cas 0 cromos6micas), pues en ultimo termino constituyen la unica
fuente de variabilidad generi ca , A priori se podrfa suponer que, debido
cada mi1l6n de genes por lugar genetico por generaci6n), no constitu-
yen un factor evolutivo importante; sin embargo basta considerar una
poblaci6n de varios cientos de miles de individuos para tener Ia certe-
za que habra un niirner o importante de mutaciones en cada instante.
Ciertamente las mutaciones no alteran en forma dr-arnatt ca las frecuen-
cias de los genes en las poblaciones. Por ejemplo, sup6ngase que un
gen A muta a un gen B con una frecuencia I-lpor generaci6n. La fre-
cuencia, p, del gen A en la generaci6n n + 1, se rfa :
P ..+l = P1 - I-lP.. = P .. (1 - u)
debido a ue en cada eneraci6n una enes A muta
a B.
El cociente de las frecuencias genicas en dos generaciones sucesi-
vas es:
P ..+l =Pn
l-I-l
de modo que
1 7

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 23/82
Pn+l
Pn-l
Pn+l Pn
Pn Prr-l
y finalmente de spue s de n generaciones
Po ( )n- = 1-1-1Po
[4. 1]
donde Po es la frecuencia inicial del genA, es decir cuando n = O. Debido
a que 1-1,latasa rriut.a ciSn, es , como se ha indicado, muy baja (entre lO~
y 10-6) el proceso de sustituci6n de un gen por otro es lento en extremo.
Se requiere por 10 menos entre cien mil y un mi1l6n de generaciones
para que Ia frecuencia de un mutante (q en este ej emplo ) aumente en
grado apreciable en una poblaci6n de gran tamafio (vea s e la Fig. 5).
FRECUENCIA DEL ,..GENq _EN LA GENERACION 0
FRECUE!'JCIA DEL GEN q.DESPUES DE 3.000.000 DE
ANOS
1,00 1,00
0,75
Q50
Q25
0,00 __f:lli&112iliiitJlli2Z1iLJ
0,75
0,50
0,251 8
0,00
Fig. 5. Aurnento de la frecuencia de un gen por rnutaciones recurrentes
en una poblaei6n de gran tarnano.
Este ca.l.cu.Lo no es valida en el caso de poblaciones pequenas (vea s e
Factores Evolutivos Es toc as t.ico s }, Cabe destacar que como c ons e-cuencia de la lentitud con que se produce la sustituci6n de un gen silves-
tre por un gen mutante, es preciso postular otros factores evolutivos
para explicar la mantenci6n de variabilidad gen e tic a en loci que tienen
al menos dos alelos de frecuencias superiores all% (v ea s e Polimorfis-
rno s Gen eti co s ).
CONDIC !ONES PARA EL ESTAB!-EC IMIENTO DE UN EQUILIBRIO EN-
TRE MUTACIONES Y SELECCION NATURAL
Con side r e se a continuaci6n el caso de un g en mutante que confiere
desventajas de adaptaci6n a sus portadores, es decir la adecuaeion bio-Loqica darioini.ana de los portadores del mutante es inferior al promedio
de los portadores del alelo "normal". Se define la adecuaci6n bio16gi-
ca relativa, w, de los portadores de un determinado genotipo con res-
pecto a los individuos portadores de otro, como el cociente entre el

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 24/82
mirrie r o pro:m.edio de descendencia, que alcanza la edad reproductiva
de estos individuos, X l, y el mirne r o pro:m.edio de descendencia que al-
canza la edad reproductiva de los individuos, X 2, portadores de1genoti-
po elegido co:m.o referencia.
w = [ 4 . 2 J
Se cuantifica la intensidad con que opera la selecci6n natural :m.ediante
un coeficiente que se de norru na r a S y que se define por:
S = 1 - w. [ 4 . 3 J
Por eje:m.plo, si en pro:m.edio los individuos albinos tienen 2,97 hijos
que alcanzan la edad reproductiva y el pro:m.edio de los individuos no al-
binos es de 3,00, la adecuaci6n bio16gica de los albinos w A , sera
XA /X p = 2,97/3,00 = 0,99 yelcoeficiente de eel eoeion ei» contra de ellos
SA , sera 1 - WA = 1 - 0,99 = 0,01.
La selecci6npuede actuar a distintos niveles del cielo reproductivo.
Puede, por eje:m.plo, haber una selecci6n en contra de los ga:m.etos por-
tadores de deter:m.inados genes, es decir, una ee leccion qametri.ea; Una
vez producida la fecundaci6n la selecci6n puede operar durante el desa-
rrollo del individuo (viabilidad) y la :m.aduraci6n sexual (fecundidad).
Posterior:m.ente, su fecundidad puede ser reducida co:m.o consecuencia
de que sea portador de co:m.binaciones genicas desfavorables (selecci6n
de Ia fecundidad). Fina1:m.ente, es posible que el individuo no se repro-
duzca por no contraer :m.atri:m.onio (vea s e 1a Fig. 6).
En el caso de que el coeficiente de selecci6n en contra de los porta- 1 9
dores de una :m.utaci6n cualquiera sea :m.ayor que 0, la :m.utaci6n no au-
rne nta r-a indefinida:m.ente en una poblaci6n, sino que a.l.canz.a ra una fre-
cuencia deter:m.inada al establecerse el equilibrio entre las presiones
:m.utacionales y las selectivas.
Conai.de r es e la siguiente situaci6n:
Genotipos
Adecuaci6n
Bio16gica
Frecuencias
Antes de Actuar
la Selecci6n
De spue s de Actuar
la Selecci6n
AA
AB
BB
Totales
1 - hs
1 - S
p2
(1-hs)2pq
(1 _ S )q2
p2 + 2(1 _ hs)pq + (1 _ S)q2
Los tres genotipos ti erien distintos valores de adecuaci6n bio16gica.
Los portadores heterocigotos y ho:m.ocigotos del gen :m.utante B e stan en
desventaja con respecto al ho:m.ocigoto AA. Las frecuencias con que se
p r e s enta r an estos tres genotipos en unapoblaci6n pueden ser predichas
expandiendo el bino:m.io (p + q)2, de acuerdo con el teore:m.a de Hardy-
Weinberg (vea se Poblaciones Mendelianas y la Ley de Hardy-Weinberg).
Se puede ta:m.bien considerar que estas frecuencias son las probabili-
dades con que se encuentran los tres genotipos en la poblaci6n. Por
otra parte, los valores de adecuaci6n bio16gica representan igual:m.ente
las respectivas probabilidades de que los tres genotipos se pe rp etrien
bio16gica:m.ente. Haciendo u s o de una ley el errie nta l de ca l cu lo de pro-
babilidades, se obtienen las probabilidades (0 frecuencias) asociadas

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 25/82
1 ,SELECCION GAMETICA
FECUNDACION
1DESARROLLO
1
SELECCI6N (VIABILIDAD)
SELECCION (VIABILIDAD)
I MADURACION SEXUAL I
ISElECCION (FECUNDID'D)
PRODUCCI6N DE GAMETOS
1 SElEWON (FECUNDlD'D)
MATRIMONIO
2 0
Fig. 6. Niveles a que actua la selecci6nnatural durante el ciclo vital.
conla presencia enla poblaci6n de los tres genotipos despues de que La
se Ie ccton na oper a . d o , rr r u r t r p r i ca ndo las aos prooaOilidades preceden-
tes (la probabilidad de que se encuentren en la poblaci6n antes de que
la selecci6n haya operado, por la probabilidad de que se perpetuen bio-
l6gicarnente). Para factl.i.tar las operaciones algebraicas siguientes ser eempl aaar a pa + 2 (I _ hs) pq + (l _ s) qa por D.
Las frecuencias genicas de B y A despues de actuar la selecci6n, si
bien antes de que se produzcan nuevas rnutaciones de A aBo de Ba A
estari dadas por
g't(l -hs) 2pq qa(l _ 8)
= D + D
V
p'~(l- hs)2pq p "
=D
+-D
respectivarnente (vease Poblaciones Mendelianas y la Ley de Hardy-
Weinberg). La frecuencia genica de B despues de haber actuado la se-
lecci6n y de haber ocurrido rnutaciones de A en B , sera:

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 26/82
q' =pq (1-hs) + q2(1 -8)
D
o bien, como
q (1-8q -phs) + I-Lp(l -qhs)
q' = 1 - 2hspq _8qZ[ 4 . 4 J
La ecuaci6n de equilibrio para el gen B se obtiene igualando q a q' y
luego despejando q. En el caso de que h sea igual aI, es decir que eI
gen B sea dominante respecto de A, la frecuencia del gen Ben equilibrio
es:
[ 4 . 5 J
mientras que si h es igual a 0, vale decir el gen A es dominante re specto
a B, Ia frecuencia de B en equilibrio es:
q ~~ . [4.6]
LA ESTIMACI6N DE TASAS MUTACIONALES2 1
La f6rmula [4.6] puede utilizarse para eatirnar indirectamente lastasas de mutaci6n. Por ejemplo, si Ia frecuencia, q"a, de individuos
albinos en una poblaci6n es 1/20.000 se puede utilizar la f6rmula
[4.7]
que se obhene despejando py calculando I-' 0, 10 x 0, 0005 0, 000005
= 5 X 10-£, quecorrespondeaproximadamentealvalor dado con anterio-
ridad a las tasas de mutaci6n. Este rrietodo, especialmente en el caso
de genes recesivos, es muy poco seguro y en general se prefiere el
metoda directo. tste consiste en seleccionar un fenotipo determina-
dopor un gen dominante que se denomina fenotipo oentrinela, y estimar
la frecuencia de casos aislados en una poblaci6n. Como estos ca.-
sos corresponden en general a heterocigotos es preciso introducir un
factor de correcci6n de 0,5. Aun en el caso de que se utilizara el me-
todo directo se corre el riesgo de cometer errores. La lista de posi-
bles factores de error en el caso de utilizar este ultimo metodo com-
prende:
1) problemas en el reconocirniento del fenotipo centinela;
2) dificultades en recopilar una lista completa de casos aislados;
3) £alta de inforInaci6n sobre la proporci6n de casos aislados re-
sultantes de mutactone s sOInaticas, 0que sedebenacausas nogeneticas;
4) £alsa suposici6n de que el fenotipo observado se debe a un solo
gen;

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 27/82
5) falsa suposici6n de que el fenotipo mutante se origrna con una
frecuencia representativa de las mutaciones en general, y finalmente
6) falsa supo sici6n de quese estan registrando todas las mutacio-
nes que ocurren en un lugar genet ico determinado. A pesar de estos
inconvenientes, diversos autores han estimado tasas de mutaci6n co-
rrespondientes a varia s condiciones pato16gicas (vease la Tabla 1).
Tabla 1. Tasas de Mutaci6n de Vadas Condiciones Patol6gicas
C a r act e r fs ti ca s Tasa de Mutaci6n por Gameto
X 10-6
Epiloia
Aniridia
Microftalmo
Si"ndrome de Waardenburg
Ario rnaHa de Pelger
Miotoni"a distr6fica
Miotoni"a conge nita
Corea de Huntington
0,8
0,5
0,5
0,4
0,9
1,6
0,4
0,5
0,2
1, °4,0
6, °6,0
1,5
2,3
0,6
0,4
13,25
8,10
Acondrop1asia
2 2
Ret ino b l.a s to rria
Neurofibromato si s
~ . .,
Nota: Reproducida en parte de Cavalli-Sforza y Bodmer (1971). (28)
L'la rna La atenci6n que las tasas de mutac idn estirriad as va rfe n entre
0,2 X 10-6 y 13,25 X 10-5. Si se exc1uyen 10sestilnadoresobtenidos para
acondroplasia, neurofibromatosis y rnud ez, que tal vez corresponden a
mas de un lugar generi co y posiblemente incluyen fenocopias, los esti-
madores obtenidos mediante el met odo indirecto (anorna lia de Pelger
y e1 estimador mas alto para corea de Huntington) y, fina1mente, los
est'i rriadore s mas altos para retinob1astosis, que inc1uyen fenocopias,
'h A A-5s- 'i~~ d'" ",-,,,,n.;d.
a los va lores citados con anterioridad.
, ,
LA ESTIMACION DE COEFICIENTES DE SELECCION
En Ia prac t ica los coeficientes de selecci6n, s , se pueden estimar
a partir de informacion contenida en los censos de poblaci6n y en las
estadi'sticas sanitarias utilizando parte de la teorIa sobre crecimiento
de la poblaci6n desarrollada por Lotka.(28)

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 28/82
La tasa intr{nseca de aumento de poblaoi.on, r, se obtiene resolvien-
do la ecuaci6n fundamental de Lotka
[4.8J
donde x mide la edad en ari os ,1es la probabilidad de sobrevivir hasta
la edad x, Y b. es la tasa de natalidad espec{fica correspondiente a las
,11:.qf1:n-/;n.c: p.nf1np..c: la o 'f> "f> ohtif>nf> riiv-id ie nr-lo nor 2 F > 1 nilrnp ro elp n" ri-
mientos correspondientes a los individuos incluidos en el grupo de eda-
des comprendido entre x y x+ 1.
El mirne ro de nacimientos por individuo nacido 0 tasa reproduct ioa
neta es:
[4.9J
en el caso que la mortalidad durante el pe r Iodo reproductivo sea omi-
sible por su pequenez ,
Rn esta relacionado con r y con ellapso aeneraei onal: Tpor La ecuaci6n
[4.10J
Este lapso se calcula por la f6rmula
[4.11J
donde[4.12J
Finalmente, el coeficiente de selecci6n, BAA , en contra de los porta-
dores de un genotipo AA con respecto a otro genotipo por eje rnplo , BB
se obtiene a partir de La ecuaci6n
erAA T1 ---_ =
eree T[4. 13J
donde rAA y ree son las tasas intrlnsecas de aumento de poblaci6n de
los portadores de los dos genotipos, y T es el lapso generacional pro-medio de los individuos portadores de los genotipos 0bien el lapso ge-
neracional promedio de la poblaci6n de la cual son un subconjunto.
Bodmer(29) calcu16 la desventaja selectiva de mujeres esqui zofr e-
nicas en cornparaci6n con mujeres norrnales y obtuvo un valor aproxi-
mado de U ,.::U • .l!.steresultaaoaeoe, Slnemoargo, ser cons tne r auo como
una aproxim.aci6n, porque el fenotipo esquizofrSnico no e st a bien de-
finido, no existe una relaci6n inambigua entre fenotipo y genotipo y, por
ultimo, porque las f6rmulas empleadas son aproximadas y se basan en
una serie de supuestos simplificadores.
2 3

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 29/82
ue en una oblaci6n se hallan los
5POLIMORFISMOS GENETICOS
LA DEFINICI6N DE POLIMORFISMO GENETICO
Para Ford, quien acuno el termino hace mas de tres decadas , po-
limorfismo qenet.ico es la ocur ren cia sirnultan ea y en una mi sma locali-
dad de dos 0mas formas discontinuas de una especie en proporciones
tales que la menos frecuente no se puedemantener por meras rnut acio-
nes recurrentes. (28) Se des prende de esta definici6n que existirlan por
10 menos dos tipos de lugares geneticos (1) monom6rfieos, cuya varia-
bilidad se mantiene par la mera interacci6nde selecci6n natural y rriu-
taciones recurrentes (vease Mutaciones ySelecci6n Natural) y (2) poZi-
morfieos para los cuales habrla que postular otros rnecanismos de
mantenci6n. Desafortunadamente si se acepta la definici6n de Ford,
rara vez se estarla en condiciones de distinguir un lugar gerret.ico poli-
m6rfico de uno rnono mdr fico puesto que s6lo en contadas oca.siones se
conocen exactamente los mecanismos de mantenci6n. Resulta dudo sa,
al menos por ahora, la utilidad de una definici6n basada en atributos
que se desconocen. Por convenci6n se ha optado por definir un poli-
morfismo como La ocurrencia, con frecuencia a.preci able , de dos 0 =as 2 5
alelos en un lugar gen etico en una misma poblaci6n. (28) Se ha convenido
igualmente en que son frecuencias apreciabl es las iguales 0 superiores
al 1%. De acuerdo con este cr it e rio, que se adopt ara , se dir a que un
locus genet.ico es polim6rfico en una poblaci6n, cuando los alelos tengan
frecuencias entre 0,01 y 0,99, 0biencuando la frecuencia de los hete-
rocigotos sea igual 0mayor de 2 pq = 2(0,01 x 0, 99) ~ 0,02.
Es importante saber en primer lugar si un dado polimorfismo es
tranei ior-io , es de ci r , siun alelo esta en proceso de reemplazar a otro
en una poblaci6n, 0bien si el polimorfisrno es estable. Para que un
polimorfismo rerina esta ultima caracterlstica, deben al menos parti-
cipar en su mantenci6n dos fuerzas de signo opuesto. En 10 que siguese pas ar a revista a varios mecanismos demantenci6nde polimorfismos
estables.
ALGUNOS MECANISMOS DE MANTENCI6N DE POLIMORFISMOS
ESTABLES
alelos A y B con las respectivas frecuencias p y q. A muta a B con una
tasa de mutaci6n u y B a A con una tasa v.
A I ; ; B .v
En cada generaci6n una cantidad p J . . l de genes A mutan a B y una cantidad
qv de genes BaA. Se estabilizan las frecuencias gerii cas de A y B
cuando

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 30/82
P J . . l = qv. [5. 1]
De rn.odo que la frecuencia de A en equilibrio, P., es:
P.J.. l (1 - P.)v
p,J ..l v - up;
P.J.. l + Pev V
p,(J..l+v) = v
vP. = - - .
J.. l+v [5.2J
En el caso que I.l y v sean de la rn.isrn.a rn.agnitud, Fe sera igual a 0,50.
Hasta este rn.orn.ento se ha analizado la situaci6n en que el heterocigoto
tiene una adecuaci6n bio16gica interrn.edia entre los dos horn.ocigotos
(vease Mutaciones y Selecci6n Natural). Sin ern.bargo, c on ai d er e s e elcaso en que los horn.ocigotos AA y BB tienen un valor adaptativo w : : : ; 0, y
los heterocigotos AB tienen un valor adaptativo W = 1. Intuitivarn.ente,
se puede reconocer que las frecuencias g eni c as deA y B se es t.abiHz a.ran
en 0,50, debido a que s610 los heterocigotos sobreviven. Este es el
caso de los Let-al-ee equilibrados. Fisher(56) fue el prirn.ero en recono-
cer que la desventaja de los horn.ocigotos (0 en forrn.a equivalente la
ventaja de los heterocigotos) origina un pol.irrio r fisrn.o estable.
2 6Cons ide r e s e una poblaci6n en equilibrio de Hardy- Weinberg.
Frecuencias
Genotipos
Adecuaci6n
Bio16gica Antes de Actuar
la Selecci6n
Des pue s de Actuar
la Selecci6n
AA W ~A
AB
BB
Totales
La frecuencia, pi, del gen A des pue s de actuar la s elecci6n natural sera
proporcional a
p r-o po.r ct oria a
El cociente entre arn.bas frecuencias g eni cas sera:
Lq '
p(WAAP + wAaq )
q (w~ aP + U8aq)[5. 3]

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 31/82
de modo que cuando pi = P y q I = q (p + qtablece un equilibrio estable
1), vale decir cuando se e s -
p'q
pq'
W A A P + w A sq
W A S P + w ssq
1
W AS P + W ss - %sp
p. [5.4]
En este caso el valor adaptativo de los heterocigotos W AS = 1, de modo
que SAB = 0, ya que, por definici6n, 8 = 1 - W (veas e Mutaciones y Se-
lecci6n Natural). N6tese aderna s que SA A = 1- W A A = W AS - W A A Yss s = 1-
- W e I ! = W AS - %8. Si se reemplazan los valores adaptativos en la expre-
si6n [5.4] por los coeficientes de selecci6n, se tiene:
p. = SSB
SA A +8ss
[5. 5]
De modo que la frecuencia, P ., del gen A en equilibrio depender a exc1u-
si.vamente de los coeficientes de selecci6n en contra de los hornocigotosAA y BB. De modo anal ogo la frecuencia, q., del gen Ben equilibrio sera:
[5.6]8A A +8 ss
CONDICIONES PARA LA ESTABILIDAD DE UN POLIMORFISMO
Exanlinemos a continuaci6n las condiciones necesarias para la es-
tabilidad de un polimorfismo, analizando 1araz6n de carnbio de las fre-
cuencias gerrica s por generaci6n. (<8) Sup6ngase que,
6p = pi -P,
donde pi YP son las frecuencias del gen A en dos generaciones sucesi-
vas. Si t s p es pos i.ti.vo, la frecuencia genica de A tiende a aumentar, y
a disminuir. Utilizando la ecuaci6n 5. 3] se
2 7
puede escribir
pi [5.7]
Si se reemplaza W AA ' W A S YW ss por 1 -SAA, 1Y1 -sss respectivamente, se
obtiene:

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 32/82
6p p' -p
6p p' -p
6p p' -p
P'p((l-sAA)P +q] +q[p + (l-ssB)q]
p ' (5. 8J
p(l-sMP) - P(1-SAAp2-ssat)
1- SA,.p2 - 8aat
pq( -8 AAP + 8ae q )
1 -8AAp2 -8aaq2
(5. 9J
N6tese que 1a expresi6n entre corchetes es siern.pre positiva, de modo
que el signo de p d ep erid er a exclusivarn.entede (-8AAP + 8ssq). Estacan-
tidad puede ser reescrita, utilizando1as ecuaciones [5. 5J Y [5. 6J, co-
rno :
2 8
De modo que
(8 AA + SBa )(Pe - pl.
6p p' p [5. 10]
Ahora se est a en condiciones de exarni n ar varias alternativas. Si los
coeficientes de se1ecci6n en contra de los horn.ocigotos, 8A Y sss, son
rn.ayores que cero, se tiene e1 caso de ventaja het eroci.qot:a 0 sobredo-
minancia en adeouaaion bioZ6gica. Es preferib1e no us ar e1 te.rrrrin o
heterosis, que s e refiere a la superioridad observada en los individuos
de 1a F1cuando se cruzan dos Iinea s puras. Dobzhansky(21) distingue 1a eu-
heterosis, que se refiere a la ventaja en adecuaci6n bio16gica de los hi'-
bridos, y la exuberanci.a hibx-i.da, en que los hibridos poseen caracterls-
ticas flsicas superiores a las de los individuos de razas puras, como,
por ejern.p10, rn.ayor t arnan o 0mayor peso, pero no tienen necesaria-
rn.ente una mayor adecuaci6n bio16gica. Mas arin , en 1a rnay or Ia de los
casos estos hibridos no son fertiles.
Si se exarn.ina rna s detenidarn.ente 1a ecuaci6n [5.10J se observa
que e1 signo de t s p depende de los signos de (8AA + Sse) Yde i», - p), pues-
to que, corn.o se ha dicho

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 33/82
siempre sera positivo. Si SAA Ysse son positivos, y la frecuencia p del
gen A es mayor que P ., la frecuencia de A en equilibrio, I : : . p sera nega-
tivo y la frecuencia de A disrrrinuira hasta alcanzar la frecuencia de
equilibrio dada por [5.4]. En caso contrario, cuando p es menor que
P., 6p sera positivo, y por consiguiente, la frecuencia de A aurne nta ra
hasta alcanzar nuevamente la frecuencia de equilibrio.
Si los coeficientes de selecci6n SAA Ysse son negativos, es decir si
el heterocigoto est a en desventaja, la cantidad (SAA + sse) sera negativa.
Por consiguiente, si la frecuencia p, del gen A, es mayor de P., P sera
positivo y la frecuencia gerrica de A seguira aumentando a.lej and ose cada
vez mas de la frecuencia genica en equilibrio. Si , por el contrario, P
es menor de P., t s p sera negativo yP seguira disminuyendo. En el caso
de que p sea igual a P., se es tabIe cera un equilibrio ine stable. Es obvio
que, siSAA Ysse son iguales a cero, no hay selecci6n natural en opera-
cion.
TIPO DE EQUILIBRIO
ESTABLE INESTABLE
,--~
~
~
2 9
AA AB BS AA AS SS
WAA
WAB W
BBWAA
WAB
WBB
<1 1 <1 >1 1 >1
Fig. 7. Condiciones para la estabilidad
de un polimorfismo.
EL COEFICIENTE DE RETORNO AL EQUILIBRIO
Cabe hacer notar que si I : : . p es funci6n lineal de p (situaci6n que se
cumple en un intervalo apreciable de valores de p) la ecuaci6n [5.10]
se podrIa reescribir
ts p = -b (p - Pel, [5.11]
donde b es el coeficiente de regresi6n 0 la pendiente de la recta de re-
gresi6n. Este coeficiente mide la »eei.l-ienci a de un equilibrio estable,
es decir, su capacidad de volver a un estado de equilibria, tras haber

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 34/82
3 0
sido desplazado de til. Malecot("") ha denominado al coeficiente b , eoef'i-:
ci.ent:e de r-etorno al equilibrio.
En forma similar se puede representar la raz6nde cambio de p en
los casos de equilibrio eatable determinadoa por presiones mutaciona-
les recurrentes que operan en sentido inverso.
! : J p [5.1Z](IJ + v) (p - P.)·
Resulta evidente que en este caso el coeficiente de retorno es muy
bajo, puesto que depende de las tasas mutacionales, que, como se ha
visto, se estimande una magnitud de 10-6 a 10-5 por gameto y por gene-
raci6n. Se puede entonces concluir que el equilibrio estable determi-
nado por el interjuego de presiones mutacionales antag6nicas tiene una
resiliencia menor que el debido a laventajadel heterocigoto, salvo enel caso en que los coeficientes de selecci6n sean de la mis:ma magnitud
que las tasas de rnut a ci on,
NIVELES DE VARIABILIDAD POLIM6RFICA
La existencia de niveles en extremo elevados de heterogeneidad ge-
n etica en poblaciones naturales de distintos organismos, incluso el
hombre, ha side uno de los descubrimientos mas i:mportantes de la ge-
n etica experimental de poblaciones. Shaw,(23) mediante tecn ica s elec-
t r ofo r et.ic aa , en cont r o que 6 de 8 deshidrogenasas eran poli:m6rficas
en elroedor Peromuscue . Estudios similares en Drosophila peeudoobe-cura realizados por Lewontin y Hubby(24) yen Homo por Harris(26) co-
rroboraron los hallazgos iniciales de Shaw y contribuyeron a demos-
trar que cercade1300/0 de los loci gen eti cos responsables de enzimas al-
canzan niveles poli:m6rficos en estos tres organis:mos. Es ad ernaa pro-
bab leq ue se subestime marcadamente la variabilidad existente utilizando
tecnicas eIec t r-ofor eticae , Lewontin(3) supone que s 610se detecta ide la
variabilidad real por medio de estas tecni cas , de modo que en principio
s e podrla conjeturar que cerca del 900/0de los Zoci geneticos son poli-
m6rficos. Los hallazgos de Shaw, Lewontin, Hubby y Harris :mostra-
ron que la hip6tesis c Ia sic a sobre la :magnitud de variabilidad presente
en poblaciones naturales, s ustentada por Muller y colaboradores, esfalsa y que Ia escuela balancista, encabezada por Dobzhansky, estaba
en 10 cierto (vease Antecedentes Hist6ricos).
Uno de los pri:meros poli:morfismos detectados en poblaciones h.u-
manas y en monos antropoides, ad em as de los grupos y tipos sangulneos3D
Se han descrito otros en poblaciones hu:manas, tales co:mo la visi6n
an6mala de colores, la capacidad de percibir el olor de determinadas
sustancias qufmtc a s , la cornpoai c idn de cerumen, los antfg eno s conte-
nidos en las ce Iul.a s sangufneas (antfgenos eritrocitarios y leucocitarios
principalznente), y algunas protei'nas s eric as y enzimas eritrocitarias
(veans e las Tablas Z y 3).
5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 35/82
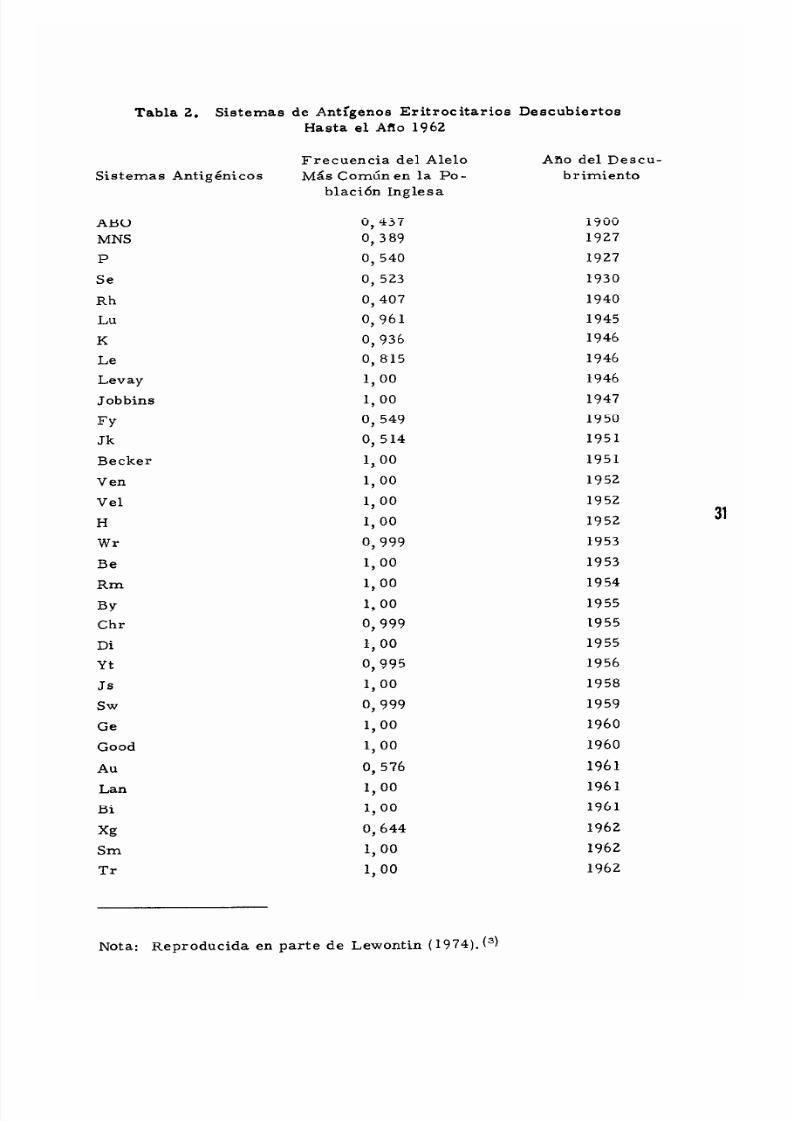
Tabla 2. Sistemas de Antlgenos Eritrocitarios Descubiertos
Hasta el AtIo 1962
Frecuencia del Alel0 Ano del Descu-
Sistemas Anttg eni cos Mas Corntin en la Po- brimientoblaci6n Inglesa
ABO 0,437 1900
MNS 0,389 1927
P 0,540 1927
Se 0,523 1930
Rh 0,407 1940
Lu 0,961 1945
K 0,936 1946
Le 0,815 1946
Levay 1,00 1946
Jobbins 1,00 1947
Fy 0,549 1950
Jk 0,514 1951
Becker 1,00 1951
Ven 1,00 1952
Vel 1,00 1952
H 1,00 1952
3 1
Wr 0,999 1953
Be 1,00 1953
Rm 1,00 1954
Bv 1 00 1955
Chr 0,999 1955
Di 1,00 1955
Yt 0,995 1956
Js 1,00 1958
Sw 0,999 1959
Ge 1,00 1960
Good 1,00 1960
Au 0,576 1961
Lan 1,00 1961
.I:H 1, UU '7U'
Xg 0,644 1962
Sm 1,00 1962
Tr 1,00 1962
Nota: Reproducida en parte de Lewontin (1974). (3)

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 36/82
Tabla 3. Protefnas Sangufneas de las Cuales Hay Maa de Una
Variante Descubierta Antes del At!o 1968
ProtelnaAno del
Descubrimiento
Ndrne ro de
Variantes
Me todo de
Distinguirla
Hemoglobina
Cadena alpha
3 2
Cadena beta
Cadena gamma
Cadena delta
Supresiones
Haptoglobina
Cadena alpha
Cadena beta
Grupos Gm
Transferina
Seudocolines-
terasa
Estearasa locus El
Estearasa locus Ea
Protelna G-C
Albrirrrina
Inv
Fosfatasa placental
alcalina
Lipoprotelnas AG
Lipoprotelnas LP
Glucos a- 6-fosfato
deshidrogenasa
Anidrasa carb6nica
Estearasa eritro-
citaria
Fosfatasa acida
Catalasa
6-Fosfato gluconato
deshidrogenasa
Mioglobina
Fosfoglucomutasa
Fosfoglucomutasa
Fosfoglucomutasa 11
Fosfoglucomutasa III
Adenilatokinas a
Lacta sa oesnloro-
genasa
Subunidad A
Subunidad B
Amilasa
Fibrin6geno
26
46 +
5
4
3
13
10 +
3
26 +18 +
3 +2 +
6 +
7 +
3 +
9 +5
2
25 +
5
2
5 +
3
7 +2
8 +
5 +2 +4 +
5
1
4 +
3
Secuencia de
Arrri n oa c id o s
Electroforesis
Inmuno16gico
Electroforesis
Inhibici6n
Electroforesis
Electroforesis
Electrofor esis
Inmuno16gico
Electroforesis
Inmuno16gico
Inmuno16gico
Electroforesis
Electroforesis
Electrofor esis
Electroforesis
Electrofor esis
Electroforesis
Electrofor esis
Electroforesis
Electroforesis
Electrofor esis
Electrofor esis
Electroforesis
Electroforesis
Electroforesis
Electroforesis
1949
1955
1956
1957
1957
1959
1960
1960
1961
1961
1961
1962
1962
1962
1963
1963
1963
1963
1964
1965
1965
1965
1965
1965
1965
1965
continua

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 37/82
Continuaci6n
ProtelnaNdrriero de Metodo de Ano del
Variantes Distinguirla Descubrirniento
AHa, glucoprotelna
ac ida 2 + Electroforesis 1965
Beta lipoprotelna 2 Electroforesis 1965
Galactosa-l-fosfato
uridil transferasa 2 + Electrofor esis 1966
Malato deshidroge-
nasa (soluble) 2 Electroforesis 1966
Protelna Xm 2 + Inrnuno Iogico 1966
AHa antitripsina 3 Electrofor esis 1966
Malato deshidroge-
nasa (mitocondrial) 2 + Electroforesis 1967
Glutati6n reductasa 2 + Electroforesis 1967
Ceruloplasmina 3 + Electrofor esi s 1967
Peptidasa A 4 + Electrofor esi s 1967
Peptidasa B 4 Electrofor es is 1967
Hipoxantina -guanina
PR transferasa 3 Estabilidad calor 1967
G'lut.arni c oxa.Iac et ico
transaminasa 2 Electroforesis 1967
NADH diaforasa 2 Electroforesis 1968 3 3
Fosfohexosoisome-
rasa 10 + Electroforesis 1968
,"VLo.; .wo." l,"VLC'Ho." ~ ,~""" "v.. uue. ,,"u:.o Ll"""" <11e.:; ce uue. 'e."~1d."-
te corruin, Reproducida en parte de Childs y der Kaloustian
(1968). (40)
POLIMORFISMOS POSIBLEMENTEMANTENIDOS POR VENTAJA HE-
TEROCIGOTA
Las fuerzas evolutivas que manti enen Ia heterogeneidad genetica
descrita no son cono cida s arin, salvo contadas excepciones. Entre los
rne cani srnos que han recibido atenci6n de preferencia esta la sobredo-
rrrirrancia en adecuaci6n bioIogi ca. Los ejemplos mejor conocidos de
1 ' ," ,1 ,1 '.:1. • \.sr , .r- ~J
terocigota, son los de las hemoglobinas. Hay varios indicios de
que los heterocigotos AS tienen mayor valor adaptativo que los in-
dividuos homocigotos SS y que los individuos normales. (33) Breve-
mente, entre las Ifne as indicativas se pueden mencionar que (1) se ha
encontrado una correlaci6n positiva entre Ia frecuencia del gen S y la
distribuci6n de la malaria, (31) (2) Allison, en 1954, infect6 a 15 indivi-
duos normales y a 15 individuos heterocigotos voluntarios con Plasmo-
dium falciparwn y encontr6 que 14 individuos normales adqui rfan la

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 38/82
enfennedad, en contraste con s610 2 casos entre los heterocigotos; (3)
se obtuvo un rnayor recuento de pa r a sitos por unidad voIurnen de sangre
entre individuos norrn.ales que entre heterocigotos; y (4) fina.Irriente , en-
tre los pacientes hospitalizados con rna Iar ta , una rnayorla significativa
de individuos resultaron ser hornocigotos AA . Los polimorfisrnos res-pecto de otras hemogIobina s ano r rna le s , el de la anerrria medit er r anea
y la glucosa 6 fosfato deshidrogenasa, aca so sean rnant erridos por rne-
canisITlos semejantes.
Si todos los Loci. genett cos fueran rrrant erridos a niveles po lirndr ficos
en forrria independiente por ventaja heterocigota, debier arrios postular
una diferencia de adecuaci6n bio16gica exagerada entre el heterocigoto
multiple y el promedio de la poblaci6n, 10que implicarla la aceptaci6n de
una carga genet lc a segregacionalinconcebibleITlente alta (vea se Consan-
guinidad). En teorla, estas dificultades pueden ser eliminadas supri-
rniendo la suposici6n de que los Loci. geneticos acttian en Ior rna indepen-diente. Sved, Reed y Bodm er , (32) King(33) y Milkrnan(34) han postulado
que si existe un Hrrrit e superior 0 urnb r al para la adecuaci6n bio16gica,
se puede mantener un niirne r o mayor de Loei. gen eti cos po l'i.rnor fi cos en
las poblaciones por ventaja heterocigota sin que ello obligue a aceptar
una carga segregacional excesiva.
OTROS FACTORES QUE ORIGINAN POLIMORFISMOS
3 4
Es Irnpor tarite destacar que hayvarios otros factores que determinan
equilibrios estables. Entre enos cabe rnencton ar la eeleoeion natural
dependiente de La freeueneia. En este ca so la adecuaci6n bio16gica delos portadores de un deterITlinado genotipo d eperide ra de su frecuencia
relativa en la poblaci6n. Se pueden considerar otras posibilidades, co-
mo la seleeei6n dependiente de la densidad de poblaei6n, la seleeei6n
de signos opuestos en los sexos, la eompensaei6n reproduetiva, el equi-
l-ibr-io entre preeiones evolutivas eietemat-ioae (tales COITlOl f luio ge-
nico) y La dei-iva qenet-ioa; 0 cualquier corrrbinacidn de ellas. Hay que
destacar que si un deterITlinado gen de un grupo de enlace se mantrene
en equilibrio en una poblaci6n 0 bien favorecido por conferir a su por-
tador una ITlayor adecuaci6n. bio16gica, los genes cercanos en el mismo
grupo de enlace se r an "t-ranepotrtadoe" por el gen en cuesti6n.
Cabe suponer que las fuerzas selectivas necesarias para establecer
un polimorfismo han actuado en el pas ado y que algunos Loci. genet'icos
polfrndr fico s son rel-iquiae de or-iei.e seleetivas previas.(29)
Esta hip6tesis es atractiva puesto que la tasa de aparici6n de nue-
vos poHrrior fi s rnos tal vez sea ITlaSalta que la tasa de eHrni.nacidn , en•• , ., .J .,. _, • .J A.J ..(.
'1" J:' ",',
preciso recordar que nurne rosos poHrnor Hsrnos pueden ser transito-
r ioe , 0 sea que un gen este en proceso de r eemp'laaa r a otro en la pobla-
ci6n, 0 bien que se origine un pol irnor Ii arno por la incorporaci6n de ge-
nes nuevos a una poblaci6n por inITligraci6n.
Las frecuencias pol.i.rndrfi cas que presentan en poblaciones hurnanas
los genes de que depende la degustaci6n de la PTC, se ha intentado ex-
plicarlas por una asociaci6n con bocio enderrrico, Por otra parte, la
mantenci6n de los poHrnor fi srnoe Rh yABO se ha tratado de relacionarla

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 39/82
con mecanismos de selecci6n por incompatibilidad, con compensaci6n
reproductiva y con enfermedades. Por des gracia, la asociaci6n del
sistema ABO con ulcera duodenal y gastrica es tan leve que este rne ca-
nismo no basta para mantener el polimorfismo.
Una de las mayores dificultades que persisten para elucidar los me-
canismos de mantenci6n de polimorfismos es la imposibilidad de de-
tectar pequena s diferencias en adecuaci6n bio16gica entre los genotipos
correspondientes a sistemas polim6rficos. Sepuede demostrar que se
requieren muestras de cerca de 300.000 individuos para detectar, por
ejernpl o, una diferencia del 10/0en una segregaci6n mendeliana de 1: 1.
El problema de la mantenci6n de Loci: poHrnor Hcos es motivo de
acalorado debate y uno de los problemas centrales de la genett ca de po-
blaciones por las implicaciones evolutivas que tiene. La probabilidad
que una proporci6n importante de la variabilidadgenetica observada en
poblaciones naturales sea selectivamente neutra ha permitido postular
modelos de evoluci6n no darwiniana. El dilema no ha sido resuelto en
forma satisfactoria y en ciertas ocasiones el rnismo conjunto de datos
ha sido utilizado por los" seleccionistas" y por los" neutralistas" para
apoyar sus respectivos puntos de vista. Por ejemplo, Kimura y Ohta(l)
partidarios de la teor Ia neutralista 0 neocIas ica , concluyen que la rna-
yorIa de los polimorfismos son selectivamente neutros, y se mantienen
en las poblaciones por un equilibrio entre factores evolutivos direccio-
nales (mutaciones) y por deriva genetica. Una proporci6n muchomenor
serIa mantenida por sobredominancia en adecuaci6n bio16gica 0por otro 3 5
tipo de selecci6n, y por ultimo, una pequenafracci6n estarla represen-
tada por polimorfis:mos transitorios. Los autores aludidos le atribu-
yen gran i:mportancia evolutiva a esta pequ ena fracci6n.
LA RELAJACI6N DE LA SELECCI6N NATURAL
rencias significativas de la frecuencia de casos de discro:matopsias con-
genitas en grupos de poblaci6n de abor fgenes de Africa, de Australia
y de America, y poblaciones tecno16gica:mente mas avanzadas de Euro-
pa. La frecuencia es de un pr ornedro, en poblaciones aborlgenes, de
1,50/0, mientras que en poblaciones europeas alcanza un 7%.(35-$) Para
explicar este hecho, Neel y Post (3S}suponen que los genes de pobIacio-
nes tecno16gica:mente avanzadas causantes de las di scrorrrat ops i.as con-
genitas, han experi:mentado un aumento en grado sustancial por efecto
de un me cani srno de relaiaoion de La eeleceion natural en contra de 105
individuos afectados. Post(37) ha agregado otros ejemplos de caracte-
rIst.ic as sometidas a control geneti co parcial que podrlan tarnb ien estar
aurnentandoenpoblaciones "civilizadas" mediante eIfen6meno de rela-
jaci6n de la selecci6n natural. Son ellas: rni opfa , ano:mahas del septum
nasal y apertura anormalmente pequena del conducto auditivo externo.
A la luz de recientes descubrimientos dentro del marco de la bio-
qufrrric a , la hip6tesis de la relajaci6n de la selecci6n natural podrIa
te6ricamente tener i:mplicaciones medicosociales importantes. Por
ejernplo, la fenilquetonuria, enfermedad autos6mica recesiva, puede
ser hoy diagnosticada en for:ma precoz. Estopermite poner a los indi-

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 40/82
con mecanismos de selecci6n por incompatibilidad, con compensaci6n
reproductiva y con enfermedades. Por des gracia, la asociaci6n del
sistema ABO con ulcera duodenal y gastrica es tan leve que este rne ca-
nismo no basta para mantener el polimorfismo.
Una de las mayores dificultades que persisten para elucidar los me-
canismos de mantenci6n de polimorfismos es la imposibilidad de de-
tectar pequena s diferencias en adecuaci6n bio16gica entre los genotipos
correspondientes a sistemas polim6rficos. Sepuede demostrar que se
requieren muestras de cerca de 300.000 individuos para detectar, por
ejernpl o, una diferencia del 10/0en una segregaci6n mendeliana de 1: 1.
El problema de la mantenci6n de Loci: poHrnor Hcos es motivo de
acalorado debate y uno de los problemas centrales de la genett ca de po-
blaciones por las implicaciones evolutivas que tiene. La probabilidad
que una proporci6n importante de la variabilidadgenetica observada en
poblaciones naturales sea selectivamente neutra ha permitido postular
modelos de evoluci6n no darwiniana. El dilema no ha sido resuelto en
forma satisfactoria y en ciertas ocasiones el rnismo conjunto de datos
ha sido utilizado por los" seleccionistas" y por los" neutralistas" para
apoyar sus respectivos puntos de vista. Por ejemplo, Kimura y Ohta(l)
partidarios de la teor Ia neutralista 0 neocIas ica , concluyen que la rna-
yorIa de los polimorfismos son selectivamente neutros, y se mantienen
en las poblaciones por un equilibrio entre factores evolutivos direccio-
nales (mutaciones) y por deriva genetica. Una proporci6n muchomenor
serIa mantenida por sobredominancia en adecuaci6n bio16gica 0por otro 3 5
tipo de selecci6n, y por ultimo, una pequenafracci6n estarla represen-
tada por polimorfis:mos transitorios. Los autores aludidos le atribu-
yen gran i:mportancia evolutiva a esta pequ ena fracci6n.
LA RELAJACI6N DE LA SELECCI6N NATURAL
rencias significativas de la frecuencia de casos de discro:matopsias con-
genitas en grupos de poblaci6n de abor fgenes de Africa, de Australia
y de America, y poblaciones tecno16gica:mente mas avanzadas de Euro-
pa. La frecuencia es de un pr ornedro, en poblaciones aborlgenes, de
1,50/0, mientras que en poblaciones europeas alcanza un 7%.(35-$) Para
explicar este hecho, Neel y Post (3S}suponen que los genes de pobIacio-
nes tecno16gica:mente avanzadas causantes de las di scrorrrat ops i.as con-
genitas, han experi:mentado un aumento en grado sustancial por efecto
de un me cani srno de relaiaoion de La eeleceion natural en contra de 105
individuos afectados. Post(37) ha agregado otros ejemplos de caracte-
rIst.ic as sometidas a control geneti co parcial que podrlan tarnb ien estar
aurnentandoenpoblaciones "civilizadas" mediante eIfen6meno de rela-
jaci6n de la selecci6n natural. Son ellas: rni opfa , ano:mahas del septum
nasal y apertura anormalmente pequena del conducto auditivo externo.
A la luz de recientes descubrimientos dentro del marco de la bio-
qufrrric a , la hip6tesis de la relajaci6n de la selecci6n natural podrIa
te6ricamente tener i:mplicaciones medicosociales importantes. Por
ejernplo, la fenilquetonuria, enfermedad autos6mica recesiva, puede
ser hoy diagnosticada en for:ma precoz. Estopermite poner a los indi-

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 41/82
6CONSANGUINIDAD
MATRIMONIOS CONSANGUINEOS
Se dice que dos individuos son consanguIne os cuando tienen a.I menos
un antepasado corridn. Se podrIa postular que en algiin grado todos los
individuos de Ia rni sma especie son cons ang ufn eo s, aunque no se este en
condiciones de reconstruir sus Hneas genea16gicas que convergen en un
antepasado comiin. Sin embargo, al estudiar los efecto s de la consan-
guinidad se cent rara La atenci6n pormera convenci6n en aque llo s ante-
pasados comunes que se remonten a cuatro generaciones. No se tend ran
en cuenta los ant epa sa.dos remotos porque en tales casos Ia consangui-
nidad es diHci.l de verificar Y, por 10 tanto, el termino pierde sentido.
La descendencia de una pareja cons arigufne a es endoqama . Los individuo s
end6gamos pueden ser portadores de un doble juego de los genes del
antepasado comiin de sus padres. Por consiguiente, pueden ser homo-
cigotos respecto a genes i.dent iooe en origen. Es precisodistinguir ge-
nes identicos en origende genes identicos en funci6n. En principio, los
primeros son copia s de un mismo gen perteneciente aalgiin antepasado
comiin y los iiltimos songenes similares, sibien provienen de eventos 3 7
mutacionales distintos.
En el caso que el antepasado cormin haya sido portador unico de
algun gen recesivo "desfavorable", es posible que descendientes en-
d6gamos de este individuo sean portadores de un juego doble del
mismo gen. Serra de esperar, por 10tanto, que los individuos end6-
gamos tuvieran una probabilidad mayor de sufrir enfermedades gene-
ticas recesivas.
Tal vez sea esta una de las tantas razones de que en va rios parses
prohiban ciertos matrimonios cons ang ufne os . La iglesia catoIica , por
ejemplo, prohibe los matrimonios incestuosos entre padres e hijos yentre
hermanos, yexige una dispensa para celebrar matrimonios entre tfos y
sobrina s, tras y sobrinos, primos hermanos y primos en segundo grado,
entre otros. Por otra parte, ciertas sociedades favorecen 0han favoreci-
do los rria.t r-i.rn.oni os cons ang ufne os : tal es el ca so de los Incas del Peru,
que durante varias generaciones se casa ron con hermanas, y el de los Fa-
raones egipcios. Ba sando se en ciertas creencias populares, que proba-
blemente surgieron como consecuencia de la observaci6nde ciertos casos
de descendencia end6gama afectada por enfermedades hereditarias rece-
sivas, nurnerosas personas rechazan los matrimonios consangufne os.
Incluso en la literatura hispanoamericana cont.ernpo rane a se encuentran
ejemplos de este rechazo, como se advierte enalgunos pa sa je s de lano-
vela" Ci.enAfios de Soledad", del colombiano Gabriel GarcIa Marquez.
EL COEFIC IENTE DE ENDOGAMIA
Los individuos pueden tener diversos grado s de endogamia, debido
a que las relaciones biol6gicas entre los padres pueden va ria r , El

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 42/82
coeficiente de endoqamia representa una forma de valorar estas diferen-
cias. Sewall Wright,(41) a quien se debe el concepto, defini6 el coe-
ficiente de endogamia como el grado de "correlacion entre gametos en
proceso de union", y dedujo la siguiente formula general para calcu-
larlo
donde FAes el coeficiente de endogamia de cualquier antepasado co rmin
que enlaza las l{neas ancestrales materna y paterna. Se designan co-
mo nd Y n. el mime ro de generaciones desde la madre y el padre a este
antepasado comiin. La contribuci6n de cada enlace de las genea logIas
materna y paterna corresponde a
(r:(l + FA).
EI coeficiente es Ia aurna de estas contribuciones. Mas recientemente,
Malecod42) ha definido el coeficiente de endogamia como La probabili-
dad de que un individuo posea, en un mi srno locus de sus cromosomas
h0H101ogos, des genes idintic()t; en Cl1,igen,es dech, plovenientes de un
solo evento mutacional. Se puede demostrar que el coe£iciente de co-
rrelaci6n precedente equivale a esta probabilidad.
3 8
En 10 que sigue se intenta ra deducir el coeficiente de endogamia por
procedimientos tradicionales, a fin de presentar luego el metodo de A.
Kudo que ofrece ventajas importantes. Consideremos el caso de 1, pa-
dre de 3 y 4, abuelo de 5 y 6 y bisabuelo de P, el propositus (Fig. 8).
III
II
I
Fig. 8. Genealogia q ue ilus-
tra el calculo del coeficien-
te de endogamia, s eg ti n e l m e -todo tradicional.
Se parte del supuesto que 1 posee en un determinado locus el gen A. La
probabilidad de que t rans mi.ta este gen a 3 es 1/2 (suponiendo que 1es un

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 43/82
individuo diploide no rrnal y que el locus es auto sdmtco ): Ia probabilidad
de que 3 10 t r an s mi.ta a 5 es 1/2, y fi na.Irne nte , que 510 transInita a P
es 1/2. Po r 10 tanto, la probabilidad de que una c opi.a del gen A este
presente en P, COInO conseeuencia de su transInisi6n a t rave s de 3 y 5,
es 1/8, es decir , el producto de las probabilidades de los t r e s paso s .
De igual modo, se puede cal cul a r las probabilidades del paso del gen de
1 a 4, de 4 a 6 y de 6 a P. Debido a que hay dos sendas independientes
que unen a 1 con P , la probabilidad que A sea t r an s rni.t ido por las dos
sendas dando eOInO resultado queP sea ho rno ci goto para genes identicos
en origen, es:
y en fo r ma mas general
donde n, es el niirne r-o de generaciones que separan al padre de P del
antepasado cormin 1, Y nd es el mirne r o de generaciones que se pa r ari a
la rnad r e de P del rrri In a e as c '
ci6n aepar a a P de su padre y otra 10 s epa r a de su rnad r e). Sin e rnba r »
go 1 posee otro gen en el rrri srno locus, digamos B, respecto del cual son
validas las mi srna s consideraeiones. Si estamos aho r a interesados en
caleular la probabilidadde que P seahomocigoto para cualquiera de los
dos genes presentes en 1 (es decir A 0 B ) se obtiene:
2 ( ~ r , + n d + 2 = ( t ) " , + n d + 1
Ahora bien, si3 y 4 tienen madre cormin, 2 en el e sq uerna , las considera-
c iorie s hechas con respecto a los genes de 1 son igualmente v.Hidas
respecto de 2. F'Ina.lrrient.e, la probabilidad (F) de que P sea homo c i.goto
para copias de los euatro genes presentes en 1 6 2 sera:
( )
n + n + 1F=L:t
Sd[6.1 J
En esta f6rInula, la sumatoria, L:, incluye las diferentes sendas que unen a
P con sus antepasados ccmune s , 1 y 2. En el caso de que 1 6 2 (0 ambos)
fueran enddgarno s , ser fa preciso rnod iftc a r la f6rInula [6. 1J para teneren consideraci6n la probabilidad (ahora rnayo r ) de que 1 (y/o 2) sean
horno ci.goto s. En e ste caso, la contribuci6n de una de las sendas cor res-
pondientes al antepasado COInUn1 (6 2) al coeficiente de endogamia sera:
[ 6 . 2 J
a rnbo s 1 Y 2 fue r an enddgarno s se t.end rfa :
[ 6 . 3 J
La modtfic ac idn hecha se puede explicar haciendo referencia a la figu-
r a 8. Hemos considerado que P podr fa ser homocigoto para dos copias
de un gen presente en 1, y reconocemos eneste caso dos posibilidades,
vale decir los genotipos AAyBB. Sise supone que A y B fueran a su vez
3 9

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 44/82
copias de un mismo gen presente en al.gun antepasado de 1, P t.a.rribien
se rfa homocigoto para copias de un rnis rno gen en el ca so que sus ge-
notipos fueran AB 0BA. La probabilidad que esto ocurriera se rfa , ra-
zonando en forma similar a Laanterior
La probabilidad de que A y B sean identicos es FH • De modo que, si se
sabe que 1 es end6gamo y su coeficiente de endogamia es FH, se tiene:
p(AA) = ( ~ f + n d + 2 ,
(~ )nB+nd+ . a ,
p(BB) "
p(AB 0BA) = F H [ C ~ ) n . + n d + 2 + ( t ) " · + n d + 2 ]
Si se consideran estas alternativas en forma simult anea
F ' = p(AA) + p(BB) + p(AB 0 BA)
se tiene:
[6.4J
4 0
Este metodo puede ser aplicado en situaciones de diferente complejidad,
sin embargo en el caso de que existan multiples antepasados comunes y
multiples formas de parentesco, es diffc Il enumerar todas las sendasposibles, tal como Loexige el rnetod o, Para evitar esta dificultad se
ha elaborado otro procedimiento, como elde Kudo, (43) en el cualla pro-
babilidad de cometer errores se reduce en grado considerable.
EL METODO DE KUDOPARA CALCULAR EL COEFICIENTE DE EN-
La senda (ve aae La Fig. 8) 1, 3, 5 la denornina Kudo l.inea paterna
y la senda 1, 4, 6, Z{nea materna. Estas dos forman un par de Z{neas
anoeetiral.ee, Aderna s , como las Ifneas paterna y materna contienen el
individuo que se ha optado por denominar 1, diremos que forman una
pveei l la, contrastando, por ejemplo, con las sendas L, 3, 5 y 2, 4, 6
que no contienen individuos comunes. La presilla definida po r 1, 3, 5
y 1, 4, 6 es de longitud L, dondeL = n.+7l,j +1. Sup6ngase, para ilus-
trar con un ejemplo el procedimiento a emplear, que se esta interesado
en computar el coeficiente de endogamia del individuo P en La genealo-gIa que aparece en Lafigura 9.
Es conveniente asigna r mirner o s i.rnpar e s a los hombres que he-
mos simbolizado par cuadrados, y mirnero s pares a las mujeres
representadas por esferas. Se comenaa i-a por enurne ra r en forma ex-
haustiva las Ifne a s paternas y maternas (Fig. 9). Esta enurneraci6n
cornprende en este caso 16 pares de Hne a s ancestrales qlle cornien-
zan en el par 1-3-9 y 2-5-11 Y terrninan en el par 1-4-8 y 2-6-10.
Hay que hacer notar adema s que cada par de Hne as ancestra1es esta
representado por una unica celda en un ordenarniento de 4 X4.
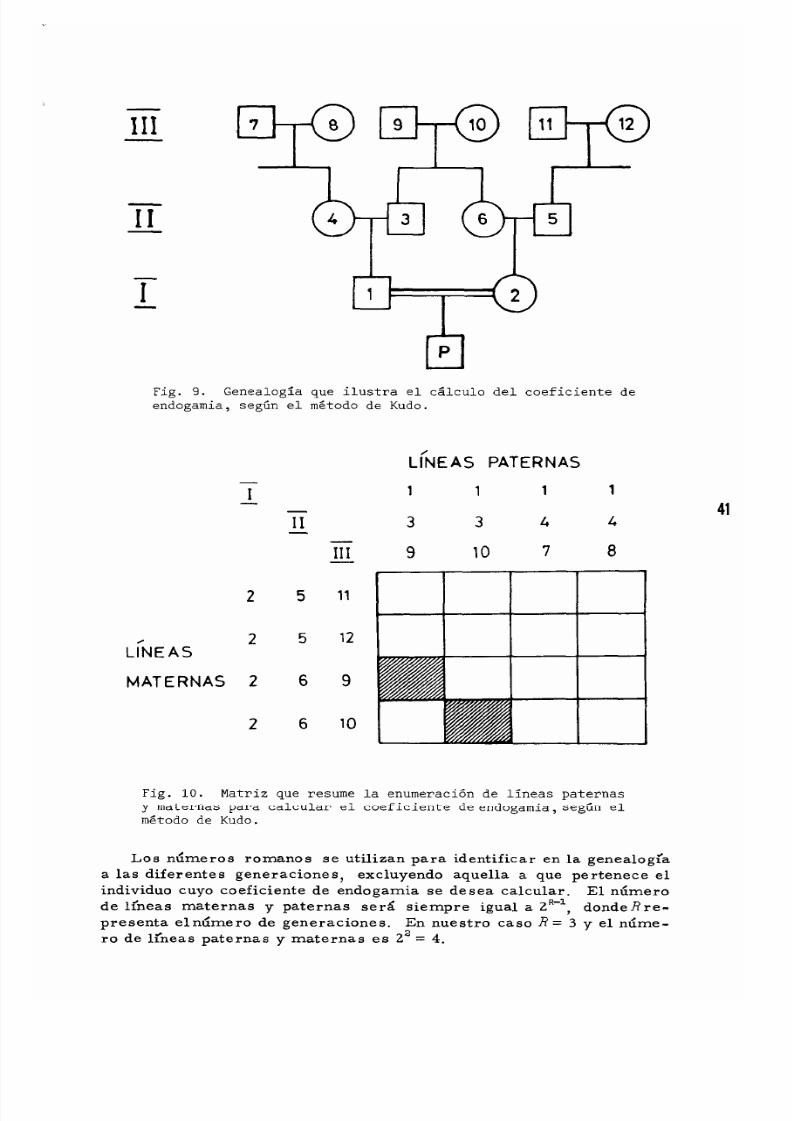
5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 45/82
I I I
I I
I
Fig. 9. Genealogia q ue ilustra el calculo del coeficiente de
endogamia, segun el metoda de Kudo.
II 3 3 4 4
III 9 10 7 8
2 5 11
.- 2 5 12
LlNEAS
MATERNAS 2 6 9
2 6 10
,-
LlNEAS PATERNAS
4 1
Fig. 1 0. Matriz q ue resume la enumeraci6n de lineas p aternas
metodo de Kudo.
Los mirne ro s rornanos se utilizan para identificar en la genealogfa
a las diferentes generaciones, excluyendo aqueUa a que pertenece el
individuo cuyo coe£iciente de endogarrria se desea ca.lcul.ar . El mirnero
de Ifnea s rriater na s y paternas sera ai ernpr e igual a 2R-\ donde R re-
presenta el mirner-o de generaciones. En nuestro caso R = 3 Yel mirne-,
ro de Ifneae paternas y rrrate rnas es 22 = 4.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 46/82
A continuaci6n se pro cede a revi sa r en forma sistematica todas las
celdas correspondientes apares de li"neas ancestrales en que un indivi-
duo detenninado aparezca ai.mul.tarieamente incluido en las Ifnea s pa-
terna y materna. Considereselaprimerali'nea paterna (1, 3, 9). Esta
li'nea no comparte individuos con La p r irne r a Ifnea paterna (2, 5, 11), ni
ta rnpoco con la segunda (2, 5, 12) 0 con Ia cuar ta (2, 6, 10), pe ro sf
comparte un individuo, el numero 9, con Ia tercera Ifnea materna (2,
6, 9). Sombrearnos Ia celda que corresponde a Ia intersecci6n de estas
Ifnea s (las Ifnea s 1-3-9 y 2-6-9). Se continua revisando las celdas y
sombreando las celdas apropiadas hasta completar el examen de todo s
los pares de Hnea.s genea16gicas. En este ejemplo, se encuentra sola-
mente una celda mas, que corresponde a la Ifnea paterna 1-3 -lOy a La
Lfnea materna 2-6-10. El coeficiente de endogamia, F, se calcula se-
gun Kudo:
F Numero de celdas sombreadas2(Ndrner o total de celdas)
[6.5]2(16)
1
16
Es posible cornpr oba r Ia validez de esta formula considerando nueva-
mente el par de Ifne aa ancestrales 1-3-9 y 2-6-9. En este caso, 9 es
el segundo antecesor del padre 1 y el segundo de la madre 2. Por 10
tanto, La contribuci6n de 9 al coeficiente de endogamia debe ser:
4 2El numero de celdas que se deben aornbr ear debido a que 9 es cormin
a las Ifnea s materna y paterna es:
N6tese que Ia formula mas general ser Ia ,
[6.6J
donde R representa de nuevo el mirriero de generaciones. Si se halla el
cociente de estos numeros, es decir el de l a contribuci6n que 9 hace al
valor de F y el mirne ro de celdas que se deben sombrear por ser 9 cormina las li"neas materna y paterna, se tiene:
mere de celdas.
La f6rmula [6. 5J puede utilizarse para aquellas situaciones en que
el 0 los antepasados comunes no sean end6gamos. En el caso en que
10sean, algunas celdas deben sornbrea r se doblemente porque un par
de li'neas ancestrales puede tener dos individuos en comfin. EI coefi-
ciente de endogamia se calcula en este caso por:

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 47/82
Ndrnero de celdas som.breadas + Ndrnero de celdasdoblem.ente som.breadas
(6.7]F 2(Num.ero total de ceIda s)
EL EFECTO DE LA ENDOGAMIAA NIVEL POBLACIONAL
, .vU''Hu<a",,,,,, UUd. P ULLLUUCLeu uuo U'd.C;C;lUU"''' , ""CL '"uupLe-
tam.ente endogarna y la otra en la cual los m.atrim.onios ocurren alazar.
Deslgnese por F la probabilidad de que un individuo endoga.rno sea
hom.ocigoto para un gen determ.inado i.dent'ico en origen. La Er accion
andcgama de la pobIacion constituye una parte F de la pobla cion total y
la fracci6n pamnfctica constituye la parte 1 -F.
La endogam.ia prolongada trae com.o consecuencia un aum.ento de los
individuos hom.ocigotos en Ia pob.la.cidn, de m.odo que en Ia fracci6n en-d6gam.a F afc anz.ar a un valor de 1, y todos los individuos de esta frac-
ci6n seran hom.ocigotos. En el caso que se considere unlugar genetico
con dos alelos codom.inantes A y B, la frecuencia con que se encuentran
los hOm.oclgOtOS7f7f'DD sera ue 1! y Y. respect+ , -y LCLue<ueu-
cia de los genes A y B sera igualm.ente p y q.
Por ot ra parte, en la fracci6n panm.lctica las frecuencias genotlpi-
cas esta ran distribuidas de acuerdo con el teorem.a de Hardy- Weinberg,
es decir, los individuos AA teridrari una frecuencia dep<l, los individuos AB
de 2 pq v, finalm.ente, los BB, de q<l. Recordem.os que la fracci6n en- 4 3
d6gam.a representa una proporci6n F de La poblaci6n total y la fracci6npanm.lctica una proporci6n 1 - F. Si se sum.an los genotipos de las 2
fracciones, es decir, si se obtiene la frecuencia de los 3 genotipos en
Ia poblaci6n total resulta:
Frecuencias en Frecuencias en Frecuencias en
Genotipos la Fracci6n la Fracci6n Ia Poblaci6n
End6gam.a Panm.lctica Total
A A Fp (1 _F)p<l p(p + Fq)
AB 0 (1 - F)2pq 2pq (1 - F)
BB Fq (1 _F)q<l q(q + Fp)
Es de i.nt er e s hacer notar que las frecuencias de los 3 genotipos
van a depender del nivel de endogam.ia de la poblaci6n. Sin ernba r go,
la frecuencia de los genes A y B no se altera por las desviaciones de la
panm.una. Por otra parte, la frecuencia de heterocigotos de Ia pobla-
ci6n decrece gradualm.ente con La endogam.ia hasta llegar a cero. Lom.ism.o sucede en el caso b.i.potetico de una poblaci.on subdividida de ta-
rriafio finito donde no act iia la seIeccion natural, el flujo genico ni las
m.utaciones recurrentes, bajo el efecto de la deriva genetica (veas e
Factores Evolutivos Es toca s ticos}.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 48/82
EL COEFICIENTE DE ENDOGAMIADE LAPOBLACI6N
Por 10 cornrin se define un coeficiente de endogamia promedio de los
individuos que constituyen una poblaci6n. Este coeficiente, que se de-
signa por 0: es igual a
[ 6 . 8 J
donde Pl es la frecuencia relativa de los individuos end6gamos que tie-
nen un determinado coeficiente de endogamia Fl' En la Tabla 4 se
pueden observar los valores de 0: de distintas poblaciones latinoameri-
cana s ,
CARGASGENETICAS
En 1950, H. J. Muller propuso el concepto de carqa qenetri.ca al re-
ferirse ala perdida de cigotos debida a genes que se mantienen en las
poblaciones mediante el equilibrio entre mutaciones y selecci6nnatural.
Mas en concreto propuso el concepto de carqa mutiaci.onal., Ocho afio s
despues, Crow de£ini6 la cal ga genetica COlllO 1adislllinuci611relativa
del valor adaptativo 0 adecuaci6n biol6gica promedio de una poblaci6n
con respecto al valor adaptativo que tendrfa si todos sus integrantes fue-
ran portadores de genotipos con valor adaptativo 6ptimo. Expresado
algebraicamente
4 4L [ 6 . 9 J
donde L representa la carga gerret ica, wmax el valor adaptativo 6ptimo y
weI valor adaptativo promedio. En el caso de que w max = 1
L = 1 - w . [6.10J
La carga genetic a que proviene de genes rnantenidos en una pobla-
ci6n por sobredominancia en adecuaci6n bio16gica (vease Polimorfis-
mos Geneticoe} se denornina carqa de eeqreqao ion, para diferenciarla
de la carga mutacional.
Considerese el caso de un gen letal recesivo cuyo heterocigoto tie-
ne una adecuaci6n biol6gica igual a Ia del homocigoto dorninante.
Genotipo FrecuenciaAdecuaci6n Contribucione s
Bio16gica ala Carga
A P 1 0
Aa 2pq 1 0
aa q2 0 q2
El valor de adecuaci6n biol6gica promedio de la poblaci6n sera p2 X 1 ++ 2pq x 1 + q2 X 0= p2 + Zpq y la carga mutacional sera q2.
Generalizando, las contribuciones ala carga mutacional de un Locue
con dos alelos en una poblaci6n panrnfctica seran:
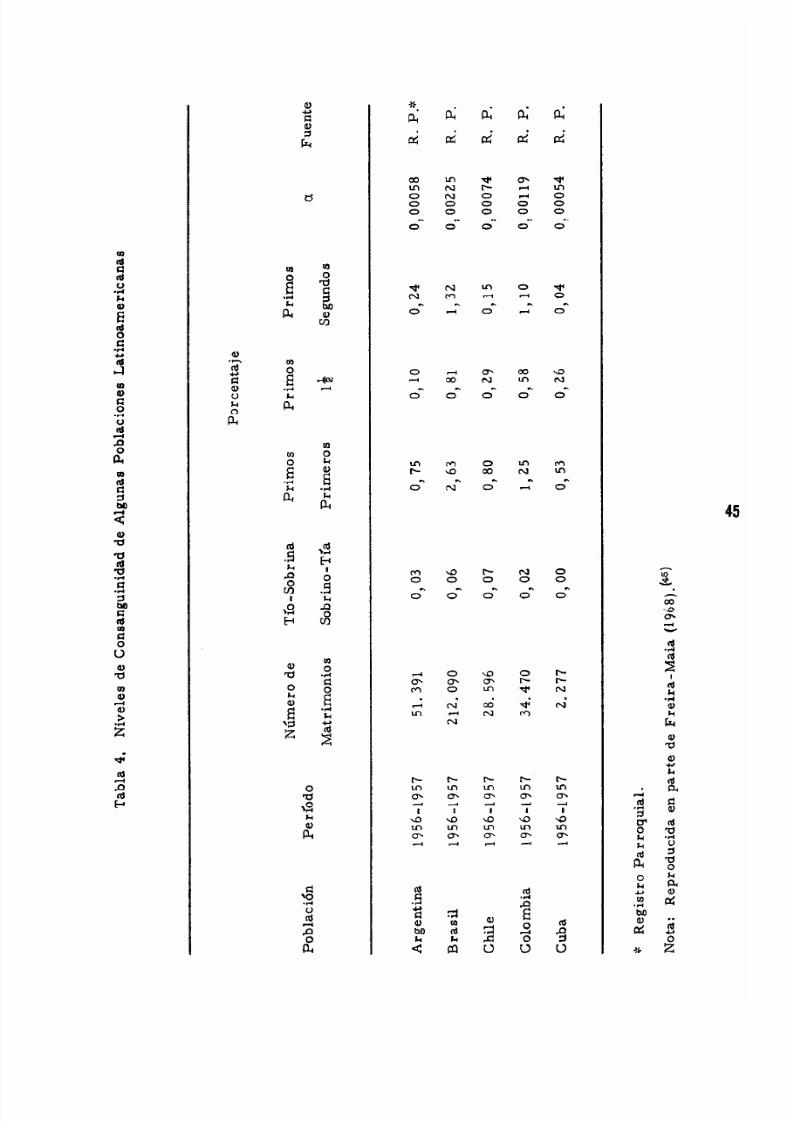
5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 49/82
q)
* p . ; p . ; p . ; p . ;. . .p . .:: i
q)
~0 :: 0 :: r . i r . i 0 ::
' "00 II) ~ 0"- ~II) N r- . . . . . . II)
tl 0 N 0 . . . . . 00 0 0 0 00 0 0 0 0
0 0 0 0 0
UI
I'll . .:: II)
I'll
S0
v '0 ~ N II) 0 ~. . . c :
' ". . . . ~ N r<' I . . . . . . . . . . . . 0
q) ' " lIIl - -a p . . q) 0 . . . . . . 0 . . . . . 0
I'llCI)
0r ::
. . . .q). . .j .;; .,
. . . .8
0 . . . . . 0"- 00 -.0
UII : : i r l J O l . . . . . 00 N II) Nq) . . . . . . . . -) o . . . 0 0 0 0 0
r ::~ c,
0
u p . .I'll. . . . ..tl III0
III 0p . .0 . . . II)
" "0 II)
" "II . S S r- . -.0 00 N II)
I'll -:: i . . . . . . . 0 N 0 . . . . 0
:I p . . . . .tIIJ p . .
4 5. .~q)
'0 III ~~'0 . SI'll . . . r - <'0
.tlI
" "-.0 r- . N 0
~. . . . 0 0 0 0 0 0 .1:: 0 I : : i. . . . CI) . . .
0 0:I I . . . 0 0 00 0tIIJ ~S .o
d 0-.0
I'll r - < CI) 0"-
UI. . . . .
I : : i -nI.
U. . . .
., nI
C D q) 0 ; : g'0 '0 . . . . . . . . 0 -.0 0 r- .
I : : i 0"- 0"- o- r-. r-. I
'"0 0
" "0 II) ~ N
nIq) . . .
S '" '. . . . q) . . . . . . N c O . , . ; N. . . .
q)
S . . . .q)
> . . . II) . . . . . . N
" " . ,z ,~ . . . . N
' "n I
~ q)
'0
.; q)
. . . .III ' ". . . . r- . r- . r-. r-. r- . atl 0 II) II) II) II) II)
~'0 0"- e- o- 0"- o-I : : i
\;..I I I I I :I
C I)
. . . -.0 -.0 -.0 -.0 -.0q) II) II) II) II) II) s n I
p . . e- 0"- o- 0"- o- '0. , . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . U
n I :I
p . . '00
0 . . .I : : i~ . . . A.
«I . . . . q)\Q . . . .
III 0 ::. . . . . .~
. . .o 1 : 1 :;j eenI. . . . . C I)
. , C I)
0 n I C I)
. . a t> O nI ; a . g 0 :: n I. . . . . . .0 . . . . . . 0 0p . . ~ Itl U U o
*Z

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 50/82
Genotipo FrecuenciaAdecuaci6n Contribuciones
Bio16gica ala Carga
A A p2 1
°B 2pq - hs 2pqhs
BB q2 - s sq2
De modo que la carga mutacional total sera sq (2ph + q). En general,
si h es muy pequefio, la carga sera aproximadamente sq2 ~}.l, Y si se
aproxima a r , L ~ 2}.l. De rnanera que la carga mutacional oscil.ara en-
tre estos valores extremos y depende del valor de h.
Las contribuciones de los tr e s genotipo s a Ia ca r ga de mutaci6n en
una poblaci6n end6gama aer an por otra parte
Genotipo FrecuenciaAdecuaci6n Contribucione s
Bio16gica a la Carga
AA p2( I - F) +Fp 1
°B 2pq (1 - F) - hs 2pq(1 - F)hs
BB q2(1 -F)+Fq 1 - s sq2(1 - F) + sFq
4 6De modo que la carga total sera:
L = 2pq (1 -F)hs + sq2(l -F) + sFq.
Esta ecuaci6n puede escribirse tambien
L - [sq2 + 2pqhs] + [sq - 2pqhs _sq2JF [6.11J
y sustituyendo las cantidades entre corchetes por a. y b., se obtiene:
L = a. + b.F. [6.l2J
En una poblaci6n panrnfcttca , donde F 0,
y en una poblaci6n end6garna, donde F
EI cociente
ha servido de base a un extenso desarrollo te6rico iniciado por Morton,
Crow y Miiller(44)destinado a diferenciar entre car gas geneti.ca s de muta-
ci6n yde segregaci6n. Estos autores esperaba n, mediante una estrategia

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 51/82
que se a.naIizara a continuaci6n, resolver Ia disputa entre balancistas y
clasicistas (vease Antecedentes Hist6ricos).
Es posible demostrar quela carga de segregaci6n al igual que La de
mutaci6n es una funci6n lineal del coeficiente de endogamia F. Tene-
mos en el caso de la carga de segregaci6n.
Genotipo FrecuenciaAdecuaci6n
Biol6gica
Contribucione s
ala Carga
A A
AB
BB
p2( 1 -F) + pF2pq(1 - F)
q2(1 -F) + qF
1- S
1
1 - t
sp2(l -F) + pF
°q2(l-F) +qF
La carga total es:
[6.13J
Ysi se sustituye de nuevo las expresiones entre corchetes por a. y b.
L a. + b.F. [6.14J
Cuando F ° nuevamente4 7
Lp = a.y cuando F =
L. a. + b•.
El cociente de ambas cargas geneticas de segregaci6n es:
De acuerdo con Morton, Crow y Muller es posible en principio diferen-
ciar entre car gas geneticas demutaci6n y cargas geneticas de segrega-
ci6n a base de los valores nurnericos esperados de los cocientes:
L.y;.
En efecto, serIa de esperar en el caso de Loci gene;ticos cuya variabi-
lidad se mantenga por equilibrio entre mutaciones y selecci6n natural
(Zoci c1asicos) que:
1es decir que el cociente sea al menos igual a 2h. Si se esta dispuesto
a aceptar que h enpromedio varIa de 0,02 a 0,03, el cociente no debie-
ra ser inferior a 15. Si h vale 0, es decir uno de los genes del locus
dia.Ielico es totalmente recesivo, el cociente debiera ser aun mas alto.
Espe cfftcarnente:

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 52/82
L. = , f i : i 8 J . l J . l 8 = !!~00Lp
si se considera que J .l = 10-4.
En el caso de loci geneticos cuya variabilidad se rnantenga por ven-
taja heterocigota (locibalanceados)2at
L.
7;=ap + tp
ap2 + tq2= - - - 2.
ata + t
Serra de esperar, por 10 tanto, que el valor del cociente fuera nota-
blemente inferior.
Si bien desde el puntodevista te6rico Ia estrategia de Morton, Crow
y Muller es atractiva, por desgracia en la practica no dio los resulta-
dos esperados. En numerosos estudios se ha tratado de estimar los
cocientes
L e
I;
relativos a distintas poblaciones humanas, en especial utilizando esti-
madores de mortalidad. Sin embargo los valores obtenidos son en pro-
medio cercanos a 5,7, es decir, tienen valores que se encuentran casi
exactamente entre los esperados para cargas de mutaci6n y de segre-
gaci6n.8
Se han postulado otros tipos de cargas geriet.icae como, por ejem-
plo, la carg a debida a la incomp atibilidad, la carg a sustitutiva, yaun
otras. No se anaIi.zaran estas cargas por las dificultades de suevalua-
cion e interpretacion.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 53/82
7
CONSTITUCION GENETICA Y MEDIO AMBIENTE
CONSIDERACIONES GENERALES
La manera en que la constituci6n genetrca humana se modifica por
influencia del medio ambiente ha sido tema de prolongada disputa ini cia-.
da hace mas de cien afios y que todavla continua. Por desgracia en va »
rias oportunidades esta poIernica se ha manifestado en terminos vitri6-
licos y poco constructivos. A Francis Galton (vea se Antecedentes
Hist6ricos) corresponde el merito de haber sido el primero en llamar
la a.tencion sobre el hecho de que la variabilidad presente en las pobla-
ciones humanas se debe a la interacci6n de las ca racte rfs t.icas innatas
con las circunstancias medio ambientales en que ocurre el crecimien-
to y desarrollo de los individuos. Con posterioridad a Galton varios
investigadores han descrito meticulosamente, y a veces con fatigado-
res detalles, Ia complejidad de dicha interacci6n. Sobre este punto se
destacan los trabajos de Hogben(46) yHaldane. (47.48)Este ultirrlO en par-
ticular se ha referido a la l6gica con que debe enfocarse el problema.
Lewontin(4S) sostiene que el objeto de las investigaciones relacionadas
con la interacci6n de la constituci6n gerie tica con el medio ambiente es 4 9
elucidar las causas de la plasticidad fenotlpica. Por .~esgracia este
objetivo, equivalente al ana Ii sis de las normae de reacc-z-on de un geno-
tipo con diversos medio ambientes y de diferentes genotipos frente a un
medio ambiente, es por el momento diffciI de abordar. Se han realiza-
do estudios de esta Indole en animales de experimentacion(SO) yen ve-
getales.(Sl)
Debido a las complejidades inherentes al ana Iisis de las ca.usas de
Ia plasticidad fenotlpica se ha intentado abo rda r el problema haciendo
una pa rticion de las desviaciones fenotlpicas individuales del valor fe-
notfpi co medio, es decir un a.nalisis de varianza en vez de un ana Ii sis
de caus as , Es evidente que ambos ana Ii sis son diferentes y su objeto
es dar, salvo contadas excepciones, respuesta a preguntas distintas.
Los diferentes estimadores del componente genett co de lavariabilidad,
obtenidos por el rrietod o de partici6n de varianza, tienen por 10tanto
un usa restringido y estan limitados temporal y espacialmente. En ge-
neral son 5610 utf.Ies en el ca so que no hay interacci6n entre los distin-
tos genottpo s y rriedio amblentes. EspecIflcamente eI anahsIs de va-
rianza corresponde al anaIisis de causas cuando todos los genotipos
va rfan en igual forma con respecto a diversos medio ambientes. Indi-
cios preliminares sugieren, sin embargo, que esta coyuntura es poco
comun.(49)
Neel y Schull, (52) enfocando el problema desde un punto de vista te6-
rico, mencionan la posibilidad de que dos genotipos que esten expuestos
ados medios diferentes originen seis respuestas fenotlpicas. El nume-
ro de respue sta s posibles aumenta a medida que se considere un mayor

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 54/82
nurrie r o de genotipos y medio ambientes. En el cas 0 de diez genotipos
y diez medio ambientes pueden originarse 7,09 X lOl44 respuestas posi-
bles. La formula general para calcular el mrm e r o de respuestas posi-
bles en el caso de m genotipos independientes y n medio ambientes es
(mn)!
mIn![7. l]
En esta formula se consideran los distintos genotipos subyacentes a
una caracterlstica de va r ia cion continua, como estatura, presion arte-
rial 0 inteligencia. Si se consideran sin ultarie arne n te varias caracte-
rlsticas, el nurne ro de respuestas aumenta aun mas y se puede calcular
mediante la formula
((mn) !) km!n!k!
[7.2J
donde my n tienen el rnis rn o significado que en [7. IJ Yk representa el
mirne ro de caracte r fs ticas conside radas.
En 10 que sigue se presentan algunos rno de Ios de herencia de ca r a c -
terlsticas de va r ia ci on continua, basados en el rrie tod o de la pa r tici on
de la varianza fenotlpica, sin entrar en detalles sobre la genet ica for-
mal de sistemas po Ii.geni c os , El lector interesado los encont ra ra en el
libro "Curso Basico de Gen et ica Hurna na!' de Ro thharnrne r y Cruz-Coke.
5 0
EL MODELO CLASICO DE HERENCIA POLIGENICA
La determinacion qenetrioa de una caracterlstica se expresa por el
cociente entre la varianza qenet ica, VG, y la varianza fenottpica total,
VF, 0 sea:
DC _G_ •
VF[7.3J
A su vez la varianza geriet ica s e puede subdividir en una varianza
aditiva, V A , y en una varianza residual, VR , es decir:
V G = V~ + V R [7.4J
Ad ern a s se puede definir la heredabilidad por el cociente de la va-
rianza genetica aditiva y lavarianzafenotlpicatotal; se la de s tgna po r h''
[7.5J
La determinacion genetic a t.arribien se denomina heredabilidad en senti-
do amplio yes siempre igual 0mayor a la heredabilidad ahora definida
y que ta.rrrbi e n se llama heredabilidad en sentido restringido (vea s e
Falconer(53)).
La varianza residual, VR, puede a su vez subdividirse en una varian-
za debida a los efectos de dominancia y una varianza deb ida a la epis-
tasis que puede haber entre los distintos genes que participan en el ca-
racter objeto de estudio. Es decir:

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 55/82
[ 7 . 6 J
La varianza fenotlpica total, VF, aba r ca a derria s de la varianza ge-
netica, V , a la varianza medio ambi.ential., VMA
[ 7 . 7 J
De acuerdo con Cavalli-Sforza y BodITler(28) la varianza rn ed i.o arrr-,
biental puede a su vez subdividirse en una varianza individual, V[ND, que
es la que caracteriza a los individuos pertenecientes a una fa.rn.iIi.a, una
varianza familiar, VF A M , quecaracteriza a dife r ente s farrri Iia.s deunITlis-
ITlOnivel socioe condrrri.c o , una varianza eoc-i.oeeonomiea ; VSE , caracterls-
tica de los distintos niveles s o cioe c on crrri c o s dentro de un grupo etnico-
cultural y, po r fin, una varianza etmi.cocu l : tiural., V E e , que ca racte riza a
los distintos grupos etnicoculturales.
Lo expuesto derrrue s tr-a de rriodo fehaciente que el p r ob Ierna de eva-
luar las contribuciones de la cons ti tuc iSn gene ti ca y el rrredio arribien te
" 1" va r-i.aci Sn fenotirri ca "" cornn1"'j",,irnn Hav 011" t"n"l' nl'''''''nt .. n11"
el m.od elo presentado no representa por el morne nto ma s que unaapro-
xirria ciSn idealizada a la realidad. Es posible, ade ma s , que apa r te de
las cont r ibuci.one s rne di o arnbi errta Ie s y gene tica s , haya factores alea-
torios que de te r-mrnan la apa r-ici on de una caracterlstica. Esto iITlpli-
ca que haya un cierto g rado de 'i.ndete r-rrrini srn o 0" ruido" en el desarro-
llo de los ca r a ct e r es bajo corrs ide r a ci.on , Es evidente que es preciso
ta mb ien rrienc iona r los e rrores de rrie dicion que pueden desviar los es - 5 1
ti m ado r e e de varianza en el caso que sean sis terria ti cos , Hasta el ITlO-
rne nto se ha considerado que si la dete r rrrina ciori gene tica de una carac-
terlstica asciende por e jerrip lo al 60%, el 40% restante de la varianza
se debe a factores m.e dio a rn bi e n ta Ie s , s e n ci l la.rrrerrte por s ub s tr a c cidn ,
Sin erriba r g o, la e s t irria cion separada, utilizando el ITletodo de los con-
juntos fa.rn.iIi a r e s , de las contribuciones g enet ica s y rrie di.oa.rnbierrta Ie s ,
no ha proporcionado los resultados esperados. Al SUITlar las contribu-
ciones geriet ica s y rried i o arnb ie nta Ie s (es ttrna.da s independienteITlente)
a la varianza total de presion sangulnea en una pob la ci Sn de Detroit,
Schull [c orriurri ca c i.on personal) ha obtenido cifras inferiores a l 100%.
Estos resultados podrlan indicar la influencia de los factores antes
rrie nc'i on ad o s y s e repres entan por e.
F'ina Irrie nt e , la i.nt e r ac cIon gene ticovrned io arribi.errta l a que nos
hemos referido va , puede en principio se r incorporada a la varianza
rrie dio ambi enta l en el m.od e Io general que se ha estado considerando.~. -, ~
GME , 1"
[ 7 . S J
Las serias Ii'm i.ta ciories del rn e to do de la pa r tic i.ori de varianza
aconsejan cautela en el uso de e s tirna.do r e s basados en este ITletodo.
Se p r e s enta r a n no obstante varias estrategias rne todo ldg Lca s elabora-
das para disecar algunas de las contribuciones rn.edio arnb ie nta le s y
gene tica s a la variabilidad £enotlpica.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 56/82
ALGUNOS M~TODOS,PARA ESTIMAR LA HEREDABILIDAD Y LA DE-
TERMINACION GENETICA
Los rnetodoe mas corrninrnerrte utilizados son:
1) Estudios con mellizos. Estos aba rcan la compa ra cion de melli-
ZOS monocig6ticos con dicigcticos criados juntos y por separado.
2) Estudios de hijos adoptivos. En este caso el rne todo consiste
en cornpa ra r los hijos adoptivos con sus padres adoptivos con quienes
se supone que comparten el ambiente, y los hijos de los padres adopti-
vos que comparten con sus padres el medio ambiente yparte de la cons-
ttt ucton gerie tica .
3) Estudios familiares. Estos se ba san en la corripa racicn de los
hijos entre sI, y con sus padres y abuelos. Las relaciones que existen
entre hermandades, y entre padres e hijos, se expresan corminrrierrte
por medio de coeficientes de correIacion,
4) Estudios basados en coniuntos familiares. En este rne todo se
destacan las relaciones familia res horizontales, a dife rencia del meto-
do anterior en que se hace hincapi e en las relaciones familiares verti-
cales.
5 2
Es preciso hacer notar que la mayor parte de estos rnet odos han
sido elaborados por geneticistas, de modo que la estrategia ha cons is -
tido en partir de la htpote sis de que una caracter1stica estadetermina-
da por el medio ambiente, y rechazar luego esta h'ipot esis medio am-
bientalista, tomando como base un juicio probabil1stico, en favor de una
hi.pSte si.s gerietica alterna.
Todos estos rne todo s, aparte de los problemas discutidos en rela-
cion con la pa rticion de varianza tienen a lavez ventajas ydesventajas.
Resulta por 10 tanto dif1cil recomendar alguno en especial. El metoda
a emplear depe nde ra en gran parte de la caracter1stica cuya determi-
nacion gene tica se desea estimar. En general, la cornpa racion de me-
llizos rnonoci.goticos y dici.gcticos criados juntos y por separado pare-
ce un rrietodo aceptable, pero por desgracia es diffciI recopilar una
muestra 10 bastante extensa. El rriet odo de los hijos adoptivos resulta
igualmente apropiado; sin embargo en pocos casos se t'iene i.nforrna cicn
sobre los padres biologicos de los hijos adoptivos, y por otra parte,
las agencias encargadas de coordinar las adopciones se esfuerzan pa-
ra que los nifios sean adoptados por familias de condiciones econorrri >
cas y culturales sirnia res a las de sus padres biologicos , 10 que
tiende a reducir las contribuciones medio ambientales a Ia varianza
fenot1pica.
Los estimadores de determinaci6n gerieti ca obtenidos a partir de
correlaciones familiares verticales pueden resultar desvirtuados por
varios motivos, entre ellos los cambios en las condiciones ambientales
a que han estado expuestos padres, hijos 0abuelos. Resulta por el mo-
mento mas conveniente recurrir a otros rn.etodos , como el de conjuntos
familiares, en que se destacan las relaciones familiares horizonta-
les.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 57/82
UTILIZACION DE MELLIZOS MONOCIGOTICOSYDICIGOTICOSPARA
ESTIMAR LA DETERMINACION GENETICA
Los mellizos rnonoctg orfcos , como sunombre indica, resultan de la
fision de un cigoto u ovulo fecundado, mientras los mellizos dici.goticosprovienen de dos cigotos, es decir, de dos ovulos fecundados por dos
espermios diferentes. Como consecuencia de su origen, los mellizos
rnono cigStic oe ecn gene ticarrierrte identicos, 10 que implicar1a que las di-
ferencias que se observan entre ellos se deben a la influencia del medio
ambiente.
Por otra parte, los mellizos d'icigSti co s comparten, en promedio,
la mitad de las caracter1sticas hereditarias, y por 10 tanto, es de es-
pe r a r que no se asemejen entre sf mas que dos hermanos. De acuerdo
con 10 expuesto serfa factible obtener un estirnador de la determinacion
genetica de una caracter1stica restando la varianza entre mellizos mo-nocdgStlco s , VMM , que se supone es medio ambiental, de la varianza en-
tre mellizos di ci got icos , VMD , que hemos supuesto medio ambiental y
gene tica a la vez , y luego expresar el resultado en funcion de la varian-
za entre mellizos dtc igStfco s , es decir
[7.9J
Es importante hacer notar que la cantidad H no equivale exactamen-
te a la heredabilidad, en el sentido arnp lio , definida anteriormente. 5 3
Se ha sostenido que los mellizos rnonocrgSt.icos tienen mas proba-
bilidades de compartir ciertas condiciones medio ambientales por sus
semejanzas rnorfoldgica s , de ca r ac te r , de edad y de sexo, entre otras.
Esto traer1a como consecuencia que la varianza entre rrie Il.izos mono-
cigoticos podlla incluit conllibuciones del xlledio anrbienle y los esti-
madores de determinacion genetica obtenidos aparecer1an abultados.
Para cont ra r res ta r este efecto se han comparado mellizos monocigoti-
cos criados juntos y por separado.
ESTIMACION DE LA DETERMINACION GENETICA A PARTIR DE ES.
TUDIOS CONHIJOS ADOPTIVOS
Una estrategia similar a la corrrparacion de mellizos monocig6ti-
cos criados en el mismo hogar 0 en hogares separados, se ha seguido
contrastando las correlaciones entre padres e hijos adoptivos con las
correlacjones entre padres e bijos b;o16gicos por ejemplo, Neel y
Schull(6:3)proporcionan i.nfor-macicn sobre estas correlaciones para los
resultados de dos tests de inteligencia (vease la Tabla 5).
LA OBTENCION DE ESTIMADORES DE LA DETERMINACION GENE.
TICA A PARTIR DE CORRELACIONES FAMILIARES
Fisher,(64) en su trabajo cIasico sobre la co r r eIa cion entre parien-
tes bajo la s upo sici.on de herencia mendeliana (vea se Antecedentes His-
t6ricos), demostr6 que el coeficiente de cor reIacion entre padres e hijos
es igual a

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 58/82
Tabla 5, Correlaciones (r) Entre los CoeflcIentee de Inteligencia de
Padres e Hijos Adoptivos y Padres e Hijos Bio16gicos
Padres PadresHijos Adoptivos Hijos Bio16gicos Test Referenda
r N r N
Padre 0,07 178 0,45 100 MA Burks (1928)
Madre 0,19 204 0,46 105 MA Burks (1928)
Padre 0, 15 178 0,51 175 Otis Leahy (1935)
Madre 0,20 186 0,51 191 Otis Leahy (1935)
Nota: Reproducido en parte de Neel y Schull (1954). (52)
[7.10J
y que 1a correlaci6n entre he rrnanos es:
tVA + tv o[7.11J
5 4
Cada una de estas correlaciones tienen un valor rriaxirno de 0, 5
cuando la varianza debida a dorrrinancia y la varianza rnedio arribienta l
son iguales a 0. Estas f6rITlulas pe rrndt.en est irna r la heredabilidad de
una caracterlstica basarido s e en correlaciones entre pad r es e hijos yhe r-m ano s .
Si se multip li can los dos terITlinos de la p rirrrer a ecuaci6n por 2 y
arnbos terITlinos de la segunda ecuaci6n por 4, ya continuaci6n se res-
ta la priITlera de Ia segunda, se obtiene
2 [t~A JVA
V F
4 [tVA +~VoJ2VA +Vo
V F V F
2V, +Vo _ VA 2V A Vo-~-+-
VF VF V F V F V F
VoDG
DG = 4rh - 2rph.
EL METODO DE LOS CONJUNTOS FAMILIARES
[7.12J
El rne todo de conjuntos farni Ila res para estirna r la heredabilidad en
el sentido restringido se enfoca ra en re Iacion con un tipo especlfico de
conjunto farrriIia r , pero el procedirrriento puede hacerse extensivo con
facilidad a casos en que el conjunto sea rnas rrurneroso 0 bien las rela-
ciones farrriIi.ar es sean distintas.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 59/82
Cons Idere ee, siguiendo a Schull (coznunicaci6n personal) un conjun-
to fazniliar que consista de un caso lndice (I) (el propositus) obtenido al
aza r de una poblaci6n panmfct ica., uno de sus he rrnano s (ll), un prirno
herznano (P) y un individuo no eznparentado elegido al azar (R) cuya edad
y sexo sean iguales al del caso fndi ce . Las razones de la elecci6n deeste conjunto pueden encontrars e en Schull y colaboradores. (83) Breve-
rne nte , este conjunto proporciona estiznadores de heredabilidad que con-o. ~, _L'
• • " 'U " 'U "v•• "'''".vu''''' l' ,
Cons Ide rese una znuestra al azar de observaciones obtenidas de I, H,
P y R; la regresi6n znUltiple de I en H, P y R esta dada p~r:
X I = a + f 3 ~ H + f 3 P X P + f 3 R X R + e .
Los coeficientes de regresi6n parcial f 3 se obtienen:
[ : ][ W ~
e v H •
W < J [ w , ]= e v H • e v • • e V .R e v I •
e V HR eV P R eV R R e V I
[7.13J
[7.14J
donde e vH., por ejernp lo , siznboliza la covarianza entre elherznanoyel
prirno herznano de los propositi.
Cada terznino del lade derecho de la ecuaci6n [7. 14J puede expre-
sarse en terzninos de la varianza aditiva (VA) l de dozninancia ( V o ) y zne-
dio aznbiental ( V M A ) de la siguiente rriane ra: (:as 5 5
eV H H = e v • • = eV R R = VA + Vo + VM A
e v H • = e v r • '" VA /8
eV H R '" e V ' R '" e V I R '"0
V V I H A f < - o f <±.
Despue s de efectuar algunas operaciones algebraicas sencillas la
ecuaci6n [7. 14J queda reducida a
donde VMes la varianza znedio aznbiental y h
2la heredabilidad.
A su vez la ecuaci6n reducida da origen a
8H= (h2/2 - (h
2)2/64 + y/ 4)/ (1 _ (h2)2/64)
s, '" (h2/8 _ (h2N16 _h
2Y/32)/(1 _ (h
2)2/64),

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 60/82
donde yes meramente una forma ecoriorrrica de escribir V O / ( V A + V o +
+ V M A )·
Al insertar en la segunda el valor de y obtenido de la prime ra ecua-
cion resulta:
o bien (7.1SJ
donde h ; a , S p y SHson los estimadores de los pa r arne t r os en cues tion ,
La varianza del estimador de hZ puede demostrarse (Chakraborty, co-
municaci6n personal) que es igual a
- a -V(hz) = 64(1 - S H ) V (S p) + 64S ~Var S H + lZ8S p(1 - S H ) COV (S HS p) *
(1 - ~H)4 [7.16J
5 6
* La fundamentaci6n de esta f6rmula a.parece ra en breve en otra parte.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 61/82
8FACTORES EVOLUTIVOS ESTOCASTICOS
EL CONCEPTO DE DERIVA GENETICA
En un capi'tulo previo (vease Mutaciones y Selecci6n Natural) se ana-
liz6 el efecto que algunos factores evolutivos direccionales, especlfica-
mente las mutaciones y la selecci6n natural, tienen sobre los cambios
de la frecuencia genica en una poblacion de una generaci6n a Ia siguien-
teo Nos referiremos a continuaci6n al efecto de los factores evolutivos
estocas t'i cos . En 1921, Hagedoorn y Hagedoorn observaron que el nu-mero de individuos destinados a ser los progenitores de una nueva ge-
neraci6n es por 10general rnerior que el nurnero de individuos que for-
man la especie. Concluyeron que algunos genes se pierden al aaa r
como consecuencia de este hecho, originando una reducci6n de la va-
riabilidad gene tica potencial incluso en ausencia de la selecci6n natu-
ral. Estos geneticistas holandeses observaron que:
"El grupo de individuos elegidos al azar pa ra integrar lageneraci6n
parental es en general, pe r o siempre ocasionalmente, menor, en for-
ma sustancial, que el mirner o de individuos de su especie. En cada 5 7
caso en que individuos excepcionales que sonportadores de genes anor-males 0 carecen de genes normales comunes a la mayori'a sean exclui-
dos por a za r de la gene ra cidn parental, la variabilidad total disminuye" .
Los Hagedoorn se lirnitaron a presentar el concepto en forma in-
tuitiva sin recisarlo de manera adecuada. Fisher (56) uien denomin6
el fenorrieno efeato de Haqedooim, yen fecha posterior Wright,(67) que
10denorrrino deriva qenetrica al.eatox-ia, elaboraron las primeras apro-
ximaciones a una soluci6n rrraternat ica del problema. En 1955, Motoo
Kimura publico la soIucion completa para el caso de sistemas multi-
a.Ie Ii.c o s ,
UNPEQUENO EXPERIMENTO QUE ILUSTRA LA DERIVA
Con el objeto de ilustrar el efecto de la de riva genetica aleatoria (0
simplemente deriva genetical haremos un paquefio experimento. Co-
rrriencese considerando un lugar genetico con dos a:elos, .A y BJy para
simp 1lC r e exper'men sin er ,
conjunto de gametos, en vez de una poblaciSn de individuos. Con ante-
rioridad (vease Poblaciones Mendelianas y la Ley de Hardy-Weinberg)
se ha demostrado que la uni6n al aza r de gametos es equivalente a l apa-
reamiento al azar 0 panmixia.
Prosiguiendo se tienen 2 tipos de gametos, el de los que contienen
el gen A y que se rep re senta ra por bolillas rojas y el de los que contie-
nen el gen B y que se r ep r ea enta r a por bolillas negras. Se comenza ra
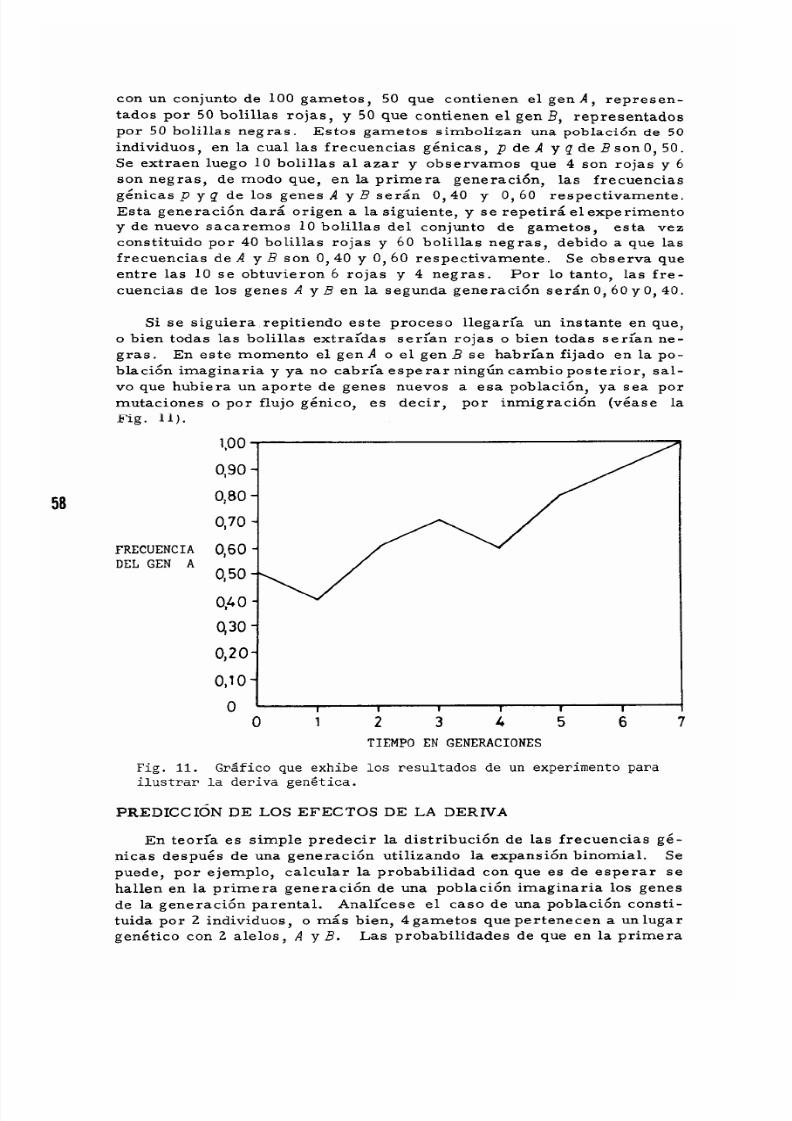
5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 62/82
con un conjunto de 100 gametos, 50 que contienen el gen A , represen-
tados por 50 bolillas rojas, y 50 que contienen el gen B, representadospor 50 bolillas negras. Estos gametos sirriboIfzan una poblaci6n de 50
individuos, en la cual las frecuencias genicas, p de A y q de B son 0,50.
Se extraen luego 10 bolillas al azar y observarnos que 4 son rojas y 6
son negras, de modo que, en la primera generaci6n, las frecuencias
genicas p y q de los genes A y B seran 0,40 y 0,60 respectivarnente.
Esta generaci6n dara origen a la siguiente, y se r-epetira el experimento
y de nuevo sacaremos 10 bolillas del conjunto de gametos, esta vez
constituido por 40 bolillas rojas y 60 bolillas negras, debido a que las
frecuencias de A y B son 0,40 Y0,60 respectivamente.. Se observa que
entre las 10 se obtuvieron 6 rojas y 4 negras. Por 10 tanto, las fre-
cuencias de los genes A y Ben la segunda generaci6n seran0,60y0,40.
Si se siguiera repitiendo este proceso llegarla un instante en que,
o bien todas las bolillas extraldas serlan rojas 0bien todas serlan ne-
gras. En este momenta el gen A 0el gen B se habrlan fijado en la po-
blaci6n imaginaria y ya no cabrla esperar rringiin cambio posterior, sal-
vo que hubiera un aporte de genes nuevos a esa poblaci6n, ya sea por
mutaciones 0por £lujogenico, es decir, por inmigraci6n (vease la
1,00
0,90
5 80,80
0,70FRECUENCIA 0,60DEL GEN A
0,50
0,40
0,30
0,20
0,'0
00 2 3 4 5 6 7
TIEMPO EN GENERACIONES
Fig. 11. Grafico que exhibe los resultados de un experimentopara
ilustrar la deriva genetica.
PREDICCION DE LOS EFECTOS DE LA DERIVA
En teorla es simple predecir la distribuci6n de las frecuencias ge-
nicas despues de una generaci6n utilizando la expansi6n binomial. Se
puede, por ejemplo, calcular la probabilidad con que es de esperar se
hallen en la primera generaci6n de una poblaci6n imaginaria los genes
de la generaci6n parental. Analfcese el caso de una poblaci6n consti-
tuida por 2 individuos, 0mas bien, 4gametos quepertenecen a unlugar
genetico con 2 alelos, A y B . Las probabilidades de que en la primera

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 63/82
generaci6n esten presentes las distintas com.binaciones de genes A 0 B ,
suponiendo que estos genes tienen en la generaci6n parental frecuencias
p y q, se obtiene m.ediante la f6rm.ula
Pr = ~ 'r 'prqn-r(h ; - r)' . [8.1 J
donde r es el nurnero de genes de unm.ism.o tipo (A 0 B ) que es de espe-
rar se hallen presentes y k es el mime ro total de gametos (en el ejem-
plo 4).
Sustituyendo en esta f6rm.ula los valores de h; y r, y suponiendo que
en la generaci6n parental la frecuencia p de A y la q de B sean iguales
a 0,50, se obtiene el valor de 0,0625. La probabilidad de que 3 genes
sean de un tipo y uno del otro es, haciendo las sustituciones correspon-
dientes, de 0,25, Ypor fin, la probabilidad de que se encuentren 2 ge-
nes A y 2 genes B es de 0,375. El calculo de las probabilidades de queesteri presentes los 2 tipos de genes en las generaciones sucesivas se
com.plica gradualm.ente, y en la pra ctica resulta difi'cil obtener resul-
tados correctos por el procedim.iento em.pleado. Sin em.bargo, es per-
fectam.ente posible calcular las distribuciones m.ediante com.putadores
electr6nicos.
Kim.ura, (68) obtuvo la soluci6n rnaterriatica com.pleta para ha cer pre-
dicciones sobre la distribuci6n de frecuencias genicas despues de n ge-
neraciones. Las ecuaciones de Kim.ura son enextrem.o com.plejas y su
presentaci6n cae fuera de los alcances de esta m.onografi'a. La proba- 5 9
bilidad de fijaci6n 0 perdida de un gen increm.enta con el tiem.po, y 5610se acerca al valor rnaxrmo de spue s de un lapso prolongado. Kim.ura
obtuvo la siguiente aproxim.aci6n de la proporci6n de poblaciones enque
se podrfa esperar la fijaci6n de un gen en funci6n del tiem.po, t, m.edi-
do en generaciones y con una frecuencia inicial Po del gen A
[8.2J
donde N es el tarnafio poblacional.
Ewens(60) propone la siguiente f6rm.ula para obtener el tiempo pro-
media de fijaeion, T, en generaciones
T = we-po log. Po + (1-Po)10g.(l-Po)J.[8.3J
La expresi6n entre corchetes es igualaO, 69paravalores de P igual
a 0,50. En este caso, T, el tiem.po prom.edio de fijaci6n, es 2, B N,donde N reoresenta nuevam.ente el tarn.afiode la poblaci6n. Para valo-
res de p m.enores, T dism.inuye. Por ejem.plo, para p igual 0,005, T
es igual a 0,03 N. En reswnen, se puede decir que el tiem.po prom.edio
de fijaci6n de un gen depende de la frecuencia genica inicial y del ta-
rnano de la poblaci6n. Com.oprim.era aproxim.aci6n sup6ngase que el
tiem.po prom.edio de fijaci6n, y sin que haya m.utaci6n, selecci6n naturalni flujo genico, es aproxim.adam.ente de 2N.
LA PROBABILIDAD DE FIJACI6N DE UN GEN MUTANTE
En un capi'tulo previo (vease Mutaci6n y Selecci6n Natural) se ha
considerado el efecto que tienen las m.utaciones sobre los cam.bios de

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 64/82
la frecuencia g erri ca en poblaciones de ta.rriafi o ili:mitado. Es interesan-
te analizar el destino de un :mutante en una poblaci6n de tarna.no finito.
Si, por eje:mplo, se considera unatasa de:mutaci6n de 10-<;y unapobla-
ci6n de 10.000 individuos, cabrla esperar que apareciera uno de estos
genes :mutantes cada 1.500 afio s , Por otra parte, la probabilidad de
que se repita exacta:mente 1a :mis:ma :mutaci6n es re:mota, a pesar de
que ciertos experi:mentos hechos con el bacteri6fago T4 parecen indicar
que, en ciertos lugares de la rnoIec uIa de acido desoxirribonucleico,
las :mutaciones son rna s frecuentes.
La probabilidad de que !ina1mente se produzca la fijaci6n de un gen
:mutante en una poblaci6n de ta.mano reducido depe nd e r-a de varios fac-
tores. Para hacer los ca lc ulo s correspondientes es preciso conocer en
pri:mer lugar el valor selectivo del heterocigoto, porque durante una
parte importante de la vida del gen, Elste pe rrnane ce ra en estado hete-
rocigoto. El destino de un gen :mutante unico fue estudiado pri:mero por
Fisher(56) y rna s adelante por Ki:mura. (59) La probabilidad de fijaci6n Psera funci6n de Po la frecuencia inicial, de s , el coeficiente de selec-
ci6n del heterocigoto y de N, el tarriafio de la poblaci6n. Si se supone
una frecuencia inicial de Po = ~, la probabilidad de fijaci6n sera igual a:2N
P ( f N )1 _e-a•
[S.4J1 _e-4N.
Esta f6r:mula no e s valida sis 0, pue s to que en ese cas o
P ( P o ) s« . [S.5J
6 0
DERIVA GENETICA EN POBLACIONES SUBDIVIDIDAS
Hasta ahora se han hecho predicciones generales sobre la distribu-
ci6n de frecuencias gern ca s esperadas de s pue s de n generaciones en po-
blaciones de tarna ho finito. A continuaci6n se arra.Iiza r a el efecto de la
deriva g erie ti ca en una poblaci6n subdividida.
Corrri enc e s e considerando el caso de una poblaci6n dividida en If
subpoblaciones discretas, cada una con frecuencias genrc a s distintas
para dos alelos A y B, con el rnismo mime ro de individuos y en equilibrio
de Hardy-Weinberg. Se tiene entonces,
Poblaci6n Frecuencia Gem ca Frecuencias Genotlpicas
A A AB BB
1 P l p~ 2P l1h q?
2 Pa p~ 2p->q2 q~
Ifa
2PkQk q ;k p o .
El prornedio p de la frecuencia del gen A en la poblaci6n total es:
p = L : P I
If
[S.6J
y la varianza de la frecuencia gentca en la poblaci6n total es:
2 L : P : . l 1 -2
a = J r ; - P • [S.7]

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 65/82
La frecuencia promedio de heterocigotos AB es:
[8.8J
Utilizando la ecua cion [8.7] se puede reescribir la ecua cion [8.8J de
la siguiente forma:
H(AB):3
2pq (1 - pq ). [8.9J
POI oLIa pal te, la £1ecaencia prollIedio de los hOXIlocigotos sera igual
a
H(AA)
H(BB)
[8. 10]
[8.11J
Se advie rte que en Ia pob la ci Sn total subdividida hay una deficiencia 6 1
de heterocigotos igual a
Esta cantidad es una medida directa de la heterogeneidad entre
las subpoblaclOnes que Integran una poblaclC>n total baJo el efecto de
Ia deriva gen etica , Es interesante notar que esta cantidad se deno-
mina varianza estandarizada 0 de Wahund, y equivale a F, el coeficien-
te de endocruzamiento (vease Endogamia) y a f, el coeficiente de pa-
rentezco si se prescinde del aporte de mutaciones que pueda haber
ocurrido.
La varianza de Wahlund aumenta con el tiempo, y es posible esta-
blecer, cuando el mime ro de generaciones es pequefio con rela ciori a N,
que
[8.l2J
Sewall Wright fue el primero que llarno la at.en cion sobre la posible
importancia evolutiva derivada de la subdivision de una pob la ci on en gru-
pos parcialmente ai slados . Wright sugirio que la subdivision puede
acelerar el proceso evolutivo: (1) favoreciendo la adapta cion a nichos
ecologicos locales, (2) inc rementando la va riabilidad genetf ca al pe r-
mitir el establecimiento de genotipos condistinta ade cua.ci Sn darwinia-
na en diferentes nichos ecoIog icos , y (3) favoreciendo nuevas combina-
ciones favorables de genes.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 66/82
MODELOS DE ESTRUCTURA POBLACIONAL
Se han disefiado varios modelos t.e oricos para predecir la variabi-
lidad gerret'ica esperada en una pob la ciSn subdividida y parcialmente
aislada bajo el efecto de las fluctuaciones alazar de las frecuencias ge-
nicas determinadas por el t.arnafio finito de las subpoblaciones y bajo
el efecto amortiguador del flujo genico existente entre elIas. EI mode-
10mas simple propuesto po r S. Wright en 1943 se denomina modele in-
su"lar, y describe una pob la cion dividida en subpoblaciones, cada una de
tarnano N. En cada gene ra cion cada subpobla cdon recibe una proporci6n
m de sus genes de un acervo c orrnin de genes que se supone es de tama-
no infinito y de frecuencias genicas constantes, mientras que una pro-
porci6n 1 - m de sus genes proviene de la generaci6n anterior de la mis-
ma subpoblaci6n. La varianza genet ic a en las condiciones de este
modelo aumenta debido a la deriva genett ca y es contrarrestada por el
flujo genico hasta que finalmente alcanza un estado de equilibrio en que
se cumple:
[8.13J1 + 4 Nm '
.1
1
pq
es la varianza estandarizada de Wahlund.
6 2 Por ser el modelo insular francamente irrealista, pronto se propusie-
ronotros: por Wright en 1943, 1946yI951;(61-63)
por Ma Iecot en 1945,1950 y 1967;(64,65,66)por Kimura y Weiss en 1964(67)y por Bodmer
y Cavalli-Sforza en 1968.(68) La innovaci6n adoptada en estos modelos
consiste sobre todo en que se considera la distancia que separa a las
subpoblaciones, a diferencia del modeloinsular, en que cada subpobla-
ci6n recibe el mismo aporte de genes que las restantes. Se pueden cla-
sificar los modelos anteriormente mencionados en dos tipos: (1) uno
continuo, en que la densidad de poblaci6n es constante en cada punto del
area habitada, y (2) otro discontinuo en que las subpoblaciones estan
localizadas en los puntos de intersecci6n de un enrejado irnaginario.
De ambos tipos de modelos hay por 10 menos do s versiones, (1) una
unidimensional en que las subpoblaciones estan localizadas en una llnearecta y (2) una versi6n bidimensional en que estan localizadas en una
superficie. El modelo continuo se denomina de "ai.el.amiento por dis-
tanoia", y el modelo discontinuo es designado de "paso de piedra en
piedra", En el modelo continuo la densidad de poblaci6n por area reem-
plaza aN, el tarnafio de poblaci6n de cada subpoblaci6n, y la densidad
tipo de las distancias entre los lugares de nacimiento de padres e hijos,
r eerrrpra.z.a am .. ;:,~ supone que esta cnstrao ucion es no rrna r. J.Jeacuer-
do con Ma Iecot,(95)en el modelo continuo unidimensional el coeficiente
promedio de parentesco entre vecinos es, en equilibrio,
1fo = ,1 + 4aoi2b
[8.14J
donde b representa el coeficiente de retorno al equilibrio, 0 la densi-
dad y a la desviaci6n tipo de las distancias entre los lugares de naci-
miento de padres e hijos.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 67/82
La version unidimensional del modelo dis continuo considera que ca-
da pob la ci Sn coris ta de N individuos, contribuyendo cada s ubpob lac idn
una f'r a cc ion de - t individuos a las dos vecinas. En su version bidimen-
sional esta f r a c cion es de ~. En el modelo dis continuo, N reemplaza a6 y el coeficiente promedio de parentesco entre subpoblaciones e s :
1 + 4Nb7~
Bodmer y Cavalli-Sforza(68) elaboraron un tercer tipo de modelo
que es bastante mas realista que los considerados antes y que se deno-
mina de "las matx-ioes miqracional.ee", En este modelo las subpoblacio-
nes pueden ser de distinto tarnafio e intercambiar individuos en distintas
proporciones. Ad erria s se puede considerar la i.nrni.g r-aci dn de indivi-
duos desde las poblaciones marginales a la pob la ci on subdividida. De
acuerdo con Bodmer y Cavalli-Sforza(68) el equilibrio se establece
. t o [8. l5J
cuando
[8.16J
donde V e representa la varianza de las trans formaciones angulares de
las frecuencias gerii ca s que se pueden obtener a partir de una matriz
e s to ca s tica que resume toda la informacion migratoria. La matriz es-
to ca s tica migratoria permitirla predecir en cada gene r ac idn la varia-
bilidad genetica en una pob la ci on subdividida. Estos va Io r e s esperados,
expresados en frecuencias genicas, pueden corrrpa r a r s e con los valores
observados obtenidos por medio del a na Ii sis de genes marcadores. 6 3
Este rn od e lo , como los derrra s , se b a s a en el supuesto que la diversifi-
cacdSng enetf ca es contrarrestada por el flujo genico entre las pobla-
ciones, mutaciones recurrentes y s e Ie c cion de tipo lineal. Cabe men-
cionar que la estructura migratoria se supone constante a 10 largo del
tiempo.
- ~TAMANO DE POBLACION EFECTIVO
Se ha supuesto hasta ahora que (1) N, el tarnano de la pob la ci.on no
cambia con el tiempo, (2) que las generaciones son discretas y (3) que
los dos sexos contribuyan en igual medida a la gene r a ci cn siguiente.No cabe duda que estos supuestos son poco realistas en el ca s o de po-
blaciones humanas. Resulta, por 10 tanto, esencial darle una mayor
generalidad a los modelos, de modo que no nos veamos obligados a
hacer estas suposiciones. El problema se resuelve al menos enparte,
definiendo un tcanaiio pobl aeional: efec tiiio, N.. El tarna rio poblacional
efectivo es aquel tarnano de p ob Ia ci on que da origen a oscilaciones al
azar de las frecuencias genicas similares a las que originarla el tama-
no de p ob Ia cion N, en el caso que los supuestos arriba enumerados se
cumplieran. En el ca s o que estos no se currip lan , N, el ta rnafio de la po-
b Ia ciori que estima mediante un censo de pob la ci.on sobreestimarla a
Ne, el tarriafio efectivo, de modo que este ultimo sera siempre menorque N.
En el caso que N va rfe de gene ra c ion en gene ra ci on , Ne es igual a:
N. [8. 17J1 1 1N l + N z + ... + N k

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 68/82
que corresponde a la media armoni ca de los tarriafios de pob laci.Sn en
distintos instantes.
Un caso especial de va rIa ciSn del tarnano de pobla cion es el Hama-
do efeeto de fundador 0ef'eo to de cuel.Lo de botiel.La, En esta si.tuacionun grupo muy escaso de individuos da origen a una nueva poblacion .
Durante la mayor parte de su evo l.ucion el hombre vi.vio en grupos rela-
tivamente pequenos , cuya existencia dependio primordialmente de la
caza y de la r-ecoIecc.ion. Este tipo de economla impone Hrrrit e s al ta-
rriafio de las comunidades humanas. Al exceder cierto umbral rrurrreri>
co, es posible que una 0 dos familias emigren y funden una nueva
comunidad. Los descendientes de estas familias pueden presentar
frecuencias gerii.cas diferentes de las de la poblacion que les dio origen,
puesto que los emigrantes no seran portadores de ciertos genes pre-
sentes en la gene ra cion parental.
El tarnano efectivo de la poblaci6n, en el caso de que haya genera-
ciones sobrepuestas, se calcula de acuerdo con Nei e Imaizumi(69)
[8.18j
en donde t es la media de la edad de reproduccion (en ano s ) y N " , es el
niirne ro de individuos que alcanzan la edad reproductiva en cada afio .
Por ultimo, en caso que los sexos contribuyan en diferente proporcion
a la gene ra cicn siguiente
6 4 4MF
N. = ---N+F [8.19J
Sin duda es probable que las tres situaciones analizadas, es decir,
fluctuaciones en el tarn ano de pobla ciorr, generaciones superpuestas y~. c..,
'-v".~~vu'-~v"c" uc"1-'~v1-'v~'- <=u. u ,
tanea.
En este caso es conveniente calcular tres ta.rna.fios de pob l.acion
efectivos, N., a partir de las formulas dadas, y luego halIar la media
de estos valores.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 69/82
9ESTIMACION DE FRECUENCIAS OiNICAS
ESTADISTICA Y GENETICA DE POBLACIONES
El desarrollo de la Genetica de Poblaciones Hum.anas ha estado {n-
tim.am.ente asociado al desarrollo de la esta dfat.l ca desde principios de
siglo. Prom.inentes geneticistas de poblaciones, tales com.o Galton,
Pearson, Fisher, Haldane y Wright, entre otros, contribuyeron deci-
didam.ente a echar los cim.ientos de la teorIa esta.dfs tica , A Galton,
por ejem.plo, se debe el concepto de regresi6n al prom.edio, y Pearson
invent6 el chi-cuadrado. Las contribuciones de Fisher a la esta.df'sti ;
ca son, por otra parte, num.erosas y am.pliam.ente conocidas y Haldane
en 1941, al encontrarse trabajando en problem.as de ligam.ento al cro-
m.osom.a X, us6 por prim.era vez en bi ologfa una ecuaci6n de grado 22.
Lo anterior se explica debido a que las rna tematf ca s y la estadi's cica
son fundam.entales para el estudio de la gene tica de poblaciones hum.a-
nas , ya que estas no pueden ser objeto de m.anipulaci6n por parte del
investigador. Norm.alm.ente este se ve en el caso de analizar datos
de experim.entos "naturales" a diferencia del geneticista experim.ental,
que esta en condiciones de planear experim.entos controlados. 6 5
Es fundam.ental para resolver esta dificultad reducir un conjunto
con m.ucha frecuencia ininteligible de datos a unos pocos nrirne ros que
resum.an los hechos esenciales. En otras palabras, es im.portante ob-
tener de un conju nto en form.aci6n que corresponde a una mueetira, una
cantidad que se denom.ina eetadie tico , Cuando uno de estos se utiliza
para estimar el valor de una ca racte r-Fstlca de la poblaci6n pasa a de-
nom.inarse un estimador. El valor real de la caracter-r'stica de la po-
blaci6n se denom.ina pax-dmet»o, Aderna s de la estim.aci6n de pa rarne -
tros, el geneticista de poblaciones se ve ante un segundo problem.a que
consiste en determ.inar la conform.idad de un conjunto de observaciones
con una hip6tesis genetica, es decir, poner a prueba hip6tesis ge-
neticas.
Entre los pa r-arriet ros que con rna s frecuencia esta interesado en
estim.ar el geneticista de poblaciones se encuentran, de acuerdo con
Schull (com.unicaci6n personal):
1) la estim.aci6n de frecuencias genicas;
2) la estim.aci6n de la proporci6n de individuos que llevan un rasgo
a partir de m.atrim.onios de un tipo especial, y
3) la estim.aci6n de las frecuencias de recom.binaci6n de genes li-
gados.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 70/82
En 10 que sigue se cent ra ra la atenci6n en Ia estimaci6n de fre-
cuencias genicas, por ser el pa rarnet r o esencial, recomendando a la vez
al lector interesado en adquirir mas informaci6n revisar el capttul.o
13 del texto de Neel y Schull (1954).
Si se esta interesado en estimar la frecuencia de un gen 0 un grupo
de genes e ape cffi co s , se debe disponer de un modele que describa el
comportamiento del gen 0 genes, de los genotipos y de los £enotipos en
la pobl a cidn que interesa y de la cual se ha obtenido una muestra. El
rriodeIo cornrinrnente emp1eado parte del supuesto de que genotipos y £e-
notipos se distribuyen de acuerdo con el teorema de Hardy-Weinberg
(vea se Poblacione s Mendelianas y Ia Ley de Hardy-Weinberg).
PROPIEDADES DE UN ESTIMADOR
Se admite que un buen estimador debe poseer al menos las prime-r a s tres de las siguientes ca r-acte r Istica s ;
1) Consistencia. Segrin esta propiedad la probabilidad de uri esH-
mador debe converger hacia el pa rarnet r o a medida que el ta.rna.no de
la muestra aumenta.
2) Imparcialidad. El valor promedio de los estimadores derivados
de un universo infinito de mue at ra s debe r a ser identico al pa r-arnet ro ,
Es decir, e1 valor esperado de un estimador debe ser igual al pa r arne-.
t r o ,6 6
3) Eficiencia. El estimador debe tener una varianza tan pequefia al
menos como la de otro estimador consistente.
4) SU£iciencia. Un estimador 0 una combiriac idn de estimadores es
suficiente si proporciona, en cada caso dado, toda la infoIIllaci6n que
es posible obtener de una muestra de observaciones.
Por 10 cormin existen varios estimadores consistentes e imparcia-
les para una determinada attua cidn: sin embargo s610algunos se ran efi-
cientes y acaso ninguno sea sU£iciente. Hasta el momento hemos enu-
merado las propiedades que se desea r fa que tuv ie r a un estimado r , peronos encontramos aun el problema de como obtenerlo.
ESTIMACION DE FRECUENCIAS GlfNICAS DE LOCI DIALELICOS CO-
DOMINANTES
Entre los va r io s metodos disponib1es, e1 mas utH e s eillamado de
maximaverosimiUtud. Para ilustrarlo supdnga se que hay presentes 2
fo r rna s alelicas, AyE, en una poblac idn de gametos, correspondientes
a un Locue genetico. Supdnga se , ad erna s , que estos 2 alelos estan en
una pr opo r ci.dn de 1:3; sin embargo, no se sabe cual de los 2 es mas
frecuente. La frecuencia del gen A (0 del gen E ) podr Ia ser 0,25 0 0,75.
Pr ocSda ee a estimar la frecuencia del gen A en Ia pob.lac idn (el para-
metro) a partir de una muestra. Elfjanse 3 gametos al azar y sup6nga-
se que ninguno es portador del gen. La probabilidad de que en una
muestra de 3 gametos ninguno sea portador del gen A, dado que La

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 71/82
frecuencia del gen A es 0,25, se puede calcular utilizando la expansi6n
binomial (vease Fa cto r es EvoluHvosEstocasticos)yes igual a 0,42188.
Po r otra parte, la probabilidad de que ningrin gameto sea portador del
gen A en una muestra de 3, suponiendo que La frecuencia del gen A sea
0,75, es igual a 0,01563, es de cir , bastante menor que Ia primera.
Comparando las dos probabilidades, se concluye que la frecuencia del
gen A en Ia poblaci6n debe ser igual a 0,25 Y» por 10 tanto, la frecuencia
del gen B igual a 0,75. Los estimadores de frecuencias genicas obte-
nidas son esHmadores de maxima verosimilitud. En general, el meto-
do consiste en elegir a.quel estimador que maximiza r ir.»), donde r ese1 ndrne ro de sucesos iavorables y pes el pa r arnet r o que se desea es-
timar. En el capitulo 3 se propuso estimar la frecuencia de a le lo s co-
dominantes contando lo s directamente en la muestra, es decir siA y B
son 2 alelos codominantes,
p
q = (nAB + 2 n a a ) / 2 N [ 9 . 2 J
donde p y q son las £recuencias de los genes A y B , nAA y nAB son las
frecuencias de los genotipos A A y A B , y I v ' es el talnano de la mues-
t r a , Es posible derno atr a r que este es un estimador de maxima vero-
similitud. La va r-i.ancia de p sera:
V(p) = pq/2N. [ 9 . 3 J
ESTIMACION DE FRECUENCIAS GENICAS DE LOC I DIALELICOS DO. 6 1
MINANTES
Hasta aho ra hemos con sider ado sistemas dialelicos sin dominancia;
sin embargo sup6ngase un lugar genetico con 2 alelos A v «, donde A
es dominante sobre a y los genotipos AA y Aa corresponden al mismo
fenotipo. En una muestra de N individuos, n , , _ corresponden al £enotipo
A - (pueden ser genotfpi.ca rnente AA oAa ), y n .. corresponden a1 fenotipo
a a . Los estimadores de maxima verosimi.litud (MV) de las frecuencias
p de A y q de a se ran:
p = 1 - q Y q = . /n.a! [ 9 . 4 J
La varianza del estimador sera:
V(q) = (1 -q'V4N. [9. 5J
ESTIMACI6N DE FRECUENCIAS GENICAS DE LOC I CODOMINANTES
LIGADOS AL SEXO
Otro ca so es el de genes codominantes ligados al sexo. Sup6ngase
que una muestra de N individuos esta compuesta de N H hombres y N Mmujeres. De las N M mujeres, nAA tienen el fenotipo A A , nAB el fenotipo
AB y naB e1 fenotipo BB . De los N H hombres, nAY tienen el fenotipo AY y
nay el fenotipo BY . Los estimadores MV de las frecuencias de los genes
A yB, P y q res pe ctivamente , son:

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 72/82
p [9.6J
q [9.7J
y
V{p) [9.8 J
, ,
ESTIMACIONDEFRECUENCIASGENICAS DE LOCI DOMINANTES LI-
GADOS AL SEXO
Ahora se c on sIde r a r a la situaci6n de genes dominantes ligados a.I
sexo. En una muestra de N individuos, N M mujeres y N H hombres, nA -
mujere s tienen el fenotipo A - y naa tienen e1 fenotipo aa . Entre 10s hom-
bres nA Y tienen el fenotipo AY y n;.y e1 fenotipo aY . El estimador MV de
las frecuencias de A v «, respectivamente p y q, sera:
p = l-q
y
q
y
[9.10J
ESTIMACI6N DEFRECUENCIAS GENICAS DE LOCI MULTIALELICOS
6 8Finalmente c on s i.de r e se el ca s o de un locus gene ti.c o con 3 alelos
entre los cuales hay relaciones de dominancia. Un buen ejemplo es
el sistema de antlgenos eritrocitarios ABO. Deslgnense a los 3 alelos
por I\ jB e IO. r e jB son dominantes sobre ZOy codominantes entre
ellos. Deslgnense adernas las frecuencias relativas (proporciones)
con que se observan los 4 fenotipos posibles por A, B, 0 Y AB Y las
frecuencias de los 3 alelos por P, q yr. Estimadores ineficientes de
p, q y r senan:
r [9.11 J
q'
l-/(B+O)
1 -I(A+O).
[9.l2J'
[9.13 J
Se pueden obtener estimadores eficientes, equivalentes al estirna-
dor MV, haciendo la siguiente correcci6n:
D = 1- ,_ r',
Estirnadores eficientes de P, q y r, que se representan por P'~, q»
y r* respectivamente, se obtienen:
i= p' (l+D/2) [9.14J
q* = q' (1 + D/2) [9.1SJ
r ' : ' (r' +D/2)(1 +D/2). [9.16J

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 73/82
BIBLIOGRAFIA
(I) KIMURA, M. y OHTA, T. Theoretical Aspects of Population
Genetics, Princeton, Princeton, N. J., 219 pag s , (1971).
(2) DOBZHANSKY, T. Genetics and the Origin of Species, Columbia
Univ. Press, Nueva York, N. Y., 364 pag s , (1951).
(3) LEWONTIN, R. The Genetics Basis of Evolutionary Change,
Columbia Univ. Press, Nueva York, N. Y., 346 pag s , (1974).
(4) SANTALO, L. A. Probabilidad eInferencia Es ta.dfat ic a , Monogra-
fra no. 11 de 1a Serie de Mat ernat.ica , Depar-tarrierrto de Asuntos
Cientr£kos, Or gallizad6n de los Estados Americanos, vvashington,
D. C., 133 pag s. (1970).
(5) KARDINER, A. y PREBLE, E. They Studied Man, Mentor Books,
Nueva York, N. Y., 255 pag s, (1961).
(6) LEWONTIN, R. The Concept of Evolution. En: International 6 9
Encyclopedia of Social Science, D. L. Sills (editor), MacMillanFree Press, Nueva York, N. Y., vol. 5, 202 pag s , (1968).
(7) MALTHUS, T. A Sununary View of the Principle of Population
(1830). En: Three Essays on Population, Mentor Books, Nueva
York, N. Y.
(8) MAYR, E. Animal Species and Evolution, Harvard Univ. Press,
Carnb ridge , Mass., 797 pag s , (1963).
(9) MENDEL, G. "Versuche tiber Pflanzenhybriden", Verh. Natur-
forsch., Brunn, 4 (1865), (Aber.), 3-47 (1866).
(10) FOCKE, W. O. Die P1anzenmischlinge, ein Beitrag zur Biologie
der Gerwachse, Gebr. Borntraeger, Be r Hn , 569 pag s, (1881).
(11) FISHER, R.A. Ann. Sci.,l, 115(1936).
(12) WRIGHT, S. Mendel's Ratios. En: The Origin of Genetics, Stern
y Sherwood (editores), Freeman, San Francisco, Calif., 179
pag s, (1966).
(13) CASTLE, W. E. Proe . Amer. Acad. Arts s-c., 35, 233 (1903).
(14) JENNINGS, H.S. Genetics, 1,53 (1916).
(15) JENNINGS, H.S. Genetics, Z , 97 (1917).

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 74/82
(16) ROBBINS, R.B. Genetics, Z , 489 (1971).
(17) ROBBINS, R.B. Genetics, 3 , 73 (1918).
(18) ROBBINS, R.B. Genetics, 3 , 390 (1918).
(19) DUNN, L. C. Am. J. Hwn. Genet. , 14 (1), 1 (1962).
(20) MULLER, H. J. Am. J. Hum, Geneti, , Z(2), III (1950),
(21) DOBZHANSKY, T. cotd. Spring Harbor Symp. Quant. Biol., ZO,
1 (1955).
(22) KING, J. L. Ann. Hum, Genet . , 38, 507 (1975).
(23) SHAW, C. R. Science, 149,936 (1965).
(24) LEWONTIN, R. C. y HUBBY, J. L. Genetics, 54, 595 (1966).
(25) HARRIS, H. Proc . Roy. Soc. London, ser, B., 164, 289 (1966).
(26) WEINBERG, W. Z. Indukt. Abstamm. vererbunqel.. , 1, 377, 440;
Z , 276 (1909).
7 0 (27) HALDANE, J.B.S. Proo . CambridgePhilos. Soc.,Z3,363(1926).
(28) CAVALLI-SFORZA, L. L. y BODMER, W. F. The Genetics of
Hum.an Populations, Freem.an, San Francisco, Calif., 965 pags.
(1971 ).
(29) BODMER, W. F. Proo . Nat. Acad. Sci. USA., 59 (3), 690 (1968).
(30) FISHER, R. A., FORD, E. B. Y HUXLEY, J. Nature, 144, 750
(1939).
(31) ALLISON, A. C. Trans. Roy. Soc. Trap. Med. Hyg., 48, 312(1954).
(32) SVED, J. A., REED, T. E. y BODMER, W. F. Genetics, 55, 469
(1967).
n'>. \ UTl\Tr! TTl:, 1;1; 4A'>.I)Qh7\
(34) MILKMAN, R. D. Genetics, 55, 493 (1967).
(35) IGLESIAS, R. y COVARRUBIAS, E. Rev. Med. Chile, 90, 866
(1962).
(36) POST, R.H. Euqen; Q., 9,131(1962).
(37) POST, R. H. Humanqenetrilc, 13, 253 (1971).

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 75/82
(38) NEEL, J.V. y POST, R. H. Eugen. Q., 10, 33 (1963).
(39) CRUZ-COKE, R. Color Blindness: An Evolutionary Approach,
Thomas, Springfield, Il1., 156 pag s , (1970).
(40) CHILDS, B. y DER KALOUSTIAN, V. NeuJEngl. J. Med., 279,
(1968).
(41) WRIGHT, S. Am. sae., 56, 330 (1922).
(42) MALECOT, G. Les Mathematiques de L'Heredite, Masson, Pa r fs
(1948).
(43) KUDO, A. Am. J. Hum. Gene t ,, 14, 426 (1962).
(44) MORTON, N. E. , CROW, J. F. y MULLER, H. J. Proc , Nat. Acad.Sci. USA., 42, 855 (1956).
(45) FREIRE-MAlA, N. Eugen. Q., IS, 22 (1968).
7 1
(46) HOGBEN, L. Nature and Nurture, Allen and Unwin, Lond r es , 143
pag s. (1933).
(47) HALDANE, J. B.S. Brkenntini:e , 6, 346 (1936).
(48) HALDANE, J. B. S. Ann. Eugen. (Londres), 13, 197 (1940).
(49) LEWONTIN, R. Am. J. Human Genet • , 26, 400 (1974).
(50) DOBZHANSKY, T. y SPASSKY, B. Genetics, 29, 270 (1944).
(51) CLAUSEN, J., BECK, D. D. y HEISEY, W. M. Carnegie Inst.
Washington PuN., 520, 1 (1940).
(52) NEIL, J. V. YSCHULL, W. Human Heredity, Chicago Univ. Press,
Chicago, Ill., 361 pags , (1954).
(53) FALCONER, D. S. Introduction to Quantitative Genetics, Oliverand Boyd, Edinburgh, 365 pag s. (1960).
(54) FISHER, R. A. Trans. Roy. Soc. Edinburgh, 52, 399 (1918).
(55) SCHULL, W. J., HARBURG, E., ERFURT, J. C., SCHORK, M.A.
v RICE R. J. Chronic Dis. 23 83 (1970).
(56) FISHER, R.A. Proc. Roy. Soc. Edinburgh, 42, 321 (1922).
(57) WRIGHT, S. Genetics, 16, 97 (1931).
(58) KIMURA, M. Evolution, 9, 419 (1955).
(59) KIMURA, M. Genetics, 47, 713 (1962).

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 76/82
(60) EWENS, W. J. PopulationGenetics, Methuen, Londres, 147 pag s.
(1969).
(61) WRIGHT, S. Genetics, 28, 114 (1943).
(62) WRIGHT, S. Genetics, 31, 39 (1946).
(63) WRIGHT, S. Ann. Eugen. (Londres), 15, 322 (1951).
(64) MALECOT, G.
(65) MALECOT, G.
(66) MALECOT, G.
4, 317 (1967).
Compt. Rend. Acad. Sci. rr., 221, 340 (1945).
Ann. Univ. Lyon, Sci. Sect. A, 13, 37 (1950).
Proc. Fifth Berkeley Symp. Math. Statist. Frob.
(67) KIMURA, M. y WEISS, G.H. Genetics, 49, 561 (1964).
(68) BODMER, W. F. Y CAVALLI-SFORZA, L. L. Genetics, 59, 565
(69) NEI, M. e IMAIZUMI, Y. Heredity, 21, 183 (1966).
BIBLIOGRAFLA ADICIONAL RECOMENDADA
7 2Se r ec orrri enda a los lectores que, qui z as motivados por esta breve
rnon og r affa., deseen profundizar en eI terna de la gen et ica de poblacio-
nes hurnana s consultar los siguientes textos:
NEEL, J. V. Y SCHULL, W. J. Hurnan Heredity, Univ. of Chicago
Press, Chicago, Ill., 361 pag s, (1954).
LI, C. C. Population Genetics, Univ. of Chicago Press, Chicago, Ill.,
366 pag s , (1955).
CAVALLI-SFORZA, L. L. Y BODMER, W. F. The Genetics of Hurnan
Populations, F'r e ernan , San F'r anc isc o , Calif., 965 pag s. (1971).
ALLENDE, J. E. Bi o sfnte s is de Pr otefaa s y el C6digo Gen et ico , Mo-
nog r affa no. 10 de la Serie de Btol.ogfa , Depa r tarrrento de Asuntos
Cientr£icos, Organizaci6n de los Estados Arne r-ican os , Washington,
D. C., 132 pag s, (1972).
REISSIG, J. L. La Gerie ti c a y la Revoluci6n en las Ciencias Bio16gicas.
Mcn og r affa no. 1 de la Serie de Bf o logfa , Depa.rt arnento de Asuntos
Cientr£icos, Organizaci6n de los Estados Arne r Icanos , Washington,
D. C., 54 pag s , (1975).
STERN, C. Principle of Human Genetics, F'r e errian , San Francisco,
caur. , 891 pag s , (1973).
ROTHHAMMER, F. y CRUZ-COKE, R. Curso Basico de Genet lc a Hu-
rnana , Ediciones de la Universidad de Chile, Santiago, Chile (1977).

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 77/82
El siguiente libro da informaci6n sobre trabajos realizados en po-
blaciones humanas latinoamericanas:
Salzano, F. M. (Editor). The Ongoing Evolution of Latin American Popu-
lations, Charles C. Thomas, Springfield, Ill., 717 pags. (1971).
En las revistas cientrficas que se enumeran a continuaci6n aparecen
arhculos relaclonados con la genehca de poblaclones humanas.
American Journal of Human Genetics
Annals of Human Biology
Annals of Human Genetics
American Journal of Physical Anthropology
Humangenetik
Human Biology
Human HereditySocial Biology
7 3

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 78/82
AORADEC IMIENTOS
Nu:rnerosos colegas e instituciones colaboraron en la preparaci6n
de la presente rnonogr affa. Danko Brncic, Raul Fernandez y Carlos
Valenzuela leyeron y criticaron la versi6n co:rnpleta del :rnanuscrito ori-
7 4 ginal. Alvaro Rodrfguez revis6 varios caprtulos. Partei:rnportante de
los conceptos expresados se originaron en conversaciones sostenidas
con colegas de la Universidad de Chile, de la Universidad de Michigan,de la Universidad de Texas y de la UniversidadAustralde Chile. A to-
dos debo expresar :rni gratitud. Adema s , deseo agradecer a Xi:rnena
Aguilar y a Mary Lopez la labor de dacti log r affa yaM. Angel Hinojosa
la preparaci6n de las ilustraciones.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 79/82
Publicadas
COLECCION DE MONOGRAFlAS CIENTIFICAS
Serie de matematica
N° 1. La Re vo luci on en las Ma te rnat ica s Escolares, por el Con-
sejo Nacional de Maestros de Maternatf ca s de los Estados
Unidos de Arrie r i.ca .
N° 2. Espacios Vectoriales y Geometrla Ana lfttc a , por Luis A.
Santa16.
Estructuras Algebraicas I, por Enzo R. Gentile.
Historia de las Ideas Modernas en la Ma terna tica , por Jose
B,abini.
Alg eb r a Lineal, por Orlando E. VillalTIayor.
Algebra Linear e Geometria Euclidiana, por Alexandre
Augusto Mal tills Rodrigues.
N° 3.
N° 4.
N° 5.
N° 6.
N° 7.
N° 8.
N° 9.
N° 10.
N° 11.
N° 12.
N° 13.
N° 14.
N° 15
N° 16.
N° 17.
Serie de flsica
N° 1.
N° 2.
N° 3.
N° 4.
N° 5.
N° 6.
N° 7.
N° 8.
N° 9.
N° 10.
N° 11.
N° 12.
El Concepto de Nurne r o , por Cesar A. Trejo.
Funciones de Variable Compleja, por JOse 1. Nieto.
Introducci6n a la Topologla General, por Juan Horvath.
F'unc oe s Reais, por Djairo G. de Figueiredo.
Probabilidad e Inferencia Est<l;_dlstiea, por Luis A. Santa16.
Estructuras Algebraicas II (Algebra Lineal), por Enzo R.
Gentile.
La Revoluci6n en las Ma.ternat ica s Escolares (Segunda Fa- 7 5
se), por Howard F. Fehr, John Carrip y Howard Kellogg.
Estructuras Algebraicas III (Grupos Finitos), por Horacio H.
O'Brien.
Tntroducciona la Teorla de Grafo~, por Fausto A ToranzDs
Estructuras Algebraicas IV (Algebra Multilineal), por
Artibano Micali y Orlando E. V'i.Ll a.rnay or .
Irit r od uc a o a Analise Funcional: Es pa.c os de Banach e Ca c ulo
Diferencial, por Leopoldo Nachbin.
Coneepto Moderno del Nuc le o , por D. Allan Bromley.
Panorama de la Astronomia Moderna, por Felix Cernuschi
y Sayd Codina.
La Estructura Electr6nica de los S6lidos, po r Leopoldo M.
Falicov.
F'Is ica de PartIculas, por Igor Saavedra.
ExperilTIento, Razonamiento y Creaci6n en F'Is ica , por Felix
Cernuschi.
Semieonduetores, por George Bemski.
Aceleradores de Partlculas, por Fernando Alba Andrade.
F'fs ica Cuan tica , po r Onofre Rojo y Harold V. Mcintosh.
La Radiaci6n C6smica, por Gast6n R. Mejia y Carlos
Aguirre.
As t r offs ica , por Carlos Jasehek y Mercedes C. de Jaschek.
Ondas, por Os ca r J. Bressan y Enrique Gaviola.
EI Laser, por Mario Garavaglia.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 80/82
Serie de qufmica
N° 1.
N° 2.
N° 3 .N° 4.
N° 5.
N° 6.
N° 7.
N° 8.
N° 9.
N° 10.
N° II .
N° 12.
N° 13.
N" 14.
N° IS .
N° 16.
N° 17.
7 6
Cinetica Oufmi ca Elemental, por Harold Behrens Le Bas.
Bfo ene rg eti.ca , por Isaias Raw y Walter Colli.
Ma c r orn o Ie cu Ia s , por Alejandro Paladini y Mois e s Burachik.Mecanismo de las Reacciones Or ganfc a s , por Jorge A.
Brieux.
Elementos Encadenados, por Jacobo Gorne z eLa r-a.
Ens eria.nz a de la QUlmica Experimental, por Francisco Giral.
Fotoqui"mica de Gases, por Ralf-Dieter Penzhorn.
Irit r oduc ci on a laGeoqui"mica, por Felix Gonz.a le z =Bono r+no.
Resonancia Ma g ne tica Nuclear de Hi d r og eno , por Pedro
Joseph-Nathan.
Cromatografi"a Lfquida de Alta Presion, por Harold M.
McNair y Benjami"n Esquivel H.
Actividad Optica, Dispersion Rotatoria Optica y Dicroi"smoCircular en Qui"mica Or ga ni ca , por Pierre Crabbe.
Espectroscopia Infrarroja, por Jesus Morcillo Rubio.
Polarografi"a, por Alejandro J. Arvi"a y Jorge A. Bolzan.
Paramagnehsmo Electronlco, por Juan A. McMlllan.
Irit r od u.cc i.Sn a la Estereoqui"mica, por Juan A. Garbarino.
Cromatografi"a en Papel y en Cap a Delgada, por Xorge
A. Domi"nguez.
Int r od uc ci dn a la Espectrometri"a de Masa de Sustancias
Or gan ica s , por Otto R. Gottlieb y Raimundo Braz Filho.
Serie de biologfa
N° 1.
N° 2.
N° 3,
N° 4.
N° 5.
N° 6.
N° 7.
N° 8.
N° 9.
:w 10.
N° II .
N° 12.
N° 13.
N° 14.
N° 15.
La Gerie tt ca y la Revo Iuci on en las Ciencias B'io Icg Ica s ,
por Jose Luis Reissig.
Bases Eco lSgica.s de la Exp lota ci on Agropecuaria en la
America Latina, por Guillermo Mann F.
La Taxonomi"a y la Re vo l.uc icn en las Ciencias Bdo Iogica s ,
por Eli"as R. de la Sota.
Principos Basicos para la Ensenanza de la Biologi"a, por
Oswaldo Frota-Pessoa.
A Vida da CHula, po r Renato Basile.
Microorganismos, por J.M. Gutierrez-Vazquez.
Principios Generales de Microbiologfa, por Norberto J.
Palleroni.
Los Virus, por Enriqueta Pfz a r r o-Bua r ez y Gamba.
Irrt r od uc ci on a la Ecologi"a del Bentos Marino, por Manuel
Vegas Velez.
Bioslntesis de Plotelnas y el Codigo Gem::tico, pOI Jorge
E. Allende.
Fundamentos de Inmunologi"a e Inmunoqui"mica, por Felix
Cordoba Alva y Sergio Estrada-Parra.
Ba cte riSfag os , por Romilio Espejo T.
Biogeografla de America Latina, por Angel L. Cabrera y
Abraham Willink.
Re Ia ci on Hue sped c Pa r a sio . Mecanismo de Patogenicidad
de los Microorganismos, po r Manuel Rodri"guez Leiva.
Ge.netic a de Poblaciones Hurnanas, por Francisco
Rothharnmer.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 81/82
En preparacion
Serie de mat emat ica
Estructuras Algebraicas V (Teorla de Cuerpos), por Darfo J. Picco.
Estructuras Algebraicas VI (Estructuras de Algebras), por Artibano
Micali.
Btomaternattca , por Alejandro Engel.
Iritroduccion al Analrsf s , por Manuel Balanzat.
Irit roducciSn a la Integral de Lebesgue en la Recta, por Juan Antonio
Gatica.
Ent roducciSn a los Espacios de Hilbert, po r Jose 1. Nieto.
Irrtr oduccion a la Cornputa cl.c n, por Jaime Michelow.
Programaci6n Lineal, por Fernando 1. Garagorry.
Serie de flsica
Oceanografla F'fsica , por Luis E. Herrera
Teorla de Fluidos en Equilibrio, por Antonio E. Rodrfguez y Roberto
E. Caligaris.
.Aplicacao da Teoria de Grupos na Espectroscopia Raman e do Infra-
Vermelho, por Jorge Humberto Nicola y Anildo Bristoti.
Teorla Estadlstica de la Materia, por Antonio E. Rodr Iguez y
Roberto E. Caligaris.
Geoflsica, por Alvaro F. Espinosa.
Serie de qufmica7 7
Fotometrla de Llama por Errrision, por Juan Ramfrez Munoz.
Fotometrla de Llama po r Absor cion Atormca , por Juan Rarn fr e z
Munoz.
Fluorescencia Atomica, por Juan Rarrrfrez Munoz.
T'er'modi.riarrrica QUlmica, por Jaime Cases.
Cromatograffa de Gases, por Harold M. McNair.
Slntesis Oz-ganfca, por Eduardo Sanchez.
Ca.tjiIisis Hornogerrea, por Eduardo Humeres A.
Cata Iisis Hete rogenea, por Sergio Droguett.
Fuerzas Intermoleculares, por Mateo Dlaz Pena.
Introducci6n a la Electroqulmica, por Dionisio Posadas.
Corrosion, por Jose Rodolfo Galvele.
Coloides, por Javier Garfias.
QUlmica de SueIos , por Elemer u. Bornemisza.
Irit roduccron a la Metalurgia Ffs ica , por Joaquln Herriaez MarIn,
Procesos Microbianos Aerobios de Importancia Industrial, por
Carlos Casas-Campillo.
Introducci6n ala EcoIisiologla Vegetal, por Ernesto Medina.
Etologla: EI Estudio del Comportamiento Animal, por Raul Vaz -
Ferreira.
Oi.togerie'tica Basica y Biologla de los Cromosomas, por H. Cardoso.
C'itogeriet'ica Ultraestructural y la Biologla Molecular de los Cro-
mosomas, por R. Wettstein y J. Roberto Sotelo.

5/14/2018 Genetica de Poblaciones Humanas - slidepdf.com
http://slidepdf.com/reader/full/genetica-de-poblaciones-humanas 82/82
Anali sis de Sistemas en Ecologla, por Gilberto C. Gallopln.
Ecologla de Poblaciones Animales, por Jorge E. Rabinovich.
Sistemas Eco16gicos y el Hombre, por Ariel E. Lugo y Greg Morris.
Biologla Celular de la Transformaci6n Maligna, por Manuel Rieber.
Comportamiento y Aprendizaje, por Hector Maldonado y Josue A.
Nunez.
Principios Basicos de laContracci6nMuscular, por
Transporte a Tr ave s de la Membrana Celular,
Garrahan y Alcides Rega.
Duplicaci6n Cromos6mica y Heterocromatina a Nivel Molecular
y Cito16gico, por Nestor O. Bianchi.
Germinaci6n, por Luiz Gouvea Labouria u.
Carlos Caputo.
por Patricio J.
7 8
Nota: Las personas interesadas en adquirir estas obras debendirigir-
se a la Unidad de Ventas y Promoci6n, Organizaci6n de los Es-
tados Americanos, Washington, D. C., 20006 0a las Oficinas de
la Secretarla General de la OEA en el pals respectivo.





























