Embed Size (px)
Citation preview

This article was downloaded by: [UZH Hauptbibliothek / Zentralbibliothek Zürich]On: 05 July 2014, At: 22:44Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
New Zealand Journal of Crop andHorticultural SciencePublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnzc20
Genotypic and environmental variationin bulb composition of New Zealandadapted onion (Allium cepa) germplasmJ. A. McCallum a , D. G. Grant b , E. P. McCartney c , J. Scheffer d ,M. L. Shaw c & R. C. Butler ca New Zealand Institute of Crop & Food Research Limited , PrivateBag 4704, Christchurch, New Zealand E-mail:b Hybrid Seed Company NZ Ltd , Patumahoe, RD 3, Pukekohe, NewZealandc New Zealand Institute of Crop & Food Research Limited , PrivateBag 4704, Christchurch, New Zealandd New Zealand Institute of Crop & Food Research Limited , CroninRd, Pukekohe, New ZealandPublished online: 22 Mar 2010.
To cite this article: J. A. McCallum , D. G. Grant , E. P. McCartney , J. Scheffer , M. L. Shaw & R. C.Butler (2001) Genotypic and environmental variation in bulb composition of New Zealand adaptedonion (Allium cepa) germplasm, New Zealand Journal of Crop and Horticultural Science, 29:3,149-158, DOI: 10.1080/01140671.2001.9514173
To link to this article: http://dx.doi.org/10.1080/01140671.2001.9514173
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or

howsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek /
Zen
tral
bibl
ioth
ek Z
üric
h] a
t 22:
44 0
5 Ju
ly 2
014

New Zealand Journal of Crop and Horticultural Science, 2001, Vol. 29: 149-1580014-0671/01/2903-0149 $7.00 © The Royal Society of New Zealand 2001
149
Genotypic and environmental variation in bulb compositionof New Zealand adapted onion (Allium cepa) germplasm
J. A. MCCALLUM1
D. G. GRANT2
E. P. MCCARTNEY1
J. SCHEFFER3
M. L. SHAW1
R. C. BUTLER1
1New Zealand Institute of Crop & FoodResearch Limited
Private Bag 4704Christchurch, New Zealandemail: [email protected]
2Hybrid Seed Company NZ LtdPatumahoe, RD 3Pukekohe, New Zealand
3New Zealand Institute of Crop & FoodResearch Limited
Cronin RdPukekohe, New Zealand
Abstract Bulb pungency, soluble solids content,and storage quality of 154 lines from the New Zea-land Institute of Crop & Food Research Limitedonion (Allium cepa L.) breeding programme wereevaluated at Pukekohe and Lincoln, New Zealand inthe 1998/99 season. Within-site variation was con-trolled by an experimental design incorporating sys-tematically allocated mild and pungent check linesrather than blocking. Spatial trends in bulb compo-sition were detected and adjustments made to mod-els accordingly, suggesting that this is an appropriate
H00039Received 28 September 2000; accepted 16 May 2001
and efficient means of compensating for within-sitevariation in onion trials. The lines surveyed includedmany that were early, but few that were mild and/orhad low soluble solids. Correlation between solublesolids content and storage loss was observed at bothsites. The correlation observed between maturitydate and frequency of rots at Pukekohe was prob-ably the result of greater impacts of onion thrip dam-age and wet weather on late-maturing lines. Weconclude that although selection for storage andearliness in this programme have been effectivelower pungency may have been lost from mild in-troductions through lack of deliberate selection.
Keywords Allium cepa; onion; pungency; storage;REML; plant breeding
INTRODUCTION
Onions (Allium cepa L.) are New Zealand's mostvaluable vegetable crop. In 1999 over 200 000 tworth NZ$100 million FOB were exported. Over thelast 50 years domestic and export onion productionhave been dominated by cultivars derived from thebrown globe storage cultivar 'PukekoheLongkeeper' (PLK), also known as 'Creamgold'.PLK is thought to have originated from growerselections from 'Straw Spanish' or 'Brown Spanish'varieties (Merry 1967). PLK has exceptional storagelife and is ideal for shipping to distant markets.However, it is desirable to reduce this reliance on anarrow genetic base and broaden the diversity ofonion germplasm grown in New Zealand to widenthe markets that can be served by local producers.
The New Zealand Institute of Crop & FoodResearch Limited (formerly DSIR Crop ResearchDivision) onion breeding programme has beenintroducing, evaluating, and developing oniongermplasm from diverse sources for many years tobroaden the base of adapted breeding material. In1981 an effort to broaden the range of mild, early,and red onion varieties was initiated (Grant 1983;Grant & Carter 1988). This has resulted in a range
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek /
Zen
tral
bibl
ioth
ek Z
üric
h] a
t 22:
44 0
5 Ju
ly 2
014

150 New Zealand Journal of Crop and Horticultural Science, 2001, Vol. 29
of lines that are now relatively well adapted to NewZealand production environments and may beevaluated for open-pollinated or hybrid cultivarusage (Grant et al. 1997).
Onion varieties vary widely in composition, fromthose with firm bulbs of high dry matter content tothose with soft bulbs of low dry matter content, andfrom high to low pungency. The flavour of onionsis determined by the interaction between the sulfurcompounds that impart pungency and thecarbohydrate content. Typically, firm storage oniontypes such as PLK from production areas in higherlatitudes (intermediate to long-day) are highlypungent and have high dry matter with low solublesugars. By contrast, onions selected for lowerlatitudes (short-day varieties) generally have lowerpungency, lower dry matter, and relatively moresimple sugars that impart sweetness (fructose andsucrose). The basis for this association betweenstorage, pungency, and dry matter content is not yetfully understood, though recent genetic (Lin et al.1995; Simon 1995) and physiological (Randle &Bussard 1993) studies have indicated that it couldbe altered through selection. Preliminary results oflinkage mapping suggest that there is a geneticlinkage between loci controlling pungency and drymatter content (Galmarini et al. 2000).
Pungency is strongly influenced by sulfur supply(Randle & Bussard 1993), but there is lessinformation about the effects of environment onsweetness and dry matter content (Randle 1992b).Pungency can be determined by taste testing but thisis complicated by after-taste and is more efficientlyachieved by biochemical analysis (Platenius & Knott1941).
The traditional approach to removingenvironmental influences when comparing theperformance of breeding lines has been to replicateover blocks and environments. An alternative formore economical comparison amongst largenumbers of lines or in early generation selection isto use check plots as controls for environmentalvariation. In this study we employed a design usingcheck lines and residual maximum likelihood(REML) analysis (Gilmour et al. 1997) to permitadjustment for within-field variation and to allowcomparison of a large number of onion breedinglines in a two-site unreplicated trial. The aim of thiswork was to evaluate variation in composition andsome agronomic traits in the Crop & Food Researchonion collection, to assess the results of earlierselection and identify lines for future breeding,conservation, and genetic studies.
MATERIALS AND METHODS
GermplasmA sample of 154 lines from the Crop & FoodResearch onion collection (Grant & Carter 1988)was used to represent the range of moderately towell-adapted material in the programme. Thismaterial had been mass selected for at least threegenerations at Pukekohe and/or Lincoln. Notablegroups within these included selections fromJapanese varieties; selections from crosses betweenthese and 'Early Longkeeper' (ELK); selectionsfrom mild North American 'Grano' and 'Granex'types; derivatives of crosses between PLK types andvarious red lines; male sterile lines selected fromNorth American public breeding programmes(Goldman et al. 2000); and a wide variety ofselections from PLK and PLK-derived populations.The pungent check line was the Crop & FoodResearch PLK selection 'Cron 16' and the mildcheck line was an inbred derived from the 'Grano'selection 'Houston'.
Experimental design and cultural practicesThe experimental design used single replicates of the154 test lines grown at two sites and replication oftwo check lines to estimate within-site variability.The experimental unit was a single plot. Balancedallocation of check plots was carried out bycombining and re-permuting Latin square designs(Pearson & Hartley 1969), to give a total mild pluspungent check rate of 20%. The Pukekohe trial of200 plots was laid out in a 10 row x 20-column blockwith two mild and two check plots allocated per rowand one of each allocated per column. At Lincoln204 plots were laid out in a six row x 34 columnblock with check plots allocated as follows: 12columns with either mild or pungent check and 10with one of each; four rows with four of each check;and two rows with three of each. This designrequired 404 analyses to survey 156 lines, in contrastto a complete randomised block design with threereplicates which would have required 936.
The experiment was sown at the Crop & FoodResearch Pukekohe site (lat. 37° 12'; Patumahoe clayloam) on 7 July 1998 and at the Lincoln site (lat. 43°39'; Wakanui silt loam) on 10 September 1998. Seedwas sown in 1.5 m x 3 m beds with five rows at215 mm spacing using an Ojyord cone seeder drillfitted with Stan Hay coulters. Rows were hand-thinned at the 3-leaf stage to 50-60 mm spacing.Crop management followed standard districtpractices (Lancaster et al. 1995). Crop maturity was
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek /
Zen
tral
bibl
ioth
ek Z
üric
h] a
t 22:
44 0
5 Ju
ly 2
014

McCallum et al.—Bulb composition of New Zealand onion germplasm 151
recorded as the date when 50% of tops had fallenover (Brewster 1990). Bulbs were lifted on 26-27January at Pukekohe, between 25 February and 2March in Lincoln and left to cure for 7-10 days inthe field. At Pukekohe bulbs were topped by handbefore harvest. After topping and harvesting bulbswere graded into large (>70 mm), medium (50-70 mm), small (30-50 mm), and rejects (picklers<30 mm, bolters, and rotted bulbs). A representativesample of 12 bulbs was taken from each plot forcomposition analysis, and remaining bulbs werestored under ambient conditions in sheds on site. At2-monthly intervals for 6 months any bulbs showingsprouting or rots were removed and these storagedefects recorded as frequencies.
Analytical methodsTops, bottom, and outer skins were first removed andthen a vertical segment (1/4 -1/8 depending on size)was removed from each bulb in the sample. Thesewere combined and juiced in a Champion Juicer(PlastaketMfg. Co., Lodi, California, United States).Juice soluble solids concentration (SS) wasmeasured as Brix % in a digital refractometer. Bulbdry matter was also estimated by oven drying theremaining bulb tissue at 80°C. Juice pungency wasmeasured as total pyruvic acid (PA) after at least 10min standing at room temperature and expressed asmicromoles pyruvate per gram fresh weight or perml of juice (Schwimmer & Weston 1961).Background pyruvate levels were not estimated. Todetermine the influence of extraction method wedetermined PA levels in extracts prepared fromsamples of a single plot of each check line by juicerextraction with extracts prepared by blending tissuein an equal volume of water in a Wareing blender(Hamilton et al. 1999).
Statistical analysisAnalysis of composition and agronomic traits wascarried out using residual maximum likelihood(REML) procedures with Genstat 5, release 4.1(1997), using the methods described by Verbyla etal. (1999), Welham & Cullis (1999), and Gilmouret al. (1999). There are two aspects to the analysis-spatial de-trending and estimation of genetic (inter-line) variances between and within sites. Theanalysis was carried out in two stages, site by site,and then both sites together. (1) Genetic variances:in this analysis, the lines were divided into checklines (pungent, mild) and test lines. The differencebetween the pungent check, mild check, and themean of the test lines was fitted as a fixed effect.
The variation amongst the test lines was thenassessed by including test line as a random effect.Thus, the main parameters estimated are the meansfor mild and pungent checks, and the overall meanand genetic variance of the test lines. (2) Examiningspatial patterns: the data were adjusted to accountfor trends in the field by fitting smooth splinesseparately across the rows and/or the columns, wherethe degree of smoothness is determined by the data.For a spline to be fitted, a linear trend across the rows(or columns) must also be included. For some of thedata sets, a linear trend alone was sufficient toremove gross field trends. For computationalpurposes linear or spline trends were fitted at bothsites in both directions. (3) Both sites combined: thespatial and genetic components of the analyses aremodelled as above, but in addition interactionsbetween site and check line, and between site andtest line are fitted. The site-test line interactionenables modelling of different genetic variances foreach site, as well as a genetic correlation betweensites to be estimated. An extra model component wasalso included to allow for different underlyingrandom ("residual") variation at the two sites. Thecross-line analyses were carried out with and withoutspatial adjustment. The genetic variances estimatedafter removal of spatial variation can be consideredmeasures of population variability from a"standardised environment". The proportion ofgenetic variability was calculated for PA and SS ateach site by expressing genetic variance as aproportion of total variance.
Analysis of inter-relationships between bulbcomposition, maturity and storage data was carriedout by correlation and graphical analysis usingMinitab 10.
RESULTS
ClimateThe 1998/99 growing season at Lincoln was drier,sunnier, and less windy than average. Meantemperature in the period September-March was14.2°C, comparable to the long-term average (1971-91) of 13.9°C, and total rainfall over this period was195 mm compared to a long-term average of314 mm. At Pukekohe the July-January period waswarmer, wetter, and less windy than long-termaverages (1972-99). Mean temperature in this periodwas 15.3°C, compared to the long-term average of
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek /
Zen
tral
bibl
ioth
ek Z
üric
h] a
t 22:
44 0
5 Ju
ly 2
014
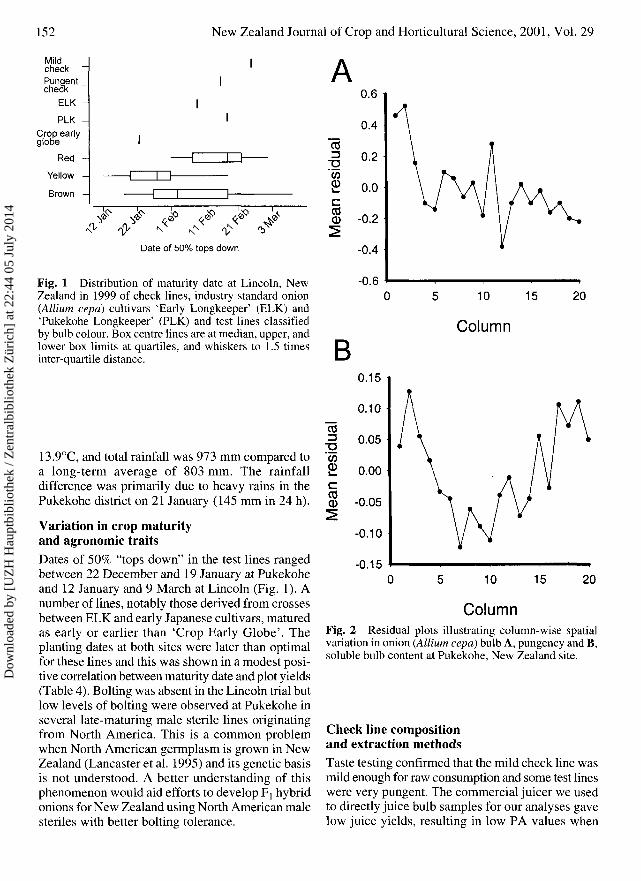
152 New Zealand Journal of Crop and Horticultural Science, 2001, Vol. 29
Mildcheck "Pungentcheck
ELK -
PLK -Crop earlyglobe
Red -
Yellow -
Brown -
A
Date of 50% tops down
Fig. 1 Distribution of maturity date at Lincoln, NewZealand in 1999 of check lines, industry standard onion(Allium cepa) cultivars 'Early Longkeeper' (ELK) and'Pukekohe Longkeeper' (PLK) and test lines classifiedby bulb colour. Box centre lines are at median, upper, andlower box limits at quartiles, and whiskers to 1.5 timesinter-quartile distance.
13.9°C, and total rainfall was 973 mm compared toa long-term average of 803 mm. The rainfalldifference was primarily due to heavy rains in thePukekohe district on 21 January (145 mm in 24 h).
Variation in crop maturityand agronomic traitsDates of 50% "tops down" in the test lines rangedbetween 22 December and 19 January at Pukekoheand 12 January and 9 March at Lincoln (Fig. 1). Anumber of lines, notably those derived from crossesbetween ELK and early Japanese cultivars, maturedas early or earlier than 'Crop Early Globe'. Theplanting dates at both sites were later than optimalfor these lines and this was shown in a modest posi-tive correlation between maturity date and plot yields(Table 4). Bolting was absent in the Lincoln trial butlow levels of bolting were observed at Pukekohe inseveral late-maturing male sterile lines originatingfrom North America. This is a common problemwhen North American germplasm is grown in NewZealand (Lancaster et al. 1995) and its genetic basisis not understood. A better understanding of thisphenomenon would aid efforts to develop Fj hybridonions for New Zealand using North American malesteriles with better bolting tolerance.
0.6
0.4
"cö
2 °'2to
2 0.0
I -0.2
-0.4
-0.6
B
as
0.15
0.10
0.05
2 0.00c
CD -0.05
-0.10
-0.15
5 10 15
Column
20
10
Column
10 20
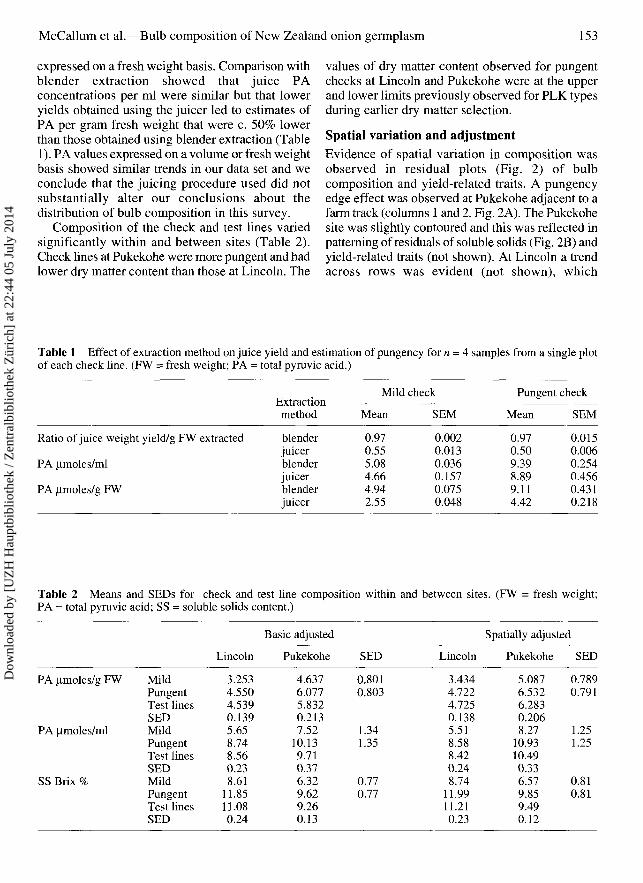
Fig. 2 Residual plots illustrating column-wise spatialvariation in onion (Allium cepa) bulb A, pungency and B,soluble bulb content at Pukekohe, New Zealand site.
Check line compositionand extraction methodsTaste testing confirmed that the mild check line wasmild enough for raw consumption and some test lineswere very pungent. The commercial juicer we usedto directly juice bulb samples for our analyses gavelow juice yields, resulting in low PA values when
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek /
Zen
tral
bibl
ioth
ek Z
üric
h] a
t 22:
44 0
5 Ju
ly 2
014
McCallum et al.—Bulb composition of New Zealand onion germplasm 153
expressed on a fresh weight basis. Comparison withblender extraction showed that juice PAconcentrations per ml were similar but that loweryields obtained using the juicer led to estimates ofPA per gram fresh weight that were c. 50% lowerthan those obtained using blender extraction (Table1). PA values expressed on a volume or fresh weightbasis showed similar trends in our data set and weconclude that the juicing procedure used did notsubstantially alter our conclusions about thedistribution of bulb composition in this survey.
Composition of the check and test lines variedsignificantly within and between sites (Table 2).Check lines at Pukekohe were more pungent and hadlower dry matter content than those at Lincoln. The
values of dry matter content observed for pungentchecks at Lincoln and Pukekohe were at the upperand lower limits previously observed for PLK typesduring earlier dry matter selection.
Spatial variation and adjustmentEvidence of spatial variation in composition wasobserved in residual plots (Fig. 2) of bulbcomposition and yield-related traits. A pungencyedge effect was observed at Pukekohe adjacent to afarm track (columns 1 and 2, Fig. 2A). The Pukekohesite was slightly contoured and this was reflected inpatterning of residuals of soluble solids (Fig. 2B) andyield-related traits (not shown). At Lincoln a trendacross rows was evident (not shown), which
Table 1 Effect of extraction method on juice yield and estimation of pungency for n = 4 samples from a single plotof each check line. (FW = fresh weight; PA = total pyruvic acid.)
Ratio of juice weight yield/g FW extracted
PA umoles/ml
PA umoles/g FW
Fxtractionmethod
blenderjuicerblenderjuicerblenderjuicer
Mild
Mean
0.970.555.084.664.942.55
check
SEM
0.0020.0130.0360.1570.0750.048
Pungent
Mean
0.970.509.398.899.114.42
check
SEM
0.0150.0060.2540.4560.4310.218
Table 2 Means and SEDs for check and test line composition within and between sites. (FW = fresh weight;PA = total pyruvic acid; SS = soluble solids content.)
PA umoles/g FW
PA umoles/ml
SS Brix %
MildPungentTest linesSEDMildPungentTest linesSEDMildPungentTest linesSED
Lincoln
3.2534.5504.5390.1395.658.748.560.238.61
11.8511.080.24
Basic adjusted
Pukekohe
4.6376.0775.8320.2137.52
10.139.710.376.329.629.260.13
SED
0.8010.803
1.341.35
0.770.77
Lincoln
3.4344.7224.7250.1385.518.588.420.248.74
11.9911.210.23
Spatially adjusted
Pukekohe
5.0876.5326.2830.2068.27
10.9310.490.336.579.859.490.12
SED
0.7890.791
1.251.25
0.810.81
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek /
Zen
tral
bibl
ioth
ek Z
üric
h] a
t 22:
44 0
5 Ju
ly 2
014
154 New Zealand Journal of Crop and Horticultural Science, 2001, Vol. 29
paralleled the layout of irrigation lines. Applicationof spatial adjustment led to significant reductions inerror (i.e., unexplained plot-to-plot) variance.
Genetic and environmentalvariation in pungency and soluble solids
There were notable differences in residual (random)variances for PA and SS between sites. Residualvariance for PA was higher at Pukekohe, but residualvariance for SS was greater at Lincoln. The lines
exhibited similar trends between sites to those shownby the checks, with lower SS and higher PA atPukekohe (Table 2). The proportion of geneticvariability estimated for PA and SS (Table 3) aresimilar to broad-sense heritability values estimatedby others using much narrower samples ofgermplasm (Simon 1995). Distribution of bulbcomposition in the sample of lines was skewedtoward higher dry matter and pungency, with fewlines milder and lower in SS than the pungent check
Table 3 Test line variance and within-site estimates of genetic variability obtained from REML without("basic adjusted") and with ("spatially adjusted") spatial adjustment. (FW = fresh weight; PA = total pyruvic acid;SS = soluble solids content.)
PA umoles/g FW
PA umoles/ml
SS Brix %
Residual varianceGenetic varianceGenetic variabilityResidual varianceGenetic varianceGenetic variabilityResidual varianceGenetic varianceGenetic variability
Lincoln
0.2070.2800.580.59
0.8180.580.611.340.69
Basic adjusted
Pukekohe Correlation
0.4840.2790.371.460.9160.3860.171.210.87
0.404
0.515
0.95
Lincoln
0.2060.2770.570.6040.7700.560.591.350.69
Spatially adjusted
Pukekohe (
0.4550.2740.381.1791.1080.4840.171.180.88
Correlation
0.442
0.588
0.94
Table 4 Correlations between bulb composition, storage measures, and maturity for 156 onion (Allium cepa) lines atPukekohe (P) and Lincoln (L), New Zealand sites. Pyruvic acid (PA) and soluble solids (SS) are spatially adjusted.Notable correlations are highlighted in bold. Rot % is frequency of rots up to 2 months after harvest. Sprout % isfrequency of sprouted bulbs up to 6 months after harvest. Maturity is date of 50% "tops down". (FW = fresh weight;PA = total pyruvic acid; SS = soluble solids content.)
PA umoles/g FWPA umoles/ml
SS Brix %
Maturity
Rot %
Sprout %
T2 0%
Yield
PLPLPLPLPLPLPLP
PA umoles/g PA SSFW umoles/ml Brix %
L P L
0.680.85 0.710.59 0.85 0.790.32 0.47 0.400.29 0.45 0.37
-0.26-0.06-0.16-0.20 0.00-0.03
P L P
0.450.41 0.990.00-0.11-0.180.11-0.03-0.12
0.03 -0.22 -0.04 -0.19 -0.43 -0.45-0.18-0.01-0.05 0.11-0.21-0.26
Maturity Rot
L
0.690.010.52
-0.21 -0.35 -0.25 -0.34 -0.64 -0.58 -0.06-0.10-0.08 0.01
0.19 0.32 0.250.19 0.11 0.07
-0.23 0.09-0.15
0.00-0.37-0.350.29 0.56 0.49
0.080.10
P L
0.060.50 0.090.15 0.550.09 0.180.22-0.39
% Sprout %
P L P
0.010.20 0.360.05-0.89-0.32
0.06 0.51 0.51-0.31-0.28-0.27-0.44-0.48-0.800.09 0.24 0.19
-0.24 0.02-0.20-0.02 0.19 0.140.450.27
0.55 -0.320.39-0.26
0.21-0.37-0.120.04-0.34-0.18
T20 % Yield
L P L
0.400.41 0.090.35 0.16 0.92
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek /
Zen
tral
bibl
ioth
ek Z
üric
h] a
t 22:
44 0
5 Ju
ly 2
014
McCallum et al.—Bulb composition of New Zealand onion germplasm 155
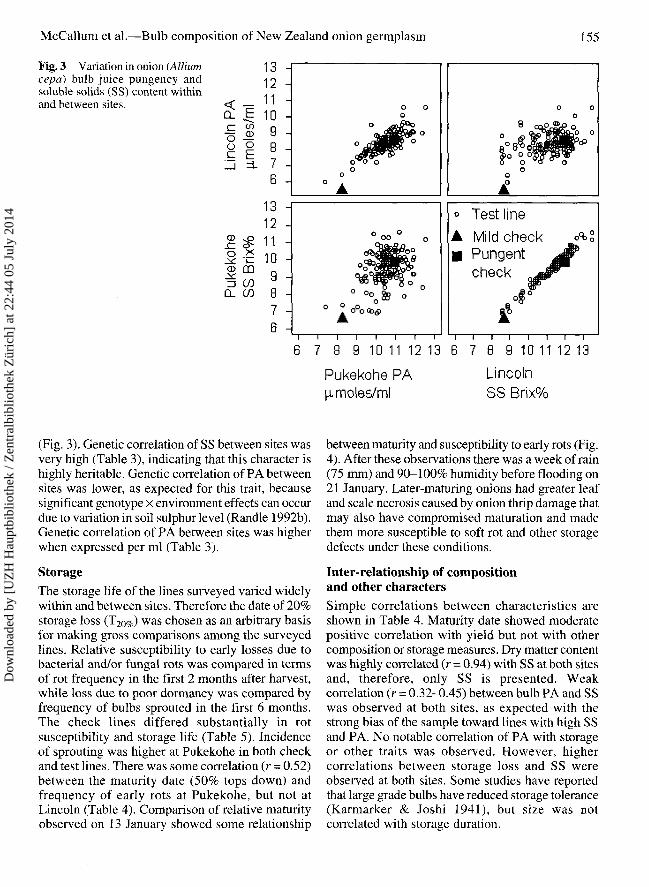
Fig. 3 Variation in onion (Alliumcepa) bulb juice pungency andsoluble solids (SS) content withinand between sites.
131211109876
131211109
r 1o> ûû
Q_ 00 8
76
O 00
o o o
° A
1 1 1 1 1 1 1 1
0 00
8 o ®_o
oo
A
° Test lineA Mild check A g• Pungent j J *
check agßr
6 7 8 9 10 11 12 13 6
Pukekohe PAH-moles/rnl
7 8 9 10 11
Lincoln
SS Brix%
12 13
(Fig. 3). Genetic correlation of SS between sites wasvery high (Table 3), indicating that this character ishighly heritable. Genetic correlation of PA betweensites was lower, as expected for this trait, becausesignificant genotype x environment effects can occurdue to variation in soil sulphur level (Randle 1992b).Genetic correlation of PA between sites was higherwhen expressed per ml (Table 3).
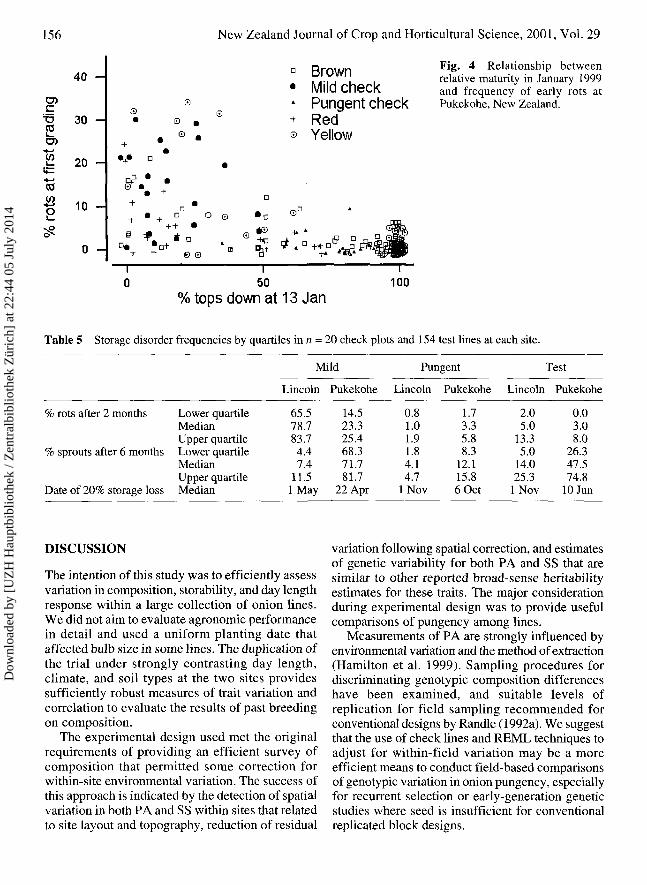
StorageThe storage life of the lines surveyed varied widelywithin and between sites. Therefore the date of 20%storage loss (T20%) was chosen as an arbitrary basisfor making gross comparisons among the surveyedlines. Relative susceptibility to early losses due tobacterial and/or fungal rots was compared in termsof rot frequency in the first 2 months after harvest,while loss due to poor dormancy was compared byfrequency of bulbs sprouted in the first 6 months.The check lines differed substantially in rotsusceptibility and storage life (Table 5). Incidenceof sprouting was higher at Pukekohe in both checkand test lines. There was some correlation (r = 0.52)between the maturity date (50% tops down) andfrequency of early rots at Pukekohe, but not atLincoln (Table 4). Comparison of relative maturityobserved on 13 January showed some relationship
between maturity and susceptibility to early rots (Fig.4). After these observations there was a week of rain(75 mm) and 90-100% humidity before flooding on21 January. Later-maturing onions had greater leafand scale necrosis caused by onion thrip damage thatmay also have compromised maturation and madethem more susceptible to soft rot and other storagedefects under these conditions.
Inter-relationship of compositionand other charactersSimple correlations between characteristics areshown in Table 4. Maturity date showed moderatepositive correlation with yield but not with othercomposition or storage measures. Dry matter contentwas highly correlated (r = 0.94) with SS at both sitesand, therefore, only SS is presented. Weakcorrelation (r = 0.32-0.45) between bulb PA and SSwas observed at both sites, as expected with thestrong bias of the sample toward lines with high SSand PA. No notable correlation of PA with storageor other traits was observed. However, highercorrelations between storage loss and SS wereobserved at both sites. Some studies have reportedthat large grade bulbs have reduced storage tolerance(Karmarker & Joshi 1941), but size was notcorrelated with storage duration.
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek /
Zen
tral
bibl
ioth
ek Z
üric
h] a
t 22:
44 0
5 Ju
ly 2
014
156 New Zealand Journal of Crop and Horticultural Science, 2001, Vol. 29
CO
.8
e
40 -
30 -
£ 20 -
10 -
0 -
©
o ©
+ + © ©
BrownMild checkPungent checkRedYellow
Fig. 4 Relationship betweenrelative maturity in January 1999and frequency of early rots atPukekohe, New Zealand.
n i0 50
% tops down at 13 Jan100
Table 5 Storage disorder frequencies by quartiles in n = 20 check plots and 154 test lines at each site.
Mild Pungent Test
Lincoln Pukekohe Lincoln Pukekohe Lincoln Pukekohe
% rots after 2 months Lower quartile 65.5 14.5 0.8 1.7 2.0 0.0Median 78.7 23.3 1.0 3.3 5.0 3.0Upper quartile 83.7 25.4 1.9 5.8 13.3 8.0
% sprouts after 6 months Lower quartile 4.4 68.3 1.8 8.3 5.0 26.3Median 7.4 71.7 4.1 12.1 14.0 47.5Upper quartile 11.5 81.7 4.7 15.8 25.3 74.8
Date of 20% storage loss Median 1 May 22 Apr INov 6 0ct INov lO Jun
DISCUSSION
The intention of this study was to efficiently assessvariation in composition, storability, and day lengthresponse within a large collection of onion lines.We did not aim to evaluate agronomic performancein detail and used a uniform planting date thataffected bulb size in some lines. The duplication ofthe trial under strongly contrasting day length,climate, and soil types at the two sites providessufficiently robust measures of trait variation andcorrelation to evaluate the results of past breedingon composition.
The experimental design used met the originalrequirements of providing an efficient survey ofcomposition that permitted some correction forwithin-site environmental variation. The success ofthis approach is indicated by the detection of spatialvariation in both PA and S S within sites that relatedto site layout and topography, reduction of residual
variation following spatial correction, and estimatesof genetic variability for both PA and SS that aresimilar to other reported broad-sense heritabilityestimates for these traits. The major considerationduring experimental design was to provide usefulcomparisons of pungency among lines.
Measurements of PA are strongly influenced byenvironmental variation and the method of extraction(Hamilton et al. 1999). Sampling procedures fordiscriminating genotypic composition differenceshave been examined, and suitable levels ofreplication for field sampling recommended forconventional designs by Randle (1992a). We suggestthat the use of check lines and REML techniques toadjust for within-field variation may be a moreefficient means to conduct field-based comparisonsof genotypic variation in onion pungency, especiallyfor recurrent selection or early-generation geneticstudies where seed is insufficient for conventionalreplicated block designs.
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek /
Zen
tral
bibl
ioth
ek Z
üric
h] a
t 22:
44 0
5 Ju
ly 2
014

McCallum et al.—Bulb composition of New Zealand onion germplasm 157
The differences observed in bulb composition andstorage between sites may be caused by severalfactors. The January 1999 flooding at Pukekohe andhigh levels of onion thrips increased incidence of rotsin later maturing lines and could have shortenedstorage life in others. Earlier work has demonstratedthat water during curing increases rot frequency(Wright & Grant 1997). The higher meantemperatures of bulb storage at Pukekohe mayshorten bulb dormancy compared to Lincoln(Yamaguchi et al. 1957). The sharp contrast in drymatter content between the same genotypes grownat Lincoln and Pukekohe may also have influencedstorage. There is anecdotal evidence that restrictingirrigation (and thus dry matter content) may improvestorage quality.
It is likely that the variation in pungency withinand between the two sites is a result of differencesin sulfur availability between the Lincoln loam andPukekohe volcanic soils. There is evidence that soilsulfur can also affect carbohydrate composition(Randle 1992b). Comparison of pungency figuresbetween studies is difficult as a result of the varietyof juice extraction methods used (Hamilton et al.1999) but we suggest our pyruvate values expressedon a fresh weight basis be doubled to compare withthose quoted by Dean (1999).
The germplasm used to broaden the base of theCrop & Food Research onion collection has includeda wide variety of material from a more geneticallydiverse "short-day" background (Bark & Havey1995). It was expected that this would provide agreater variety of bulb composition, as well as otheragronomic traits, to extend the variation present inPLK germplasm. The recent emphasis duringdevelopment of the Crop & Food Research breedingprogramme has been primarily on storage andearliness (Grant & Carter 1988). This is reflected bythe number of early lines with excellent storage,notably selections from crosses between ELK andJapanese cultivars. However, despite introducing,selecting and inter-crossing milder onion varieties inthe Crop & Food Research programme, few mildlines have been retained that would be suitable forfresh market use. Red lines were mostly similar incomposition to the pungent PLK check, suggestingthat they are primarily suitable for export storagemarkets than fresh markets.
The skewing of composition toward lines withhigh soluble solids and pungency is to be expectedwith the emphasis on PLK-derived lines in theprogramme, although it may be a result of traitcorrelation. A relatively strong association between
SS content and storability is evident within thegenotypes surveyed. High dry matter, lowerpungency lines would be clearly preferable for eitherexport or local fresh markets that require a milderonion, but are rare in the sample surveyed. Loss ofmildness has possibly occurred because selection forstorage life and thus higher dry matter has been verystrong under the storage conditions employed andselection for higher pungency has occurred throughgenetic linkage or pleiotropism (Galmarini et al.2000). Larger population sizes, minimal storageselection, and biochemical and/or marker-basedselection for desired pungency and dry matterphenotypes in early generations are needed to enableselection of lines with desired quality.
PLK is world renowned for its storage quality,although it was derived from varieties that arethemselves not known for their quality. The mild andhumid conditions that prevail in the Pukekohe areanot only favour fungal and bacterial diseases thatcompromise storage, but also physiological changesin bulbs that lead to sprouting and spoilage. Wespeculate that the exceptional storage properties ofPLK have been achieved because prolonged massselection for storage quality was carried out on basepopulations with suitable variation in an area wherethe climate is extremely unfavourable for onionstorage. Breeders overseas as a source of superiorstorage genetics are exploiting PLK germplasm. Thechallenge for New Zealand breeders is to maintainthe quality of PLK while introgressing desirablevariation for composition and disease quality fromother sources. This will be aided by improvedknowledge of the genetics of these characters andnew technologies to manipulate them usingmolecular markers.
ACKNOWLEDGMENTS
This work was funded by the New Zealand Foundationfor Research, Science and Technology ProgrammeCO2808.
REFERENCES
Bark, O. H.; Havey, M. J. 1995: Similarities and relation-ships among populations of the bulb onion asestimated by nuclear RFLPs. Theoretical and Ap-plied Genetics 90: 407-414.
Brewster, J. L. 1990: Physiology of crop growth andbulbing. In: Rabinowitch, H. D.; Brewster, J. L.ed. Onions and allied crops. Vol. 1. Boca Raton,CRC Press Inc.
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek /
Zen
tral
bibl
ioth
ek Z
üric
h] a
t 22:
44 0
5 Ju
ly 2
014

158 New Zealand Journal of Crop and Horticultural Science, 2001, Vol. 29
Dean, W. B. 1999: What is a sweet onion? Onion WorldMay/June: 18.
Galmarini, C. R.; Goldman, I. L.; Havey, M. J. 2000:Quantitative trait loci controlling solid content,pungency and antiplatelet activity of onion (Al-lium cepa L.). Plant & Animal Genome VIIIConference, Town & Country Hotel, San Diego,CA, United States.
Genstat 5 Committee 1997: Genstat 5, Release 4.1, Ref-erence Manual Supplement. Lawes AgriculturalTrust, Rothamsted Experimental Station.
Gilmour, A. R.; Cullis, B. R.; Verbyla, A. P. 1997: Ac-counting for natural and extraneous variation inthe analysis of field experiments. Journal of Ag-ricultural, Biological, and Environmental Statis-tics 2(1): 269-293.
Gilmour, A. R. C ; Welham, S. J.; Thompson, R. 1999:ASREML Manual.
Goldman, I. L.; Schroek, G.; Havey, M. J. 2000: Historyof public onion breeding programs in the UnitedStates. Plant Breeding Reviews 20: 68-103.
Grant, D. G. 1983: Onions. In: Wratt, G. S.; Smith, H. C.ed. Plant breeding in New Zealand. Christchurch,Butterworths NZ Ltd. Pp. 165-170.
Grant, D. G.; Carter, B. 1986: Onion breeding in theDSIR. Proceedings of the Agronomy Society ofNew Zealand 16: 115-117.
Grant, D. G. Carter, B. V. 1988: Progress in the evalua-tion and use of onion germplasm in New Zealand.Proceedings of the 4th Eucarpia Allium Sympo-sium, Wellesborne, United Kingdom.
Grant, D. G.; McCartney, E. P.; Stuart, C. S. 1997:Breeding and evaluation of intermediate day-length storage onion hybrids. Second InternationalSymposium on Edible Alliaceae, Adelaide,Australia.
Hamilton, B. K.; Yoo, K. S.; Pike L. M. 1999: Juiceextraction methods make difference in pungencyestimation of onion. Onion World July/August:25-26.
Karmarkar, D. V.; Joshi, B. M. 1941: Investigations onthe storage of onions. Indian Journal of Agricul-tural Science 11: 82-85.
Lancaster, J. E.; McCartney, E. P.; Jermyn, W. A.;Johnstone, J. V. 1995: Identification of onioncultivars for commercial production in Canter-bury, New Zealand. New Zealand Journal of Cropand Horticultural Science 23: 299-306.
Lin, M.; Watson J. F.; Baggett J. R. 1995: Inheritance ofsoluble solids and pyruvic acid content of bulbonions. Journal of the American Society for Hor-ticultural Science 120: 119-122.
Merry, D. M. E. 1967: Years of care and selection pro-duced the noted Pukekohe onion. New ZealandJournal of Agriculture 114: 46-49.
Pearson, E. S.; Hartley, H. O. ed. 1969: Biometrika tablesfor statisticians. London, Biometrika Trust.
Platenius, H. K.; Knott, J. E. 1941: Factors affectingonion pungency. Journal of Agricultural Research62: 371-380.
Randle, W. M. 1992a: Sulfur nutrition affects nonstructuralwater-soluble carbohydrates in onion germplasm.HortScience 27: 52-55.
Randle, W. M. 1992b: Sampling procedures to estimateflavor potential in onion. HortScience 27: 1116-1117.
Randle, W. M.; Bussard, M. L. 1993: Pungency andsugars of short-day onions as affected by sulfurnutrition. Journal of the American Society forHorticultural Science 118: 766-770.
Schwimmer, S., Weston, W. J. 1961: Enzymatic develop-ment of pyruvic acid in onion as a measure ofpungency. Journal of Agricultural and FoodChemistry 9: 301-304.
Simon, P. W. 1995: Genetic analysis of pungency andsoluble solids in long-storage onions. Euphytica82: 1-8.
Verbyla, A. P.; Cullis B. R.; Kenward, M. G.; Welham, S.J. 1999: The analysis of designed experimentsand longitudinal data by using smoothing splines.Applied Statistics 48: 269-300.
Welham, S. J.; Cullis, B. R. 1999: Analysis of correlateddata using REML. Australasian Genstat Confer-ence, Genstat99, Melbourne, Australia.
Wright, P. J.; Grant, D. G. 1997: Effects of culturalpractices at harvest on onion bulb quality andincidence of rots in storage. New Zealand Journalof Crop and Horticultural Science 25: 353-358.
Yamaguchi, M.; Pratt, H. K.; Morris, L. L. 1957: Effectof storage temperatures on keeping quality andcomposition of onion bulbs and on subsequentdarkening of dehydrated flakes. Proceedings ofthe American Society of Horticultural Science69: 421-426.
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek /
Zen
tral
bibl
ioth
ek Z
üric
h] a
t 22:
44 0
5 Ju
ly 2
014




![RESEARCH Open Access Genotypic variation in sorghum [Sorghum … · 2018-06-03 · RESEARCH Open Access Genotypic variation in sorghum [Sorghum bicolor (L.) Moench] exotic germplasm](https://img.pdfslide.net/doc/110x75/5f03a3af7e708231d40a0b15/research-open-access-genotypic-variation-in-sorghum-sorghum-2018-06-03-research.jpg)
























