Embed Size (px)
Citation preview

Gibberellic Acid-Induced Pollen Mortality and Abnormal
Microsporogenesis in Safflower
Girjesh Kumar and Preeti Srivastava*
Department of Botany, Plant Genetics Laboratory, University of Allahabad, Allahabad 211002, India
Received May 12, 2009; accepted July 6, 2009
Summary The present research work was undertaken to assess the effect of gibberellic acid (GA3)applied exogenously to the Safflower plant for the study of pollen behavior. A cytological characteri-zation in safflower from meiosis to pollen formation revealed that whereas meiosis was recorded tobe normal in control plants with normal microsporogenesis, GA3-treated plants possessed disturbedtetrad configurations of pollen. GA3 caused deductions in pollen yield and increased the formation ofabnormal meiotic products. Our study elucidates that the microspore sacs, instead of having normalfour microspores, possessed 1 (monad), 2 (dyads), 3 (triads) and even up to 5 (polyads) microspores,which may produce unreduced gametes at later stages. A significant variation in the degree of pollenmortality was also displayed amongst these pollens. Another interesting feature noticed during thestudy was the fusion among tetrads due to wall dissolution.
Key words Gibberellic acid (GA3), Microsporogenesis, Pollen sterility, Polyad, Safflower.
Safflower is one of the most important oil crops in the world due to the favorable fatty acidcomposition of the oil with regard to human consumption. The seeds contain 35–50% oil, 15–20%protein and 35–45% hull fraction (Rahamatalla et al. 2001). The oil is high in linolenic acid, an un-saturated fatty acid that aids the lowering of blood cholesterol levels (Cosge et al. 2007). Besidesthis, the plant has gained significant interest amongst scientists as a drought-tolerant crop, which iscapable of obtaining moisture from levels not available to the majority of crops (Weiss 2000). Sincenot much work on the cytology of the plant has been done so far, due to the poor staining capabilityand stickiness of chromosomes, the present work enumerates the influence of gibberellic acid onthe reproduction capacity of the plant via the study of pollen grains, as the pollen sterility system isuseful in hybrid breeding and obtaining pollen sterile female plants (Baydar 2002). It is well knownthat every sample of angiosperm pollen may contain several percent of dead grains, although thereasons of pollen mortality are insufficiently studied (Drake et al. 1998). Safflower plants treatedwith exogenous GA3 applications to the buds before flowering produced no or insufficient pollenbecause of inhibition of microsporogenesis (Baydar 2000, 2001) as can be evidenced from ourstudy also. For economical and practical hybrid seed production in safflower, attempts have beenmade to benefit from genetic or synthetic pollen sterility (Heaton and Knowles 1982, Baydar 2001).Thus the present study is an attempt to analyse the effect of GA3 treatment on the pollen sterility ofsafflower.
Materials and methods
Procurement of seeds and chemicalsSeeds of safflower (Carthamus tinctorius L.) were obtained from the National Bureau of Plant
Genetic Resources, New Delhi (India) and gibberellic acid was obtained from Merck, India.
* Corresponding author, e-mail: [email protected]
© 2009 The Japan Mendel Society Cytologia 74(2): 171–176, 2009
Gibberellic acid treatmentHealthy and uniform-sized seeds of safflower were selected and presoaked in distilled water
for 2 h. These seeds were then transferred in their respective pots for germination. On the onset ofbudding, the plants were sprayed with 100 ppm, 200 ppm, and 300 ppm concentrations of GA3 3times at a regular interval of 7 days. Untreated plants were considered as control. Cytogeneticalanalysis was carried out using 2% acetocarmine. A detailed study of pollen was also carried to eval-uate the pollen yield and deviations from the normal course in pollen arrangement. Photomicro-graphs were taken using a Nikon image capturing system.
Results
Cytogenetical evaluationThe cytogenetical study of the plant suggests that the chromosome pairing is normal at diaki-
nesis with 12 bivalents. Data regarding the induction of meiotic abnormalities at metaphase andanaphase after GA3 treatment with successive sprays has been presented in Table 1. After the treat-ment there is an increment in chromosomal aberrations at various stages, although the frequency ofdifferent anomaly varied. Data given reveals that the frequency of metaphasic abnormalities wasmore than that of anaphasic anomalies.
Pollen studyMicrosporogenesis was not affected in the control plants but plants treated with GA3
possessed highly disturbed microsporogenesis and the formation of a high percentage of abnormalmicrospores was observed. Results obtained during the present investigation elaborate that GA3
could prove to be an effective chemical in upbringing pollen mortality and inducing successfulpollen sterility. Spraying with 100 ppm did induce sterility, although not to the amount observedwith 200 and 300 ppm concentration treatment. 200 ppm with 2 sprays produced 50% pollen sterili-ty and 300 ppm dose proved to be very effective in inducing pollen sterility (Table 1). Significantparts of the study were the variation in microspore arrangement/types, the degree of existence ofdead pollen grains in microspores, and the rare occurrence of cytomixis (via cytoplasmic channels,cell fusion and syncyte formation) as presented in Table 2 and picturized in Plates 1, 2.
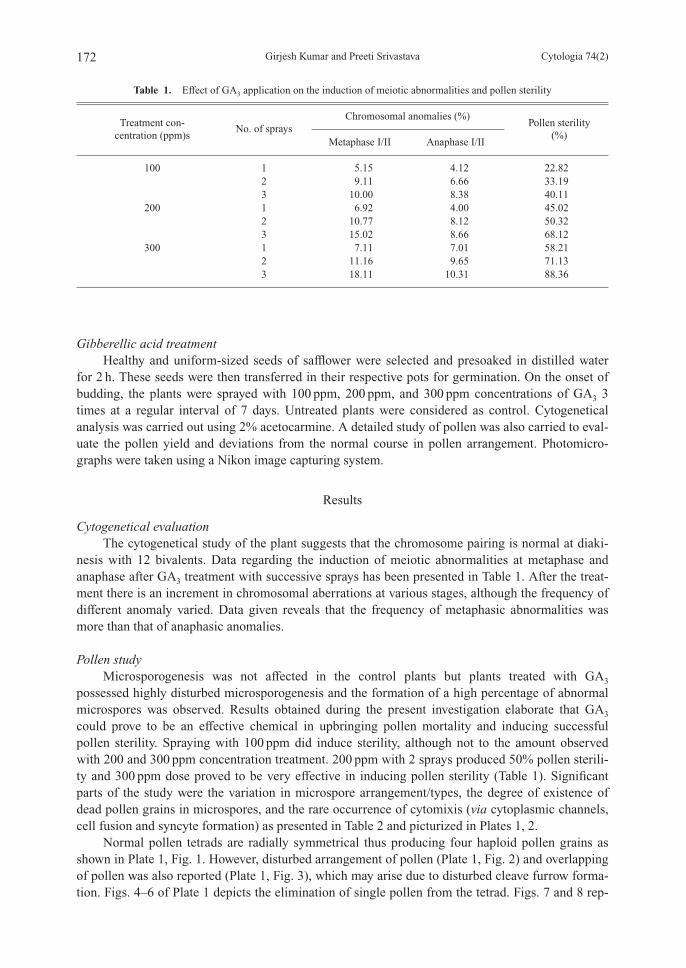
Normal pollen tetrads are radially symmetrical thus producing four haploid pollen grains asshown in Plate 1, Fig. 1. However, disturbed arrangement of pollen (Plate 1, Fig. 2) and overlappingof pollen was also reported (Plate 1, Fig. 3), which may arise due to disturbed cleave furrow forma-tion. Figs. 4–6 of Plate 1 depicts the elimination of single pollen from the tetrad. Figs. 7 and 8 rep-
172 Cytologia 74(2)Girjesh Kumar and Preeti Srivastava
Table 1. Effect of GA3 application on the induction of meiotic abnormalities and pollen sterility
Treatment con-Chromosomal anomalies (%)
Pollen sterilityNo. of sprays
centration (ppm)sMetaphase I/II Anaphase I/II
(%)
100 1 5.15 4.12 22.822 9.11 6.66 33.193 10.00 8.38 40.11
200 1 6.92 4.00 45.022 10.77 8.12 50.323 15.02 8.66 68.12
300 1 7.11 7.01 58.212 11.16 9.65 71.133 18.11 10.31 88.36
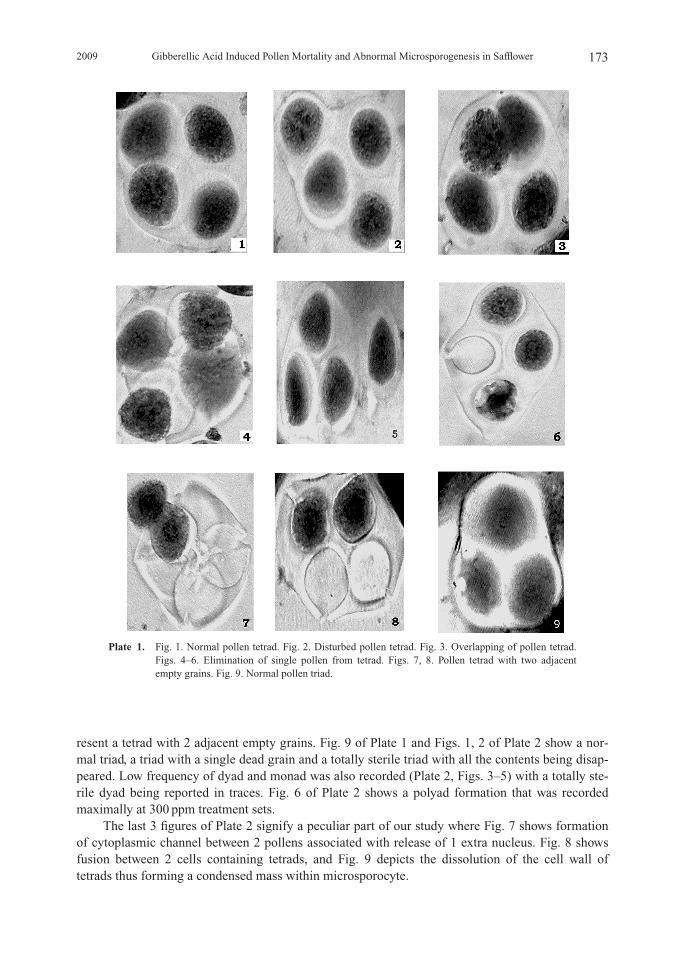
resent a tetrad with 2 adjacent empty grains. Fig. 9 of Plate 1 and Figs. 1, 2 of Plate 2 show a nor-mal triad, a triad with a single dead grain and a totally sterile triad with all the contents being disap-peared. Low frequency of dyad and monad was also recorded (Plate 2, Figs. 3–5) with a totally ste-rile dyad being reported in traces. Fig. 6 of Plate 2 shows a polyad formation that was recordedmaximally at 300 ppm treatment sets.
The last 3 figures of Plate 2 signify a peculiar part of our study where Fig. 7 shows formationof cytoplasmic channel between 2 pollens associated with release of 1 extra nucleus. Fig. 8 showsfusion between 2 cells containing tetrads, and Fig. 9 depicts the dissolution of the cell wall oftetrads thus forming a condensed mass within microsporocyte.
2009 173Gibberellic Acid Induced Pollen Mortality and Abnormal Microsporogenesis in Safflower
Plate 1. Fig. 1. Normal pollen tetrad. Fig. 2. Disturbed pollen tetrad. Fig. 3. Overlapping of pollen tetrad.Figs. 4–6. Elimination of single pollen from tetrad. Figs. 7, 8. Pollen tetrad with two adjacentempty grains. Fig. 9. Normal pollen triad.
Discussion
Compound pollen grains in the form of dyad, triad, tetrad, polyad, or pollinium occur in morethan 56 families of angiosperms (Knox and McConchie 1986). These studies may be correlated toours where abnormal microspore formations in the form of monad, dyad, triads, and abnormal arrangement of tetrads and polyads were obtained.
The tetrad is the product of the particular spatial orientation of the spindle with respect to eachother during anaphase II. At the end of telophase II a cleavage furrow is formed between the 4 mei-otic products and a tetrad is formed (Becerra et al. 2002). Variation from normal tetrad arrangementhas been reported in many plants, which may arise due to disturbances in spindle behavior to divide
174 Cytologia 74(2)Girjesh Kumar and Preeti Srivastava
Plate 2. Fig. 1. Triad with a single dead grain. Fig. 2. Completely sterile triad. Fig. 3. A dyad. Fig. 4. Asterile dyad. Fig. 5. A monad. Fig. 6 polyad. Fig. 7 polyad with elimination of 1 nucleus and cyto-plasmic channel formation among 2 microspores. Fig. 8. Fusion between 2 cells containing tetrads.Fig. 9. Dissolution of cell wall of tetrads thus forming a condensed mass within microsporocyte(Syncyte).
the tetrads normally and thus results in the formation of abnormal meiotic products. The deviationfrom normal tetrad arrangement is caused by parallel and fused spindles that form dyads and that oftriads through the formation of tripolar spindle (Becerra et al. 2002, Singhal et al. 2008). In thetetrad stages of meiosis a monad, dyads and triads were reported in Sweet Potato by Becerra et al.(2002) which can be correlated to our study where such deviations were also noticed. The forma-tion of dyads and triads was also reported in Paspalum (Pagliarini et al. 1999) was due to absenceof cytokinesis at the end of meiosis I. However formation of polyads occurs when absence of spin-dle in second meiosis at metaphase and anaphase were not observed; instead II cytokinesis occursin prophase II cells after cytokinesis I and the final product of meiosis was completely abnormalwith a predominance of polyads with microspores having abortive pollen grains (Pagliarini 2000).Such a report validates the observations found in our study. During our investigation some pollenwith dead grains were also observed at various stages, such as at tetrad, triad and dyad. Such resultswere also obtained by Berdinikov et al. (2002) in Typha latifolia L. where they reported pollenclumps with empty tetrads where all anther contents perished for some or other reason. They re-ported tetrad with 2 dead pollen grains which may result mostly from non-disjunctionin anaphase I,and those with 1 pollen grain from non-disjunction at anaphase II, thus making tetrad analysis inef-fective for estimating the rate of gametophytic lethals (Berdinikov et al. 2002). Copenhaver et al.(2000) discovered that a microsporocyte heterozygous for a recessive gametophytic lethal shouldresult in a tetrad with 2 dead grains as found in our study (Figs. 7, 8 of Plate 1). Occurrence ofmonad and total elimination of 1 genome may result in the production of haploids (Becerra et al.2002). Interesting feature of the study was the formation of cytoplasmic channels between pollen,which may be manifested as an effect of chemical used on the concerned plant. Cell fusion between2 tetrads might be a probable cause of male sterility in the plant. Fusion of PMCs was also reportedby Mehra and Kalia (1973) and Mendes- Bonato et al. (2001) where they interpreted such fusion tobe the originator of syncyte formation, which further gave rise to abnormal sized pollen grains. Thecause of syncyte formation may be diverse and can be presumed to be due to chemicals such as X-rays, temperature, culture conditions (Stern 1946, Pantulu and Manga 1971, Peeters t al. 1985) andgenetic factors (Pantulu and Manga 1971, Rao and Koduru 1978, Rao et al. 1991, Nirmala and Rao1996). Nirmala and Rao (1996) even reported that these factors are a major cause of male sterility.Thus our study strongly suggests that GA3 is a potent chemical for the induction of male sterility orpollen sterility in safflower, which was in concordance with the studies conducted by Shuster(1969), and Seetharam and Kusumkumari (1974, 1975) on sunflowers by spraying GA3 at bud initi-ation stage.
2009 175Gibberellic Acid Induced Pollen Mortality and Abnormal Microsporogenesis in Safflower
Table 2. Effect of GA3 on microsporogenesis of safflower
TreatmentMicrosporogenesis anomalies Cytomixis
No. of concentration
sprays Normal Disturbed Triad Diad Monad Polyad
Channel Cell Syncyte (ppm)s
tetrad tetrad formation fusion formation
100 1 88.00 3.32 2.18 — 0.11 — 0.33 0.11 —2 83.13 3.81 4.12 0.85 — — 0.20 0.30 —3 72.22 5.57 4.00 1.38 — 0.36 0.62 0.52 —
200 1 79.11 5.01 5.05 1.22 — 1.98 — 0.54 —2 75.00 5.36 5.72 2.00 0.22 2.45 1.16 0.35 0.883 68.82 7.35 7.50 2.16 — 4.16 0.88 0.82 —
300 1 55.12 7.36 6.00 — 1.11 6.01 0.65 0.12 0.932 48.21 8.00 7.16 1.98 — 6.18 — 0.42 1.133 38.18 8.12 7.85 3.11 2.00 6.35 1.02 1.77 2.12

References
Baydar, H. 2000. Gibberellik asidin aspir (Cathamus tinctorius L.) de erkek kisirlik, tohum verimi ile yag ve yag asetlerisentezi uzerine etkisi. Turk. J. Biol. 24: 159–168.
— 2001. Gibberellik asit ile aspir (Carthamus tinctorius L.) de pollen ksirliginin uyarilmasi. Turkiye IV. Tarla BitkeleriKongresi, 17–21 Eylul. Terkidag. pp. 61–65.
— 2002. Effects of Gibberellic acid treatment for pollen sterility induction on physiological activity and endogenous hormone levels of seeds in Safflower. Turk. J. Biol. 26: 235–239.
Becerra Lopez-Lavelle, L. A. and Orjeda, G. 2002. Occurrence and Cytological Mechanism of 2n pollen formation inTetraploid Accession of Ipomoea batatas (Sweet Potato). The American Genetic Association 93: 185–192.
Berdnilkov, V. A., Kosterin, O. E. and Bogdanova, A. 2002. Mortality of pollen grains may result from the errors of meiosis:study of pollen tetrads in Typha latifolia L. Heredity 89: 358–362.
Copenhaver, G. P., Keith, K. C. and Reuss, D. 2000. Tetrad analysis in higher plants. A budding technology. Plant Physiol.124: 7–16.
Cosge, B., Gurbuz, A. and Kirlan, M. 2007. Oil content and fatty acid composition in some safflower (Carthamus tinctoriusL.) varieties sown in spring and winter. Int. J. Eng. and Nat. Sci. 1: 11–15.
Drake, J. W., Charlesworth, D. and Crow, J. F. 1998. Rates of spontaneous mutation. Genetics 148: 1667–1686.Heaton, T. C. and Knowles, P. F. 1982. Inheritance of male sterility in Safflower. Crop Science 22: 520–522.Knox, R. B. C. and McConchie, A. 1986. Structure and function of compound pollen. In S. Blackmore and I. K. Ferguson
[eds.], Pollen and spores, form and function, Academic Press, London. pp. 264–282.Mehra, P. N. and Kalia, V. 1973. Accessory chromosomes and multinucleate pollen mother cells in Saccharum benghalense
Retz. complex. Nucleus 16: 75–78.Mendes-Bonato, A. B., Pagliarini, M. S., Silva, N. and Valle, C. B. 2001. Meiotic instability in invaders plants of signal
grass Brachiaria decumbens Stapf. (Gramineae). Genet. Mol. Biol. 23: 619–625.Nirmala, A. and Rao, P. N. 1996. Genesis of chromosomal numerical mosaism in higher plants. Nucleus 39: 151–175.Peeters, J.P., Griffiths, J.F. and Wilkes, G. 1985. In vivo karyotypic modifications following spontaneous cell fusion in maize
(Zea mays L.). Can. J. Genet. Cytol. 27: 580-585.Pagliarini, M. S., Freitas, P. M., Carraro, L. R., Adamowski, E. V. and Batista L. A. R. 1999. Failure of cytokinesis and 2n
gamete formation in Brazilian accessions of Paspalum. Euphytica 108: 129–135.—, — and Batista, L. A. R. 2000. Chromosome stickiness in meiosis of a Brazilian Paspalum accessions. Cytologia 65:
288–294.Pantulu, J.V. and Manga, V. 1971. Monofactorial “multiploid sporocytes” condition induced by EMS in pearl millet.
Genetica.42: 214-218.Rahamatalla, A. B., Babiker, E. E., Krishna, A. G. and EI Tinay, A. H. 2001. Changes in fatty acid composition of safflower
(Carthamus tinctorius L.) cultivars grown in the Mediterranean region of Turkey. J. Agron. Crop. Sci. 189:359–360.
Rao, M. K. and Koduru, P. R. K. 1978. Cytogenetics as a factor for syncyte formation and male sterility in Pennisetumamericanum. Theor. Appl. Genet. 3: 1–7.
—, Nirmala, A. and Ranganadham P. 1991. Cytogenetical studies of plasmodial sporocytes, chromatin disintegration, desynapsis and centromere breakage in pearl millet. Cytologia 56: 165–171.
Seetharam, A. and Kusuma Kumari, P. 1974. Gibberellic acid induced male sterility in sunflower. Sci. Cul. 40: 398–399.— and — 1975. Induction of male sterility in sunflower. Indian J. Genet. 35: 136–138.Shuster, W. 1969. Beobachtungen uber mannlicne sterilliat beidax conrenblume (Helianthus annus L.) ansgeolost durch
genetische. Physiologissche and induszierte chemische factoren. Theor. Appl. Genet. 39: 261–273.Singhal, V. K., Kaur, D. and Kumar, P. 2008. Effect of cytomixis on the pollen size in Seabuckthorn (Hippophae rhamnoides
L., Elaeagnaceae). Cytologia 73: 167–172.— and Kumar, P. 2008. Impact of cytomixis on meiosis, pollen viability and pollen size in wild populations of Himalayan
Poppy (Meconopsis aculeate Royle). J. Biosci. 33: 371–380.Stern, H. 1946. The formation of polynucleated pollen mother cells. Heredity 37: 46–50.Weiss, E. A. 2000. Oilseed crops (second edition). Victoria-Australia. Blackwell Science Ltd., Oxford. p. 364.
176 Cytologia 74(2)Girjesh Kumar and Preeti Srivastava





























