Embed Size (px)
Citation preview

Genotyping of Mycobacterium tuberculosis and
Mycobacterium leprae ancient DNA
A thesis submitted to The University of Manchester for the degree of
Doctor of Philosophy
in the Faculty of Science and Engineering
2020
Ammielle A. Kerudin
School of Earth and Environmental Sciences

2
Table of Contents
List of Tables ................................................................................................................... 5
List of Figures .................................................................................................................. 7
List of Abbreviations ..................................................................................................... 10
Abstract .......................................................................................................................... 13
Declaration ..................................................................................................................... 14
Copyright ....................................................................................................................... 14
Dedication and Acknowledgement ................................................................................ 15
Chapter 1: Introduction .................................................................................................. 16
1.1 Aims and objectives ............................................................................................. 16
1.2 The biology of tuberculosis and leprosy .............................................................. 18
1.3 Skeletal changes in tuberculosis and leprosy ....................................................... 25
1.4 History of tuberculosis and leprosy based on historical documents and skeletal
changes evidence ........................................................................................................ 31
1.4.1 History of tuberculosis..................................................................................... 31
1.4.2 History of leprosy ............................................................................................ 34
1.4.3 The concern in using historical documents as evidence of ancient disease .... 36
1.5 Ancient DNA........................................................................................................ 38
1.5.1 Ancient DNA background ............................................................................... 38
1.5.2 Characteristic of ancient DNA ........................................................................ 38
1.5.2.1 Fragmentation ........................................................................................... 39
1.5.2.2 Miscoding lesions ..................................................................................... 40
1.5.2.3 Blocking lesions ........................................................................................ 41
1.6 Ancient DNA studies of tuberculosis and leprosy ............................................... 42
1.6.1 Ancient DNA studies of tuberculosis .............................................................. 42
1.6.1.1 Case confirmation ..................................................................................... 42
1.6.1.2 Origin and evolution of tuberculosis ......................................................... 44
1.6.2 Ancient DNA studies of leprosy ...................................................................... 50
1.6.2.1 Case confirmation ..................................................................................... 50
1.6.2.2 Origin and evolution of leprosy ................................................................ 52
Chapter 2: Materials and methods ................................................................................. 56
2.1 Archaeological samples........................................................................................ 56
2.2 Authentication regimes ........................................................................................ 66
2.3 Bone scraping and crushing ................................................................................. 67
2.4 Ancient DNA extraction....................................................................................... 68
2.5 PCR assays screening for ancient DNA preservation .......................................... 69

3
2.6 Next generation sequencing ................................................................................. 73
2.6.1 DNA library preparation .................................................................................. 73
2.6.2 Target enrichment: in-solution target hybridization capture ........................... 76
2.6.3 Quality control and quantification of sequencing libraries .............................. 78
2.7 Bioinformatics analysis ........................................................................................ 80
2.7.1 Merging of paired reads and removal of adapter sequences ............................ 82
2.7.2 Mapping to reference genome ......................................................................... 83
2.7.3 Cleaning and sorting reads (PicardTools) ........................................................ 83
2.7.4 Metagenomic content analysis ......................................................................... 84
2.7.5 Sequence variant analysis ................................................................................ 84
Chapter 3: Study of M. tuberculosis aDNA in archaeological remains from Yorkshire,
England. ......................................................................................................................... 85
Part I: MTBC aDNA screening by polymerase chain reaction. ..................................... 85
3.1 Introduction .......................................................................................................... 85
3.2 Results .................................................................................................................. 87
3.2.1 St Andrew Fishergate 6.................................................................................... 95
3.2.2 St Andrew Fishergate 277 ................................................................................ 99
3.2.3 St Andrew Fishergate 339................................................................................ 99
3.2.4 St Helen on the walls 6003 ............................................................................ 100
3.2.5 East Heslington 229 ....................................................................................... 102
3.2.6 Wetwang Slack 2 ........................................................................................... 102
3.2.7 Wetwang Slack 7 ........................................................................................... 104
3.2.8 Wharram Percy 26 and Wharram Percy 1600 ............................................... 104
3.2.9 Addingham 134 and Addingham 223 ............................................................ 106
3.2.10 Addingham 103 ............................................................................................ 106
3.2.11 Melton 5319 ................................................................................................. 109
3.2.12 Hickleton 46 ................................................................................................. 109
3.2.13 St Giles by Brompton Bridge 1542 .............................................................. 111
3.2.14 Sewerby 34 .................................................................................................. 111
3.3 Discussion........................................................................................................... 112
3.3.1 MTBC positive samples ................................................................................. 112
3.3.2 Contamination ................................................................................................ 113
3.3.3 Failure of MTBC aDNA detection ................................................................ 116
3.3.4 Samples for Next Generation Sequencing ..................................................... 118
Chapter 4: Study of M. tuberculosis aDNA in archaeological remains from Yorkshire,
England. Part 2: Next Generation Sequencing. ............................................................ 120

4
4.1 Introduction ........................................................................................................ 120
4.2 Results ................................................................................................................ 121
4.2.1 Shotgun sequencing...................................................................................... 121
4.2.2 Target hybridization capture .......................................................................... 143
4.3 Discussion .......................................................................................................... 144
4.3.1 Efficiency of shotgun sequencing in isolating endogenous DNA ................. 144
4.3.2 Taxonomical content of the archaeological samples ..................................... 146
4.3.3 Target enrichment sequencing strategy ......................................................... 150
4.3.4 Possible mixed infection in sample St Andrew Fishergate House 6 ............. 151
4.3.5 MTBC aDNA detection in bone remains from Yorkshire ............................ 151
Chapter 5: Genotyping of Mycobacterium leprae ancient DNA from mediaeval England
...................................................................................................................................... 153
5.1 Background of study .......................................................................................... 153
5.2 Publication draft ................................................................................................. 155
5.2.1 Introduction ................................................................................................... 156
5.2.2 Material and methods .................................................................................... 159
5.2.2.1 Skeletons ................................................................................................. 159
5.2.2.2 Ancient DNA regime .............................................................................. 162
5.2.2.3 DNA extraction, PCR and sequencing.................................................... 162
5.2.2.4 Data analysis ........................................................................................... 164
5.2.3 Results ........................................................................................................... 164
5.2.4 Discussion ...................................................................................................... 167
5.3 Supplementary information ................................................................................ 175
Chapter 6: Conclusion.................................................................................................. 190
6.1 The extent to which the objectives have been addressed: objective one
(tuberculosis) ............................................................................................................ 190
6.2 The extent to which the objectives have been addressed: objective two
(leprosy)…………………………………………………………………………….192
6.3 Limitations of the thesis and future work........................................................... 192
6.4 Ethical issues raised by this work ...................................................................... 193
References .................................................................................................................... 195
Appendices ................................................................................................................... 218
Word counts: 47, 645 words

5
List of Tables
Table 2.1: A complete list of all bone remains studied for the preservation of MTBC
aDNA……………………………………………………………………..59-65
Table 2.2: List of primers used in the PCR screening for MTBC aDNA presence..........72
Table 3.1: Result summary for all PCR assays tested on all samples, using original DNA
extract and 10-fold diluted DNA…….…………………....……...............88-92
Table 3.2: Sanger sequence result summary for 15 samples with a positive band in at
least one of the PCR assays…………………………………………...…93-94
Table 4.1: The result summary of the shotgun read mapping against the M. tuberculosis
reference genome……………….……………………………….…………123
Table 4.2: The number of reads assigned to each super kingdom, genus and species of
interest by MEGAN for sample St Andrew Fishergate 253……………..…124
Table 4.3: The number of reads assigned to each super kingdom, genus and species of
interest by MEGAN for sample St Andrew Fishergate 6……………...…...127
Table 4.4: The number of reads assigned to each super kingdom, genus and species of
interest by MEGAN for sample St Helen-on-the-Walls 5494…….….…….130
Table 4.5: The number of reads assigned to each super kingdom, genus and species of
interest by MEGAN for sample Helen-on-the-Walls 6003………………...133
Table 4.6: The number of reads assigned to each super kingdom, genus and species of
interest by MEGAN for sample Hickleton 46……………………….....…..135
Table 4.7: The number of reads assigned to each super kingdom, genus and species of
interest by MEGAN for sample Wetwang Slack 185…………….…..…….137
Table 4.8: The number of reads assigned to each super kingdom, genus and species of
interest by MEGAN for sample Wetwang Slack 7………………..…..……139
Table 4.9: The number of reads assigned to each super kingdom, genus and species of
interest by MEGAN for sample Wetwang Slack 8………………..………..141
Table 4.10: Comparison of shotgun and target enrichment NGS results for St Andrew
Fishergate 6.……………………………………………………….……….143
Table 4.11: The DNA library concentrations for the 8 samples subjected to shotgun
NGS………………………………………………………………..……….144
Table 4.12: Comparison of the 20 most abundant species in each sample as determined
by BLAST and MEGAN analysis of the reads obtained by shotgun
sequencing………………………………………………...………..…148- 149
Table 5.1. Details of skeletons and samples that were taken………………….………161
Table 5.2. Results of RLEP PCRs……………………………………………..………165
Table 5.3. Genotype assignments……………………………………………..………166
Table S5.1. Detailed osteological report………………………………………;;….…184

6
Table S5.2. Summary statistics for Illumina sequencing following enrichment of
samples by in-solution hybridization……………………………………….185
Table S5.3. Identities in the C21, C48 and R5046 genomes of the 215 SNPs known in
modern M. leprae strains………………………………………………..…186
Table S5.4. Unique variations (highlighted in green) present in the C21, C48 and/or
R5046 genomes………………………………..………………….....187-189

7
List of Figures
Figure 1.1: The mechanism of tuberculosis infection in cells and granulomas formation
in active and latent tuberculosis infection…………………………………20
Figure 1.2: The bone changes indicative of tuberculosis infection in skeletal
remains………………………………...…………………………………..27
Figure 1.3: The bone changes of leprosy on foot……………………………………...29
Figure 1.4: The genotyping scheme of MTBC members using the katG463 and gyrA95
markers. …………………………………………………………...………45
Figure 1.5: MTBC phylogenetic lineage 1 to 7……………………………………..…46
Figure 1.6: The distribution of different leprosy subtypes around the world………….54
Figure 2.1: The origin locations of the Yorkshire archaeological remains……………58
Figure 2.2: A simplified scheme of make up of dual-indexed DNA library
fragments………………………………………………………………….74
Figure 2.3: The relationship between the SPRI beads-to-template ratios to fragment size
selection……………………………………………………………………79
Figure 2.4: The bioinformatics analysis flows performed on different samples………81
Figure 2.5: Three outcome from paired-end data from Illumina sequencing by
AdapterRemoval v2……………………………………….……………….82
Figure 3.1: Gel electrophoresis results for the sample St Andrew Fishergate 6 showing
positive bands for three markers………………………………..….………96
Figure 3.2: The alignment of St Andrew Fishergate 6 Sanger clone sequences of IS6110
123 bp and nested 92 bp PCR product against M. tuberculosis H37Rv
reference sequence……………………………….….……………………..97
Figure 3.3: The alignment of St Andrew Fishergate House 6 Sanger clone sequences of
gyrA and Pks 15/1 PCR product against M. tuberculosis H37Rv reference
sequence……………………………………………………………………98
Figure 3.4: The alignment of St Andrew Fishergate House 339 clone sequence of gyrA
PCR product against the reference M. tuberculosis H37Rv sequence…….99
Figure 3.5: gyrA PCR amplification for sample St Helen-on-the-Walls 6003……..…100
Figure 3.6: The alignment of St Helen on the Walls 6003 clone sequences from the gyrA
PCR assay to the M. tuberculosis H37Rv reference sequence…..……….101
Figure 3.7: The alignment of Heslington East 229 clone sequences from the IS6110 123
bp PCR assay to the M. tuberculosis H37Rv reference sequence…….…101
Figure 3.8: The alignment of Wetwang Slack 2 clone sequences from the gyrA PCR
assay against the M. tuberculosis H37Rv reference sequence……….….103

8
Figure 3.9: The alignment of Wetwang Slack 7 clone sequences from the IS6110 123 bp
PCR assay against the M. tuberculosis H37Rv reference sequence...........103
Figure 3.10: The alignment of Wharram Percy 26 clone sequences from the gyrA PCR
assay against the M. tuberculosis H37Rv reference
sequence……………………………………………………………….....105
Figure 3.11: The alignment of Wharram Percy 1600 clone sequences from the gyrA PCR
assay against the M. tuberculosis H37Rv reference sequence.…..............105
Figure 3.12: The alignment of Addingham 134 and Addingham 223 clone sequences
from the IS6110 first step, 123 bp PCR assay against the M. tuberculosis
H37Rv reference sequence…………………………….…….….………...107
Figure 3.13: The alignment of Addingham 103 clone sequences from the gyrA PCR
assay against the M. tuberculosis H37Rv reference sequence…….……...108
Figure 3.14: The alignment of Melton 5319 clone sequences from the gyrA PCR assay
against the M. tuberculosis H37Rv reference sequence……...…………..108
Figure 3.15: The alignment of Hickleton 46 clone sequences from the gyrA PCR assay
against the M. tuberculosis H37Rv reference sequence…………………..110
Figure 3.16: IS6110 123 bp and nested 92 bp PCR amplification of sample Sewerby
34………………………………………………………………………...111
Figure 4.1: The 20 species with highest read number assigned by MEGAN for sample
St Andrew Fishergate 253………………………………………………..126
Figure 4.2: The 20 species with highest read number assigned by MEGAN for sample St
Andrew Fishergate 6……………………………………………………..129
Figure 4.3: The 20 species with highest read number assigned by MEGAN for sample St
Helen-on-the-Walls 5494………………………………………………..131
Figure 4.4: The 20 species with highest read number assigned by MEGAN for sample St
Helen-on-the-Walls 6003………………………………………………..134
Figure 4.5: The 20 species with highest read number assigned by MEGAN for sample
Hickleton 46…………………………………………………….……….136
Figure 4.6: The 20 species with highest read number assigned by MEGAN for sample
Wetwang Slack 185……………………………………………………...138
Figure 4.7 The 20 species with highest read number assigned by MEGAN for sample
Wetwang Slack 7………………………………………………………...140
Figure 4.8 The 20 species with highest read number assigned by MEGAN for sample
Wetwang Slack 8………………………………………….…………..…142
Figure 4.9: The electropherogram showing the DNA library size distribution of the
sample Wetwang Slack 185……………………………….….………….145
Figure 5.1: Locations of the sites from which skeletal samples were obtained….…..159

9
Figure S5.1: Skeleton C21…………………………………………………………178
Figure S5.2: Skeleton C35…………………………………………………………178
Figure S5.3: Skeleton C48…………………………………………………………179
Figure S5.4: Skeleton C227………………………………………………………..179
Figure S5.5: Skeleton R5046………………………………………………………180
Figure S5.6: Skeleton R5256………………………………………………………181
Figure S5.7: Skeleton H3726………………………………………………………182

10
List of Abbreviations
A. mirum – Actinosynnema mirum
A. sulfonivorans - Arthrobacter sulfonivorans
AD – Anno Domini
aDNA – Ancient DNA
AMS – Accelerator mass spectrometry
ATP – Adenosine 5’-triphosphate
BB – Borderline borderline
BC – Before Christ
BCE – Before common era
BL leprosy – Borderline lepromatous leprosy
BLAST – Basic local alignment search tools
BQSR – Base quality score recalibration
BSA – Bovine serum albumin
BT leprosy – Borderline tuberculoid leprosy
BWA – Burrows-Wheeler aligner
C. woesei –- Conexibacter woesei
CD4+ – Cluster of differentiation 4
CD8+ – Cluster of differentiation 8
CDC – Centres for disease control and prevention
DNA – Deoxyribonucleic acid
dNTPs – Deoxyribonucleotide triphosphate
dsDNA – Double stranded deoxyribonucleic acid
E. coli – Escherichia coli
EB buffer – Elution buffer
EDTA – Ethylenediaminetetraacetic acid
F. Johnsoniae – Flavobacteria Johnsoniae
GATK – Genome analysis toolkit
GTCF – Genomic Technologies Core Facility
gyrA –DNA gyrase subunit A
H. sapiens – Homo sapiens
HIV – Human immunodeficiency virus
IFN-γ – Interferon gamma
IL-10 – Interleukin 10
IL-2 – Interleukin 2
IL-4 – Interleukin 4
IL-5 – Interleukin 5
IL-12 – Interleukin 12
IS1081 – Insertion sequence 1081
IS6110 – Insertion element 6110
K. flavida – Kribella flavida
K. setae – Kitasatospora setae
katG – Catalase peroxidase
K. albida – Kutzneria albida
LCA – Lowest common ancestor
LSPs – Large sequence polymorphisms

11
M. africanum – Mycobacterium africanum
M. bovis – Mycobacterium bovis
M. caprae – Mycobacterium caprae
M. fascicularis – Niastella koreensis
M. microti – Mycobacterium microti
M. pinnipedii – Mycobacterium pinnipedii
M. tuberculosis – Mycobacterium tuberculosis
MDR-TB – Multi-drug resistance tuberculosis
MEGAN – MEtaGenome Analyzer
MIRU – microsatellites or microbial interspersed repetitive units
MOTT – Mycobacterium other than tuberculosis
MPC – Magnetic particle collector
MRCA – Most recent common ancestor
MTBC – Mycobacterium tuberculosis complex
N. dassonvillei – Nocardiopsis dassonvillei
N. dassonvillei – Nocardiopsis dassonvillei
N. dokdonensis – Nocardioides dokdonensis
N. moscoviensis – Nitrospora moscoviensis
NERC – National Environment Research Council
NGS – Next generation sequencing
PCR – Polymerase chain reaction
PGG – Principal genetic group
Pks 15/1 – Polyketide synthase 15/1
qPCR – Quantitative polymerase chain reaction
R. tataouinensis – Ramlibacter tataouinensis
RD2 – Region of difference 2
RD7 – Region of difference 7
RLEP – Mycobacterium leprae repetitive element
S. amylolyticus – Sandaracinus amylolyticus
S. avermitilis – Streptomyces avermitilis
S. bingchenggenesis – Streptomyces bingchenggenesis
S. cellulosum – Sorangium cellulosum
S. denitrificans – Steroidobacter denitrificans
S. espanaensis – Saccharothrix espanaensis
S. fulvissmus – Streptomyces fulvissmus
S. hindustanus – Streptoalloteichus hindustanus
S. laurentii – Streptomyces laurentii
S. roseum – Streptosporangium roseum
S. venezuelae – Streptomyces venezuelae
S. violaceusniger – Streptomyces violaceusniger
SAM – Sequence alignment mapping
SNPs – Single nucleotide polymorphisms
SPRI – Solid phase reverse immobilization
T. actinomycetes – Thermophilic actinomycetes
T. bispora – Thermobispora bispora
T. curvata – Thermomonospora curvata

12
TbD1 – M. tuberculosis specific deletion
Th1 – Helper T cell type 1
Th2 – Helper T cell type 2
TLR – Toll-like receptors
UV – Ultraviolet
V. paradoxus – Variovorax paradoxus
VNTRs – Variable number tandem repeats
WGS – Whole genome sequencing
WHO – World Health Organization
XDR-TB – Extensively drug-resistant tuberculosis

13
Abstract
The overall aim of this study was to employ a biomolecular technique – ancient DNA
(aDNA) – to study two ancient diseases that were endemic in Europe (and therefore
Britain) during the medieval period: tuberculosis and leprosy. In humans, the diseases
are caused by M. tuberculosis and M. leprae, respectively – both of which are members
of the M. tuberculosis complex (MTBC). Skeletal manifestations of both diseases may
develop in bone remains, which can be recognized using osteological analysis. In some
cases, however, the skeletal changes are ambiguous. Ancient DNA methods are used for
case confirmation and to answer historical questions such as the spread, origin and
evolution of disease. The first objective of this thesis was to determine whether the
MTBC aDNA detection frequency is high enough to plan a larger study to test
hypotheses such as possible strain differences in urban and rural areas, as it has been
suggested that urbanization assists the spread of tuberculosis, enhancing its virulence. To
meet this objective, 60 skeletal remains from 16 different locations in Yorkshire,
England were studied. All samples were screened for MTBC aDNA presence and 8
samples were selected for next-generation sequencing (NGS). In the PCR assay
screening, only 1 sample produced a positive MTBC amplification. However, when
subjected to NGS, this sample together with the other 7 samples did not produce enough
sequence reads to allow genome comparisons. An attempt to compare metagenomic
content between urban and rural sites was also performed. There was no specific
difference in metagenomic content between urban and rural samples. Based on the PCR
analysis, the sample St Andrew Fishergate 6, dated to the early 14th century AD, showed
evidence of possible tuberculosis infection. NGS analysis further revealed a possible M.
tuberculosis and M. leprae mixed infection, albeit with insufficient read coverage to
determine genome sequence polymorphism. The second objective was to use NGS to
determine the genotype of the M. leprae strains present in skeletons from two mediaeval
sites, at Chichester and Raund Furnells, both in England. This study served as a
continuation for the previous confirmation by PCR of leprosy in these skeletons. The
samples were further subjected to whole M. leprae genome target enrichment before
subsequent high-throughput sequencing. For all 3 historical M. leprae isolates, at least
70% genome sequence coverage was obtained, with a mean read depth of 4-10x. The
near-complete genome sequences that were obtained allowed subtype identification for
each of the ancient M. leprae isolates. Two mediaeval samples from Chichester
belonged to the 3I subtype, which is typical of ancient Northern European and
contemporary North American isolates. Meanwhile, an M. leprae isolate from Raunds
was identified as belonging to the 3K subtype – the first example of this subtype
identified in Britain. Transmission of the M. leprae 3K subtype to Britain is suggested to
have been associated with the travels of crusaders and pilgrims to the Holy Land during
the mediaeval period. The overall conclusion of the work is that although M. leprae
aDNA is well preserved in skeletal remains showing osteological signs of leprosy, the
same is not true for MTBC preservation in skeletons showing indications of
tuberculosis. To test hypotheses such as the effect of urbanisation on tuberculosis, a high
frequency of MTBC detection must be achieved, but this is complicated by the very
nature of ancient DNA itself – highly fragmented, low endogenous DNA copy, presence
of environmental contaminants – and by the possibility of low bacterial load in skeletons
at the time of death. In projects where the testing of a high number of samples is
required, more stringent selection criteria must be imposed to minimize the impact of
destructive analysis.

14
Declaration
No portion of the work referred to in the thesis has been submitted in support of an
application for another degree or qualification of this or any other university or other
institute of learning
Copyright
i. The author of this thesis (including any appendices and/or schedules to this
thesis) owns certain copyright or related rights in it (the “Copyright”) and
s/he has given The University of Manchester certain rights to use such
Copyright, including for administrative purposes.
ii. Copies of this thesis, either in full or in extracts and whether in hard or
electronic copy, may be made only in accordance with the Copyright,
Designs and Patents Act 1988 (as amended) and regulations issued under it
or, where appropriate, in accordance with licensing agreements which the
University has from time to time. This page must form part of any such
copies made.
iii. The ownership of certain Copyright, patents, designs, trademarks and other
intellectual property (the “Intellectual Property”) and any reproductions of
copyright works in the thesis, for example graphs and tables
(“Reproductions”), which may be described in this thesis, may not be owned
by the author and may be owned by third parties. Such Intellectual Property
and Reproductions cannot and must not be made available for use without the
prior written permission of the owner(s) of the relevant Intellectual Property
and/or Reproductions.
iv. Further information on the conditions under which disclosure, publication
and commercialisation of this thesis, the Copyright and any Intellectual
Property and/or Reproductions described in it may take place is available in
the University IP Policy (see
http://documents.manchester.ac.uk/DocuInfo.aspx?DocID=24420), in any
relevant Thesis restriction declarations deposited in the University Library,
The University Library’s regulations (see
http://www.library.manchester.ac.uk/about/regulations/) and in the
University’s policy on Presentation of Theses.

15
Dedication and Acknowledgement
This work is dedicated to my parents, Kerudin Masintai and Rusinang Sibul who
inspired and taught me that nothing is more precious than education in life. You both
inspired me, prayed for me and supported me in a way that nobody could ever have done
and most importantly, it is your love that kept me going.
There are so many people whom I would like to thank – those who made my PhD
journey possible, bearable and enjoyable even – at times.
First, I submit my heartiest gratitude to my respected supervisor, Emeritus Prof Terry
Brown for his guidance, constant support and most of all his positivity and trust in me. I
could not have asked for a better supervisor. I would also like to thank my advisors, Dr
Richard Preziosi and Dr Russell Garwood for their guidance and support.
Secondly, I would like to show my appreciation to MARA who funded my PhD study –
which made it possible for me to pursue my PhD studies.
And to my family: brothers – Halley and Frankie, sister in laws, nephews and nieces; my
extended family and my family in law, thank you for your unwavering love, support and
encouragement which made it possible for me to go through some rough days along the
way.
To everyone in Brown’s group – thank you for your encouragement and support; for
treating me warmly for the last four years. Especially to Konstantina, Jannine, and Romy
who trained me in the lab; introduced and taught me about the “ancient DNA world”.
And to Kamalliawati for her lending ears, her house door that is always open for me and
for her constant encouragement. Most of all, thank you for making my thesis submission
possible – I wouldn’t have made it without you.
And to wrap this up, my biggest thanks to my husband, Benedict who have been
supportive from day one and for putting up with my rants and my emotional days. Most
of all, thank you for your patience throughout the years.
To my dear Estelle, you are my source of strength.

16
Chapter 1: Introduction
The history of mankind has been intertwined with infectious diseases. Tuberculosis and
leprosy, which are both contagious illnesses of ancient origins, have greatly affected the
course of world history. These diseases are still massive threats to human health to this
date, albeit their centuries of existence and the availability of treatments (World Health
Organization 2018a; World Health Organization 2018b). Their persistence in human
populations attracted studies from a wide range of fields. In sync to the notion: “the past
informs the present” (Brown & Barnes 2015, p.144), in the ever-growing discipline of
palaeopathology, ancient DNA is appropriately employed to answer historical questions
about palaeodiseases (Monot et al. 2005; Bos et al. 2014; Kay et al. 2015; Schuenemann
et al. 2018). The first isolation and detection of ancient DNA, from a dried muscle of
extinct Equus quagga in 1984, immediately ignited interest in the vast potential of this
tool in archaeology (Higuchi et al. 1984). Less than a decade later, Spigelman and
Lemma successfully isolated and detected the first microbial pathogen ancient DNA: M.
tuberculosis through the polymerase chain reaction (PCR) (Spigelman & Lemma 1993).
Since then, the study of ancient DNA in palaeopathology has not just been limited to
disease confirmation but also applied in answering broader and deeper questions, for
example, spread of disease through phylogeography and evolution of disease (Bos et al.
2014; Donoghue et al. 2015; Kay et al. 2015; Schuenemann et al. 2018).
1.1 Aims and Objectives
The overall aim of this study is to employ a biomolecular technique – ancient DNA – to
study two ancient diseases that were endemic in Europe (and therefore Britain) during
the medieval period: tuberculosis and leprosy. This chapter will focus on the background
of both ancient diseases studied: tuberculosis and leprosy. The biology of infection will
be described, followed by the manifestation of each disease in skeletons, which allows
the identification of the diseases in archaeological skeletons. The ability of tuberculosis
and leprosy to leave “fingerprint” changes in bones allows the identification of each
disease in skeletal remains using osteological methods. The limitation of this method
will be explained, which will then bring the ancient DNA method in to the picture. This
will be followed by the description of ancient DNA. The next section will describe the
ancient DNA study that has been done for both tuberculosis and leprosy to this date,

17
which includes case confirmation and evolutionary and disease origin studies. Chapter 2
describes the materials and methods performed as well as their relevance for this study.
The research described in the thesis had two objectives. The first objective was to
determine if the success rate of M. tuberculosis complex aDNA detection is high enough
to make it worthwhile to plan larger projects to test hypotheses such as possible strain
differences in urban and rural areas, as it has been suggested that urbanization assists the
spread of tuberculosis, enhancing its virulence (Comas & Gagneux 2011). Therefore, in
order to explore the association of urbanization and M. tuberculosis genotypes,
archaeological bones from urban and rural locations in Yorkshire, the majority with
tuberculosis indicative lesions which have been diagnosed by previous various
osteological studies, were examined. Yorkshire was selected because York was the
second largest city after London, during the 14th-15th centuries AD, and underwent rapid
urbanization during this time. The study of bones with tuberculosis lesions from
Yorkshire is split into 2 chapters: Chapter 3 and Chapter 4. The work described in
Chapter 3 aimed to screen all bone remains for the preservation of MTBC aDNA
through use of PCR assays of four targets selected based on their specificity for the
MTBC genome. The presence of MTBC aDNA is verified by cloning of PCR bands and
subsequent Sanger sequencing and matching the obtained sequences to the reference M.
tuberculosis H37Rv sequence. Based on the results obtained from Chapter 3, bone
remains were selected for next generation sequencing (NGS) using two sequencing
strategies, shotgun and target enrichment, which is described in Chapter 4. In Chapter 4,
the sequences obtained were then evaluated by various bioinformatic methods.
The work described in Chapters 3 and 4 showed that there is not extensive preservation
of MTBC aDNA in the bones from Yorkshire that were studied. The project was
therefore extended to include one additional objective. This additional objective aimed
to use NGS to determine the genotype of the M. leprae strains present in bones from
Chichester and Raund Furnells, both in England. This study serves as a continuation for
the previous confirmation by PCR of leprosy from Chichester and Raund Furnells, both
in England (Müller 2008). This work is described in Chapter 5. Leprosy was endemic in
Europe before it declined during the 16th century for unknown reasons, remaining only
in certain parts of Europe, human leprosy completely vanishing from Britain. Therefore,
archaeological remains are the only source of information on leprosy in Britain.
Identifying the genotypes of the M. leprae strains detected in the bones from Chichester
and Raund Furnells, as well as identifying new polymorphisms from each sequence in

18
comparison with the modern M. leprae TN strain (the reference genome that is used in
comparisons of M. leprae strain variability), will give an indication of the diversity of M.
leprae strains present in Britain during the medieval period, as well as giving clues about
the spread of leprosy to this part of the world. Finally, Chapter 6 describes the
conclusion of the works presented in this thesis.
1.2 The biology of tuberculosis and leprosy
Tuberculosis is one of the ten most common causes of global death (World Health
Organization 2018a). The World Health Organization (WHO) reported 1.7 million
fatalities caused by this disease alone, while there were 10.4 million people who
contracted the disease in 2016. Tuberculosis is also the leading cause of death among
HIV patients, accounting for 40% death in the same year (World Health Organization
2018a). This chronic granulomatous disease is caused by an obligate bacterium in
humans and animals (Brites & Gagneux 2015). There are seven closely related species
of Mycobacterium known with the ability of cause this disease: M. tuberculosis, M.
bovis, M. africanum, M. canettii, M. caprae, M. pinnipedii and M. microti (Wirth et al.
2008; Homolka et al. 2012). These pathogens are together known as the group members
of Mycobacterium tuberculosis complex (MTBC). The majority of infections in humans
and animals are caused by M. tuberculosis and M. bovis respectively (Bouwman et al.
2012). M. tuberculosis is transmitted through airborne droplets containing the pathogen
bacilli from an individual with active pulmonary tuberculosis infection, in which the
lung will be the primary site of infection (CDC 2013). On the other hand, the
transmission of M. bovis usually occurs through ingestion of contaminated milk and
meat, typically from cattle, where the gut will in turn become the main site of infection
(Atkins 2000; Waddington 2006). This is no longer a significant route of transmission
today, but bovine tuberculosis was a major problem in Britain from 1850 to 1950 where
it was responsible for approximately 800,000 deaths in the population (Atkins 2000;
Atkins 2008).
Tuberculosis bacilli are transmitted through droplet nuclei which will traverse the
mucociliary system and reach the alveoli of the lungs (CDC 2013). In this process, the
first line of defence would be the mucus-secreting goblet cells in the upper part of the
airways where the bacilli will be trapped on the mucus and subsequently “removed”
upward in accordance to the coordination by the cilia present on the cell surfaces

19
(Knechel 2009). The “unfiltered” bacteria will have the opportunity to invade the
alveolar spaces before they are subsequently phagocytized by the alveolar macrophages
(Pieters 2008). Following ingestion by macrophages, the mycobacteria will start to
multiply at a very slow rate (Knechel 2009). The macrophages will attempt to destroy
the bacteria by releasing cytokines and proteolytic enzymes (Flesch & Kaufmann 1993).
This cytokine production will trigger immune response from T cells through the
recognition of the mycobacteria antigen presented on the surface of macrophages. The
accumulation of immune cells including T cells, B cells and macrophages will aid the
formation of granulomas surrounding the M. tuberculosis, in an individual with good
cell-mediated immunity (Figure 1.1-a) (Pai et al. 2016). This will slow the replication of
the mycobacteria and prevents their spread. The micro-environment will contain the
bacteria but in the majority of cases, a complete eradication of the pathogen will not
occur but instead, they will adapt and survive (Forrellad et al. 2013). The low pH and
oxygen level conditions in the micro-environment will promote latency where the bacilli
survive within the granulomas but do not establish active infection to the host until the
immune system is compromised (Wayne & Hayes 1996; Cardona & Ruiz-Manzano
2004). The bacilli can stay in dormant stage, encapsulated in the calcified lesion, for
years, and even for a lifetime (Lin & Flynn 2010). In contrast, in hosts with a weaker
immune system, the infection will not be contained, therefore primary progressive
tuberculosis infection will be established (Figure 1.1-b) (Pai et al. 2016). The risks of
acquiring active tuberculosis infection is higher in HIV and diabetes patients,
malnourished people, as well as those who consume tobacco, with 8% of the cases
worldwide associated with smoking (World Health Organization 2018a). More than 95%
of tuberculosis cases and fatalities occur in the developing world; 45% and 25% of the
new cases are in Asia and Africa, respectively (World Health Organization 2018a).
Tuberculosis infection that occurs in the lungs is termed pulmonary tuberculosis
(Campbell & Bah-Sow 2006). The symptoms of pulmonary tuberculosis include cough
and sputum production, difficulties in breathing, haemoptysis, gradual wright loss, fever,
wasting, malaise, and anorexia in some cases (Campbell & Bah-Sow 2006). The typical
diagnostic methods for pulmonary tuberculosis include radiologic study, sputum direct
microscopy and culture and molecular detection (Ryu 2015). Although the lungs are the
primary site of tuberculosis infection, the bacilli can escape the lungs and spread to the
other parts of the body through the bloodstream and the lymphatic system (Ramirez-
Lapausa et al. 2015). Extrapulmonary tuberculosis can affect any part of the body but the
typical sites of infection are the lymph node, pleura, bones and joints or other skeletal

20
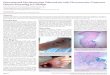
Figure 1.1: The mechanism of tuberculosis infection in cells and granulomas formation in active and latent tuberculosis infection. (a)
Inhaled tubercle bacilli enter the alveolar space of the lungs and bacteria elimination will be attempted by alveolar macrophages. In case of
unsuccessful bacteria elimination, the pathogens will invade the interstitial tissue of the lung. T cell priming will be initiated through the action of
either dendritic cells or monocytes – transporting the bacilli to pulmonary lymph nodes. This event will “attract” immune cells such as T cells and
B cells to the lung parenchyma leading to a subsequent granuloma formation. (b) M. tuberculosis replication occurs within the granuloma,
leading to high bacterial load. Failure of infection containment leads to bacteria dissemination to body organs. At this stage, the bacilli have the
ability to spread through the bloodstream or re-enter the respiratory tract to initiate similar immune reactions. In this infection phase, the host now
has active tuberculosis – will show symptoms and with the ability to infect other individuals (figure taken from Pai et al. 2016: 5).

21
parts, central nervous system, genitourinary system and abdomen (Golden & Vikram
2005). Extrapulmonary tuberculosis most frequently occurs in the lymph node and the
most frequent presentation is cervical lymphadenopathy but the involvement of the
inguinal, axillary, mesenteric, mediastinal and intramammary regions are also possible
(Golden & Vikram 2005). Approximately 35% of extrapulmonary tuberculosis cases
affect the skeletons; frequently spondylitis, followed by tuberculosis arthritis
concentrated on the joints (weight-bearing) and extraspinal manifestation (Golden &
Vikram 2005; Vanhoenacker et al. 2009). This type of tuberculosis involvement will be
discussed further in the next section. The regime treatment of a supposedly drug-
susceptible tuberculosis patient is a multi-drug combination consisting of rifampin,
isoniazid, pyrazinamide and ethambutol (Horsburgh et al. 2015). The inclusion of
ethambutol is not desired especially in young children due to its toxicity; it is usually
omitted in the treatment once drug-susceptibility is confirmed. The standard duration of
treatment for drug-susceptible patients is six months (Horsburgh et al. 2015). The multi-
drug combination regimen has been quite successful in treating tuberculosis patients; 53
million deaths were prevented between years 2000 and 2016 (World Health
Organization 2018a). However, the emergence of multi-drug resistance tuberculosis
(MDR-TB) and extensively drug-resistant tuberculosis (XDR-TB) halted the progression
towards zero tuberculosis cases (World Health Organization 2018a; World Health
Organization no date). According to the World Health Organization, there were
approximately 490,000 people with MDR-TB, with 600,000 newly reported cases of
rifampin drug resistance (World Health Organization 2018a). The treatment for
multidrug-resistant (MDR) tuberculosis is often complex and must be adjusted
specifically to the individuals by expert physicians according to drug-susceptibility
results performed using culture or DNA testing methods (Lange et al. 2014). Due to the
emergence of MDR-TB and XDR-TB, it is apparent that discoveries of new drugs and
vaccines are crucial. In addition, an effective public health system is also needed in order
to keep tuberculosis spread in check (Russell et al. 2010)

22
The second ancient disease of interest in this study is leprosy. This chronic infectious
disease causes manifestation on the skin, upper respiratory tract, peripheral nerves and
sometimes the eyes (Yassin et al. 1975; Lastória & de Abreu 2014). According to the
data gathered from 145 countries, there were 173,358 leprosy sufferers and 216,108 new
cases reported in 2016 alone (World Health Organization 2018b). Prior to 2008, M.
leprae was the sole leprosy pathogen known, before the identification of the second
leprosy causative pathogen: Mycobacterium lepromatosis (Han et al. 2008). Apart from
the majority infection observed in humans, leprosy infection has also been observed in
nine-banded armadillos in southern United States, chimpanzees, sootey mangabey
monkeys, and British Isles red squirrels (Meyers et al. 1991; Truman et al. 2011; Avanzi
et al. 2016). M. leprae is an extremely slow growing pathogen with a doubling time of
12.5 days (Jacobson & Krahenbuhl 1999). Similar to M. tuberculosis, this pathogen is
also an obligate intracellular bacterium (Groathouse et al. 2006). The study of M. leprae
is made difficult due to inability of its cultivation in normal cell culture media (Davis et
al. 2013). In vivo, leprosy has been studied using mouse foot-pad inoculation technique
and on immunocompromised mice, but neither systems are as successful as the
armadillo as an animal model for leprosy (Shepard 1960; Rees 1966; Kirchheimer &
Storrs 1971; Kirchheimer et al. 1972).
Although the precise path of M. leprae transmission is yet to be proven, an
overwhelming number of studies suggest that leprosy infection may spread through
nasal discharge containing M. leprae bacilli from untreated leprosy patients (Rees &
McDougall 1977; de Wit et al. 1993; Martinez et al. 2011). The bacilli will then make
their way into the healthy individual’s body through the respiratory route. Another
possible way for the bacilli to gain entry is through skin contact with a leprosy patient
(Satapathy et al. 2005). The bacilli enter the healthy individual’s body, possibly through
the skin and nose, and the host innate immune system will recognize the “foreign body”
and be triggered. This occurs through recognition of the mycobacterial lipoproteins by
the Toll-like receptors (TLRs) on macrophage and monocyte surfaces (Walker &
Lockwood 2006). In the case of M. leprae infection, recognition by the TLR2/1
heterodimer will allow the differentiation of monocytes into both dendritic cells and
macrophages (Modlin 2010). This will be followed by naïve T cell activation through
antigen presentation by dendritic cells to trigger more Th1 response (Walker &
Lockwood 2006).

23
Rather than M. leprae virulence, the clinical manifestation of this disease is determined
by the immune status of the individuals affected (Jin et al. 2018). One of the
classifications of leprosy discriminates between tuberculoid and lepromatous leprosy, as
well as the borderline intermediate infection, based on the resistance of the patient
towards the infection (Ridley & Jopling 1966).
An individual with stronger immune status will be able to “fight” the infection and will
only manifest the “mild” tuberculoid leprosy infection (Jacobson & Krahenbuhl 1999).
In this case, intense phagocytic activity will be triggered in lesions through the
secretions from Th1 T-cells (IL-2, lymphotoxin-α and interferon (IFN)-γ) (Walker &
Lockwood 2006; Bobosha et al. 2014). The cytokine-triggered macrophages will form
granulomas together with lymphocytes (Wang, Maeda, et al. 2013). Inside the
granulomas, T cells will produce granulysin, an antimicrobial protein (Walker &
Lockwood 2006). For these individuals, the CD4+ cells will outnumber the CD8+ cells
(Modlin 1994). Therefore, the infection will be halted by “trapping” the M. leprae inside
well-formed granulomas.
In contrast to tuberculoid leprosy, the granuloma is not so well-formed in lepromatous
leprosy, thus, making it difficult to “trap” the M. leprae bacilli (Wang, Maeda, et al.
2013). This is due to the different types of cytokines produced: IL-4, IL-5 and IL-10
(Yamamura et al. 1991). The cytokines IL-4 and IL-10 are capable of downregulating
TLR2 on monocytes while the latter will inhibit IL-12 production (secretion of which
will allow the activation of naïve T cells) (Zumla & James 1996; Walker & Lockwood
2006). The Th2 immune response will be activated while suppressing that of Th1, which
is responsible for initiation of the macrophage response (Modlin 1994). This type of
leprosy infection is characterized by the absence of granulomas and lack of successful
cell-mediated immunity.
The third type or degree of leprosy infection is intermediate or borderline infection
(Ridley & Jopling 1966). For this spectrum, the immunology response can change from
tuberculoid to lepromatous (White & Franco-Paredes 2015). Three types of borderline
leprosy infections are: borderline borderline (BB), borderline lepromatous (BL) and
borderline tuberculoid (BT) (Ridley & Jopling 1966). The borderline tuberculoid
spectrum of infections is the most common type of leprosy, although the exact
immunology is still not well-understood (Ankad 2018).

24
The clinical representation of leprosy depends on the strength of the host’s immune
system to fight M. leprae infection (Jin et al. 2018). In tuberculoid leprosy, there are
very few lesions found (Zumla & James 1996). The lesions are usually plaques and
macules with sharply demarcated edges (Lynnerup & Boldsen 2012). The appearance of
the lesions is usually hairless and hypoesthetic, which is due to the damage on the
dermal nerve fibres (Gunatilake & Settinayake 2004). The spread of infection to the
peripheral nerves such as the tibia, fibula and ulna is possible, however the damage is
limited. The effects on the nerves of a tuberculoid leprosy patient may include noticeable
enlargement of the area where the peripheral nerves are affected (Gunatilake &
Settinayake 2004). This is due to the granulomatous inflammation on the nerves which
in turn cause sensory and motor loss, resulting in the inability to detect pain (Walker &
Lockwood 2006). The swelling nerves could also experience further impairment as a
result of the entrapment inside the fibro-osseous tunnels.
In lepromatous leprosy, the early signs of disease include skin changes in the form of
macules distributed uniformly and widely on the face predominantly, and some are also
distributed on the upper limbs of the body (Lynnerup & Boldsen 2012). The edges of
these macules are almost undistinguishable and usually coupled with hypopigmentation
and redness on the skin (Gunatilake & Settinayake 2004; Reibel et al. 2015). In addition,
the face skin may thicken when the disease is neglected and untreated, and this results in
an appearance known as “leonine face” (Walker & Lockwood 2006). Dermal
involvement is pronounced in leprosy patients in this pole, which may give rise to
“glove and stocking” neuropathy (Sabin & Ebner 1969). The peripheral nerves may be
affected at a later stage (Lynnerup & Boldsen 2012). Skin lesions characteristic of
patients with borderline leprosy have characteristics in between those of tuberculoid and
lepromatous leprosy.
M. leprae shows a strong tropism in macrophages and Schwann’s cells of the peripheral
nervous systems (Reibel et al. 2015). Inside the cells, the pathogen bacilli will actively
proliferate, causing cell deterioration and failure to regenerate (Spierings et al. 2000).
Untreated leprosy nerve involvement will eventually result in deformity and disability,
sometimes irreversible (Lastória & de Abreu 2014). This occurs through sensory loss as
a result of nerve damage which in turn causes unnoticed injury, which leads to
secondary infections, causing damages in tissues (White & Franco-Paredes 2015). The
patient will eventually become disabled from the loss of motor function.

25
Multidrug combination regimen was introduced in 1982 which includes rifampicin,
clofazimine and dapsone (Jacobson & Krahenbuhl 1999; Reibel et al. 2015). The
duration of the course of treatment is still in debate; WHO has fixed the duration for 12
months, however many still argue that the duration is not sufficient for the infection to
resolve (Kumar et al. 2013; Malathi & Thappa 2013).
1.3 Skeletal changes in tuberculosis and leprosy
The field of palaeopathology has traditionally relied heavily on macroscopic inspection
to recognize ancient diseases in bone remains. Macroscopic or visual examination
provide direct evidence of such diagnosis in skeletal remains. It is crucial to understand
the effects of diseases in the human body, especially the skeleton, in a modern clinical
sense in order for the diagnosis to be made with skeletal remains (Roberts and
Manchester 2010). In bone remains, the osteological changes caused by infectious
diseases could be in the form of bone destructions, bone formation, or a mixture of both
(Roberts and Manchester 2010). Skeletal involvements are also common in modern
clinical samples, in the post-antibiotic era (Steyn et al. 2013; Steyn & Buskes 2016). The
lesion distribution on the skeleton is an important clue to identify the type of infection
the individual had suffered. Typically, palaeopathology researchers are performing the
osteological analysis using macroscopic and radiological methods but the histological
approach has become increasingly popular.
The skeletal changes in tuberculosis and leprosy are a manifestation of a chronic, long-
established infection in a relatively healthy individual (Zink et al. 2001; Tayles &
Buckley 2004; Adachi et al. 2006; Rubini, Zaio & Roberts 2014; Suzuki et al. 2014;
Steyn & Buskes 2016; Inskip et al. 2017). The tuberculosis bone changes occur in an
individual with a secondary tuberculosis infection, where the infection of a person with
latent tuberculosis is reactivated (Ortner 2003; Roberts & Manchester 2010). In
secondary tuberculosis infection, the tubercle bacilli spread haematogenously within the
bones to other parts of the body (Roberts & Manchester 2010). The bacilli will
predominantly reside in the skeletal areas where the circulatory and metabolic rates are
high inside the skeleton, e.g. in the haemopoietic or red marrow (Ortner 2003).
Regardless of the age of individuals, the red bone marrow can be found in all bones
including the vertebrae, sternum and ribs (Prabhakar et al. 2009).

26
Tuberculosis infection in skeletal remains can be recognized from bone destruction in
the spine, especially the lower thoracic and lumbar vertebrae (Roberts and Manchester
2010). In most cases, the bone changes are severe which involves abscess development
inside the vertebrae which in turn perforate into the abdomen or chest, and the affected
spine will finally collapse (Figure 1.2 – a) (Roberts & Manchester 2010; Holloway et al.
2013). In archaeological remains, result of the spinal collapse will cause an angular
deformity of the spine. This is the most common type of skeletal tuberculosis, which
accounts for 25-60% of infections (Roberts & Manchester 2010). Skeletal lesions on the
vertebrae are usually shown as osteolytic lesions on the frontal area of the lower thoracic
and the upper lumbar, in which the number of affected vertebrae is usually between one
to four (Holloway et al. 2011; Rasouli et al. 2012). Following the spine, the knee and hip
are the second part of the body that are frequently infected which make up to 10-20 to
15-30% of skeletal tuberculosis cases other than spine involvement, respectively (Ortner
2003; Roberts & Manchester 2010). The tuberculosis infections on these joints are
characterised by fibrous fixation also known as ankylosis (Roberts & Manchester 2010;
Saraf & Tuli 2015). However, great care has to be taken on confirming tuberculosis
diagnosis based on this evidence as it may also be caused by pyogenic osteomyelitis,
brucellosis and fungal infections (Holloway et al. 2011).
Other than that, there are other non-specific bone changes of tuberculosis. These include
new bone formation giving rise to rib lesions, which are increasingly common in post-
antibiotic era (Steyn & Buskes 2016). The tuberculosis lesions on the ribs could be the
result of chronic pulmonary tuberculosis (Figure 1.2 – b) (Kelley & Micozzi 1984;
Matos & Santos 2006). These lesions are typically subtle with the characteristics of
periosteal new bone formation as well as bone resorption (Roberts & Buikstra 2003).
The lesions usually result from bloodstream spread of bacilli, but can also be caused by
direct extension from neighbouring tuberculosis foci and extension from abscesses
within the paravertebral region (Ortner 2003; Roberts & Manchester 2010). New bone
formation is the result of the response of bone tissue to traumatic insults or pathological
infection. The bone infection will cause an inflammatory response that will trigger the
production of new bone on top of the periosteal bone surface, which sometimes can
result in a different colour to the bone ‘layer’ (Figure 1.2 -b). In 2003, Ortner
mentioned that rib tuberculous changes normally occur in 9% of tuberculosis sufferers
(Ortner 2003). In minor cases, dactylitis, which is toe or fingers inflammation, and skull
involvements are also characteristics used for tuberculosis diagnosis (Holloway et al.
2011; Steyn & Buskes 2016). Although these non-specific bone changes have been

27
Figure 1.2: The bone changes indicative of tuberculosis infection in skeletal remains.
(a) The destructive spinal lesions and collapse of vertebrae which is considered pathognomonic
for tuberculosis. The red arrow indicates where the vertebral bodies collapse occurred, which
results in spinal angular deformity in people suffering from spinal tuberculosis. (b) Rib bones
showing new bone formation, as shown by the yellow arrows. New bone formation is one of the
characteristics of probable tuberculosis infection in skeletal remains. (Images from Charlotte
Roberts’ personal collection, personal communication 2019).
(a)
(b)

28
considered more and more in future studies, especially the rib lesions, caution should be
taken as to never treat these as a definitive evidence of tuberculosis infection (Roberts
and Manchester 2010). The non-specific bone changes can also be caused by other
conditions. Endocranial new bone formation could also be caused by induced
meningitis. Similarly, rib lesions can also be attributed to other diseases such as
metastases, unspecific osteomyelitis, pneumonia, bronchiectasis and mycosis (Nicklish
et al. 2012).
Characterisation of bone changes in skeletal remains suspected to be infected by leprosy
was first undertaken by a Danish physician Vilhelm Møller-Christensen in 1953 (Møller-
Christensen 1953; Lynnerup & Boldsen 2012). Leprosy is a slow progressing, chronic
infection which explains its strong manifestation on the skeleton as written by Lynnerup
and Boldsen (2012), “leprosy is a disease one dies with rather than of”. Bone lesions of
leprosy infection in archaeological remains are recognizable on the skull, the extremities
of the hands and feet as well as the lower legs (Thappa et al. 1992; Roberts &
Manchester 2010; Mohammad et al. 2016). As previously mentioned, M. leprae bacilli
attack the peripheral nerves, causing functional loss (Gunatilake & Settinayake 2004).
This will subsequently cause deformities, with most affected regions being the upper and
lower limbs due to muscle paralysis on these regions. Consequently, anaesthesia will
occur on the hands and feet, thus allowing secondary infection to manifest, causing
tissue necrosis due to the unnoticed injuries. In turn, this will give rise to an appearance
of what is known as “clawed hands” (Ortner 2003; Gunawan et al. 2017). On the foot,
the loss of bones on the distal phalanges also may occur, giving rise to a noticeable
“pencil ends” configuration on the metatarsals (Figure 1.3) (Roberts & Manchester
2010). This appearance is associated with the bone atrophy that always takes place
starting from the distal margins of the hands or feet (Barnetson 1951). This bone atrophy
could then proceed to the proximal bones. For example, on the foot, bone absorption
may begin on the distal phalanx, and proceed to the proximal bones, resulting in the loss
of all or some of the phalanges. Further progress of the bone absorption could lead to the
pencilling of the metatarsals, as depicted in Figure 1.3. Although not as frequently as on
the foot, the same pattern of bone absorption can occur on the hand (Ankad et al. 2011).
The bone absorption that typically starts from the distal fingertip, will cause a defect that
looks like a V-shape or “sharpened” fingertip (Lu et al. 2015). Similarly, the worsening
and further progression of the bone absorption will eventually lead to the disappearance

29
of the distal phalanx or will leave behind irregular looking bone remains. The bone
absorption will progress to proximal bones.
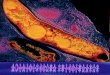
Figure 1.3: The bone changes of leprosy on the foot.
The image below shows the ‘pencilling’ appearance of the foot metatarsals and loss of some of
the foot phalanges. As a comparison, the image on top is showing the normal foot appearance in
a modern skeleton. The blue brackets are showing the metatarsal bones, while the red brackets
are showing the phalanges in both normal and affected bones. The yellow arrows depict the
affected bones where the loss of phalanges and pencilling of metatarsal bones has occurred.
(Image from Charlotte Roberts’ personal collection, personal communication, 2019).

30
On the skull, the most recognizable leprosy change is what is termed as rhinomaxillary
syndrome (Ortner 2003). This term was introduced by Andersen and Manchester (1992)
in place of ‘facies leprosa’ proposed by Møller-Christensen in 1978. The former is a
preferred term in palaeopathology as the latter is also used to describe the soft tissue
changes and therefore will not best fit the palaeopathology contexts (Andersen &
Manchester 1992). Rhinomaxillary syndrome is exclusively the result of lepromatous
and near-lepromatous leprosy, therefore the presence of all elements of the bone changes
is pathognomonic of these conditions (Andersen & Manchester 1992; Nerlich & Zink
2008). The tendency of M. leprae to invade the cooler regions of the face explains the
lesion distribution on the mucosal membranes and the cooler exposed skin area
(Andersen & Manchester 1992). The bony lesions in rhinomaxillary syndrome can be
observed in the nasal cavity, the maxilla, on the oral surface of the palatine and alveolar
processes, without any effect shown on the mandible. The absence of lesions in the
mandibular region is explained by the difference in temperature in the regions affected.
Perforation is also possible on the palate, accompanying the inflammatory pitting on
both sides of it (Roberts & Manchester 2010). Apart from that, incisor teeth loss also
may occur as a result of the alveolar bone loss on the upper jaw in that region of the
teeth. The effects on the nasal spine and nasal aperture will include absorption and
remodelling, respectively.
There are always limitations in any discipline, and this is particularly the case with
palaeopathology. This has been highlighted by Wood and colleagues (1992), who
described the “osteological paradox”, which summarises the uncertainties that arise
when it is attempted to use skeletal evidence to study the health and disease status of a
past population. The author highlighted three major problems in palaeopathology. These
are the non-fixed state of the population (so a single or small group of skeletons from
one point in time is not representative of a large population that might be undergoing
change due to migration and mixing with other populations), selective mortality (not all
people with a disease died because of that disease), and the individual variations in the
risks of contracting disease and death (so skeletons with palaeopathological lesions
might represent only a biased proportion of population as a whole). These are the
limitations that are considered impossible to overcome. Other limitations that should be
taken into consideration is that the ‘dead populations’ being studied cannot be
considered to be representative of the living populations (Roberts and Manchester 2010).
Besides, the diagnosis confirmation is heavily dependent on the state of preservation of

31
the skeletal remains. Furthermore, bone changes in acute diseases might be difficult to
detect as the individuals might have died quickly from the disease without having
enough time to develop bone changes.
The most challenging aspect of ancient disease diagnosis based on osteological
observation is the lack of bone lesions specific for certain diseases. The abnormal bone
lesions observed could be the result of different diseases, therefore affecting the
accuracy of the differential diagnosis (Klaus 2017). This is particularly apparent in the
diagnosis of tuberculosis as the non-specific bone lesions could also be a result of other
diseases as previously described in this section. Furthermore, even if there are
pathognomonic bone changes that can diagnose a disease correctly, it would still be
dangerous to use this information to infer the disease status of the population from which
the studied skeleton originated. This information would still suffer from the osteological
paradox. Therefore, to overcome this, it is useful to incorporate other methods such as
molecular study of a skeleton to identify the disease or diseases suffered by that
individual in the past. The biomolecular study of skeletal remains, specifically the use of
ancient DNA, will be further described in the following section 1.6. This section
explains how such studies may complement and add precision to palaeopathological
studies of both tuberculosis and leprosy.
1.4 History of tuberculosis and leprosy based on historical
documents and skeletal changes evidence
1.4.1 History of tuberculosis
Historical documents – written or pictorial – are the key sources which can indicate the
antiquity of certain diseases, albeit the interpretations should be made with caution. The
first convincing historical evidence of tuberculosis appeared in the Chinese literature,
documented by Emperor Shennong of China, dating to approximately 2700 BCE
(Tripathy 2015). This medical text mentioned “xulao bing” which can be interpreted as
“weak consumption”, thought to describe tuberculosis. Medical papyrus from Egypt
such as the Ebers Papyrus (1500 BCE) is the other well-known ancient document which
outline diseases and treatments affecting populations in that area during that period
(Khalil & Richa 2014). A few authors are convinced that this document might be

32
describing tuberculosis. However, Chalke (1962), in much earlier writing is convinced
otherwise. The same differing opinions are held regarding the Bible references about
tuberculosis. Daniel and Daniel (1999) suggested that two verses from the Old
Testament are describing tuberculosis, but Chalke has stated that the descriptions from
the Bible are ambiguous and there is not sufficient information to support the description
of tuberculosis such as is present today (Chalke 1962; Daniel & Daniel 1999).
Throughout different periods, tuberculosis has been called by many names. Tuberculosis
was known during the Classical Greece period as ‘phthisis’, according to the records in
Greek ancient literature, during the period of Hippocrates (460-370 BC) (Moonan 2018).
He was one of the most well-known Greek physicians, who stated in his records that
‘phthisis’ was the most typical disease where death is almost inevitable during that time.
In addition, Hippocrates also described the symptoms of ‘phthisis’ in his Book 1 ‘of the
Epidemics’ including fever, coughing accompanied by concentrated sputa, colourless
urine, and loss in desire to consume food and drinks (Firth 2014). His book also noted
that people who severely suffering from ‘phthisis’ were in the age range between 18 and
35. Hippocrates was in agreement with others who believed at this time that the nature of
‘phthisis’ transmission was hereditary (Lakhtakia 2013). However, the famous Greek
philosopher Aristotle believed that this disease is contagious rather than hereditary, in
contrary to other contemporaries during that period. After Hippocrates, another
renowned physician, Galen, described phthisis, emphasising the presence of “lung
ulcers”, throat or thorax, and body consumption by pus in addition to the other
symptoms already described by Hippocrates (Rosenthal 2013). Some treatment methods
were also outlined such as the use of opium to induce sleep and to numb pains,
bloodletting and recommended diets (Tripathy 2015).
In Europe, tuberculosis was endemic in the 17th century AD and continued to be so for
two centuries, which is why it was known as the “Great White Plague” (Barberis et al.
2017). During this period, tuberculosis was the leading cause of death and whoever
contracted the disease was considered to have received a death sentence, as fatality
occured in almost all instances of infection. The spread of the disease agent was
aggravated by the living conditions in Europe during that time, which were overcrowded
and unsanitary (Roberts & Manchester 2010; Tripathy 2015). In the 17th century AD, the
Bills of Mortality were available in London as a record of mortalities together with their
diagnosis – with tuberculosis being the most common cause of death (Matossian 1985).
Later, from the 19th century onwards, the Registrar General’s statistics provide a quite

33
accurate representation of the disease progress in both Wales and England, which show a
stable drop in the prevalence of tuberculosis in these regions except during the World
Wars (Davies et al. 1999). The 19th century AD was the period of tuberculosis
romanticism (Daniel 2006). During this time, tuberculosis claimed many lives among
the glamorous, talented, smart and good-looking young individuals in society. The
characteristics of tuberculosis sufferers, pale and thin, added to the “romantic” aspect of
the disease, which was considered stylish during that time. In fact, tuberculosis is
depicted in many arts. Portrait of individuals with hunched-back and thin, pale looking
young women were often depicted during this era (Roberts & Buikstra, 2003).
There are many historical documents available throughout the centuries about
tuberculosis. However, precaution should be taken in using or verifying this information.
The accuracy of the tuberculosis diagnosis is questionable; there are instances where the
disease was wrongly assigned (Davies 1998). Diseases with similar symptoms should be
taken into consideration when confirming these diagnoses. For example, non-
tuberculous pneumonia and bronchitis may exhibit similar symptoms to tuberculosis
(Ukil 1940). In addition, the credibility of the individuals who assessed the disease must
also be taken into consideration (Roberts & Buikstra, 2003).
Other than historical documents, the antiquity of tuberculosis can also be inferred
through osteological analysis as described in section 1.3. The earliest skeletal evidence
in different locations based on the study of skeletal changes is described here. The
earliest skeletal evidence of tuberculosis is a female skeleton showing spondylitis found
in the Neolithic level of Arma dell’Aquila Cave in Liguria, Italy (Canci et al. 1996). The
age of the skeleton (5800±90 years BP) was determined based on the 14C radiocarbon
dating of a neighbouring grave located on the same horizon. There is also evidence of
skeletal tuberculosis in the Old World at the Late Neolithic tell settlement site in the
South of Hungary dated 4932-4602 cal BC (Masson et al. 2015). The earliest skeletal
evidence in Britain was found at Tarrant Hinton, Dorset, at an Iron Age site (400-230
BC) (Mays & Taylor 2003). Evidence of tuberculosis in Asia comes from relatively
recent sites, from China, dated to the second century BC, Japan (454 BC – 124 AD) and
Thailand (300 BC – 300 AD) (Suzuki & Inoue 2007; Stone et al. 2009). Plenty of
tuberculosis evidence is available from this period onwards including skeletal evidence
found in France, Lithuania and Austria dated to the 4th century (Stone et al. 2009). In the
Americas, skeletal evidence for tuberculosis mounted throughout the 20th century AD
(Stone et al. 2009). In South America, one of the earliest convincing pieces of evidence

34
of tuberculosis from human remains is from the Andean populations of Peru and
Northern Chile (Allison et al. 1981). Meanwhile in North America, all the skeletons with
evidence of tuberculosis are dated after 1000 AD (Roberts & Buikstra 2003).
Confirmation of ancient tuberculosis cases based on additional biomolecular evidence
will be discussed further in section 1.6.1.1.
1.4.2 History of leprosy
Similar to tuberculosis, leprosy also has existed since antiquity. The earliest mentions of
this disease can be traced back from ancient Egypt (1550 BC), India (600 BC); and
China (4th century BC – 217 BC) (Leung 2009; Robbins et al. 2009). Many authors
believe that the Ebers Papyrus from 16th century Egypt includes a description of leprosy
infection (Robbins et al. 2009). However, a few scholars are sceptical, based on the
accuracy of the translation of this manuscript (Mark 2002). In fact, the notion was
previously rejected and faced strong criticism due to a lack of consistency between the
disease characteristics as described and the symptoms of modern leprosy as we know
today. It was later agreed that the disease description is more fitting to gas gangrene
compared to leprosy (Mark 2002). Sushruta Samitha (600 BC), an ancient Indian
medical document, provided descriptions of disease characteristics that almost all agree
with advanced lepromatous leprosy symptoms (Dharmendra 1947; Jacob & Franco-
Paredes 2008). These include sensation loss, ulceration and disfigurement of limbs,
collapsing of the nose, and falling off of fingers (Roberts & Manchester 2010). Sushruta
Samitha is the only ancient medical writing that gives the most precise description of
leprosy. These texts were estimated to be compiled around 600 BC but the writings are
believed to contain information which was acquired from a much earlier period
(Dharmendra 1947). There are other Indian ancient literatures that might have mentioned
this disease; these are the Laws of Manu and the Atharava Veda (Jacob & Franco-
Paredes 2008). However clinical descriptions of the disease are absent, therefore there is
no significance confidence to confirm that the disease described was leprosy
(Dharmendra 1947). A description of leprosy from China has been acquired from a 3rd
century bamboo book which outlined similar disease characteristics as described in the
Sushruta Samitha (Roberts & Manchester 2010). The presence of these ancient medical
documents with such articulate descriptions of the disease raises the possibility that
leprosy might have existed in the Far East and India from a very early period, as the

35
authors seem to have a deep clinical understanding and judgement of the disease
(Roberts & Manchester 2010). Based on the presence of such detailed and accurate
medical writing and the presence of clinically skilled individuals, it is quite possible that
leprosy was prevalent in these regions during that period of time (Roberts & Manchester
2010). It has also been suggested that leprosy is referred to in the Book of Leviticus, in
the mention of the word tzaraat, but this proposal was later dismissed as a
mistranslation, and it is now accepted that the tzaarat mentioned in the bible was a range
of skin diseases and not the leprosy that we know of today (Trautman 1984; Mark 2002;
Grzybowski & Nita 2016).
In Europe, the prevalence of leprosy infection peaked in the 11th to 14th centuries AD
(Mendum et al. 2014). Leprosy was feared and stigmatized due to the disability and
disfigurement that it caused as well as the lack of understanding of this disease during
that time (Bennett et al. 2008; Sermrittirong & Van Brakel 2014). Leprosy was seen as a
punishment from God and the patients were isolated in a specific area or “facility” called
a leposarium, to avoid contagion from the infected individuals (Donoghue et al. 2015).
During the Middle Ages in Europe, people who were suffering from leprosy infection
were required to wear distinguishing attire, and had to ring bells to let people know that
they were close by (Bennett et al. 2008). In addition, they were also expected be on the
side of the road where the wind does not blow from, as people were so afraid of this
disease and they shunned whoever was suffering from it (Hussain 2007). Leprosy, due to
its unpleasant outcome, was viewed as a blasphemy and even thought to be inherited.
Due to its dramatic representation and high prevalence throughout history, leprosy has
inspired many art works (Gron 1973). Leprosy has been one of the most misunderstood
diseases and still is in certain regions of the world (Sermrittirong & Van Brakel 2014).
Luckily, the discovery of the causative pathogen M. leprae in 1874 by Dr. Gerhard
Armauer Hansen of Norway was the most important moment in the history of leprosy as
it opened a pathway to the understanding of this disease (Irgens 1984; Trautman 1984;
Ghosh & Chaudhuri 2015).
The earliest convincing evidence of leprosy in human remains is from Balathal India,
dated to 2,000 BC (Robbins et al. 2009). This might support the speculation that the
Mediterranean leprosy brought to Europe by the armies of Alexander the Great
originated in the Indo-Gangetic basin, which was not supported by any skeletal evidence
before in the review by Stone and colleagues (2009). One of the earliest evidence of
leprosy in skeletal remains has also been observed at the Dakleh Oasis in Egypt (250

36
BC) (Dzierzykray-Rogalski 1980) and in Israel and Nubia (Stone et al. 2009). More
leprosy bone studies will be elaborated further in section 1.6.2.
1.4.3 The concern in using historical documents as evidence of ancient
disease
Historical texts are crucial in the process of deciphering diseases in historic populations
(Metcalfe, 2007). The writers who lived in the past can provide a clue about the culture
of societies during that time, which then can assist the palaeopathological analysis of
human remains. In some historical texts, the signs and symptoms of diseases are
recorded clearly, and therefore can provide a direct source of information. In
palaeopathological studies, the exclusion of historical written documents will pose extra
challenges to the research itself and might cause inaccuracy in the interpretation
(Mitchell 2017). However, it is also worth noting that cautions must be taken in
interpreting historical documents as evidence of diseases in the past.
The differences in belief and perspective during the historic period might put the study at
risk of misinterpretation (Mitchell 2011). As the ancient medical texts are derived from
various civilizations, it is expected that they were written according to the perspective of
the belief in that geographical area during that time (Roberts 1971). Consequently, this
might give rise to a different description that can be easily misinterpreted from the
context of what we know in this modern medical period. Therefore, it is important to
consider the time when the historical documents were written and the interpretation must
be made by someone with an expertise in the beliefs and perspective of the ancient
civilization (Mitchell 2017).
Furthermore, the evidence of disease must be obtained by reading the original version of
the historical documents (Mitchell 2011; Mitchell 2017). Modern translations or any
quotes from secondary sources are best to be avoided where possible. The concern with
using documents from modern translation is that the translator might not have a medical
background, therefore affecting the accuracy of the disease’s descriptions. Therefore, it
is highly desirable that palaeopathologists work hand in hand with medical historians
who are sufficiently versed in the linguistics to make sure that the original texts are
properly translated and interpreted (Mitchell 2017). In addition, we also cannot be sure
that the medical writers have witnessed the disease during their lifetime or simply just

37
copied it from the previous records of their predecessors. If this is the case, it will give a
wrong indication of the timeline of the disease – or if it really has existed in the
population during the time the document was written. The skills and competency of the
medical personnel who produced the documents cannot be verified. It could be someone
who never had any medical training. By considering all these concerns, there is a lot of
room for disease misinterpretation.
The nature of the written sources and artwork also must be taken into account when
making an interpretation about past diseases. In the investigation of ancient diseases, it is
sensible to start by looking at historical medical texts. When using such document, the
historian must consider when they were written, the person who wrote them and the
purpose for producing or writing such a document (Mitchell et al. 2017). Typically, such
texts are written and being presented to someone high up. Therefore, the writer might
have the intention of impressing this person in power in order to gain something in
return, for example lucrative posts or career advancement (McVaugh, 2006). It is
possible that the text being presented might have been subject to alteration in order to
impress whoever the text was being presented to. Meanwhile, non-medical texts are
often written by non-medical practitioners with only basic medical knowledge
(Robinson 2003). Therefore, although they may not have as much bias as the medical
texts, the disease descriptions that were written could be quite vague and following
personal observations on only the obvious and clearest symptoms and signs. In the case
of histories, the text usually covers a long time range, therefore some of the chronicles
might have been copied from oral tales or much older written sources (Mitchell 2017).
Other documents that can be the source of disease information are biographies, personal
letters and diaries. These types of document can be highly illuminating as sometimes the
diseases are described in detail – how they, their friends or families feel or experience
when they suffered from the disease. Births and deaths registers could also provide good
information about the severity of a disease in the past – the statistic of disease in the past
in a particular region (Wrigley et al. 1997). However, these could also be the subject of
disease misinterpretation and alteration, depending on the reason for why such document
is being produced.
In short, there are many historical documents available which recorded diseases in the
past and they are no doubt great sources of information. But caution must be taken when
making interpretations by thinking carefully about what the document means to the
writer and to whom the document is being presented, and when was the document

38
written; artworks included. When using historical documents, it is also important that the
person is frank about how confident he or she is about the accuracy of the interpretation
(Mitchell 2017). Some documents might only mention a few symptoms and some
symptoms described could be ambiguous and could be present in more than one disease.
Therefore, the choice of words is important and remarks about other possible diseases
should be made.
1.5 Ancient DNA
1.5.1 Ancient DNA background
Genetic information from archaeological biological remains can be preserved and stored
in the form of ancient DNA (aDNA), which then can be assessed and studied (Higuchi et
al. 1984; Spigelman & Lemma 1993). The ancient DNA research field began in 1984
with the isolation of DNA from a preserved Equus quagga skin; this is a zebra-like
animal which became extinct 100 years previously (Higuchi et al. 1984). This
publication provoked interest to study aDNA in much wider types of archaeological
specimens but this was initially hindered by the difficulties in analysing the extremely
low amounts of fragmented and degraded nucleic acid materials from ancient specimens
(Brown & Brown 1992). The development of the polymerase chain reaction (PCR) by
Kary Mullis in 1985 opened a new pathway to study ancient DNA more efficiently as it
allows rapid amplification of a single fragment of DNA, which is perfect for aDNA
which typically exists at a very low copy number (Mullis et al. 1986). This led to the
first application of PCR to amplify aDNA from a 7000-year-old brain (Pääbo et al.
1988). Fast forward to this date, aDNA has been extracted from various archaeological
samples and studied using the current state-of-the-art sequencing technologies (Green &
Speller 2017).
1.5.2 Characteristic of ancient DNA
The DNA molecules of an organism are in constant threat of chemical “attack” at all
times. Fortunately, this is countered by enzymatic repair mechanisms when the organism
is still living to protect its genome integrity (Dabney, Meyer & Pääbo 2013). However,
upon the death of the organism, the genome becomes exposed to various “destroying

39
agents” that may affect its stability such as intracellular nucleases. This is termed
autolysis (Brown & Brown 2011). This, together with two other degradation factors,
environmental effects and microbial attack, influence the state of aDNA preservation
(Burger et al. 1999; Turner-Walker 2008; Kistler et al. 2017). Those biomolecules that
manage to escape the degrading autolysis activity will stand a possibility of being
archaeologically preserved (Brown & Brown 2011). However, they will be subjected to
other less rapid but persistent decaying processes due to the environment, both physical
and chemical factors (Turner-Walker 2008). The chemical agents include oxygen and
water, which are highly reactive and possess the ability to promote oxidative and
hydrolytic reactions, respectively. In addition, the by-products released from the
degradation of one type of biomolecule might themselves trigger decay of another
biomolecular type (Brown & Brown 2011). The physical factors involved in
biomolecular decay include geological, ultraviolet (UV) and cosmic radiations, which
possess the ability to significantly affect the integrity of biomolecules in both living cells
and post-mortem (Cadet & Wagner 2013). However, it is possible to disrupt the
complete destruction of the DNA molecules under favourable circumstances, for
example by freezing or desiccation of specimens (Nicholls 2005). The third degradation
factor, microbial, is also important as several types of microbe are attracted towards
organic material contained in biological remains as this may provide nutrients and an
energy source. In order to “feed” on the organic materials, microbes will secrete certain
enzyme(s) whose function is to break down the organic materials, including DNA,
leading to their degradation (Turner-Walker 2008). The extent of microbial-facilitated
biomolecular destruction depends on the microflora population existing in the burial
environment. Due to the presence of these destructive factors and the absence of repair
mechanisms after death, typical features of ancient DNA are (i) short fragment size due
to degradation; (ii) the presence of DNA replication blocking lesions; and (iii) the
presence of miscoding lesions (Dabney, Meyer & Pääbo 2013), as described below.
1.5.2.1 Fragmentation
Desiccation or anoxic conditions are thought to be the most suitable for DNA
preservation as these exclude the presence of DNA-damaging water and oxygen
(Nicholls 2005; Brown & Barnes 2015). Water targets the β-N-glycosidic bond resulting
in the release of purine or pyrimidine bases from the nucleotide sugar component,

40
consequently creating apurinic or apyrimidinic sites (Kim & Wilson 2012). These sites
are vulnerable to chemical attack, which will then lead to DNA fragmentation, creating
short fragments (Dabney, Meyer & Pääbo 2013). According to the results of in vitro
studies with contemporary DNA specimens, it has been proposed that DNA
fragmentation is caused by hydrolytic depurination. Subsequently, single-strand breaks
will occur due to hydrolysis of the phosphate backbone. These studies provide
supporting evidence to correlate temperature to fragmentation rate (Lindahl &
Andersson 1972; Lindahl & Nyberg 1972). Accordingly, thermal history was proved to
be a useful determinant in predicting the state of biomolecular survival in fossil bones;
the assumption is that DNA depurination is the main determinant of DNA degradation
(Smith et al. 2003). During the preservation period, DNA molecules that are contained
in archaeological remains undergo fragmentation. In result, the aDNA molecules
acquired from archaeological specimens are relatively short (Pääbo 1989). The average
fragment length of aDNA isolated from dried animal muscle tissue is between 100 to
200 base pairs; the maximum fragment length is 500 base pairs (Pääbo 1989). Studies
have shown that the size of aDNA fragments or the degree of fragmentation is
independent of the age of the archaeological specimen (Pääbo 1989; Hagelberg & Clegg
1991; Kistler et al. 2017). Instead of a time factor, it seems that the rapidity of tissue
desiccation after the time of death is more likely to be the determinant of the degree of
DNA fragmentation (Pääbo 1989). The aDNA decay model established for mammal
bones proposed by Kistler and colleagues (2017) suggests that DNA fragmentation
reaches a threshold rapidly after death and then slows down. In contrast, for DNA loss
throughout the preservation period, the process of bulk diffusion out of the sample might
be the major factor. By saying this, this group emphasized the importance of creating
optimum conditions in a closed system in order to improve the DNA preservation.
1.5.2.2 Miscoding lesions
Miscoding lesions are typical of aDNA as a result of hydrolytic deamination of
nucleotide bases (Dabney, Meyer & Pääbo 2013). These will cause errors of “reading”
by DNA polymerase during amplification resulting in incorporation of incorrect
nucleotides during PCR. Cytosine (C) has the highest susceptibility towards
deamination, which will produce uracil (U). During DNA duplication with a strand
containing a miscoding lesion used as template, uracil will direct adenine (A)

41
incorporation that subsequently causes C to T or G to A transition in the amplified PCR
product. Hofreiter and colleagues demonstrated the presence of modified deoxycytidine
in aDNA extracts from animal teeth and bones with ages ranging between 25,000 to
50,000 years (Hofreiter et al. 2001). The treatment of the aDNA template with uracil N-
glycosylase significantly lowered the numbers of substitutions. In their ancient DNA
damage model, using Neandertal mammoth DNA, Briggs and colleagues concluded that
nucleotide misincorporations in aDNA arise mainly because of cytosine deamination,
and are aggravated in single stranded molecules, which are typical for aDNA (Briggs et
al. 2007).
1.5.2.3 Blocking lesions
One of the effects of polynucleotide modification is inhibition of DNA polymerase
progress along the aDNA template strand (Brown & Brown 2011). This is due to
“blocking lesions” which inhibit DNA replication in living cells and may cause eventual
death to cells. However, in living cells, the repair mechanisms usually act accordingly
before such consequences occur (Karanam et al. 2013). However, these mechanisms will
cease to function post mortem, which results in the accumulation of blocking lesions
(Gilbert et al. 2003). If an aDNA sample with blocking lesions were to be used in PCR,
then shorter products are expected, as the Taq DNA polymerase will not be able to copy
the template to full length due to the inhibition from the blocking lesions (Feuillie et al.
2014). The majority of blocking lesions are caused by purine and pyrimidine oxidation
(Dabney, Meyer & Pääbo 2013). Some of the powerful oxidation agents known are
cosmic rays, and derivatives of geological radiation such as super-oxide, hydrogen
peroxide and hydroxyl radicals (Brown & Brown 2011). Oxygen is known as the less
reactive oxidant. Oxidation may disrupt the purine or pyrimidine ring structure and
break it open (Kim & Wilson 2012). This process also may cause purines on the
opposite strands to be dimerized. Maillard reactions may occur which may consequently
cause DNA-protein cross-links. These cross-links are effective blocking lesions which
can in turn prevent DNA amplification by Taq DNA polymerase (Brown & Brown
2011). Other than oxidation, modifications of guanines has also been suggested to be
one of the factors causing blocking lesions (Heyn et al. 2010). Other than cross-links,
the attachment of a peptide to a polynucleotide could also be sufficient to act as a
“blocker”.

42
1.6 Ancient DNA studies of tuberculosis and leprosy
Ancient DNA can be potentially recovered from any form of living remains: humans,
plants, animals. The sources of archaeological aDNA are reviewed by Green and Speller
(2017). Due to the variety of its sources, albeit hindered by preservation state and
molecule damage, aDNA has evolved as an important tool in archaeology. Ancient DNA
has been used to study myriad topics and answer historical questions including the
identification of sex and the relationship between archaeological skeletons; the study of
diets of the people in antiquity; the study of agricultural origin and spread; the
technology used in prehistoric periods; the history of early human origins and
migrations; and the study of ancient disease or palaeopathology (Brown & Brown 2011).
The aDNA application that will be discussed in detail here is the study of past diseases:
tuberculosis and leprosy. To this date, the study of both leprosy and tuberculosis has
been focused on case confirmation and the origin and evolution of both diseases. As
described in section 1.3, skeletal manifestations of both tuberculosis and leprosy are
providing a perfect opportunity to study both diseases using osteological methods.
However, this method does have limitations. For both diseases, not all infected
individuals show skeletal manifestations (Zink et al. 2001), and even for those skeletons
that do show bone changes, the lesions could be ambiguous and could be caused by
other types of disease that give rise to similar bone changes (Roberts & Manchester
2010). Therefore, aDNA studies may offer more precise disease identification by trying
to detect the presence of the pathogen that caused the infection. Evidently, the success of
aDNA disease detection methods depends on the preservation state of the skeletons
themselves.
1.6.1 Ancient DNA studies of tuberculosis
1.6.1.1 Case confirmation
The majority of aDNA studies of past diseases have been done on tuberculosis. This is
owing to the fact that tuberculosis is viewed as one of the most important palaeodiseases
in terms of its long co-existence with human populations as shown by the lesions in
archaeological remains, as well as the fatality burden that still exists today, despite the
previous success of the multi-drug treatments. Ancient tuberculosis has been the focus of

43
aDNA studies as it can cause identifiable bone changes and its DNA can persist in
archaeological remains despite long preservation periods (Spigelman & Lemma 1993).
The aDNA study of MTBC can provide an insight to the origin and evolution of this
ancient disease, which in turn might lead to a better understanding of this disease. As
described earlier, tuberculosis may cause skeletal changes in affected individuals. Out of
all individuals with active tuberculosis infections, only about 3-5% will develop skeletal
manifestation (Roberts & Buikstra 2003). From this 3-5%, approximately 50% will
develop spine tuberculosis. Therefore, the study of tuberculosis aDNA is typically
performed by extracting DNA from the vertebrae and sometimes the ribs – the ribs may
also show lesions in the case of tuberculosis infection (Mays & Taylor 2003; Taylor et
al. 2005).
The typical method of M. tuberculosis complex ancient DNA detection in archaeological
remains is by PCR assays utilising target regions that are thought to be specific to
MTBC members. The two earliest targets that have been used to screen for the presence
of MTBC are the multi-copy targets, insertion sequences IS6110 and IS1081 (Taylor et
al. 1996; Zink et al. 2001). These markers were thought to enhance the likelihood of
ancient TB detection as they are highly conserved within MTBC members as well as
their multi-copy nature. In each bacterium, IS6110 and IS1081 exist in zero to 26 copies
and five to six copies respectively (van Soolingen et al. 1992; Alonso et al. 2013).
However, there are some limitations in using IS6110 as the genetic marker. It is absent
in certain M. tuberculosis strains particularly in some Asian regions, where more than
10% of strains lack IS6110 (Brown & Brown 2011). Furthermore, there is a constant
suspicion about the lack of specificity of IS6110 as a marker (Müller et al. 2015). In
2015, Müller and colleagues published a study which confirmed the presence of IS6110
in ‘mycobacteria other than tuberculosis’ (MOTT) in archaeological remains from
Roman England and medieval Scotland. This study raised a doubt of using IS6110 as the
sole target in confirming archaeological detection of tuberculosis; usage of other more
specific tests is recommended when screening for the presence of ancient TB DNA.
Also, IS6110 and IS1081 do not allow discrimination between MTBC members, which
has prompted the study of single copy markers that might be to be used to identify
individual species in the MTBC group (Coros et al. 2008). One of these markers is the
oxyR pseudogene which harbours a single nucleotide polymorphism (SNP) that has the
discriminative power to distinguish M. bovis from the remaining members of the MTBC
(Sreevatsan et al. 1996). The presence of an A nucleotide at the 285th nucleotide position
of this gene confirms the M. bovis identity while the presence of G points out the

44
identity of the bacterium as being one of the other members of the complex (Sreevatsan
et al. 1996). More recently, two more genes harbouring distinguishing SNPs were
discovered and applied to ancient tuberculosis detection and discrimination: katG and
gyrA (Sreevatsan et al. 1997). The study of variable regions in 100 strains of M.
tuberculosis, M. bovis, M. africanum, M. microti and M. canettii led to the detection of
an M. tuberculosis specific deletion (TbD1) which allows discrimination between
“ancient” and “modern” strains of M. tuberculosis (Brosch et al. 2002). Another single
copy marker that is routinely used in M. tuberculosis aDNA detection is RD2 – currently
known as RD7 – although this is not as frequently used as IS6110 (Gordon et al. 1999).
This particular region can distinguish M. tuberculosis from M. bovis due to its intact
nature in M. tuberculosis but absence in the M. bovis genome.
PCR screening using IS6110, IS1081 and TbD1 confirmed the earliest case of
tuberculosis, dating from 9250 to 8160 years ago (Hershkovitz et al. 2008). The skeletal
remains belonged to a woman and an infant buried together, and were recovered from
Atlit-Yam in the East Mediterranean. Adult and infant aDNA were extracted from rib
and long bone, respectively. The study of aDNA in archaeological remains has brought
about exciting discoveries about this disease in the past. In 1994, Salo and colleagues
successfully amplified IS6110 from naturally mummified lung tissues from a body
which belonged to a 1000-year-old female recovered in Southern Peru (Salo et al. 1994).
This evidence pointed to the presence of human tuberculosis in the New World during
the pre-Columbian period.
1.6.1.2 Origin and evolution of tuberculosis
The study of MTBC variations has resulted in the attempts to reconstruct the origin and
evolution of tuberculosis. The genotyping studies performed in archaeological
specimens are benefiting from the genomic variation studies in contemporary MTBC
isolates.
One of such studies was performed by Sreevatsan and colleagues (1997) which
successfully identified two important SNP markers: katG and gyrA, which allow the
genotyping of MTBC strains into three main principal groups (PGG) : PGG1 to PGG3.
They looked at 26 structural genes in 842 modern MTBC global isolates and
successfully identified the two SNP markers. The discrimination lies in nucleotide

45
mutations in the codon positions 463 and 95 of the katG and gyrA genes, respectively.
The T to G nucleotide substitution on the 463rd position of katG will result in a leucine
to arginine amino acid change. Similarly, the C to G nucleotide substitution at the 95th
codon position of gyrA gene will result to a threonine to serine amino acid substitution.
The distribution of MTBC isolates is depicted in Figure 1.4 below. Interestingly, all M.
microti, M. africanum and M. bovis isolates were categorized specifically PGG1, while
M. tuberculosis isolates are spread across group PGG1 to group PGG3. It was then
highlighted that the species in PGG1 are ancestral to PGG2 and PGG3 as there is more
genetic variations observed – it was assumed that these pathogens were allowed to
accumulate more genetic variations by a longer time of evolution. However, the lack of
neutral mutations was highlighted in this study, as up to 95% of nonsynonymous
mutations are harboured in the antibiotic resistance genes which are under a strong
positive selection pressure (Sreevatsan et al. 1997).
Figure 1.4: The genotyping scheme of MTBC members using the katG463 and
gyrA95 markers. (taken from Sreevatsan et al. 1997: p. 9,871).
Later, a study performed by Bos and research members (2002) have discovered a
specific deletion in RD region and TbD1 gene that can be used to separate between
modern and ancient M. tuberculosis isolates. The classification is based on the absence
or presence of the deletion in the MTBC genome. Another important study of
contemporary MTBC isolates was published by Filliol and colleagues in 2006, where
they successfully classified MTBC into phylogenetically distinct groups made up of six
lineages described as SNP cluster groups (SCGs) as well as five subgroups (Filliol et al.
2006). The M. bovis isolates are contained in the seventh group.

46
Figure 1.5: MTBC phylogenetic lineages 1 to 7.
The ‘modern’ and ‘ancient’ MTBC can be distinguished based on the presence or absence of 7bp
deletions within the TbD1 region. Each branch is showing different MTBC lineages (L1-L7)
which is also associated to the geographical origins of the individuals where the MTBC isolates
were extracted from. (Taken from Brites et al. 2015).
Next, an additional phylogenetic lineage was recognized giving rise to 7 M. tuberculosis
lineages: L1-L7 (Figure 1.5), where M. bovis is now typically being called lineage 8
(Comas et al. 2013). It was revealed that there is a strong link between the place of birth
of the individual and the geographical origin of the M. tuberculosis sample. Lineage 1
typically occurs in the Indian Ocean and the Philippines, lineage 2 is linked to East Asia
tuberculosis, lineage 3 is assigned to both East Africa and India, lineage 4 is for
tuberculosis in Europe and North and South America, whereas lineage 5 occurs
predominantly in West Africa (Witas et al. 2015). Interestingly, lineage 4 is the most
typical group of M. tuberculosis and lineage 7 has only been found in the Ethiopian
region or among recent emigrants from Ethiopia. M. tuberculosis lineage genotyping has
been performed with several archaeological specimens. In 2012, Bouwman and
colleagues performed target enrichment for 247 SNPs of which 214 sites were

47
successfully covered in the subsequent sequencing. From this study, an individual from
the 19th century whose body was recovered from St George’s Crypt in Leeds, West
Yorkshire, England was identified to harbour a genotype belonging to lineage 2, SNP
type (ST) 14/40 and SCG 6 (Bouwman et al. 2012). Another British MTBC aDNA study
was also performed by Müller and colleagues (2014b) revealing that multiple genotypes
of M. tuberculosis were present in this region between the 18th – 19th centuries.
Many studies have been performed to decipher the origin and evolution of MTBC – both
using contemporary and ancient isolates. The most common misconceptions of the
evolution of tuberculosis is that human tubercle bacilli were derived from M. bovis
(Kapur et al. 1994; Stead et al. 1995). This assumption was initiated by the zoonotic
acquisition of tuberculosis infection by humans where the disease can be transmitted
through the ingestion of meat or milk products that have been contaminated by M. bovis.
This idea is propagated even further by the fact that M. tuberculosis has a highly
specialized niche in humans, while M. bovis have a wide host spectrum including cattle,
goats and sheep. The initial theory was that a strain of M. bovis successfully infected a
human, followed by successful spread within human populations. This led to co-
evolution of the tubercle bacillus with human hosts for millennia that resulted in a highly
specialized niche inside the human body, finally giving M. tuberculosis. Furthermore,
the earliest DNA evidence of human tuberculosis is only 9,250-8,160 years ago – which
is a skeleton from the now submerged Atlit-Yam site in the Eastern Mediterranean
(Hershkovitz et al. 2008). Meanwhile, the animal tuberculosis biomolecular evidence
points to a much older case – 17,000 before present (BP) isolated from the an extinct
long-horned bison recovered from the Natural Trap Cave in Wyoming, North America
(Rothschild et al. 2001). This theory however was proposed prior to the availability of
the whole genome sequence (WGS) of M. tuberculosis. The WGS of M. tuberculosis
allowed a comparative genomics study, which successfully uncovered variable genomic
regions among the MTBC members. Among the variable genomic regions uncovered is
the M. tuberculosis specific deletion 1 (TbD1) that is deleted in M. tuberculosis H37Rv
but is intact in the other members of the MTBC. In a set of experiments involving 100
strains of MTBC including M. tuberculosis, M. bovis, M. africanum, M. canettii and M.
microti, 20 variable regions were studied by Brosch and colleagues (2002), including the
TbD1. The MTBC member strains were collected from hosts from diverse global
locations. In this study, it was further found that some of the M. tuberculosis strains have
the deletion in the TbD1 region. In addition, this study also showed that M. bovis
harbours more deletions in its genome in comparison to M. tuberculosis; and based on

48
the M. bovis AF2122/97 (cattle strain) near-complete genome, it was shown that M.
bovis has a smaller genome compared to M. tuberculosis. It was speculated that the
tubercle bacilli that are most similar to the precursor of M. tuberculosis are human
pathogens, not an animal pathogen (M. bovis) – contrary to previous beliefs.
Meanwhile, MTBC origin is also being extensively studied using contemporary and
ancient isolates – with results for the date of origin varying in range from 70 kya to less
than 6,000 years ago. In an earlier publication, Kapur and colleagues (1994) studied
eight short target sequences in 31 modern M. tuberculosis isolates from all parts of the
world. The result revealed a very low nucleotide diversity among the M. tuberculosis
isolates. The calculation of evolutionary time scale based on the number of variations in
four genes suggested that the divergence among the modern isolates might have begun
15,300 to 20,400 years ago. This timeline coincides approximately to the
palaeomigration of humans into the New World. In addition, this time estimate also
coincides with the beginning of cattle domestication which convinced the authors that
rather than M. tuberculosis, M. bovis was the more ancestral species. However, this
study is not without its limitations. It did not take into account the diversity levels in the
other members of the MTBC, hence weakening the argument.
Later, a study by Sreevatsan and colleagues (1997) proposed a recent evolutionary
bottleneck event – with the speciation time predicted to have occurred around 15,000-
20,000 years ago. This assumption was based on the small allelic diversity observed in
the tubercle bacilli. An African origin of MTBC was proposed and several other studies
are also in line with this theory (Gutierrez et al. 2005; Hershberg et al. 2008; Wirth et al.
2008). The age of the M. tuberculosis last common ancestor was estimated by looking at
the accumulation rate of synonymous substitutions. By counting the synonymous
substitution rate, Gutierrez and colleagues (2005) estimated that M. tuberculosis has
existed since 3 million years ago. This coincides with the presence of hominids in East
Africa 3 million years ago – supporting the theory of the emergence of tubercle bacilli
from Africa. From there, it was speculated that the bacilli underwent early
diversification followed by the spread of successful clones to the other parts of the world
following human migrations out of Africa. However, the evidence from archaeological
specimens is not in line with this theory. Bos and colleagues (2014) proposed that the
age of the M. tuberculosis complex most recent common ancestor (MRCA) is less than
6,000 years ago. This conclusion was derived from an aDNA study of a Pre-Columbian
skeleton aged approximately 1,000-years-old. The Peruvian ancient genomes are more

49
closely related to animal lineages; closest to M. pinnipedii strains – restricted to seals
and sea lions. This provides a plausible theory of seal and sea lions as the possible route
of tuberculosis pathogen entry to the New World. The study confirmed pre-Columbian
MTBC infection in South America.
Another recent comprehensive study of ancient M. tuberculosis was published in 2015
by Kay and colleagues. This group has successfully obtained genome sequences of 14
ancient M. tuberculosis isolates through a shotgun metagenomics method, without prior
target enrichment (Kay et al. 2015). The 18th century Hungarian M. tuberculosis isolates
were extracted from human remains, most of which had undergone natural
mummification, recovered from a Dominican church in Vac. The genome sequences
depicted that all 14 ancient M. tuberculosis isolates belonged to phylogeny lineage 4.
This lineage characterization, which is also known as the Euro-American lineage, was
also confirmed by the 7 bp nucleotide deletion within the pks15/1 gene region. A
phylogenetic analysis performed with 1,582 unassembled genomes of other identified
lineage 4 members revealed that at least 12 M. tuberculosis strains were present in this
region during that time. Kay et al. (2015) also dated the divergence time for lineage 4 by
utilizing four high-coverage M. tuberculosis ancient genomes. The mutation rate of this
lineage is estimated to be 5.00 x 10-8 substitutions per nucleotide in one year. This is in
line with the hypothesis from the previous historical M. tuberculosis study (Bos et al.
2014), which suggested the age of the last common ancestor to be not more than 6,000
years ago. Mixed-infection from multiple M. tuberculosis genotypes was also shown in
this study.
In contrary to the hypothesis derived from contemporary MTBC isolates, in historical M.
tuberculosis isolates, Bos et al. (2014) and Kay et al. (2015) predicted the age of the last
common ancestor to be less than 6,000 years ago. However, this is not in line with
several historical tuberculosis cases that are older than 6,000 years old supported by
osteological and biomolecular evidence (Rothschild et al. 2001; Hershkovitz et al. 2008;
Nicklisch et al. 2012). This discrepancy could be influenced by the errors made in the
assumptions in the phylogenetic dating. Kay and colleagues (2015) also proposed that
contamination could also be the cause of this discrepancy. Looking at the current
theories of tuberculosis origin, it seems that more studies are needed to obtain a
conclusion that is consistent for both the contemporary and historical study of
tuberculosis.

50
1.6.2 Ancient DNA studies of leprosy
1.6.2.1 Case confirmation
Similar to tuberculosis, the earliest studies of leprosy ancient DNA were mainly
focussed on case confirmation as an extension of the osteological diagnosis approach.
Rafi and colleagues detected the first ancient M. leprae DNA in 1994 from a 600 AD
archaeological bone recovered from a grave at the Monastery of St John the Baptist,
located at the river Jordan which is where Jesus is believed to have been baptised by
John (Rafi et al. 1994). The detection was performed by PCR amplification of a part of
the 36 kDa antigen and 65 kDa protein encoding genes, the first of which is specific to
M. leprae and the second of which is found in all mycobacteria (Hartskeerl et al. 1989;
Telenti et al. 1993). PCR amplification of the 36 kDa gene gives a 530 bp product,
which is relatively large for ancient DNA amplification, considering the fragmented and
damaged nature of ancient DNA. In 2001, this problem was addressed by Donoghue and
colleagues by designing M. leprae specific primers for a nested PCR which targets the
repetitive element RLEP, and which gives an outer product of 129 bp and a 99 bp
product for the nested PCR (Donoghue et al. 2001). M. leprae detection by PCR of this
repetitive element was demonstrated by Woods and Cole (1990) with modern isolates,
and then subsequently used by Yoon and colleagues (Yoon et al. 1993) with clinical M.
leprae isolates. The repetitive element was first used for detection of M. leprae from
archaeological remains by Donoghue et al. (2001). Today, RLEP is the most frequently
used target to detect M. leprae in archaeological remains. The repetitive element RLEP
is dispersed in at least 28 copies throughout the genomes of modern M. leprae isolates
(Woods & Cole 1990), and is attractive for ancient M. leprae detection as its multi-copy
nature increases the chance of DNA fragment preservation.
The oldest case of leprosy confirmed with ancient DNA is from The Tomb of the Shroud
in Hinnom Valley, Mount Zion (Matheson et al. 2009). The location is associated with
the traditional Akeldama, mentioned in the Bible as the “Field of Blood”. The age of the
skeletal remains is estimated to be from 2025±28 years BP as estimated by the AMS
radiocarbon method. Interestingly, one of the skeletons is not only positive for M. leprae
DNA but also tested positive for M. tuberculosis DNA which indicates co-infection
(Matheson et al. 2009). The presence of these two pathogenic mycobacteria indicates
immune-suppression, which is one of the characteristics of a person with lepromatous or

51
multi-bacillary spectrum leprosy. This skeleton gave positive PCR results for both the
nested RLEP assay and the 18 kDA gene, although the product for the 18 kDa PCR was
in low amounts due to the single copy nature of the marker. The same marker was used
to detect M. leprae DNA in skeletal remains from a South German ossuary and a
Hungarian cemetery dated approximately to 1400-1800 AD and the 10th century AD,
respectively (Haas et al. 2000). The Germany samples were taken from skulls while the
Hungarian sample was from the hard palate and both gave positive RLEP1 and RLEP3
amplifications. In the same study, DNA from the hands and feet were also examined but
did not produce sufficient amplification products for M. leprae detection. The results
suggest that peripheral parts of a skeleton displaying signs of leprosy may contain
significantly fewer bacteria, or it could be that the peripheral bone changes are the result
of secondary infection. This also suggests that the rhinomaxillary alteration is a result of
direct M. leprae bacterial involvement. The first ancient leprosy case from Britain was
from Orkney, Scotland (Taylor et al. 2000). The archaeological remains recovered from
the 13-14th Norse Christian cemetery located at Newark Bay showed lepromatous
leprosy skeletal changes. Screening with the RLEP PCR produced positive amplification
for the extract from the skull bones but not the clavicle, left scapula, vertebra, or femur.
This again, seems to align to the result obtained by Haas and colleagues (Haas et al.
2000). On the other hand, a study of 12th century AD skeletal remains recovered from
the fortress site of Capilla y Castillo de San Jorge in Spain reported successful M. leprae
detection from metacarpal bones following RLEP PCR (Montiel et al. 2003). The
positive identification was also supported by restriction analysis with ClaI which detects
a site present in the RLEP region as well as Sanger sequencing.
Later studies have detected more leprosy cases from global locations. The earliest case
of leprosy confirmed by osteological evidence of high confidence is from the Roman
period (Rubini, Erdal, et al. 2014). A skeleton recovered from Martellona, Italy, was of a
child aged between 4 to 5 years with pronounced rhinomaxillary syndrome strongly
suggestive of a well-developed leprosy infection. This is the youngest individual with a
confirmed case of leprosy. It is quite surprising to observe convincing skeletal
manifestations in such young individuals, considering leprosy is a chronic and slow
progressive infection. However, one of the proposed explanations for this was that the
M. leprae strain that existed at this time was more aggressive compared to contemporary
M. leprae (Rubini, Erdal, et al. 2014). Another childhood leprosy case that was reported
in the same study, describing a 4-5 month old infant from Kovuklukaya, Turkey, with
some indication of chronic inflammation in the bone, is even more surprising as the

52
leprosy case was confirmed by molecular evidence (Rubini, Erdal, et al. 2014). These
skeletal remains are estimated to age from the 8th to 10th centuries AD, during the
Byzantine period. Both cases most probably result from in utero haematogenous
transmission of the infection from a pregnant mother who was suffering from leprosy
infection. In addition, past leprosy cases have also been confirmed through molecular
evidence in Japan, from archaeological remains recovered from the Hatanai site located
in the Honshu Island of Japan (Suzuki et al. 2010). The analysis of the excavation sites
estimated the age of the bone remains to be most likely from the mid-18th to the early
19th century AD. The bones show characteristics of the lepromatous type of leprosy,
with rhinomaxillary syndrome (Suzuki et al. 2010). Another study by Watson and
Lockwood (2009) confirmed leprosy cases in Croatia (8th-9th century AD), Denmark
(1275-1560 AD) and the United Kingdom (900-1000 AD). The positive detections were
from the rhino-maxillary area (Croatia), palatine bone (Denmark) and rhino-maxillary,
tibia, and metatarsal (United Kingdom). Positive M. leprae detection has also been
obtained from a Byzantine skeleton dated 300-600 AD from Bet Guvrin, Israel
(Spigelman & Donoghue 2001).
1.6.2.2 Origin and evolution of leprosy
Most of the past leprosy case confirmation studies described previously were performed
by amplification of the RLEP region. Whole genome sequencing of the modern M.
leprae TN strain from Tamil Nadu, India, provided an opportunity to explore additional
sites of the genome that could be studied and applied to ancient M. leprae isolates (Cole
et al. 2001). The 3,268,210 bp genome of M. leprae has a G+C content of only 57.8%,
which is much lower than that of M. tuberculosis, whose genome is 4,411,531 bp with
65.6% G+C content. In fact, the G+C content of M. leprae is the lowest of all known
mycobacteria (Singh & Cole 2011). From the genome sequence analysis done by Cole
and colleagues, only 49.5% of the M. leprae genome codes for proteins, while 27% and
23.5% of the genome are pseudogenes and non-coding DNA, respectively (Cole et al.
2001). From a comparison of the M. leprae and M. tuberculosis genomes, under the
assumption that these were at one point of time in a state of similar topology, it was
concluded that the M. leprae genome has undergone significant downsizing, and may
have lost around 2,000 genes since the last common ancestor of the two species
(Eiglmeier et al. 2001). The high number of pseudogenes together with the increasing

53
number of insertion sequences (IS) and reduced G+C content are attributes of reductive
evolution, which can be linked to the extreme obligate nature of M. leprae (Singh &
Cole 2011). The reason for the massive gene inactivation could be the highly specific
niches occupied by the bacteria, such as Schwann’s cells, where M. leprae will not be
subject to competitive pressure from other microbes. Gomez-Valero and colleagues
(2007) suggested that the shaping of M. leprae into a highly specialized pathogen was
due to massive gene reduction which most likely happened at the same time as the
pseudogenization events.
The sequencing of the whole M. leprae TN genome allowed further exploration of
regions and targets that can be used in genotyping. Selected genes, pseudogenes and
non-coding regions were compared between the genomes M. leprae TN and Brazilian
strains (Monot et al. 2005). This comparative genomic study resulted in the discovery of
three unique SNPs that can classify M. leprae isolates into four groups: SNPs type 1 to
4. These SNPs, at positions 14,676, 1,642,875 and 2,935,685, interestingly show strong
association with the geographic origins of patients. SNP type 1 is mostly shown by
patients from Asia, East Africa and the Pacific region, SNP type 4 is typically found in
the Caribbean and West Africa, while SNP type 3 is the typical of the M. leprae type
infecting the people in Europe, North Africa and the Americas, as demonstrated in
Figure 1.6 (Monot et al. 2005). Fascinatingly, SNP type 2 is the least typical type,
infection of M. leprae with this type being found only in Malawi, Ethiopia, Nepal or
North India, and New Caledonia (Monot et al. 2005). Based on this scheme, it is
suggested that leprosy originated in the Near East or in Eastern Africa. Subsequently,
successive human migrations have been responsible for the dissemination of this disease
worldwide. This scheme was soon used to classify ancient M. leprae isolates from
Europe, including the United Kingdom, Denmark and Croatia (Watson & Lockwood
2009). This study showed that all ancient M. leprae isolates genotyped from these three
European countries (476 AD to 1350 AD) belong to SNP type 3 (Watson & Lockwood
2009). A comprehensive study performed by Monot and colleagues (2009) involved the
comparative analysis of four complete genomes of modern M. leprae strains: M. leprae
TN from India, M. leprae Br4923 from Brazil, M. leprae Thai 53 from Thailand, and M.
leprae NHDP63 from the United States. The analysis revealed extremely low sequence
diversity among strains: they are

54
Figure 1.6: The distribution of four different leprosy SNP types around the world
and the prediction of human migrations.
Each SNP type is depicted in a different colour. SNP-type 1: yellow, SNP-type 2: orange, SNP-
type 3: purple, SNP-type 4: green. The arrows are showing the proposed routes of migrations,
inferred from SNP analysis – the colour is similar to the circles for each SNP-type. The
estimated time of migration (in years) is shown in grey arrows. The estimation was made based
on genetic, archaeological and anthropological studies (Figure taken from Monot et al. 2005).
99.995% identical. There were only 215 polymorphic sites and 5 pseudogenes identified.
The evaluation of polymorphic sites classified M. leprae isolates into 16 subtypes (1A-
1D, 2E-2H, 3I-3LM, 4N-4O) that are closely related to the geographical location of their
hosts. This subtyping scheme is able to shed light on the trade routes and migration
patterns of humans in the past, as depicted in Figure 1.6 (Monot et al. 2005). Based on
the construction of a phylogenetic tree of the present M. leprae isolate sequences, the
genotype of the ancestral M. leprae strain was estimated as between type 2 and 3. This
result supports the previous assumption that M. leprae in East Africa could belong to
type 2. This type 2 East African strain then produced the type 1 strain that moved east to
Asia following human migrations. From there, M. leprae spread to the west, towards the
Middle East then Europe, in type 3 form before finally giving rise to the type 4 that can
be found in West Africa, the latter movement believed to be through the slave trade. The
introduction of leprosy to the Americas was most probably through European
immigrants instead of crossings of humans via the Bering straits (Monot et al. 2009). A
genome-wide comparison of five medieval M. leprae strains, each with more than 80%
genome coverage, discriminated M. leprae isolates into five distinct branches, 0-4, each

55
associated with a particular subtype (Schuenemann et al. 2018). The most ancestral
branch 0 is associated with subtype 3K which can be found in contemporary M. leprae
strains isolated in China and New Caledonia.

56
Chapter 2: Materials and methods
2.1 Archaeological samples
The archaeological bones described here are the samples studied in Chapter 3 and 4. The
samples investigated in Chapter 5 is described separately in each respective chapter.
Sixty skeletal samples from sixty individuals dated from Roman to the post-Medieval
periods were studied (Table 2.1, Table S2.1). Rib fragments were mainly analysed from
the 60 skeletons from 16 different locations in Yorkshire. The samples originated from
urban and rural locations of Yorkshire. The exact location in each site is shown in
Figure 2.1. Seven of the sites are in York city, therefore the samples from the area are
classified as “urban”. The sample from East Heslington is still considered urban
although it is not located inside the city of York. This classification is based on its
proximity to York in comparison to the other rural sites. It could have been a separate
community in the past but the closeness to York made it possible for people to be
exposed to the same strain of MTBC that was circulating in the city during that period.
The archaeological bones were collected by Dr Darlene Weston as part of the National
Environment Research Council (NERC) project on “The Palaeopathology of
Tuberculosis in Britain and Europe” (NE/E015697/1, 2007–2010), subsequently
continued as “Palaeopopulation genomics of Mycobacterium tuberculosis
(NE/K012185/1, 2013–2016). Two from the 60 bone remains: 3 Driffield Terrace 13 and
East Heslington 229 have previously been studied and reported by Dr Romy Muller and
colleagues (2014a). The remaining 58 samples were studied for the first time in this
project. DNA extraction was performed from the bone powder in the aDNA lab facility
at the Manchester Institute of Biotechnology. In my work, PCR assays were used to test
for MTBC aDNA preservation, using markers which are supposedly specific to the
MTBC genome: the insertion sequence IS6110, and the gyrA, Rv0083 and Pks 15/1 loci
(Bouwman et al. 2012; Alonso et al. 2013; Müller et al. 2014a). Although the specificity
of the multi-copy region IS6110 is questionable, it is still the most robust target for
MTBC aDNA detection so far (Müller et al. 2015). Therefore, as per the
recommendation by Muller et al. (2015), the screening was accompanied by the other
three single copy regions, which are deemed specific to the MTBC genome. Nested PCR
was incorporated in the IS6110 assay to enhance the detection power and increase

57
detection specificity. The three single-copy targets have previously been used for SNP
(gyrA, Rv0083) and indel (Pks15/1) genotyping (Müller et al. 2014b).

58
(b)
(a)
Figure 2.1: The origin locations of the Yorkshire archaeological remains. (a) The rural sites
which are marked by red placemarks and one of the urban sites marked by the yellow
placemarks. The red circle marks the city of York location where the rest of the urban samples
were taken from, (b) The urban sites in the city of York marked by yellow placemarks. Some of
the placemarks overlap with each other. Images are extracted from Google Earth.
York

59
Burial location Period Context Skeleton
number
Parts of skeletons
showing
tuberculosis
lesions or non-
specific lesions
Part of
skeleton
studied
Reference
York Minster Early
mediaeval
Urban
cemetery
1 Control Right rib Lee n.d.
15 Lytic lesions on
sacrum and lower
thoracic
vertebrae.
Periosteal bone
formation and
lytic lesions on
the ribs
Right shaft
femur
Fishergate House Late
mediaeval
Urban
cemetery
86 Periosteal new
bone formation
on the right ribs
Right rib Holst 2005
98 Rib lesions Left rib
108 Lytic lesions on
lateral portions of
lower thoracic
and lumbar
vertebral bodies
and lesions on the
ribs
Left rib

60
135 Rib lesions Left rib
147 Control Rib
149 Control Rib
St Andrew Fishergate Late
mediaeval
(Early 14th
century AD)
Urban
cemetery
6 Lytic lesions on
sacrum and lower
thoracic
vertebrae.
Periosteal bone
formation and
lytic lesions on
the ribs
Rib Stroud & Kemp 1993
277 Rib lesions Rib
286 Rib lesions Rib
296 Periosteal new
bone formation
on bodies of L1-
L3 vertebrae.
Lytic lesions
observed on body
of L1 vertebrae
Rib
323 Destruction of L5
vertebrae.
Periosteal new
bone formation
and lytic lesions
on the ribs
Fragment
from
proximal
right tibia

61
339 Lytic lesions on
the thoracic and
lumbar vertebrae
Rib
384 Rib lesions
accompanied by
endocranial
lesions
Rib
34 Control Rib
131 Control Rib
253 Control Rib
St Helen-on-the-Walls Late
mediaeval
Urban
cemetery
5000 Left os coxa
which is a
separated element
from mixed
context
Rib Dawes & Magilton 1980
5844 Lesions on the
left ribs and the
L4 and L5
vertebrae
Left rib
6003 Lower thoracic
and lumber
vertebrae
Rib
5494 Control Rib

62
East Heslington Late Roman
(3-8th
century AD)
Burial
associated
with high-
status
Roman villa
in rural
location
229 Lower thoracic
vertebrae
Lumbar
vertebrae
body
Holst 2008; Neal & Roskams 2012
3 Driffield Terrace Roman
(Early 3rd
century AD)
Urban
cemetery
37 Rib lesions Rib Caffell & Holst 2012; Muldner et al. 2011
54 Rib lesions Rib
13 None Rib
15 Rib lesions Rib
6 Driffield Terrace Roman
(Early 3rd
century AD)
Urban
cemetery
19 Rib lesions Rib Caffell & Holst 2012
22 None Rib
St Peter’s Huddersfield Post
mediaeval
Urban
churchyard
5 Rib lesions Right rib Cafell & Holst 2008b
7 Rib lesions Right
femur
17 Control Proximal
end of left
humerus
Wetwang Slack Iron Age Rural
cemetery
1 Rib lesions Right rib Dent 1984
2 Destruction of left
hip
Rib

63
5 Lytic lesions on
thoracic vertebrae
T12 and lumbar
vertebrae L2
Rib
6 Lytic lesions on
upper thoracic
vertebrae
Rib
8 Lytic lesion on
right ilium
Rib
9 Psoas abscess on
the left femur
Rib
3 Control Rib
4 Control Rib
7 Control Rib
Settlement 185 N/A Vertebrae Dent 1984
360 N/A Vertebrae
415 N/A Vertebrae
Sewerby Anglo-
Saxon (mid-
6th – 7th
century AD)
Rural
cemetery
34 Calcified pleura Skull
fragment
Hirst 1985
44 Control Mid-shaft
left radius

64
St Giles by Brompton
Bridge
Late
mediaeval
(12th-15th
century AD)
Rural church
cemetery
1288 Rib lesions Rib
fragment
Cardwell et al. 1995
1531 Rib lesions Rib
fragment
1542 Control Left
humerus
fragment
Alicy Hill Early
mediaeval
(7th century
AD)
Rural
cemetery
1044 Pott’s spine with
lytic lesions on
T11 and T12
vertebrae
Rib Hall & Whyman 1996
1043 Control Rib
Wharram Percy Late
mediaeval
Rural
cemetery
26 Left elbow Distal left
ulna
Mays et al. 2007
1600 Control Femur
fragment
Addingham Early
mediaeval
(670-
990AD)
Rural
cemetery
134 Pott’s spine Rib
fragment
Adams 1996
223 Rib lesions Rib
fragment
103 Control Rib
fragment
Melton Roman Rural 4297 Rib lesions Left rib Caffell & Holst 2008a

65
cemetery 2554 Rib lesions Right rib
5319 Control Left tibia
Hickleton Late
mediaeval –
early post
mediaeval
Rural 46 Spine Vertebrae Manchester & Roberts 1986
Table 2.1: A complete list of all bone remains studied for the preservation of MTBC aDNA. This table showing the skeleton dates, site origins, skeleton
parts showing lesion or non-specific lesions (and control) and skeleton parts sampled in the study. N/A: information not available

66
2.2 Authentication regimes
As discussed previously, the damaged, fragmented and low copy nature of ancient
DNA demands protocols to be performed in a very careful manner (Dabney, Meyer, et
al. 2013). DNA templates of these characteristics are very prone to exogenous DNA
contamination (Knapp et al. 2015). The typical sources of contamination are from the
environment and DNA of the personnel handling the archaeological remains which
effortlessly outcompete the very little and damaged endogenous aDNA recovered
(Noonan et al. 2005; Poinar et al. 2006). There is a risk to introduce contaminants at
any stage of the experiments (Gruber 2015). The study was carefully tailored to make
sure there is a balance between minimizing the risk of contamination in samples and
maximizing the yield of endogenous DNA.
All experimental protocols prior to PCR amplification in the PCR thermocycler were
performed in physically isolated clean rooms dedicated to ancient DNA at the
Manchester Institute of Biotechnology. The clean rooms are located on a different
floor (lower ground) to the laboratory where all the “modern” or “post PCR
amplification” experimental works is carried out on the second floor (previously first
floor) in the building. There are two clean rooms dedicated for experimental works
using archaeological bone remains as starting materials: a human bone aDNA
extraction room and a human bone aDNA PCR mixture preparation room. Both rooms
are physically isolated from each other. Access to these rooms is restricted to trained
lab personnel and the inevitable occasional equipment maintenance, and is locked at all
times except when in use. Each room is designed to have an antechamber at the anterior
of the clean room which acts as contamination buffer zone and gowning area before
entering the clean room. The room is supplied with ultra-filtered air which circulates in
the room in a positive displacement manner. This manner of air circulation will avoid
the uptake of possibly-contaminated unfiltered air from the outside of the clean room.
In addition, the clean rooms are also equipped with ultraviolet (UV) lights (254 nm);
the rooms are UV irradiated for at least four hours after each use.

67
Appropriate protective clothing is worn before entering the clean room; the gowning is
performed in the antechamber. The protective clothing includes disposable
polypropylene coverall, surgical mask, hair cover/net, goggle, disposable shoe covers
and two layers of gloves with the outside layer being changed regularly. Benches and
surfaces inside the aDNA room are cleaned with 5% sodium hypochlorite, followed by
70% ethanol. The more delicate equipment surfaces such as centrifuges and weighing
balances are cleaned with DNA Away (Molecular Bioproducts). Smaller equipment
such as pipettes, consumables and selected reagents were UV irradiated (254 nm,
120,000 µJ cm-2 for 2 × 5 min, with 180° rotation between the two exposures) before
use. All DNA extraction protocols were performed in a Class II biological safety
cabinet. Meanwhile, in a physically separate aDNA dedicated room, PCR mixtures
were prepared in a laminar flow cabinet. This room is restricted for PCR mixture
preparation before the amplification is performed in a distant modern laboratory
dedicated to modern samples and post-PCR experiments. In addition, the PCR clean
room is also used for DNA library mixture preparation, again prior to the amplification
step.
2.3 Bone scraping and crushing
The archaeological bone samples studied in Chapter 3 and Chapter 4 have previously
being scraped and crushed by personnel in Brown lab: Dr Abigail Bouwman and Dr
Romy Muller. Meanwhile, the samples described in Chapter 5 have been scraped,
crushed, extracted and screened for M. leprae presence by Dr Romy Muller as part of
her Master’s project. The starting materials used in my project were DNA extracts with
confirmed positive detection of M. leprae aDNA. The protocol described in this section
explains how the bone powder was prepared.
The part of bone of interest, usually the part showing lesions, was cut using a small
hacksaw while the remaining bone fragments, if any were kept. The exposed surface
area of the bones was scraped with a scalpel, to remove the outer 1 mm, before placing
in a small sampling bag and UV irradiating on both sides for approximately 5 minutes
on each side. This is followed by bone crushing. Finally, the bone powder was weighed
and transferred to sterile 1.5 mL Eppendorf tubes for subsequent DNA extraction.

68
2.4 Ancient DNA extraction
This procedure was performed on the bone powder samples described in Chapter 3 and
4. The ancient DNA extraction procedure performed in this study follows the protocols
described by Dabney and colleagues with a few modifications (Dabney, Knapp, et al.
2013). The two days DNA extraction used 200 mg of bone powder in each extraction
batch. Each extraction batch included four different samples and two extraction blanks.
In cases where DNA extraction was performed on only one or two samples, only one
extraction blank was included. For the extraction blanks, similar reagents were added,
and the same protocols were performed but no sample was added to the tube, not even
water.
On the first day of extraction, the 200 mg bone powder was mixed with extraction
buffer in a Falcon™ 50 mL conical centrifuge tube. The extraction buffer comprises
0.45 M Ethylenediaminetetraacetic acid (EDTA), 0.25 mg/ mL molecular biology
grade Proteinase K (New England BioLabs) and Invitrogen™ UltraPure™
DNase/RNase-free distilled water to 2 mL final volume. Next, the samples were
incubated overnight (18 to 24 hours) in the dark in a water bath set to 37°C, with
constant agitation at 200 rpm. The incubation step is aiding the DNA release from the
bones by enhancing the powder digestion (Rohland & Hofreiter 2007).
On the second day of DNA extraction, the incubated solution was centrifuged at 6,000
rpm in an Eppendorf centrifuge for 5 minutes. This step was repeated twice by slightly
turning the Falcon™ 50 mL conical centrifuge tube containing the extraction solution
each time. After the third time of centrifuging, the supernatant was transferred to a new
Falcon™ 50 mL conical centrifuge tube and mixed with 12 mL binding/PB buffer
(Qiagen). The addition of binding buffer will aid the efficiency of DNA binding to the
spin-column membrane. This step and the subsequent steps were performed following
the instructions for the Qiagen MinElute kit with a few amendments. Now, the total
volume of the supernatant-binding buffer mix solution is 14 mL which exceeded the
maximum volume of the spin-columns provided by the manufacturer (Qiagen).
Therefore, in order to accommodate this, the Qiagen-provided spin-column was
connected to a reservoir and attached to a Falcon™ 50 mL conical centrifuge tube.

69
Next, the binding apparatus was centrifuged at 6,000 rpm for 5 minutes. Subsequently,
the spin column was detached from the binding apparatus and placed into a collection
tube provided by the manufacturer (Qiagen). To dry, the spin-column was centrifuged
for 1 minute at 6,000 rpm speed. This is followed by two washing steps with 750 mL
washing/PE buffer (Qiagen). In between the washing steps, the spin-column was
centrifuged at 3,300 g for 1 minute. Lastly, two rounds of elution were performed by
adding 30 µL elution/EB buffer at each step, yielding 60 µL total volume of DNA
extract.
2.5 PCR assays screening for ancient DNA preservation
This step was performed for all samples mentioned in this study except for the samples
in Chapter 5, as the presence of M. leprae aDNA has already been confirmed in a
previous study (Müller 2008). To maintain the “ancient” nature of the DNA extract
used as the template, the PCR mix was prepared in the PCR clean room before
amplification in the modern laboratory. To maintain the one-way workflow, the
subsequent experiments post-PCR amplification were all performed in the modern
laboratory.
All DNA extract samples obtained from archaeological skeletons mentioned in Chapter
3 and 4 were screened for MTBC aDNA presence. In order to avoid contamination, a
positive control of M. tuberculosis DNA was not included in this study, which means
there will be no source of modern M. tuberculosis contamination, though there is still
the possibility of cross-contamination from M. tuberculosis DNA positive samples. In
each round of PCR amplification using the ancient DNA extract, two negative controls
(the PCR mix with no added DNA extract) were included.
The PCR assays for MTBC aDNA presence screening targeted four regions of the
MTBC genome, both multi-copy and single-copy regions. The first region targeted was
the multi-copy insertion sequence IS6110, which is typically used in MTBC aDNA
detection (Salo et al. 1994; Rothschild et al. 2001). The insertion sequence IS6110 can
exist in up to 26 copies per genome (Alonso et al. 2013). However, in rare cases, some
M. tuberculosis isolates have no copies (Huyen et al. 2013). The multi-copy nature
increases the chance of preservation of this target region in archaeological bone

70
remains and hence increases the sensitivity of the PCR assay. To further increase the
detection signal, a nested PCR which amplifies a 92 bp fragment within the 123 bp
product from the first round of PCR was incorporated in the study (Konomi et al.
2002). Moreover, the presence of the band from the nested PCR in the gel
electrophoresis increases the confidence that the PCR band signal obtained from the
first PCR step was indeed from MTBC aDNA.
Apart from the insertion sequence IS6110, three single-copy targets were also included
in the MTBC aDNA preservation study. These targets are gyrA, Rv0083 and
polyketide synthase pks15/1, which have been used in MTBC strain discrimination in
previous studies (Müller et al. 2014b). Both gyrA and Rv0083 contain discriminative
SNPs while insertion or deletion of the polyketide synthase 15/1 enables MTBC strains
to be classified (Bouwman et al. 2012).
The PCR mixture with a total volume of 30 µL contained 1 x Amplitaq Gold 360
Master Mix (Applied Biosystems), 10% v/v 360 GC enhancer (Applied Biosystems),
0.5 ng/µL of bovine serum albumin (BSA) (New England Bio Labs), 400 nM of
forward and reverse primers and water. The purpose of adding BSA is to minimize the
PCR inhibitions (Forbes & Hicks 1996; Abu Al-Soud & Rådström 2000). The primer
sequences used in each PCR assay are depicted in Table 2.2. Since the amount of
endogenous DNA in the extract is not known, 3 to 6 µL DNA extract was used as the
template. For the IS6110 nested PCR, 1 µL of PCR product from the first step
amplification was used as the template. PCR inhibitors are often a problem in ancient
DNA studies, as these may have been co-extracted therefore hindering the PCR
amplification process. Hence, apart from the original undiluted DNA, a 10-fold diluted
DNA template was prepared for each sample and tested by adding 6 µL of this template
in the PCR mix.
For the first step of the nested IS6110 PCR, and for the gyrA, Rv0083 and Pks 15/1
PCRs, the cycle conditions were set as: 95ºC for 5 minutes followed by 45 cycles of
95ºC for 45 seconds, annealing temperature (Table 2.2) for 45 seconds and extension at
72ºC for 45 seconds. Lastly, the final extension was performed at 72ºC for 7 minutes.
Identical cycle conditions were set up for the IS6110 nested PCR, but instead of 45
cycles, 25 cycles were performed. The IS6110 nested PCR amplification was
performed in a separate thermal cycler machine to the other four assays, as the template
used is the already-amplified PCR product from IS6110, therefore considered as no

71
longer “ancient”. This is to minimize the risk of contamination. The PCR products were
loaded into a 1.5% agarose gel electrophoresis which was stained with GelRed Nucleic
Acid Gel stain (Biotium, Inc). A UV transilluminator was used to visualize any PCR
bands in the agarose gel. Bands that are believed to be the correct or near to the correct
size were purified using the MinElute PCR purification kit following the standard
supplier procedure (Qiagen). In the case of multiple bands produced from the same
sample, and where one of the bands is of correct of near correct size, the band was
excised and the DNA purified with the MinElute Gel Extraction kit following the
standard supplier procedure (Qiagen).

72
Table 2.2: List of primers used in the PCR screening for MTBC aDNA presence. Four MTBC specific targets were used in the MTBC aDNA presence
screening. The primer sequences, annealing temperatures and amplicon sizes are shown.
Target Primer pair sequence 5'-3' Annealing
temperature
(°C)
Amplicon
size (bp)
Reference
IS6110 first step F- CCTGCGAGCGTAGGCGTCGG 68 123 (Thierry et al. 1990)
R- CTCGTCCAGCGCCGCTTCGG
IS6110 nested PCR F- TCGGTGACAAAGGCCACGTA 58 92 (Taylor et al. 1996)
R- TTCGGACCACCAGCACC T
gyrA F- CCGGTCGGTTGCCGAGACCA 68 104 (Bouwman et al. 2012)
R- GCGGGTAGCGCAGCGACCAG
Rv0083 F- GCCACCGCCCCGACCAC 69 110 (Bouwman et al. 2012)
R- GTCACCCACACCGCCGAGTC
Pks 15/1 F- ATCTCGCCGAAATCACCCACG 67 92-99 (Bouwman et al. 2012)
R - CGTACCAGCCCCCGCAGAG

73
Once purified, the PCR products were then cloned in XL-1 Blue competent Escherichia
coli cells (Agilent Technologies) using the blunt-end system CloneJET PCR Cloning kit
(ThermoFisher Scientific) protocols. Positive clones were screened using pJET vector
specific primers: pJET forward primer
(5’-CGACTCACTATAGGGGAGAGCGGC-3’) and pJET reverse primer (5’-
AAGAACATCGATTTTCCATGGCAG-3’). The 20 µL PCR mix comprises 200 nM of
each primer mentioned, 1x Taq PCR buffer and 0.5 units of Taq DNA polymerase, both
from New England Biolabs, and 200 µM of dNTP mix. Cycle conditions were: 95ºC
initial denaturation for 3 minutes followed by 30 cycles of denaturation at 94ºC for 30
seconds, annealing at 60ºC for 30 seconds, and extension at 72ºC for 30 seconds. Lastly,
the final extension was set to 72ºC for 7 minutes. Next, the positive clones were purified
and sent for Sanger sequencing at GATC Biotech (now Eurofins Genomics). The
resulting Sanger sequences were visualised and analysed in Geneious 8.1
(https://www.geneious.com, Kearse et al., 2012). In Geneious, the sequences were
trimmed and aligned to the M. tuberculosis H37Rv reference sequence for each region.
The possible identities for the sequences which failed to match the reference sequence
were determined using the NCBI Basic Alignment Search Tool (BLAST).
2.6 Next generation sequencing
2.6.1 DNA library preparation
For the samples in Chapter 3 and 4, depending on the PCR evaluation of MTBC aDNA
presence, appropriate samples were selected to be further studied using next generation
sequencing (NGS). For the samples in Chapter 5, all samples deemed positive for M.
leprae aDNA from the previous study (Müller 2008), were subjected to NGS. For all the
samples subjected to NGS, the DNA libraries were prepared following the double
indexing strategy for Illumina Genome Analyzer sequencing. Following the
development of high throughput DNA sequencing, it is a routine to include multiplexed
samples (pooling multiple libraries together) per run to increase cost efficiency (Rohland
& Reich 2012). In Illumina sequencing, the multiplexing strategy usually allows simple
identification by including an embedding index (short fragment of unique sequence)
within one of the adapters (Kircher et al. 2012). However, one of the drawbacks of this
approach is the risk of falsely assigning the resulting sequences to their original samples,
which is highly undesirable in a genotyping study. Therefore, a double-indexing method,

74
in which indexes are placed in both universal adapters at both ends of the sequence, was
utilized in DNA library preparation in this study in order to improve multiplex
sequencing accuracy (Kircher et al. 2012).
Figure 2.2: A simplified scheme of make up of dual-indexed DNA library fragments.
Adapters (green) are ligated to both ends of the DNA insert. In the indexing PCR amplification,
P7 and P5 adapters (light and dark blue); each contain different indices (yellow), will be attached
to the insert.
The DNA library was prepared to obtain fragments which contain compenents as
depicted in Figure 2.2 above. The DNA library preparation was preformed in house,
rather than outsourcing, to make sure the conditions comply to ancient DNA
requirements. The DNA library preparation involves a series of stages including blunt-
end repair, adapter ligation, adapter fill-in, and the final indexing PCR. The protocols
followed the steps described by Meyer and Kircher (Meyer & Kircher 2010). All first
part of the procedure was performed in the clean room before the subsequent
amplification steps were performed in the modern laboratory. All tubes used were DNA
LoBind tubes which minimize the sticking of DNA to the walls. The DNA templates

75
were not subjected to any DNA shearing, as per the typical modern DNA library
preparation protocols, as ancient DNA is already fragmented.
Firstly, blunt-end repair was performed to remove or fill the overhanging 5’- and 3’-
ends. These actions are completed by T4 DNA polymerase while the T4 polynucleotide
kinase is attaching the 5’-phosphates. In this step, 1 x Buffer Tango (ThermoFisher
Scientific), dNTPs (100 µM each), 1 mM adenosine 5’-triphosphate (ATP)
(ThermoFisher Scientific), 0.5 U/µL T4 polynucleotide kinase (ThermoFisher Scientific)
and 0.1U/µL T4 DNA polymersae (ThermoFisher Scientific) and 25 µL DNA template
were combined together to make up 50 µL reaction. Next, the mix solution was
incubated at 25ºC for 15 minutes followed by 12ºC at 5 minutes in a thermal cycler.
Subsequently, the solution was purified following the standard MinElute PCR
purification kit (Qiagen) protocols and eluted in 20 µL volume.
Secondly, the two adapters, P5 and P7 were attached to the ends of the blunt-end
repaired DNA templates through the actions of T4 DNA ligase. The reaction mix with a
total volume of 50 µL comprises 1 x DNA ligase buffer (ThermoFisher Scientific), 5%
polyethylene glycol 4000 (PEG-4000) (ThermoFisher Scientific) and 0.125 U/µL T4
DNA ligase (ThermoFisher Scientific) was prepared and combined together with 20 µL
purified and blunt-end repaired DNA template from the previous step. This solution was
incubated at 22ºC for 30 minutes. Similarly to the previous step, after this incubation the
solution was purified using the MinElute PCR purification kit (Qiagen) and eluted to
obtain 20 µL final volume.
Next, nicks produced from the previous step were filled-in with Bst polymerase in an
adapter fill-in reaction. This reaction was prepared by mixing 1 X ThermoPol reaction
buffer (New England Biolabs), dNTPs (250 µM each) and 0.3 U/µL Bst polymerase,
large fragment (New England Biolabs) which was then combined together with the 20
µL eluate from the previous step to make up a 40 µL reaction. There was no purification
step necessary in this stage as the Bst polymerase is inactivated by heating to 80°C. Prior
to the next step, the indexing PCR, the number of molecules in the DNA library has to
be quantified in order to calculate the PCR cycle number needed to obtain a sufficient
amount of the DNA library. The samples were quantified by quantitative PCR (qPCR)
against standards with known concentration. The qPCR mix which was prepared in a 96
well plates consists 1 x LightCycler® 480 SYBR Green I Master (Roche Life Science),
200 nM of each primer which are complementary to the universal adapter sequence: IS7
primer 5’–ACACTCTTTCCCTACACGAC-3’ and IS8 primer 5’-

76
GTGACTGGAGTTCAGACGTGT-3’ (Meyer & Kircher 2010). DNA sample of
volume 1 µL was then added to the mix to give a 20 µL reaction volume. The reactions
were run in a LightCycler® 480 Instrument II (Roche Life Science) with cycles at: 95°C
for 10 minutes, followed by 45 cycles of 95°C for 15 seconds, 59°C for 25 seconds and
72°C for 35 seconds. In addition, the amount of DNA was also measured in the original
DNA sample with a Qubit® fluorometer (ThermoFisher Scientific) following the
dsDNA High Sensitivity Assay (ThermoFisher Scientific) protocol. Determination of the
optimal PCR cycle number was then carried out as described by Meyer and Kircher
(2010).
Finally, the P7 and P5 adapters with indexes embedded were attached at both ends of the
library by PCR amplification. The whole library volume was used as the template in 7 x
25 µL reactions consisting of 1 x KAPA HiFi HotStart Uracil + ReadyMix (Roche Life
Science), 200 nM of each P5 and P7 indexing primers. The amplification was performed
in a thermal cycler with initial denaturation for 4 minutes at 95°C, followed by the
optimum cycle number (determined in the previous step) of denaturation at 98°C for 20
seconds, annealing at 63°C for 30 seconds, and elongation or extension at 72°C for 30
seconds; lastly, final extension was performed at 72°C for 1 minute. The amplified
library was purified following the MinElute PCR purification kit (Qiagen) standard
protocols to give a final elution volume of 25 µL. The next step will depend on the NGS
starategy that is to be performed to the DNA library: shotgun sequencing or target
capture. For shotgun sequencing, there are no further steps needed before pooling except
for quality control and quantification before sending for sequencing.
2.6.2 Target enrichment: in-solution target hybridization capture
This study used the in-solution DNA capture strategy for whole genome target
enrichment. The baits targeting the whole genome of M. tuberculosis and M. leprae were
designed in house in Brown’s lab group. These baits were supplied as part of the
myBaits® custom target kit from Arbor Biosciences (formerly MYcroarray®). They
were designed based on the whole genome sequence of the M. tuberculosis ancestor
genome and the M. leprae TN genome (Cole et al. 2001; Comas et al. 2013) . The baits
are 80 bp in length and have 2X tiling density, which means each base of the genome is
covered twice by different baits.

77
The target capture protocols were performed in the modern laboratory as the indexed
libraries used as template have already been PCR amplified. The steps were performed
following the standard myBaits® in-solution sequence capture protocols. The indexed
DNA libraries were used as templates with the total of 25 µL volume reduced to 7 µL
using the Eppendorf® centrifugal vacuum concentrator. In the first step, which is the
hybridization step, the DNA library templates are denatured into single strands, followed
by addition of adapter blockers, which allows the baits to hybridize to their
complementary targets. This was performed by preparing the hybridization mix and the
blockers mix into two separate Eppendorf LoBind 0.5 mL tubes. The hybridization
contained HYB#1, HYB#2, HYB#3, HYB#4, RNase block and the baits in 18.5 µL final
volume. Meanwhile, the blockers mix comprising BLOCK#1, BLOCK#2, BLOCK#3
and the 7 µL sequencing library were combined together in a separate 0.5 ml tube. The
tube containing the blockers mix was incubated in the thermal cycler at 95°C for 5
minutes to allow DNA library denaturation and the adapter blocking actions. Next, both
tubes containing the blockers mix and hybridization mix were incubated at the
hybridization temperature (60-65°C) for 5 minutes. After the 5 minutes, the
hybridization mix was then transferred to the blockers mix and the incubation resumed
for 36 hours to allow the baits to hybridize to the sequencing templates. The samples
described in Chapter 5 were hybridized at 65°C while the samples described in Chapter
4 were hybridized at 60°C.
After the 36 hours incubation, it is expected that the baits bound to the complementary
sequencing libraries will form bait-target hybrids. In the hybrid bind step, the solution
mix from the previous step will be combined with streptavidin-coated magnetic beads,
Dynabeads® MyOne™ Streptavidin C1 (ThermoFisher Scientific), which will bind to
the bait-target hybrids. The hybrids that are bound to the magnetic beads are pelleted in a
magnetic particle collector (MPC) while the non-hybrids (exogenous DNA) will remain
in the solution and will be removed in the washing steps. In this step, beads were first
washed three times with Binding Buffer which had been allowed to equilibrate to room
temperature prior to use. The washed beads were suspended in 70 µL Binding Buffer
before the capture reaction was transferred to it in the water bath. The capture reaction-
beads mix was incubated at the hybridization temperature for 30 minutes with occasional
tube agitation. Next the beads (which are now bound to the bait-target hybrids) were
pelleted in the MPC, the supernatant (which contains non-target DNA) was removed and
the beads were washed with Wash Buffer 2.2 that had already been incubated at the
hybridization temperature for at least 45 minutes before use. This step was repeated

78
three times before the beads were eventually eluted in 30 µL EB buffer. The post-
capture DNA libraries were amplified using the beads suspension solution as template.
A 35 µL PCR mix comprising 1 X KAPA HotStart ReadyMix (Roche) and 500 nM of
each IS5 and IS6 primer was added to the 15 uL post-capture template forming a 50 µL
post-capture PCR reaction. Two tubes for each sample were included in the
amplification. In the thermal cycler, the templates were pre-denatured at 98°C for 2
minutes followed by 8-14 cycles of denaturation at 98°C for 20 seconds, annealing at
60°C for 30 seconds and elongation at 72°C for 30 seconds. Lastly, the final elongation
was carried out at 72°C for 5 minutes. The amplified post-capture PCR products of the
same sample from the two tubes were combined together. The beads were pelleted in the
MPC and the supernatant was collected for purification with MinElute PCR purification
kit (Qiagen) following the standard protocols. The final elution volume in EB buffer
(Qiagen) was 20 µL. The protocols are repeated when performing double capture to
increase the yield even further, although caution should be taken as there may be some
loss of sequence targets during the procedures. The samples described in Chapter 4 were
subjected to double capture while the samples in Chapter 5 had only a single
hybridization capture performed.
2.6.3 Quality control and quantification of sequencing libraries
Quality control was performed after the indexing PCR (Section 2.6.1) for shotgun
sequencing or after the hybridization capture (2.6.2) for a target enriched library. The
NGS was performed using both the Illumina HiSeq 2500 and Illumina HiSeq 4000
sequencing techonology depending on the instrument availability at the Genomic
Technologies Core Facility (GTCF), at the University of Manchester. Adapter dimers
may affect the cluster efficiency and the subsequent reads obtained (Kircher et al. 2011).
Therefore, it is important to monitor the presence of adapter dimers in the sequencing
libraries. The presence of these adapter dimers was assayed using the Bioanalyzer 2100
(Agilent Technologies) instrument. The DNA High Sensitivity kit (Agilent
Technologies) allows the inspection of the fragment size distribution in the sequencing
DNA library. In case of a DNA library with high content of adapter dimers, the adapter
dimers were removed following the SPRIselect (Beckman Coulter) standard protocols.
The SPRI-based chemistry allows the selection of longer fragments while getting rid of
shorter ones (Figure 2.3). The sequencing library volume was adjusted to 50 µL. A 1.2x

79
ratio to template (60 µL) of SPRIselect (Beckman Coulter) was then added to the
sequencing library. The SPRI beads were pelleted in the MPC where the clear
supernatant (containing smaller fragments) was discarded. The pellets were washed with
180 µL of 85% ethanol, and the 85% ethanol then removed and the pellet allowed to dry.
The pellet was eluted in 20-25 µL EB buffer, placed in the MPC and the supernatant
kept as it contain the desired sequencing libraries.
Figure 2.3: The relationship between the SPRI beads-to-template ratios to fragment size
selection. The 1.2X SPRI beads-to-template ratio was used to filter out the adapter dimers from
the sequencing library in this study (figure taken from Beckman Coulter 2012).
The libraries from multiple samples were pooled together in an equimolar ratio before
being sent for sequencing. For this purpose, the sequencing libraries were quantified
with the qPCR based KAPA library quantification kit Illumina® platforms (Roche). Pre-
diluted DNA standards with 452 bp length, flanked by P5 and P7 primers (similar to the
primers in the Illumina flow cell) with concentrations from 20 pM to 2x10-4 pM were
used to generate a standard curve. The concentration of the DNA library was calculated
by plotting against the standard curve using absolute quantification methods. A dilution
series was prepared for the sequencing library and used as the template in a qPCR. The
reactions were prepared in a 96 well plate with each well containing a 20 µL reaction
comprising 1X LightCycler® 480 SYBR Green I Master (Roche Life Science), 200 nM
of each primer – IS5 and IS6, and 4 µL of diluted library template. At least duplicates
were prepared for both sample and standard reactions. The qPCR was performed in the
LightCycler® 480 Instrument II (Roche Life Science). The cycle conditions were: initial

80
denaturation for 5 minutes at 95°C; 35 x amplification cycles with denaturation at 95°C
for 15 seconds, annealing at 60°C for 25 seconds, and extension at 72°C for 35 seconds;
followed by melt curve analysis at 95°C for 5 seconds, 55°C for 1 minute, increasing to
97°C with 7 acquisition per °C. In the analysis, the absolute quantification method was
used to generate a standard curve from the DNA standards. The standard curve was
verified to have a ∆Cq value between 3.1 to 3.6, a reaction efficiency range of 90% to
110% ,and a R2 value ≥0.99. A standard curve with one criterion lying outside of the
range will require repeating as the library conecntration calculation will be doubted. In
the concentration calculation, the standard curve is used to convert the concentration of
diluted library to pM. Next, the average size-adjusted concentration was calculated for
each library dilution, and the concentration converted to nM. The average library
fragment length used in this calculation is obtained from the results of the Bioanalyzer
analsysis. Based on the concentration value (in nM) obtained for each library, pooled
sequencing reactions were prepared in final volume of 20 µL to be sent for NGS at he
Genomic Technologies Core Facility (GTCF) of the University of Manchester. The
samples described in Chapter 4, for shotgun sequencing, were run in an Illumina HiSeq
2500 with paired end sequencing at 100 bp length. The samples described in Chapter 4
for target capture sequencing and the samples described in Chapter 5 were run using the
Illumina HiSeq 4000 platform with paired end sequencing with 75 bp length on each
pair. The sequencing facility performed all the sequencing protocols and the raw data
were received in the form of demultiplexed fasta sequence files.
2.7 Bioinformatics analysis
The bioinformatics analysis was performed in a Linux-based operating system based on
Debian, Ubuntu 16.04. The starting raw data from the sequencer were obtained as
demultiplexed FASTQ files. The analysis flows that were performed are depicted in
Figure 2.4. Each sample was processed in a different pathway based on the outcome of
downstream analysis. The paired-end sequencing produced read 1 and read 2, one from
each end. These two reads can be aligned together, and depending on the length, they
can be merged or collapsed together forming a single collapsed read (Lindgreen 2012).
This allows better alignment to the reference sequence, improving the entire data set.
FASTQ file format has four lines with the first line containing the sequencer information
and sequence identifier, the second line showing the nucleotide sequence which could be

81
either A, C, T, G or N, the third line usually a plus (+) sign which acts as a separator,
and the fourth or the last line is the quality scores of each base (Cock et al. 2010). The
quality score is encoded in Phred +33 format. The quality score is an estimation of the
probability that the base called in that position is incorrect.
Figure 2.4: The bioinformatics analysis flows performed on different samples. Purple:
analysis performed on all samples. Green: analysis performed on samples described in Chapter
4. Pink: analysis performed on samples described in Chapter 5.

82
2.7.1 Merging of paired reads and removal of adapter sequences
The FASTQ data still contain the adapter sequences which will interfere with
downstream analysis such as mapping. Therefore, removing these is a crucial first step
in the data analysis. This was performed using the AdapterRemoval v2 program
(Schubert et al. 2016). This program not only trims the adapter sequences but it also
allows the merging or collapsing of two overlapping paired reads.
Figure 2.5: Three outcome from paired-end data from Illumina sequencing by
AdapterRemoval v2. (A) Pair 1 and pair 2 read mates are longer than 75 or 100 bp (depending
on the sequencer used) therefore do not overlap with each other, (B) Read sequence mates
overlap at minimum 5 bp will be collapsed forming a single sequence, (C) Read mates are both
overlap with each other and to the adapter sequence. Adapted from (Lindgreen 2012).
Paired-end sequencing reads a DNA fragment from both ends. The lengths of the reads
produced are 100 or 75 bp depending on the sequencer used. In the case of longer target
fragments, the read mates do not overlap with each other, and therefore are kept in
separate pair1 and pair2 FASTQ files (Figure 2.5-A). Read sequence mates that overlap
at 5 or more bases will be merged together forming a single collapsed sequence (Figure
2.5-B). The mismatch rate that is allowed within the overlapping region is 1/3; if this

83
rate is exceeded, the read mates will not be merged into a single collapsed read. In
Figure 2.5-A and B, there is no adapter contamination. In Figure 2.5:C, as the sequence
is very short, the read mates not only overlap with each other, they also overlap with the
adapter sequence causing adapter contamination. The two reads will be merged into one
collapsed read, and the adapter sequence contamination will be removed. However, if
the resulting collapsed sequence is less than 25 bp length it was removed. The stretches
of Ns and low quality bases from the 5’ and 3’ termini were trimmed (Schubert et al.
2016).
2.7.2 Mapping to reference genome
In the M. tuberculosis study, the M. tuberculosis ancestor genome (Comas et al. 2013)
was used as the reference, while in the M. leprae study the reads were mapped against
the M. leprae TN genome (Cole et al. 2001) sequence. The mapping was performed
using the Burrows-Wheeler Aligner 0.7.12 (bwa) programme (Li & Durbin 2009).
Firstly, the reference genome was indexed. Pairs 1 and 2 from the uncollapsed reads
were mapped separately to the reference genome, unlike the collapsed reads. The
resulting file is in Sequence Alignment/Mapping (SAM) format (Li et al. 2009). This
format stores read mapping information in respect to the reference genome. BWA-ALN
algorithm was used in the alignment with disabled seeding and maximum edit distance
of 0.1. Subsequently, BWA sampe and BWA samse was performed to generate the SAM
file from paired reads and collapsed/merged reads, respectively.
2.7.3 Cleaning and sorting reads (PicardTools)
PicardTools (http://broadinstitute.github.io/picard) CleanSam option was performed for
soft clipping. After that, the coordinates of the reads in respect to the reference genome
were added using SortSam of PicardTools. The mapped reads were extracted into a new
separate file using Samtools with the SAMtools view option. Read duplicates were
removed using the PicardTools MarkDuplicates option. After this step, the subsequent
analysis that was performed depended on the result obtained. If the number of reads
obtained is sufficient for genotyping analysis (Chapter 5), the programs used are
depicted in pink as shown in Figure 2.4 (will be described in section 2.7.5). Otherwise,
the analysis will be similar to what is depicted in green, as the metagenomics contents
identification will be performed instead (will be described in section 2.7.4).

84
2.7.4 Metagenomic content analysis
In the metagenome content analysis, only the collapsed reads were used in the analysis.
BEDTools (Quinlan & Hall 2010) was used for file conversion from BAM to Fastq files.
FASTQ files were converted to FASTA files using the Seqtk programme
(https://github.com/lh3/seqtk). Once the file has been converted to FASTA file, it was
then run in Basic Local Alignment Search Tools (BLAST) to identify the possible
identity of each read against the NCBI database. The BLAST output was then visualized
in MEtaGenome Analyzer 5 (MEGAN 5) (Huson et al. 2007) to identify the taxonomic
profile of each sample. In MEGAN 5, the identification of a sample’s taxonomical
content is estimated based on the lowest common ancestor (LCA) concept. In this
concept, if multiple taxa are found in the BLAST search for one read, the lowest shared
ancestor will be assigned as the taxon identity for this read. The LCA parameters were
set to min score: 0; max expected: 1.0e-7; top percent: 0; min support percent: 0; min
support: 1; LCA percent: 100. This is the last step in the analysis for the samples studied
in Chapter 4. For the analysis performed in Chapter 5, the subsequent steps will be
described in section 2.7.5.
2.7.5 Sequence variant analysis
To study the sequence variants in each sample, the BAM file containing reads mapped to
the reference genome is converted to a FASTA file and the sequence identity determined
by BLAST. Once the possible identity of each read is identified in BLAST, the resulting
file was visualized in MEGAN to view the taxonomic content. Using an in house perl
script, a FASTA file which only contains the reads assigned to M. leprae in MEGAN
was extracted from the original mapped read BAM file. The in house script was also
used to prepare the FASTQ file for subsequent analysis with the Genome Analysis
Toolkit (GATK) 3.6 (McKenna et al. 2010). Base Quality Score Recalibration (BQSR)
was performed in GATK 3.6 before the recalibrated BAM file was visualized in
Geneious® 8.1 (Kearse et al. 2012) for polymorphism analysis. Unique polymorphisms
in the microbial isolates were confirmed if the read has at least 5-fold depth in that
position and variant frequency of at least 80%.

85
Chapter 3: Study of M. tuberculosis aDNA in
archaeological remains from Yorkshire, England.
Part I: MTBC aDNA screening by polymerase chain reaction.
3.1 Introduction
Chapter 3 and Chapter 4 focus on the biomolecular study done on 60 archaeological
skeletal remains originating from 16 different locations in Yorkshire, England. The aim
is to determine whether the success rate in MTBC aDNA detection is high enough to
make it worthwhile to plan a larger project to test hypotheses such as possible strain
differences in urban and rural areas. Chapter 3 describes the preliminary study
undertaken to screen for the presence of MTBC aDNA in each of the bone samples
using PCR assay and Sanger sequencing. Based on the PCR assay results, the samples
deemed positive for MTBC aDNA preservation were subjected to NGS – which is the
focus in Chapter 4.
York – the location of origin for 32 of the samples – was a thriving city during the
Roman era. It was founded by the Roman Ninth Legion (Legio IX Hispana) in ca. 71 AD
and they remained there until the 4th century AD (Ottaway 2004). After the Romans had
left, the regional context of York was left blank until 627 AD (Palliser 2014). Not much
is known about this city for three centuries, from around 400 to 700 AD. Although the
city was not completely abandoned, its urbanization had definitely shrunken by the fifth
century AD. In the late sixth century AD, immigrants who were coming across the North
Sea, known as the Angles or English, occupied York districts and this period is also
known as the Anglo-Saxon period. Fast forwards to 866 AD, the town was captured by
the Vikings who decided to remain and settle there; the name of the town was later
changed to Jórvík (Addyman 1980). Jórvík became the capital of a new Viking kingdom
in 870s to 980s AD, resulting in the growth of York into a bigger town than it was
previously (Palliser 2014). During the ‘Viking period’, Jórvík was more heavily
populated following the development of a large commercial zone located between the
two important rivers in the town and the fortress (Palliser 2014). The Normans
conquered York in the 11th century AD and it was made the northern operations base by
William the Conqueror (Bartlett 2000). York was a thriving port and successful
manufacturer during the mediaeval period and was well-connected to Europe. This

86
attracted many other craftsmen to live in York. In addition, hospitals were run by the
church during the mediaeval period in York.
However, the arrival of the Black Death in 1349 AD caused a drastic decline in the
population of York (Nuttgens 2007). Despite the plagues which struck this town several
more times, York remained as one of the most important towns in the northern region of
England until the 16th century AD. York ceased to be the most important town in the 17th
century AD due to competition from neighbouring places (Sheahan & Whellan 1855;
Sellers 1987). Nevertheless, York retained its importance especially in the middle 18th
century AD (Sheahan and Whellan 1855). The industrial revolution resulted in the
booming of other towns in Yorkshire. During the beginning of the 19th century, York
remained as a market town and main distributing hub for its rural hinterlands
(Armstrong 1974). It was not until towards the middle 19th century that the population in
York grew rapidly again when it became the centre of railway transportation. The
housing condition was overcrowded, especially after the arrival of Irish immigrants who
escaped the potato famine (MacRaild 1999). The living conditions are imagined to be
overcrowded, unsanitary and dirty. The population continued to grow rapidly but the
living conditions were better in the 20th century AD due to clearing efforts done by the
York council (Rawnsley & Singleton 1995).
Based on the above history, York did undergo rapid urbanization in the past.
Urbanization has been suspected to assist the spread of tuberculosis or even to promote
the evolution to more aggressive MTBC strains (Comas & Gagneux 2011). The
inclusion of both urban and rural skeletal remains provides an opportunity to attempt
sequence comparisons between the two origins of MTBC isolates.

87
3.2 Results
The summary of the PCR results is depicted in Table 3.1. The positive and negative
detections given in the table refer to the agarose gel observations. The presence of a
band of similar or near-exact to the expected length is marked positive. For the samples
without any bands or unspecific amplification, they are marked as negative. All the
bands with sizes similar or near-exact to the expected length were cloned and Sanger
sequenced. The resulting sequences were compared to the M. tuberculosis H37Rv
reference sequence. The sequence that matches the reference sequence is deemed
positive and vice versa for the sequence that did not match the reference sequence. This
result is indicated in the bracket next to the positive/negative mark for the agarose gel
band result (Table 3.1).
Fifteen samples from 9 different locations produced a band with size similar or near to
the expected band size. All samples from York Minster, Fishergate House, 3 Driffield
Terrace, 6 Driffield Terrace, St Peter Huddersfield and Ailcy Hill produced negative
PCR amplification for all four PCR targets. The results for the suspected PCR positive
samples are further described below and the Sanger sequencing result is summarised in
Table 3.2.

88
Sample
origin
Sample
number
IS6110 gyrA Rv0083 Pks 15/1
123bp 92 bp
Undiluted
DNA
1:10
diluted
DNA
Undiluted
DNA
1:10
diluted
DNA
Undiluted
DNA
1:10
diluted
DNA
Undiluted
DNA
1:10
diluted
DNA
Undiluted
DNA
1:10
diluted
DNA
York Minster 15 - - - - - - - - - -
1 - - - - - - - - - -
Fishergate
House
147 - - - - - - - - - -
98 - - - - - - - - - -
86 - - - - - - - - - -
149 - - - - - - - - - -
108 - - - - - - - - - -
135 - - - - - - - - - -
St Andrew
Fishergate
6 -a/+(+)b +(+)a/+(+)b +(+)a/+(+)b +(+)a/+(+)b -a/-b -a/+(-)b -a/-b -a/-b -a/-b -a/+(+)b
253 - - - - - - - - -
131 - - - - - - - - - -
277 - - - - - - - +(-) - -

89
34 - - - - - - - - - -
323 - - - - - - - - - -
296 - - - - - - - - - -
384 - - - - - - - - - -
339 - - - - - +(-) - - - -
286 - - - - - - - - - -
St Helen on
the Walls
5494 - - - - - - - - - -
6003 - - - - +(-) n.d. - - - -
5000 - - - - - - - - - -
5844 - - - - - - - - - -
East
Heslington
229 - +(-) - - - - - - - -
3 Driffield
Terrace
15 - - - - - - - - - -
37 - - - - - - - - - -
13 - - - - - - - - - -
54 - - - - - - - - - -
6 Driffield 19 - - - - - - - - - -

90
Terrace 22 - - - - - - - - - -
St Peter
Huddersfield
5 - - - - - - - - - -
17 - - - - - - - - - -
7 - - - - - - - - - -
Wetwang
Slack
1 - - - - - - - - - -
2 - - - - - +(-) - - - -
3 - - - - - - - - - -
6 - - - - - - - - - -
4 - - - - - - - - - -
5 - - - - - - - - - -
7 +(-) - - - - - - - - -
8 - - - - - - - - - -
9 - - - - - - - - - -
185 - - - - - - - - - -
360 - - - - - - - - - -
415 - - - - - - - - - -

91
Sewerby 44 - - - - - - - - - -
34 +(n.d.) +(n.d) - - - - - - - -
St Giles by
Brompton
Bridge
1288 - - - - - - - - - -
1531 - - - - - - - - - -
1542 - +(-) - - - - - - - -
Ailcy Hill 1043 - - - - - - - - - -
1044 - - - - - - - - - -
Wharram
Percy
26 - - - - +(-) - - - - -
1600 - - - - +(-) - - - - -
Addingham 103 - - - - - +(-) - - - -
134 - +(-) - - - - - - - -
223 - +(-) - - - - - - - -
Melton 4297 - - - - - - - - - -
5319 - - - - +(-) - - - - -
2554 - - - - - - - - - -
Hickleton 46 - - - - +(-) - - - - -

92
Table 3.1: Result summary for all PCR assays tested on all samples, using original DNA extract and 10-fold diluted DNA.
+ PCR band similar or close to the size of the expected PCR product length observed in gel electrophoresis (single band or multiple bands)
- No band produced, or the size of the produced band/bands is not similar to the length of the expected PCR product
n.d. Not done, (+) Sanger sequence identity matched the M. tuberculosis H37Rv reference sequence
(-) Sanger sequence identity did not match the M. tuberculosis H37Rv reference sequence
(a) First extraction; (b) second extraction - only for those samples with two extractions performed

93
Sample PCR assay Fragment length (bp) Number of mismatches to the reference genome
(bp)
Possible sequence identity by BLAST Pairwise identity E-value
St Andrew Fishergate 6
IS6110 123 bp 123 0-1 n.d. n.d. n.d.
IS6110 92 bp 92 0 n.d. n.d. n.d.
gyrA 104 6 Mycobacterium peregrium partial gyrA sequence
98.40% 1.33 e-21
Pks 15/1 91 3-4 M. tuberculosis and M. bovis 92.20% 5.76 e-11
St Andrew Fishergate 277
Rv0083 109 n.a. Nocardioides sp. JS614 80.00% 1.31 e-4
St Andrew Fishergate 339
gyrA 104 6 Partial gyrA gene from an uncultured bacterium: ancient DNA from St Peter's
Collegiate Church 28-a,b isolate
96.90% 1.62 e-20
St Helen on the walls 6003
gyA 104 8-10 Mycobacterium vaccae 95.30% 8.61 e-19
East Heslington 229
IS6110 123 bp 123 16 Unidentified partial IS6110-like insertion sequence; isolate IS6110-like sequence type
B
100.00% 3.9 e-23
Wetwang Slack 2 gyrA 104 11 Uncultured bacterium partial gyrA gene of St Shchekavitsa 8-a-e ancient isolates
90.60% 4.3 e-15
Wetwang Slack 7 IS6110 123 bp 118 14 (and 5 bp gaps) Unidentified partial IS6110-like insertion sequence; isolate IS6110-like sequence type
B
91.50% 4.60 e-23

94
Wharram Percy 26
gyrA 104 7 Mycobacterium chelonae 98.40% 1.33 e-21
Wharram Percy 1600
gyrA 104 9 Mycobacterium chelonae 95.30% 6.90 e-19
Addingham 134 IS6110 123 bp 123 16 Unidentified partial IS6110-like insertion sequence; isolate IS6110-like sequence type
B
98.80% 6.85 e-31
Addingham 223 IS6110 123 bp 123 16 Unidentified partial IS6110-like insertion sequence; isolate IS6110-like sequence type
B
98.80% 3.68 e-31
Addingham 103 gyrA 104 6 Mycobacterium peregrinum 98.40% 1.35 e-21
Melton 5319 gyrA 104 8 and 12 Variant 1 and 2: Mycobacterium chelonae Variant 1: 96.9%, Variant 2: 90.9%
Variant 1: 1.62 e-20 , Variant 2: 4.36 e-15
Hickleton 46 gyrA 104 8 Mycobacterium chelonae 96.90% 1.62 e-20
St Giles by Brompton Bridge
1542
IS6110 123 bp 134 49 Streptomyces lunaelactis 97.80% 2.72 e-35
Table 3.2: Sanger sequence result summary for 15 samples with a positive band in at least one of the PCR assays.
n.d. Not done. BLAST search was not performed as the product sequence completely matches the reference genome.
n.a. Not aligned. The sequence cannot be aligned to the reference genome in Geneious due a large number of mismatches.

95
3.2.1 St Andrew Fishergate 6
This sample produced a positive PCR amplification in the second nested step of the
IS6110 assay when the original undiluted DNA template was used (Figure 3.1-a).
Positive amplification was achieved in both the first and nested step of IS6110 where
the 10-fold diluted DNA was used as template (Figure 3.1-a). In this extraction, the
other three single-copy target PCR assays produced negative amplification (not shown).
A second DNA extraction was performed; positive amplification was observed in the
first and nested step of the IS6110 assay in both instances where undiluted and 10-fold
diluted DNA were used as templates (Figure 3.1-b). Positive amplification was
obtained from 10-fold diluted DNA in the gyrA PCR assay but not from the undiluted
DNA (Figure 3.1-b). A very faint band was shown in the agarose gel from the 10-fold
diluted DNA amplification for the Pks 15/1 PCR assay, and similar to the gyrA assay,
no band was produced when the undiluted DNA was used as template (Figure 3.1-b).
The Sanger sequences of the cloned IS6110 PCR product when aligned to the M.
tuberculosis H37Rv reference genome revealed a complete match in 5 clone sequences,
and one mismatch each at inconsistent positions in 4 clones for the first extraction. On
the other hand, all IS6110 nested PCR sequences show a complete match to the
reference genome (Figure 3.2-a). For the second St Andrew Fishergate 6 extraction, all
clone sequences for both the IS6110 123 bp and nested PCRs show a complete match
to the reference genome with no mismatch observed (Figure 3.2-b). The sequence
identities are also supported by BLAST analysis. The 3 clone sequences from the gyrA
PCR assay of the second DNA extraction revealed 6 mismatches from the 64 bp
sequence (after trimming of primers) in consistent positions for all the clones (Figure
3.3-a). BLAST search revealed the highest sequence similarity to Mycobacterium
peregrium partial gyrA sequence (Table 3.2). Meanwhile, the clone sequences from
Pks 15/1 PCR amplification of St Andrew Fishergate 6 second extraction contained
three consistent mismatches in all clones, and another one shared mismatch in 2 clones
(Figure 3.3-b). Based on the BLAST search result, the highest sequence match was
shown against M. tuberculosis and M. bovis (Table 3.2).

96
M A B C D
100 bp
200 bp
IS6110 - 123 bp IS6110 - 92 bp
M A B C D E F G H
200 bp
100 bp
IS6110 - 123 bp IS6110 - 92 bp gyrA Pks 15/1
Figure 3.1: Gel electrophoresis results for the sample St Andrew Fishergate 6 showing
positive bands for three markers. (a) DNA extraction 1. A and C: undiluted DNA as
template; B and D: 10-fold diluted DNA as template. (b) DNA extraction 2. A, C, E and G:
undiluted DNA as template, B, D, F and H: 10-fold diluted DNA as template. In both 1 and 2,
M: DNA ladder.
(a)
(b)

97
Figure 3.2: The alignment of St Andrew Fishergate 6 Sanger clone sequences of IS6110 123 bp and nested 92 bp PCR product against M. tuberculosis
H37Rv reference sequence. (a) St Andrew Fishergate 6 DNA extraction 1, (b) St Andrew Fishergate 6 DNA extraction 2. Primer sequences are omitted from
the alignment. Mismatches are highlighted in blue.
(a)
(b)

98
Figure 3.3: The alignment of St Andrew Fishergate House 6 Sanger clone sequences of gyrA and Pks 15/1 PCR product against M. tuberculosis H37Rv
reference sequence. (a) gyrA and (b) Pks 15/1 DNA extraction 2. Primer sequences are omitted from the alignment. Mismatches are highlighted in blue
(a)
(b)

99
3.2.2 St Andrew Fishergate 277
Skeleton St Andrew Fishergate House 277 produced multiple bands in the Rv0083
assay, one with a length close to the expected 110 bp PCR product from this assay
(Figure S3.1). The faint lower band was cut from the agarose gel, purified and cloned
into competent E. coli cells. The clone Sanger sequences show a PCR product of 109
bp, shorter than the expected fragment length (not shown). The alignment to the
reference M. tuberculosis H37Rv sequence shows only 74.2% pairwise identity.
BLAST search revealed that Nocardioides sp. JS614 as the closest match to the clone
sequences. However, the e-value for this match is only 1.31e-4 (Table 3.2).
3.2.3 St Andrew Fishergate 339
St Andrew Fishergate produced a positive band in the gyrA PCR assay, while showing
negative amplifications in the other PCR assays. The positive band was produced from
the amplification of the 10-fold diluted DNA template. It was faint (Figure S3.2) but
had an apparent length close to 104 bp. However, the sequence length spanned by both
forward and reverse primers had 6 mismatches with the reference sequence (Figure
3.4). The possible identity of the amplified sequence based on BLAST analysis is the
partial gyrA gene from an uncultured bacterium: ancient DNA from St Peter’s
Collegiate Church 28-a,b isolate (Müller et al. 2016).
Figure 3.4: The alignment of St Andrew Fishergate House 339 clone sequence of gyrA
PCR product against the reference M. tuberculosis H37Rv sequence. Primer sequences are
omitted from the alignment. Mismatches are highlighted in blue.

100
M A
200 bp
100 bp
3.2.4 St Helen on the walls 6003
Figure 3.5: gyrA PCR amplification for sample St Helen on the Walls 6003. A: undiluted
DNA template. M is the DNA ladder with the 100 bp and 200 bp marked in the figure. Faint
lower band is shown inside the red box.
St Helen on the Walls produced double bands in the gyrA assay when undiluted DNA
was tested, one of which with had a size close to the expected gyrA PCR product
(Figure 3.5). The 10-1 DNA dilution was not tested for this particular assay for this
sample. This sample tested negative for the other PCR assays. Clone sequences
revealed a 104 bp PCR product, which matched the expected product length of the
gyrA target. However, the sequence did not correspond to the reference M. tuberculosis
sequence. There are 8 consistent mismatches throughout all 9 clones, and one mismatch
that is present in 6 clones (Figure 3.6). Two clones show one mismatch each, in
different positions. BLAST search shows a closest match to Mycobacterium vaccae
(Table 3.2).

101
Figure 3.6: The alignment of St Helen on the Walls 6003 clone sequences from the gyrA PCR assay to the M. tuberculosis H37Rv reference sequence.
The reference sequence is shown at the top, while the mismatches are highlighted in blue. Primer sequences are omitted from the alignment and BLAST search.
Figure 3.7: The alignment of East Heslington 229 clone sequences from the IS6110 123 bp PCR assay to the M. tuberculosis H37Rv reference sequence.
The reference sequence is shown at the top, while the mismatches are highlighted in blue. Primer sequences are omitted from the alignment and BLAST search.

102
3.2.5 East Heslington 229
Skeleton East Heslington 229 produced multiple bands in the first step of the IS6110
PCR assay. The lowest band as marked in Figure S3.3 shows a faint possible positive
amplification. However, the subsequent IS6110 nested 92 bp PCR assay did not
produce positive amplification. The lowest band from the first step PCR assay was
excised from the gel, cloned and Sanger sequenced. Clone sequences revealed a
product with 123 bp length, however the sequence did not match the reference M.
tuberculosis H37Rv. There are 16 mismatches within the region spanned between the
forward and reverse primers, consistently found across all clones (Figure 3.7). The
closest match of this sequence is to the unidentified partial IS6110-like insertion
sequence; isolate IS6110-like sequence type B, which is reported in Müller et al.
(2015).
3.2.6 Wetwang Slack 2
The Wetwang Slack 2 skeleton produced multiple bands for the gyrA PCR
amplification, in which the length of the lower band can be approximated to 104 bp
(Figure S3.4). The other PCR assays tested for this sample produced negative
amplification – absence of band or bands with differing length to the expected product
size. The amplicon produced by the Wetwang Slack 2 sample is 104 bp as confirmed
by the clone Sanger sequences. However, the sequence does not correspond to the
reference. There are 11 mismatches to the reference within the 64 bp sequence – after
the primers are removed (Figure 3.8). Based on the BLAST search result, the closest
sequence match is to the uncultured bacterium partial gyrA gene of St Shchekavitsa 8-
a-e ancient isolates which was reported by Müller and colleagues (2015).

103
Figure 3.8: The alignment of Wetwang Slack 2 clone sequences from the gyrA PCR assay against the M. tuberculosis H37Rv reference sequence.
Disagreements to the reference sequence are highlighted in blue. Forward and reverse primer sequences are omitted from this figure as well as in the BLAST
search.
Figure 3.9: The alignment of Wetwang Slack 7 clone sequences from the IS6110 123 bp PCR assay against the M. tuberculosis H37Rv reference
sequence. Disagreements to the reference sequence are highlighted in blue while the gaps are shown in red. Forward and reverse primers are omitted from the
alignment and for the BLAST search.

104
3.2.7 Wetwang Slack 7
The undiluted DNA of the sample Wetwang Slack 7 produced a band with a size that
matches the expected 123 bp product with undiluted DNA as the template (Figure
S3.5). The subsequent nested PCR did not produce positive amplification. The clone
Sanger sequence however confirmed that the amplicon size is 118 bp, 5 bp shorter than
the expected product (Figure 3.9). The closest identity match of this sequence is to the
unidentified partial IS6110-like insertion sequence, isolate IS6110-like sequence type B
(Müller et al. 2015). Wetwang Slack 7 produced negative amplification for the IS6110
nested 92 bp PCR assay and the other 3 targets.
3.2.8 Wharram Percy 26 and Wharram Percy 1600
Both sample Wharram Percy 26 and Wharram Percy 1600 showed positive
amplification for the gyrA target (Figure S3.6) while showing negative amplification
for the other three targets. The positive amplification was obtained from the undiluted
DNA as template. The clone Sanger sequences revealed a 104 bp amplicon from both
samples, similar to the expected product length. However, the nucleotide sequences do
not correspond to the reference sequence. Wharram Percy 26 clone sequences show 7
mismatches to the reference sequence (Figure 3.10). The possible sequence identity
determined by BLAST is matched to Mycobacterium chelonae. The Wharram Percy
1600 clone sequence contain 9 mismatches against the reference sequence (Figure
3.11). Similar to Wharram Percy 26, the best BLAST match was assigned to M.
chelonae.

105
Figure 3.10: The alignment of Wharram Percy 26 clone sequences from the gyrA PCR assay against the M. tuberculosis H37Rv reference sequence. The
disagreements to the reference sequence are highlighted in blue. The forward and reverse primer sequences were trimmed from the alignment and were not
included in the BLAST search query.
Figure 3.11: The alignment of Wharram Percy 1600 clone sequences from the gyrA PCR assay against the M. tuberculosis H37Rv reference sequence.
The disagreements to the reference sequence are highlighted in blue. The forward and reverse primer sequences were trimmed from the alignment and were not
included in the BLAST search query.

106
3.2.9 Addingham 134 and Addingham 223
Both of samples Addingham 134 and Addingham 223 gave positive amplification in
the IS6110 first step, 123 bp PCR assay using the 10-fold diluted DNA as template
(Figure S3.7). The samples amplified multiple bands, with one of the bands showing a
length close to the expected amplicon size. These samples, however, produced negative
amplification in the subsequent nested IS6110 PCR assay and in PCR of the other 3
targets. Sanger sequences of product clones verified the amplicon size of 123 bp from
both samples, matching the expected fragment size. However, the nucleotide sequences
did not match the M. tuberculosis H37Rv reference. Both Addingham 134 and
Addingham 223 show 16 mismatches within the 83 bp amplicon (primer sequences
omitted) (Figure 3.12). BLAST search suggested the unidentified partial IS6110-like
insertion sequence, isolate IS6110-like sequence type B as the possible identity of the
amplicons from both samples.
3.2.10 Addingham 103
The sample Addingham 103 showed positive amplification in the gyrA PCR assay
using the 10-1 diluted DNA as template (Figure S3.8). This sample produced negative
amplification for the other targets. Sanger sequences from clones confirmed the
presence of a 104 bp amplicon which matches the expected product length. The
nucleotide sequences, however, do not correspond to the M. tuberculosis H37Rv
reference as there are 6 mismatches within the region spanned between the forward and
reverse primers (Figure 3.13). BLAST search revealed Mycobacterium peregrinum as
the possible sequence identity with 98.4% pairwise identity and e-value of 1.35e-21.

107
(a)
(b)
Figure 3.12: The alignment of Addingham 134 and Addingham 223 clone sequences from the IS6110 first step, 123 bp PCR assay against the M.
tuberculosis H37Rv reference sequence. (a) Addingham 134, (b) Addingham 223. The disagreements to the reference sequence are highlighted in blue.
Forward and reverse primer sequences are omitted from this figure as well as in the BLAST search.

108
Figure 3.13: The alignment of Addingham 103 clone sequences from the gyrA PCR assay against the M. tuberculosis H37Rv reference sequence. The
disagreements to the reference sequence are highlighted in blue. Forward and reverse sequence are omitted from this figure as well as in the BLAST search.
Figure 3.14: The alignment of Melton 5319 clone sequences from the gyrA PCR assay against the M. tuberculosis H37Rv reference sequence. The
disagreements to the reference sequence are highlighted in blue. Forward and reverse sequence are omitted from this figure as well as in the BLAST search.

109
3.2.11 Melton 5319
The amplification of the Melton 5319 undiluted DNA sample produced a faint band in
the agarose gel with length that matches the expected amplicon length (Figure S3.9).
The presence of a 104 bp product is confirmed by the clone Sanger sequences. Two
sequence variants are produced: one with 8 mismatches and one with 12 mismatches
against the reference sequence (Figure 3.14). Both do not correspond to M.
tuberculosis. The most possible identity for both sequence variants is Mycobacterium
chelonae as determined by BLAST search. The first sequence variant (with 8
mismatches) has 96.9% pairwise identity with 1.62e-20 e-value, while the second
sequence variant (with 12 mismatches) has 90.9% pairwise identity with 4.36e-15 e-
value.
3.2.12 Hickleton 46
Positive amplification of the gyrA target was produced from undiluted DNA of the
Hickleton 46 sample (Figure S3.10). A faint band at the expected position can be
observed after the amplification of the 10-1 diluted DNA of this sample. No positive
amplification was obtained from the other PCR assays. Sanger sequencing from clones
revealed the presence of a 104 bp amplicon, matching the expected PCR product
length. The nucleotide sequences, however, do not correspond to the M. tuberculosis
H37Rv reference as there are 8 mismatches within the region spanned between the
forward and reverse primers (Figure 3.15). The BLAST search suggests M. chelonae as
the most possible identity of the sequence rather than M. tuberculosis. This claim is
supported by 96.9% pairwise identity and 1.62e-20 e-value: the probability that the
matches occurred by chance.

110
Figure 3.15: The alignment of Hickleton 46 clone sequences from the gyrA PCR assay against the M. tuberculosis H37Rv reference sequence. The
disagreements to the reference sequence are highlighted in blue. Forward and reverse sequence are omitted from this figure as well as in the BLAST search.

111
IS6110
123 bp
IS6110
Nested 92 bp
M A B C D
200 bp
100 bp
3.2.13 St Giles by Brompton Bridge 1542
The sample St Giles by Brompton Bridge produced multiple bands in the IS6110 123 bp
PCR assay; one of the bands produced from the 10-1 DNA dilution template had a band
length close to the expected size (Figure S3.11). Sanger sequences revealed the amplicon
size to be 134 bp, which is 11 bp larger than the expected PCR product. In the alignment
against the M. tuberculosis H37Rv sequence, 49 disagreements were found, including
gaps (not shown). The closest sequence match in a BLAST search was Streptomyces
lunaelactis (Table 3.2). This bacterium has previously been isolated from moonmilk
speleothem from a cave (Maciejewska et al. 2015).
3.2.14 Sewerby 34
Figure 3.16: IS6110 123 bp and nested 92 bp PCR amplification of sample Sewerby 34. A
and C: undiluted DNA template, B and D: 10-1 diluted DNA template. M is showing the DNA
ladder with 100 bp and 200 bp bands indicated in the figure.

112
Positive amplification seemed to be produced from both the undiluted and 10-1 diluted
DNA template of Sewerby 34 for the IS6110 123 bp assay (Figure 3.16). However, the
sequence identity cannot be verified as the product could not be cloned. The cloning was
not pursued further as the nested PCR did not produce any positive amplification;
therefore, it is unlikely that the identity is MTBC.
3.3 Discussion
3.3.1 MTBC positive samples
In this study, a sample is considered MTBC-positive if positive amplification supported
by a Sanger sequence that matches the M. tuberculosis reference is obtained in at least
one of the assays. Exception is made for the small number of mismatches that can be
accounted for as miscoding lesions in the aDNA targets (Müller et al. 2014a). From the
60 archaeological remains studied, only seven of them produced positive amplification
in the first step of the IS6110 PCR assay: St Andrew Fishergate 6, East Heslington 229,
Wetwang Slack 7, Addingham 134, Addingham 223, St Giles by Brompton Bridge 1542
and Sewerby 34. In the subsequent nested PCR of this target, only St Andrew Fishergate
6 produced positive amplification, with a match to M. tuberculosis identity supported by
Sanger sequencing. The other six samples gave products that did not match the reference
sequence, albeit the product length was similar to the expected amplicon length with the
exception of Wetwang Slack 7 and St Giles by Brompton Bridge 1542. These other six
samples also produced negative amplification in the nested PCR, which can be attributed
to the dissimilarity of the sequence produced from the first IS6110 PCR step.
The results of the MTBC preservation screening therefore suggested that only one of the
60 samples contain MTBC aDNA: St Andrew Fishergate 6. The positive band from both
steps of the IS6110 PCR assay was confirmed as MTBC through Sanger sequencing.
There was one nucleotide discrepancy in each of the three clones for the 123 bp product
which is likely to be a result of miscoding lesions which are common in ancient DNA
templates (Brown & Brown 2011; Dabney, Meyer & Pääbo 2013). These mismatches
were only present in three of the clones and occurred at inconsistent positions. In

113
addition, these mismatches did not appear in the clone sequences derived from the
nested PCR amplicon. Therefore, the nucleotide discrepancies are very unlikely to be
true polymorphisms. This is further supported by the additional presence of clone Sanger
sequences for the amplicons from both steps of the IS6110 PCR from the second DNA
extraction which completely matched the reference sequence without any presence of
polymorphisms. The 7 bp deletion in the Pks15/1 region was also detected in this
sample. The absence of these 7 nucleotides is associated with the Euro-American lineage
of M. tuberculosis and has also been found in other ancient isolates (Bouwman et al.
2012). Three mismatches appeared in each of the clone sequences, which again can be
attributed to miscoding lesions (Müller et al. 2014a). Interestingly all the mismatches are
C → T substitutions which is a hallmark attribute of an ancient miscoding lesion.
Hydrolytic deamination will result in nucleotide modification, resulting in errors when
read by DNA polymerase during DNA replication (Dabney, Meyer, Pääbo 2013).
Cytosine is the primary target of such modification: deamination, which will form uracil
that will further incorporate adenine during the DNA duplication. This will cause either
a C → T or G → A substitution depending on which strand of the DNA is being
amplified (Dabney, Meyer, Pääbo 2013). In the gyrA PCR of St Andrew Fishergate 6,
however, despite the length of PCR product matching that of the expected amplicon, the
sequence did not agree with the M. tuberculosis reference. Instead, the most likely
identity as determined by BLAST analysis is to M. peregrinum. This sequence could
therefore have originated from a contaminant. M. peregrinum is a member of the
Mycobacterium fortuitum complex; one of the members of the rapidly growing non-
tuberculous mycobacteria. The M. fortuitum complex have previously been isolated from
the environment such as soil and water-related sources (Nagao et al. 2009).
3.3.2 Contamination
Three samples produced ambiguous results in the first round of amplification directed at
the insertion sequence IS6110. These samples produced near-exact length amplicons,
which is an indication of MTBC presence: the samples are East Heslington 229,
Addingham 134 and Addingham 223. However, all the samples failed to produce
positive amplification in the subsequent nested PCR round. Sanger sequences from all
three samples are similar to the MTBC reference sequence, but with 16 mismatches; the
positions of these mismatches consistent within the clones from each individual sample.

114
It is tempting to associate the similarity to cross-contamination between these samples.
However, firstly, except for Addingham 134 and Addingham 223, East Heslington 229
was processed in a different DNA extraction and PCR screening batch. Therefore, it is
unlikely that cross-contamination occurred in samples that were handled at different
times. In addition, all three samples produced multiple bands in the gel electrophoresis,
but the numbers and sizes of the unspecific bands were inconsistent across all 3 samples.
The closest match for these three sequences, according to the BLAST results, is the
unidentified partial IS6110-like insertion sequence, sequence type B, which was reported
by Müller et al. (2015). It can be argued that the sequence might have arisen from
contamination as the samples described by the study mentioned were processed in the
same facility. However, the sequence match is not 100% identical to the previously
reported sequence type B. All three samples show one mismatch relative to the sequence
type B, with the mismatch for Addingham 134 and Addingham 223 located in the same
position, but not in the similar position for sample East Heslington 229. It is possible
that the sequence is a product of a contaminant present in the burial environments, which
would explain the sequence similarity for the samples collected from Addingham but not
from East Heslington (Yang & Watt 2005).
The IS6110 PCR for samples Wetwang Slack 7 and St Giles by Brompton Bridge 1542
produced amplicons of 118 bp and 134 bp, respectively. The closest match for the
sequence from the Wetwang Slack 7 sample is the unidentified IS6110-like sequence
type B. Other than the 5 bp deletion in Wetwang Slack 7, there is only one mismatch
found compared to the unidentified IS6110-like sequence type B. Meanwhile, the
sequence from the sample St Giles by Brompton Bridge 1542 matched to Streptomyces
lunaelactis. This actinobacterium species was previously isolated from moonmilk
deposit of a cave (Maciejewska et al. 2015). Bacteria from the genus Streptomyces have
been isolated from various environmental sources worldwide, where they dominate soil
populations; 106 to 109 Streptomyces cells can be isolated from a gram of soil (Barka et
al. 2016).
Furthermore, eight samples produced ambiguous results for the gyrA PCR. Similar to
the samples discussed previously, the correct-length amplicons have different sequences
compared to the M. tuberculosis reference. Two of the samples best matched to the
unspecific gyrA sequence obtained from ancient isolates previously reported by Müller
and colleagues (2015). The samples St Andrew Fishergate 339 and Wetwang Slack 2
gave sequences that are similar but not identical to the gyrA amplicon sequence obtained

115
from ancient DNA from St Peter’s Collegiate Church 28-a-b and ancient DNA from St
Shchekavitsa 8-a-e, respectively. Although the ancient isolates which both samples were
best matched to were processed in the same facility, the sequences are not identical, with
at least 2 mismatches in each sample. The other six samples (St Helen-on-the-Walls
6003, Wharram Percy 26, Wharram Percy 1600, Addingham 103, Melton 5319 and
Hickleton 46) all produced amplicons with sequence best match to mycobacterium
species. The three species the samples matched to are M. vaccae, M. peregrinum and M.
chelonae. All three mycobacterium species are classified as mycobacterium other than
tuberculosis (MOTT) (Ho et al. 2012; Thomson et al. 2013). M. vaccae is a natural soil
non-pathogenic bacterium (Ho et al. 2012). The contamination of environmental bacteria
is entirely plausible as the skeletons were left in contact with soil during the preservation
years (Wilbur et al. 2009). Meanwhile, M. chelonae and M. peregrinum are both part of
the rapidly growing mycobacteria (RGM) group (Fernández-Roblas et al. 2000). M.
chelonae, although typically found in the environment, in water and soil, is also shown
to be able to cause clinical infection in humans. In fact, in the contemporary era, this
bacterium has been shown as a contaminant on hospital equipment (Shimoide et al.
1995). The clinical manifestation of this bacterium infection include skin and soft tissue
infections and, sometimes, pulmonary diseases and infections in joints, bones and
muscles in immunosuppressed patients (Brown‐Elliott et al. 2001).
The successful amplification in the first step of the IS6110 and gyrA PCR with the
correct amplicon length but in fact non-MTBC DNA sequence demonstrates the
difficulties in screening of M. tuberculosis DNA from archaeological remains. There are
so far 188 identified species with valid names within the Mycobacterium genus; the
majority of these are environment bacteria, typically found in soil and water and only a
small number being pathogenic species (Gupta et al. 2018). The species within this
genus are divided into two groups: slow growing mycobacterium, which take more than
7 days to form colonies, and rapid growing mycobacterium, taking less than 7 days for
colony formation. The environmental mycobacteria are also known as ‘mycobacteria
other than tuberculosis’ (MOTT). One of the contamination sources from M.
tuberculosis study is the MOTT bacteria that are present in the soil environment of the
burials and, for some, might have invaded the skeletal remains (Müller et al. 2016). The
DNA from these MOTT bacteria can be amplified as shown in this study. This is raising
a question on the specificity of the PCR assays used to screen MTBC DNA from
archaeological remains (Wilbur et al. 2009). The PCR assays used to detect the presence
MTBC DNA are considered specific for these species from clinical samples (Eisenach et

116
al. 1990). However, considering the complex bacterial content of archaeological
specimens, this might not hold true as demonstrated by a previous study (Müller et al.
2015) and now, in this study. The first round of the IS6110 PCR assay, as well as the
gyrA and Rv0083 PCRs have demonstrated the ability to produce amplicons of the
expected length, however with non-MTBC sequences. In the case of gyrA target, most
of the amplicons obtained in this study are from MOTT. This highlights the importance
of performing sequencing verification of the amplicon as it is clear that these targets are
not solely amplifying MTBC aDNA.
PCR assays were designed to incorporate AmpliTaq Gold and BSA in the reaction mix
as it was shown that AmpliTaq Gold has greater efficiency in PCR while the BSA may
reduce the effect of inhibitors on the amplification (Forbes & Hicks 1996; Abu Al-Soud
& Rådström 2000; Pandey et al. 2012). In addition to the original undiluted DNA, 10-1
diluted DNA also used as a template in the PCR screening. The purpose of testing 10-1
diluted DNA is to further minimize the effect of inhibitors in the amplification efficiency
(Wilson 1997). Two of the main challenges of ancient DNA studies are the low copy of
available endogenous DNA and the presence of PCR inhibitors (Dabney, Meyer &
Pääbo 2013). From the 16 samples with positive bands, only 2 of them show positive or
positive-like amplification from both undiluted and 10-1 diluted DNA templates. In the
undiluted DNA, the amount of endogenous DNA is maximized, but unfortunately so is
the amount of the PCR inhibitors. While in the 10-1 diluted DNA, the effect of the PCR
inhibitors will be reduced but the number of the endogenous DNA molecules will be
reduced as well. This could be the reason why some samples only produced positive or
positive-like amplification with one of the DNA templates.
3.3.3 Failure of MTBC aDNA detection
Based on the osteological studies performed previously, most of the archaeological
skeletons studied here are suggested to be positive for tuberculosis infection (Table 2.1).
However, based on the PCR screening results, using four targets supposedly specific for
MTBC, only one sample, St Andrew Fishergate 6, shows evidence for the presence for
MTBC aDNA. This raises a question on whether these individuals were not infected
with tuberculosis or whether the detection failures were caused by any other factors.
Even the samples which show Pott’s lesion, which is considered to be a pathognomonic

117
sign of tuberculosis infection, did not show any evidence for MTBC aDNA. There are
several factors that might contribute to the failure of MTBC aDNA amplification. The
most important determinant is the preservation of the archaeological bone remains. The
skeletons could be in poor preservation state – resulting in low endogenous DNA
amounts (Taylor et al. 1996; Donoghue 2011). The low endogenous DNA amount
combined with the presence of environmental contaminants will contribute further to the
complication of endogenous DNA amplification (Wilbur et al. 2009; Müller et al. 2016).
Environmental organisms may contaminate the skeletal remains throughout the
preservation years (Tsangaras & Greenwood 2012). The presence of abundant
environmental organisms including MOTT may result in positive amplification as
depicted in this study (Wilbur et al. 2009). In addition, the bacterial load of infectious
pathogen at the time of death could also have contributed to the failure of the MTBC
aDNA amplification (Barnes & Thomas 2006). It could be that a high bacterial load is
necessary in order for the pathogen DNA to be retained in the bones throughout the
preservation history. The inability to obtain positive amplification from the samples
cannot therefore dispute the presence of tuberculosis infection in the individuals studied.
Furthermore, the archaeological remains selected in this study comprised of skeletal
bone showing tuberculosis lesions and normal bones without any lesions. Majority of the
non-control specimen studied showing lesions on ribs; others are showing either
vertebrae, hips or endocranial lesions. The generally accepted pathognomonic bone
change of tuberculosis is the destructive lesions observed on the vertebrae, typically on
the lumbar spine or lower thoracic (Roberts 2015). There is an absence or very little
involvement of new bone formations. Other than the spine, the effect can also be seen in
the knee and hips, although other parts of the body can also be affected. The bone
change in ribs is classified as non-specific tuberculosis lesions. Although previous
studies have observed the potential of rib lesions as an indicative for tuberculosis in
archaeological remains, the bone change on this part of the body can also be the result of
any other types of diseases. Therefore, it is also possible that the non-specific lesions
observed on the majority of the bones studied here were caused by other diseases.
Caution must be taken to conclude a diagnosis based on non-specific bone changes
alone. The biomolecular analysis method is incorporated in ancient tuberculosis
diagnosis confirmation for this reason. However, one also should not rely completely on
an ancient DNA study result as the occurrence of false negatives are possible due to the
factors highlighted above. By performing both osteological and biomolecular analysis
together on archaeological remains it might be possible to increase the confidence of the

118
palaeodisease diagnosis. It is dangerous to draw a definite conclusion on whether
tuberculosis infection is absence or present based on either type of analysis alone
(Roberts 2015).
3.3.4 Samples for Next Generation Sequencing
Eight samples were selected for NGS. Ideally, the samples to be chosen should be those
with positive MTBC aDNA detection, confirmed by PCR assay and Sanger sequencing.
However, as previously shown, only St Andrew Fishergate 6 gave positive results for the
presence of MTBC aDNA. Therefore, another seven samples were selected from the
‘negative’ samples. The use of ‘negative’ samples is justified because PCR failure can
arise if a sample contains aDNA that is too short to amplify, or where contaminants are
present, and under these circumstances NGS following DNA capture can still be
successful (Templeton et al. 2013).
The main selection criterion for the eight samples used for NGS remained the
comparison between rural and urban sites. In choosing these eight samples, other
considerations were the presence or absence of lesions and, when lesions were present,
the part of the skeleton affected, the choice made to include a range of examples in case
aDNA preservation was affected by any of these factors. Practical considerations such as
the amount of sample material available were also taken into account. However, the
main interest is still to compare MTBC aDNA between samples retrieved from urban
and rural locations.
From the eight samples selected for NGS, four samples are from urban and rural sites
respectively. From the four urban samples, two are from St Andrew Fishergate and the
other two are from St Helen-on-the-Walls. From the first location, a sample with
tuberculosis lesion (St Andrew Fishergate 6, which gave positive PCR results) and a
control bone (St Andrew Fishergate 253) were chosen. Meanwhile, from the second
urban location, St Helen-on-the-Walls 6003 and St Helen-on-the-Walls 5494 were a
bone with tuberculosis lesion and a control bone respectively. St Helen-on-the-Walls
6003 was chosen as the skeletal remains show lesions on the lower thoracic and lumbar
vertebrae which is highly likely to be caused by tuberculosis, albeit these samples tested
negative for MTBC aDNA in the PCR screening.
From the rural samples, one is from Hickleton and the remaining three are from
Wetwang Slack, the latter being a site with relatively large amounts of available sample

119
material, so it would be possible to repeat NGS experiments if necessary. The three
samples from Wetwang Slack include rib bones with lesions (Wetwang Slack 8) and
without lesions (Wetwang Slack 7). The third sample that was included from this site is
Wetwang Slack 185, which is a sample from a vertebrae bone. Sample 46 from
Hickleton was chosen as this individual was showing spine lesions and the vertebra was
used for the study. Spinal lesions are generally accepted as the pathognomonic sign of
tuberculosis infection, therefore there is a probability that this individual had indeed
contracted tuberculosis, although the PCR tests showed otherwise. The selection for the
next samples for NGS (if any) would be based on the NGS results of these samples.
More samples would have been chosen if the NGS results (Chapter 4) had been more
promising.

120
Chapter 4: Study of M. tuberculosis aDNA in archaeological
remains from Yorkshire, England. Part 2: Next Generation
Sequencing.
4.1 Introduction
In the early ‘age’ of ancient DNA study, the focus was mainly directed towards the
detection of the presence of the organism of interest, which was routinely performed
through PCR (Spigelman & Lemma 1993; Salo et al. 1994; Haas et al. 2000). Before the
advent of the high throughput sequencing technologies, the acquisition of aDNA
sequence was highly challenging owing to its very low endogenous DNA concentration,
degraded and short fragment length (Rizzi et al. 2012). The largest amount of genomic
aDNA sequence generated by the Sanger method was the 26,861 bp sequence from an
extinct Pleistocene cave bear (Noonan et al. 2005). The pioneering of NGS has
accelerated the aDNA field with the first NGS performed on an ancient specimen
producing 13 million bp of DNA sequence from the extinct woolly mammoth;
representing a 480-fold of increase compared with the cave bear genome data reported
by Noonan and colleagues (2005) previously (Poinar et al. 2006). NGS has
revolutionized the study of MTBC ancient DNA as well. Hybridization capture directed
at polymorphic regions of the M. tuberculosis genome in a 19th century English skeleton
managed to identify the genotype as closely associated to MTBC strains that were
thought to be common in North America in the early 20th century, albeit very rare today
(Bouwman et al. 2012). Metagenomics NGS performed from an archaeological tissue
sample from a mummy recovered from Vac, Hungary produced 32x M. tuberculosis
genome coverage which enabled the identification of mixed infection originating from
two different M. tuberculosis genotypes (Chan et al. 2013). Metagenomics study of other
18th century Hungarian archaeological remains also showed infection from different M.
tuberculosis genotypes during this period of time (Kay et al. 2015). From these

121
examples, it is shown that metagenomics and hybridization capture can be applied as
NGS strategies in studying ancient MTBC DNA.
Both metagenomics and target enrichment through in-solution capture were used as high
throughput sequencing strategies in this study. Metagenomics or shotgun sequencing
was performed with eight samples as indicated in Chapter 3. The shotgun strategy is
used to allow the identification of all known organisms in the archaeological remains.
The sample St Andrew Fishergate 6 was also subjected to whole M. tuberculosis genome
target enrichment to further increase the reads from M. tuberculosis.
The eight samples subjected to NGS were St Andrew Fishergate 253, St Andrew
Fishergate 6, St Helen-on-the-Walls 5494, St-Helen-on-the-Walls 6003, Hickleton 46,
Wetwang Slack 185, Wetwang Slack 7 and Wetwang Slack 8. The first four samples
mentioned originated from urban locations while the remaining four were excavated
from rural parts of Yorkshire. These are all subjected to shotgun sequencing methods. In
addition to this, the sample St Andrew Fishergate 6, which tested positive for MTBC
aDNA in the PCR screening described in Chapter 3, was also subjected to target
enrichment through in-solution hybridization capture method before sequencing using
NGS. All of these samples were sequenced using the Illumina HiSeq platform.
4.2 Results
4.2.1 Shotgun sequencing
The computational tools and parameters used to analyse the sequence of all samples
were the same to ensure consistency. The sequences were pre-processed using the
programs explained in Section 2.7. In both the shotgun and hybridization studies, only
the collapsed reads were used in the subsequent analyses to ensure consistency across all
samples. The collapsed reads were aligned to the M. tuberculosis ancestor reference
genome (Comas et al. 2014).
The shotgun sequencing yielded 1.2 x 107 to 6.0 x 107 collapsed reads for all samples
except for St Andrew Fishergate 6 and Wetwang Slack 185 (Table 4.1). However, less

122
than 1% of the total reads mapped to M. tuberculosis for all samples. Even the sample St
Andrew Fishergate 6, which produced positive PCR screening of MTBC aDNA
(Chapter 3), has only 0.0476% of reads mapped to M. tuberculosis, this being 1,210
reads before duplicate removal and only 60 reads after duplicate removal. The highest
mapped read percentage is Wetwang Slack 185, which had the lowest number of total
collapsed reads. As well as investigating the number of reads mapped to M. tuberculosis,
the metagenome content of the samples was also explored. This was done using the
NCBI Blastn tool to assign each read to its respective taxonomy. In the BLAST search,
the minimum e-value was assigned at 1x10-7 to avoid a match occurring only by chance.
The word size, which is defined by the sensitivity of the search, was set to 20, which
mean that 20 “words” have to match between the query and the database to initiate the
extension to match. The results were then visualized and analysed in MEGAN 5. Due to
computational power limitations, running the 10s of millions of reads in MEGAN, a read
subset was extracted randomly from each file to be used in downstream analysis, except
for St Andrew Fishergate 6 and Wetwang Slack 185 where it was computationally
possible to run all reads. The exact bioinformatics steps to perform these analyses were
explained in detail in section 2.7.

123
Archaeological sample Number
of reads
Reads mapped to M. tuberculosis % of reads
mapped to M.
tuberculosis
Before duplicate reads
removals
After duplicate
reads removal
St Andrew Fishergate 253 22,143,918 5,747 2,815 0.0260
St Andrew Fishergate 6 2,541,970 1,210 60 0.0476
St Helen-on-the-Walls 5494 15,385,188 802 149 0.0052
St Helen-on-the-Walls 6003 57,912,949 7,344 4,493 0.0127
Hickleton 46 35,541,718 3,751 1,947 0.0106
Wetwang Slack 185 4,662 9 9 0.1931
Wetwang Slack 7 30,548,281 13,669 4,543 0.0447
Wetwang Slack 8 12,549,272 5,200 2,355 0.0414
Table 4.1: The result summary of the shotgun read mapping against the M. tuberculosis reference genome. The read numbers that passed the
pre-processing quality control, the number of reads mapped to the M. tuberculosis reference genome and their respective percentages are shown in the
table. The number of reads mapped to the M. tuberculosis reference genome before duplicate removal was used to calculate the percentage.

124
The taxonomic content of each sample was visualized in MEGAN 5. In MEGAN 5, firstly,
the number of reads corresponding to each superkingdom (i.e. Bacteria, Eukaryota,
Archaea, Viruses) was determined. This was calculated based on the total number of reads
with identifiable taxonomy. Next, the percentage of reads of the organism of interest in the
respective superkingdom was calculated. In Bacteria superkingdom, the percentage of
Mycobacterium genus, the intermediate level MTBC and M. tuberculosis species were
calculated in respect to the amount of the bacterial reads rather than the total read number.
The same analysis was performed for Homo sapiens: the percentage of H. sapiens reads
was calculated based on the overall Eukaryota reads – not the total read number.
St Andrew Fishergate 253
Super Kingdom
Number of
reads Percentage
Bacteria 39031 50.47%
Mycobacterium 2852 7.31%
MTBC 94 0.24%
M. tuberculosis 3 0.001%
Archaea 743 0.90%
Viruses 0 0.00%
Eukaryota 37559 48.57%
H. sapiens 2565 6.83%
Table 4.2: The number of reads assigned to each superkingdom, genus and species of interest
by MEGAN for sample St Andrew Fishergate 253. The superkingdom is highlighted in yellow,
genus in blue, species in green while the intermediate level MTBC is highlighted in red.

125
A subset of collapsed reads was investigated in BLAST for the identification of the
possible organisms that the reads belonged to. For the extract from skeleton 253 from St
Andrew Fishergate, 48.57% and 50.47% of the assigned reads belonged to the Eukaryota
and Bacteria superkingdoms, respectively (Table 4.2). Mycobacterium genus accounted for
7.31% of the total bacteria content of the extract, with only 0.24% (94 reads) assigned to
MTBC. There were no reads assigned to virus DNA. From the overall eukaryote reads,
6.83% belong to H. sapiens. In the identification of the 20 species dominating the reads
(Figure 4.1), H. sapiens shows the highest number of reads followed by Thermomonospora
curvata. Most of the organisms with the most abundant reads are soil-dwelling bacteria:
Sandaracinus amylolyticus, Sorangium cellulosum, Streptomyces bingchenggenesis,
Kitasatospora setae, Streptomyces cattleya, Streptomyces violaceusniger, Nocardiopsis
dassanovillei and Rhosopseudomonas palustris (Lampky 1971; Ichikawa et al. 2010; Sun
et al. 2010; Mohr et al. 2012; Wang et al. 2013; McMurry & Chang 2017). Interestingly,
the 20th most abundant species is Ramilibacter tataouinensis, a novel desert bacterium
which was isolated from fragments of meteorite submerged in sands of Tunisian desert
showing properties of persistence in desert environments (Strobel et al. 2012).

126
Figure 4.1: The 20 species with the highest read number assigned by MEGAN for sample St Andrew Fishergate 253. The number of reads
assigned to each species is shown on the top of the bars in the figure.

127
St Andrew Fishergate 6
Table 4.3: The number of reads assigned to each superkingdom, genus and species of interest
by MEGAN for sample St Andrew Fishergate 6. The superkingdom is highlighted in yellow,
genus in blue, species in green while the intermediate level MTBC is highlighted in red.
The majority of the reads from skeleton 6 from St Andrew Fishergate originated from
bacteria (91.06%); followed by 8.76% from eukaryotes, 0.17% archaea and no reads from
viruses (Table 4.3). From the overall reads of bacteria, only 0.30% were identified as
belonged to the Mycobacterium genus, and only one read was matched to MTBC. Homo
sapiens accounted for 32.3% of the Eukaryota reads. The most abundant reads in sample
St Andrew Fishergate 6 originated from Streptosporangium roseum , a soil-dwelling
non-pathogenic, non-motile spore-producing bacterium (Nolan et al. 2010). The second
most abundant reads originated from H. sapiens (Figure 4.2). In addition, other than H.
sapiens, sequences from mammals are also present in high proportions in this sample.
These include Macaca fascicularis and Ovis canadensis which are Southeast Asian
monkey and North American bighorn sheep, respectively (Higashino et al. 2012). Most of
the organism with abundant reads in this sample are naturally occurring soil bacteria:
Super
Kingdom
Number of
reads Percentage
Bacteria 390318 91.06%
Mycobacterium 1182 0.30%
MTBC 1 0.00%
M. tuberculosis 0 0.00%
M. leprae 229 0.06%
Archaea 743 0.17%
Viruses 0 0.00%
Eukaryota 37559 8.76%
H. sapiens 12106 32.23%

128
Streptomyces lydicus, Saccharopolyspora erythraea, Streptomyces iranensis, Lysobacter
enzymogen, Conexibacter woesei and Streptomyces vietnamensis (Zhu et al. 2007; Pukall
et al. 2010; de Lima Procópio et al. 2012; Qian et al. 2013). It is worth noting that most of
the species are from the Streptomyces genus, representing the largest genus within the
Actinobacteria class. Streptomyces are typically found in soil and decaying vegetation (de
Lima Procópio et al. 2012). Three of the most abundant species are also found in the
sample from the same site, St Andrew Fishergate 253: T. curvata, N. dassonvillei and
Kutzneria albida.

129
Figure 4.2: The 20 species with the highest read number assigned by MEGAN for sample St Andrew Fishergate 6. The number of reads
assigned to each species is shown on top of the bars in the figure.

130
St Helen-on-the-Walls 5494
Super
Kingdom
Number of
reads Percentage
Bacteria 42585 76.95%
Mycobacterium 131 0.31%
MTBC 2 0.00%
M. tuberculosis 0 0.00%
Archaea 397 0.72%
Viruses 27 0.05%
Eukaryota 12329 22.28%
H. sapiens 3834 31.10%
Table 4.4: The number of reads assigned to each superkingdom, genus and species of interest
by MEGAN for sample St Helen-on-the-Walls 5494. The superkingdom is highlighted in yellow,
genus in blue, species in green while the intermediate level MTBC is highlighted in red.
The reads from the sample St Helen-on-the-Walls 5494 are dominated by bacteria and
eukaryotes with 76.95% and 22.28%, respectively (Table 4.4). From the overall bacterial
population in the sample, only 0.31% of the reads were classified to Mycobacterium. Only
two of the overall reads were identified as belonging to MTBC. Archaea and viruses were
identified in the sample with abundance percentages of 0.72 (397 reads) and 0.05 (27
reads), respectively. No reads were specifically matched to M. tuberculosis. The
taxonomy composition of sample St Helen-on-the-Walls 5494 revealed H. sapiens as the
most abundant reads in the sample (Figure 4.3). A. sulfonivorans and K. flavida are both
bacteria from the Actinobacteria class that naturally occur in soil (Park et al. 1999;
Mongodin et al. 2006). Interestingly, in addition to H. sapiens, two other primates were
also identified to be among the most abundant reads in this sample: M. fascicularis and
Pan troglodytes. M. fascicularis is the species name for monkey, typically found in
Southeast Asia while P. troglodytes is referring to chimpanzee which is

131
Figure 4.3: The 20 species with the highest read number assigned by MEGAN for sample St Helen-on-the-Walls 5494. The number of reads
assigned to each species is shown on top of the bars in the figure.

132
also known as the African apes (Luke & Verma 1995). It is suggested as the closest
existing relatives to humans. Flavobacterium johnsoniae together with Pedobacter
heparinus and Niastella koreensis are common inhabitants of soil environments (Agarwal
et al. 1997; Weon et al. 2006; Han et al. 2009). A few of the species with the most reads
have been identified to be able to cause infection in humans: Pseudomonas stutzeri,
Arcobacter butzleri, Flavobacterium psychrophilum and Sphingobacterium sp. ML3W
(Lalucat et al. 2006; Arguello et al. 2015; Smith et al. 2015; Rochat et al. 2017). P. stutzeri
was shown to be associated to septicemia, bacteremia, bone infection, endocarditis, eye
infection, meningitis, skin infection, ventriculitis, urinary tract infection and pneumonia
(Lalucat et al. 2006). However, this bacterium was identified to be low in virulence and the
small number of deaths reported to be associated with this pathogen is still doubtful. The
20th most abundant read in sample St Helen-on-the-Walls 5494 came from the species
Sphingobacterium sp. ML3W which previously has been isolated from patients who
suffers from chronic respiratory infection (Smith et al. 2015).

133
St Helen-on-the-Walls 6003
Super
Kingdom
Number of
reads Percentage
Bacteria 481912 98.15%
Mycobacterium 1691 0.93%
MTBC 0 0.00%
M. tuberculosis 0 0.00%
Archaea 4584 0.93%
Viruses 64 0.01%
Eukaryota 4418 0.90%
H. sapiens 720 16.30%
Table 4.5: The number of reads assigned to each superkingdom, genus and species of interest
by MEGAN for sample Helen-on-the-Walls 6003. The superkingdom is highlighted in yellow,
genus in blue, species in green while the intermediate level MTBC is highlighted in red.
The BLAST search result visualized in MEGAN revealed that 98.15% of the successfully
assigned reads belong to the Bacteria superkingdom (Table 4.5). From the overall bacterial
content, 0.93% are assigned to the Mycobacterium genus but none of the reads is
specifically assigned to the MTBC. Archaea and viruses comprise 0.93% and 0.01% of the
reads, respectively. Only 0.90% from the reads are from Eukaryota: 16.3% of these are
from H. sapiens. The most abundant read in the sample belongs to F. johnsoniae, a
bacterium most commonly found in soil and freshwater which also gave abundant reads for
sample St Helen-on-the-Walls 5494 (Figure 4.4). Similar to the three samples described
previously, most of the species with the most abundant reads are naturally found in the
environment, especially soil: K. flavida, Nocardoides sp. JS614, A. sulfonivorans,
Sterptomyces pristinaespiralis, Nocardiodes dokdonensis, N. koreensis, S. amylolyticus, V.
paradoxus, Rhodoplanes sp. Z2-YC6860 and Arthrobacter sp. Strain FB24. Interestingly,
Ramilbacter tataouinensis, which has 1,759 reads assigned, has been previously shown to
have a strong adaptation to a desert lifestyle (de Luca et al. 2011).

134
Figure 4.4: The 20 species with the highest read number assigned by MEGAN for sample St Helen-on-the-Walls 6003. The number of reads
assigned to each species is shown on top of the bars in the figure.

135
Hickleton 46
Super
Kingdom
Number of
reads Percentage
Bacteria 564847 97.04%
Mycobacterium 1254 0.22%
MTBC 0 0.00%
M. tuberculosis 0 0.00%
Archaea 3621 0.62%
Viruses 2004 0.34%
Eukaryota 11619 2.00%
H. sapiens 3424 29.47%
Table 4.6: The number of reads assigned to each superkingdom, genus and species of interest
by MEGAN for sample Hickleton 46. The superkingdom is highlighted in yellow, genus in blue,
species in green while the intermediate level MTBC is highlighted in red.
The identifiable reads from the sample Hickleton 46 comprise of 97.04% bacteria, 2.0%
eukaryotes, 0.62% archaea and 0.34% virus sequences. From the bacteria, 1,254 reads
(0.22%) are assigned to Mycobacterium but none of them were identified as MTBC (Table
4.6). In the taxonomical classification performed by MEGAN, the most abundant reads in
the sample originated from the species Streptomyces albus (Figure 4.5). This organism has
been isolated from a wide range of environmental niches. In addition, this microorganism
was shown to be able to cause mycetoma infection in humans (Martín et al. 2004).
Mycetoma is a progressively destructive chronic inflammatory disease. The foot is the
most commonly affected part, but other regions of the body can be infected as well. More
than half of the 20 most abundant species in the sample belong to the Streptomyces genus.
As previously described, Streptomyces sp. are predominantly found in soil and decaying
vegetation (de Lima Procópio et al. 2012). Another soil-inhabiting bacterium, K. setae is
also abundant; this particular microorganism has similar morphology and lifestyle to the
species within the Streptomyces genus (Ichikawa et al. 2010).

136
Figure 4.5: The 20 species with the highest read number assigned by MEGAN for sample Hickleton 46. The number of reads assigned to each
species is shown on top of the bars in the figure.

137
Wetwang Slack 185
Table 4.7: The number of reads assigned to each superkingdom, genus and species of interest
by MEGAN for sample Wetwang Slack 185. The superkingdom is highlighted in yellow, genus
in blue, species in green while the intermediate level MTBC is highlighted in red.
There were very few reads produced from sequencing of the sample Wetwang Slack 185.
The majority (96.77%) of the identified reads were assigned to bacteria (Table 4.7). There
were 10 reads assigned to the Mycobacterium genus accounting for 1.34% of the overall
bacteria read number. However, none of these reads were identified as belonging to MTBC
aDNA. There were no virus sequences identified in the sample. Meanwhile, archaea and
eukaryote reads were identified with 0.52 and 2.52% abundance, respectively. From the
reads identified as belonging to eukaryota, only one read was assigned to H. sapiens.
Similar to sample St Andrew Fishergate 6, the most abundant reads in this sample were
assigned to S. roseum (Figure 4.6). T. bispora is also present abundantly as in sample St
Andrew Fishergate 6. Interestingly, M. leprae was identified as the third most abundant
species in the sample. A number of species are identified as soil-dwelling bacteria:
Kocuria rhizopila and Amycolatopsis orientalis. K. rhizopila was shown to be able to cause
human infections in a few instances, particularly of the bloodstream (Becker et al. 2008;
Moissenet et al. 2012). Another soil-inhabiting bacterium is the Amycolatopsis orientalis.
Six of the most abundant species in the sample belong to the Streptomyces genus – typical
soil-dwelling bacteria. Species belong to the genus Nocardiodes are widespread in aquatic
and terrestrial environments such as soil as wastewater.
Super
Kingdom
Number of
reads Percentage
Bacteria 748 96.77%
Mycobacterium 10 1.34%
MTBC 0 0.00%
M. tuberculosis 0 0.00%
Archaea 4 0.52%
Viruses 0 0.00%
Eukaryota 21 2.72%
H. sapiens 1 4.76%

138
Figure 4.6: The 20 species with the highest read number assigned by MEGAN for sample Wetwang Slack 185. The number of reads assigned to
each species is shown on top of the bars in the figure.

139
Wetwang Slack 7
Super
Kingdom
Number of
reads Percentage
Bacteria 763098 99.45%
Mycobacterium 1803 0.24%
MTBC 78 0.01%
M. tuberculosis 1 0.00%
Archaea 1970 0.26%
Viruses 12 0.00%
Eukaryota 2210 0.29%
H. sapiens 576 26.06%
Table 4.8: The number of reads assigned to each superkingdom, genus and species of interest
by MEGAN for sample Wetwang Slack 7. The superkingdom is highlighted in yellow, genus in
blue, species in green while the intermediate level MTBC is highlighted in red.
The majority of the reads obtained from the Wetwang Slack 7 sample originated from
bacteria with 0.29% from the overall population from Mycobacterium (Table 4.8). There
were 78 reads assigned to the MTBC group but only one of them was specifically assigned
to M. tuberculosis. The remaining reads were assigned to archaea (0.26%) and eukaryotes
(0.29%) of which 26.06% of the overall read population was assigned to H. sapiens.
There are also a small number of reads assigned to viruses (12 reads). In the determination
of the abundant species in the sample, S. roseum was shown to exhibit the most reads
(Figure 4.7). Two of the most abundant species are from the Lysobacter genus. Species in
this genus are ubiquitous in water and soil. The other two species, which have not been
described previously, Streptomyces sp. 769 and Catenulispora acidiphila, are soil-dwelling
bacteria.

140
Figure 4.7: The 20 species with the highest read number assigned by MEGAN for sample Wetwang Slack 7. The number of reads assigned to
each species is shown on top of the bars in the figure.

141
Wetwang Slack 8
Super Kingdom
Number of
reads Percentage
Bacteria 724453 99.36%
Mycobacterium 1664 0.23%
MTBC 30 0.00%
M. tuberculosis 1 0.00%
Archaea 2375 0.33%
Viruses 38 0.01%
Eukaryota 2239 0.31%
H. sapiens 274 12.24%
Table 4.9: The number of reads assigned to each superkingdom, genus and species of interest
by MEGAN for sample Wetwang Slack 8. The superkingdom is highlighted in yellow, genus in
blue, species in green while the intermediate level MTBC is highlighted in red.
The reads from the sample Wetwang Slack 8 mostly originated from bacteria (99.36%), of
which 0.23% was from the Mycobacterium genus, with 30 reads are assigned to MTBC
(Table 4.9). Only one read was assigned to M. tuberculosis. Some of the reads are assigned
to eukaryotes (0.31%), archaea (0.33%) and viruses (0.01%). From the eukaryote reads,
12.24% were assigned to H. sapiens. The most abundant read in sample Wetwang Slack 8
is from S. roseum (Figure 4.8); similar to Wetwang Slack 7, Wetwang Slack 185 and St
Andrew Fishergate 6. Nineteen of the 20 most abundant species in sample Wetwang Slack
8 are also found in the other previous samples. The only unique abundant species in this
sample is the bacterium Micromonospora narathiwatensis, which has been reported to be
found widely in the environment including water, sandstone, soils, mangrove sediments
and root nodules (Thawai et al. 2018).

142
Figure 4.8 The 20 species with the highest read number assigned by MEGAN for sample Wetwang Slack 8. The number of reads assigned to
each species is shown on top of the bars in the figure.

143
4.2.2 Target hybridization capture
Target enrichment was performed on sample St Andrew Fishergate 6 for both the M.
tuberculosis and M. leprae genomes separately. Target enrichment directed at M. leprae
was performed because 229 reads were assigned to M. leprae in the shotgun sequencing of
this sample. The results of shotgun sequencing and M. tuberculosis and M. leprae target
enrichment are summarized in Table 4.10.
Sequencing
strategy
Total read
number
Reads mapped to reference genome Percentage
Before duplicate
removal
After duplicate
removal
Shotgun sequencing 2,541,970 1,210 60 0.05%
M. tuberculosis
target capture
43,199,554 4,528 1,108 0.01%
M. leprae target
capture
23,600,572 898,728 4,104 3.81%
Table 4.10: Comparison of shotgun and target enrichment NGS results for St Andrew
Fishergate 6.
The percentage of reads mapped to the M. tuberculosis reference genome after shotgun
sequencing was higher than those from the library subjected to M. tuberculosis enrichment.
However, a higher percentage of reads mapped to the M. leprae reference genome than to
M. tuberculosis. None of the three NGS strategies gave sufficient reads to enable data to be
obtained on the genotypes of the M. tuberculosis and M. leprae bacteria in the sample.

144
4.3 Discussion
4.3.1 Efficiency of shotgun sequencing in isolating endogenous DNA
The maximum percentage of endogenous (M. tuberculosis) DNA recovered from any
sample was 0.19%, from Wetwang Slack 185. However, this figure could be affected by
bias as this sample produced very few reads, reflecting the low concentration of Wetwang
Slack 185 DNA that was added to the pool that acted as the template for library
preparation (Table 4.11). Both St Andrew Fishergate 6 and Wetwang 185 exhibit the
lowest initial DNA library concentration. During the pooling of the 8 samples into a single
sequencing library, the other 6 samples were diluted so that they can be pooled in an
equimolar ratio. However, based on the electropherogram result (Figure 4.9), a high
proportion of adapter dimers is present in Wetwang Slack 185 DNA library – as indicated
by the fluorescence peak at the 109 bp position. Although all samples were pooled in an
equimolar ratio, most of the reads produced from this sample could be from the adapter
dimers contamination, which explains the very small number of reads that were obtained.
Sample DNA library concentration (nM)
St Andrew Fishergate 6 25.29
St Andrew Fishergate 253 665.15
St Helen-on-the-Walls 5494 166.29
St Helen-on-the-Walls 6003 556.31
Hickleton 46 468.75
Wetwang Slack 185 89.36
Wetwang Slack 7 623.84
Wetwang Slack 8 920.32
Table 4.11: The DNA library concentrations for the 8 samples subjected to shotgun NGS.
These concentration values were used to pool the samples into a single NGS library in an
equimolar ratio. St Andrew Fishergate 6 and Wetwang Slack 185 have the lowest concentration
among all samples.

145
Figure 4.9: The electropherogram showing the DNA library size distribution of the sample
Wetwang Slack 185. The fluorescence signal (FU) shown on the y-axis, is plotted against the
DNA library size indicated on the x-axis. The detection of the fluorescence signal is dependent on
the number of sequences with the respective sizes present in the sample.
The second-highest percentage of endogenous content is from St Andrew Fishergate 6,
Wetwang Slack 7 and Wetwang Slack 8, each with approximately 0.04% of endogenous
DNA. The very low endogenous DNA recovery could be due to several factors. The first
problem could be the presence of environmental DNA contaminants, which may
outcompete the endogenous DNA (Noonan et al. 2005; Der Sarkissian et al. 2014; Llamas
et al. 2016). In shotgun sequencing, the entire DNA that is present in the sample will be
sequenced. Some of the microorganisms in the burial soil might not have been successfully
removed during the pre-extraction protocols and their DNA will be included in the
sequencing library. The soil microorganism DNA might be in a better condition compared
to the endogenous aDNA and will probably be more abundant in the sample (Green et al.
2008; Knapp & Hofreiter 2010). As shown by the taxonomy determination using MEGAN,
the bacteria DNA content is at least 91% in all six samples: St Andrew Fishergate 6, St
Helen-on-the-Walls 6003, Hickleton 46, Wetwang Slack 185, Wetwang Slack 7 and
Wetwang Slack 8. Sample St Andrew Fishergate 253 has an almost equal content of

146
bacteria and eukaryote reads and this is reflected in the species determinations with H.
sapiens being the most abundant species in the sample. Similarly, with St Helen-on-the-
Walls 5494 where again there is a relatively low proportion of bacterial reads in the
sample, H. sapiens is identified as the most abundant species. However, in each sample,
the majority of the reads are from environmental bacteria. All the samples studied here
were recovered from grave burials and therefore have had long years of contact with soil.
Therefore, shotgun sequencing might not be the best strategy to obtain endogenous aDNA
from archaeological specimens which have been buried in this way (Der Sarkissian et al.
2014). Shotgun sequencing is probably much more suited for much better preserved
archaeological remains (Gilbert et al. 2007).
4.3.2 Taxonomical content of the archaeological samples
The collapsed read datasets from shotgun sequencing were examined by BLAST and
MEGAN in order to assign reads to species. The resulting taxonomies for six of the
samples contained at least 1400 species, the exceptions being St Andrew Fishergate 6 (293
species) and Wetwang Slack 185 (77 species). The number of species identified in the
other samples was 2,365 for St Andrew Fishergate 253; 1,467 species for St Helen-on-the-
Walls 5494; 2,631 species for St Helen-on-the-Walls 6003; 2,181 species for Hickleton 46;
1,916 species for Wetwang Slack 7 and 1,820 species for Wetwang Slack 8.
The taxonomies contained several species that are not expected to have come into contact
with the skeletons, such as mammals including monkeys and bighorn sheep, as well as
some bacteria from specialised environments such as desert soil. Some of these anomalies
are due to the absence from the NCBI database, which is searched by BLAST, of many
species, especially bacteria species that have not been studied yet (Santamaria et al. 2012;
Breitwieser et al. 2019). In these cases, the read is assigned to its closest match in the
database, which will be a species related to the one from which the read was obtained,
though possibly a species that lives in a different environment (Porter & Beiko 2013). This
means that some of the species that are identified are incorrect, but these incorrect
identifications should be consistent, so identification of the same set of species in two

147
skeletons suggests that the taxonomic contents of those two skeletons are similar.
Comparisons between skeletons could therefore show differences in the preservation
conditions, for example, whether this is different in urban and rural areas.
Due to the high number of species in each taxonomy, only the 20 most abundant species in
each sample were studied and compared between each sample. All the species that are
shared by at least two samples are depicted in Table 4.12. There is no specific pattern
observed between urban and non-urban regions. However, there are 6 species that were
present abundantly in urban but not non-urban samples which are Sorangium cellulosum,
Sandaracinus amylolyticus, Rhodoplanes sp. Z2-YC6860, Ramlibacter tataouinensis,
Conexibacter woesei and Macaca fascisularis. S. cellulosum, S. amylolyticus,
Rhodoplanes sp. Z2-YC6860 and C. woesei are all found in soils (Lampky 1971; Pukall et
al. 2010; Mohr et al. 2012; Rosa et al. 2018). R. tataouinensis was isolated from meteorite
buried in Tunisia desert sand and adapted to this lifestyle, and so this identification is
presumably, in fact, an unidentified related bacterium present in soil. The reads identified
as Macaca are probably degraded human DNA reads, which contain miscoding lesions
resulting in misidentification. There were also five species that were found in both of the
St Helen-on-the-Walls samples, these are Flavobacteria johnsoniae, Kribela flavida,
Arthrobacter sulfonivorans, Variovorax paradoxus and Arthrobacter sp. strain FB24. All
of these are common soil bacteria.
Meanwhile for the samples from non-urban regions, Streptomyces sp. S10, Streptomyces
avermitilis, Streptomyces venezuelae, Streptomyces laurentii, and Streptomyces sp. Mg1
are the unique abundant organisms which are present only in Hickleton and Wetwang
Slack sample. The Streptomyces genus contains many soil-dwelling bacteria and these
identifications are not unusual. These taxonomies show that there are some similarities
between samples from the same site and some similarities among urban and rural samples
(Table 4.12). However, because the total number of reads is low for all samples, it is not
possible to compare the different taxonomies with endogenous DNA preservation and it is
unknown if the environmental bacteria affect the preservation of ancient DNA in a sample.

148
Sample/shared species St Andrew Fishergate253
St Andrew Fishergate 6
St Helen-on-the-
Walls 5494
St Helen-on-the-Walls
6003
Hickleton 46
Wetwang Slack 185
Wetwang Slack 7
Wetwang Slack 8
Thermonospora curvata yes yes yes yes yes
Kutzneria albida yes yes yes
Nocardiopsis dassonvillei yes yes yes yes
Flavobacteria Johnsoniae yes yes
Kribela flavida yes yes
Nitrospora moscoviensis yes yes yes yes yes
Arthrobacter sulfonivorans yes yes
Steroidobacter denitrificans yes yes yes yes yes yes
Variovorax paradoxus yes yes
Arthrobacter sp. Strain FB24 yes yes
Sorangium cellulosum yes yes
Streptomyces cattleya yes yes
Candidatus Nitrospira defluvii yes yes yes yes
Homo sapiens yes yes yes yes yes
Streptomyces sp. S10 yes yes
Kitasatospora setae yes yes yes yes
Streptomyces fulvissmus yes yes
Streptomyces avermitilis yes yes
Streptomyces venezuelae yes yes
Streptomyces laurentii yes yes
Streptomyces sp. Mg1 yes yes
Streptosporangium roseum yes yes yes yes
Thermobispora bispora yes yes yes yes
Kribella flavida yes yes

149
Nocardioides dokdonensis yes yes
Streptoalloteichus hindustanus yes yes yes yes
Streptomyces violaceusniger yes yes
Streptomyces bingchenggenesis
yes yes yes yes
Actinosynnema mirum yes yes yes
Kibdelosporangium sp MJ126-NF4
yes yes
Saccharothrix espanaensis yes yes yes
Niastella Koreensis yes yes yes
Frankia sp. Eul1c yes yes
Saccharopolyspora erythraea yes yes
Frankia sp. EAN1pec yes yes
Sandaracinus amylolyticus yes yes
Rhodoplanes sp. Z2-YC6860 yes yes
Nocardiopsis dassonvillei yes yes yes
Ramlibacter tataouinensis yes yes
Frankia sp. Eul1c yes yes
Conexibacter woesei yes yes
Macaca Fascicularis yes yes
Niastella koreensis yes yes
Number of species unique to sample
2 5 8 3 8 10 5 1
Table 4.12: Comparison of the 20 most abundant species in each sample as determined by BLAST and MEGAN analysis of the reads
obtained by shotgun sequencing. Yes: species is present abundantly in the sample, yellow: species is present abundantly in at least one skeleton in
all locations (urban and rural), blue: the abundant species is only present in the urban locations, pink: the abundant species is only present in the rural
locations, green: the presence of the abundant species is specific to a single location.

150
Pathogenic bacteria are present abundantly in two samples: St Helen-on-the-Walls 5494
and Wetwang Slack 185. The A. butzleri, a pathogenic bacterium species is abundantly
found in skeleton 5494 from St Helen-on-the-Walls. This microorganism has been
recognized as an emerging pathogen where it was associated with a persistent watery
diarrhoea case in a patient with chronic lymphocytic leukaemia (Arguello et al. 2015).
Two infectious bacteria species were identified in Wetwang Slack 185 sample. M. leprae
is the causative agent of leprosy, while K. rhizophila has been previously inflicted to a
persistent bloodstream infection and it was recognized as an emerging “micrococcus”
(Becker et al. 2008; Moissenet et al. 2012). The death of individuals St Helen on the
Walls 5494 and Wetwang Slack 185 could possibly be associated to these pathogens.
However, care should be taken in deriving such a conclusion. First, the manifestation
and nature of preservation of these pathogens in bone remains are not known. The
identities of these pathogens are not confirmed – the results may be a detection of
similar (but not yet identified) environmental bacteria. Secondly, the M. leprae and K.
rhizophila species account for only 6 and 4 reads in sample Wetwang Slack 185,
respectively. This is due to the low read number obtained from this sample as described
in section 4.4.1. This is not sufficient to draw a conclusion about the cause of death of
individual 185 from Wetwang Slack.
The most abundant read in sample Hickleton 46 is from the species S. album which was
shown to able to cause mycetoma infection in humans (Martin et al. 2004). Interestingly,
mycetoma or also previously known as Madura foot is an alternative diagnosis for
leprosy in skeletons (Hershkovitz et al. 1993). It is possible that the individuals St
Helen-on-the-Walls 5494, Wetwang Slack 185 and Hickleton 46 could have contracted
infections from the pathogens found in the samples but survived. However, the data
presented here are not sufficient to draw such a conclusion.
4.3.3 Target enrichment sequencing strategy
In sample St Andrew Fishergate House 6, the percentage of endogenous DNA recovered
after target enrichment did not differ that much in comparison to the shotgun sequencing
result. It was suggested that the success of target enrichment, especially in-solution
hybridization capture, is vastly dependent on the amount of endogenous DNA in the
original sample; an endogenous DNA amount of at least 1% in the overall DNA content
will be more likely to yield successful results after target enrichment (Cruz-Dávalos et

151
al. 2018). In addition, low clonality and low complexity of starting samples are
important factors in determining the success of target enrichment – especially for
archaeological specimens.
4.3.4 Possible mixed infection in sample St Andrew Fishergate House 6
The skeleton 6 from St Andrew Fishergate showed positive amplifications, verified by
Sanger sequencing as being M. tuberculosis, for at least two of the MTBC PCRs in each
of two DNA extraction batches. From the PCR result, it can be inferred that this sample
does contain MTBC aDNA. However, the result from the shotgun sequencing did not
show many reads mapped to the M. tuberculosis reference genome. The taxonomical
content determination of this sample showed that more reads were assigned to M. leprae
compared to M. tuberculosis. PCR was performed to screen for the presence on M.
leprae aDNA in the sample, but no amplification was shown. Separate target capture
was attempted for both M. tuberculosis and M. leprae, giving enriched libraries that
produced 1,108 and 4,104 reads that mapped to M. tuberculosis and M. leprae,
respectively, with a higher endogenous percentage for M. leprae. This suggests a
possible mixed infection in St Andrew Fishergate 6. Co-infection of M. tuberculosis and
M. leprosy has been demonstrated by Donoghue and colleagues (2005). The mixed-
infection was shown in bone remains from 1st century AD Israel, 4th century AD
(Roman) Egypt, 10th century AD Hungary and mediaeval (10-13th century AD) Sweden
(Donoghue et al. 2005). These cases are older than skeleton 6 from St Andrew
Fishergate (early 14th century AD). The latter is around the time where the prevalence of
leprosy peaked in Britain before it disappeared from this region. However, the number
of reads obtained was not enough to provide sufficient coverage to give any genotype
information for either Mycobacterium.
4.3.5 MTBC aDNA detection in bone remains from Yorkshire
The objective of the study performed in Chapter 3 and Chapter 4 was to determine if the
frequency of MTBC aDNA detection is high enough to plan for a much larger project to
test the hypothesis that there were strain differences between urban and rural areas.
Testing this hypothesis would require a much larger sample size and higher frequency of
positive results than was achieved in this thesis.

152
In the PCR screening, only one of the 60 samples produced a positive result of MTBC.
Other samples produced non-specific amplification, even though some of these skeletons
showed destructive lesions on the vertebrae – a specific bone change indicative of
tuberculosis. Eight samples were sequenced in order to compare the metagenomic
content and to see if there is any MTBC aDNA that can be detected by the NGS.
However, the NGS results showed an extremely low number of MTBC reads, even in
the PCR positive sample: St Andrew Fishergate 6. The frequency of the MTBC aDNA
detection is not high enough to enable endogenous sequence comparison between
different samples. The target capture is expected to increase the number of endogenous
read output from the samples. However, this is not the case in this study. Perhaps this is
due to extremely low copies of endogenous DNA in the sample to begin with. To study
broader hypotheses a much larger sample size would be required in order to obtain a
high enough aDNA detection frequency to allow different strains of MTBC to be
detected and compared. In addition, much better-preserved specimens might be
considered for such studies.
The study described in Chapters 3 and 4 highlight the difficulties in detecting and
isolating MTBC aDNA due to environmental contaminants. In addition, the study also
demonstrates the non-specific results of PCR assays that are supposedly specific to
MTBC alone. Caution should be taken in confirming the presence of tuberculosis
infection based on the skeletal lesions alone, especially the non-specific lesions.
Similarly, caution has to be taken when making disease identification based on positive
PCR results without sequencing authentication.

153
Chapter 5: Genotyping of Mycobacterium leprae ancient DNA
from mediaeval England
5.1 Background of study
This chapter focuses on a genotyping study of M. leprae isolates, which were
responsible for the leprosy endemic in mediaeval England. Leprosy was endemic in
Britain during the mediaeval period and the peak infection occurred in the 13th century
AD (Mendum et al. 2014). In the 15th century AD, human leprosy started to decline
before disappearing in the 16th century AD, although leprosy has recently been detected
in British red squirrels (Avanzi et al. 2016). The same timeline is reflected in Europe,
except for some regions where there is still some leprosy cases reported (Ramos et al.
2016). The absence of contemporary cases in Britain makes it difficult to know which
strains and genotypes were present in this region. Moreover, M. leprae is an extremely
host-dependent pathogen that cannot be cultured in normal culture medium, which
makes leprosy study even more difficult (Groathouse et al. 2006). Therefore, the ability
to study aDNA from archaeological remains provides an opportunity to access genomic
information of past M. leprae strains, especially where there is a zero modern human
leprosy cases as such in Britain.
Here, whole genome target enrichment was used as the NGS strategy, as it was shown to
substantially increase the proportion of M. leprae endogenous DNA in a sequencing
library in previous studies (Bos et al. 2014; Schuenemann et al. 2018). The three English
samples from Chichester and Raunds were tested positive for M. leprae aDNA in a
previous study, confirming the osteological observations (Müller 2008). This provided
the opportunity to retrieve genome sequences of the M. leprae strains present in this
region during that period of time. By comparing the Chichester and Raunds samples
with these other ancient samples, it will be possible to explore the diversity of M. leprae
infections in the past and the spread of different strains.

154
Here, this study is presented in form of a complete manuscript that has been submitted
and accepted, subject to revision for publication in Journal of Archaeological Science.
The roles of each author are outlined as follow.
Authors’ contributions: The osteological examination, DNA extraction and PCR
screening were performed by Dr Romy Muller as part of her Master’s project which was
supervised by Dr Christopher Knusel in 2008 at University of Bradford. She also
prepared the initial DNA library at Manchester Institute of Biotechnology during her
employment as Postdoctoral Research Associate in Brown’s lab. The following DNA
library target enrichment, final DNA library preparation and bioinformatics data analysis
and interpretation were performed by myself. The first draft of the article was written by
myself before being reviewed by my supervisor, Prof Terry Brown. The bone remains to
be studied were provided by Dr Jo Buckberry.

155
5.2 Publication draft
Ancient Mycobacterium leprae genomes from the mediaeval sites of Chichester and
Raunds in England
Ammielle Kerudina, Romy Müllera, Jo Buckberryb, Christopher J. Knüselc, Terence A.
Browna,*
a School of Earth and Environmental Sciences, Manchester Institute of Biotechnology,
University of Manchester, Manchester M1 7DN, UK
b Biological Anthropology Research Centre, School of Archaeological and Forensic
Sciences, University of Bradford, Bradford BD7 1DP, UK
c UMR5199 PACEA, Bâtiment B8, Allée Geoffroy Saint Hilaire, CS 50023, Pessac
Cedex, France 33615
Corresponding author
Email address: [email protected]

156
Abstract
We examined seven skeletons from mediaeval contexts from three sites in England for
the presence of Mycobacterium leprae DNA, each of the skeletons displaying
osteological indicators of leprosy. Polymerase chain reactions directed at the species-
specific RLEP multicopy sequence produced positive results with three skeletons, these
being among those with the clearest osteological signs of leprosy. Following in-solution
hybridization capture, sufficient sequence reads were obtained to cover >70% of the M.
leprae genomes from these three skeletons, with a mean read depth of 4–10. Two
skeletons from a mediaeval hospital in Chichester, UK, dating to the 14th–17th centuries
AD, contained M. leprae strains of subtype 3I, which has previously been reported in
mediaeval England. The third skeleton, from a churchyard cemetery at Raunds Furnells,
UK, dating to the 10th to mid-12th centuries AD, carried subtype 3K, which has been
recorded at 7th–13th century AD sites in Turkey, Hungary and Denmark, but not
previously in Britain. We suggest that crusaders or pilgrims to the Holy Land might have
been responsible for the transmission of subtype 3K from southeast Europe to Britain.
Keywords: Ancient DNA, Leprosy, Mediaeval England, Mycobacterium leprae,
Palaeopathology
5.2.1 Introduction
Leprosy is a slowly progressive, chronic granulomatous disease caused by
Mycobacterium leprae (Hansen, 1874) and potentially, in a minority of cases, by the
more recently characterised agent described as Mycobacterium lepromatosis (Han et al.,
2008). The primary symptoms are granulomas of the skin, peripheral nerves and
respiratory tract, but sometimes the eyes, bones and nasal cartilage are also affected
(Britton and Lockwood, 2004). The bacilli accumulate in the extremities of the body,
invading the Schwann cells causing nerve damage followed by a gradual sensory loss
and eventually leading to deformities and disabilities (Masaki et al., 2013). A multi-drug
regime comprising dapsone, rifampicin and clofazimine has been used successfully to
treat 16 million leprosy patients over the last twenty years, but new infections are
frequent with 210,671 leprosy cases reported in 2017 (World Health Organisation,

157
2018). With the highest incidence of new cases occurring in northeast South America,
central Africa and the Indian subcontinent, leprosy is classified as a ‘neglected tropical
disease’ (Lenk et al., 2018). Although the disease itself is curable, leprosy-related
deformities and disabilities are irreversible, especially when treatment has been delayed
(Britton and Lockwood, 2004). Some 2–3 million people worldwide display post-leprosy
disfigurements, and many are subject to the social discrimination referred to as leprosy
stigma, which in the past was driven by misunderstandings regarding transmission of the
disease, and which still persists today in some parts of the world (Grzybowski et al.,
2016).
Leprosy is one of the oldest diseases known to humankind. Although ambiguous,
textual references to skin diseases in the Indian Atharva Veda and Laws of Manu (2000–
1500 BC) (Bloomfield, 2004) and the Egyptian Ebers papyrus (1550 BC) (Hulse, 1972)
have been identified as leprosy, and there are more recent accounts of the disease dating
from the 6th century BC to 1st century AD from India (Bhishagratna, 1996), China
(McLeod and Yates, 1981; Leung, 2008), Greece (Pinhasi et al., 2005) and Rome
(Roberts and Manchester, 2010). Additional evidence is provided by palaeopathological
examination of archaeological skeletons for the osteological manifestations of the
disease that can be observed in the hands, feet, facial bones, tibiae and fibulae of
affected skeletons (Roberts and Manchester, 2010). The oldest skeleton displaying such
lesions dates to 2000 BC, from Rajasthan in northwest India (Robbins et al., 2009), in
accordance with the Indian textual references from the same period. It has been
suggested that the disease was brought to Europe and Northern Africa by the armies of
Alexander the Great, with their return from the Indian campaign in 327–326 C (Roberts
and Manchester, 2010). There is skeletal evidence of leprosy in Egypt at 200 BC
(Dzierzykray-Rogalski, 1980) and in western Europe from the 4th century AD (Reader,
1974). However, the disease appears to have been uncommon in Europe until the
mediaeval period, when skeletons displaying lesions become more abundant (Roberts
and Manchester, 2010). In Britain, the prevalence of leprosy peaks in the 13th century
AD and then declines during the 15th century AD before becoming uncommon again
from the 16th century AD onwards (for a review of the osteological evidence for Britain,
see Roberts, 2002), possibly because of improved social conditions combined with the
development of enhanced resistance to the disease among the human population
(Schuenemann et al., 2013). The decline is mirrored in continental Europe (Bennike,
2002), although the disease persisted in some parts of Norway and elsewhere until the
19th century AD (Boldsen, 2001).

158
About 5% of leprosy cases develop skeletal changes, and the lesions used in
osteological assessment of the disease can be ambiguous. An important adjunct to
palaeopathology has therefore been provided by the detection and sequencing of M.
leprae DNA, which is sometimes preserved in archaeological skeletons displaying
osteological lesions and has also occasionally been detected in skeletons free from such
lesions (Donoghue et al., 2017). Initially ancient DNA typing was used mainly to
support osteological identifications of leprosy (Rafi et al., 1994; Taylor et al., 2000,
2006; Donoghue et al., 2001, 2005, 2015; Inskip et al., 2015), but with increasing
knowledge of genomic diversity among extant M. leprae strains it has become possible
to contextualise ancient DNA data within an evolutionary scheme for the bacterium
(Schuenemann et al., 2018). The M. leprae genome is 3.27 Mb, substantially smaller
than the 4.42 Mb genome of Mycobacterium tuberculosis, and contains relatively high
number of pseudogenes, indicative of reductive evolution (Singh and Cole, 2011).
Different strains show high sequence similarity, with only a small number of variations
in the form of indels and single nucleotide polymorphisms (SNPs) (Monot et al., 2009).
The SNP variations were initially used to divide modern isolates into four main types
and 16 subtypes called 1A–1D, 2E–2H, 3I–3M and 4N–4P. With the addition of more
sequences, this classification has become elaborated into a phylogenetic scheme
comprising six main branches, with branches 1 and 2 corresponding to types 1 and 2,
respectively, branch 3 to subtype 3I, branch 4 to the type 4 strains and also subtypes 3L
and 3M, and branches 5 and 0 to different variants of subtype 3K (Schuenemann et al.,
2013, 2018). Among modern isolates, variants display geographical partitioning with
branch 1 associated with south and east Asia, branch 2 with south and southwest Asia,
branch 3 with Central and North America, branch 4 with west Africa and South
America, and branches 5 and 0 with east Asia (Monot et al., 2009; Schuenemann et al.,
2013). However, these present-day distributions do not reflect the full complexity of M.
leprae distribution in the past, especially in mediaeval Europe where subtypes within
branches 2, 3, 4 and 0 have been identified in skeletons dating from the 5th–14th centuries
AD (Singh and Cole, 2011).
Although M. leprae aDNA has been reported from a number of British sites
(reviewed by Donoghue et al., 2017), sufficient data for subtype identification has only
been obtained from six skeletons from the St Mary Magdalen leprosarium in Winchester
(Scheunemann et al., 2013; Taylor et al., 2013; Mendum et al., 2014; Roffey et al.,
2017) and one skeleton from a cemetery in Great Chesterford, Essex (Scheunemann et
al., 2018). Three of the Winchester skeletons yielded subtype 3I and the other three, as

159
well as the Great Chesterford sample, were subtype 2F. To extend the geographical
range of our knowledge of ancient M. leprae subtypes in Britain, we carried out a
biomolecular examination of seven skeletons from three sites from mediaeval England
(Fig. 5.1), each displaying pathological lesions indicative of leprosy though with varying
degrees of ambiguity. We report M. leprae genome sequences for three of these
skeletons. Two of the genomes correspond to subtype 3I, previously known in Britain,
but the third is novel to Britain and highlights the role that individual mobility might
have played in adding complexity to the phylogeography of M. leprae in mediaeval
Europe.
Figure 5.1: Locations of the sites from which skeletal samples were obtained.
5.2.2 Material and methods
5.2.2.1 Skeletons
Samples were selected, with permission, from the collection of the Biological
Anthropological Research Centre, University of Bradford, UK, based on various criteria.

160
First, skeletons that clearly show pathological rhinomaxillary changes indicative of
leprosy were identified. Given that these changes are pathogonomic for lepromatous
leprosy, bilateral and symmetrical non-specific changes in the lower limb and feet of
those skeletons are hypothesized to be associated with the disease as well. Additionally,
skeletons were sought that showed non-specific lesions in the lower limbs and feet as
commonly seen in leprosy but where no rhinomaxillary alterations could be recorded,
either because they were not present or they could not be observed due to the state of
preservation of the skeleton. The distribution of these lesions, as well as the fact that no
other alterations were found which would suggest a different aetiology, made the
differential diagnosis of leprosy for these skeletons likely. The decision about which
skeletal element and, in case of bilateral skeletal involvement, which side of the body
would be sampled, was based on whether or not destruction was justifiable given the
importance of the specimens for future studies.
Based on these criteria, samples were taken from seven skeletons from three sites
(Table S5.1, Supplementary Note). Skeletons C21, C35, C48 and C227 were excavated
in 1989 from a cemetery that had belonged to the Hospital of St James and St Mary
Magdalene, Chichester, UK. The original hospital was founded c.1118 AD and housed
leprosy sufferers until being dissolved in 1442 AD and converted to an almshouse when
the prevalence of leprosy declined in the UK (Magilton et al., 2008). Based on
osteological data, C21 was a young adult male, C35 an adult of indeterminate sex, C48 a
mature adult male, and C227 an adult, likely male. All four skeletons were dated to the
14th–17th century AD, based on historical documents and associated pottery (Magilton et
al., 2008). Skeletons R5046 and R5256 were males of 17–25 and 25–35 years,
respectively, from the late Anglo-Saxon churchyard cemetery at Raunds Furnells,
Northamptonshire, UK, excavated during 1977–1985. Stratigraphical analyses and
radiocarbon dating suggested that the churchyard cemetery was in use from the mid-10th
until mid-12th centuries (Boddington, 1996). Individual H3726 was a less well-preserved
and incomplete male skeleton of 25–35 years from a cemetery in the southwest area of
the cathedral at Hereford, UK, excavated in 1993 (Stone and Appleton-Fox, 1996). The
cemetery was in use from the 12th–16th centuries AD.

161
Table 5.1: Details of skeletons and samples that were taken.
Site Skeleton Sex, age at death Leprosy indicatorsa Elements sampleda
Rhinomaxillary
changes
Sub-periosteal new
bone formation
Other
changes
Chichester C21 Young adult male Yes Yes Yes Tibia, metatarsus
C35 Adult, indeterminate
sex
No Yes Yes Tibia
C48 Mature adult Yes Yes No Tibia, fibula
C227 Male(?) adult Yes Yes Yes Calcaneus, phalanx
Raunds R5046 Male, 17–25 years Yes Yes Yes Fibula
R5256 Male 25–35 years No Yes No Tibia, fibula, new bone
formation
Hereford H3726 Male 25–35 years Viscerocranium
absent
Yes Yes Tibia, fibula
a For details of pathology and elements sampled, see Supplementary Note (summary of archaeological sites, pathological lesions of skeletons, and elements that
were sampled) and Supplementary Table S5.1 (detailed osteological report).

162
Skeletons C21, C48 and C227 from Chichester and the Raunds skeleton R5046
displayed rhinomaxillary changes and other lesions indicative of leprosy (Table S5.1,
Supplementary Note, Supplementary Table S5.1). The fourth Chichester skeleton, C35,
and individual R5256 from Raunds did not show the typical rhinomaxillary changes but
both did have sub-periosteal new bone formation on other skeletal elements. Only parts
of the left lower limb were recovered from the Hereford skeleton H3726 and so
rhinomaxillary changes could not be assessed. However, the pattern of sub-periosteal
new bone formation on the lower limbs and feet suggested a possible leprous infection.
5.2.2.2 Ancient DNA regime
DNA extractions, PCRs and Illumina library preparations were performed in two
physically separated laboratories within the specialized ancient DNA research facility
at the University of Manchester. Each laboratory was supplied with ultra-filtered air
under positive displacement. After each use, benches and equipment were
decontaminated by UV irradiation and by cleaning with 5% hypochlorite acid, 70%
ethanol and DNA Away (Molecular Bioproducts). Small equipment, plasticware and
UV-stable reagents were decontaminated by UV irradiation (254 nm, 120,000 mJ cm–2
for 2 5 min, with 180° rotation between the two exposures) before use. Aqueous
solutions were similarly irradiated for 15 min. Personnel wore a disposable forensic
suit, face mask, hair net, goggles, two layers of gloves and disposable shoe covers at all
times. DNA extractions were accompanied by two blanks (normal extraction but
without skeletal material) per five samples and every set of 5–7 PCRs was
accompanied by at least two blanks (set up with water rather than DNA extract).
5.2.2.3 DNA extraction, PCR and sequencing
Bone samples were taken using a hacksaw or electronic drill by personnel wearing
protective clothing, including forensic suits, hair nets, face masks and two pairs of
sterile gloves. Samples were placed in sterile plastic bags and stored under dry and cool
conditions and transferred to the ancient DNA facility. The bone surfaces were

163
decontaminated by mechanical removal of the outer 1–2 mm of each sample, followed
by UV irradiation (254 nm, 120,000 mJ cm–2) for 2 5 min, with 180° rotation
between the two exposures (Bouwman et al., 2006). Bone samples were then placed in
a DNA-free plastic bag wrapped in a sterile piece of aluminium foil and crushed into
fine powder. DNA was extracted from 0.2 g of bone powder by standard methods
(method D of Bouwman and Brown, 2002; Dabney et al., 2013).
An initial screening for presence of M. leprae DNA was carried out by hemi-nested
PCR directed at the RLEP repetitive element, in 50 µl reactions comprising 2.5 µl of
DNA extract or 1.0 µl of first round PCR product, 1 AmpliTaq Gold PCR Master Mix
(ThermoFisher Scientific), 2 mM MgCl2, 200 µM dNTPs, 200 ng each primer, 1%
bovine serum albumin and 1.25 units AmpliTaq Gold DNA polymerase (ThermoFisher
Scientific). The primers for the first PCR (forward: 5´–
CACCTGATGTTATCCCTTGC–3´; reverse: 5´–ATCATCGATGCACTGTTCAC–3)
amplified a 133 bp fragment, and the second PCR (forward: 5´–
CATTTCTGCCGCTGGTATC –3´; reverse as for first PCR) amplified a 111 bp
fragment. Cycling conditions were 7 min at 95°C, followed by 35 cycles each
consisting of 1 min at 56°C, 1 min at 72°C, 1 min at 94°C, and a final cycle at 56°C for
1 min and 72°C for 10 min. PCR products were analysed by agarose gel electrophoresis
and directly purified using the QIAquick PCR product purification kit (Qiagen) prior to
Sanger sequencing (GATC Biotech, Cologne).
Dual-indexed libraries for Illumina sequencing were prepared from positive
samples. No DNA fragmentation step was performed as ancient DNA is already highly
degraded. Library preparation included a blunt-end repair step but no A-tailing,
followed by purification using the MinElute PCR purification kit (Qiagen), with elution
in 20 µl. Subsequent adapter ligation was performed using p5 and p7 adapters at a
concentration of 0.2 µM (Meyer and Kircher, 2010). Nicks from the previous step were
filled in with Bst polymerase before quantification by qPCR (Roche LightCycler 480)
and fluorimetry (Qubit 2.0) to determine the number of cycles required for the
subsequent indexing PCR. Sample-specific barcodes were added by double-indexing
(Kircher et al., 2012), using KAPA HiFi Uracil+ (Kapa Biosystems). Samples were
then pooled in equimolar ratios and sequenced from both ends in a single flow cell
(Illumina HiSeq 4000). As well as shotgun sequencing, samples were also sequenced
after enrichment by in-solution hybridization capture (MYcroarray) according to the

164
manufacturer's instructions for degraded samples. RNA baits were transcribed from 80-
mer oligonucleotides complementary to the M. leprae TN genome to give an array with
2 tiling density. Sequence data are curated at the European Nucleotide Archive under
study accession number PRJEB31393.
5.2.2.4 Data analysis
Raw sequencing data were pre-processed with AdapterRemoval 2.1 (Schubert et al.,
2016) to remove adapter sequence remnants, trim low quality bases and merge paired-
end reads. Reads of at least 25 bp which formed pairs with at least 11 bp overlap, and
non-overlapping pair mates of >25 bp, were retained in separate files. The paired-end
reads were then mapped to the M. leprae TN genome with BWA 0.7.12 (Li and Durbin,
2009). The alignments were cleaned by soft clipping, sorted based on coordinate with
Picard Tools (http://broadinstitute.github.io/picard), and mapped reads with a
quality score of at least 20 extracted using SAMtools 0.1.19 (Li et al., 2009). Read
duplicates were removed using the MarkDuplicates option in Picard Tools. The
mapped reads with duplicates removed were converted to Fasta files and tested by
BLAST (Altschul et al., 1990) with the outputs visualised using MEGAN 6 (Huson et
al., 2016). Base quality score recalibration was performed with GATK 3.6 (McKenna
et al., 2010) using the non-human genome method. The recalibrated alignments
containing the reads that mapped to M. leprae and were verified by BLAST were
visualized using Geneious 8.1.9 (Kearse et al., 2012). Polymorphisms were considered
genuine if supported by at least 5 coverage and a variant frequency of at least 80%.
5.2.3 Results
Samples (Table 5.1) were screened for the presence of M. leprae DNA by hemi-
nested polymerase chain reactions (PCRs) directed at the multicopy RLEP element,
which is believed to be specific to this species (Braet et al., 2018) and has previously
been used for detection of M. leprae ancient DNA (Donoghue et al., 2017). The first-
round PCRs provided products of the correct size, as judged by agarose gel
electrophoresis, for the two samples (tibia and metatarsus) taken from skeleton C21, the

165
two samples (tibia and fibula) from C48, and the single sample (fibula) from R5046
(Table 5.2). The second-round PCRs provided bands of the expected sizes from the
same samples, and no others. The results were replicated with a second set of PCRs on
the same extracts. None of negative controls (extraction blanks and PCR blanks)
revealed amplification products. Direct sequencing of the PCR products verified their
identity as genuine RLEP amplicons.
Table 5.2: Results of RLEP PCRs.
Skeleton Sampled element PCR resultsa
C 21 Tibia +,+
Metatarsus +,+
C 35 Tibia –,–
C 48 Tibia +,+
Fibula +,+
C227 Calcaneus –,–
Phalanx –,–
R5046 Fibula +,+
R5256 Tibia –,–
Fibula –,–
New bone formation –,–
H3726 Tibia –,–
Fibula –,–
a Result of first hemi-nested PCR, result of second hemi-nested PCR.

166
Samples from the three positive skeletons – C21, C48 and R5046 – were further
studied by Illumina sequencing. Shotgun sequencing was attempted for all three
samples but less than 0.001% of the reads mapped to the M. leprae TN genome, which
was insufficient for genome analysis. In-solution hybridization capture was therefore
used to enrich the samples for M. leprae sequences. Enrichment dramatically increased
the numbers of reads mapping to the reference genome (Supplementary Table S5.2),
with >70% of the genome covered for each of the samples and a mean read depth of 4–
10. The data enabled the ancient strains to be assigned to M. leprae genotypes (Table
5.3) (Monot et al., 2009), revealing that C21 and C48 belong to subtype 3I and R5046
to subtype 3K.
Table 5.3 Genotype assignments.
Skeleton SNP positiona Type SNP positiona Subtype
14,676 1,642,875 2,935,685 413,902 1,133,492 2,312,059 3,267,975
C21 C T C 3 G T C G I
C48 C T C 3 G T C G I
R5046 C T C 3 G G G G K
a SNP positions according to the M. leprae TN genome sequence.
Comparisons between different modern strains of M. leprae have revealed a total of
215 polymorphic sites (Monot et al., 2009). These sites were examined in the ancient
M. leprae genomes to determine whether the SNP version that was present was the
same as in the M. leprae TN reference sequence, or was the alternative SNP version
present in some other modern genomes (Supplementary Table S5.3). Of the three
ancient genomes, R5046 was the most greatly diverged from M. leprae TN, with 119 of
the 183 SNPs (65.0%) that were covered by the ancient sequence displaying the version
not present in the reference genome. In comparison, 53.1% and 56.1% of the SNPs

167
covered in the C21 and C48 genomes, respectively, had the non-reference version. The
greater dissimilarity between R5046 and M. leprae TN reflects the greater phylogenetic
distance between subtype 3K and subtype 1A, to which TN belongs (Schuenemann et
al., 2018). An additional 41 sites, comprising 30 SNPs and 11 indels, were specific to
the three ancient genomes reported here (Supplementary Table 5.4). Of the 30 SNPs,
18 were present only in the R5046 genome, and 7 and 5 were unique to C21 and C48,
respectively. None of the 30 SNPs were present in all three ancient genomes. Of the
eleven indels, five were specific to R5046 and six were present in all three samples,
four of the latter in pseudogenes.
5.2.4 Discussion
We examined seven skeletons from mediaeval contexts of three sites in England,
each of the skeletons displaying osteological indicators of leprosy, though with
different degrees of ambiguity. We identified M. leprae DNA in three skeletons and
following enrichment by in-solution hybridization obtained sufficient sequence data to
assign skeletons C21 and C48 to subtype 3I and skeleton R5046 to subtype 3K.
Although C21 and C48 came from the same cemetery, and were curated together for 25
years prior to DNA analysis, we can be confident that both contain endogenous M.
leprae DNA (as opposed to cross-contamination between the skeletons or
contamination from a single external source) as their M. leprae genome sequences are
non-identical.
Each of the three skeletons that produced positive results had extensive osteological
indications of leprosy, including rhinomaxillary changes, sub-periosteal new bone
formation, and other lesions on various skeletal elements. Of the samples that produced
negative results, C227 had a pathological condition most likely indicative of leprosy, in
particular pencilling of the fifth metatarsal with complete resorption of the head and
distal part of the diaphysis in the right foot as well as a slight pitting of the palate.
However, this skeleton displayed relatively poor physical preservation, indicating that
the failure to detect M. leprae DNA was possibly due to biomolecular degradation
before the skeleton was excavated. Skeletons C35 and R5256 did not display
rhinomaxillary changes, weakening the diagnosis of leprosy in both cases. The

168
pathology of the other parts of skeleton C35 could be ascribed to a treponemal disease
or other infection, and R5256 displayed new bone formation on the ossa coxae and left
scapula, skeletal elements that are not usually involved in leprosy infection, possibly
suggesting a systemic condition other than leprosy. Skeleton H3726 comprised only
fragments of the left lower limb, so the pathological evidence for leprosy was rather
weak, and unsupported by the DNA evidence. Overall, the results confirm those of
other groups (Rafi et al., 1994; Taylor et al, 2000, 2006, 2013; Donoghue et al., 2001,
2005, 2015; Inskip et al., 2015) by emphasising the value of ancient DNA analysis as a
means of providing independent support to palaeopathological identifications of
leprosy.
M. leprae strains previously reported from mediaeval Britain and Ireland have been
assigned to subtypes 2F and 3I (Taylor et al., 2013, 2018; Schuenemann et al., 2013,
2018; Mendum et al., 2014), the latter corresponding to branch 3 in the more recent
phylogenetic classification (Schuenemann et al., 2018). The discovery of subtype 3I in
two skeletons from Chichester, dating to the 14th–18th centuries AD, is therefore
consistent with these previous studies. Subtype 3K, however, has not previously been
reported in Britain. In modern M. leprae, this subtype is associated with east Asia, in
particular Japan, China, the Philippines and New Caledonia (Schuenemann et al.,
2018). Among ancient specimens it has been detected in a Turkish skeleton from the
8th–9th centuries AD (Erdal, 2004), three skeletons from Hungary, from the 7th–10th
centuries AD (Pálfi et al., 2002; Molnár et al., 2006; Schuenemann et al., 2018), and
another from 11th–13th century AD Denmark (Schuenemann et al., 2018). The R5046
skeleton is from a similar period (10th to mid-12th centuries AD) as these other
European detections, but is the most westerly in location, and hence the most distant
from the modern distribution of the subtype. The distribution pattern raises the
intriguing possibility that the individual represented by skeleton R5046 did not contract
leprosy in Britain but instead had travelled to continental Europe and/or Asia and
contracted the disease there. It has previously been suggested that human mobility
along the Silk Route was responsible for bringing subtype 3K to eastern Europe from
its supposed centre of origin in east Asia (Monot et al., 2009). During the Anglo-Saxon
period, up until the 10th century AD, there was also extensive mobility between Britain
and continental Europe, especially of educated clerics who taught and held religious
positions in various European countries (Palmer, 2009). Additionally, the 11th century
AD marks the beginning of the crusades by which European adventurers attempted to

169
gain control of the Christian Holy Land of Jerusalem. One of the routes taken by the
crusaders, and by pilgrims, to reach the Holy Land from western and central Europe
began in Vienna and passed along the Danube and the Via Diagonalis to
Constantinople, traversing Hungary, Serbia and Bulgaria (The Way to Jerusalem,
2018). It is therefore possible that leprosy of subtype 3K was transmitted to Britain and
other parts of western Europe by crusaders and pilgrims who had travelled to the Holy
Land and back via this route.
Acknowledgements
We thank Anthea Boylston (University of Bradford) for assistance with the
osteological examination, and Keri Brown (University of Manchester, UK) for
assistance with sampling. This work was funded by a studentship awarded by Majlis
Amanah Rakyat (MARA) to A.K. and by the University of Bradford and the University
of Manchester.
References
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basic local
alignment search tool. J. Mol. Biol. 215, 403–410. https://doi.org/10.1016/S0022-
2836(05)80360-2.
Bennike, P., 2002. Vilhelm Moller-Christensen: his work and legacy, in: Roberts, C.A.,
Lewis, M.E., Manchester, K. (Eds.), The Past and Present of Leprosy: Archaeological,
Historical, Palaeopathological and Clinical Approaches. BAR International Series
1054, Archaeopress, Oxford, pp. 135–144.
Bhishagratna, K.K, 1996. English Translation of Sushruta Samhita (ca 600 BC). Bose,
Calcutta.
Bloomfield, M., 2004. Hymns of the Atharva Veda. Kessinger Publishing, Whitefish.
Boddington, A., 1996. Raunds Furnells: the Anglo-Saxon Church and Churchyard.
Archaeological Report 7, English Heritage.
Boldsen, J.L., 2001. Epidemiological approach to the paleopathological diagnosis of
leprosy. Am. J. Phys. Anthropol. 115, 380–387. https://doi.org/10.1002/ajpa.1094.

170
Bouwman, A.S., Brown T.A., 2002. Comparison between silica-based methods for the
extraction of DNA from human bones from 18th–mid-19th century London. Ancient
Biomol. 4, 173–178. https://doi.org/10.1080/1358612021000028470.
Bouwman, A.S., Chilvers, E.R., Brown, K.A. & Brown T.A., 2006. Identification of
the authentic ancient DNA sequence in a human bone contaminated with modern DNA.
Am. J. Phys. Anthropol. 131, 428–431. https://doi.org/10.1002/ajpa.20411.
Braet, S., Vandelannoote, K., Meehan, C.J., Brum Fontes, A.N., Hasker, E., Rosa, P.S.,
Lucena-Silva, N., Rigouts, L., Suffys, P.N., De Jong, B.C., 2018. The repetitive
element RLEP is a highly specific target for detection of Mycobacterium leprae. J.
Clin. Microbiol. 56, e01924-17. https://doi.org/10.1128/JCM.01924-17.
Britton, W.J., Lockwood, D.N.J., 2004. Leprosy. Lancet 363, 1209–1219.
https://doi.org/10.1016/S0140-6736(04)15952-7 (2004).
Dabney, J., Knapp, M., Glocke, I., Gansauge, M.T., Weihmann, A., Nickel, B.,
Valdiosera, C., García, N., Pääbo, S., Arsuaga, J.L., Meyer, M., 2013. Complete
mitochondrial genome sequence of a Middle Pleistocene cave bear reconstructed from
ultrashort DNA fragments. Proc. Natl. Acad. Sci USA 110, 15758–15763.
https://doi.org/10.1073/pnas.1314445110.
Donoghue, H.D., Holton, J., Spigelman, M., 2001. PCR primers that can detect low
levels of Mycobacterium leprae DNA. J. Med. Microbiol. 50, 177–182.
https://doi.org/10.1099/0022-1317-50-2-177.
Donoghue, H.D., Marcsik, A., Matheson, C., Vernon, K., Nuorala, E., Molto, J.E.,
Greenblatt, C.L., Spigelman, M., 2005. Co-infection of Mycobacterium tuberculosis
and Mycobacterium leprae in human archaeological samples: A possible explanation
for the historical decline of leprosy. Proc. R. Soc. B 272, 389–394.
https://doi.org/10.1098/rspb.2004.2966.
Donoghue, H.D., Taylor, G.M., Marcsik, A., Molnar, A., Palfi, G., Pap, I., Teschler-
Nicola, M., Pinhasi, R., Erdal, Y.S., Velemínsky, P., Likovsky, J., Belcastro, M.G.,
Mariotti, V., Riga, A., Rubini, M., Zaio, P., Besra, G.S., Lee, O.Y., Wu. H.H.,
Minnikin, D.E., Bull, I.D., O'Grady, J., Spigelman, M., 2015. A migration-driven
model for the historical spread of leprosy in mediaeval Eastern and Central Europe.
Infect. Genet. Evol. 31, 250–256. https://doi.org/10.1016/j.meegid.2015.02.001.
Donoghue, H.D., Taylor, G.M., Stewart, G.R., Lee, O.Y.-C., Wu, H.H.T., Besra, G.S.,
Minnikin, D.E., 2017. Positive diagnosis of ancient leprosy and tuberculosis using
ancient DNA and lipid biomarkers. Diversity 9(4), 46.
https://doi.org/10.3390/d9040046.
Dzierzykray-Rogalski, T., 1980. Paleopathology of the Ptolemaic inhabitants of
Dakhleh Oasis (Egypt). J. Hum. Evol. 9, 71–72. https://doi.org/10.1016/0047-
2484(80)90044-5.

171
Erdal, Y.S., 2004. Health status of the Kovuklukaya (Boyabat/Sinop) people and its
relations with their lifestyle. Anadolu Araştirmalari 22, 169–196.
Grzybowski, A., Sak, J., Pawlikowski, J., Nita, M., 2016. Leprosy: social implications
from antiquity to the present. Clin. Dermatol. 34, 8–10,
https://doi.org/10.1016/j.clindermatol.2015.10.009.
Han, X.Y., Seo, Y.H., Sizer, K.C., Schoberle, T., May, G.S., Spencer, J.S., Li, W.,
Nair, R.G., 2008. A new mycobacterium species causing diffuse lepromatous leprosy.
Am. J. Clin. Pathol. 130, 856–864. https://doi.org/10.1309/AJCPP72FJZZRRVMM.
Hansen, G.H.A., 1874. Undersøgelser Angående Spedalskhedens Årsager. Norsk Mag.
Laegervidenskaben. 4, 1–88.
Hulse, E.V., 1972. Leprosy and Ancient Egypt. Lancet 2, 1024.
https://doi.org/10.1016/S0140-6736(72)92636-0.
Huson, D.H., Beier, S., Flade, I., Górska, A., El-Hadidi, M., Mitra, S., Ruscheweyhm
H.J., Tappu, R., 2016. MEGAN community edition – interactive exploration and
analysis of large-scale microbiome sequencing data. PLoS Comput. Biol. 12(6),
e1004957. https://doi.org/10.1371/journal.pcbi.1004957.
Inskip, S.A., Taylor, G.M., Zakrzewski, S.R., Mays, S.A., Pike, A.W.G., Llewellyn, G.,
Williams, C.M., Lee, O.Y., Wu, H.H., Minnikin, D.E., Besra, G.S., Stewart, G.R.,
2015. Osteological, biomolecular and geochemical examination of an early Anglo-
Saxon case of lepromatous leprosy. PLoS ONE 10, e0124282.
https://doi.org/10.1371/journal.pone.0124282.
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S., Buxton,
S., Cooper, A., Markowitz, S., Duran, C., Thierer, T., Ashton, B., Meintjes, P.,
Drummond, A., 2012. Geneious basic: an integrated and extendable desktop software
platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–
1649. https://doi.org/10.1093/bioinformatics/bts199.
Kircher, M., Sawyer, S., Meyer, M., 2012. Double indexing overcomes inaccuracies in
multiplex sequencing on the Illumina platform. Nucl. Acids Res. 40(1), e3.
https://doi.org/10.1093/nar/gkr771.
Lenk, E.J., Redekop, W.K., Luyendijk, M., Fitzpatrick, C., Niessen, L., Stolk, W.A.,
Tediosi, F., Rijnsburger, A.J., Bakker, R., Hontelez, J.A.C., Richardus, J.H., Jacobson,
J., Le Rutte, E.A., de Vlas, S.J., Severeens, J.L., 2018. Socioeconomic benefit to
individuals of achieving 2020 targets for four neglected tropical diseases
controlled/eliminated by innovative and intensified disease management: Human
African trypanosomiasis, leprosy, visceral leishmaniasis, Chagas disease. PLoS Negl.
Trop. Dis. 12(3), e0006250. https://doi.org/10.1371/journal.pntd.0006250.
Leung, A.K.C., 2008. Leprosy in China: a history. Columbia University Press, New
York.

172
Li, H., Durbin, R., 2009. Fast and accurate short read alignment with Burrows-Wheeler
transform. Bioinformatics 15, 1754–1760.
https://doi.org/10.1093/bioinformatics/btp698.
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., Marth, G.,
Abecasis, G., Durbin, R., 2009. The sequence alignment/map format and SAMtools.
Bioinformatics 25, 2078–2079. https://doi.org/10.1093/bioinformatics/btp352.
Magilton. J., Lee, F., Boylston, A., 2008. ‘Lepers Outside the Gate’: Excavation at the
Cemetery of the Hospital of St James and St Mary Magdelene, Chichester, 1986-87 and
1993. CBA Research report 158, Council for British Archaeology, York.
Masaki, T., Qu, J., Cholewa-Waclaw, J., Burr, K., Raaum, R., Rambukkana, A., 2013.
Reprogramming adult Schwann cells to stem cell-like cells by leprosy bacilli promotes
dissemination of infection. Cell 152, 51–67. https://doi.org/10.1016/j.cell.2012.12.014.
McKenna, A., Hanna, M., Banks, E., Sivachenko, A., Cibulskis, K., Kernytsky, A.,
Garimella, K., Altshuler, D., Gabriel, S., Daly, M., DePristo, M.A., 2010. The Genome
Analysis Toolkit: a MapReduce framework for analysing next-generation DNA
sequencing data. Genome Res. 20, 1297–1303. https:/.doi.org/10.1101/gr.107524.110.
McLeod, K., Yates, R., 1981. Forms of Ch'in law: an annotated translation of the Feng-
chen shih. Harv. J. Asiat. Stud. 41, 111–163. https://doi.org/10.2307/2719003.
Mendum, T.A., Schuenemann, V.J., Roffey, S., Taylor, G.M., Wu, H., Singh, P.,
Tucker, K., Hinds, J., Cole, S.T., Kierzek, A.M., Nieselt, K., Krause, J., Stewart, G.R.,
2014. Mycobacterium leprae genomes from a British mediaeval leprosy hospital:
towards understanding an ancient epidemic. BMC Genomics 15, 270.
https://doi.org/10.1186/1471-2164-15-270.
Meyer, M., Kircher, M., 2010. Illumina sequencing library preparation for highly
multiplexed target capture and sequencing. Cold Spring Harb. Protoc. 2010(6),
pdb.prot5448. https://doi.org/10.1101/pdb.prot5448.
Molnár, E., Marcsik, A., Bereczki, Z., Donoghue, H.D., 2006. Pathological cases from
the 7th century in Hungary, in Paleopathology Association, 16th European Meeting,
Scientific Program and Abstracts. Supplement to Paleopathology Newsletter, pp. 36–
37.
Monot, M., Honoré, N., Garnier, T., Zidane, N., Sherafi, D., Paniz-Mondolfi, A.,
Matsuoka, M., Taylor, G.M., Donoghue, H.D., Bouwman, A., Mays, S., Watson, C.,
Lockwood, D., Khamesipour, A., Dowlati, Y., Jianping, S., Rea, T.H., Vera-Cabrera,
L., Stefani, M.M., Banu, S., Macdonald, M., Sapkota, B.R., Spencer, J.S., Thomas, J.,
Harshman, K., Singh, P., Busso, P., Gattiker, A., Rougemont, J., Brennan, P.J., Cole,
S.T., 2009. Comparative genomic and phyleogeographic analysis of Mycobacterium
leprae. Nat. Gen. 42, 1282–1289. https://doi.org/0.1038/ng.477.

173
Pálfi, G., Zink, A., Haas, C.J., Marcsik, A., Dutour, O., Nerlich, A.G., 2002. Historical
and palaeopathological evidence of leprosy in Hungary, in: Roberts, C.A., Lewis, M.E.,
Manchester, K. (Eds.), The Past and Present of Leprosy: Archaeological, Historical,
Palaeopathological and Clinical Approaches. BAR International Series 1054,
Archaeopress, Oxford, pp. 205–209.
Palmer, J.T., 2009. Anglo-Saxons in a Frankish World, 690–900. Brepols, Turnhout.
Pinhasi, R., Foley, R., Donoghue, H.D., 2005. Reconsidering the antiquity of leprosy.
Science 312, 846. https://doi.org/10.1126/science.312.5775.846a.
Rafi, A., Spigelman, A., Stanford, J., Lemma, E., Donoghue, H., Zias, J., 1994. DNA of
Mycobacterium leprae detected by PCR in ancient bone. Int. J. Osteoarchaeol. 4, 287–
290. https://doi.org/10.1016/S0140-6736(94)92494-5.
Reader, R., 1974. New evidence for the antiquity of leprosy in early Britain. J.
Archaeol. Sci. 1, 205–207. https://doi.org/10.1016/0305-4403(74)90043-0.
Robbins, G., Tripathy, M., Misra, V.N., Mohanty, R.K., Shinde, V.S., Gray, K.M.,
Schug, M.D., 2009. Ancient skeletal evidence for leprosy in India (2000 B.C.). PLoS
ONE 4(5), 1-8. https://doi.org/10.1371/journal.pone.0005669.
Roberts, C., Manchester, K., 2010. The Archaeology of Disease, third ed. The History
Press, Stroud.
Roberts, C.A., 2002. The antiquity of leprosy in Britain: the skeletal evidence, in:
Roberts, C.A., Lewis, M.E., Manchester, K. (Eds.), The Past and Present of Leprosy:
Archaeological, Historical, Palaeopathological and Clinical Approaches. BAR
International Series 1054, Archaeopress, Oxford, pp. 213–222.
Roffey, S., Tucker, K., Filipek-Ogden, K., Montgomery, J., Cameron, J., O’Connell, T.,
Evans, J., Marter, P., Taylor, G.M., 2017. Investigation of a medieval pilgrim burial
excavated from the leprosarium of St Mary Magdalen Winchester, UK. PLoS Negl.
Trop. Dis. 11(1), e0005186. https://doi.org/10.1371/journal.pntd.0005186.
Schubert, M., Lindgreen, S., Orlando, L., 2016. AdapterRemoval v2: rapid adapter
trimming, identification, and read merging. BMC Res. Notes 9, 88.
https://doi.org/10.1186/s13104-016-1900-2.
Schuenemann, V.J., Singh, P., Mendum, T.A., Krause-Kyora, B., Jager, G., Bos, K.I.,
Herbig, A., Economou, C., Benjak, A., Busso, P., Nebel, A., Boldsen, J.L., Kjellström,
A., Wu, H., Stewart, G.R., Taylor, G.M., Bauer, P., Lee, O.Y.-C., Wu, H.H.T.,
Minnikin, D.E., Besra, G.S., Tucker, K., Roffey, S., Sow, S.O., Cole, S.T., Nieselt, K.,
Krause, J., 2013. Genome-wide comparison of mediaeval and modern Mycobacterium
leprae. Science 341, 179–183. https://doi.org/10.1126/science.1238286.
Schuenemann, V.J., Avanzi, C., Krause-Kyora, B., Seitz, A., Herbig, A., Inskip, S.,
Bonazzi, M., Reiter, E., Urban, C., Dangvard Pedersen, D., Taylor, G.M., Singh, P.4,

174
Stewart, G.R., Velemínský, P., Likovsky, J., Marcsik, A., Molnár, E., Pálfi, G.,
Mariotti, V., Riga, A., Belcastro, M.G., Boldsen, J.L., Nebel, A., Mays, S., Donoghue,
H.D., Zakrzewski, S., Benjak, A., Nieselt, K., Cole, S.T., Krause, J., 2018. Ancient
genomes reveal a high diversity of Mycobacterium leprae in mediaeval Europe. PLoS
Pathog. 14(5), e1006997. https://doi.org/10.1371/ journal.ppat.1006997.
Singh, P., Cole, S.T., 2011. Mycobacterium leprae: genes, pseudogenes and genetic
diversity. Future Microbiol. 6, 57–71, https://doi.org/10.2217/fmb.10.153.
Stone, R., Appleton-Fox, N., 1996. A View from Hereford's Past: a Report on the
Archaeological Excavation in Hereford Cathedral Close in 1993. Logaston Press,
Eardisley.
Taylor, G.M., Widdison, S., Brown, I.N., Young, D., 2000. A mediaeval case of
lepromatous leprosy from 13th–14th century Orkney, Scotland. J. Archaeol. Sci. 27,
1133–1138. https://doi.org/10.1006/jasc.1999.0532.
Taylor, G.M., Watson, C.L., Bouwman, A.S., Lockwood, D.N.J., Mays, S.A., 2006.
Variable number tandem repeat (VNTR) typing of two palaeopathological cases of
lepromatous leprosy from mediaeval England. J. Archaeol. Sci. 33, 1569–1579.
https://doi.org/10.1016/j.jas.2006.02.008.
Taylor, G.M., Tucker, K., Butler, R., Pike, A.W.G., Lewis, J., Roffey, S., Marter, P.,
Lee, O.Y., Wu, H.H., Minnikin, D.E., Besra, G.S., Singh, P., Cole, S.T., Stewart, G.R.,
2013. Detection and strain typing of ancient Mycobacterium leprae from a mediaeval
leprosy hospital. PLoS ONE 8, e62406. https://doi.org/10.1371/journal.pone.0062406.
Taylor, G.M., Murphy, E.M., Mendum, T.A., Pike, A.W.G., Linscott, B., Wu, H.,
O'Grady, J., Richardson, H., O'Donovan, E., Troy, C., Stewart, G.R., 2018. Leprosy at
the edge of Europe – biomolecular, isotopic and osteoarchaeological findings from
medieval Ireland. PLoS ONE 13, e0209495. https://doi.
org/10.1371/journal.pone.0209495.
The Way to Jerusalem, 2018. Historical routes: routes to the Holy Land.
http://www.waytojerusalem.com/en/historical-routes/routes-to-the-holy-land.
World Health Organization, 2018. Global leprosy update, 2017: reducing the disease
burden due to leprosy. Wkly Epidemiol. Rec. 93, 445–456.
https://apps.who.int/iris/handle/10665/27

175
5.3 Supplementary information
Ancient Mycobacterium leprae genomes from the mediaeval sites of Chichester and
Raunds in England
Ammielle Kerudina, Romy Müllera, Jo Buckberryb, Christopher J. Knüselc, Terence A.
Browna,*
a School of Earth and Environmental Sciences, Manchester Institute of Biotechnology,
University of Manchester, Manchester M1 7DN, UK
b Biological Anthropology Research Centre, School of Archaeological and Forensic
Sciences, University of Bradford, Bradford BD7 1DP, UK
c UMR5199 PACEA, Bâtiment B8, Allée Geoffroy Saint Hilaire, CS 50023, Pessac
Cedex, France 33615
Supplementary Information
Supplementary Note 1. Summary of archaeological sites, pathological lesions of
skeletons, and elements that were sampled.
Supplementary Table S5.1. Detailed osteological report. – see Kerudin et al Table
S1.xlsx.
Supplementary Table S5.2. Summary statistics for Illumina sequencing following
enrichment of samples by in-solution hybridization.

176
Supplementary Table S5.3. Identities in the C21, C48 and R5046 genomes of the 215
SNPs known in modern M. leprae strains.
Supplementary Table S5.4. Unique variations (highlighted in green) present in the
C21, C48 and/or R5046 genomes.

177
Supplementary Note 1
Details of archaeological sites, pathological lesions of skeletons, and elements that
were sampled
Chichester. The Chichester site is situated outside the old city walls in the city of
Chichester, West Sussex, UK. The cemetery from which the skeletons were recovered
once served the mediaeval hospital of St. James and St. Mary Magdalene which was
used from approximately the early 12th until the 17th century AD (Magilton et al.,
2008). As indicated from contemporary documents, since its foundation at
approximately 1118 AD until 1442 AD, the hospital was used as a leprosarium, and
later became an almshouse. The original excavation was conducted in 1986–87 and
recovered 330 individuals, while ongoing work in 1993 revealed a further 44 skeletons.
The skeletons recovered from the cemetery date from around 1300–1700 AD, based on
associated pottery as well as on historical documents. For analytical reasons, the
cemetery was divided into two main areas, A and B, further subdivided into A1, and
A2, and B1 and B2. Skeleton C227 was excavated from Area B1, and the other three
from Area A2. It is suspected that Area A predates Area B.
Skeleton C21. Palaeopathological examination of skeleton C21 revealed rhinomaxillary
changes as well as new bone formation in several parts of the skeleton, in particular in
the lower limbs. Additional changes in the hands and feet were also suggestive of a
leprous infection. For DNA analysis, the middle third of the diaphysis of the left tibia
and the first metatarsal of the left foot (apart from its proximal articular surface) were
selected. The tibia shows new sub-periosteal striated compact bone formation,
diffusely, along the entire diaphysis (Fig. S5.1A), and the metatarsal shows porous and
compact sub-periosteal new bone formation (Fig. S5.1B).

178
Skeleton C35. Apart from a slight alveolar recession, no signs of rhinomaxillary
syndrome were found in this individual. However, both tibiae and fibulae show sub-
periosteal new bone formation. Especially, the fibulae and the right tibia are osteitic in
appearance and the right tibia and fibula were ankylosed. The distribution of these
pathological alterations, together with signs of infection in the left foot (the right foot is
missing) and bony exostoses on the left talus, may be due to leprosy. Therefore, the
proximal part of the broken distal diaphysis of the right fibula was chosen for analysis
(Fig. S5.2). This element shows sub-periosteal new porous compact bone formation,
partially striated and partially with overlying woven bone formation and is heavily
osteitic. Even though broken post-mortem, it is obvious that the most distal part of the
distal right fibula was ankylosed with the tibia through the interosseus ligament.
Skeleton C48. Rhinomaxillary changes as seen in leprosy were present and further
pathological alterations throughout the infracranial skeleton are hypothesised to be
associated with the disease. Sub-periosteal new bone formation can be seen in both
tibiae and fibulae, in the calcanei and in other bones of the left foot. In particular, the
left metatarsals are heavily osteitic and show areas of lytic destruction. Furthermore,
the tarsals of the left foot show dorsal tarsal bars. For DNA analysis, a part of the distal
end of the diaphysis was selected from both left tibia and fibula. The tibia shows
subperiosteal new striated porous compact bone formation diffusely spread along the
entire diaphysis. The lateral and postero-lateral aspects of the middle to distal third of
Fig. S5.1. Skeleton C21. (A) Sub-periosteal new compact bone formation on the diaphysis of the left tibia
(medial view). (B) Sub-periosteal new compact bone formation on the left first metatarsal (plantar view).
Fig. S5.2. Skeleton C35. Sub-periosteal new compact
bone formation with overlying woven bone on the right
fibula (medial view). Note the osteitic appearance and
that the distal head of the fibula was ankylosed with the
tibia before post-mortem breakage.

179
Fig. S5.4. Skeleton C227. (A) Plantar
view of the proximal phalanx of the right
tarsal digit I showing slight new porous
compact bone formation. (B) Lateral
view of the right calcaneus showing sub-
periosteal new compact bone formation.
Fig. S5.3. Skeleton C48. (A) Lateral view of the left
tibia showing diffuse, compact and striated sub-periosteal
new bone formation. Note the slightly osteitic
appearance in the distal to middle third of the diaphysis.
(B) Anterior aspect of the left fibula showing diffuse sub-
periosteal new bone formation. The new bone formation
is porous and compact. Note the osteitic appearance.
the diaphysis are affected most severely, with a slightly osteitic appearance in the distal
part laterally (Fig. S5.3A). Also the fibula exhibits diffuse subperiosteal new bone
formation, porous, compact and striated in appearance. The new bone formation is most
severe in the distal up to the middle part of the diaphysis, demonstrating osteitic
swelling (Fig. S5.3B).
Skeleton C227. This individual is badly preserved with only parts of the maxillae, parts
of the right lower limbs and part of the distal condyle of the femur present. Except
pitting of the palate, no rhinomaxillary changes suggestive of leprosy were recorded.
However, infectious alterations of the right foot that resulted in ankylosis of several
bones could be observed. New porous compact bone formation was also noted on the
right first metatarsal as well as on the calcaneus. The proximal phalanx of the right first
metatarsal, showing slight new porous compact bone formation (Fig. S5.4A), and the
distal part of the right calcaneus which, overall, showed new compact bone formation
(Fig. S5.4B), were chosen for DNA analysis.
Raunds. The late Anglo-Saxon church and churchyard of Raunds Furnells is situated in
East Northamptonshire, south of the River Nene. Excavation of the church and the
associated cemetery was conducted between 1977 and 1985 (Boddington, 1996). Based
on stratigraphic analyses and radiocarbon dating, the cemetery was in use from the
mid-10th until the late 11th or mid-12th centuries AD. Excavation of the graveyard

180
Fig. S5.5. Skeleton R5046.
Left fibula showing new
striated compact bone
formation.
recovered the remains of 363 individuals, most of these in a fairly good state of
preservation. Skeleton R5046 was buried on the southeastern edge of the graveyard
(zone 5), and R5256 in central zone 1. Both burials were orientated east-west with the
head directed westwards.
Skeleton R5046. This individual shows slight rhinomaxillary changes suggestive of
lepromatous leprosy. Bilateral pathological alterations in the lower limbs could further
be attributed to the disease. In particular, the left tibia shows sub-periosteal new
compact bone formation, partially striated and partially with overlying woven bone.
This sub-periosteal reaction is most severe on the lateral but also medial aspect of the
shaft. For the right tibia only a small plaque of new bone formation at the mid-shaft
medially could be recorded. New striated compact bone was also found on the left
fibula, spreading diffusely along the entire shaft. The distal shaft is affected most
severely and possesses a swollen osteitic appearance. For DNA analysis, the distal part
of the diaphysis of the fibula was selected (Fig. S5.5). As the proximal edge of this part
was treated with glue this area was removed before crushing of the bone.
Skeleton R5256. This skeleton presents a questionable case of leprosy. No pathological
alterations suggestive of rhinomaxillary syndrome could be observed. However, sub-
periosteal new bone formation as often seen in leprosy was found throughout the lower
extremities, including tibia and fibulae as well as the femora and bones of the right
foot. Sub-periosteal new bone formation was also found on the ossa coxae and on the
left scapula, skeletal elements that are usually not involved in leprous infection. For
DNA analysis, the distal third of the diaphysis of the left fibula showing sub-periosteal
new bone formation was selected (Fig. S5.6A). In addition, a window was excised and
collected from the antero-medial part of the distal diaphysis of the left tibia as this also
possessed sub-periosteal new bone formation (Fig. S5.6B). During the excision, a small
piece of new woven bone flaked off and was also tested for DNA.

181
Hereford. Hereford is located in the west of England, north of the River Wye. An
archaeological excavation was conducted on the southwest corner of the cathedral in
the summer of 1993 in preparation for construction of a new library to re-house the
historical treasures of Hereford cathedral (Stone and Appleton-Fox, 1996). The
excavation revealed parts of a cemetery associated with the cathedral that was in use
for much of the mediaeval period. As well a grave pit filled with the bones of an
estimated 5000 individuals, dated to the beginning 12th century, the excavation revealed
approximately 1100 additional individuals. These individuals were most likely buried
between the end of the 12th century and the closing down of the cemetery in the 16th
century when, according to historical documents, it became a garden. Many of the
skeletons are incomplete due to disturbances by later burials, including around 200
individuals excavated from three mass graves dating to the late 14th or early 15th
centuries who may have been victims of the plague. Individual H3726 was recovered
from the southeastern edge of cemetery, orientated in the east-west direction with the
head at the western end.
Skeleton H3726. This skeleton is badly preserved. Only fragments of the left lower
limb are present, comprising the femur, tibia and fibula. Both tibia and fibula show sub-
periosteal new bone formation diffusely spread along the entire diaphysis. For DNA
analysis, a piece of bone was cut from the distal part of the distal fragment of the
diaphysis of the tibia, which exhibits a patch of new bone formation on its lateral aspect
(Fig. S5.7A). A second sample was taken from the distal fragment of the diaphysis of
the fibula (Fig. S5.7B).
Fig. S5.6. Skeleton R5256. (A) Lateral view of the
diaphysis of the left fibula showing sub-periosteal new
bone formation. (B) Anterior view of the left tibia
exhibiting sub-periosteal new bone formation.

182
Fig. S5.7. Skeleton H3726. (A) Fragmented left tibia showing sub-periosteal new bone formation diffusely
spread along the entire diaphyses. Note the patch of new bone formation on the lateral aspect of the distal
diaphysis. (B) Fragmented left fibula exhibiting sub-periosteal new bone formation diffusely spread along the
diaphysis.

183
References
Boddington, A., 1996. Raunds Furnells: the Anglo-Saxon Church and Churchyard.
Archaeological Report 7, English Heritage.
Magilton. J., Lee, F., Boylston, A., 2008. ‘Lepers Outside the Gate’: Excavation at the
Cemetery of the Hospital of St James and St Mary Magdelene, Chichester, 1986-87
and 1993. CBA Research report 158, Council for British Archaeology, York.
Stone, R., Appleton-Fox, N., 1996. A View from Hereford's Past: a Report on the
Archaeological Excavation in Hereford Cathedral Close in 1993. Logaston Press,
Eardisley.

184
Supplementary Table S5.1
Detailed osteological report.
see Kerudin et al Table S1.xlsx.

185
Supplementary Table S5.2
Summary statistics for Illumina sequencing following enrichment of samples by in-solution
hybridization.
Statistic C21 C48 R5046
Non-duplicate reads mapped to M. leprae 174,013 150,929 437,081
Pairwise identity (%) 99.3 99.0 98.4
Mean depth of coverage 4.4 3.6 10
% of reference genome covered 71.8 73.4 86.6

186
Supplementary Table S5.3
Identities in the C21, C48 and R5046 genomes of the 215 SNPs known in modern M. leprae
strains.
SNP version C21 C48 R5046
Version in the M. leprae TN genome 76 72 64
Version not in the M. leprae TN genome 86 92 119
SNPs not covered in the ancient sequence 53 51 32

187
Supplementary Table S5.4
Unique variations (highlighted in green) present in the C21, C48 and/or R5046 genomes.
Type Position in M. leprae
TN sequence
Variation
Locus
M. leprae TN C21 C48 R5046
Indel 202,441 (G)3 (G)3 (G)3 (G)2 ML0141
SNP 285,608 C C C A ML0214
SNP 389,111 C not covered T C Intergenic
SNP 523,366 G G not covered A RLEP
Indel 639,281 T(6) T(6) T(6) (T)7 Intergenic
SNP 698,809 G G G A ML0575
SNP 727,160 G T G G Intergenic
SNP 792,321 A A A T moxR3 (pseudogene)
SNP 865,934 C C C T ML0723 (pseudogene)
SNP 924,308 C C A C ML0779
SNP 938,796 G G A G ML0794 (pseudogene)
Indel 944,192 – A A A PPE (pseudogene)
SNP 954,575 G A G G ML0805 (pseudogene)

188
Indel 958,229 (C)2 (C)3 (C)3 (C)3 ML0809 (pseudogene)
SNP 1,288,067 G G G T ML1113
SNP 1,353,076 G G G A fadA4
SNP 1,445,832 C T C C Intergenic
SNP 1,451,503 T T not covered C RLEP
Indel 1,477,964 T T T – Intergenic
SNP 1,561,467 C C C G PPE (pseudogene)
SNP 1,605,242 A C A A ML1345 (pseudogene)
SNP 1,828,126 A A A G ML1514 (pseudogene)
Indel 1,849,027 (C)2 (C)3 (C)3 (C)3 PPE (pseudogene)
Indel 1,912,457 – C C C mpt53 (pseudogene)
Indel 2,127,771 – not covered not covered C Intergenic
SNP 2,330,059 T T T C Repeat region
SNP 2,470,760 C T C C ML2079 (pseudogene)
Indel 2,486,597 (T)8 (T)8 (T)8 (T)7 ML2090 (pseudogene)
SNP 2,568,277 G G G A Intergenic
SNP 2,615,157 A A A C Intergenic
SNP 2,710,194 A A A G ML2286 (pseudogene)
SNP 2,719,838 C C C T acS (pseudogene)

189
SNP 2,778,223 G C G G ML2340 (pseudogene)
SNP 2,796,573 C not covered C T ML2353
Indel 2,893,092 – T T T Intergenic
SNP 2,953,004 C C T C ML2478
Indel 3,100,774 (C)3 (C)4 (C)4 (C)4 Intergenic
SNP 3,164,216 C C C A Intergenic
SNP 3,221,222 T T T C ML2676 (pseudogene)
SNP 3,253,040 G A G G ML2699
SNP 3,263,734 C C T C ML2707

190
Chapter 6: Conclusion
Ancient DNA is recognised to be a very valuable tool in palaeopathology as it can not
only confirm the presence of diseases in the past but also shed light on the origin, spread
and evolution of palaeodiseases. Ancient DNA, albeit a fairly new field of research,
holds tremendous potential in understanding diseases in the past. The completed work
presented here contributes important insights in the study of tuberculosis and leprosy. It
highlights the danger of assuming the authenticity of a MTBC PCR fragment without
characterization by sequencing. Consequently, this raises questions about the specificity
of the PCR markers routinely used to confirm the presence of MTBC ancient DNA,
confirming the suspicion raised by a past study (Müller et al. 2016). This thesis serves as
a good reminder of the need to for stringent methods when using aDNA to perform
disease identification in palaeopathology. It also demonstrates the extent of exogenous
DNA presence in an archaeological sample acquired by microbial contamination
throughout the preservation years or even during sample handling. Furthermore, this
thesis presents an important addition to our knowledge of leprosy through the discovery
of a new M. leprae subtype that has never been reported in mediaeval England before.
This discovery sheds new light on the spread of leprosy to England in the past.
6.1 The extent to which the objectives have been addressed:
objective one (tuberculosis)
The first objective of this thesis was to determine whether the MTBC aDNA detection
frequency is high enough to plan a larger study to test hypotheses such as possible strain
differences in urban and rural areas. This objective was addressed by screening 60
archaeological bone samples collected from rural and urban locations in Yorkshire,
England, for the presence of MTBC aDNA. This work was done in two stages, the first
stage being the screening of MTBC aDNA presence using PCR assays and further
verification with Sanger sequencing. One sample from the 14th century AD, St Andrew
Fishergate 6, showed evidence of MTBC aDNA presence, supporting the osteological
observation of tuberculosis indicative lesions on the vertebrae and the ribs of this
skeleton. Fourteen other samples gave exact or near-length PCR fragments but none of
these matched the MTBC sequence, demonstrating the importance of sequence
authentication to verify MTBC aDNA presence in archaeological bones, and questioning

191
the specificity of the markers routinely used in MTBC aDNA detection, especially
IS6110 (Muller et al. 2016). However, inclusion of the nested IS6110 PCR increased the
specificity of detection for this marker in this study, as St Andrew Fishergate 6, was the
only sample that showed positive amplification in both the first step and nested PCR,
and was also the only sample whose amplified fragment matched the MTBC reference
sequence.
The second stage in the work addressing the first objective involved NGS of eight
selected samples from stage one, all of which were subjected to shotgun sequencing and,
in the case of St Andrew Fishergate 6 to additional enrichment using the hybridization
target method. The number of reads mapped to the M. tuberculosis reference sequence
after shotgun sequencing was between 0.0052-0.1931% of the total. These low
percentages indicate that a large portion of the sequencing capacity is dominated by the
environmental DNA, as revealed by taxonomical classification of the reads. This study
suggests that the shotgun approach might be better suited for much better-preserved
archaeological specimens, for example, naturally mummified remains (Sabin et al.
2020). However, the hybridization capture target enrichment procedure did not
significantly increase the percentage MTBC reads for St Andrew Fishergate 6, although
the parallel M. leprae capture carried out with this sample provided M. leprae reads,
suggesting a possible mixed infection between the two pathogens. Mixed infection has
been reported in much earlier cases (Donoghue et al. 2005) than St Andrew Fishergate 6
(early 14th century AD).
In summary, the MTBC aDNA detection frequency obtained in this study was not high
enough to enable a larger scale study to be planned to test hypotheses such as possible
strain differences in urban and rural areas in historic Yorkshire. The negative results
could be due the preservation conditions resulting in a high amount of aDNA
breakdown, or possibly the bacterial load (the number of MTBC bacteria present in the
bone) at time of death was low, in which case even under good preservation conditions
there might not be enough aDNA to detect. Another possibility is that M. tuberculosis
was not be present in some of the samples. Thirty eight samples with lesions associated
with tuberculosis gave no evidence of MTBC aDNA. Although these results cannot be
used to completely refute the probability of these individuals ever being infected with
tuberculosis, they suggest that a differential diagnosis should also be considered for
these skeletal remains.

192
6.2 The extent to which the objectives have been addressed:
objective two (leprosy)
The second objective of the thesis was to use NGS to determine the genotype of the M.
leprae strains present in skeletons from two mediaeval sites, at Chichester and Raund
Furnells, both in England. This objective was addressed by typing the polymorphic sites
within the M. leprae genome to determine the strain and diversity of the ancient samples
previously suggested to contain M. leprae aDNA through PCR screening. Near complete
whole genome sequences were generated by target enrichment NGS allowing the
genotypes of three historical M. leprae isolates to be determined. The two isolates from
Chichester belonged to the typical European subtype – 3I. Meanwhile, the first 3K
subtype, dating to the 10th to mid-12th century AD, from historical M. leprae samples in
Britain is reported here. Based on the phylogeographic scheme of M. leprae isolates, the
transmission of the 3K subtype was suggested to be associated with the travels of
crusaders and pilgrims to the Holy Land during the mediaeval period.
6.3 Limitations of the thesis and future work
This study was not without its challenges and limitations. The main limitation arose
from the very nature of ancient DNA itself. As previously explained, physical and
chemical damages exerted on the bone remains during their preservation history will
cause DNA fragmentation, resulting in short fragments which could be missed during
the DNA extraction. This will subsequently lead to insufficient amounts of template for
PCR amplification and NGS. Skeleton remains recovered from grave burials are also at
high risk of environmental contamination that may interfere with PCR and NGS
analysis. This burial method allows close contact of skeletons to soil, and hence the
accumulation of soil microorganisms within the bones over time during the preservation
period. As demonstrated from the tuberculosis NGS results, this type of environmental
contamination leads to the sequences being dominated by exogenous bacteria, so there
are very few authentic aDNA reads.
DNA fragmentation and contamination of samples with environmental bacteria were not
limitations in addressing the second objective of the thesis. Each of the three samples
that had previously tested positive with M. leprae PCRs gave enough NGS sequences to
allow polymorphic sites in the M. leprae genome to be typed and the strains identified.

193
These results suggest that it is worth while carrying out future work using NGS to type
M. leprae aDNA with samples showing indications of leprosy, and the results of this
work could add more information on the strains of leprosy present in mediaeval Britain,
and tell more about how leprosy spread to Britain. The success in obtaining M. leprae
genome sequences reported in this thesis appears to agree with previous work (e.g.
Schuenemann et al. 2013, 2018) where a high proportion of bones showing indications
of leprosy could be sequenced, but it is difficult to make a comparison because negative
results are not often published.
The complications caused by DNA fragmentation and contamination did limit the
conclusions that could be drawn with regard to the first objective. Only one of 60
samples gave authentic MTBC sequences after PCR, and none of the eight samples that
were tested by NGS gave enough sequences for genome comparisons. Because negative
results are not often published, it is difficult to compare the high degree of negative
results reported in this thesis with other work, although Müller et al. (2014a) had
negative results with 43 of 77 British and European samples (1st-19th centuries AD)
tested by PCR. The limitation is that it was not possible to determine if the high
proportion of negative results is due to problems with the preservation and
contamination of the samples, or the absence of MTBC aDNA due to low bacterial load
at time of death. It would be interesting in future projects to compare hybridization
capture of both MTBC and human aDNA with the same samples, to see if the degree of
preservation of the two types of aDNA are comparable. If samples with good human
aDNA preservation still give negative results when tested for MTBC aDNA, then it
would suggest that the MTBC bacterial load in the samples was too low for aDNA
detection.
6.4 Ethical issues raised by this work
Naturally, in order to achieve the higher number of M. tuberculosis aDNA detections
needed to test hypotheses such as differences between strains in urban and rural
communities, more archaeological specimens could be used. However, the main ethical
concern in ancient DNA study, which is destructive sampling, should not be ignored
(Kaestle & Horsburgh 2002; Wilbur et al. 2009). Archaeological bone materials are seen
as historically precious due to their irreplaceable nature, and therefore should be

194
protected properly. Inevitably, the study of aDNA is destructive, because it requires part
of the skeleton to be taken and crushed. One of the recommendations to address this
issue is to only study the samples with the highest likeliness to yield positive results
(Kaestle & Horsburgh 2002). However, in an MTBC study, satisfying this requirement
is complicated by the fact that only 3-5% of active tuberculosis infections result in
skeletal manifestation (Roberts & Buikstra 2003).
Ancient DNA is still a fairly new interdisciplinary field, with the first ancient DNA
extraction performed only 36 years ago (Higuchi et al. 1984). Understandably, more
work is needed in order to “perfect” the way of studying ancient DNA without
compromising the ethical issues. Destructive analysis is obviously a valid concern, but
the tremendous potential of ancient DNA should not be ignored. However, the results
presented in this thesis suggest that in future studies, sample selection should be made
more stringent, so that valuable material is not needlessly destroyed. In previous studies,
M. tuberculosis aDNA has been successfully isolated from bones without lesions (Baron
et al. 1996; Müller et al. 2014a). One of the ways to address the concerns about
destruction would therefore be to sample bones that are available in a large number in
the skeleton, such as ribs, even if these do not themselves show lesions. If this approach
is combined with use only of skeletons with tuberculosis pathognomonic lesions, or
from sites with clear historical reports or evidences of tuberculosis, then it might be
possible to obtain sufficient MTBC NGS results for large scale studies, without
destroying large amounts of important material. To do this, it is important for historians,
osteologists, and molecular biologists to communicate with one another and to work
together to fill in the gaps in this field of research (Brown and Brown 1992; Squires et
al. 2019).

195
References
Abu Al-Soud, W. & Rådström, P., 2000, ‘Effects of amplification facilitators on
diagnostic PCR in the presence of blood, feces, and meat.’, Journal of Clinical
Microbiology, 38(12), 4463–70.
Adachi, N., Suzuki, T., Sakaue, K., Takigawa, W., Ohshima, N. & Dodo, Y., 2006,
‘Kinship analysis of the Jomon skeletons unearthed from a double burial at the
Usu-Moshiri site, Hokkaido, Japan’, Anthropological Science, 114(1), 29–34.
Adams, M., 1996, 'Excavation of pre-conquest cemetery at Addingham, West
Yorkshire', Medieval Archaeology, 40(1), 151-191.
Addyman, P.V., 1980, ‘Eburacum, Jorvik, York’, Scientific American, 242(3), 76-87.
Agarwal, S., Hunnicutt, D.W. & McBride, M.J., 1997, ‘Cloning and characterization of
the Flavobacterium johnsoniae (Cytophaga johnsonae) gliding motility gene,
gldA.’, Proceedings of the National Academy of Sciences of the United States of
America, 94(22), 12139–44.
Allison, M.J., Gerszten, E., Munizaga, J., Santoro, C. & Mendoza, D., 1981,
Tuberculosis in Pre-Columbian Andean populations, in Buikstra J.E. (ed.),
Prehistoric tuberculosis in the Americas, 49–61, Northwestern University
Archaeological Program, Evanston Illinois.
Alonso, H., Samper, S., Martín, C. & Otal, I., 2013, ‘Mapping IS6110 in high-copy
number Mycobacterium tuberculosis strains shows specific insertion points in the
Beijing genotype’, BMC Genomics, 14(1), 422.
Andersen, J.G. & Manchester, K., 1992, ‘The rhinomaxillary syndrome in leprosy: A
clinical, radiological and palaeopathological study’, International Journal of
Osteoarchaeology, 2(2), 121–129.
Ankad, B.S., 2018, ‘Dermoscopy of borderline tuberculoid leprosy’, International
Journal of Dermatology, 57, 74–76.
Arguello, E., Otto, C.C., Mead, P. & Babady, N.E., 2015, ‘Bacteremia caused by
Arcobacter butzleri in an immunocompromised host’, Journal of Clinical
microbiology, 53(4), 1448–51.
Armstrong, A., 1974, Stability and change in an English county town: a social study of
York 1801-51, Cambridge University Press, New York USA.
Atkins, P., 2008, ‘Fear of animal foods: a century of zoonotics.’, Appetite, 51(1), 18–21.
Atkins, P.J., 2000, Milk consumptions and tuberculosis in Britain, 1850-1950, in Fenton
A. (ed.), Order and disorder: the health implications of eating and drinking in the
nineteenth and tweentieth centuries, 83–95, Tuckwell Press, East Linton.
Avanzi, C., Del-Pozo, J., Benjak, A., Stevenson, K., Simpson, V.R., Busso, P.,
McLuckie, J., Loiseau, C., Lawton, C., Schoening, J., Shaw, D.J., Piton, J., Vera-
Cabrera, L., Velarde-Felix, J.S., McDermott, F., Gordon, S. V., Cole, S.T. &
Meredith, A.L., 2016, ‘Red squirrels in the British Isles are infected with leprosy
bacilli’, Science, 354(6313), 744–747.
Ankad, B.S., Hombal, A., Rao, S. & Naidu, V.M., 2011, 'Radiological changes in the
hands and feet of leprosty patients with deformities', Journal of Clinical and
Diagnostic Research, 5(4), 703-707.
Barberis, I., Bragazzi, N.L., Galluzzo, L. & Martini, M., 2017, ‘The history of
tuberculosis: from the first historical records to the isolation of Koch’s bacillus’,
Journal of Preventive Medicine and Hygiene, 58, E9–E12.

196
Barka, E.A., Vatsa, P., Sanchez, L., Gaveau-Vaillant, N., Jacquard, C., Meier-Kolthoff,
J.P., Klenk, H.-P., Clément, C., Ouhdouch, Y. & van Wezel, G.P., 2016,
‘Taxonomy, physiology, and natural products of Actinobacteria’, Microbiology and
Molecular Biology Reviews, 80(1), 1–43.
Barnes, I. & Thomas, M.G., 2006, ‘Evaluating bacterial pathogen DNA preservation in
museum osteological collections’, Proceedings of the Royal Society of London B:
Biological Sciences, 273(1587), 645–53.
Barnetson, J., 1951, 'Pathogenesis of bone changes in neural leprosy', International
Journal of Leprosy, 19(3), 297-307.
Baron, H., Hummel, S. & Herrmann, B., 1996, ‘Mycobacterium tuberculosis complex
DNA in ancient human bones’, Journal of Archaeological Science, 23, 667-671.
Bartlett, R., 2000, England under the Norman and Angevin Kings 1075 – 1225, Oxford
University Press, Oxford United Kingdom.
Becker, K., Rutsch, F., Uekotter, A., Kipp, F., Konig, J., Marquardt, T., Peters, G. & von
Eiff, C., 2008, ‘Kocuria rhizophila adds to the emerging spectrum of Micrococcal
species involved in human infections’, Journal of Clinical Microbiology, 46(10),
3537–3539.
Beckman Coulter, 2012, SPRIselect User Guide.
Bennett, B.H., Parker, D.L. & Robson, M., 2008, ‘Leprosy: steps along the journey of
eradication’, Public Health Reports , 123(2), 198–205.
Bobosha, K., Wilson, L., van Meijgaarden, K.E., Bekele, Y., Zewdie, M., van der Ploeg-
van Schip, J.J., Abebe, M., Hussein, J., Khadge, S., Neupane, K.D., Hagge, D.A.,
Jordanova, E.S., Aseffa, A., Ottenhoff, T.H.M. & Geluk, A., 2014, ‘T-Cell
regulation in lepromatous leprosy’, PLoS Neglected Tropical Diseases, 8(4), e2773.
Bos, K.I., Harkins, K.M., Herbig, A., Coscolla, M., Weber, N., Comas, I., Forrest, S.A.,
Bryant, J.M., Harris, S.R., Schuenemann, V.J., Campbell, T.J., Majander, K.K.,
Wilbur, A.K., Guichon, R.A., Wolfe Steadman, D.L., Cook, D.C., Niemann, S.,
Behr, M.A., Zumarraga, M., Bastida, R., Huson, D., Nieselt, K., Young, D.,
Parkhill, J., Buikstra, J.E., Gagneux, S., Stone, A.C. & Krause, J., 2014, ‘Pre-
Columbian Mycobacterial genomes reveal seals as a source of New World human
tuberculosis’, Nature, 514(7523), 494–497.
Bouwman, A.S., Kennedy, S.L., Müller, R., Stephens, R.H., Holst, M., Caffell, A.C.,
Roberts, C.A. & Brown, T.A., 2012, ‘Genotype of a historic strain of
Mycobacterium tuberculosis’, Proceedings of the National Academy of Sciences of
the United States of America, 109(45), 18511–18516.
Breitwieser, F.P., Lu, J. & Salzberg, S.L., 2019, ‘A review of methods and databases for
metagenomic classification and assembly’, Briefings in Bioinformatics, 20(4),
1125-1136.
Briggs, A.W., Stenzel, U., Johnson, P.L.F., Green, R.E., Kelso, J., Prüfer, K., Meyer, M.,
Krause, J., Ronan, M.T., Lachmann, M. & Pääbo, S., 2007, ‘Patterns of damage in
genomic DNA sequences from a Neandertal’, Proceedings of the National
Academy of Sciences, 104(37), 14616–14621.
Brites, D. & Gagneux, S., 2015, ‘Co-evolution of Mycobacterium tuberculosis and
Homo sapiens’, Immunological Reviews, 264(1), 6–24.

197
Brosch, R., Gordon, S. V., Marmiesse, M., Brodin, P., Buchrieser, C., Eiglmeier, K.,
Garnier, T., Gutierrez, C., Hewinson, G., Kremer, K., Parsons, L.M., Pym, A.S.,
Samper, S., van Soolingen, D. & Cole, S.T., 2002, ‘A new evolutionary scenario
for the Mycobacterium tuberculosis complex’, Proceedings of the National
Academy of Sciences of the United States of America, 99(6), 3684–3689.
Brown‐Elliott, B.A., Wallace Jr., R.J., Blinkhorn, R., Crist, C.J. & Mann, L.B., 2001,
‘Successful treatment of disseminated Mycobacterium chelonae infection with
Linezolid’, Clinical Infectious Diseases, 33(8), 1433–1434.
Brown, T.A. & Barnes, I.M., 2015, ‘The current and future applications of ancient DNA
in quaternary science’, Journal of Quaternary Science, 30(2), 144–153.
Brown, T.A. & Brown, K.A., 1992, ‘Ancient DNA and the archaeologist’, Antiquity,
66(250), 10–23.
Brown, T.A. & Brown, K.A, 2011, Biomolecular acrhaeology: an introduction, John
Wiley & Sons Ltd, West Sussex United Kingdom.
Burger, J., Hummel, S., Hermann, B. & Henke, W., 1999, ‘DNA preservation: a
microsatellite-DNA study on ancient skeletal remains’, Electrophoresis, 20(8),
1722–1728.
Cadet, J. & Wagner, J.R., 2013, ‘DNA base damage by reactive oxygen species,
oxidizing agents, and UV radiation’, Cold Spring Harbor Perspectives in Biology,
5(2), a012559–a012559.
Caffell, A. & Holst, M., 2008a, ‘Osteological analysis: Melton A63 East Yorkshire’,
York Osteoarchaeology No. 0507, Unpublished.
Caffell, A. & Holst, M., 2008b, ‘Osteological analysis: St Peter's Church, Huddersfield,
West Yorkshire’, York Osteoarchaeology No. 0508, Unpublished.
Caffell, A. & Holst, M., 2012, ‘Osteological analysis: 3 and 6 Driffield Terrace, York,
North Yorkshire', York Osteoarchaeology No. 0212. Unpublished.
Campbell, I.A. & Bah-Sow, O., 2006, ‘Pulmonary tuberculosis: diagnosis and
treatment’, British Medical Journal, 332, 1194–1197.
Canci, A., Minozzi, S. & Borgognini Tarli, S.M., 1996, ‘New evidence of tuberculous
spondylitis from Neolithic Liguria (Italy)’, International Journal of
Osteoarchaeology 6, 497–501.
Cardona, P.-J. & Ruiz-Manzano, J., 2004, ‘On the nature of Mycobacterium
tuberculosis-latent bacilli’, The European Respiratory Journal, 24(6), 1044–51.
Cardwell, P., Chundun, Z., Costley, S., Hall, J., Hanson, J., Huntley, J., Jamfrey, C.,
Maxwell, R., Speed, G., Simpson, R. & Stallibrass, S., 1995, 'Excavation of the
hospital of St Giles by Brompton Bridge, North Yorkshire', Archaeological Journal
152, 109-245.
CDC, 2013, Chapter 2: Transmission and pathogensis of tuberculosis, in Core
curriculum on tuberculosis: what the clinician should know (6th edn.), 19–44,
Centers for Disease Control and Prevention, Atlanta Georgia, USA.
Chalke, H.D., 1962, ‘The impact of tuberculosis on history, literature and art’, Medical
History, 6(4), 301–18.
Chan, J.Z.-M., Sergeant, M.J., Lee, O.Y.-C., Minnikin, D.E., Besra, G.S., Pap, I.,
Spigelman, M., Donoghue, H.D. & Pallen, M.J., 2013, ‘Metagenomic analysis of
tuberculosis in a mummy’, New England Journal of Medicine, 369(3), 289–290.

198
Cock, P.J.A., Fields, C.J., Goto, N., Heuer, M.L. & Rice, P.M., 2010, ‘The Sanger
FASTQ file format for sequences with quality scores, and the Solexa/Illumina
FASTQ variants’, Nucleic Acids Research, 38(6), 1767–71.
Cole, S.T., Eiglmeier, K., Parkhill, J., James, K.D., Thomson, N.R., Wheeler, P.R.,
Honoré, N., Garnier, T., Churcher, C., Harris, D., Mungall, K., Basham, D., Brown,
D., Chillingworth, T., Connor, R., Davies, R.M., Devlin, K., Duthoy, S., Feltwell,
T., Fraser, A., Hamlin, N., Holroyd, S., Hornsby, T., Jagels, K., Lacroix, C.,
Maclean, J., Moule, S., Murphy, L., Oliver, K., Quail, M.A., Rajandream, M.A.,
Rutherford, K.M., Rutter, S., Seeger, K., Simon, S., Simmonds, M., Skelton, J.,
Squares, R., Squares, S., Stevens, K., Taylor, K., Whitehead, S., Woodward, J.R. &
Barrell, B.G., 2001, ‘Massive gene decay in the leprosy bacillus’, Nature, 409,
1007–1011.
Comas, I., Coscolla, M., Luo, T., Borrell, S., Holt, K.E., Parkhill, J., Malla, B., Berg, S.,
Thwaites, G., Yeboah-Manu, D., Bothamley, G., Mei, J., Wei, L., Bentley, S.,
Harris, S.R., Niemann, S., Diel, R., Aseffa, A., Gao, Q., Young, D. & Gagneux, S.,
2013, ‘Out-of-Africa migration and Neolithic co-expansion of Mycobacterium
tuberculosis with modern humans’, Nature Genetics, 45(10), 1176–1182.
Comas, I. & Gagneux, S., 2011, ‘A role for systems epidemiology in tuberculosis
research’, Trends in Microbiology, 19(10), 492–500.
Coros, A., DeConno, E. & Derbyshire, K.M., 2008, ‘IS6110, a Mycobacterium
tuberculosis complex-specific insertion sequence, is also present in the genome of
Mycobacterium smegmatis, suggestive of lateral gene transfer among mycobacterial
species’, Journal of Bacteriology, 190(9), 3408–10.
Cruz-Dávalos, D.I., Nieves-Colón, M.A., Sockell, A., Poznik, G.D., Schroeder, H.,
Stone, A.C., Bustamante, C.D., Malaspinas, A.-S. & Ávila-Arcos, M.C., 2018, ‘In-
solution Y-chromosome capture-enrichment on ancient DNA libraries’, BMC
Genomics, 19(1), 608.
Dabney, J., Knapp, M., Glocke, I., Gansauge, M.-T., Weihmann, A., Nickel, B.,
Valdiosera, C., García, N., Pääbo, S., Arsuaga, J.-L. & Meyer, M., 2013, ‘Complete
mitochondrial genome sequence of a middle Pleistocene cave bear reconstructed
from ultrashort DNA fragments’, Proceedings of the National Academy of Sciences
of the United States of America, 110(39), 15758–15763.
Dabney, J., Meyer, M. & Pääbo, S., 2013, ‘Ancient DNA damage’, Cold Spring Harbor
Perspectives in Biology, 5(7), a012567.
Daniel, T.M., 2006, ‘The history of tuberculosis’, Respiratory Medicine, 100(11), 1862–
1870.
Daniel, V.S. & Daniel, T.M., 1999, ‘Old Testament biblical references to tuberculosis’,
Clinical Infectious Diseases, 29(6), 1557–1558.
Davies, P.D.O., 1998, ‘A little history of tuberculosis’, 3–10. Available at:
http://www.evolve360.co.uk/Data/10/Docs/19/19Davies.pdf.
Davies, R.P.O., Tocque, K., Bellis, M.A., Remmington, T. & Davies, P.D.O., 1999,
‘Historical declines in tuberculosis in England and Wales: improving social
conditions or natural selection?’, The International Journal of Tuberculosis and
Lung Disease, 3(12), 1051-1054.
Davis, G.L., Ray, N.A., Lahiri, R., Gillis, T.P., Krahenbuhl, J.L., Williams, D.L. &
Adams, L.B., 2013, ‘Molecular assays for determining Mycobacterium leprae
viability in tissues of experimentally infected mice’, PLoS Neglected Tropical
Diseases, 7(8), e2404.

199
Dawes, J.D. & Magilton, J., 1980, ‘The Cemetery of St. Helen-on-the-Walls, Aldwark.
12/1’, York: Council for British Archaeology.
de Lima Procópio, R.E. de, Silva, I.R. da, Martins, M.K., Azevedo, J.L. de & Araújo,
J.M. de, 2012, ‘Antibiotics produced by Streptomyces’, The Brazilian Journal of
Infectious Diseases, 16(5), 466–471.
de Luca, G., Barakat, M., Ortet, P. Fochesato, S., Jourlin-Castelli, C., Ansaldi, M., Py,
B., Fichant, G., Coutinho, P.M., Voulhoux, R., Bastien, O., Maréchal, E., Henrissat,
B., Quentin, Y., Noirot, P., Filloux, A., Méjean,V., DuBow, M.S., Barras, F.,
Barbe, V., Weissenbach, J., Michalcescu, I., Verméglio, A., Anchouak, W. &
Heulin, T., 2011, ‘The cyst-dividing bacterium Ramlibacter tataouinensis TTB310
genome reveals a well-stocked toolbox for adaptation to a desert environment’,
PLoS ONE, 6(9), e23784.
Dent, J.S., 1984, Wetwang Slack: an Iron Age cemetery on the Yorkshire Wolds, M. Phil
dissertation, University of Sheffield.
Der Sarkissian, C., Ermini, L., Jonsson, H., Alekseev, A.N., Crubezy, E., Saphiro, B. &
Orlando, L., 2014, 'Shotgun microbial profiling of fossil remains', Molecular
Ecology, 23, 1780-1798.
de Wit, M.Y., Douglas, J.T., McFadden, J. & Klatser, P.R., 1993, ‘Polymerase chain
reaction for detection of Mycobacterium leprae in nasal swab specimens’, Journal
of Clinical Microbiology, 31(3), 502–6.
Dharmendra, 1947, ‘Leprosy in ancient Indian medicine’, International Journal of
Leprosy, 15(4), 424–430.
Donoghue, H.D., 2011, ‘Insights gained from palaeomicrobiology into ancient and
modern tuberculosis’, Clinical Microbiology and Infection, 17(6), 821–829.
Donoghue, H.D., Holton, J. & Spigelman, M., 2001, ‘PCR primers that can detect low
levels of Mycobacterium leprae DNA’, Journal of Medical Microbiology, 50, 177–
182.
Donoghue, H.D., Marcsik, A., Matheson, C., Vernon, K., Nuorala, E., Molto, J.E.,
Greenblatt, C.L. & Spigelman, M., 2005, ‘Co-infection of Mycobacterium
tuberculosis and Mycobacterium leprae in human archaeological samples: a
possible explanation for the historical decline of leprosy’, Proceedings of the Royal
Society of London B: Biological Sciences, 272(1561), 389–394.
Donoghue, H.D., Spigelman, M., O’Grady, J., Szikossy, I., Pap, I., Lee, O.Y.-C., Wu,
H.H.T., Besra, G.S. & Minnikin, D.E., 2015, ‘Ancient DNA analysis – An
established technique in charting the evolution of tuberculosis and leprosy’,
Tuberculosis, 95, S140–S144.
Dzierzykray-Rogalski, T., 1980, ‘Paleopathology of the Ptolemaic inhabitants of
Dakhleh Oasis (Egypt)’, Journal of Human Evolution, 9(1), 71–74.
Eiglmeier, K., Parkhill, J., Honoré, N., Garnier, T., Tekaia, F., Telenti, A., Klatser, P.,
James, K.D., Thomson, N.R., Wheeler, P.R., Churcher, C., Harris, D., Mungall, K.,
Barrell, B.G. & Cole, S.T., 2001, ‘The decaying genome of Mycobacterium
leprae.’, Leprosy Review, 72(4), 387–98.
Eisenach, K.D., Cave, M.D., Bates, J.H. & Crawford, J.T., 1990, ‘Polymerase chain
reaction amplification of a repetitive DNA sequence specific for Mycobacterium
tuberculosis.’, The Journal of Infectious Diseases, 161(5), 977–81.

200
Fernández-Roblas, R., Esteban, J., Cabria, F., López, J.C., Jiménez, M.S. & Soriano, F.,
2000, ‘In vitro susceptibilities of rapidly growing mycobacteria to telithromycin
(HMR 3647) and seven other antimicrobials’, Antimicrobial Agents and
Chemotherapy, 44(1), 181–2.
Feuillie, C., Merheb, M.M., Gillet, B., Montagnac, G., Daniel, I. & Hänni, C., 2014,
‘Detection of DNA sequences refractory to PCR amplification using a biophysical
SERRS assay (Surface Enhanced Resonant Raman Spectroscopy)’, PloS ONE,
9(12), e114148.
Filliol, I., Motiwala, A.S., Cavatore, M., Qi, W., Hazbón, M.H., del Valle, M.B, Fyfe, J.,
García-garcía, L., Rastogi, N., Sola, C., Zozio, T., Guerrero, M.I., León, C.I.,
Crabtree, J., Angiuoli, S., Eisenach, K.D., Durmaz, R., Joloba, M.L., Rendón, A.,
Sifuentes-Osornio, J., de León, A.P., Cave, M.D., Fleischmann, R., Whittam, T.S.
& Alland, D., 2006, ‘Global phylogeny of Mycobacterium tuberculosis based on
Single Nucleotide Polymorphism ( SNP ) analysis : insights into tuberculosis
evolution, phylogenetic accuracy of other DNA fingerprinting systems, and
recommendations for a minimal standard SNP Set’, Journal of Bacteriology,
188(2), 759-772.
Firth, J., 2014, ‘History of tuberculosis part 1 - phthisis, consumption and the white
plague’, History, 22(2), 29–35.
Flesch, I.E.A. & Kaufmann, H.E., 1993, ‘Role of cytokines in tuberculosis’,
Immunobiology, 189, 316–339.
Forbes, B.A. & Hicks, K.E., 1996, ‘Substances interfering with direct detection of
Mycobacterium tuberculosis in clinical specimens by PCR: effects of bovine serum
albumin’, Journal of Clinical Microbiology, 34(9), 2125–8.
Forrellad, M.A., Klepp, L.I., Gioffré, A., y García, J.S., Morbidoni, H.R., de la Paz
Santangelo, M., Cataldi, A.A. & Bigi, F., 2013, ‘Virulence factors of the
Mycobacterium tuberculosis complex’, Virulence, 4(1), 3–66.
Ghosh, S. & Chaudhuri, S., 2015, ‘Chronicles of Gerhard-Henrik Armauer Hansen’s life
and work’, Indian Journal of Dermatology, 60(3), 219–21.
Gilbert, M.T.P., Hansen, A.J., Willerslev, E., Rudbeck, L., Barnes, I., Lynnerup, N. &
Cooper, A., 2003, ‘Characterization of genetic miscoding lesions caused by
postmortem damage’, American Journal of Human Genetics, 72(1), 48–61.
Gilbert, M.T.P., Tomsho, L.P., Rendulic, S., Packard, M., Drautz, D.I., Sher, A.,
Tikhonov, A., Dalen, L., Kuznetsova, T., Kosintsev, P., Campos, P.F., Higham, T.,
Collins, M.J., Wilson, A.S., Shidlovskiy, F., Buigues, B., Ericson, P.G.P.,
Germonpre, M., Gotherstrom, A., Iacumin, P., Nikolaev, V., Nowak-Kemp, M.,
Willerslev, E., Knight, J.R., Irzyk, G.P., Perbost, C.S., Fredrikson, K.M., Harkins,
T.T., Sheridan, S., Miller, W. & Schuster, S.C., 2007, ‘Whole-genome shotgun
sequencing of mitochondria from ancient hair shafts’, Science, 317(5846), 1927–
1930.
Golden, M.P. & Vikram, H.R., 2005, ‘Extrapulmonary tuberculosis: an overview',
American Family Physician, 72(9), 1761–1768.
Gomez-Valero, L., Rocha, E.P.C., Latorre, A. & Silva, F.J., 2007, ‘Reconstructing the
ancestor of Mycobacterium leprae: the dynamics of gene loss and genome
reduction’, Genome Research, 17, 1178–1185.
Gordon, S.V, Brosch, R., Billault, A., Garnier, T., Eiglmeier, K. & Cole, S.T., 1999,
‘Identification of variable regions in the genomes of tubercle bacilli using bacterial
artificial chromosome arrays’, Molecular Microbiology, 32(3), 643–55.

201
Green, E.J. & Speller, C.F., 2017, ‘Novel substrates as sources of ancient DNA:
prospects and hurdles’, Genes, 8(7), 180.
Green, R.E., Malaspinas, A.-S., Krause, J., Briggs, A.W., Johnson, P.L.F., Uhler, C.,
Meyer, M., Good, J.M., Maricic, T., Stenzel, U., Prüfer, K., Siebauer, M., Burbano,
H.A., Ronan, M., Rothberg, J.M., Egholm, M., Rudan, P., Brajković, D., Kućan, Ž.,
Gušić, I., Wikström, M., Laakkonen, L., Kelso, J., Slatkin, M. & Pääbo, S., 2008,
‘A complete Neandertal mitochondrial genome sequence determined by high-
throughput sequencing’, Cell, 134(3), 416–426.
Groathouse, N.A., Brown, S.E., Knudson, D.L., Brennan, P.J. & Slayden, R.A., 2006,
‘Isothermal amplification and molecular typing of the obligate intracellular
pathogen Mycobacterium leprae isolated from tissues of unknown origins’, Journal
of Clinical Microbiology, 44(4), 1502–1508.
Gron, K., 1973, ‘Leprosy in literature and art’, International Journal of Leprosy, 41(2),
249–283.
Gruber, K., 2015, ‘Here, there, and everywhere: from PCRs to next-generation
sequencing technologies and sequence databases, DNA contaminants creep in from
the most unlikely places’, EMBO Reports, 16(8), 898–901.
Grzybowski, A. & Nita, M., 2016, ‘Leprosy in the bible’, Clinics in Dermatology, 34, 3–
7.
Gunatilake, S.B. & Settinayake, S., 2004, ‘Lepsory’, Practical Neurology, 4, 194–203.
Gunawan, H., Haryati, N.S. & Suwarsa, O., 2017, 'Swan neck deformity mimicking
claw hand caused by arthritis in leprosy', Case Reports in Dermatology, 9(3), 231-
235.
Gupta, R.S., Lo, B. & Son, J., 2018, ‘Phylogenomics and comparative genomic studies
robustly support division of the genus Mycobacterium into an emended genus
Mycobacterium and four novel genera’, Frontiers in Microbiology, 9, 67.
Gutierrez, M.C., Brisse, S., Brosch, R., Fabre, M., Omaïs, B., Marmiesse, M., Supply, P.
& Vincent, V., 2005, ‘Ancient origin and gene mosaicism of the progenitor of
Mycobacterium tuberculosis’, PLoS Pathogens, 1(1), e5.
Haas, C.J., Zink, A., Palfi, G., Szeimies, U. & Nerlich, A.G., 2000, ‘Detection of leprosy
in ancient human skeletal remains by molecular identification of Mycobacterium
leprae’, American Journal of Clinical Pathology, 114, 428–436.
Hall, R.A. & Whyman, M., 1996, 'Settlement and Monasticism at Ripon, North
Yorkshire, from the 7th to 11th Centuries A.D.', Medieval Archaeology, 40(1), 62-
150.
Hagelberg, E. & Clegg, J.B., 1991, ‘Isolation and characterization of DNA from
archaeological bone.’, Proceedings of the Royal Society of London B: Biological
Sciences, 244(1309), 45–50.
Han, C., Spring, S., Lapidus, A., Rio, T.G. Del, Tice, H., Copeland, A., Cheng, J.-F.,
Lucas, S., Chen, F., Nolan, M., Bruce, D., Goodwin, L., Pitluck, S., Ivanova, N.,
Mavromatis, K., Mikhailova, N., Pati, A., Chen, A., Palaniappan, K., Land, M.,
Hauser, L., Chang, Y.-J., Jeffries, C.C., Saunders, E., Chertkov, O., Brettin, T.,
Göker, M., Rohde, M., Bristow, J., Eisen, J.A., Markowitz, V., Hugenholtz, P.,
Kyrpides, N.C., Klenk, H.-P. & Detter, J.C., 2009, ‘Complete genome sequence of
Pedobacter heparinus type strain (HIM 762-3T)’, Standards in Genomic Sciences,
1(1), 54–62.

202
Han, X.Y., Seo, Y.H., Sizer, K.C., Schoberle, T., May, G.S., Spencer, J.S., Li, W. &
Nair, R.G., 2008, ‘A new Mycobacterium species causing diffuse lepromatous
leprosy’, American Journal of Clinical Pathology, 130(6), 856–864.
Hartskeerl, R.A., de Wit, M.Y.L. & Klatser, P.R., 1989, ‘Polymerase chain reaction for
the detection of Mycobacterium leprae', Microbiology, 135(2357–2364).
Hershberg, R., Lipatov, M., Small, P.M., Sheffer, H., Niemann, S., Homolka, S., Roach,
J.C., Kremer, K., Petrov, D.A, Feldman, M.W. & Gagneux, S., 2008, ‘High
functional diversity in Mycobacterium tuberculosis driven by genetic drift and
human demography’, PLoS Biology, 6(12), e311.
Hershkovitz, I., Arensburg, B. & Speirs, M., 1993, 'Leprosy or madura foot? The
ambigious nature of infectious disease in palaeopathology: reply to Dr.
Manchester', American Journal of Physical Anthropology', 91, 251-253.
Hershkovitz, I., Donoghue, H.D., Minnikin, D.E., Besra, G.S., Lee, O.Y.-C., Gernaey,
A.M., Galili, E., Eshed, V., Greenblatt, C.L., Lemma, E., Bar-Gal, G.K.,
Spigelman, M., Kahila Bar-Gal, G. & Spigelman, M., 2008, ‘Detection and
molecular characterization of 9,000-year-old Mycobacterium tuberculosis from a
Neolithic settlement in the Eastern Mediterranean.’, PloS one, 3(10), e3426.
Heyn, P., Stenzel, U., Briggs, A.W., Kircher, M., Hofreiter, M. & Meyer, M., 2010,
‘Road blocks on paleogenomes-polymerase extension profiling reveals the
frequency of blocking lesions in ancient DNA’, Nucleic Acids Research, 38(16),
e161.
Higashino, A., Sakate, R., Kameoka, Y., Takahashi, I., Hirata, M., Tanuma, R., Masui,
T., Yasutomi, Y. & Osada, N., 2012, ‘Whole-genome sequencing and analysis of
the Malaysian Cynomolgus macaque (Macaca fascicularis) genome’, Genome
Biology, 13(7), R58.
Higuchi, R., Bowman, B., Freiberger, M., Ryder, O.A. & Wilson, A.C., 1984, ‘DNA
sequences from the quagga, an extinct member of the horse family’, Nature,
312(5991), 282–284.
Hirst, S.M., 1985, ‘An Anglo-Saxon inhumation cemetery at Sewerby, East Yorkshire’,
York University Archaeological Publications 4.
Ho, Y.S., Adroub, S.A., Abadi, M., Alwan, B. Al, Alkhateeb, R., Gao, G., Ragab, A.,
Ali, S., van Soolingen, D., Bitter, W., Pain, A. & Abdallah, A.M., 2012, ‘Complete
genome sequence of Mycobacterium vaccae type strain ATCC 25954’, Journal of
Bacteriology, 194(22), 6339–40.
Hofreiter, M., Jaenicke, V., Serre, D., Haeseler, A. Von & Paabo, S., 2001, ‘DNA
sequences from multiple amplifications reveal artifacts induced by cytosine
deamination in ancient DNA’, Nucleic Acids Research, 29(23), 4793–4799.
Holloway, K.L., Henneberg, R.J., de Barros Lopes, M. & Henneberg, M., 2011,
‘Evolution of human tuberculosis: a systematic review and meta-analysis of
paleopathological evidence’, HOMO, 62(6), 402–458.
Holloway, K.L., Link, K., Ruhli, F. & Hennenberg, M., 2013, 'Skeletal lesions in human
tuberculosis may sometimes heal: and aid to palaeopathological diagnoses', PLoS
ONE, 8(4). e62798.
Holst, M., 2005, Artefacts environmental evidence: the human bone, in Spall, C.A. &
Toop, N.J. (eds) Report on Excavations: July 2000–July 2002, Blue Bridge Lane
and Fishergate House: York, APC Monograph Series.

203
Holst, M., 2008, ‘Osteological analysis: Heslington East, York’, York Osteoarchaeology
No. 1108, Unpublished.
Homolka, S., Projahn, M., Feuerriegel, S., Ubben, T., Diel, R., Nübel, U. & Niemann,
S., 2012, ‘High resolution discrimination of clinical Mycobacterium tuberculosis
complex strains based on single nucleotide polymorphisms’, PLoS ONE, 7(7),
e39855.
Horsburgh, C.R., Barry, C.E. & Lange, C., 2015, ‘Treatment of tuberculosis’, The New
England Journal of Medicine, 373(22), 2149–2160.
Huson, D.H., Auch, A.F., Qi, J. & Schuster, S.C., 2007, ‘MEGAN analysis of
metagenomic data’, Genome Research, 17(3), 377–386.
Hussain, T., 2007, ‘Leprosy and tuberculosis: an insight-review’, Critical Reviews in
Microbiology, 33(1), 15–66.
Huyen, M.N.T., Tiemersma, E.W., Kremer, K., de Haas, P., Lan, N.T.N., Buu, T.N.,
Sola, C., Cobelens, F.G.J. & van Soolingen, D., 2013, ‘Characterisation of
Mycobacterium tuberculosis isolates lacking IS6110 in Viet Nam’, The
International Journal of Tuberculosis and Lung Disease, 17(11), 1479–1485.
Ichikawa, N., Oguchi, A., Ikeda, H., Ishikawa, J., Kitani, S., Watanabe, Y., Nakamura,
S., Katano, Y., Kishi, E., Sasagawa, M., Ankai, A., Fukui, S., Hashimoto, Y.,
Kamata, S., Otoguro, M., Tanikawa, S., Nihira, T., Horinouchi, S., Ohnishi, Y.,
Hayakawa, M., Kuzuyama, T., Arisawa, A., Nomoto, F., Miura, H., Takahashi, Y.
& Fujita, N., 2010, ‘Genome sequence of Kitasatospora setae NBRC 14216T: an
evolutionary snapshot of the family Streptomycetaceae’, DNA Research, 17(6),
393–406.
Inskip, S., Taylor, G.M., Anderson, S. & Stewart, G., 2017, ‘Leprosy in pre-Norman
Suffolk, UK: biomolecular and geochemical analysis of the woman from Hoxne’,
Journal of Medical Microbiology, 66(11), 1640–1649.
Irgens, L.M., 1984, ‘The discovery of Mycobacterium leprae. A medical achievement in
the light of evolving scientific methods’, The American Journal of
Dermatopathology, 6(4), 337–43.
Jacob, J.T. & Franco-Paredes, C., 2008, ‘The stigmatization of leprosy in India and its
impact on future approaches to elimination and control.’, PLoS Neglected Tropical
Diseases, 2(1), e113.
Jacobson, R.R. & Krahenbuhl, J.L., 1999, ‘Leprosy’, Lancet, 353(9153), 655–660.
Jin, S.-H., Ahn, K.J. & An, S., 2018, ‘Importance of the immune response to
Mycobacterium leprae in the skin’, Biomedical Dermatology, 2(1), 1.
Kaestle, F. A. & Horsburgh, K. A., 2002, ‘Ancient DNA in anthropology: methods,
applications, and ethics’, Yearbook of Physical Anthropology, 45, 92-130.
Kapur, V., Whittam, T.S. & Musser, J.M., 1994, ‘Is Mycobacterium tuberculosis 15,000
Years Old?’, The Journal of Infectious Diseases, 170(5), 1348–1349.
Karanam, K., Loewer, A. & Lahav, G., 2013, ‘Dynamics of the DNA damage response:
insights from live-cell imaging’, Briefings in Functional Genomics, 12(2), 109–17.
Kay, G.L., Sergeant, M.J., Zhou, Z., Chan, J.Z.-M.Z.-M., Millard, A., Quick, J.,
Szikossy, I., Pap, I.I., Spigelman, M., Loman, N.J., Achtman, M., Donoghue, H.D.
& Pallen, M.J., 2015, ‘Eighteenth-century genomes show that mixed infectoins
were common at time of peak tuberculosis in Europe’, Nature Communications,
6(6717), 1–9.

204
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S., Buxton,
S., Cooper, A., Markowitz, S., Duran, C., Thierer, T., Ashton, B., Meintjes, P. &
Drummond, A., 2012, ‘Geneious Basic: an integrated and extendable desktop
software platform for the organization and analysis of sequence data’,
Bioinformatics, 28, 1647–1649.
Kelley, M.A. & Micozzi, M.S., 1984, ‘Rib lesions in chronic pulmonary tuberculosis’,
American Journal of Physical Anthropology, 65(4), 381–386.
Khalil, R.B. & Richa, S., 2014, ‘When affective disorders were considered to emanate
from the heart: the Ebers Papyrus’, American Journal of Psychiatry, 171(3), 275.
Kim, Y.-J. & Wilson, D.M., 2012, ‘Overview of base excision repair biochemistry.’,
Current Molecular Pharmacology, 5(1), 3–13.
Kircher, M., Heyn, P. & Kelso, J., 2011, ‘Addressing challenges in the production and
analysis of illumina sequencing data’, BMC Genomics, 12, 382.
Kircher, M., Sawyer, S. & Meyer, M., 2012, ‘Double indexing overcomes inaccuracies
in multiplex sequencing on the Illumina platform’, Nucleic Acids Research, 40(1),
e3.
Kirchheimer, W.F. & Storrs, E.E., 1971, ‘Attempts to establish the armadillo (Dasypus
novemcinctus Linn.) as a model for the study of leprosy. I. Report of lepromatoid
leprosy in an experimentally infected armadillo’, International Journal of Leprosy
and Other Mycobacterial Diseases, 39(3), 693–702.
Kirchheimer, W.F., Storrs, E.E. & Binford, C.H., 1972, ‘Attempts to establish the
Armadillo (Dasypus novemcinctus Linn.) as a model for the study of leprosy. II.
Histopathologic and bacteriologic post-mortem findings in lepromatoid leprosy in
the Armadillo’, International Journal of Leprosy and Other Mycobacterial
Diseases, 40(3), 229–42.
Kistler, L., Ware, R., Smith, O., Collins, M. & Allaby, R.G., 2017, ‘A new model for
ancient DNA decay based on paleogenomic meta-analysis’, Nucleic Acids
Research, 45(11), 6310–6320.
Klaus, H.D., 2017, 'Paleopathological rigor and differential diagnosis: case studies
involving terminology, description, and diagnostic frameworks for scurvy in
skeletal remains', International Journal of Palaeopathology, 19, 96-110.
Knapp, M. & Hofreiter, M., 2010, ‘Next generation sequencing of ancient DNA:
requirements, strategies and perspectives’, Genes, 1(2), 227–243.
Knapp, M., Lalueza-Fox, C. & Hofreiter, M., 2015, ‘Re-inventing ancient human DNA’,
Investigative Genetics, 6(1), 4.
Knechel, N.A., 2009, ‘Tuberculosis: pathophysiology, clinical features, and diagnosis’,
Critical Care Nurse, 29(2), 34–43.
Konomi, N., Lebwohl, E., Mowbray, K., Tattersall, I. & Zhang, D., 2002, ‘Detection of
Mycobacterial DNA in Andean mummies’, Journal of Clinical Microbiology,
40(12), 4738–40.
Kumar, A., Girdhar, A. & Girdhar, B.K., 2013, ‘Twelve months fixed duration WHO
multidrug therapy for multibacillary leprosy: incidence of relapses in Agra field
based cohort study’, The Indian Journal of Medical Research, 138(4), 536–40.
Lakhtakia, R., 2013, ‘Of animalcula, phthisis and scrofula: historical insights into
tuberculosis in the pre-Koch era’, Sultan Qaboos University Medical Journal,
13(4), 486–490.

205
Lalucat, J., Bennasar, A., Bosch, R., García-Valdés, E. & Palleroni, N.J., 2006, ‘Biology
of Pseudomonas stutzeri’, Microbiology and Molecular Biology Reviews, 70(2),
510–47.
Lampky, J.R., 1971, ‘Distribution of Sorangium cellulosum’, Applied Microbiology,
22(5), 937–938.
Lange, C., Abubakar, I., Alffenaar, J.-W.C., Bothamley, G., Caminero, J.A., Carvalho,
A.C.C., Chang, K.-C., Codecasa, L., Correia, A., Crudu, V., Davies, P., Dedicoat,
M., Drobniewski, F., Duarte, R., Ehlers, C., Erkens, C., Goletti, D., Günther, G.,
Ibraim, E., Kampmann, B., Kuksa, L., de Lange, W., van Leth, F., van Lunzen, J.,
Matteelli, A., Menzies, D., Monedero, I., Richter, E., Rüsch-Gerdes, S., Sandgren,
A., Scardigli, A., Skrahina, A., Tortoli, E., Volchenkov, G., Wagner, D., van der
Werf, M.J., Williams, B., Yew, W.-W., Zellweger, J.-P., Cirillo, D.M. & TBNET,
2014, ‘Management of patients with multidrug-resistant/extensively drug-resistant
tuberculosis in Europe: a TBNET consensus statement’, The European Respiratory
Journal, 44(1), 23–63.
Lastória, J.C. & Morgado De Abreu, M.A.M., 2014, ‘Leprosy: review of the
epidemiological, clinical, and etiopathogenic aspects-Part 1’, Anais Brasileiros de
Dermatologia, 89(2), 205–218.
Lee, F., No date, ‘The pathological report on selected skeletons from the Anglo-Saxon
levels at York Minster’, Calvin Wells Laboratory University of Bradford,
Unpublished.
Leung, A.K.C., 2009, ‘Introduction’, in Leung A.K.C. (ed.), Leprosy in China: a
history, 1–16, Columbia University Press, New York.
Li, H. & Durbin, R., 2009, ‘Fast and accurate short read alignment with Burrows-
Wheeler transform’, Bioinformatics, 15, 1754–1760.
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., Marth, G.,
Abecasis, G. & Durbin, R., 2009, ‘The Sequence Alignment/Map format and
SAMtools’, Bioinformatics, 25(16), 2078–2079.
Lin, P.L. & Flynn, J.L., 2010, ‘Understanding latent tuberculosis: a moving target’, The
Journal of Immunology, 185(1), 15–22.
Lindahl, T. & Andersson, A., 1972, ‘Rate of chain breakage at apurinic sites in double-
stranded deoxyribonucleic acid’, Biochemistry, 11(19), 3618–3623.
Lindahl, T. & Nyberg, B., 1972, ‘Rate of depurination of native deoxyribonucleic acid’,
Biochemistry, 11(19), 3610–3618.
Lindgreen, S., 2012, ‘AdapterRemoval: easy cleaning of next-generation sequencing
reads’, BMC Research Notes, 5, 337–341.
Llamas, B., Valverde, G., Fehren-Schmitz, L., Weyrich, L.S., Cooper, A. & Haak, W.,
2016, ‘From the field to the laboratory: controlling DNA contamination in human
ancient DNA research in the high-throughput sequencing era’, Science and
Technology of Archaeological Research, 3(1), 1–14.
Luke, S. & Verma, R.S., 1995, ‘Human (Homo sapiens) and chimpanzee (Pan
troglodytes) share similar ancestral centromeric alpha satellite DNA sequences but
other fractions of heterochromatin differ considerably’, American Journal of
Physical Anthropology, 96(1), 63–71.
Lu, Z., Wu, J. & Cheng, G., 2015, 'Leprosy', in Li, H. (ed), Radiology of Infectious
Diseases: Volume 2, 131-142, Springer, Dordrecht.
Lynnerup, N. & Boldsen, J., 2012, ‘Leprosy (Hansen’s disease)’, in Grauer A.L. (ed.), A
Companion to Paleopathology, 458–471, Wiley-Blackwell, Oxford UK.

206
Maciejewska, M., Pessi, I.S., Arguelles-Arias, A., Noirfalise, P., Luis, G., Ongena, M.,
Barton, H., Carnol, M. & Rigali, S., 2015, ‘Streptomyces lunaelactis sp. nov., a
novel ferroverdin A-producing Streptomyces species isolated from a moonmilk
speleothem’, Antonie van Leeuwenhoek, 107(2), 519–531.
MacRaild, D. M., 1999, Irish migrants in modern Britain, 1750-1922, St. Martin’s Press
Inc, New York USA.
Malathi, M. & Thappa, D.M., 2013, ‘Fixed-duration therapy in leprosy: limitations and
opportunities’, Indian Journal of Dermatology, 58(2), 93–100.
Manchester, K. & Roberts, C.A., 1986, ‘Palaeopathological evidence of tuberculosis and
leprosy in Britain’, SERC Report (Grant 337.367).
Mark, S., 2002, ‘Alexander the Great, geafaring, and the spread of leprosy’, Journal of
the History of Medicine and Allied Sciences, 57(3), 285–311.
Martín, M.C., Manteca, A., Castillo, M.L., Vázquez, F. & Méndez, F.J., 2004,
‘Streptomyces albus isolated from a human actinomycetoma and characterized by
molecular techniques’, Journal of Clinical Microbiology, 42(12), 5957–60.
Martinez, T.S., Figueira, M.M.N.., Costa, A.V., Gonçalves, M.A., Goulart, L.R. &
Goulart, I.M.B., 2011, ‘Oral mucosa as a source of Mycobacterium leprae infection
and transmission, and implications of bacterial DNA detection and the
immunological status’, Clinical Microbiology and Infection, 17(11), 1653–1658.
Masson, M., Bereczki, Z., Molnár, E., Donoghue, H.D., Minnikin, D.E., Lee, O.Y.-C.,
Wu, H.H.T., Besra, G.S. & Bull, I.D., 2015, ‘7000 year-old tuberculosis cases from
Hungary – osteological and biomolecular evidence’, Tuberculosis, 95, S13–S17.
Matheson, C.D., Vernon, K.K., Lahti, A., Fratpietro, R., Spigelman, M., Gibson, S.,
Greenblatt, C.L., Donoghue, H.D. & Zissu, B., 2009, ‘Molecular exploration of the
first-century tomb of the Shroud in Akeldama, Jerusalem’, PLoS ONE, 4(12),
e8319.
Matos, V. & Santos, A.L., 2006, ‘On the trail of pulmonary tuberculosis based on rib
lesions: results from the human identified skeletal collection from the Museu
Bocage (Lisbon, Portugal)’, American Journal of Physical Anthropology, 130(2),
190–200.
Matossian, M.K., 1985, ‘Death in London’, The Journal of Interdisciplinary History,
16(2), 183–197.
Mays, S., Harding, C. & Heighway, C., 2007, ‘Wharram XI: The churchyard York’,
York University Press.
Mays, S. & Taylor, G.M., 2003, ‘A first prehistoric case of tuberculosis from Britain’,
International Journal of Osteoarchaeology, 13(4), 189–196.
McKenna, A., Hanna, M., Banks, E., Sivachenko, A., Cibulskis, K., Kernytsky, A.,
Garimella, K., Altshuler, D., Gabriel, S., Daly, M. & DePristo, M.A., 2010, ‘The
Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation
DNA sequencing data’, Genome Research, 20(9), 1297–1303.
McMurry, J.L. & Chang, M.C.Y., 2017, ‘Fluorothreonyl-tRNA deacylase prevents
mistranslation in the organofluorine producer Streptomyces cattleya’, Proceedings
of the National Academy of Sciences of the United States of America, 114(45),
11920–11925.
McVaugh, M. R., 2006, The rational surgery of Middle Ages, SISMEL. Edizioni del
Galluzzo, Florence.

207
Mendum, T.A., Schuenemann, V.J., Roffey, S., Taylor, G., Wu, H., Singh, P., Tucker,
K., Hinds, J., Cole, S.T., Kierzek, A.M., Nieselt, K., Krause, J. & Stewart, G.R.,
2014, ‘Mycobacterium leprae genomes from a British medieval leprosy hospital:
towards understanding an ancient epidemic’, BMC Genomics, 15(1), 270.
Metcalfe, N.H., 2007, ‘A description of the methods used to obtain information on
ancient disease and medicine and how the evidence has survived’, Postgraduate
Medical Journal, 83(984):655-658.
Meyer, M. & Kircher, M., 2010, ‘Illumina sequencing library preparation for highly
multiplexed target capture and sequencing’, Cold Spring Harbor Protocols,
2919(6), pdb.prot5448.
Meyers, W.M., Gormus, B.J., Walsh, G.P., B, B.G. & B, H.G., 1991, ‘Naturally
acquired and experimental leprosy in nonhuman primates’, The American Journal
of Tropical Medicine and Hygiene, 44(4 Pt 2), 24–27.
Mitchell, P.D., 2011, ‘Retrospective diagnosis and the use of historical texts for
investigating disease in the past’, International Journal of Palaeopathology, 1(2),
81-88.
Mitchell, P.D., 2017, ‘Improving the use of historical written sources in
palaeopathology’, International Journal of Palaeopathology, 19, 88-95.
Modlin, R.L., 1994, ‘Th1-Th2 paradigm: insights from leprosy', The Journal of
Investigative Dermatology, 102(6), 828–32.
Modlin, R.L., 2010, ‘The innate immune response in leprosy’, Current Opinion in
Immunology, 22(1), 48–54.
Mohammad, W., Malhotra, S.K. & Garg, P.K., 'Clinico-radiological correlation of bone
changes in leprosy patients presenting with disabilities/deformities', Indian Journal
of Leprosy, 88(2), 83-95.
Mohr, K.I., Garcia, R.O., Gerth, K., Irschik, H. & Muller, R., 2012, ‘Sandaracinus
amylolyticus gen. nov., sp. nov., a starch-degrading soil myxobacterium, and
description of Sandaracinaceae fam. nov’, International Journal of Systematic and
Evolutionary Microbiology, 62(Pt 5), 1191–1198.
Moissenet, D., Becker, K., Mérens, A., Ferroni, A., Dubern, B. & Vu-Thien, H., 2012,
‘Persistent bloodstream infection with Kocuria rhizophila related to a damaged
central catheter’, Journal of Clinical Microbiology, 50(4), 1495–8.
Møller-Christensen, V., 1953, ‘Location and excavation of the first Danish leper
graveyard from the Middle Ages - St. Jørgen’s Farm, Naestved’, Bulletin of the
History of Medicine, 27(2), 112–123.
Mongodin, E.F., Shapir, N., Daughtery, S.C., DeBoy, R.T., Emerson, J.B., Shvartzbeyn,
A., Radune, D., Vamathevan, J., Riggs, F., Grinberg, V., Khouri, H., Wackett, L.P.,
Nelson, K.E. & Sadowsky, M.J., 2006, ‘Secrets of soil survival revealed by the
qenome sequence of Arthrobacter aurescens TC1’, PLoS Genetics, 2(12), e214.
Monot, M., Honoré, N., Garnier, T., Araoz, R., Coppée, J.-Y.Y., Lacroix, C., Sow, S.,
Spencer, J.S., Truman, R.W., Williams, D.L., Gelber, R., Virmond, M., Flageul, B.,
Cho, S.-N.N., Ji, B., Paniz-Mondolfi, A., Convit, J., Young, S., Fine, P.E.,
Rasolofo, V., Brennan, P.J. & Cole, S.T., 2005, ‘On the origin of leprosy’, Science,
308(5724), 1040–1042.

208
Monot, M., Honoré, N., Garnier, T., Zidane, N., Sherafi, D., Paniz-Mondolfi, A.,
Matsuoka, M., Taylor, G.M., Donoghue, H.D., Bouwman, A., Mays, S., Watson,
C., Lockwood, D., Khamispour, A., Dowlati, Y., Jianping, S., Rea, T.H., Vera-
Cabrera, L., Stefani, M.M., Banu, S., Macdonald, M., Sapkota, B.R., Spencer, J.S.,
Thomas, J., Harshman, K., Singh, P., Busso, P., Gattiker, A., Rougemont, J.,
Brennan, P.J. & Cole, S.T., 2009, ‘Comparative genomic and phylogeographic
analysis of Mycobacterium leprae’, Nature Genetics, 41(12), 1282–1289.
Montiel, R., García, C., Cañadas, M.P., Isidro, A., Guijo, J.M. & Malgosa, A., 2003,
‘DNA sequences of Mycobacterium leprae recovered from ancient bones’, FEMS
Microbiology Letters, 226(2), 413–4.
Moonan, P.K., 2018, ‘Tuberculosis—the face of struggles, the struggles we face, and the
dreams that lie within’, Emerging Infectious Diseases, 24(3), 592–593.
Müldner, G., Chenery, C. & Eckardt, H., 2011, ‘The ‘headless Romans’: multi-isotope
investigations of an unusual burial ground from Roman Britain’, Journal of
Archaeological Science, 38,280–290.
Müller, R., 2008, The biomolecular archaeology of leprosy, PhD thesis, University of
Bradford.
Müller, R., Roberts, C.A. & Brown, T.A., 2014a, ‘Biomolecular identification of ancient
Mycobacterium tuberculosis complex DNA in human remains from Britain and
continental Europe’, American Journal of Physical Anthropology, 153, 178–189.
Müller, R., Roberts, C.A. & Brown, T.A., 2014b, ‘Genotyping of ancient
Mycobacterium tuberculosis strains reveals historic genetic diversity’, Proceedings
of the Royal Society of London B: Biological Sciences, 281 (17181), 20133236.
Müller, R., Roberts, C.A. & Brown, T.A., 2015, ‘Complications in the study of ancient
tuberculosis: non-specificity of IS6110 PCRs’, Science and Technology of
Archaeological Research, 1(1), 1–8.
Müller, R., Roberts, C.A. & Brown, T.A., 2016, ‘Complications in the study of ancient
tuberculosis: presence of environmental bacteria in human archaeological remains’,
Journal of Archaeological Science, 68, 5–11.
Mullis, K., Faloona, F., Scharf, S., Saiki, R., Horn, G. & Erlich, H., 1986, ‘Specific
enzymatic amplification of DNA in Vvtro: the polymerase chain reaction’, Cold
Spring Harbor Symposia on Quantitative Biology, 51, 263–273.
Nagao, M., Sonobe, M., Bando, T., Saito, T., Shirano, M., Matsushima, A., Fujihara, N.,
Takakura, S., Iinuma, Y. & Ichiyama, S., 2009, ‘Surgical site infection due to
Mycobacterium peregrinum: a case report and literature review’, International
Journal of Infectious Diseases, 13(2), 209–211.
Neal, C. & Roskams, S., 2012, ‘Assessment report Heslington East, Volumes 1 and 2’,
Archaeology Data Service archive, http://archaeologydataservice.ac.uk.
Nerlich, A.G. & Zink, A.R., 2008, Past leprae, in Raoult, D. & Drancourt, M. (eds.),
Paleomicrobiology: Past human infections, 99–123, Springer-Verlag, Berlin
Heidelberg.
Nicholls, H., 2005, ‘Ancient DNA comes of age’, PLoS Biology, 3(2), e56.
Nicklisch, N., Maixner, F., Ganslmeier, R., Friederich, S., Dresely, V., Meller, H., Zink,
A. & Alt, K.W., 2012, ‘Rib lesions in skeletons from early Neolithic sites in central
Germany: on the trail of tuberculosis at the onset of agriculture’, American Journal
of Physical Anthropology, 149(3), 391–404.

209
Nolan, M., Sikorski, J., Jando, M., Lucas, S., Lapidus, A., Glavina Del Rio, T., Chen, F.,
Tice, H., Pitluck, S., Cheng, J.-F., Chertkov, O., Sims, D., Meincke, L., Brettin, T.,
Han, C., Detter, J.C., Bruce, D., Goodwin, L., Land, M., Hauser, L., Chang, Y.-J.,
Jeffries, C.D., Ivanova, N., Mavromatis, K., Mikhailova, N., Chen, A., Palaniappan,
K., Chain, P., Rohde, M., Göker, M., Bristow, J., Eisen, J.A., Markowitz, V.,
Hugenholtz, P., Kyrpides, N.C. & Klenk, H.-P., 2010, ‘Complete genome sequence
of Streptosporangium roseum type strain (NI 9100T)’, Standards in Genomic
Sciences, 2(1), 29–37.
Noonan, J.P., Hofreiter, M., Smith, D., Priest, J.R., Rohland, N., Rabeder, G., Krause, J.,
Detter, C., Pääbo, S. & Rubin, E.M., 2005, ‘Genomic sequencing of Pleistocene
cave bears’, Science, 309(5721), 597–600.
Nuttgens, P., 2007, The History of York: From the earliest times to the year 2000,
Blackthorn Press, York United Kingdom.
Ortner, D.J., 2003, Identification of pathological conditions in human skeletal remains,
2nd edn, Academic Press, San Diego USA.
Ottaway, P., 2004, Roman York, Tempus, Stroud.
Pääbo, S., 1989, ‘Ancient DNA: extraction, characterization, molecular cloning, and
enzymatic amplification’, Proceedings of the National Academy of Sciences of the
United States of America, 86, 1939–1943.
Pääbo, S., Gifford, J.A. & Wilson, A.C., 1988, ‘Mitochondrial DNA sequences from a
7000-year old brain’, Nucleic Acids Research, 16(20), 9775–87.
Pai, M., Behr, M.A., Dowdy, D., Dheda, K., Divangahi, M., Boehme, C.C., Ginsberg,
A., Swaminathan, S., Spigelman, M., Getahun, H., Menzies, D. & Raviglione, M.,
2016, ‘Tuberculosis’, Nature Reviews Disease Primers, 2, 16076.
Palliser, D.M., 2014, Medieval York: 600-1540, 1st edn, Oxford University Press, New
York USA.
Pandey, R.K., Singh, D.P., Sudhakar, G., Thangaraj, K. & Rao, V.R., 2012,
‘Standardization of PCR conditions for an ancient DNA amplification’,
International Journal of Human Sciences, 9, 1.
Park, Y-H., Yoon, J-H., Shin, Y.K., Suzuki, K-I., Kudo, T., Seino, A., Kim, H-J., Lee, J-
S. & Lee, S.T., 1999, 'Classification of 'Nocardioides fulvus' IFO 14399 and
Nocardioides sp. ATCC 39419 in Kribbella gen. nov., as Kribbella flavida sp. nov.
and Kribbella sandramycini sp. nov., International Journal of Systematic
Bacteriology, 49, 743-752.
Pieters, J., 2008, ‘Mycobacterium tuberculosis and the macrophage: maintaining a
balance’, Cell Host & Microbe, 3(6), 399–407.
Poinar, H.N., Schwarz, C., Qi, J., Shapiro, B., Macphee, R.D.E., Buigues, B., Tikhonov,
A., Huson, D.H., Tomsho, L.P., Auch, A., Rampp, M., Miller, W. & Schuster, S.C.,
2006, ‘Metagenomics to paleogenomics: large-scale sequencing of mammoth
DNA’, Science, 311(5759), 392–394.
Porter, M.S. & Beiko, R.G., 2013, ‘SPANNER: taxonomic assignment of sequences
using pyramid matching of similarity profiles’, Bioinformatics, 29(15), 1858–64.
Prabhakar, M., Ershler, W.B. & Longo, D.L., 2009, ‘Bone marrow, thymus and blood:
changes across the lifespan’, Aging Health, 5(3), 385–393.

210
Pukall, R., Lapidus, A., Glavina Del Rio, T., Copeland, A., Tice, H., Cheng, J.-F., Lucas,
S., Chen, F., Nolan, M., Bruce, D., Goodwin, L., Pitluck, S., Mavromatis, K.,
Ivanova, N., Ovchinnikova, G., Pati, A., Chen, A., Palaniappan, K., Land, M.,
Hauser, L., Chang, Y.-J., Jeffries, C.D., Chain, P., Meincke, L., Sims, D., Brettin,
T., Detter, J.C., Rohde, M., Göker, M., Bristow, J., Eisen, J.A., Markowitz, V.,
Kyrpides, N.C., Klenk, H.-P. & Hugenholtz, P., 2010, ‘Complete genome sequence
of Conexibacter woesei type strain (ID131577)’, Standards in Genomic Sciences,
2(2), 212–9.
Qian, G., Wang, Y., Liu, Y., Xu, F., He, Y.-W., Du, L., Venturi, V., Fan, J., Hu, B. &
Liu, F., 2013, ‘Lysobacter enzymogenes uses two distinct cell-cell signaling
systems for differential regulation of secondary-metabolite biosynthesis and colony
morphology’, Applied and Environmental Microbiology, 79(21), 6604–16.
Quinlan, A.R. & Hall, I.M., 2010, ‘BEDTools: a flexible suite of utilities for comparing
genomic features’, Bioinformatics, 26(6), 841–842.
Rafi, A., Spigelman, M., Stanford, J., Lemma, E., Donoghue, H. & Zlas, J., 1994, ‘DNA
of Mycobacterium leprae detected by PCR in ancient bone’, The Lancet,
343(8909), 287–290.
Ramirez-Lapausa, M., Menendez-Saldana, A. & Noguerado-Asensio, A., 2015,
‘Extrapulmonary tuberculosis: an overview’, Journal of Spanish Prison Health, 17,
3–11.
Ramos, J.M., Romero, D. & Belinchón, I., 2016, ‘Epidemiology of leprosy in Spain: the
role of the international migration’, PLoS Neglected Tropical Diseases, 10(3),
e0004321.
Rasouli, M.R., Mirkoohi, M., Vaccaro, A.R., Yarandi, K.K. & Rahimi-Movaghar, V.,
2012, 'Spinal tuberculosis: diagnosis and management', Asian Spine Journal, 6(4),
294-308.
Rawnsley, S. L. & Singleton, F. B., 1995, A History of Yorkshire, 3rd edn, Phillimore,
Chichester.
Rees, R.J.W., 1966, ‘Enhanced susceptibility of thymectomized and irradiated mice to
infection with Mycobacterium leprae’, Nature, 211(5049), 657–658.
Rees, R.J.W. & McDougall, A.C., 1977, ‘Airborne infection with Mycobacterium leprae
in mice’, International Journal of Leprosy, 44(1&2), 99–103.
Reibel, F., Cambau, E. & Aubry, A., 2015, ‘Update on the epidemiology, diagnosis, and
treatment of leprosy’, Médecine et Maladies Infectieuses, 45(9), 383–393.
Ridley, D.S. & Jopling, W.H., 1966, ‘Classification of leprosy according to immunity. A
five group system’, International Journal of Leprosy, 34(3), 255–273.
Rizzi, E., Lari, M., Gigli, E., Bellis, G. De & Caramelli, D., 2012, ‘Ancient DNA
studies: new perspectives on old samples’, Genetics Selection Evolution, 44(1), 21.
Robbins, G., Tripathy, V.M., Misra, V.N., Mohanty, R.K., Shinde, V.S., Gray, K.M. &
Schug, M.D., 2009, ‘Ancient skeletal evidence for leprosy in India (2000 B.C.)’,
PLoS ONE, 4(5), e5669.
Roberts, C.A. & Buikstra, J.E., 2003, The bioarchaeology of tuberculosis : a global
perspective on a re-emerging disease, University Press of Florida, Gainesville
Florida.
Roberts, C. & Manchester, K., 2010, The archaeology of disease, 3rd edn, The History
Press, Stroud Gloucestershire.

211
Roberts, C.A., 2015, 'Old World tuberculosis: evidence from human remains with a
review of current research and future prospects', Tuberculosis, 95, S117-S121.
Roberts, R.S., 1971, The use of literary and documentary evidence in history of
medicine, in Clarke, E. (ed), Modern methods in the history of medicine, 36-56,
Athlone Press, London.
Robinson, C.F., 2003, Islamic Historiography, Cambridge University Press,Cambridge.
Rochat, T., Fujiwara-Nagata, E., Calvez, S., Dalsgaard, I., Madsen, L., Calteau, A.,
Lunazzi, A., Nicolas, P., Wiklund, T., Bernardet, J.-F. & Duchaud, E., 2017,
‘Genomic Characterization of Flavobacterium psychrophilum serotypes and
development of a multiplex PCR-based serotyping scheme', Frontiers in
Microbiology, 8, 1752.
Rohland, N. & Hofreiter, M., 2007, ‘Ancient DNA extraction from bones and teeth’,
Nature Protocols, 2(7), 1756–1762.
Rohland, N. & Reich, D., 2012, ‘Cost-effective, high-throughput DNA sequencing
libraries for multiplexed target capture’, Genome Research, 22(5), 939–946.
Rosa, L.T., Springthorpe, V., Bianconi, M.E., Thomas, G.H. & Kelly, D.J., ‘Massive
over-representation of solute-binding proteins (SBPs) from the tripartite
tricarboxylate transporter (TTT) family in the genome of the α-proteobacterium
Rhodoplanes sp. Z2-YC6860’, Microbial Genomics, 4(5), e000176.
Rosenthal, M., 2013, ‘Tuberculosis: ancient history, modern scourge’, Journal of
Ancient Diseases & Preventive Remedies, 1(2), 1–4.
Rothschild, B.M., Martin, L.D., Lev, G., Bercovier, H., Bar‐Gal, G.K., Greenblatt, C.,
Donoghue, H., Spigelman, M. & Brittain, D., 2001, ‘Mycobacterium tuberculosis
Complex DNA from and extinct bison dated 17,000 years before the present’,
Clinical Infectious Diseases, 33(3), 305–311.
Rubini, M., Erdal, Y.S., Spigelman, M., Zaio, P. & Donoghue, H.D., 2014,
‘Paleopathological and molecular study on two cases of ancient childhood leprosy
from the Roman and Byzantine empires’, International Journal of
Osteoarchaeology, 24, 570–582.
Rubini, M., Zaio, P. & Roberts, C., 2014, ‘Tuberculosis and leprosy in Italy. New
skeletal evidence’, HOMO, 65(1), 13–32.
Russell, D.G., Barry, C.E. & Flynn, J.L., 2010, ‘Tuberculosis: what we don’t know can,
and does, hurt us’, Science, 328(5980), 852–856.
Ryu, Y.J., 2015, ‘Diagnosis of pulmonary tuberculosis: recent advances and diagnostic
algorithms’, Tuberculosis and Respiratory Diseases, 78(2), 64–71.
Sabin, S., Herbig, A., Vågene, A.J., Ahlström, T., Bozovic, G., Arcini, C., Kühnert, D.
& Bos, K.I., 2020, ‘A seventeenth-century Mycobacterium tuberculosis genome
supports a Neolithic emergence of the Mycobacterium tuberculosis complex’,
Genome Biology, 21(201).
Sabin, T.D. & Ebner, J.D., 1969, ‘Patterns of sensory loss in lepromatous leprosy’,
International Journal of Leprosy, 37(3), 239–248.
Salo, W.L., Aufderheide, A.C., Buikstrat, J. & Holcomb, T.A., 1994, 'Identification of
Mycobacterium tuberculosis DNA in a pre-Columbian Peruvian mummy',
Proceedings of the National Academy of Sciences of the United States of America,
91(6),2091-2094.

212
Santamaria, M., Fosso, B., Consiglio, A., Caro, G. De, Grillo, G., Licciulli, F., Liuni, S.,
Marzano, M., Alonso-Alemany, D., Valiente, G. & Pesole, G., 2012, ‘Reference
databases for taxonomic assignment in metagenomics’, Briefings in Bioinformatics,
13(6), 682–695.
Saraf, S.K. & Tuli, S.M., 2015, 'Tuberculosis of hip: a current concept review', Indian
Journal of Orthopaedics, 49(1), 1-9.
Satapathy, J., Kar, B.R. & Job, C.K., 2005, ‘Presence of Mycobacterium leprae in
epidermal cells of lepromatous skin and its significance’, Indian Journal of
Dermatology, Venereology and Leprology, 71(4), 267–269.
Schubert, M., Lindgreen, S. & Orlando, L., 2016, ‘AdapterRemoval v2: rapid adapter
trimming, identification, and read merging’, BMC Research Notes, 9(1), 88.
Schuenemann, V.J., Avanzi, C., Krause-Kyora, B., Seitz, A., Herbig, A., Inskip, S.,
Bonazzi, M., Reiter, E., Urban, C., Dangvard Pedersen, D., Taylor, G.M., Singh, P.,
Stewart, G.R., Velemínský, P., Likovsky, J., Marcsik, A., Molnár, E., Pálfi, G.,
Mariotti, V., Riga, A., Belcastro, M.G., Boldsen, J.L., Nebel, A., Mays, S.,
Donoghue, H.D., Zakrzewski, S., Benjak, A., Nieselt, K., Cole, S.T. & Krause, J.,
2018, ‘Ancient genomes reveal a high diversity of Mycobacterium leprae in
medieval Europe’, PLoS Pathogens, 14(5), e1006997.
Schuenemann, V.J., Singh, P., Mendum, T.A., Krause-Kyora, B., Jäger, G., Bos, K.I.,
Herbig, A., Economou, C., Benjak, A., Busso, P., Nebel, A., Bolsden, J.L.,
Kjellström, A., Wu, H., Stewart, G.R., Taylor, G.M., Bauer, P., Lee, O.Y.C., Wu,
H.H.T, Minnikin, D.E., Besra, G.S., Tucker, K., Roffey, S., Sow, S.O., Cole, S.T.,
Nieselt, K. & Krause, J., 2013, 'Genome-wide comparison of Medieval and modern
Mycobacterium leprae', Science, 341, 179-183.
Sellers, M., 1987, ‘York in the Sixteenth and Seventeenth centuries’, The English
Historical Review, 12 (47) 437-447.
Sermrittirong, S. & Van Brakel, W.H., 2014, ‘Stigma in leprosy: concepts, causes and
determinants’, Leprosy Review, 85, 36–47.
Sheahan, J. J. & Whellan, T., 1855, History and topography of the city of York; The
Ainsty Wapentake; and the East Reading of Yorkshire; Embracing a general review
of the early history of Great Britain, and a general history and description of the
county of York (Volume 1), John Green Beverley, Great Britain.
Shepard, C.C., 1960, ‘The experimental disease that follows the injections of human
leprosy bacilli into foot-pads of mice’, The Journal of Experimental Medicine,
112(3), 445–54.
Shimoide, H., Anzai, E., Murata, Y., Kusajima, K., Ichihara, H., Takano, T., Hirayama,
N., Sato, N. & Kobayashi, Y., 1995, ‘Contamination of flexible fiberoptic
bronchoscopes with Mycobacterium chelonae linked to an automated endoscope
disinfection machine-on the relationship between the presence of the organism in
the intestinal tract and contamination of disinfection machine, and a case of
gallbladder and bile duct infection with M. chelonae’, Kekkaku (Tuberculosis),
70(10), 571–7.
Singh, P. & Cole, S.T., 2011, ‘Mycobacterium leprae: genes, pseudogenes and genetic
diversity’, Future Microbiology, 6(1), 57–71.
Smith, C.I., Chamberlain, A.T., Riley, M.S., Stringer, C. & Collins, M.J., 2003, ‘The
thermal history of human fossils and the likelihood of successful DNA
amplification’, Journal of Human Evolution, 45(3), 203–217.

213
Smith, S.A., Krasucki, S.P., McDowell, J. V & Balke, V.L., 2015, ‘Complete genome
sequence of Sphingobacterium sp. strain ML3W, isolated from wings of Myotis
lucifugus infected with white nose syndrome’, Genome Announcements, 3(1),
e01477-14.
Spierings, E., De Boer, T., Zulianello, L. & Ottenhoff, T.H., 2000, ‘The role of Schwann
cells, T cells and Mycobacterium leprae in the immunopathogenesis of nerve
damage in leprosy’, Leprosy Review, 71 Suppl, S121–S129.
Spigelman, M. & Donoghue, H.D., 2001, ‘Brief communication: unusual pathological
condition in the lower extremities of a skeleton from ancient Israel’, American
Journal of Physical Anthropology, 114, 92–93.
Spigelman, M. & Lemma, E., 1993, ‘The use of the polymerase chain reaction (PCR) to
detect Mycobacterium tuberculosis in ancient skeletons’, International Journal of
Osteoarchaeology, 3(2), 137–143.
Squires, K., Booth, T. & Roberts, C.A., 2019, The ethics of sampling human skeletal
remains for destructive analyses, in Squires, K., Errickson, D. & Márquez-Grant,
N. (eds), Ethical approaches to human remains. Springer, Cham.
Sreevatsan, S., Escalante, P., Pan, X., Gillies, D.A., Siddiqui, S., Khalaf, C.N.,
Kreiswirth, B.N., Bifani, P., Adams, L.G., Ficht, T., Perumaalla, V.S., Cave, M.D.,
Embden, J.D. van & Musser, J.M., 1996, ‘Identification of a polymorphic
nucleotide in oxyR specific for Mycobacterium bovis.’, Journal of Clinical
Microbiology, 34(8), 2007–10.
Sreevatsan, S., Pan, X.I., Stockbauer, K.E., Connell, N.D., Kreiswirth, B.N., Whittam,
T.S. & Musser, J.M., 1997, 'Restricted structural gene polymorphism in the
Mycobacterium tuberculosis complex indicates evolutionarily recent global
dissemination', Proceedings of the National Academy of Sciences of the United
States of America, 94, 9869-9874.
Stead, W.W., Eisenach, K.D., Cave, M.D., Beggs, M.L., Templeton, G.L., Thoen, C. &
Bates, J.H., 1995, ‘When did Mycobacterium tuberculosis infection first occur in
the New World? An important question with public health implications’, Clinical
Commentary, 151, 1267–1268.
Steyn, M & Buskes, J., 2016, 'Skeletal manifestations of tuberculosis in modern human
remains', Clinical Anatomy, 29, 854-861.
Steyn, M., Scholtz, Y., Botha, D. & Pretorius, S., 2013, 'The changing face of
tuberculosis: trends in tuberculosis-associated skeletal changes', Tuberculosis, 93,
467–474.
Stone, A.C., Wilbur, A.K., Buikstra, J.E. & Roberts, C.A, 2009, ‘Tuberculosis and
leprosy in perspective’, American Journal of Physical Anthropology, 140 Suppl,
66–94.
Strobel, T., Al-Dilaimi, A., Blom, J., Gessner, A., Kalinowski, J., Luzhetska, M., Pühler,
A., Szczepanowski, R., Bechthold, A. & Rückert, C., 2012, ‘Complete genome
sequence of Saccharothrix espanaensis DSM 44229T and comparison to the other
completely sequenced Pseudonocardiaceae’, BMC Genomics, 13(1), 465.
Stroud, G., & Kemp, R.L., 1993, ‘Cemeteries of the church and priory of St. Andrew,
Fishergate, part 2’, York Archaeological Trust.

214
Sun, H., Lapidus, A., Nolan, M., Lucas, S., Rio, T.G. Del, Tice, H., Cheng, J.-F., Tapia,
R., Han, C., Goodwin, L., Pitluck, S., Pagani, I., Ivanova, N., Mavromatis, K.,
Mikhailova, N., Pati, A., Chen, A., Palaniappan, K., Land, M., Hauser, L., Chang,
Y.-J., Jeffries, C.D., Djao, O.D.N., Rohde, M., Sikorski, J., Göker, M., Woyke, T.,
Bristow, J., Eisen, J.A., Markowitz, V., Hugenholtz, P., Kyrpides, N.C. & Klenk,
H.-P., 2010, ‘Complete genome sequence of Nocardiopsis dassonvillei type strain
(IMRU 509)’, Standards in Genomic Sciences, 3(3), 325–36.
Suzuki, K., Saso, A., Hoshino, K., Sakurai, J., Tanigawa, K., Luo, Y., Ishido, Y., Mori,
S., Hirata, K. & Ishii, N., 2014, ‘Paleopathological evidence and detection of
Mycobacterium leprae DNA from archaeological skeletal remains of Nabe-kaburi
(head-covered with iron pots) burials in Japan’, PloS ONE, 9(2), e88356.
Suzuki, K., Takigawa, W., Tanigawa, K., Nakamura, K., Ishido, Y., Kawashima, A.,
Wu, H., Akama, T., Sue, M., Yoshihara, A., Mori, S. & Ishii, N., 2010, ‘Detection
of Mycobacterium leprae DNA from achaeological skeletal remains in Japan using
whole genome amplification and polymerase chain reaction’, PLoS ONE, 5(8),
e12422.
Suzuki, T. & Inoue, T., 2007, ‘Earliest evidence of spinal tuberculosis from the
Aneolithic Yayoi period in Japan’, International Journal of Osteoarchaeology, 17,
392–402.
Tayles, N. & Buckley, H.R., 2004, ‘Leprosy and tuberculosis in Iron Age Southeast
Asia?’, American Journal of Physical Anthropology, 125, 239–256.
Taylor, G.M., Widdison, S., Brown, I.N., Young, D. & Molleson, T., 2000, ‘A
Mediaeval case of lepromatous leprosy from 13–14th century Orkney, Scotland’,
Journal of Archaeological Science, 27(12), 1133–1138.
Taylor, G.M., Young, D.B. & Mays, S.A., 2005, ‘Genotypic analysis of the earliest
known prehistoric sase of tuberculosis in Britain’, Journal of Clinical
Microbiology, 43(5), 2236–2240.
Taylor, M.G., Crossey, M., Saldanha, J. & Waldron, T., 1996, ‘DNA from
Mycobacterium tuberculosis identified in Mediaeval human skeletal remains using
polymerase chain reaction’, Journal of Archaeological Science, 23(5), 789–798.
Telenti, A., Marchesi, F., Balz, M., Bally, F., Bottger, E. & Bodmer, T., 1993, ‘Rapid
identification of Mycobacteria to the species level by polymerase chain reaction
enzyme analysis’, Journal of Clinical Microbiology, 31, 175–178.
Templeton, J.E.L., Brotherton, P.M., Llamas, B., Soubrier, J., Haak, W., Cooper, A. &
Austin, J.J., 2013, 'DNA capture and next-generation sequencing can recover whole
mitocholdrial genomes from highly degraded samples for human identification',
Investigative Genetics, 4(1), 26.
Thappa, D.M., Sharma, V.K., Kaur, S. & Suri, S., 1992, 'Radiological changes in hands
and feet in disabled leprosy patients: a clinico-radiological correlation', Indian
Journal of Leprosy, 64(1), 58-66.
Thawai, C., Kanchanasin, P., Ohkuma, M., Kudo, T. & Tanasupawat, S., 2018,
‘Identification and antimicrobial activity of Micromonospora strains from Thai peat
swamp forest soil’, Journal of Applied Pharmaceutical Science, 8(4), 119–125.
Thierry, D., Brisson-noël, A., Vincent-lévy-frébault, V., Nguyen, S., Guesdon, J.L &
Gicquel, B., 1990, ‘Characterization of a Mycobacterium tuberculosis Insertion
Sequence, IS6110, and its application in diagnosis’, Journal of Clinical
Microbiology 28(12), 2668–2673.

215
Thomson, R., Tolson, C., Carter, R., Coulter, C., Huygens, F. & Hargreaves, M., 2013,
‘Isolation of nontuberculous Mycobacteria (NTM) from household water and
shower aerosols in patients with pulmonary disease caused by NTM’, Journal of
Clinical Microbiology, 51(9), 3006–11.
Trautman, J.R., 1984, ‘A brief history of Hansen’s Disease’, Bulletin of the New York
Academy of Medicine, 60(7), 689–695.
Tripathy, S.N., 2015, Tuberculosis manual for obstetricians and gynecologists, 1st edn,
Jaypee Brothers Medical Publisher, New Delhi India.
Truman, R.W., Singh, P., Sharma, R., Busso, P., Rougemont, J., Paniz-Mondolfi, A.,
Kapopoulou, A., Brisse, S., Scollard, D.M., Gillis, T.P. & Cole, S.T., 2011,
‘Probable zoonotic leprosy in the Southern United States’, New England Journal of
Medicine, 364(17), 1626–1633.
Tsangaras, K. & Greenwood, A.D., 2012, ‘Museums and disease: using tissue archive
and museum samples to study pathogens’, Annals of Anatomy - Anatomischer
Anzeiger, 194(1), 58–73.
Turner-Walker, G., 2008, The chemical and microbial degradation of bones and teeth,
in Pinhasi, R. & Mays, S., (eds.), Advances in Human Palaeopathology, 3–29,
Wiley, Chichester.
Ukil, A.C., 1940, ‘Chronic non-tuberculous infections of the lung’, The Indian Medical
Gazette, 75(10), 581–588.
Vanhoenacker, F.M., Sanghvi, D.A. & Backer, A.I. De, 2009, ‘Imaging features of
extraaxial musculoskeletal tuberculosis’, The Indian Journal of Radiology &
Imaging, 19(3), 176–86.
van Soolingen,, Hermans, P.W., de Haas, P.E. & van Embden, J.D., 1992, ‘Insertion
element IS1081-associated restriction fragment length polymorphisms in
Mycobacterium tuberculosis complex species: a reliable tool for recognizing
Mycobacterium bovis BCG’, Journal of Clinical Microbiology, 30(7), 1772–7.
Waddington, K., 2006, The bovine scourge: meat, tuberculosis and the public’s health,
1860s-1914, The Boydell Press, Woodbridge.
Walker, S.L. & Lockwood, D.N.J., 2006, ‘The clinical and immunological features of
leprosy’, British Medical Bulletin, 77–78(1), 103–121.
Wang, H., Maeda, Y., Fukutomi, Y. & Makino, M., 2013, ‘An in vitro model of
Mycobacterium leprae induced granuloma formation.’, BMC Infectious Diseases,
13, 279.
Wang, X.-J., Zhang, B., Yan, Y.-J., An, J., Zhang, J., Liu, C.-X. & Xiang, W.-S., 2013,
‘Characterization and analysis of an industrial strain of Streptomyces
bingchenggensis by genome sequencing and gene microarray’, Genome, 56(11),
677–689.
Watson, C.L. & Lockwood, D.N.J., 2009, ‘Single nucleotide polymorphism analysis of
European archaeological M. leprae DNA’, PLoS ONE, 4(10), e7547.
Wayne, L.G. & Hayes, L.G., 1996, ‘An in vitro model for sequential study of shiftdown
of Mycobacterium tuberculosis through two stages of nonreplicating persistence’,
Infection and Immunity, 64(6), 2062–2069.

216
Weon, H.-Y., Kim, B.-Y., Yoo, S.-H., Lee, S.-Y., Kwon, S.-W., Go, S.-J. &
Stackebrandt, E., 2006, ‘Niastella koreensis gen. nov., sp. nov. and Niastella
yeongjuensis sp. nov., novel members of the phylum Bacteroidetes, isolated from
soil cultivated with Korean ginseng’, International Journal of Sytematic and
Evolutionary Microbiology, 56(8), 1777–1782.
White, C. & Franco-Paredes, C., 2015, ‘Leprosy in the 21st century’, Clinical
Microbiology Reviews, 28(1), 80–94.
Wilbur, A.K., Bouwman, A.S., Stone, A.C., Roberts, C. A., Pfister, L.-A., Buikstra, J.E.
& Brown, T.A., 2009, ‘Deficiencies and challenges in the study of ancient
tuberculosis DNA’, Journal of Archaeological Science, 36(9), 1990–1997.
Wilson, I.G., 1997, ‘Inhibition and facilitation of nucleic acid amplification’, Applied
and Environmental Microbiology, 63(10), 3741–51.
Wirth, T., Hildebrand, F., Allix-Béguec, C., Wölbeling, F., Kubica, T., Kremer, K., van
Soolingen, D., Rüsch-Gerdes, S., Locht, C., Brisse, S., Meyer, A., Supply, P. &
Niemann, S., 2008, ‘Origin, spread and demography of the Mycobacterium
tuberculosis complex’, PLoS Pathogens, 4(9), e1000160.
Witas, H.W., Donoghue, H.D., Kubiak, D., Lewandowska, M. & Gładykowska-
Rzeczycka, J.J., 2015, ‘Molecular studies on ancient M. tuberculosis and M. leprae:
methods of pathogen and host DNA analysis’, European Journal of Clinical
Microbiology & Infectious Diseases, 34(9), 1733–49.
Wood, J.W., Milner, G.R., Harpending, H.C. & Weiss, K.M., 1992, 'The osteological
paradox: problems of inferring prehistoric health from skeletal samples', Current
Anthropology, 33(4), 343-370.
Woods, S.A. & Cole, S.T., 1990, ‘A family of dispersed repeats in Mycobacterium
leprae’, Molecular Microbiology, 4(10), 1745–51.
World Health Organization, 2018a, Tuberculosis [Fact Sheets].
World Health Organization, 2018b, Leprosy [Fact Sheets].
World Health Organization, no date, Drug-resistant TB: XDR-TB FAQ.
Wrigley, E.A., Davies, R.S., Oeppen, J.E. & Schofield, R.S., 1997, English population
history from family reconstruction, 1580-1837, Cambridge University Press,
Cambridge.
Yamamura, M., Uyemura, K., Deans, R., Weinberg, K., Rea, T., Bloom, B. & Modlin,
R.L., 1991, ‘Defining protective responses to pathogens: cytokine profiles in
leprosy lesions’, Science, 254(5029), 277–279.
Yang, D.Y. & Watt, K., 2005, ‘Contamination controls when preparing archaeological
remains for ancient DNA analysis’, Journal of Archaeological Science, 32(3), 331–
336.
Yassin, A., Shennawy, M. el, Enany, G. el, Wassef, N.F. & Shoeb, S., 1975, ‘Leprosy of
the upper respiratory tract. A clinical bacteriological, histopathological and
histochemical study of twenty cases’, The Journal of Laryngology and Otology,
89(5), 505–11.
Yoon, K.H., Cho, S.N., Lee, M.K., Abalos, R.M., Cellona, R. V, Fajardo, T.T., Guido,
L.S., Cruz, E.C. Dela, Walsh, G.P., Kim, J.D. & Kim, J.D., 1993, ‘Evaluation of
polymerase chain reaction amplification of Mycobacterium leprae-specific
repetitive sequence in biopsy specimens from leprosy patients’, Journal of Clinical
Microbiology, 31(4), 895–9.

217
Zhu, H.-H., Guo, J., Yao, Q., Yang, S.-Z., Deng, M.-R., Phuong, L.T.B., Hanh, V.T. &
Ryan, M.J., 2007, ‘Streptomyces vietnamensis sp. nov., a streptomycete with violet
blue diffusible pigment isolated from soil in Vietnam’, International Journal of
Sytematic and Evolutionary Microbiology, 57(8), 1770–1774.
Zink, A., Haas, C.J., Reischl, U., Szeimies, U. & Nerlich, A.G., 2001, ‘Molecular
analysis of skeletal tuberculosis in an ancient Egyptian population’, Journal of
Medical Microbiology, 50, 355–366.
Zumla, A. & James, G., 1996, ‘Granulomatous infections: etiology and classification’,
Clinical Infetious Diseases, 23, 146–158.

218
Appendices
Site Name Skeleton Number Sampled
Age Sex
Melton 4297 36-45 M
2554 11-13 yrs
J
5319 36-45 M
St Peter's Huddersfield
5 46+ M
7 46+ M
17 18-25 F
St Andrew Fishergate
6 20-30 M
277 5 yrs J
286 5-6 yrs J
296 18-25 M
323 16-18 M
339 30-40 M
384 4-6 yrs J
34 adult F?
131 46+ M
253 30-40 F
3 Driffield Terrace 37 36-45 M
54 26-35 M
13 16-18 M?
15 26-35 M
6 Driffield Terrace 19 26-35 M
22 36-45 M?
St Helen-on-the-Walls
5000 adult I
5844 26-35 F
6003 36-45 F
5494 46+ F
Heslington East 229 26-35 M
Wharram Percy 26 46+ F
1600 adult I
Sewerby G34 46+ M
G44 N/A M
Addingham 134 26-35 M
223 14-16 yrs
J
103 26-35 M
St Giles by Brompton Bridge
1288 26-35 M?
1531 18-25 F?
1542 26-35 M?
Wetwang Slack 1 14 yrs J
2 36-45 M
5 26-35 M
6 26-35 M
8 18-25 F
9 25-30 M

219
3 18-25 M
4 18-25 F
7 26-35 F
Hickleton 46 N/A N/A
York Minster 1 18-25 M
15 36-45 M
Ailcy Hill 1044 N/A J
1043 46+ M
Fishergate House C1163 (86) 46+ M
C1188 (98) 36-45 M
C1205 (108) 12-14 yrs
N/A
C1259 (135) 26-35 F
C1282 (147) 26-35 M
C1286 (149) 36-45 F
Table S2.1: Additional skeleton information for the 57 skeleton studied. This table is
showing the age and sex information for the Yorkshire archaeological bones studied (except for
the three samples from Wetwang Slack settlement as this information is not available). M: male,
F: female, N/A: information not available, J: juvenile, I: ambiguous.

220
M A B
200 bp
100 bp
Figure S3.1: Rv0083 PCR assay amplification for sample St Andrew Fishergate 277. A:
undiluted DNA template, B: 10-fold diluted DNA template. M is the DNA ladder with the 100
bp and 200 bp marked in the figure. Faint lower band is shown inside the red box.
Figure S3.2: gyrA PCR assay amplification for sample St Andrew Fishergate 339. A:
undiluted DNA template, B: 10-fold diluted DNA template. M is the DNA ladder with the 100
bp and 200 bp marked in the figure. Faint lower band is shown inside the red box.
M A B
200 bp
100 bp

221
M A B
200 bp
100 bp
200 bp
100 bp
A B M
Figure S3.3: IS6110 123 bp PCR amplification of the sample Heslington East 229. A:
undiluted DNA template, B: 10-1 diluted DNA template. M is showing the DNA ladder with the
100 bp and 200 bp marked in the figure. Faint lower band is shown inside the red box.
Figure S3.4: gyrA PCR amplification of the sample Wetwang Slack 2. A: undiluted DNA
template, B: 10-1 diluted DNA template. M is showing the DNA ladder with the 100 bp and 200
bp marked in the figure. Faint lower band is shown inside the red box.

222
200 bp
100 bp
A B M
M A B C D
100 bp
200 bp
Figure S3.5: IS6110-123 bp PCR amplification of the sample Wetwang Slack 7. A:
undiluted DNA template, B: 10-1 diluted DNA template. M is showing the DNA ladder with the
100 bp and 200 bp marked in the figure.
Figure S3.6: gyrA PCR amplification of the samples Wharram Percy 26 and Wharram
Percy 1600. A and C: undiluted and 10-1 diluted DNA templates of Wharram Percy 26,
respectively; B and D: undiluted and 10-1 diluted DNA template of Wharram Percy1600,
respectively. M is showing the DNA ladder with the 100 bp and 200 bp marked in the figure.

223
M A B C D
200 bp
100 bp
M A B
200 bp
100 bp
Figure S3.7: IS6110 first step PCR amplification of the samples Addingham 134 and
Addingham 223. A and C: undiluted DNA and 10-1 diluted DNA templates of Addingham 134,
respectively; B and D: undiluted DNA and 10-1 diluted DNA templates of Addingham 223,
respectively. M is showing the DNA ladder with the 100 bp and 200 bp marked in the figure.
The bands of interest are shown in the red boxes.
Figure S3.8: gyrA PCR amplification of the sample Addingham 103. A: undiluted DNA
template, B: 10-1 diluted DNA template. M is showing the DNA ladder with the 100 bp and 200
bp are marked in the figure.

224
M A B
200 bp
100 bp
M A B
200 bp
100 bp
Figure S3.9: gyrA PCR amplification of the sample Melton 5319 sample. A: undiluted DNA
template, B: 10-1 diluted DNA template. M is showing the DNA ladder with the 100 bp and 200
bp bands indicated in the figure.
Figure S3.10: gyrA PCR amplification of the sample Hickleton 46. A: undiluted DNA
template, B: 10-1 diluted DNA template. M is showing the DNA ladder with 100 bp and 200 bp
bands indicated in the figure.

225
M A B
200 bp
100 bp
Figure S3.11: IS6110 123 bp PCR amplification of the sample St Giles by Brompton Bridge
1542. A: undiluted DNA template, B: 10-1 diluted DNA template. M is showing the DNA ladder
with 100 bp and 200 bp bands indicated in the figure. The desired band is marked inside the red
box.