Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY,0021-9193/98/$04.0010
Nov. 1998, p. 5601–5611 Vol. 180, No. 21
Copyright © 1998, American Society for Microbiology. All Rights Reserved.
Gigantism in a Bacterium, Epulopiscium fishelsoni, Correlateswith Complex Patterns in Arrangement, Quantity,
and Segregation of DNAV. BRESLER,1 W. L. MONTGOMERY,2* L. FISHELSON,3 AND P. E. POLLAK2
Institute for Nature Conservation Research1 and Department of Zoology,3 G. Wise Faculty of Life Sciences,Tel Aviv University, Ramat Aviv, 69978 Tel Aviv, Israel, and Department of Biological Sciences,
Northern Arizona University, Flagstaff, Arizona 86011-56402
Received 25 February 1998/Accepted 1 September 1998
Epulopiscium fishelsoni, gut symbiont of the brown surgeonfish (Acanthurus nigrofuscus) in the Red Sea,attains a larger size than any other eubacterium, varies 10- to 20-fold in length (and >2,000-fold in volume),and undergoes a complex daily life cycle. In early morning, nucleoids contain highly condensed DNA inelongate, chromosome-like structures which are physically separated from the general cytoplasm. Cell divisioninvolves production of two (rarely three) nucleoids within a cell, deposition of cell walls around expandednucleoids, and emergence of daughter cells from the parent cell. Fluorescence measurements of DNA, RNA,and other cell components indicate the following. DNA quantity is proportional to cell volume over cell lengthsof ;30 mm to >500 mm. For cells of a given size, nucleoids of cells with two nucleoids (binucleoid) containapproximately equal amounts of DNA. And each nucleoid of a binucleoid cell contains one-half the DNA of thesingle nucleoid in a uninucleoid cell of the same size. The life cycle involves approximately equal subdivisionof DNA among daughter cells, formation of apical caps of condensed DNA from previously decondensed anddiffusely distributed DNA, and “pinching” of DNA near the middle of the cell in the absence of new wallformation. Mechanisms underlying these patterns remain unclear, but formation of daughter nucleoids andcells occurs both during diurnal periods of host feeding and bacterial cell growth and during nocturnal periodsof host inactivity when mean bacterial cell size declines.
In 1985, an unusual unicellular organism was reported fromthe gut of brown surgeonfish, Acanthurus nigrofuscus (Acan-thuridae: Teleostei), in the Red Sea (14). These organismswere named Epulopiscium fishelsoni and tentatively placed inthe kingdom Protoctista by Montgomery and Pollak (23) on thebasis of their maximum size (.600 mm), great variation inlength (;30 to .600 mm) and volume (.2,000-fold) bothwithin single hosts and during a daily cycle, daily variability innucleoid and cytoplasm structure, and a peculiar mode ofreproduction (daughter cells form within parental organismsand eventually emerge as mobile cells from the maternal en-velope). Similar and apparently related organisms representingat least 10 different morphotypes were later collected fromsurgeonfishes from the Hawaiian Islands, French Polynesia,Tuvalu, Guam, southern Japan, Papua New Guinea, the GreatBarrier Reef, and South Africa (8, 13a, 15a, 22, 24a). None ofthese organisms (here termed “epulos” to distinguish themfrom other groups of bacteria) has been cultured, so all de-scriptions of distribution or functions within hosts and descrip-tions of diel changes in size or stages in the life cycle are basedon samples from populations of epulos collected from hostssacrificed at different times of day and night.
Initial electron microscopy of epulos revealed a complexultrastructure lacking standard eukaryotic organelles (8, 14,23). Following suggestions by Clements and Bullivant (9) thatthe fine structure of cytoplasm, cell wall, and flagella indicatedthat epulos were in fact prokaryotic, Angert et al. (2) isolatedand sequenced the gene encoding the 16S rRNA subunit, plac-
ing these giant microorganisms in a group of low-G1C gram-positive bacteria related to Clostridium. In situ hybridizationwith fluorescein-labelled oligonucleotide probes based oncloned rRNA sequences confirmed the source of the rRNAgene. E. fishelsoni represents, therefore, the largest bacteriumso far described (2, 9).
Cell size also varies more than in other bacteria. If oneestimates the volume of a cigar-shaped E. fishelsoni cell asroughly the volume of two cones, each with height and radiusof the base equal to half of cell length and half of its maximaldiameter, respectively (V 5 2/3pr2h), the volume of a verylarge E. fishelsoni (;354,000 mm3 [500 by 52 mm]) is approx-imately 3,000-fold greater than that of a very small E. fishelsoni(;125.6 mm3 [30 by 4 mm]). The volume of a large epulo can,therefore, exceed the volume of a bacterium such as Esche-richia coli (;2 mm3 [1 by 2 mm]) (25) by at least 5 orders ofmagnitude.
Cellular or molecular mechanisms that may support andcontrol such exceptional variability in dimension, volume andsurface/volume ratios of E. fishelsoni are unclear. However,cells are highly mobile, vary in mean size and structure duringa 24-h period, affect the pH of the host’s gut fluids differentiallyduring day and night (suggesting metabolic changes on a dielcycle), and construct mobile daughter cells within the parentalcell (9, 14, 23, 24). The active metabolism and correspondingexpectation of genomic activity supporting synthesis of macro-molecules led us to predict correlations between DNA quantityor condensation state and cell size or stage in the daily lifecycle.
Because E. fishelsoni and related organisms have not beencultured, we relied on collections of cells from host fishessacrificed at different times of day and night, fluorescencecytochemistry, microfluorometry, and transmission electron
* Corresponding author. Mailing address: Department of BiologicalSciences, Northern Arizona University, Flagstaff, AZ 86011-5640.Phone: (520) 523-7505. Fax: (520) 523-7500. E-mail: [email protected].
5601
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
211
.223
.74.
164.

microscopy to describe changes in the functional state anddistribution of DNA and other cell constituents during themicrobe’s life cycle. Our primary objective was to study rela-tionships between DNA quantity and cell volume, as well aspossible changes in DNA distribution and functional activity ofthe nucleoid during the life cycle of E. fishelsoni. In this paper,we initially demonstrate that DNA quantity is directly propor-tional to cell volume rather than to length or surface area, andthat large quantities of DNA are almost equally divided anddistributed to developing daughter cells. We then describe howcoordinated changes in DNA condensation state and distribu-tion, nucleoid expansion, and cell wall deposition lead to theformation of daughter cells.
MATERIALS AND METHODS
Specimens of the host surgeonfish, A. nigrofuscus, were collected by net orhand spear near the Interuniversity Institute, Steinitz Marine Biological Labo-ratory, Eilat, Red Sea, Israel (see reference 21 for details of the site and thecollection techniques). Because surgeonfish are active only during the day, hostfish intended for sampling at night were held in flowing seawater prior tosacrifice; samples obtained during the day were taken shortly after fish werecollected. Epulos have not been cultured, so each fish provided a single sampleof an epulo population at the time of sacrifice.
Specimens were sacrificed by a blow to the head and immediately dissected. E.fishelsoni cells were collected from the central intestine of the fish within ;15min of host sacrifice. Cells were either examined live at the marine laboratory orwere fixed (in absolute methanol for fluorometry and in other fixatives, describedbelow, for light and electron microscopy) at various times of day and night forsubsequent analysis at our home institutions.
Lengths of epulos varied within a single host fish, and epulo size distributionchanged with time of day or night. In the field, we used an ocular grid in a Zeissbinocular microscope at 3100 to assign live cells collected at different times to;50-mm-interval length categories (,50 mm, 50 to 101 mm, and so on [see Table1]). Sixty haphazardly chosen cells from each of two fish were measured per timeperiod (total n 5 120 per period). Because we were unable to measure rapidlymoving cells precisely with the grid available to us at our field laboratory, wecreated a frequency distribution among categories and calculated an approxi-mate mean length for each time period, assuming that all cells in a particularcategory were the median length for that category (see Table 1).
Preserved cells intended for fluorometry (see below) were returned to Tel AvivUniversity. There, cells falling into several narrow size ranges (5 to 10 mm forcells ,250 mm long, 20 to 30 mm for cells .300 mm long [see Table 2]) wereselected from samples. Samples representing different collection times weretreated independently. Initially, six size categories (groups I through VI of Table2) were established, such that cells in each of groups II through VI had volumesapproximately twice those of cells in the preceding smaller group (see Table 2).A seventh group (group 0) was later added to expand the size of range ofsurveyed cells and included specimens of the smallest easily assayed E. fishelsoni(;30 mm long). Cells with lengths between ;30 and ;150 mm were excluded dueto limitations in equipment available to make precise measurements.
Because epulos collected at different times of day and night show markedvariability in the distribution and condensation state of DNA and in the size andlocation of nucleoids, initial measurements of total DNA in nucleoids were madeon cells from a sample taken at approximately 0800 h. Nucleoids in this materialwere compact, round in shape, and located at the poles of the cell. We studied60 cells from each group. Surveys of a variety of cells sampled at different timesof day or night were then made in order to describe correlations in state ordistribution of DNA and other cell components with time or stage in the epulolife cycle. We used fluorescence cytochemistry, microfluorometry, and digitalanalysis of fluorescently stained cells, the most sensitive methods to measureamount and state of DNA and RNA in single prokaryotic and eukaryotic cells (3,4, 10–13, 32, 33).
Fluorescence cytochemistry had never been applied to E. fishelsoni, so we usedtwo classical fluorescence methods to estimate relative amounts of DNA in cells:the Fuelgen reaction with acridine yellow-Schiff-type reagent (Sigma), and ber-berine sulfate (Sigma) staining after ribonuclease treatment (3, 4, 10). Schiffreagent reacts with aldehyde groups of free deoxyribose nucleotides liberated byacid hydrolysis. Because the intensity of the Fuelgen reaction depends on tem-perature and time of hydrolysis, we standardized our preparation by hydrolyzingwith 6 N HCl for 10 min at room temperature before treating with Schiff reagentfor 30 min. After washing (in 1 N HCl–10% NaHSO3–distilled water [1/1/18 byvolume] followed by distilled water), stained bacteria were mounted in nonfluo-rescent glycerine for measurements. In the second technique, we stained with a0.01% solution of berberine sulfate in ethanol for 20 min after pretreatment withribonuclease; berberine sulfate is an intercalating agent used without the hydro-lysis required by the Fuelgen technique. A conventional fluorescence objective(10 by 0.40) and measuring diaphragms corresponding to the structure beingmeasured (whole cell or nucleoid only) were used for measuring relative
amounts of DNA (Fuelgen and berberine procedures); units were arbitrary(microamperes of current). Results from these two techniques (see Table 2)validate their application in this system.
We used both morphological and cytochemical criteria following staining withacridine orange to map the locations of different cell components during differ-ent stages in the cell and life cycles. Acridine orange has been applied widely asa bacterial stain to detect and characterize soil or sediment bacteria (15, 16, 27,31), and the metachromic red or green fluorescence of acridine orange has beenused to assess viability and physiological activity of both bacteria and bacterialspores (7, 18–20, 28).
Cells stained with acridine orange were mapped according to the classes ofmolecules distinguished (those with condensed DNA or decondensed DNA,RNA enriched, general cytoplasm, cell wall [see Fig. 3 through 6]). The mor-phology of E. fishelsoni is well known from both light and electron microscopestudies (9, 14, 23) (see Fig. 1 and 2), and we scanned cells stained with acridineorange under permanent visual control. We could, therefore, distinguish fluo-rescent signals from wall, cytoplasm, and nucleoid under the light microscope.
The fluorescent signals we used for this purpose are the ratios of red and greenfluorescence (red fluorescence/green fluorescence 3 1,000 [R/G ratio]) of acri-dine orange excited at 380 to 420 nm. In all treatments and stages in the life cycle(see Table 4), ratios for wall materials remained stable at 150 to 154 and thusserved as a check on the comparability of the technique among treatments.Variation among samples in ratios measured for cytoplasm and nucleoids wasdue primarily to either RNA enrichment or the condensation state of DNA.Acridine orange fluoresces green (maximum at 530 nm with excitation at 380 to420 nm) when intercalated near A-T (or A-U) base pairs of double-strandednucleic acids (e.g., DNA and duplex sections of tRNAs) but fluoresces red(maximum at 620 nm) when associated with single-stranded nucleid acids. (Redand green autofluorescence of fixed, unstained cells was ,5% of values forstained cells and is ignored here; red fluorescence of epulos [reported in refer-ence 14] occurred when live cells were excited at 510 to 560 nm). Thus, nucleoidswith highly condensed DNA will generate lower R/G ratios than nucleoids withdecondensed DNA, those with relatively high frequency of single-stranded DNAsegments, or those enriched with RNA; in fact, any sites enriched with RNA willexhibit high R/G ratios (11–13, 33).
In order to control for RNA content in nucleoids and cytoplasm of epulos,cells taken at the same life history stage and from the same samples as thosescanned for Fig. 3 through 6 were treated with RNase (DNase-free RNase A;Sigma). Cells were incubated in RNase solution (100 U/ml) for 2 h at 37°C,washed in cold phosphate buffer, stained with acridine orange, and scanned asdescribed above. This control demonstrates the impact of RNA enrichment onratios from various samples and explains why ratios may appear to overlap fordifferent regions of the cells. For example, R/G ratios for cytoplasm prior toRNase treatment ranged from 174 to 210 but dropped to 0 after RNase treat-ment as red fluorescence was eliminated. For nucleoids, ranges of ratios pre- andpost-RNase treatment were 92 to 210 and 96 to 139, respectively, demonstratingthat ratios for DNA did not overlap with those of cytoplasm and cell wall.
We used a special microfluorometer equipped with both conventional andcontact fluorescence objectives for this work (5, 6). This included an epifluores-cence illuminator (OI-30; Leningrad Optical and Mechanical Corporation) de-signed for work with both contact objectives (tube length, 190 mm) and conven-tional objectives (tube length, 160 mm), a focusing system for contact objectives,changeable filters, rectangular changeable measuring diaphragms, and a Nikonphotomultiplier. To create digital images which show distribution and state ofcellular components at various stages of the cell cycle, we scanned longitudinaloptical sections through the central plane of cells. A fluorescence contact objec-tive (60 by 1.15), a rectangular measuring diaphragm (5 by 10 mm), and dual-wavelength microfluorometry at 530 and 620 nm allowed us to calculate the R/Gratio and plot values for each 5-by-10-mm frame. Digital images were thusproduced for uninucleoid (those with a single nucleoid) and binucleoid (thosewith two nucleoids) cells of approximately equal size from each separate timesample. These were then scanned into a computer, ranges of ratios representa-tive of different compounds were replaced by distinctive shadings, and edgeswere smoothed to present a more realistic image (see Fig. 3 and legend fordetails).
Microbes intended for light and electron microscopy were fixed (2% glutar-aldehyde–0.25 M sucrose–0.05 M cacodylate in distilled water or 2% glutaral-dehyde–0.05 M cacodylate in filtered seawater). Light micrographs were takenon Kodak Technical Pan film on a Leitz Orthomat E microscope with differentialinterference contrast (Nomarsky) optics. Specimens for transmission electronmicroscopy were postfixed in 1% OsO4, embedded in EMBED 812 resin, thinsectioned, and viewed on a JEOL 1200exII electron microscope.
Descriptive statistics were calculated, and statistical tests were performed withSystat (version 5.03) and Sigma Plot.
RESULTS
Life cycle of E. fishelsoni. Key events in the E. fishelsoni lifecycle include changes in mean cell size as well as changes innumber, size, shape and location of nucleoids and incipientdaughter cells (Table 1; Fig. 1) (23). Because cell replication
5602 BRESLER ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
211
.223
.74.
164.

events occur against a backdrop of consistent, daily changes inapparent cell function and size, we distinguish the cell cycle(restricted to cell division events) from the life cycle (whichincludes events throughout a 24-h period). We term cells witha single nucleoid uninucleoid and those with two nucleoidsbinucleoid. We have encountered E. fishelsoni cells with threenucleoids, but their extreme rarity (e.g., a single trinucleoid cellwas seen among 480 cells surveyed in 1988) leads us to ignorethem here.
In early morning, cells contain compact, round nucleoidslocated near the apices of the cells (Fig. 1A and B, 2, and 3).Nucleoids elongate during the day until, in late afternoon andevening, they attain a maximum of ;75% of the parent celllength in both uni- and binucleoid cells with lengths of .;100mm (23); nucleoids of smaller parent cells are usually ,50% ofparent cell length and, unlike nucleoids of larger cells, do notoverlap within the parent (Fig. 1C and D; similar to morpho-types E, G, or J described in reference 8). Also during the day,average cell length increases (Table 1), and nucleoids of inter-mediate lengths are encountered (Fig. 1E), eventually growingto afternoon and evening sizes, at which they make up largefractions of parent cell volume (Fig. 1F). Occasional nucleoidsin cells encountered at night (Fig. 1G and H) appeared moreoptically dense than usual (Fig. 1F). In any event, daughtercells are then released from the parent cell, which is destroyedin the process.
During the night, when the host fishes are inactive, binucle-oid cells are common (accounting for over 70% of all cells insome samples [reference 23 and this study]), as are cells thatappear to be recently released daughter cells based on theirlack of incipient daughter cells, and average cell length de-clines (Table 1). Data are presented below for DNA content ofrelatively small epulos (category 0; length, ;30 mm), but ap-parent epulos encountered late at night and in early morningmay be much smaller (Fig. 1C and D), approaching the lengthsof more commonly studied rod-shaped bacteria. These cellshave not been studied, but four observations indicate that theyare small forms of E. fishelsoni. They are absent from daytimesamples, are largely restricted to early morning samples, con-sistently produce two daughter cells, and link through recog-nizable intermediate stages to larger epulos.
General aspects of the epulo life cycle coincide with hostactivities (21). Increase in modal size of cells and nucleoidsoccurs during the day, when hosts are actively feeding and the
gut is completely filled with algal food materials; during thistime, pH of gut fluids is suppressed in areas where epulos occurin profusion (23, 24). Nocturnal declines in modal cell sizeoccur when hosts are inactive in reef shelters; no suppressionof pH in gut fluids is evident at night.
Structure and DNA content of compact nucleoids. E. fishel-soni cells collected at 0800 h included both binucleoid cells,with two compact, round nucleoids located near the poles ofthe cell (Fig. 2 and 3A), and uninucleoid cells, with a singlecompact nucleoid located at one pole (Fig. 3B). To avoidvariation in dispersion and state of DNA that might be causedby comparing different stages in the cell cycle, we initiallymeasured total DNA on cells from this morning collection(Table 2). As noted above, cells in size categories II through VIwere approximately twice the volume of cells within the pre-vious, smaller category.
Treatment of cells with fluorescent Fuelgen or RNase-ber-berine sulfate showed that nucleoids consisted of a dense ma-trix of DNA. Digital images based on acridine orange stainingyielded R/G ratios characteristic of condensed, double-strandednucleic acid (mean 5 128.3; standard deviation 5 7.8). Indi-vidual nucleoids differed somewhat in the range of measuredratios (e.g., 119 to 140 and 131 to 140 for Fig. 3A; 107 to 135for Fig. 3B) and in the specific location of particular ratioswithin a nucleoid, although values from the periphery of nucle-oids were generally higher than values from centrally locatedregions.
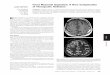
Electron micrographs of cells from the same collections(Fig. 2) support this interpretation, clearly showing condensedmaterials at the apices of cells. The two micrographs in Fig. 2represent two different fixations yet have important similari-ties. The most electron-dense materials are arrayed in elongatestrands ;200 nm in diameter, generally appear composed of acore of darkly staining material surrounded by a halo of slightlyless dense material, and exhibit striations with a periodicity of;40 nm. The position of the strands (dispersed in one section,clustered in the other) may be a function of the plane of thesection. Also, in each case the nucleoids are surrounded andseparated from the remaining cytoplasm by a line of uniden-tified material which appears continuous with a similar mate-rial at the perimeter of the cell.
Microfluorometric measurements of total DNA in nucleoidsof either Fuelgen- or berberine sulfate-treated cells from the0800-h sample demonstrated three patterns. First, DNA con-tent per cell is proportional to cell volume over a considerablerange of size (;30 to 520 mm [Table 2]), and variation in totalDNA content for cells of a given size, whether they wereuninucleoid or binucleoid, was relatively low; coefficients ofvariation ranged from 3.1% (group VI) to 17.6% (group I).
Second, uninucleoid and binucleoid cells of the same sizecontain the same amount of DNA (Table 3). For example, theratio of total DNA in binucleoid cells (Table 3) to DNA in thenucleoid of a uninucleoid cell of the same size group rangedfrom 0.96 to 1.19 in the four groups measured; the individualratios did not differ significantly from 1.0 by the chi-square test(n 5 4; ratios calculated from means of uninucleoid andbinucleoid total DNA).
Third, for binucleoid cells of a particular size, the amount ofDNA per nucleoid is approximately identical for the two nucle-oids (Table 3). DNA content generally differs little betweendaughter nucleoids of individual cells, with mean ratios ofDNA content in the larger of the two nucleoids relative to thesmaller nucleoid ranging from 1.1 to 1.3. However, chi-squaretests using individual ratios for each of the 40 cells examinedper size group found no significant deviations from 1.0 in anygroup.
TABLE 1. Lengths of epulos collected from brown surgeonfish,Eilat, Israel (Red Sea), June 1988a
Cell length(mm)
No. of cells at time of collection (h)
0700–0800
1100–1200
1400–1500
2100–2200
2400–0100
0300–0400
,50 23 1 0 6 18 3050–101 56 34 11 42 80 78
102–154 31 44 35 28 19 10155–207 8 31 35 26 2 2208–261 1 10 33 6 1 0262–314 1 0 6 6 0 0315–367 0 0 0 6 0 0
Estimated mean 89 133 175 139 79 70
a Data are from 60 haphazardly chosen cells from each of two fish per timeperiod (n 5 120 cells per time period). Modes for each time period are in boldprint. Estimated mean lengths per time period were calculated by assuming thatall cells in a category were the median length for that category (rounded to thenearest 5 mm).
VOL. 180, 1998 DNA DYNAMICS IN GIANT BACTERIA 5603
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
211
.223
.74.
164.

FIG. 1. Light micrographs of stages in the life cycle of E. fishelsoni taken under differential interference contrast illumination. Bars 5 50 mm. (A) A 237-mm-longcell with two compact, apical nucleoids (arrowheads). The cell was collected at 0640 h. (B) A 45-mm binucleoid (arrowheads) cell collected at 0640 h. (C) Uninucleoidand binucleoid cells with expanded nucleoids. The larger cell is 60 mm long. Unlike larger cells, small nucleoids (arrowheads) of small binucleoid cells do not overlapwithin the parent cell. Cells were collected at 2000 h. (D) Very small binucleoid cells with nonoverlapping nucleoids (arrowheads). The smallest cell (arrow) is 23 mmlong with nucleoids that are ,10 mm long. The cells were collected at 2000 h. (E) A 216-mm binucleoid cell with oval nucleoids (between arrowheads). Note thepresumed spirillum (arrow) with a length of ;18 mm. Cells were collected at 0915 h. (F) A 195-mm-long uninucleoid cell exhibiting presumed “caps” (see text) ofcondensed DNA (arrowhead) at apices of maximally enlarged nucleoid. Cell was collected at 1640 h. (G) A 225-mm-long uninucleoid cell with optically distinct daughtercell found only in night samples (compare with Fig. 1F). Cell was collected at 2200 h. (H) A 184-mm-long binucleoid cell with optically distinct daughter cells. Cellswere collected at 0130 h.
5604 BRESLER ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
211
.223
.74.
164.

FIG. 2. Transmission electron micrographs of compact nucleoids of E. fishelsoni collected at 0800 h. Note presumptive condensed DNA in chromosome-like bodies(arrowheads), delicate cross-striations on some of these bodies, and separation of nucleoidal material from remaining cytoplasm by structures (arrows) continuous withsimilar materials below the cell wall at the tip of the cell. Bars 5 1 mm. (A) Cell fixed in 2% glutaraldehyde–0.05 M cacodylate in filtered seawater. (B) Cell fixed in2% glutaraldehyde–0.25 M sucrose–0.05 M cacodylate in distilled water.
VOL. 180, 1998 DNA DYNAMICS IN GIANT BACTERIA 5605
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
211
.223
.74.
164.

Changes in state and distribution of DNA with stages of thecell cycle. The following sections are best interpreted in light ofthe general effects of RNase treatment on samples (Table 4).R/G ratios of the periphery of cells, interpreted as cell wallmaterials, remained unchanged at 150 to 154. Ratios for cyto-plasm dropped from high values to zero due to the loss of redfluorescence from RNA. Ratios for compact, apical nucleoids
declined slightly (128.3 to 124.8) and those for apical capsremained essentially unchanged (111.4 to 112.1), indicatinglittle single-stranded nucleic acid. In contrast, the mean ratiofor enlarged nucleoids declined from 145.2 to 136.7, indicatingthat single-stranded nucleic acid is common in such nucleoids.
(i) Morning. As noted above, in early morning most cellscontain compact, apical nucleoids (Fig. 1A and B). The DNA
FIG. 3. Digital images and graphical representation of the same images for binucleoid (A) and uninucleoid (B) E. fishelsoni cells collected at 0800 h and containingcompact nucleoids similar to those shown in Fig. 1A and B. Both cells measured ;520 by 65 mm. Cells were stained with acridine orange as described in the text. Valuesin digital images are R/G fluorescence ratios. Numbers in italics are values consistent with glycoprotein of cell walls; bold and underlined numbers are values consistentwith condensed DNA or condensed DNA plus RNA; remaining values are consistent with RNA-enriched cytoplasm, with higher values reflecting higher concentrationsof RNA. For visual clarity, ranges of values were identified for condensed DNA, decondensed DNA, general cytoplasm, RNA-enriched cytoplasm, and cell wallmaterials. Values were then replaced by distinctive shading or hatching patterns, and the edges were smoothed to more closely represent cell configuration. Subsequentfigures present only the graphical images; copies of the original digital images are available from the authors and are posted on W. L. Montgomery’s web site(http://www2.nau.edu/;wlm). In some figures generated from digital images, cell wall material appears unusually thick along sides and, particularly, apices of cells. Thisis due to readings of thick optical sections along strongly curved portions of whole cells. More accurate representations of walls occur in electron micrographs (Fig.2) (14, 23).
TABLE 2. DNA content per cell for seven size categories of E. fishelsoni cells determined by two methodsa
Group Cell length(mm)
Volume(mm3, 103) Relative volume
DNA content per cell (arbitrary units 6 95% confidence interval)
RNase 1 berberine sulfate staining Fluorescent Fuelgen reaction
Measured Relative Measured Relative
0 30–35 0.169 1.0 0.263 6 0.007 1.0I 150–160 10.72 63.2 (1.0) 17.2 6 0.9 65.4 (1.0) 22.1 6 1.0 1.00II 190–200 20.93 123.4 (1.9) 34.3 6 1.4 130.4 (2.0) 44.8 6 1.6 2.03III 240–250 40.88 241.0 (3.9) 68.2 6 3.0 259.3 (4.0) 90.75 6 2.1 4.10IV 300–320 85.74 505.5 (8.0) 137.7 6 4.7 523.6 (8.0) 177.5 6 3.8 8.03V 390–420 167.47 987.4 (15.6) 274.3 6 3.6 1,043.0 (15.9) 353.1 6 3.6 15.97VI 490–520 340.42 2,007.2 (31.7) 544.2 6 13.4 2,069.2 (31.6) 703.7 6 5.6 31.8
a Group 0, which includes the smallest specimens examined, has not been studied in detail (see subsequent tables) but is consistent with other size groups in termsof DNA quantity-volume relationships. Relative volumes and relative measures of DNA quantity per cell for the RNase plus berberine sulfate technique (see text) arecalculated, therefore, using both group 0 and group I values (parentheses) as denominators. n 5 60 cells for each group.
5606 BRESLER ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
211
.223
.74.
164.

in these nucleoids exhibited R/G ratios characteristic of con-densed DNA. Ratios for nucleoids in the samples collected at0800 h averaged 128.3 (95% confidence interval, 61.0 [Table4]). Shortly before this, in samples collected at 0640 h, averageR/G ratios were even lower (109.8 6 2.5), suggesting that DNAin the 0800-h samples was partially decondensed.
R/G ratios for the general cytoplasm and the extreme pe-riphery of cells were similar for the 0640- and 0800-h samples.High ratios characteristic of single-stranded nucleic acids, pri-marily RNA, occurred in cytoplasm at 0640 and 0800 h(182.1 6 0.2 and 184.4 6 0.5, respectively). Lower ratios char-acteristic of proteins and glycoproteins occurred along theedge of the cells (150.7 6 0.2 and 151.8 6 0.2, for the sameperiods, respectively), consistent with deposition of cell wallsand, perhaps, membrane-associated enzymes.
(ii) Late morning. In most cells collected between 1000 and1100 h, nucleoids in both uninucleoid and binucleoid cells hadenlarged and shifted from the apices toward the center of thecell (Fig. 4). Nucleoids were characterized by decondensedDNA (reflected in an increase of the R/G ratio to 140 to 148;mean, 145.2 6 0.4 [Table 4]) distributed evenly around theperiphery of the nucleoid and core areas rich in RNA (R/Gratios of 267 [Fig. 4]). This is likely the stage described andshown by Robinow and Kellenberger (26). RNA-rich areas,
with ratios in the 250 to 259 range, also occurred in the cyto-plasm external but adjacent to the nucleoids. The cytoplasmicmean R/G ratio (210.6 6 2.3) was higher than that for earlierperiods when DNA was condensed, suggesting more activeRNA synthesis and transport to the cytoplasm with deconden-sation of DNA. R/G ratios for peripheral portions of the cellremained unchanged from earlier samples (151.6 6 0.2 [Table4]).
(iii) Early afternoon. Elongation of nucleoids continues un-til those of both uninucleoid and binucleoid cells reach approx-imately 75% of cell length (23). During this period of elonga-tion, which in our samples extended until at least 1430 h, R/Gratios remained much as described for the samples from 1000to 1100 h (Fig. 4). DNA remained decondensed (ratios of 140to 149) and evenly dispersed around the periphery of thenucleoid. The interior core of the nucleoids, as well as areasimmediately external to the nucleoidal DNA layer, continuedto exhibit R/G ratios indicating RNA enrichment (245 to 255),cytoplasm external to these enriched areas remained high at;184 to 187, and the cell edge was unchanged at ;150.
(iv) Evening. Late in the day and into night (1640 and2200 h), nucleoids within both uninucleoid and binucleoid cellswere surrounded by material producing ratios previously seenonly at the edge of cells (R/G ratio, ;150) and were separated
TABLE 3. DNA content per nucleoid of uninucleoid and binucleoid cells as determined by RNase plus berberine treatment for four of sixsize groups of E. fishelsonia
Group DNA content per nucleoidof uninucleoid cells
Size of nucleoidmeasuredb
DNA content per nucleoidof binucleoid cells
Binucleoidtotal DNA L/S ratioc Bi.total/uni.
ratioc
I 20.3 (0.8) Large 10.4 (0.3) 18.4 (0.4) 1.3 (0.05) 0.96Small 8.0 (0.2)
III 70.4 (2.1) Large 43.8 (0.7) 81.6 (1.4) 1.2 (0.02) 1.19Small 37.9 (0.7)
V 272.6 (3.9) Large 145.6 (1.0) 282.4 (1.3) 1.1 (0.01) 1.04Small 136.8 (0.8)
VI 551.8 (11.0) Large 298.2 (2.5) 564.3 (4.2) 1.1 (0.01) 1.04Small 266.2 (2.8)
a Forty uninucleoid and 40 binucleoid cells of each size group were examined. Binucleoid total DNA is the mean sum of DNA content of large and small binucleoidcells. The L/S ratio is the ratio of DNA content of large binucleoid cells/DNA content of small binucleoid cells. The Bi.total/uni. ratio is the ratio of binucleoid totalDNA/uninucleoid cell DNA content. Values are means (standard errors). DNA content is given in arbitrary units.
b Differences between means of small and large nucleoids were significant at ,0.001 level for all size groups (paired-sample t test).c Ratios were not significantly different from 1.0; chi-square test, x2
0.05[39] 5 54.6 (for L/S ratios) or x20.05[3] 5 7.8 (for Bi.total/uni.ratios).
TABLE 4. Values of R/G fluorescence ratios in the nucleoids, cytoplasm, and peripheral areas of E. fishelsonia
Nucleoid type Mean R/Gratio SD SE 95% confidence
intervalMinimum R/G
ratioMaximum R/G
ratioDNA content
(arbitrary units)
Apical with condensed DNANucleoid 128.3 (124.8) 7.8 (7.2) 0.5 (0.5) 1.0 (1.0) 107 (115) 210 (136) 247 (197)Cytoplasm 184.4 (0) 6.9 0.2 0.5 174 (0) 210 (0) 812Periphery 151.8 (151.4) 2.0 (1.9) 0.1 (0.2) 0.2 (0.3) 150 (150) 154 (154) 460 (135)
Elongate with decondensed DNANucleoid 145.2 (136.7) 3.4 (2.1) 0.2 (0.2) 0.4 (0.3) 140 (133) 148 (139) 232 (138)Cytoplasm 210.6 (0) 32.4 1.2 2.3 182 (0) 267 (0) 732Periphery 151.6 (150.9) 2.0 (1.7) 0.1 (0.2) 0.2 (0.3) 150 (150) 154 (154) 393 (131)
Capped with highly condensed DNANucleoid 111.4 (112.1) 13.4 (3.9) 1.0 (0.4) 1.9 (0.8) 92 (96) 132 (115) 187 (86)Cytoplasm 204.2 (0) 24.1 1.0 2.0 182 (0) 250 (0) 547Periphery 151.5 (151.1) 2.6 (1.2) 0.1 (0.2) 0.2 (0.3) 148 (150) 154 (154) 434 (135)
a Values in parentheses are measurements on different cells taken from the same samples and treated with RNase before scanning with a microfluorometer. See Fig.3 to 6 for distributions within cells at various stages in the life cycle of condensed and decondensed DNA, general and RNA-enriched cytoplasm, and peripheralmaterials. Note the stability of measurements for peripheral materials at all stages, in contrast to changes in degree of condensation of DNA (low ratios indicate highcondensation) in the nucleoids and enrichment of cytoplasm by RNA (higher values result from enrichment).
VOL. 180, 1998 DNA DYNAMICS IN GIANT BACTERIA 5607
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
211
.223
.74.
164.

from the parent cell’s periphery by a layer of material yieldingratios characteristic of general cytoplasm (R/G ratio, 182 to186 [Fig. 5]). Shifting between fluorescence and transmittedlight microscopy demonstrated that the areas with R/G ratiosof ;150 were thickened zones surrounding nucleoids and thatthey were separated from, but structurally and visually similarto, the parental cell wall. This is consistent with electron mi-crographs which show nucleoids surrounded with thick, lami-nar material (23) (Fig. 3 and 5 and our unpublished results).We interpret this new layer as cell wall formation aroundmaturing daughter cells.
Evening samples also contained cells with well-developedwalls, with DNA evenly dispersed immediately below the walls,and with little or no cytoplasm external to the DNA layer. Such
cells were of a size and structure similar to daughter cellswithin parent cells and are interpreted as daughter cells fol-lowing their release.
(v) Early morning. In some daughter cells collected between1640 and 0500 h, the DNA located at one or both apices of thecell was very highly condensed (R/G ratio, 96 to 115; mean,111.4 6 1.9 [Table 4]) compared to the peripheral DNA (R/Gratios, 130 to 140; mean, 138.3), and DNA near the apicesappeared thicker than that along the lateral walls (Fig. 6B C).We termed these condensed, thickened areas of apical DNA“caps.”
Throughout the night, one encounters daughter cells with no(Fig. 6A), one (Fig. 6C), or two caps (Fig. 1F and 6B). In manycells collected at 0430 and 0500 h, however, the DNA was
FIG. 4. Binucleoid and uninucleoid E. fishelsoni collected at 1000 h. Dimensions: ;320 by 45 mm (A) and ;330 by 55 mm (B). Other features are as described inthe legend for Fig. 3. Note expansion of nucleoids, their shift toward the center of the cells, the concentration of decondensed DNA along periphery of nucleoid, andthe high concentration of RNA in core of nucleoids.
FIG. 5. Formation of mature daughter cells in binucleoid and uninucleoid E. fishelsoni collected at 1640 h. Dimensions: ;520 by 65 mm (A) and ;530 by 65 mm(B). Other features are as described in the legend for Fig. 3. Note formation of presumptive cell wall material around nucleoids and the separation of this wall materialfrom the parental wall by a thin layer of cytoplasm. Cells for this figure were collected at the same time as those shown in Fig. 6A through C, emphasizing that stagesin the epulo life cycle are not precisely synchronized in time and between individual host fish.
5608 BRESLER ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
211
.223
.74.
164.

arranged in a figure-8 form, with the apical caps of condensedDNA located at the top and bottom of the “8,” and withdecondensed DNA near the center of the cell displaced inward(Fig. 6B and C) to produce a pinched appearance. This pinch-ing appears to be independent of any obvious cytologicalchanges such as formation of cross walls.
Other cells sampled during these hours, also with distinctcaps of condensed DNA, exhibited complete or nearly com-plete separation of the decondensed DNA located more cen-trally (Fig. 6D and E), suggesting that this stage represents astep en route from diffuse distribution of decondensed DNA toits segregation to form daughter nucleoids. Additional conden-sation and aggregation of the DNA around the caps couldsubsequently produce the next distinct stage of condensed,apical DNA masses noted in samples from 0640 h (above).During this period of apparent dynamism with respect to DNAdistribution and state, R/G ratios for cytoplasm (204.2 6 2.0[Table 4]) and periphery (151.5 6 0.2) remained similar tothose recorded at other stages in the life cycle.
DISCUSSION
The morning nucleoid. Previously published electron micro-graphs (14, 23) show specimens fixed in late afternoon or
evening and show considerable complexity of epulo ultrastruc-ture, including a trend for the nucleoid regions to be clearlyseparated from surrounding materials. Where a structure re-sponsible for this separation has been visible, it has generallyappeared thick and occasionally laminar and has been inter-preted as wall material. Specimens used for Fig. 2 were fixedshortly after the first appearance of apical nucleoids of con-densed DNA in early morning and demonstrate that even atthis early stage in the life cycle, the nucleoid appears to beseparated from the cytoplasm. This is clearest in Fig. 2A,where a fine, smoothly curving line surrounds the nucleoid.The identity of the delineating feature and its significanceremain unclear, but spore formation in Bacillus subtilis involvesthe surrounding of an incipient spore by maternal cell mem-brane and the subsequent formation of a spore coat (17).Angert et al. (1) suggest that similar events occur duringdaughter cell and endospore formation in a close relative of E.fishelsoni, Metabacterium polyspora.
Internally, the nucleoid at this time appears dominated byelongate and possibly folded strands reminiscent of condensedchromosomes of dinoflagellates and other bacteria (see Fig. 3,4, and 27 in reference 30 and the discussion of them in refer-ence 26). We assume these structures are composed primarilyof DNA because similar materials are not evident in otherparts of the cell, and fluorescence staining with acridine orange(mean R/G ratio 5 128.3, typical of condensed DNA) andDAPI (49,6-diamidino-2-phenylindole) (reference 23 and ourunpublished results) demonstrates the presence of condensedDNA only in these nucleoids. Possible coarse aggregation ofprokaryote DNA (reviewed in reference 26) in fixatives con-taining glutaraldehyde cannot account for the condensedstrands seen in electron micrographs (Fig. 2), because dis-persed nucleoidal materials similar to more conventional viewsof bacterial nucleoids are evident in micrographs of cellstreated with the same fixatives but later in the cell cycle (14,23).
Finally, R/G ratios in zones adjacent to the nucleoids indi-cate some RNA enrichment (R/G ratio 5 193 to 210 [Fig. 3Aand B]), but values are much lower than those recorded forlate-morning cells with decondensed DNA (R/G ratio 5 250 to267). While not definitive, this suggests less transcriptionalactivity of condensed nucleic acid compared to decondensedmaterial, parallelling observations from eukaryote chromo-somes.
Treatment with RNase generated additional support for thishypothesis (Table 4). First, the highest values of R/G ratios incytoplasm occurred either in cells with enlarged nucleoids(mean R/G 5 210) or in those in transition with some DNA incondensed caps and some remaining dispersed (mean 5 204).Note that ratios for cytoplasm dropped to zero after RNasetreatment due to the loss of all red fluorescence from single-stranded nucleic acid. Second, ratios for apical nucleoids de-clined slightly (128.3 to 124.8) and those for apical caps re-mained essentially unchanged (111.4 to 112.1), supporting theinterpretation that there is little RNA in the nucleoids duringsuch periods and that DNA is condensed, double stranded, andgenerally not transcribing. In contrast, the mean ratio for theelongate nucleoids declined from 145.2 to 136.7. This decline isconsistent with the contention that RNA is common in suchnucleoids, as indicated by RNA-enriched zones within nucle-oids in Fig. 4. Digestion of RNA in cells with expanded nucle-oids left a ratio higher (136.7) than that for similarly treatedcells with compact nucleoids (112.1 and 124.8), indicating thata greater fraction of the DNA in enlarged nucleoids is insingle-stranded form than in compact nucleoids. Finally, ratiosin small areas of enlarged nucleoids (results not shown but
FIG. 6. Maturing daughter cells of E. fishelsoni collected at 1640 h (Athrough C) and 0430 h (D through E). Other features are as described in thelegend to Fig. 3. (A) Daughter cell (;390 by 45 mm) lacking caps and withdecondensed DNA evenly dispersed below the cell wall. (B) Daughter cell (;350by 45 mm) with two caps. (C) Daughter cell (;350 by 45 mm) with single cap. (D)Daughter cell (;360 by 45 mm) with two caps and DNA almost completelyseparated. (E) Daughter cell (;360 by 45 mm) with two caps and DNA com-pletely separated. Note mid-cell constriction of DNA in panels B and C thatappears unrelated to additional wall formation (contrast with Fig. 4) and con-densation of DNA in caps while that along periphery and in center of cellremains decondensed.
VOL. 180, 1998 DNA DYNAMICS IN GIANT BACTERIA 5609
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
211
.223
.74.
164.

available from the authors) declined to zero after RNase treat-ment (zeros were not used in calculations of R/G ratios forTable 4), again emphasizing the apparent concentration ofRNA within the central regions of expanded nucleoids.
Cell size, volume, and DNA content. E. fishelsoni appears tobe not only the largest known eubacterium but the most vari-able in size as well. Most of our data derive from studies onlarge cells (length, .150 mm), supplemented by work with onemuch smaller group (30 to 35 mm). Four observations make itclear that group 0 cells in fact are small E. fishelsoni. First, theproduction of one or two nucleoids or daughter cells within theparent cell as well as the elongate cigar shape of E. fishelsonicells is consistent throughout the size range and may, in fact,extend well below this range (Fig. 1C and D).
Second, length-width relationships (from which volume wasestimated) are consistent among epulo cells across the sizerange. We performed a linear regression on log10(length) andlog10(volume) data for groups 0 and I to VI (Table 2) supple-mented with data from cells of intermediate length not usedfor DNA analyses (length ranges of four groups: 46 to 55, 65to 75, 94 to 109, and 113 to 137 mm); the regression[log10(volume) 5 25.1 1 2.8 log10(length); r2 5 0.99, df 5 9]indicates consistent length-volume, and therefore length-width,relationships across the entire span of cell sizes.
Third, there is a consistent pattern in the range of sizes ofcells collected from hosts taken at different times of day andnight (Table 1) (23). Small- to medium-sized cells (length, ;30to 150 mm) prevail in the morning (0600 to 0800 h), averagelength increases during the day to a maximum in late afternoon(ca. 1600 h), and length declines thereafter to a minimum invery early morning (2400 to 0400 h). Consistent with this, verysmall (length, ,50 mm) cells are essentially absent during theday, occur in combination with a wide length range of cellsprior to midnight, often dominate epulos in fish sacrificedbetween midnight and 0400 h, and decline in frequency andeventually disappear in later morning samples.
Finally, preliminary staining of very early morning sampleswith fluorescent probes developed by Angert et al. (2) for largeE. fishelsoni yielded positive binding to small as well as inter-mediate sized cells (data not shown).
The roughly 30-fold increase in both cell volume and totalDNA for groups I to VI clearly exceeds the ;3.3-fold differ-ence or the ;10.6-fold difference expected if DNA quantitywere proportional to length or surface area, respectively. In-clusion of group 0 in calculations demonstrates a direct corre-lation between cell volume and DNA content across the entiresize range of E. fishelsoni measured (length, ;30 to 500 mm),with DNA content differing .2,000-fold among these cells. Ifstructurally similar cells in the 10-to-15-mm size range (Fig. 1)are in fact E. fishelsoni and adhere to the same pattern ofproportionality, as we suspect based on observations of inter-mediate sized cells, they would extend the ratio of cell volumeand DNA in largest to smallest cells by at least an additionalorder of magnitude (based on the regression described above,a 10-mm-long cell [the approximate size of daughter cells in thesmallest parent cell of Fig. 1D] would have a volume of ;0.005mm3, ;1/68,000 the volume and DNA content of a 500-mm-long cell.
DNA content and distribution to daughter cells. Flow cyto-metric studies of E. coli show that DNA content varies system-atically among other bacterial cells, and to some degree withcell size, but this variation usually represents only two- toeightfold increases over the amount of DNA within a singlegenome (29). We do not know the size of a unit genome in E.fishelsoni, but copy number in large cells appears to be verygreat. There are large quantities of DNA in large cells, there is
localization of DNA-specific stains to nucleoids, and fluores-cence ratios indicate condensation of DNA when electron mi-crographs show darkly staining, elongate structures in the nu-cleoids.
Many copies of a unit genome may support the growth,mobility, and apparently active metabolism described for thesegiant bacteria (9, 24, 26). Multiple copies of an entire genomewould also support rapid production of daughter cells by un-coupling potentially rate-limiting DNA replication from DNA-subdivision and other cell division events.
In epulos there is low variance in DNA content for cells ofa given size, an arithmetic increase in DNA content with cellvolume, and a ratio of DNA in the two nucleoids of binucleoidcells that is not significantly different from 1.0 (Table 3). Thus,there appears to be a mechanism for approximately equalsubdivision of large quantities of DNA between two daughtercells during the two rather distinct diurnal and nocturnalphases of the life cycle. During the day, organized subdivisionof DNA to daughter cells occurs when both parent cells andnucleoids are growing, and replication must be active to main-tain the ratio of DNA to cell volume. At night, parental cellsproduce smaller daughter cells (usually 50 to 75% of parentcell length) and replication is likely halted or perhaps proceed-ing at low levels.
The mechanism for subdivision of DNA may be more com-plex than that inferred by simple formation of equivalentdaughter cells. We have recorded E. fishelsoni cells with threedaughter cells of roughly equal size, and some morphotypesconsistently produce up to seven similarly sized daughter cells(8). We lack data on DNA content and dynamics in thesemorphotypes.
Changes in state and distribution of DNA with stages of thecell cycle. Clearly, a complex series of events attends growthand maturation of epulos. Quantitative fluorescence cytochem-istry demonstrates that DNA in the largest epulos exceedsamounts in the smallest cells by 4 to 5 orders of magnitude andprobably exceeds amounts in most other bacteria by an evengreater margin. Such large quantities of DNA could pose prob-lems for a cell undergoing the dynamic processes of DNAreplication, condensation and decondensation, and systematicdistribution to duplicate nucleoids.
Organization of DNA into discrete structures, as suggestedby electron micrographs and strong, localized fluorescence sig-nals from small, delimited morning nucleoids, may overcomesome of these problems. Structures apparent within early-morning nucleoids (collected between 0640 and 0900 h) arereminiscent of condensed chromosomes of other bacteria anddinoflagellates (26, 30), but we lack details of their fate duringthe remainder of the day. Previous electron micrographs ofsections through enlarged nucleoids and immature daughtercells generally lack evidence of such structures (14, 23), al-though diffuse, darkly staining zones are evident in the interiorcytoplasm of a daughter cell shown in a figure by Clements andBullivant (Fig. 2 of reference 9). In any event, by late morning(1000 to 1100 h), the nucleoids exhibit evidence of DNA de-condensation (increased R/G ratios) and increased transcrip-tional activity (RNA enrichment within the nucleoid and inadjacent cytoplasm), and as the day and evening progress DNAappears to continue decondensation and widespread disper-sion.
Condensation of DNA and its association into duplicatecaps by an unknown mechanism appear to presage the forma-tion of daughter nucleoids and eventually daughter cells. Con-versely, replication with formation of a single cap could be amechanism for increasing DNA content of a cell line and
5610 BRESLER ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
211
.223
.74.
164.

supporting additional increases in cell dimensions within thatline.
Such events may be carried out without interruption of nor-mal cell functions. Even as caps form from highly condensedDNA, the cells retain dispersed and probably actively tran-scribing DNA in a layer immediately adjacent to the cytoplasm(Fig. 6B through E). Large quantities of cytoplasmic RNA,reflected in our high R/G ratios for cells throughout the lifecycle, and intense, cell-wide fluorescence signals obtainedwhen cells were probed with specific 16S ribosomal subunitsequences (2), collectively suggest that cells are metabolicallyactive at all stages.
ACKNOWLEDGMENTS
Funding was provided by the Tobias Landau Foundation; grant3716-87 from the National Geographic Society; the Organized Re-search Fund of Northern Arizona University (NAU); and the Depart-ment of Zoology, Tel Aviv University.
L. Fritz performed fixations in Eilat and, assisted by M. Sellers andR. Earhart, the electron microscopy at NAU. Collections in Israel wereundertaken at the Interuniversity Institute of Eilat with the graciousassistance of Avi Baranes as well as many resident faculty, technicians,and students. Collecting permits were provided by the Nature Protec-tion Society (NPS) of Israel; particular thanks are due Nurit Popper ofthe NPS for assistance in arranging permits and facilitating work.Robyn O’Reilly prepared the computer maps of cells. GordonSoutham and several anonymous reviewers provided invaluable cri-tiques of the manuscript.
REFERENCES
1. Angert, E. R., A. E. Brooks, and N. R. Pace. 1996. Phylogenetic analyses ofMetabacterium polyspora: clues to the evolutionary origin of daughter cellproduction in Epulopiscium species, the largest bacteria. J. Bacteriol. 178:1451–1456.
2. Angert, E. R., K. D. Clements, and N. R. Pace. 1993. The largest bacterium.Nature 362:239–241.
3. Arndt-Jobin, D. J., and T. M. Jobin. 1989. Fluorescence labeling and mi-croscopy of DNA. Methods Cell Biol. 30:417–448.
4. Bohm, N., E. Sprenger, and W. Sandritter. 1973. Absorbance and fluores-cence cytometry of nuclear Fuelgen DNA. In A. A. Thaer and M. Sernetz(ed.), Fluorescence techniques in cell biology. Springer-Verlag, Berlin, Ger-many.
5. Bresler, V., and L. Fishelson. 1994. Microfluorometrical study of benzo(a)pyrene and marker xenobiotics bioaccumulation in the bivalve Donaxtrunculus from clean and polluted sites along the Mediterranean shore ofIsrael. Dis. Aquat. Org. 19:193–202.
6. Bresler, V., and V. Yanko. 1995. Chemical ecology: a new approach to thestudy of living benthic epiphytic foraminifera. J. Foraminifer. Res. 25:267–279.
7. Caron, G. N.-V., and R. A. Bradley. 1995. Viability assessment of bacteria inmixed populations using flow cytometry. J. Microsc. 179:55–61.
8. Clements, K. D., D. C. Sutton, and J. H. Choat. 1989. Occurrence andcharacteristics of unusual protistan symbionts from surgeonfishes (Acanthu-ridae) of the Great Barrier Reef, Australia. Mar. Biol. 102:403–414.
9. Clements, K. D., and S. Bullivant. 1991. An unusual symbiont from the gutof surgeonfishes may be the largest known prokaryote. J. Bacteriol. 173:5359–5362.
10. Crissman, H., and J. Steinkamp. 1990. Cytochemical techniques for multi-variate analysis of DNA and other cellular constituents, p. 227–241. In M.Melamed, T. Lindmo, and M. Mendelsohn (ed.), Flow cytometry and sort-
ing, 2nd ed. Wiley-Liss, New York, N.Y.11. Darzynkiewicz, Z. 1993. Mammalian cell cycle analysis, p. 45–69. In P. Fantes
and R. Brooks (ed.), The cell cycle: practical approaches. Oxford UniversityPress, Oxford, England.
12. Darzynkiewicz, Z., and J. Kapuscinski. 1990. Acridine orange: a versatileprobe of nucleic acids and other cell constituents, p. 291–314. In M. Mel-amed, T. Lindmo, and M. Mendelsohn (ed.), Flow cytometry and sorting,2nd ed. Wiley-Liss, New York, N.Y.
13. Domkina, L. K., V. M. Bresler, and L. N. Simanovsky. 1976. Changes in thestructure of chromatin in the nuclei of hepatocytes of rats trained to hypoxia.Bull. Exp. Biol. (Moscow) 81:370–373.
13a.Fishelson, L. Unpublished data.14. Fishelson, L., W. L. Montgomery, and A. A. Myrberg, Jr. 1985. A unique
symbiosis in the gut of tropical herbivorous surgeonfish (Acanthuridae:Teleostei) from the Red Sea. Science 239:49–51.
15. Ghiorse, W. C., and D. I. Balkwill. 1983. Enumeration and morphologicalcharacterization of bacteria indigenous to subsurface environments. Dev.Ind. Microbiol. 24:213–222.
15a.Grim, N. Unpublished data.16. Haugland, R. P. 1996. Handbook of fluorescent probes and research chem-
icals, 6th ed. Molecular Probes, Inc., New York, N.Y.17. Losick, R., P. Youngman, and P. J. Piggot. 1986. Genetics of endospore
formation in Bacillus subtilis. Annu. Rev. Genet. 20:625–669.18. McFeters, G. A. 1991. Acridine orange staining reaction as an index of
physiological activity in Escherichia coli. J. Microbiol. Methods 13:87–93.19. McFeters, G. A. 1995. Physiological assessment of bacteria using fluoro-
chromes. J. Microbiol. Methods 21:1–9.20. Millard, P. J., B. L. Roth, and C. H. Kim. 1997. Fluorescence-based methods
for microbial characterization and viability assessment. Biotechnol. Int.1:291–305.
21. Montgomery, W. L., L. Fishelson, and A. A. Myrberg, Jr. 1989. Feedingecology of surgeonfishes (Acanthuridae) in the northern Red Sea, withparticular reference to Acanthurus nigrofuscus (Forsskal). J. Exp. Mar. Biol.Ecol. 132:179–207.
22. Montgomery, W. L., and R. Galzin. 1993. Seasonality in gonads, fat depositsand condition of tropical surgeonfishes (Teleostei: Acanthuridae). Mar. Biol.115:529–536.
23. Montgomery, W. L., and P. E. Pollak. 1988. Epulopiscium fishelsoni, N.G., N.Sp., a protist of uncertain taxonomic affinities from the gut of an herbivorousreef fish. J. Protozool. 35:565–569.
24. Montgomery, W. L., and P. E. Pollak. 1988. Gut anatomy and pH in a RedSea surgeonfish, Acanthurus nigrofuscus. Mar. Ecol. Prog. Ser. 44:7–13.
24a.Montgomery, W. L., and P. E. Pollak. Unpublished data.25. Neidhardt, F. C., J. L. Ingraham, and M. Schaechter. 1990. Physiology of the
bacterial cell: a molecular approach. Sinauer, Sunderland, Mass.26. Robinow, C., and E. Kellenberger. 1994. The bacterial nucleoid revisited.
Microbiol. Rev. 58:211–232.27. Schmidt, E. L., and E. A. Paul. 1982. Microscopic methods for soil micro-
organism, p. 803–814. In A. L. Page, R. H. Miller, and D. R. Keeney (ed.),Methods of soil analysis, part 2. American Society for Agronomy and SoilScience Society of America, Madison, Wis.
28. Sharma, D. K., and D. N. Prasad. 1992. Rapid identification of viablebacterial spores using a fluorescence method. Biotech. Histochem. 67:27–32.
29. Skarstad, K., E. Boye, and H. B. Steen. 1986. Timing of initiation of chro-mosome replication in individual Escherichia coli cells. EMBO J. 5:1711–1717.
30. Spector, D. L. 1984. Dinoflagellate nuclei, p. 107–147. In D. L. Spector (ed.),Dinoflagellates. Academic Press, New York, N.Y.
31. Van Beelen, P., A. K. Fleuren-Kemila, M. P. A. Huys, A. C. P. van Montfort,and P. L. A. van Vlaardingen. 1991. The toxic effects of pollutants on themineralization of acetate in subsoil microcosms. Environ. Toxicol. Chem.10:775–789.
32. Van Dilla, M. A., R. G. Langlois, and D. Pinkel. 1983. Bacterial character-ization by flow cytometry. Science 220:620–622.
33. Yossefi, S., Y. Oschry, and L. M. Levin. 1994. Chromatin condensation inhamster sperm: a flow cytometric investigation. Mol. Reprod. Dev. 37:93–98.
VOL. 180, 1998 DNA DYNAMICS IN GIANT BACTERIA 5611
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
211
.223
.74.
164.