Embed Size (px)
Citation preview

UNITED STATES DEPARTMENT OF THE INTERIOR, Fred A. Seaton, Secretary
FISH AND WILDLIFE SERVICE, Arnie J. Suomela, Commissioner
SYSTEMATICS AND BIOLOGY OF THEGIZZARD SHAD (DOROSOMA CEPEDIANUM)
AND RELATED FISHES
By ROBERT RUSH MILLER
FISHERY BULLETIN 173From Fishery Bulletin of the Fish and Wildlife Service
VOLUME 60
PUBLISHED BY U.S. FISH AND WILDLIFE SERVICE. WASHINGTON. 1960
PRINTED BY U. S. GOVERNMENT PRINTING OFFICE, WASHINGTON 25, D.C.
For sale by the Superintendent of Documents, U.S. Government Printint Office, Washintton ]5, D.C.Price 15 cents

Library of Congress catalog card for the series, Fishery Bulletin of the Fishand Wildlife Service:
u.s. Fish a·nd Wildlife 8e1"Llice.Fishery bullet.in. v. 1-
Washington, U.S. Govt.. Print.. Off., 1881-19
v. in illus., maps (part fold.) 23-28 em.
Some ,·ols. issued in the congressional series as Senate or House(locuUlents.
BuHetins e01i11",sing \". 4i- also numbered 1-Tit.\e varies: v. 1-4U. Bulletin.Vols. 1-4!1 issued by Bureau of Fisheries (called l!'ish C01llmission,
v. 1-2:3)
1. Fisheries-U.S. 2. Fil!h-culture--U.S. I. Title.
II
SH11.A25
Libl'ar3' of Oongress
6a9.20617a 0-35239*

CONTENTSPage
Inh'oduction .. 371Ker to genera of Dorosomatinlle ._________________ 372f:enus f)orOSOtlHt .• 372Ke~' to species of DOl"osotlw ._________________ 373Do/'osoJl/a {,e.pe.diu1I-um. (LeSueur) .. 373
S~·non~·mr ._________________ 373Diagnosis ._~_______________ :~74
Desl'ription .__ ..,______________ 374CoIOl: 375Varilltion 375Studr materilll 375Ueprollllctioll lind development 376Fecumlitr 379Age and gl'owth 380Si1.p. 381
Habitat and migration 3S1~ood 382Predators ._________________ 383Ut.ili1.ation 38<:
AllUJI(lan.?e ~------------------.---------________ 384Pal'asi tes ~-------------------------_.___ _ _ 385Mortality ._________________ 385Forage va llle . .:.____ 386Uange :387
8111l11111ll'Y ._ __ _ __ _ __ _ _ _ _ 3S8Litel'lltul'e cited ._________________ 388
III

ABSTRACT
The gi7.7.a1'll shads, mal'ine ami fresh-watel' herrings of the suhfamil~' DQI'O
SOllllltinne, lire chat'adel'i7.ell alHl ke,l's aJ'e given to the seven known gener'lIalHl to the live spedes of the Amel'kan gen~ls Dorosollln (Sif/llf/losa is l'egal'ded as a subgenul'l). The s,l'stematics anll biolog~' of the eastern gi7.7.arllshafl, J)omsoll/·f/ cf?/)('difllllllll. are treated in detail. The biological infornmtionl'esults hu'ge1r fl'om a nitical surve~' of papel's (unpubJishell as well as pubIishecl"l clealing with its fec'undity. reproduction and elevelopment. Si7.e. ageamI gl'owth, IlbullClance. habitnt IIml migration. mortalitr. pal'asites Ilndprf'dntol's. allll its fooel, fOJ'age value. and utili7.ation b~' man, O,'iginal th'uwings of earl~' Iife-histltl,~, stagel! are IJres·ented. Tlti:" SI1lIIllHtl'~' l!lll.luld enablefisher~' bi ...logists concerned with inland wnterl! to gnin Il more e:olll)lrehensiveoutlook 'Ill the role of the gizzal'd "had in lil!h llllmagelllent,
IV

SYSTEMATICS AND BIOLOGY OF THE GIZZARD SHAD(DOROSOMA CEPEDIANUM) AND RELATED FISHES
By ROBERT RUSH MILLER, Curator of Fishes,Museum of Zoology, University of Michigan
The herrings of" the subfamily Dorosomatinae,known as gizzard and threadfin shads in the New""Vorld, comprise seven genera inhabitilig Asia,the Indo-Australian region, and North America.Fishery biologists dealing with management ofinland waters in eastern North America havebeen increasingly concerned about the role of thegizzard shad, DOro801lW· cepedi(UI:1(,1/I.~ in the ecology of fish populutions. Praised in some pla':esus a vahmble forage fish, this widespread species has elsewhere so overpopulated lakes thatcostly und time-consuming control measures havebecome necessary. A review of the systematicsand biology of this species 8ho~1ld make possiblea more critical assessment of its part in fish management.. The present paper is designed to summarize what is known of gizzard shads in generalnnd of D. cel1edia.nw1/. in particular.
As a gronp, the gizzard shads are characterized by t.he inferior, subterminal or terminal,toothless mouth and the gizzardlike stomach;they wel'e first recognized under the nameChatoessilllt by Glinther (1868: 381, 406). Theunit was raised to family rank, Dorosomidae,more properly Dorosomatidne, by Gill (1872:17), and was ret.ained at. t.his level for manyyears, at. least by American ichthyologists. Mostworkers now refer the group to the Clupeidae andrelegat.e the gizzard" shads at most. t.o a subfamily, Dorosomutinae, as did Berg (1940).
The body is generully short and deep (except.in t.he slender young) und moderately to stronglycompressed. The scales are thin, cycloid, andmore or less adherent; they are absent on theheu,d ancI may be present or ubsent on the backbet.ween the occiput and the dorsul fin. The abdomen is compressed to a ridge and armed withkeeled, bony scutes. The eyes hnve conspicuousadipose "eyelids," The gill membranes are sepa-
NOT.:.-Apl'roved for puhliclltion. No,'. 4. 1958. Jo'i.hery Bulletin l7:1.
rate and free from the isthmus; the gill rakersare slender, close set., and exceedingly numerous.There are five to six branchiost.egals, and thepseudo-branchiae are large.
Regan (1917: 297) induded this subfamily inthe CI.upeinae because he regarded the separationof the gizzard shads from the other genera centering about Olupea as an art.ificial arrangement.I have not st.udied t.he Old W"orld genera and canneit.her confirm nor dispute Regan's viewpoint.However, since there hns been no comprehensivestudy of the species and genera now referred tothe Dorosomatinae, the interrelationships of theOld and New ""Vorld forms should be regardedas tentative.
As a whole, the gizzard shads are migratoryfishes, primarily marine, entering fresh or brackish waters to spawn. Some species, and somepopulations of single species (such as DOT080'1na
repedif/:nm1lo and D. petenense) , are landlocked,completing their life cycle wholly in fresh water. Theil' food, except early in life, consistschiefly of minute organic partides which a,restrained by the fine, abundant gill rakers, aidedhy the accessory pharyngeal pockets (LagleI' andKraatz, 1945; Iwai 1956). The st.omach is ashort., t.hick-walled muscular struct.ure like thegizzard of a fowl; the int.estine is long and mucheonvoluted, with numerous folds on its innersnrface and hundreds of pyloric caeca extenlally.These digestive specializations further serve toussimiJa.te the minute food of these fishes.
All but t.wo of the seven genem, A'nodo-ntostoma· and Gonia.loi,/a, are further characterized byhaving the last, dorsa.l my prolonged int.o a conspicnolls, elongltte.d filament. in the adult.. OnlyD01'08onw (including S-ig·na.lo8fl) inhabits theNew '''orld; t.he other genera are found in Asiaund the Indo-Australian region.
In surveying the literature dealing with the371

372 FISHERY BULLETIN OF FISH AND WILDLIFE SERVICE
biology of Dm'o807lw, cepedi.((,ntt'ln, I have beenassisted very materially by t.he index and references in t.he libml'y of John van Oosten. Ot.herswho have aided in supplying dat.a or referencesare Reeve M. Bailey, George W. Bennett, S. M.Bower, Gerald P. Coope.r, Robert Cummins, Jr.,Alfred W. Eipper, Ralph Hile, Melvin T. Huish,Alfred Larsen, Foster D. Roszman, Milton B.Traut.man, C. R. Robins, and E. L. 'Vickliff. Iam particularly indebted to Edward N. Warner
for allowing me to publish his original drawings (figs. 2-4) and to summarize unpublished
. material from his doct.oral thesis, and likewise t.oAnt.hony Bodola for permission to use data fromhis doctoral dissertat.ion. William L. Brudonmade the drawing of the adult (fig. 1).
In the following key, the separat.ion of the AsiIttiC genera is based on the dat.a given by Regan(1917: 298, 308-317) and on t.he synopsis presented by Herre and Myers (1931: 234-236).
KEY TO GENERA OF DOROSOMATINAE
Ill.. Maxillary provided with two supramaxillary bones (the anterior one thinner and more scalelike than the posterioroneL New World (Doro8oma)
lb. A single supramaxillary bone (Old World genera) 22a. Last ray of dorsal fin produced into a conspicuous filament 32b. Last ray of dorsal fin Hot, produced into a filament or only weakly so (.4nodonto8toma and Gonialo8a) 5
3a. Maxillary flattened, wide posteriorly, without a downward curve at it,s distal tip; gill rakers of posteriorend of ceratohyalnearly as long as longest opposite gill filaments_ _________________________________ 4
3b. Maxillary little flattened, very narrow, with a downward angle at its distal tip; gill rakers of posteriorpart of ceratohyal only half or less than half as long as the opposite gill filaments __ Nematalo8a Regan
4a. Dentary normal, not reflected outward, fitting well up inside upper jaw with mouthclosed Clltpanodon LacepMe.
4b. 'Dentary anterior to point where it appears from beneath upper jaw (with mouth closed) with itssharp ramus reflected outward, its edge opposed to edge of upper jaw Kono8iru8 Jordanand Snyder
5a. Maxillary straight, thin, transversely expanded, tapering distally. Last dorsal ray little if at allproduced A nodonto8toma Blee.ker
5b. Maxillary slender, distally slightly expanded and curved downwards. Last dorsal ray notproduced . . Gonialo8a Regan
Genus Dorosoma Rafinesque
Gizzard shads, threadfin shad
Dor08om-a Rafinesque, Western Rev. and Misc. Mag.,1820: 2 (3): 171 (see Fowle-r 1945: 6,8); Ichth~'o
logia Ohiensis, 18:!O: 39. T~'pe specie-s by lllonotypy,D. 'Ilotata = D. cepedia.ll1l1n (LeSuem'). Type locality: Below the falls of the Ohio Rivel'.
.'iJignalosu EVel'lllann and Kendall, 1898. Bull. U.S. FishCOlllmis.sion, 17 (1897) : 127. Type- species, .'Jipnalosaatr.haJa.la.yae. = D. petene.n.se. (GUnther). Type localit~': Atchafalaya River, Melville, La.
(/om'11l.on 1/,mne.-The fishes of this genus havegenerally been referred to as gizzard shads beca·use of the gizzardlike muscular stomach. Thevernaculars, skipjack, hickory shad, mud shad,sawbelly, jack shad, et cetera, were formerly applied to D. cepedl(tnmn and are still used locally;t.he name Cl:lwun is used in Q,uebec. Threadfinshad has been adopted as the vernacular for D.pef;enen~e.
(Jelle1'ic cna'1'adel'.5'.-Clupeid fishes wit.h the last.ray of the dorsal fin prolonged into a slende.r fila-
ment (absent or inconspicuous III the YOlmg) ,thus resembling their marine New 'Vorld relat.ive Oplstlto'lle'l1Uf.. The mouth is small to moderate, terminal, subterminal, or inferior, t.he lowerjaw included or the jaws subequal. Mouth toot.hless in adult, but young with a row of fine teethon upper jaw. Maxillary with two supramaxillary bones. Snout short and rounded. Stomachgizzardlike., the intestine long and much convoluted, with numerous pyloric caeca. Body compressed, silvery, the abdomen armed with bonyscutes (tot.al, 23·-32). Dorsal rays 9-15; analrays 17-38; pectoral rays 12-17, petvic' rays 8;cltudal rays 19 (17 branched), rarely 17 or 18.Sca1es cycloid, thin, 40-83 along side. Vertebrae(including urostyle) 40-51.
Ra:nge.-All the species except D. s'I/1.ithl areconfin"ed to the Atlantic drainage of Nort.h and

SYSTEMATICS AND BIOLOGY OF' GIZZARD SHAD 373Middle America (Canada to Nicaragua). Theyare found from the Great Lakes-St.. LawrenceRiver basins, southern South Dakota, Nebraska,and Minnesota, and from about latitude 40° N.on the Atlantic seaboard of New Jersey andsoutheastern Pennsylvania southward to LakeNicaragua (but with discontinuous distributionfrom northern Guatemala and British Hondurasto Nicaragua). D. smithi is known only fromcoastal streams of northwestern Mexico, in Sonora, Sinaloa, and Nayarit.1
8pecies.-Five species of Doro8orna are recognized, four in the subgenus Dor0801no. and one,
D. petene'lMe, in the subgenus Signo.lo8o.. Theymay be distinguished by means of the followingkey. The resemblances between DO'l'080ma and8ignalo8Q. are numerous, the differences few, withno sharp structural gaps. The form and positionof the mouth afford the most reliable means ofdistinction. The relationships of the five species.seem t.o be better expressed by referring them allto Do·r080'lna., using subgenera to indicat.e thelesser phyletic lines.
1 The record for Na~'arlt Is based on 123 specimens In theUniversity of Michigan Museum of Zoology (UMMZ 171979)collected in 1955.
KEY TO SPECIES OF DOROSOMAIll.. Mouth terminal, ventral edge of upper jaw smooth. Fewer than 50 scales in lateral series, regularly arranged·
Anal rays 17-27, usually 20-25. Vertebrae 40-45. Atlantic slope from Florida, Tennessee, and Oklahoma westand south to northern Guatemala and British Honduras Subgenus S1'gnalosa, D. petenense (Giin,ther)
lb. Mouth subterminal or inferior, ventral edge of upper jaw with slight to pronounced notch (except in young). More~han 50 scales in lateral series, irregularly arranged. Anal rays 22-38, usually 29-35 (where range overlaps thatof S':gnalosa). Vertebrae 43-51 (47-51 where the two subgenera coexist) Subgenus Dorosoma
2a. Lateral scales 52-70, usually 58-65; scales around body 36--45; vertebrae 48-51. Atlantic drainage of easternNorth America south to Rio Panuco, Mexico D. cepedianu1I! (LeSueur)
2b. Lateral scales 70-83, usually 73-78; scales around body 46-60; vertebrae 43-48 .. 3311.. Dorsal filament long, it,s length as measured from dorsal origin 0.95 to 1.4 times, usually 1.1 to
1.3, in distance from pelvic insertion to tip of snout; anal base 1.1-1.4 in same distance; anal rays 29-38,usually 32-35. Atlantic slope of Mexico and northern Guatemala (RCo Papaloapan to Rio Usuma-cinta) D. anale Meek
:~b. Dorsal filament short, its lengt,h as measured from dorsal origin 1.4-1.85, usually 1.5-1.8, in distance frompelvic insertion to tip of snout; anal base 1.6-2.2 in same distance; anal rays 22-31, usually 23-29 4
4a. Mandible long, nearly one-half length of head; scales around caudal peduncle 20-26; dorsal raysusually 13 (12-14). Lakes Managua and Nicaragua, Nicaragua D. chaves': Meek
-lb. Mandible short, less than one-third length of head; scales around cauda! peduncle 2&-31; dorsal raysusually 11-12 (9-13, rarely 9, 10, or 13). Pacific slope of northwestern Mexico (Sonora toNayaritL ~ D. smithi Hubbs and Miller
In this work, only D01'osomo. cepediarl:ltin istreated in det.ail. Information on the systematiccharacters, growth, lmd relationships of the othert.hree species of the subgenus D01'os01na appea,redelsewhere (Miller 1950). Recent interest in thethreadfin shad, D01'080ma. (8-igna.losa.) petenense,as It fomge fish hits resulted in the experimentalplanting of this speeies in vltl'ious States (Parsonsand Kimsey, 1954; Kimsey 1954). Relativelylittle published data is avnilable on the systematicsand biology of the threadfin shad.
Dorosoma cepedianum (LeSueur)
Eastern gizzll,rd slutd, hickory shad,skipjack, mud shad
(Figures 1-4)
8yn01Wlll.y.-In the following partial synonymy, only the references to original descrip-
tions of the forms now regarded to be conspecificwith Do·rosoma. ceped';~Jrn.11'1n are given.
Megalop8 cepedia.n.a LeSueur. 1818, Jour. Acad. Nat. Sci.Phila.. 1: 361-363 (original description: markets ofBaltimore and Philadelphia, hence usually given asChesapeake and Delaware Bays).
Olupea. hetenl.ra· Rafinesque, 1818. Amer. Month. Mag.,1818: 354 (Ol"iginal descl'iption; Ohio River).
Dorosoma notata. Raflnesql1e, 1820, Western Rev. andMisc. Mag., 2: 172 (original descl'lption; falls of theOhio Rivel·).
01l.a.f.Oi!8S·1I8 elliptiC-liS Kirt.land, 1888. Rept. Zoo!. Ohio, in
Second Ann. Rept. Geol. Surv. Ohio, Columbus. 1888:169. 195 (nomen, n.//.d//.m, Ohio; same as DO/"OSOIl/II
nof(l.fa..l. 1844, Boston Jour. Nat. Hist.• 4 (2): 235237, pl. 10. fig. 1 (original description, comparisons.OCCUl'l'ence in Ohio, habits, mOl'talitl': Ohio Riverand its ·tributaries).
01l.0tO('88//8 vn8ocia.biU8 Abbott, 1861. Proc. Acad. Nat.Sci. Phila., 12 (1860) : 365-366 (Oliginal description.habits; "sturgeon pond" 2 miles below Trenton. N.J. ).

374 FISHERY BULLETIN OF FISH AND WILDLIFE' SERVICE
Mega-Zops bimacu.lata. LeSueur. 1848. in Cuvler and Valenciennes, Rist. Nat. Poiss., 21: 104 (nomel,. 11,'ltdu.m.;synonymized with Cha.toess·us cepetUamts by Valenciennes).
Dorosoma ceped·iatr./wl. ea:ile Jordan and Gilbert. 1883,Proc. U.S. Nat!. Mus.. 5 (1882): 585 (original description, based on 2 specimens, from Galveston,Tex.) .
Diagn.os1.8.-A gizzard shad with 52-70 lateralscales,2 36-45 scales around the body, 48-51 (usually 00) vertebrae, 25-36 anal rays, and preponderantly 18 + 12 ventral scutes.
The dorsal filament is comparatively long. Asin the other species of DO'rosoma, its length variesgreatly with age, and statements regarding thisstructure have little meaning unless accompaniedby data on the size of the specimen. The filamentis inconspicuous or absent in young fish, increasing in length with age up to a certain size range(around 200 mm. ~), and then decreasing in relative size.
Description.-Body depth 2.3-3.1 in standardlength (all measurements "stepped off" with apair of precision dividers); predorsaI length1.85-2.05; prepelvic length 2.0-2.35; anal originto caudal base 2.4-2.8; head length 3.0-3.9; lengthof dorsal filament 3.1-6.5 (0.8-2.0 in head
2 For method of counting scales see Miller (1950: 388-389).
length) ; length of anal base 3.2-3.9 (0.75-1.25 inhead length). Head .width 1.8-2.5 in head length;eye length 3.3-0.4; snout length 5.0-6.0; bonywidth of interorbital 3.3-4.5; length of upper jaw3.5--4.2; length of mandible 2.6-3.3; length ofcaudaI peduncle 2.4-3.5; depth of caudal peduncle2.45-3.4; length of pectoral 1.15-1.45; length ofpelvic 2.0-2.5; length of dorsal base 1.85-2.6;length of lower lobe of caudal 0.7-1.1, typicallylonger than anal base (rarely subequal).
Dorsal rays 10-13, average 11.61 in 197 specimens; anaI rays 25-36, 31.32, in 195; pectoral rays14-17,15.52, in 288 (144 individuals) ; pelvic rays8, rarely 7; and caudal rays 19. Lateral scales(first scale counted was first one lying aboveuppennost corner of gill opening) 52-70, 61.06,in 67; scales between dorsal and anal fins 19-24,21.77, in 52; scales around body (beginning withfi·rst scale just in front of left pelvic fin) 36-45,41.03, in 73; and scnJes around caudal peduncle(slenderest part) 16-20, 18.02, in 53. PrepelvicsClItes 17-20, 17.99, in 196; postpelvic sClItes 10-14,11.76, in 197; totn.l ventral sClItes 27-32, 29.74,in 196. Number of vertebrae (including urostyle) 48-51, 49.83, in 42 specimens. Gill rakersvery numerous nnd fine, those on first archnumber about 90 to 300 at standard lengths of
FIGURE I.-Adult female DOl'wot/lQ· cepedlQt~llm ~ lTMMZ 128171), 180 nUll. standard leiigth (about 9 inches. total length).from Lafayette County, Arkansas. (Drawn by W. L. Brudon.)

SYSTEMATICS AND BIOLOGY OF GIZZARD SHAD 375
20-65 mm., about 350 at. 95 mm., and 412 in aspecimen 157 mm. long.
Alt.hough the adult (fig. 1) is a deep-bodied,st.rongly compressed fish, the young (fig. 4, c) isslender, minnowlike, and nearly cylindrical. Themaxillaries on young specimens have a few minute teet:.h on t.he lower edge which are lost wit.hage.. The highly specialized digestive tract of theadult is also lacking in the young, which havean almost st.raight intestine and no pyloric eaeca.By the end of the first summe.r, however, theYOl'mg possess the specializations of the adult:
No external characteristics will reliably distinguish the sexes.
Colo·I'.-In life the body is silvery bluish overthe back and upper sides but milky white on theabdomen, and often has brassy or golden reflections from the scales. There are six to eight.horizontal, dark stripes along the upper sidesabove the level of t.he middle of the shoulderspot, extending from behind the head to the baseof the cltudal fin. The large, round dark spotbehind the opercle., so prominent in the young andhalf-grown, is lustrous purple. In adults, thedorsal fin is nearly uniformly dusky; the caudalfin is dusky but. darkened on·its outer third; the
- outer two-thirds of the anal fin is dark, the basalthird lighter, with melanophores sprinkled overmost of the fin; the pectorals and pelvics havetheir out.er halves darkened, paling basally. Thetop of the head, snout., upper jaw, and upper pa.rtof the opercle are pigmented in young and adult;the rest. of t.he head is silvery. In young-of-theyear (up to about 4.5 inches, total length), thedorsal fin is sparsely but. uniformly sprinkledwith chromntophores; the caudal fin ~s similarbut has more pigment cells; the anal, pectoral,and pelvic fins are almost unpigmented. A goodcolor plate of the species is given by Forbes andRichardson (1920, opposite p. 46).
Va:rid.tion..-Insufficient data concerning certainvltriations in this species led to t.he recognitionof at least three nominal subspecies; namely,D. c. ce.pedia:ml'ln (LeSueur), on the Atlanticslope southward and west.ward along the Gulf ofMexico; D. c. hete?'1.ffuv/. (Rafinesque), in themiddle and upper parts of the Mississippi Riversystem and the Great. Lakes-St.. Lawrence watershed; and D. c. err.ile .rordan and Gilbert., incoastal streams from Texas to northeasternMexico. As Jordan (1882: 871) stated, "The
528919 0-60-2
difference between hete1'UrU'fI'b and cepedia:n-l6m isnot great., the grettter arch of the back in cepedianwn being the main difference." It wassoon recognized that this minor feature does not.characterize these inland populat.ions of gizzardshad. The form described from Texas (Galveston) has persisted much longer in the literature (e.g., Fowler 1945: 22, 366). This nominalsubspecies, very briefly and inadequately described, was based solely' on the supposedlyslenderer body (body depth about 2.9-3.3, ratherthan 2.5:-2.7 in specimens from Indiana and SouthCarolina), whence the Latin word exlle, meaningslender. Unfort.ulUttely, t.he two type specimens(USNM No. 30913) have not been found. Although I have not. examiiled specimens· frol~lGalveston, the ratios of body depth to standardlengt.h (table 1) demonstrate that.. subspecies ofDoro80'1na. cepedianum· cannot. be recognized onthis basis.
TABLE l.-Variation in body depth in regional sample-8 ofDorosoma cepedianum
Ran~ein NumberArt'a stan ard Ratio Average of specl·
length mens(mm.)------------
Maryland to North Carollna____ . 61-211 2.3-2.95 2.65 26Middle and npper Mississippi
R.-Ort'at Lakes .~tems______ 78-215 2. 31>-2. 8S 2.62 36Western Florida to uisiana____ 84-204 2.5-,'1.1 2. 69 20Texas. __ ._. _____________________ . 56-196 2. 45-2. 9 2.69 29Northeastern Mexlco _____ ._ .. ___ 81-227 2. 4-2. 9 2.68 12
NOTE.-The depth was stepped Into the standard length .by nslng finedividers under magnillcation.
The dark shoulder spot (fig. 1) is said to disappear with age but is variably developed indifferent populations. This spot, present in youngand half-grown, is typically seen in small adultsand frequently in large ones. For example, thenominal species Olwloe881t8 lnfJoaiahllis Abbottwas based in part. on the retent.ion of this marking in large adults. This spot was well developedin a freshly preserved series of adults, 165 to204 mm. in standard length, from the EscambiaRiver, Florida.
Study '7Iwte1'ial.-The proportions used in thedescription are based on 74 specimens, between 78and 2+.7 mm. standard length, representing localities in Florida., Alabama, Mississippi, Louisiana,Texas, and northeastern Mexico, and from Oklahoma, Arkansas, Tennessee, Missouri, Kansafil,Nebraska, Iowa, Illinois, India.na, Ohio, and Michigan. Specimens from Atlantic coastal waters

376 FISHERY BULLETIN OF FISH AND WILDLIFE SERVICE
are not represented in the general description,but they were used in a study of variation in bodydepth. Counts of fin mys, scales, and ventralseutes 'are based on 51 to 200 specimens distributedfrom Michigan and Maryland southward andwestward to Tamaulipas, San Luis Potosi, andQuerptaro, Mexico. Vertebral counts are basedon 42 specimens from localities scattered over therange of the spedes.
Reproduction and de·l'elop11wll.t.-Spawningtakes place in fresh water (Gunter 1938: 71)from late winter (mid-March) through most ofthe summer (at least to August 20) in sloughs,ponds, lakes, and large rivers. A group of malesand females swimming near the surface beginto roll and tumble about each other in a mass, theeggs and sperm being ejected during this activity.The sticky eggs slowly sink to the bottom ordrift with the current, readily becoming attachedto any object t.hey may contact. At times thesmall eggs cover aquatic vegetation, particularlystreamers of green algae adhering to rocks, forcing the plants to the bottom of the pond or river.Gizzard shad generally spawn on a rising temperature although on one occasion Trautman(personal communication) found them spawningon a falling temperature at Buckeye Lake, Ohio.Langlois (1954:: 224) observed gizzal:d shadspawning along shore, at a depth of 6 to 12 inches,on May 29, 1935, in North Reservoir, Akron,Ohio; when oviposition occurred (67° F.) afemale was flanked on each side by a male.
The bulk of the populations that inhabit thewarm to temperate waters of the United States(28° to 41° N. Lat.), spawns during April, May,and .June at temperatures between about 50° and70° F., the onset of spawning varying with theseason. For example, in exp~rimental ponds atAuburn, Ala., gizzard shad hatched at the endof April in 194:1 but first appeared in the middleof March in 1942; in 1941 the last brood hatchedon August 20 but in 1942 hatching continued onlyinto July (Swingle 1949: 53). tn ChickamaugaReservoir on the Tennessee River, a few milesabove ChaUailOoga, 'Tenn., most individuals hadspawned by the last week of May in 1942; allndults eXllmined on .June 9-10 had spawned, abouttwo-thirds of the females studied .June 1-5 werespent, and very few adults had spawned priorj'o May 22 (Eschmeyer, Stroud, and .Jones,1944: 96). In Norris Reservoir, Tenn., shad
spawned in 1943 between May 18 and June 8,when surface temperatures varied from 73.5° to81.7° F.; in H/44 most of the spawning took placebetween May 15 and .June 1, with the temperaturefrom 78° to '84° F. (Dendy 1946b: 121). Gonadswere ripening near the end of January 1954 inLake Panasoffkee, Fla., with the bulk of spawninghtking place in March and April and completedin May (Moody 1957: 30,38). Near the northernlimit of its range, in Fort Randall Reservoir,S. Dak., all adults caught after .July 6, 1955, hadspawned and most of those obtained before.June 24 were green, indicating a short and comparatively late spawning period (Shields 1956:30) . Bodola 3 fmmd that. water temperatureduring development of the eggs is proba,bly moreinfluential in determining the time of peak spawning than is the water temperature immediatelypreceding the spawning period.
At Buckeye Lake, Ohio, a shallow impoundment in the Ohio canltl system, the gizzard shadbegins to spawn when the water warms to 60° F.,usually during the first part of May but varyingfrom year to year. The spawning period normally extends over about 2 weeks, with an occasional ripe female found later in the season(one was taken in the latter part of July)(Warner 1941 : 639; observations made 1938-40).
In Iowa, the species is reported to spawn inlate April or early May (Harlan and Speaker,1956: 60). In the vicinity of Greenwood, Miss.,It gravid female was taken in .June 1925 (Hildebrand and Towers, 1928: 114). In ChesapeakeBay, Md., the gizzard shad spawns in "earlysummer" (Hildebrand and Schroeder, 1928: 107).Ripe males and females were recorded from thecentral part of the Illinois River, Ill., during May(Forbes lmd Richardson, 1920: 47). A springspawning migration (dates not given) up theMississippi River is reported by Gowanloch(1933: 215).
The embryology and early life history of thegizzard sha.d have been studied by Edward N.'Warner, and reported in abstract ('Varner 1941).Dr. 1Vnrner has kindly permitted me to use matel'iltl fl'om his doctoral thesis that did not appearin the abstrli.ct, including drawings of the embryonic and larval stages (figs. 2-4).
• Bodola. An tllony. The IIf", histary of the gizzard shad, Doro80ll/.{I- cepe(lianu.m (LeSueur). ill western Lake Erie. Ph.D. thesis. Ohio State Univ",rsity. 1955: i-xl, 1-130. figs. 1-44.
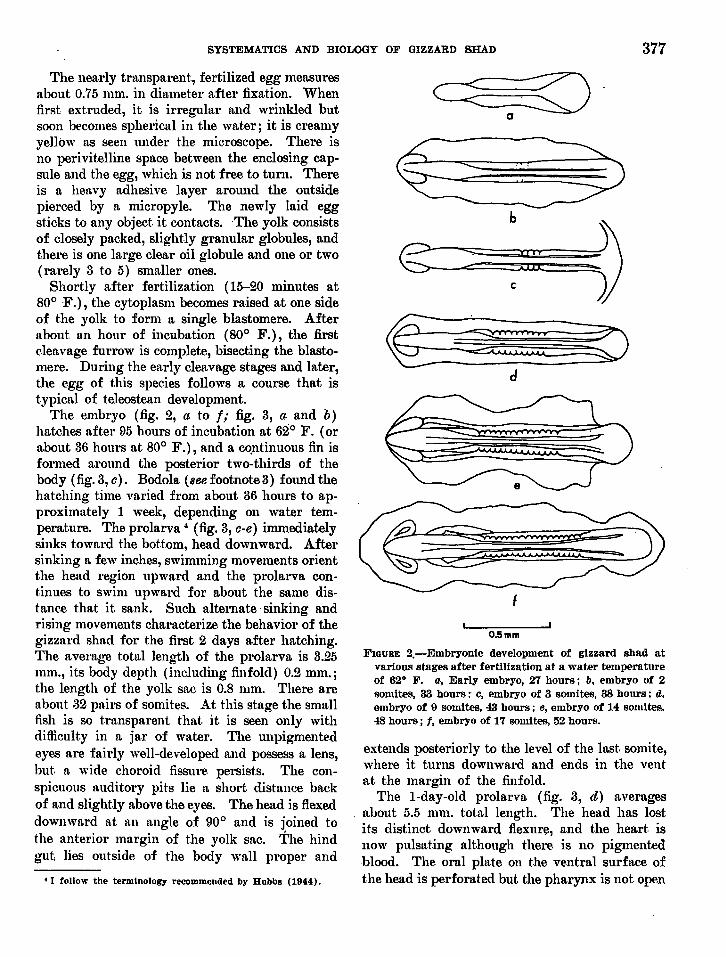
SYSTEMATICS AND BIOLOGY OF GIZZARD SHAD 377
The nearly transparent., fertilized egg measuresabout 0.75 mm. in diameter after fixation. Whenfirst ext.ruded, it is irregular and wrinkled butsoon becomes spherical in the wat.er; it is creamyyellow as seen under the microscope. There isno perivitelline space between t.he enclosing capsule and the egg, which is not free to turn. Thereis a helLvy adhesive layer around the outsidepierced by a micropyle. The newly laid eggst.icks to any object it. contacts. -The yolk consistsof closely packed, slightly granular globules, andthere is one laTge clear oil globule and one or two(rarely 3 t.o 5) smaller ones.
Short.ly after fertilization (15-20 minutes at80° F.), the cytoplasm becomes raised at one sideof the yolk to form a single blastomere. Afterabout an hour of incubation (80° F.), the firstcleavage furrow is complete, bisecting the blastomere. During the early cleavage stages and later,the egg of this species follows a course that istypical of teleostean development.
The embryo (fig. 2, a to f,. fig. 3, a· and b)hatches ltfter 95 hours of incubation at 62° F. (orabout 36 hours at 80° F.), and a co.ntinuous fin isformed around the posterior two-thirds of thebody (fig. 3, a). Bodola (Bee footnote 3) fmmd thehatching t.ime varied from about 36 hours to approximately 1 week, depending on water temperature. The prolarva 4 (fig. 3, a-e) immediatelysinks toward the bottom, head downward. Aftersinking a few inches, swimming movements orientthe helLd region upward and the prolarva continues to swim upward for about the same distance that. it sank. Such alternate' sinking andrising movements characterize the behavior of thegizzard shad for the first 2 days dter hatching.The average total length of the prolarva is 3.25mm., its body depth (including finfold) 0.2 mm.;the length of the yolk sac is 0.8 mm. There areabout 32 pairs of somites. At this stage the smallfish is so transparent tluLt it is seen only withdifficulty in a jaT of water. The unpigmentedeyes are fairly well-developed and possess a lens,but a wide choroid fissure persists. The conspicuous auditory pits lie a short distance backof and slightly above the eyes. The head is flexeddownward at au angle of 90° and is joined tothe anterior lTIltrgin of the yolk sac. The hindgut lies out.side of the body wall proper and
• I follow the terminology recommended by Hubbs (1944).
€ ii35Je ~ :~ ~~ ::3'E:§EE
d
f
O.5mm
FIGURE 2.-Embryonic development of gizzard shad atvarious stages after fertilization at a wate-r tempe-ratureof 62° F. Q" Early embryo, 27 hours; b. e-mbryo of 2somites, 33 hours ~ C, embryo of 3 somites, 38 hours: d.embryo of 9 somites, 43 hours; e, embryo of 14 somites.48 hours; f. embryo of 17 somites, 52 hours.
extends posteriorly t.o the level of the htst somite,where it turns downward and ends in the ventat the margin of the finfold.
The I-day-old prolarva (fig. 3, d) averagesnbout 5.5 mm. total length. The head has lostits distinct downward flexure, and the heart isnow pulsat.ing although there is no pigmentedblood. The oral plate on the ventral surface ofthe head is perforated but the pharynx is not open
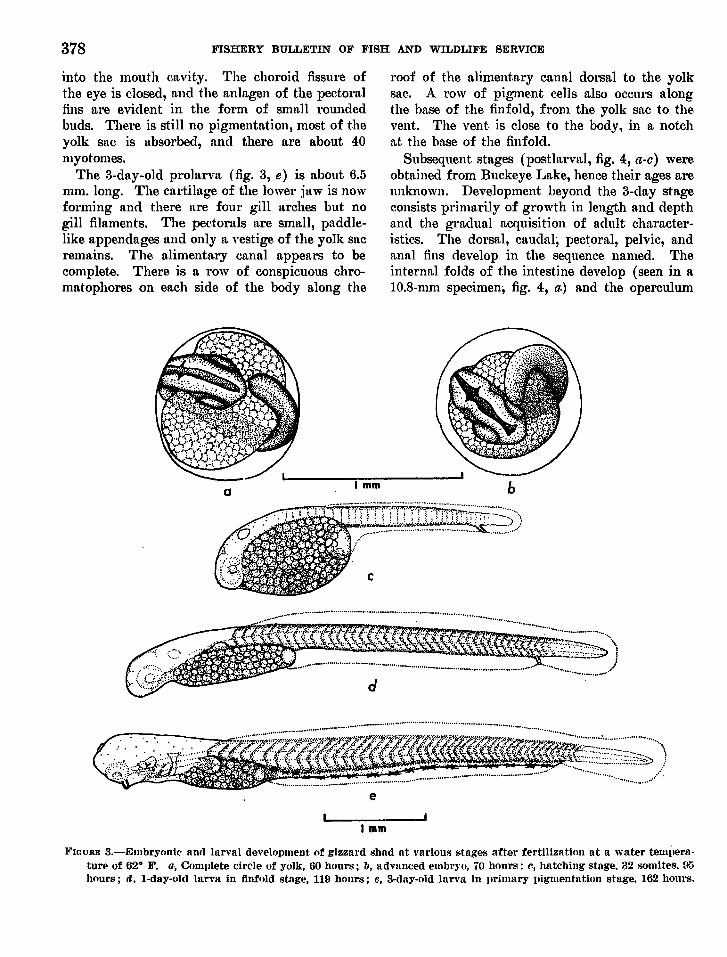
378 FISHERY BULLETIN OF FISH AND WILDLIFE SERVICE
into the mouth Cl"lVit.y. The choroid fissure ofthe eye is closed, and the anlagen of the pectoralfins are evident in the form of small roundedbuds. There is still no pigmentation, most of theyolk sac is absorbed, and there are about 40myotomes.
The 3-day-old prolarva (fig. 3, e) is about 6.5mm. long. The cartilage of the lower jaw is nowforming and there are four gill arches but nogill filaments. The pectorals are small, paddlelike appendages and only a vestige of the yolk sacremains. The a.Iimenta.ry cana.I appears to becomplet.e. There is a row of conspicuous chromatophores on each side of the body along the
roof of the alimentary cana.I dorsal to the yolksac. .A row of pigment cells also occurs alongthe hase of the fin fold, from the yolk sac to thevent. The vent is close to the body, in a notchat the base of the finfold.
Subsequent stages (postln,rval, fig. 4, a-c) wereobtained from Buckeye Lake, hence their ages areunknown. Development beyond the 3-day stageconsists primarily of growth in length and depthand the gradua.I acquisition of adult characteristics. The dorsal, caudal; pectoral, pelvic, andanal fins develop in the sequence named. Theinternal folds of the intestine develop (seen in alO.S-mm specimen, fig. 4, a.) and the operculum
b
c
I mm
-~1~;:~5)../
a
..............................: '.',o .~:.••: .
.... ;.:::::: ..:.::::.:::::..:;:.:..::::::<.~~~~:) .)...... ................................
e
Imm
1·'WURE 3.-Embryonie aOfI larval development of gizzard ,,;had at various stages after fertilization lit a water telllilel'ftture of 62° F. a·, COllllllete circle of yolk, 60 hours; 11, advllnced embr~·(.•, 70 hours: C, hatching stage. 32 somites. 95hours; fl. I-day-olc1 larYa in finfold st.nge, 119 hours; c, 3-day-old larva in llrilllary pigmentation stage. 162 hours.

SYSTEMATICS AND BIOLOGY OF GIZZARD SHAD 379
grows posteriorly to cover tIH~ gills. The auditorypi~ becomes invisible from the surface by theovergrowth of the head skeleton and lm,lsculature.The pigmentation becomes niore prominent, especilllly nlong the dorsal surface and on the dorsaland caudal fins. Bodola (see footnote 3) was \lll
able to rear the young beyond the 10th dlly afterhatching, at which time his fry were slightly moret}:l.an 6 mm. long.
The lnrvae of the gizzard shad may be distinguished from those of fishes with which theymay be confused by the long gut (1.5 to 2.0 t.imesthe total length of the fish), the retarded development of the single dorsal fin, nnd the elongated anal fin (22 rudimentary rays at 17.5 mm.,30 to 34 at 10 to 22 mm. total length ) .
FeC'lmd-ity.-In his study of the gizzard shadin Lake Erie, Bodola (see footnote 3) found considerable varilttion in the estimated number ofeggs per individual female. Two females of agegroup 1, averaging 231 mm. in standard length,had an average of 59,480 eggs; 5 females of agegroup II, 291 mm. long, averaged 378,990 eggs;3 individuals of age-group III, averaging 331mm. long, had 344,780 eggs; 2 in age-group IV,356 mm., avern,ged 308,750; and 1 female of agegroup VI, 355 mm. long, had n,n estimated 215,330eggs. Although meager, the data show that precocious shad have few eggs, that the II-groupindividuals have the most eggs, and that egg production declines with successively older groups.Fish weighing between 500 and 600 grams
FIGURE 4.-Larval development of gizzard shad collecled in plankton net, Buckeye Lake, Ohio. a, Eye-pigment s'tage.length 10.8 111m., age unknown; b, pelvic-fin stage, 17.5 mID., age unknown; 0, definitive-fin stage, 22.0 nUll., ageunknown.
528919 0-60-3

380 FISHERY BULLETIN OF FISH AND WILDLIFE SERVICE
I Based on number of annuli.
----1-------------------
(From Lagler and Applegatc. 1943: table 3J
Total length June July August Septem- 'Septem- Octoher(mm.) 24-30 I 6-21 ' 3-21) 3 ber 2 ber 28-29 'rl
1,0552.4461, 847
····--;ii- :::::::::: ::::::::::31 22 1
183 162 3464 256 22478 297 92336 481 224161 604 362112 437 21453 118 7611 52 314 11 18
4 72 2 41 _ 1
1,1132,546TotaL _ 2.233
I Five samples collected on June 16. 24 (3 samplesl. and 30., Four or the six samples taken we7C collected on July 7.3 FOUf collections made on August 3, 12. 19. and 20.
ItemAge group I
0 I II III IV V------------
Number of specimens___ ._ 242 38 84 81 25 6Size range (mm.l. ._ ... _.. 1!l-79 130-1SO 172-222 180-238 199-250 230-309Size fange (in.). ___ ._ ...... ------ 7-9 9-11 !l-12 10-13 11-15Average standard length
{mm.\. _....... _.... _._. 41 152 193 207 223 256Average totallcngth (In.I . ---~ -- 7.6 9.7 10.5 11. 3 12.8
TABLE 3.-Length distribution of 11,240 ,I/oung-of-the-lleargizzard shad in. Chicka.mauga Reservoir, Tenn., 1942
[From Eschmeyer, Stroud. and Jones, 1944: table H)
At the end of the first year of life in Fort.Rltnc!:tll Reservoir, S. Dak., the average totallength of the 1954 age group was 5.1 inches (170specimens) and of the 1953 a.ge group 7.0 inches(Shields 1956: 30). Somewhat greater averagelengt.hs are given for samples from Black River,Mo., by Plttriarche and Lowry (l953: 99-105).
The gizzard shad t.ypically matures in its sec'OIul or third year and lives for at least. 7 years,in OklahOll1lt (.Tenkins 1953a: 54) and Lake Erie(Bodola; Me footnote 3, p. 376). Pat.riltrche(1953: 249) found lO-year-old gizza.rd shad inLake 1Vappapello, Mo., but. stated tha,t in ot.herMissouri reservoirs t.he spec.ies lives no longer than5 or 6 yea,rs. Gizzltrd shad in La.ke Newnan, Fla.,average about 10 inches (totn.llength) at the endof their first year of life, about. 12.5 inches at t.heend of the second year, aml about 13.6 inches at
TABI,E 2.-Age of giZ%a~d shad and average lengths of fishin each age group in two Indiana ponds. June 26-AllgW.t28, 1940
21-26.. _.... 123 7 .. _. __ _ _........•... _..... _..'rl-32 _ 202 26 .. .. _....•. _ .. __ ._._ .•33-38 __ ..• _. 366 509 1 ..•. _._ __ . •39-14....... 546 767 10 _. __ - _. __45-50 _ 336 269 5851-56 _.. 221 213 12657-62. __ 207 187 15463-li8 __ __ 123 115 1576!l-74 _.__ 74 122 16575·~10 . 'rl 166 13581--86....... 7 105 12987-92 _ ._ .. __ 44 7793-98 .. __ 1 11 599!l-104 __ • __ . _. ..•. _ 3 26106-110..... .......•.. 2 13111-116.. _._ __ ......•. _...... 2117-122_ .. _ _._ _ _.. __ I123-128_ __ - . __ __ ..129- . __ . .
produce the most eggs. A ripe female 315 mm.long, taken in June 1925, nen.r Greenwood, Miss.,contained n.pproximately 50,000 eggs according toHildebrand and Towers (1928: 114).
Age and gl'o'wth.-Data on the age and growthof the gizzard shad Imve been summarized recently by Ca.rlandel· (1950: 24-25; 1953: 282--283)and further treated by subsequent authors. Thefollowing summary is taken in large part fromthe original papers cited therein.
In Foots Pond, Ind., growth of young-of-theyear, as noted ill samples taken on four successivedates in 1940, was as follows (standard lengthra.nge, in mm., followed by average length):19.0-44.5, 31.4 (71 specimens, June 26); 32.065.0, 57.0 (29, July 10); 45.0-79.0, 66.6 (42,August 20); and 61.0-88.0, 72.4 (70, November 13). Growth is very rapid in the first 5 to
. f\ weeks of life and gradually tapers off 'as theseason progresses (Ln.gler and Applegnte, 1943:104-105). At the end of the' first summer, anaverage total length of about 4 inches is attn.ined(Indiana and Ohio); total length is about 4.5inches in the Chesapeake Bay region, and 5.0inches in Tennessee and Oklahoma.
A summary of the age and growth of D01'O.~Ol1Ut
in Foots Pond lind Grassy Pond, Ind., as determined from samples collected between June 26and August 28, 1940, is given in table 2. Studieson gizzard shad from the Chickama.uga Reservoir, Tenn., in 1942, showed that I-year-old fishhad n. modal length of 7 inches in June and about7.5 inches by fall; 2-year-old fish were about. 8.5inches long in April; and t.he largest shad wereabout Hi inches long. Young shad (born in latP.Mayor early .June) showed an average lengthof npproximately 1.5 inches by the last week in.June, 2.5 inches by mid-August, and 3.5 inches by"late September (table 3). A summary of growthin Grand Lake, Okla., over a 13-year period(.Tenkins 1953a: 53), showed that here, as atHerringt.on Lake, Ky., first-year growth is onlyabout. 4.5 (rather than 7) inches, thus ext.endingthe time during which gizzard shad are availableas It forage fish. The a.verage length of gizzardshad in Crab Orchard Lake, an artificial impoundment. in southern Illinois, was only 4.0inches at the end of the first year, 5.4 inches atthe second year, and 6.6 inches at t.he end of thethird year (I"ewis 1953). .

SYSTEMATICS AND BIOLOGY OF GIZZARD SHAD '381
the completion of the third year-thus indicatingrapid growth coupled with a very short life Spllll.Only about 5 percent approach their third actua.Iyear of life and none over 4: yeRrs old was found(Berry 1958). .
From 70 to 80 percent of the annual growth ofgizzard ·shad in Lake Erie tu.kes place during.June, .July, and August or .July, August, andSeptember, depending on the season u.nd the ageof the fish. The species grows lit.tle if at allduring winter, when considerable loss of weight.occurs (Bodoln.: .see footnote 3).
There wns no sexua.I dimorphism in the rate ofgrowth or in the lengt.h-weight relation for 1,136gizzard shad from Beaver Dam Lake, Ill. (LagleI'and Van Meter, 1951: 357~'360).
Abnornutl growth has been reported by Hubbsand 'Whitlock '(1929), who found two extremeforms in samples taken not far apart in the Arkansas and Poteau Rivers, Okla. The abnormalsample, composed OJ,lly' of young fish, showed ateratological condition (head long, wide, andswollen; eyes In.rge; t.ails stunted) that was evideiltly related to an environmental factor orfactors.
8·ize.-The gizzard sh:ld is known to attain atotal length of 20.5 inches but does not commonly grow longer than 10 to 14 inches. In aletter dated .July 14, 1953, Alfred Larsen (Fishery Biologist, Pennsylvania Fish Commission)wrote that. specimens of Dm·o.smn((. cepedia:YlJltmkilled in Pl'esque Isle Bay early in 195:3 (see section on Mortality) ntried in length from 4 to 19inches. Mnturity may be attained at muchsma.ller sizes than commonly thought, for VIn.dykov (1945: 35) reported a mature female to be151 mm. (a little over 6 inches) in total length.Fish 10 to 13 inches long weigh about 1 pound;individuals 14 to 18 inches long, from the OhioRiver c1ra.inu.ge or Ohio, weigh 1 to 3 pounds;and the largest recorded specimen (20.5 inches)weighed 3 pounds 7 ounces (Trautman 1957:182). As in other fishes, size and weight varyconsiderably and in some localities there ismarked dwarfing.
llabltrtt ((:lId ·m.ig·l'a.tion.-The gizzard shad inhabits large rivers, reservoirs, lakes, swamps,b:lyS, borrow pits, bayous, estuaries, temporaryfloodwater pools along lar~ river courses,sloughs, and similar quiet open waters whichmay be dear to very silty. Although it is fairly
common in the relatively strong current of theupper Mississippi River, it prefers quieter waters and swarms in the sluggish lower parts ofthe same river. The adults may ascend smallerstreams or ditches to spawn and the young arelater abundant in such pla-ces if the gradient issufficiently low. In coastal rivers of Virginiathe young are present in great abundance wellupstream from brackish water (Massmann 1953).In Lake Erie the species is most plentiful at theshallow western end, over mud bottom (Nash1950: 563), particularly in protected bays andabout the mouths of tributaries (Bodola; seefootnote 3). Abundance is greatest in late summer and early fall when the populations areaugmented by young-of-the-year. The species isparticularly attracted by warm water flowingfrom industria.I plants (see section on Mortality).It is able to withstand rather high temperatures,to 35° C., and has shown no geographic variation in its a.bility to tolerate lethal temperatures(Ha,rt 1952: 28-29).
Gizzard shad a·re common in Chesapeake Bayonly during the fall months, occurring principa.Ily in brackish water near the mouths of freshwate.r. streams. The species is common or abundant in the rivers of the region throughout theyear, but very young individuals evidently do notenter brackish water (Hildebrand and Schroeder,1928: 106-108). Along the Texas coast, the species frequents the large, brackish-water bayswhere individuals may be taken throughout theyear in' ,,'aters that vary in salinity from 2.0 to:m.7 parts per thousand. Gizzard shad in thisarea prefer brackish water to sea water and, ingeneral, the sma.Ilest fish occur in the freshestwater, with size increasing as salinity increases(Gunter 1945: 30~'31).
DOl'o.solll(l· cepedia1l1t:1n is essentially an openwitter spedes, usually living at or near the surface, and the young are reported to prefer bedsof spatterdock (Nnph(M') in Foots Pond, Indiana (Hubbs and Lagler, 1943: 77). In westernLake Erie, young shad live close to shore in midsummer, usually in shallow water (Bodola; seefootnote :~). If the oxygen supply is adequate,the species may descend to depths as great as 108feet, as in Norris Reservoir, Tenn. (Cady 1945:113-114; Dendy 1945: 126; IV46a). In the CoosaRiver, Ala., gizzard shad were found in deep (25

382 FISHERY BULLETIN OF FISH AND WILDLIFE SERVICE
feet) as well as shallow water (Scott. 1951: 38:39).
Young-of-t.he-year gizzard shad t.ravel in compact schools soon after hatching, but by fall mostof the-schools disperse and few form the following spring, at least in Norris Reservoir; schooling hugely ceases by t.he time the shad are ayear old (Dendy 1946b: 121). In the Chesapea.ke Bay region there is a fall "run" in .September and October, and a corresponding spring"run" has been recorded in North Carolina. Aspring migra.t.ion, evidently a spawning run, hasbeen noted in the Mississippi River near NewOrleans, anel lake populations migrate to shallower water at spawning time. An unusual midwinter migration was noted in t.he MinnesotaRiver, from about December 1 until late January (Swanson 1932: 34).
Food.-Except for a short time aft.er hat.ching, t.his species is almost. entirely herbivorous,feeding heavily on microscopic plant life, phyt.oplankton, and algae.
At birt.h the alimentnry canal is a simple,nearly straight. tube (fig. 4, a) but by t.he timethe wild fish is about. 22 mm. long four flexureshave developed; the sect.ion between the first andsecond flexures subsequently enlarges to becomethe gizzard and caeca. a.rise from t.he duodenumby the 27.5-millimeter stage. The intestine becomes t.remendously convoluted with age; whereasit equals about. one-half the total length of theday-old fish, it becomes three times the length ofthe adult and is packed int.o an abdominal cavity only one-third that of the fish's length. Theyoung commence t.o eat about 4 or I) days afterbirt.h (Bodoln; flee footnot.e 3). Wier andChurchill (1946) have described the anatomy andhistology of t.he digestive system.
During the first few weeks of life, gizzard shadeat mai.nly protozoa and entomostraca. Contentsof the digest.ive trncts of Ia.rva.l and postlarvalshad examined by 'Warner (see Wickliff 1945:1--4) consisted almost entirely of water fleas(Bormtina.) , copepods, and a few ostracods. Theseorganisms were found in young up to about 22mm. long. Individuals 26 mm. long, with theintestine, gizzard, and liver well developed andhaving essent.ially the appearance of the adult,cont.ained largely algal plankt.on (diatoms, desmids, and colonial forms) and shelled nnd flagel-
lated protozoa (Euglena), which occur in the bottom ooze, on veget.ation and as free-floatingorganisms. At this size, Bosrntna has almostdisappenred from the diet. Food may occur inthe plllwyngea.I pockets, t.he gizzard, and the intestine (Bodolu; see footnote 3).
The algal consumpt.ion of t.he gizzard shad ledTiffnny (1922: 285) to describe these fish as"living t.ow nets." He ident.ified 150 species andvarieties of nonfilamentous algae from t.he stomach nnd intestine of specimens taken in streamsand ponds of Ohio and Illinois. To what extent. these are utilized as food is questionable,however, for Velasquez (1939) showed experimentally that 50 of the nbove species might havebeen viable after passing t.hrough the digestivet.ract.. A cert.ain amount of mud is typically ingested by the shad while feeding (hence thename, mud shad), but this material is evidentlyt.aken accidentally; however, sand may be purposefully inge.c;ted as an' aid in the maceratingaction of the gizzn.rd (Bodola.; see foot.note 3):On occasion, the species may be canniba.listic(Dendy 1946b: 119).
Earlier Tiffany (1921: 383) gave a summaryof the food eaten by young sha.d in six Ohiolakes. This consisted of microalgae, 70 to 90percent; microanimals, 0 to 15 percent; filamentous algae, 0 to 5 percent; and plant. debris, 5 to20 percent.. Mud usua.lly comprised from 10 to30 percent of t.he st.omach a.nd intestinal contents, but it was often absent. The gizzard shadis able to ut.ilize a large variety of microscopicplants and it does so consistently. Moreover, thediet is remarkably alike a.t various sizes (oncethe early zooplankt.on st.age is passed). Tiffany found that the percenta.ge composition offood eaten by shad 200 mm. long was not materia.lly different from the data given above foryoung fish, except that there was more unrecognizable debris. Examination of intestinal contents in samples from La.ke Erie showed" thatlitde food is eaten in winter amI early spring(Bodoln; see footnote 3).
In the vicinity of distilleries this species willfeed on distillery slops (cornmeal) and it mayalso occasionally take Coleoptera, univalve mollusks, young 001ivu, and spiders and water mites(Forbes 1888 : 438; Forbes and Richardson,1920: 47).

SYSTEMATICS AND BIOLOGY OF GIZZARD SHAD 383
Seven specimens from Buckeye Lake, Ohio, 22to 31 mm. in total length, taken on June 25 and.July 11, 1930, yielded 14 percent Cll1docera and86 percent algae and debris (Ewers and Boesel,1936: 61). Some of these individuals were inthe transitional stage during which zooplanktonis being replaced by phytoplankton in their diet.
Predatol's.-At a certain stage in its life history, the gizzard shad is reported to form a major part of the diet of at least 17 important gamefishes. Of value in this respect." are young-of-theyear about 2 to 5 inches long, but individuals 8inches or longer are utilized to some extent bywalleyes (8tlzostediml. '1.'itreum) , according toStroud (1949). In ponds and lakes of Ohio, itis preyed upon by white bass (Boccus chrysops),largemout.h bass (JJlic'ropte1'us sulmoides) , andwhite crappie (Po'l1!.oi!!is a.nnul.aris) (Wickliff andTraut.man, 1931: 15, 25, 37). In Foots Pond,Ind., the gizzard shad was eaten by three gars(Lepi808te'/{J5 phttosto'm!/t~, L. pI'Od1IrfU.S," and L.
OSSe1f.s) , white bass, largemouth bass, white crappie, lI.nd hlack cru.ppie (PO'1noi!!is ·nigl'oma.m.datu.s), according to LagleI', Obrecht, and Harry(1943 : 120, 122--123, 126) , and LagleI' andRicker (194::i: 59-62). Two gars, L. pl'odu.ct·uslind L. uJorse'Il.s, utilized the speeies in Texus (Bonham 1941: 359-::i60), and the skipjack (Alos((.r:hl·Y8ochlO1·i.~), north~rli pike (EsoJ.! lv.clus) , yellow bullhead (Ictnl.U'l"!t8 'lUi-taUs), and whitecrappie preyed upon the gizzard shad in Illinois(Forbes 1888: 435; Hansen 1951: 225). In theIllinois Ri,'er, Forbes (1903: 34-38,40) reportedthut walleye (.Stizo.stedion ·!,itl·emn); sauger(8tizoJStedion c,.made·ll.~e), und yellow bass (Roc(:us i'lltM'1'lIptUS) preyed upon shud between 3 and4: inches long and that this species was a veryimportant item' in their diet.
InNorris Reservoir, Tenn., Dendy (1946b :12g, 194) stnt.ed thut D. cepedia:mnn constitutesthe most important food supply for the gamefishes. These are channel catfish (Ictal!i-l'us
pUilldatu.~), largemouth bass, smallmouth bass(lllic'l'optel'lts dolom.ieu-i) , spotted bass (JJ/. punctulatu8), bluck erappie, walleye, sauger, and thefresliwuter drum (Al17odinotus g·l'wnnie'lls). Int.he Clinch River, Tenn., white bass and h1l'gemouth bass fed mostly on gizzard shad nnd brooksilversides (Eschmeyer 1944: 38).
Even the lake trout (Sa1.1'elinll.s '1Utl1Utycush)is credited with eating Dm'o8oma. in Cayuga Lake,New York. An examination of 312 stomachs thatcontained food revealed one with a gizzard shad6.7 inches long. ll The bowfin, A.1nia cah'a., alsois known to eat gizzard shud (LagleI' and Hubbs,1940).
The periodic morhtlity (see elsewhere) of gizzard shad provides an important source of foodfor numerous species of wat.erfowl. This foodcomes at a most. opportune time, when otherfoods are scarce or when waterfowl are forcedout of their normal feeding plaees by hunters.Young-of-the-year shad are fed upon at BuckeyeLake, Ohio, during the fall and winter by thelesser loo'n, horned grebe, pied-billed grebe,' whitepelican, great blue heron, American egret, snowyegret, eastern green heron, black-crO\vned nighthel'On, Americ:lll merganser, red-breasted merganser, hooded merganser, mallal;d, black duck,gadwall, bnldpate, green-winged t.e.a.I, blue-wingedteal, American pinta.il, wood duck, redhea.d, canvnsback, lesser scuup, ring-necked duck, Americangoldeneye, buftlehead, oldsquaw, king eid~r, Americun scoter, and whitewinged seoter (Trautman:':W4:0: 110-111, 155-g06). During' the relativelyopen winter of 195~53 at. Columbus, Ohio, Trautman (personal communication) stated that thehuge waterfowl concentrations in central Ohio\,"ould hlwe been impossible without the gizzardshad.
Utiliza.tion.-The gizzard shad is not esteemedfor food by man because of its soff. and rathertasteless flesh and the numerous fine bones. Thereis no evidence thnt the aborigines sought. thisspecies (Rostlund 1952: 14). In the ChesapeakeBay region it once sold fairly well to a cluss oftrade that. demanded a chellp fish. In 1921, theretail price in Balt.imore was about 5 cents It
pound. Among the commercinl fishes of Chesapellke Bay in 1920, it ranked twentieth in valuewith a cn.feh of 72,85g pounds worth $2,O1:~
(Hildebrand nnd Schroeder, 1928: 1(7). Thec:.ommercinl enteh in the Great La.kes O\'e!' It 14year period (tnble 4:) shows the erratic nntllre ofthe take.
"Galligan, Jalll~s 1'.. I'h.. llistribution of luke trout and asso"ill ted species iu l:u)'ugn Lake. M.A. thesis. Cornell Unh·erslty.1951: 72.

384 FISHERY BULLETIN OF FISH AND WILDLIFE SERVICE
I To nearest round ftgure., For State of Michigan only; final ligures for 1957 are approximate.
TABLE -t.-Commercial catch of Dorosoma cepediaullm inthe Great Lakes, 1939-57
[Excluding Lake Superior. From statistical records of U.S. Fish and Wild·lIle Service and Michigan Department of Conservation]
The species has been used to some extent inmaking guano, and in 18i4 a guano factory existed at Black POilit, above Pa.Iat.ka., Florido.(Bean 1893: 64; Goode 1884: 610). Many yearsago, on Lake Erie, it was split and salted andsporndically marketed with other low-grade fishas "lake shad'~ (Jordan 1882: 871), and in the1840's it appeared on the markets in Ohio but.was not highly rega.rded. (Kirtland 1838: 195).
"Then it becomes excessively abundant, as itdid in Black Hawk Lake, Iowa, in 1951, the gizzltrc1 shad may be used as hog food or for fieldfertilizer (Mlt<lden 1951: 185). In the Ohiownters of Lake Erie, the harvest. is limited and isrestricted to Sandusky Bay. Here one commercial drag seiner htkes shad, carp, and goldfishfor use IlS hog feed, and another drag seinerprovides enough D01·OIJOl1Ul. to freeze for trout.food (letter from Robert. Cummins, Jr., Sandusky,Ohio, .Tune 26,1953). In t.he Pennsylvania watersof LItke Erie, where shad mortality has beenhigh in recent. years, the species has been st.eamedIlnd pressed for oil and the remainder used ascattle food. Gizzard shad roe has occasionallyhee,n marketed for food in Florida (Moody 1954:147). Its ~lse IlS food for fur anima1s has beeninvestigated. Since the whole fish contains considerable amounts of the enzyme thiaminase, itmust. be cooked or fed in It special feeding schedule. Protein content. 'of the whole fish is about.
.15 percent. and fat content 12 percent, which is
-----------------
compamtively high (U.S. Fish and Wildlife Service, 1956).
The species hilS had limited use as a bait fish.Evermann (1899: 308) claimed that gizzard shadwere "of considerable importance as bait" in theAtchafa.Iaya River, Miss., but Kuhne (1939: 25)stated that since it dies very.easily it is an unsatisfactory ba.it 'minnow. According to T. H.Bean (1893: 64), the shad. "ho.s been very successfully kept. in the aquarium * * *." It has alsobeen successfully propagated in ponds as food foryoung bass.
Although the gizzard shad can hardly be classedas a sport fish it. is taken occasionally by angling,as at Lake Chautaqua, Ill. A hook baited withan angleworm, a small minnow, or even an artificia.l fly, is attractive to the shad, and Abbott(1861: 366) stnted thnt the fish afforded muchsport to juvenile anglers in New ,Jersey.
Ab-unda:nce.-In recent years, the gizzard shadhas become a problem species wherever it. has soincreased that. a detrimental affect is produced onother fishes. D01'08011Ul. has inhabited Lake Eriefor more than 100 years, yet its greatest abundance there has been nttained since about 1950,according to commercial fisherman (Bodola: seefootnote 3). Fluctuat.ing but generally inereasingnumbers in the Great Lakes, especially LakeHuron and Lake Erie, is indicated by the commercinl catch from 1939 to 1957 (table 4). In1948, 3,000 pounds were taken in one seine haulin Lake Huron, and it was noted t.hat the fishwere, being captured in' record numbers after a15-year lull in abundance (Ann Arbor News,Mich., December 1, 1948). Overpopulat.ion ist.ypica.By associated with manmade modificationof the environment (LagleI' and Van Meter, 1951).
In Carpenter Lake, Ky., during October 1954,285 pounds of gizzard shad per acre were killedwith rotenone in un impoundment of 70 acreshaving an average depth of 5 fe.et. and a maximumdepth of 11 feet; only about 50 pounds of sharIper ucre were anticipated (Bowers 1955). Amongfomge fishes, there was ll- phenOlnenal increase inabundance of gizzard shad in Clearwater Lake,Mo. The pel'cenh\ge composition rose f\'Om 2.0 in1949 (1 year after impoundment) to 57.8 in 1950;large schools of shad were seen in 1950 throughoutthe lake (Murtin and CallJpbell, 1958: 59). Sincethe species had spl"wned before the reservoir filled
Pounds I Value I
300 $18.0035 1.00
300 6.0023.000 139.00
900 44.00300 13.00600 28.00IOO 7.00600 10.00
29,400 294.0031,600 930.0020,600 614.00
4.';0 16.002,300 55.00
100 1. 002.200 32.003, iOO 41.005.180 149.004.750 95.00
126.415 2.493.00
LakesYear
1939_ ____ Huron .1940_ ____ Erie. Huron_. ._ . _1941. • _ Huron . . ._1942 Erie (Pa.)._. . _1943 Erie. Huron.. . . _1944 do ._. - _1945 do . . ._1946_ ____ __ Huron .. . _1947 • Erie, Huron . _1948_ ___ _ Huron .. _1949_ __ __ Erie. Huron. . _1950 do . . . _1951. do .' _1952. do. .' _1953_ ____ _ Huron . _19501 • Ontario. Erie. Huron _1955_ _ Huron. Mlchigan.. . __ . _1956 ' • .do • .1957 , • .do . . __ .. _
TotaL ... _. . .. .. _-.. _

SYSTEMATICS AND 'BIOLOGY OF GIZZARD SHAD 385
in 1948, the tremendous increase in 1950 resultedlargely from brood stock spawned in 1949.
Black Hawk Lake, Iowa, became so overpopulated with D01·OSO'l1W. that the game fishes weresuppressed by sheer weight of numbers of gizzardshad (Madden 1951). In 1934 there was a complete winter kill in this shallow, fertile lake of957 acres; in 1939, 105 acres were dredged so ast.o avoid further severe winter mortality. Thedepth was 6 to 7 feet except in the dredged portion, which was 9 to 16 feet. The first gizzardshad was netted in 1945 and by 1947 (when thelake was flooded by Boyer River) the populationhad reached a nuisance level. With the progressive increase of shad it was noted that bottomorganisms declined. In the fall and winter of1950-51, 10,000 adult DOl'osoma were removedby seining. Largemouth bass, walleye, and northern pike were stocked in 1951 to provide predatorst.o keep the shad in check, but in the fall of 1951llpproximately 7,360 young gizzard shad per acrewere removed from the lake. Nevertheless,through intensive netting during t.he period 195157, the poundage percentage of gizzard shad removed per haul (mostly by a 2,500-foot seine)decreased from 98.1 to 2.8; at t.he same time thepoundage percentage for carp increased from 1.8t.o 96.3-thus suggest.ing that the ecological nichevacated by the shad was rapidly assumed by thecarp (Rose 1957).
Increase of gizzard shad in 31,000-acre LakeApopka, Fla., reached the point (sometime between 1950 and 1956) where t.his species constituted more than 80 percent of the total fishpopulation (Heinen 1958). By means of selectivetreatment wit.h rotenone, an estimat.ed 3.5 millionpounds of DOl'OS011UI, were killed (November 4,1957) in the first of three projected applications.Past records indicated a marked decrease of sportfishing aft.er 1950 and a, change in the lake fromclear water and la.rge quantities of submergedvegetation to turbidity (from high plankt.onbloom) amI sparse submerged plants. Similaroverpopulations of gizzard shad have occurred inother Floridll lakes. For example, in NewnansLake, Alachua County, with 6,182 acres, an esti~
mated 1 million pounds of shad were destroyedwith rotenone (Melvin T. Huish, pe,rsona.l communication). In five of seven slmllow Floridalakes studied by Moody (1954), gizzard shad and
gars constituted about. 50 percent or more of thetota.l weights of fishes obtained by large haulseines.
In comparing fish populations in two similarOklahoma lakes, Jenkins (1957) demonstratedthat the lake containing gizzard shad had anestimated standing crop of 1,043 pounds per acreand a weight of desirable-sized fish of 466 poundsper acre, whereas the lake lacking this specieshad an estimated standing crop of 655 pounds peracre of which '608 pounds comprised desirablesized fish. The average coefficient of conditionfor six species of sunfishes in the .t.wo lakesshowed that the lake without shad was above theState average, whereas that containing Do'rosom,a.was well below the State average. This is indicative of direct compet.it.ion between centrarchids and gizzard shad in small-lake populations.
Pa,Trt8#es.-This species appears to be usuallyfree from attack by parasit.es, from which it isundoubtedly protect.ed by its herbivorous foodhabit.s. Bangham and Hunter (1939: 396) examined 5 gizzard shad from Lake Erie and foundan unidentified larval nematode in the int.estineof one young specimen. Van Cleve found buttwo species of Acanthocephala in 300 gizzardshad, and Essex and Hunter reported no parasites in more than 100 individuals from the Rockand Mississippi rivers (as cited by Bangham andHunter). In Buckeye Lake, Ohio, where gizzardshad are abundant, the young often carry a veryheavy infestation of a myxosporidian whichforms large white cysts in the body cavity. Eachof 15 adult. shad from the same lake were freeof parasites, and 10 of 12 yoimg carried manyof the encysted sporozoans (Microsporidia) described above, often giving them a "potbellied~~
appearance (Bangham 1941).il/O"l'tality.-Many gizzard sha.d die during t.he
winter, frequently in great numbers nVickliff1953). This is a rather regular and often spectacular phenomenon and on occasion may lead toa health problem, as it did when more than 1,000tons of dead and live Do'rosoma. were removedfrom Lake Erie at Erie, Pa., during January andFebruary 1953 (Walsh 1953: 2-4, 5 figs.). Themass morta.lit.y, which occurred in Presque IsleBay, was t.he greatest. in It sequence of similardellths that took place there during the decadeHl43 to 1953. The blty contains two basins, East

386 FISHERY BULLETIN OF FISH AND WILDLIFE SERVICE
and 'Vest, each of which has Rn area of about15 acres, with a maximum depth of 8 to 10 feet.The pumping of water from the East Basin t.othe 'Vest Basin by an electric power companyresulted in a, discharge of wltrmed water into the'Vest, Basin at It rate greater than 100,000 gallonsper minute. At the time of the mllSS deaths,water in t.he 'Vest Basin varied from 55 to 60° F.,whereas that in the East Basin and the bayproper was only 34-35° F. Dissolved oxygen wasnot low and, wit.h one exception, carbon dioxidelevels were not unusunl. The mortalit.y occurredin the wn·rmer 'Vest. Basin and was dearly associated with high temperature. In 1V50, the previous record year, 800 tons of shad were removedfrom the same basin (I am indebted to AlfredLarsen, Fishery Biologist, Pennsylvania FishCommission, for most of the foregoing information) .
The cause of such mort.a.lit.ies is not completelyunderstood. It has been suggested that a markeddifference between the carbon dioxide t.ension ofthe surface water and the carbon dioxide part.ialpressure of the air is a causa.I factor (Powers1938: 279) .. This has been disputed, for exampleby Ba.ker (1942: 48), and it is generally felt thatabrupt. temperature changes cause the mass deathsof this species (see also Agersborg 1930). Thisfactor was noted more than 100 years ago byKirtland (1844: 237), who described a mass mortality of DO'/,ofJ01na, in the Miami Cana.}, Ohio,after an abrUIJt wluming following prolongedeold weather. Kirtland, however, did not realizethat the deaths were prollably It direct result ofthe sudden temperature rise, for he attributedthe mortality to an inability of the species towithstand the colder, northern climate. That lowtemperature is a faetor limit.ing the distributionof the gizzard shad, however, is evidenced bytheir absence from t.he upper parts of Lake Huronand Lake Michigan and an of Lake Superior(Miller 1957: 108). Also, la.rge numbers of youngshad were seen frozen in the ice throughout. FortRandall Reservoir, S. Dak., during the winterof 1955-56, a mortality plausibly attributed tocold temperature (Shields 1956: 29).
The possibility t.hat a fungus dise.ase may belocally responsible for deat.hs of gizzllrd shad(Tra.utman, -in Cln.rk Hl42: 255; a.Iso suggested ton~e in personal conuuunication by Dr. George 'V.
Bennett) seems to me to be more likely the resultof secondary infection aft.er the shad are weakened. Bangham and Hunter (1939: 396) andBangham (1941: 442) suggested that a sporozoanparasite ma.y kill many young gizzard shad duringAugust. and Sept.ember in Buckeye Lake, Ohio.However, the shad that die are represented byseveral yea.r classes, indicating that the causalfactor (or factors) is not necessarily correlatedwith age.
The gregarious habits of this species and itspredilection for quiet Wfl.tPl''l may also lead tomass mortality. From about November 11 to 13,in 1952, a large run of shad from the DetroitRiver up t.he intake pipe (diameter, 5 feet) ofthe Pn.rke-Davis Company plant in Detroit. ledto the death, from. mechanica.I causes, of anestimated 2 to 3 t.ons of Doro8oma of rather uniform size. The reduced current velocity of theintake, as. compared with that in t.he river, is aplausible explanation for the migration (summarized from report in the files of the Institutefor Fisheries Research, Michigan Department ofConservation) .
There are two report.s of apparent mass spawning mortalit.y, both from Florida (Moody 1957:30). A marked reduct.ion of the population, without subsequent immediate recovery, occurred inApril or May toward the end of the spawningseason. A simila.r mortality took place in thethreadfin shad (D. peteneJl/je) in Florida (Berry,Huish, and Moody, 1956).
F01'fl.ge 'I.wlue.-Since the 1880's when its foodand feeding ha.bits were first st.udied by Forbes,t.he merits of the gizzard shad as a forage fishha.ve been repeatedly emphasized. Its important.qualifica tions are (1) direct utilization of phytoplankton; (2) a high reproductive capacity andllbundance; (3) genera.} freedom: from parasites;(4) a rapid mte of growth; and (5) utilizationas food by importa.nt. game fishes. It. has beenspoken of as "the most efficient biologically ofllll the forage fishes" (Hubbs 1934: 57) becauseof the shOJ,t and efficient link in its food cha.inthat. directly connects basic plant life with gamefishes.
Thus, it might seem as if t.he gizzard shadcould be c.alled the idenl fomge fish, but its suitnbility in this regard is affected by cert.ain ecological conditions. Because of rapid growth to

SYSTEMATICS AND BIOLOGY OF GIZZARD SHAD 387
a size not eaten by' predators ample breedingstocks of D01'08011Ul. are assured, but undue multiplicntion of these stocks may result, in competition with the predntors. The heavy populationsof gizzard shad in some waters may be partiallyresponsible for high turbidity ml(l this nmylead to a decrease in productivit.y of game fish(Thompson 1941: !H1, 214). The great abundance of young slmd may result. in competitionfor food wit.h the young of game fishes. Illustrntions of how shad populations can mushroomin warm, shallow bodies of water that have asoft. mud bot.tom, high t.urbidit.y, and relnt.ivelyfew predators have already been given (see section on Abunda.nc.e, p. 384:). ~ere D01'080'11UJ, findscondit.ions for existence nt an optimum and theintroduct.ion of this fish into such WItters is to beavoided if a high level of game-fish production isdesired. Wiebe (-in. Hubbs 1934: 60-61) notedthat stocking gizzard shad in ponds at Burlington,Iowa, resulted in overpopulation of t.he species.
In many other wltt.ers the gizzard shad is ahighly est.eemed forage fish. Wickliff (1933: 275)stated, "Upon the presence or absence of this fishseems to rest. t.he burden of whether or not. impounded waters in Ohio will he productive ofseveral game fish * * *.~' This author pointedout t.hat. D. cepedif.I:nU'ln does not compet.e appreeiably with other fishes in spawning sites or ingenertll habitat preference, that. it cannot. be depleted by angling, and that it. makes an a.ttrllctivebait. Its chief drnwhacks are a natural mortalit.yin t.he spring and fall and sensitivity to handling.In the deep, clear reservoirs of the Te.nnesseeRiver system, young-of-the-year gizzard shnd constitute the most important. food source of thegmne species taken from deep as well as fromshallow wat.er. Continuous cropping of this yeardass leaves just II sufficient nucleus of adultsto maintain a large numher of young, so that theforage fish-predator relation is in nearly perfect.bnlance.
In experiment.al comhinations of gizzard shadwith bluegill '( Lep01nilj Jluw/'ocldr/.l/:l) and largemouth bass (LlIic'/'opter!fs 8llbnoides) in Alabamaponds, Swingle (1949: 5:\-55) showed that. although a relatively high poundage of largemouthhass resulted from a shnd-hass combination, theyield was not sufficient to justify omission ofbluegills and, when that species was added, la-rge
shad accumulated and l:educed the size of the adultbluegills with which they competed for food.
In discussing the mallagement of Indiana, pondsfor fishing, Krumholz (1!)52: 256) wrote thatgizzard shad should not. be stocked in the pondsof that. State, but. gave no reasons.
The gizzard shad is said to be by far the mostabundant species in the larger impoundments ofOklahoma and t.o assure an ample food supplyfor game fishes throughout. the summer months(.Jenkins 1951: 9:~). The species is fast-growingand rather short-lived, and, although its rapidgrowth eliminates t.he gizzard shad from a sizerange suitable for food of the smaller game fishes,the large. crop produced each year offsets thisdisadvnnt.age (Jenkins 1953b: 36).
After a study of 22 wltrlll-water artificial lakesin Illinois, Bennett (1943: 364) wrot.e:
Gizzal'd sharI were present in 10 of the 22 lakes, Insome cases they wel'e introduced b)' floods fmm nearbystreams. In others the)' were stocked as forage fish.The)' are not satisfactol'y for this pmpose in artificiallakes because they repl'oduce in tremendous numbel'S andrapidl~' become too lal'ge to be eaten by the game fishpresent. In almost every case, large shad populationswere llssociated with small numbers of bass. In 5 of the10 lakE'S the sharI constituted from 48 to 65 percent of theentil't~ fish populations (b~' weight) and the game andpHn fish wel'e small llnd stunted,
DoroJ5O"1na, cepedianuln is a Yilluable forage fishhut. under certain conditions it is not suitable forthis purpose. In shallow, warm-water lakes withmud bottom, high turbidity, and high fertility,it is likely to get out of eontrol, even if numbersof predatory game fishes are present. This isparticularly true if the species is not· nflt.ive tosuch waters. It. is ideal for forag-e use in fluctuating impoundments (sueh as Norris Reservoir,Tenn.) where the water is deep and elear, theshoreline is abrupt., no littoral vegetation develops, there is a paucity of benthic flora andfauna but. adequate plankton, and sufficient predatory species are present to crop the young-of-theyear. It is also valuable in many waters that· aresomewhat intermediate hetween these t.wo extremes, particularly where it for111s a part. of theIlative fish fauna which also includes a number ofpredatory fishes.
RfI!lIge.-A <Jetailed discussion of the northe.rnlimit of the gizznrd slutel was published recently(Miller 1057: 105-108). Howeyer the species may

388 FISHERY BULLETIN OF FISH AND WILDLIFE SERVICE
have entered the Great Lakes, it. is known todayfrom Lake Erie, the southern parts of LakeHuron and Lake Michigan (which it has veryrecently invaded), and the Lake Ontario basin(Greeley 1940: 68). Speeimens were taken recently in the St. Lawrence River near Quebec(Vladykov 1945). The erroneous records of thisspeeies from New Brunswick have been furtherdiscussed by Scott and Crossman (1959: 30). .
The species ranges from southeastern SouthDakota and central Minnesota, the Great Lakesdrainage, and extreme southern New York, southward through t.he Mississippi River system anda.}ong t.he Atlant.ic slope to t.he Gulf coast of theUnited States and to the hasin of the Rio Panucoin eastern ]\{pxico.
That the species has also entered the artificialcanals and thus extended its range seems wellfounded. It has entered Lake Michigan by moving through the Chieago River Camtl, and itsOCCUl'l'ence nt the northern end of Cayuga Lake,N.Y., has been plausibly eredited to its transportt\lere from I.ake Erie by way of the Erie Canal.'Vright (1918: 544) wrote, "The most recent Eriecontribution in the mouths of our Ithaea streamsis the gizzard shad (Do'ro80lna cepedumum)* * *:. Greeley (1928: 95) recorded about 20specimens taken November 11, 1916, from CayugaLake (evidently the same fish referred to by"Tright.), with the notation that fish presumablyof this species were reported to have come throughthe Erie Canal one winter, many dying nnderthe ice.
SUMMARY
Because of its importance to fishery workers,the gizzard shad, DO'ro8oma cepedi~l:n1t1n: istreated in detail. Series of specimens fromthroughout its wide range form the basis for asummary of taxonomic characters and variation;no subspecies are recognized.
Dm'Q8o·lIlr1. eepedlrt:II·!WI. spawns nea.r the surfacein fresh water from about the middle of March"to the latter part of August, over a water temperature range of about 50° to 70° F. The eggsal'e sticky and demersal and, at times, may eoveraquatie plants. The fertilized egg is nearlytransparent and measures about. 0.75 mm. in diameter after fixation. The embryology und ea-rly lifehistory n.re described and figured.
The gizzard shad usually matures in its secondor third year and may live to be 10 years old,but it t.ypically does not attain an age greaterthan 5 t.o 7 years. In the northern part of itsrange, it may mature at about 6 inches in totallength. From 70 to 80 percent of its annualgrowth occurs during .Tune-August or JulySeptember, depending on the season and the ageof the fish. Fish 10 to 13 inches long weigh about1 pound. The species commonly attains totallengths bet.ween 10 and 14 inehes; the largestknown individual was 20.5 inehes long andweighed 3 pounds 7 ounces. No sexual dimorphism has been q.emonstrated for growth rateor in the length-weight relation. There are noreliable external cha.racters by which the sexesmay be distinguished.
Except for a few weeks after ha tching, D.cepediaml1/1. is almost entirely herbivorous, feeding heavily on mieroscopic plants, phytoplankton,and algae. Once the early zooplankton stage ispassed the diet. is remarkably alike at varioussizes. The species is essentially a filter feeder.
At least 17 important game fishes may eat. gizzard shad, usually the young-of-the-year about2 to 5 inches long; walleyes, however, may utilizeindividuals 8 inches or longer. Its importanceas a forage fish is emphasized by the short foodchain (direct utilization of phytoplankton), highreproductive capacity a.nd a.bundance, generalfreedom from parasites, and rapid growth rate.At times, however, the speeies so overpopulatessome waters that expensive means of control arenecessary. Gizzard shad nmy become a nuisaneein warm, shallow lakes that have a soft mudbottom, high turbidity, and relatively few predators; this is pllrtieularly true if the speeies is notnutive to sueh waters. In many other waters thegizzard shad is a highly esteemed forage fish,and it is partieularly valuable fOl' this purpose influctuating inpoundments whieh have deep andclear water, U~l ahrupt shoreline, little or no littoral vegetation, adequate .plankton (but a sparsebenthie flora and fauna), llnd suffieient predatoryfishes to crop the young-of-the-year.
LITERATURE CITED
AIIROTT, CHARLES C.1SRl. Description of II new species of Chatoes!!Ius
Cuv, il'om New Jel'ser. PI·OC. Acad. Nat. Sci.Phila., 1860. pp. 365-3Rfi,

SYSTEMATICS AND BIOLOGY OF GIZZARD BHAD 389
AGERSBORG, H. P. K.Ul30. The influenee of tempel'atm'e on fish. Eeolog.v,
vol. 11, No. I, Pl'. 136-144, figs. 1-4.BAKER, C. L.
194~. The effeets on fish of gulping atmosphel'ic airfrom waters of val'ious carbon dioxide tensions.Tennessee Acad. Sci., vol. 17, No. I, pp. 39-50,fig. 1.
BANGHaM, RALPH V.1941. Parasites from fish of Buekeye Lake, Ohio.
Ohio Jour. ScI., vol. 41, No.6, pp. 441-H8.BANGHA!>l, RALPH V., line} GEORGE W. HUNTER, III.
Ul39. Studies on fish parasites of Lake Erie. Distribution studies. Zoologica, vol. 24, pp. 385---148.
BEAN, T. H.1893. The fishes of Pennsylvania, with descriptions
of the species and notes on their common names,distribution, habits, etc. Harrisburg, viii + 149 pp.,illus.
B~~NNETT, GEORGE W.1943. Management of small al'tificial lakes. Bull.
Illinois Nat. Rist. Smv., vol. 22, pp. 357-376, figs.1-7.
BERG, LEO SEMYONOVICH,Ul40. Glassificlltion of fishes, both recent and fossil.
'L'l'av. Inst. Zool. Aead. SeL USSR, vol. 5, No.2,pp. 87-517, figs. 1-190. (Photolithogrllphed, 1947,b~' Edwards Bros. Ine.. Ann Arbor, Michigan.)
BERRY, FIIEllERICK H.1958. Age and gl'owth of the gizzard shad (Doro
sOllla lacepedi) (LeSueur), in Lake Newnan, Florida. Proc. Eleventh Ann. Conf. Southel1sternAssoc. Game and Fish Commissioners, pp. 318-331,figs. 1-6.
B~~RRY, FRJo:llio~RICK H., MELVIN T. HUISH, and HAROLDMOODY.
105G. Spawning m'JI"tality of the thl'e!ldfln shad,DQl"Osoma petenellse (Giinther), in Florida. Copeia,No.3, p. 192.
HUNHalf, KEI.SHaW,10·n. Food of gars in Texas. Tl'ans. ArneI'. Fish.
Soc., vol. 70 (1940.1, pp. 351'1-362.BOWERS, CHaRI.ES C.
1955. Seleetive poisoning of gizzard shad withrotenone. U.s. Fish and Wildlife Service, Prog.Fish-Cult., vol. 17, No.3, pp. 134-135.
CADY, EARL R.UJ45. Fish distrihution, Norris Resel'voir, Tennessee,
1943. 1. Depth distrihution of fish in NOl'ri!<Reservoit·. Rept. Reelfoot Lakl' BioI. Sta.. vol. 0,pp. 103-114, fib'S' 1-5.
CARLANllER, KENNETH D.1950. Handbook of fl'eshwatel' fishery biolog;v. Wm.
C. Bl'own Co., Dubuque, Iowa, v + 281 lip., appendiees.
1953. First sllpplellll'nt to handhook of fl'eshwatl'l'fisher~' biology. Jhid., vi + 277--4~9.
CLARK, CLARENCE F.1942. , A stll'I~· of thl> I,)ss of fish fl'Olll an al'tifieial
lake over' a wasteweil', Lake LOl'amie, Ohio. Tl·ans..7th No. Amer. Wildlife Conf., pp. 250-250, figs. 1-~.
DENllY, JACK S.1945. Fish distribution, Norris Reservoir, Tennessee.
1943. II, Depth llistl'i1JUtion of fish in relation toenvironmental faetors, Non'is Reservoil'. Rept.Reelfoot BioI. Stn., vol. 9, pp. 114-135, figs. 1-15.
1946a. Furthel' studies of depth distribution of fish,Norris Resel'voil', Tenn. Jour. Tennessee Acad.Sci., vol. 21, No. I, pp. 94-104, figs. 1--4.
1946b. Food of several species of fish, Norris Reservoir, Tennessee. Rept. Reelfoot Lake BioI. Sta"vol. 10, pp. 105-127. figs. 1-5.
ESCHMEYER, R. WILJ;.IAM.1944. Fish migl'ation into the Clinch Rivel' below
Norris Dam, Tennessee. Rept. Reelfoot I.ake BioI,Sta., vol. 8, pp.31-41.
ESCHMEYER, R. W., RICHARIl H. STROUD, amI ALDEN M.JONES,
1944. Stmlies of the fish population on the shoalarea of a TVA mainstream reservoir. Rept. Reelfoot Lake BioI. Sta., vol. 8, pp. 70-122, figs. 1-3,
EVER1UNN, BARTON WARREN.1899. Report on investigations by the United States
Insh Commission in Mississippi, Louisiana, andTexas, in 1897. Rept. U. S. ComIn. Fish and Fish.,1898, pp. 285-310, pIs. 8-36.
EWERS, LELA A., and l\f. W. BOESEL,193ft The food of some Buckeye Lake fishes. Trans.
Amel". Fish. Soc.. vol. 65 (1935), pp. 57-70.FORBES, S. A.
1888. Studies of the food of fresh-water fishes.Bull. Illinois State Lab. Nat. Rist., vol. 2, pp. 483473.
1903. The food of fishes. Bull. Illinois State Lab.Nat. Hist., ed. 2, vol. 1, No. 3~ pp. 19-70.
FORBES, STEPHEN A., llnd ROBERT E. RICHARDSON.19~0. The fishes of Illinois. Illinois Nat. Rist.
SUI'v.. ed. 2, cxxx\'i + 3!'i7 pp., figs. 1-76, 78 pIs.l. 54 in eolor) ..
FOWLER, HENRY W.19·Hi. A study of the fishes of the southern piedmont
and eIJastal plain. Acal1. Nat. Sci. Phila., Monogl".,vol. 7, i-vi + 408 pp.. figs. 1-313.
OIU" THEODORE. ,1872. Arrangement of the families of fishes, or
classes Pisces, Marsipobl'anchii, and Leptocardii.Smithsonian Misc. Coli., No. 247, i-xlvi + 1-49.
GOODE, GEORGE BROWN.1884. The fisheries and fishery industries of the
United States. U. S. Comm. Fish and Fish.. sec. 1,xxxiv + 895 pp.
GOWANLOCH, JAlfES N.1933. Fishes and fishing in Louisiana. Louisiana
Dept. COll8el'v.. Bull. 23, pp. 1-6.'36, illns.HKEEI.EY, ,J. R.
1fI28. I!'ishes of the Oswego watel'shed. New YOl'kConsel'v. Dept., Suppl., 17th Ann. Rept.. 1927, pp.84-107, col. pIs. 1-12.
1940. Fishes of the watershed with annotated list./", A biological surve~' of the Lake Ontario watershed. Suppl., 29th Ann. Rept. New York Conserv.Dept., 1939, pp. 42-81, col. pIs. 1-4, 2 figs.

390 FISHERY BULLETIN OF FISH AND WILDLIFE SERVICE
GUNTER. GORDUN.1938. Notes on invasion of fresh water b~' fish{'s of
the Gulf (If Mexico, with special reference to theMississippi-Atchafala~'a River system. Copeia.No.2. pp. 69-72.
1945. Studi{'s '011 Illlu'ine fish{'s of Texas. PubI.Inst. Mar. Sci., vol. 1. No.1. 190 pp. figs. 1-11.
GUNTHER. ALBERT.18fl8. Catalogue of th{' fishes in the British Museum.
Vol. 7. xx + 512 pp. London.HANSEN. DONALD F.
1951. Biolog~' of the whit{' crappie in Illinois. Bull.IlIinois Nat. Hist. Surv.. vol. 25. art. 4. pp. 211265. figs. 1-13, frontis.
HARLAN. JAMES R.. and EVERETT B. SPEAKER,1956. Iowa fish and fishing. State of Iowa Printing
Board, Des Moines, I'd. 3, x + 377 pp., 10 figs..22 pIs. (18 in color).
HART. J. S.195:::!. Geogl'aphic variations of some ph~'siological
and morphological characters in certain freshwaterfish. Univ, Toronto. BioI. Sel·.• No. 60, Publ. Ontario Fish. Res. Lab.. No. 72. pp. i-iv. 1-79. figs.1-18.
HEINEN. E. T,1958. For better fishing. Florida Wildlife, vol. 11,
No.9. pp. 16-18. 39. illus.HERR~;. ALBERT 'V., and GEORGE S. MYERS.
1931. Fishes from southeastel'n China and Hainan.. Lingnan S('i. Jour.• vol. 10, pp. :::!33-254.
IhLDEBRANll. SAl!>fUET. F .. and WTLLIAM C. SCHROEDER.19::?8. Fishes of Chesapeake Bay. Bull. U.S. Bur.
Fish.. vol. 43. pt. I, pp. 1-366. illus.HILDEBRAND. SAMUEL F .. and IRVING L. TOWERS.
1928. Annotatell list of fishes collected in the vicinit~' of Greenwood. Miss.. with descriptions of threenew species. Bull. U.S. Bur. Fish.• vol. 43 (1927).pp. 105-136, figs. 1-9.
HUBBS. CARL L.1934. Some experiences and suggestions on forage
fish ....ulture. Tl'anl". Amel'. Fish. Soc.. vol. 63(1933 ), pp. 53--63.
1944. Terminology of elll'I~' stages of fishes. Copeia,1943. No.4. p. :::!60.
HUBBS. Cl.UT. L .• and KARL F. LAGLER,Hl43. Annotated list of the fishes of Foots Pond.
nibson County. Indiann. Indiana Dept. Conserv..Div. Fish anel Game. Investigations of IndianaLakes and Stl·eams. vol. 2 (10421. No.4. pp. 73-83.
HUBBS. CART. I,.. lind STANLEY C. WHITLOCK.1929. Di vel'se types of young in a single species of
fish, the gizzard shad. Papers Michigan Acad.Sci., AI·tS, and Letters, vol. 10 (1928), pp. 461-482.figs. 31-40.
IWAI. TA"lIOTSU.1956. Anatomy of the phar~'ngelll pockets of the
,Tapanese gizzard shad. KOllosirus pUll~ta·tus (Temminck and Schlegel). Japanese Soc. Sci, Fish..vol. 2::?, No.1. pp. 9-11, figs. 1--4.
JENKINS. ROBERT M.1951. A fish population study of Clal'emore City
lake. Proc. Oklahoma Acad. Sci.. vol. 30 (1949),pp. 8-!-93.
1953a. Growth histol'ies of the principal fishes inGrand Lake W'The Chel'okees), Oklahoma,through thit·teen years of impoundment. Oklahoma Fish. Res. Lab. Rept.. No. 34, pp. iii + 87,figs. 1-19 (mimeo.).
1953b. A pre-impoundment sm'vey of Fort GibsonResel'voir, Oklahoma (summel' 1952). OklahomaFish. Res. Lab. Rept., No. 29. 53 pp. (mimeo.).
1957. The effect of gizzard shad on the fish population of a smaII Oklahoma lake, Tl'ans. ArneI'. Fish,Soc.. vol. 85 (1955). pp, 58-74, figs. 1-6.
JORDAN, DAVID S.1882. Report on the fishes of Ohio. Ohio Geol.
Surv., vol. 4, pt, 1, pp. 737-1002.KIMSEY, J. B.
1054. The introduction of the redeye black bass andthe threadfin shad into California. California Fishand Game. vol. 40, No.2. pp. 203-204,
KIRTLAND, JARED P.1838. Rep(ll't on the zoology of Ohio. Ann, Rept.
Geol. Sun. State of Ohio. vol. 2. pp. 157-197.1844. Descriptions of the fishes of Lake Erie. the
Ohio River and theil' tributal·ies. Boston Jour,Nat. Hist., vol. 4, pp. ~1-240. pIs. 9-11.
KRIT"lfHOLZ, LoUIS A.19.'12. Management of Indiana ponds for fishing.
Jour. Wildlife l\:Igmt., vol. 16, No.3. !lp. 254-257.KUHNE, EUGENE R.
1939. A guide to the fishes of Tennessee and themid-south. Div. Game and Fish.• Tennessee Dept.Conserv.• 124 pp.• 81 figs.
LAGLER. KARJ. 1<'., CARL H. OBRECHT. and GEORGE V. HARRY,1943. The foO(I and 'habits of gal'S (Lcpi8osfeu8 spp,)
considered in relation to fish management. IndianaDept. Conserv.• Div. Fish and Game. Investigationsof Indiana Lakes I1ml Streams. vol. 2 (1942), No.8,pp. 117-135. fig. 1.
LAGT..ER. KARL F .• and FIIANCES V. HURBS,1940. Food of the long-nosed gar (Lepisosteus OSSCItS
o.r.yu.rus) and the bowfin (A.mia calt-al in southernMichigan. Copeia. No.4. pp. 239-240.
LAGLER. KARL F .• and HARRY VAN METER,1951. Abundance and growth of gizzard shad, Doro
soma cepedianum (LeSuem' I , in a small IllinoisLake. Jour. Wildlife Mgmt., vol. 15. No.4. pp. 357360.
LAGI.ER, KARJ, F .. and VERNON C. ApPLEGATE.1943. Age lind growth of the gizzard shad. Doro
soma re/JCdi" 11"11111 (LeSueur I. with a d iseussion ofits value as a buffer and as forage of game fishes.Indiana Dept. Consel'v., Div. Fish and Game. Investigations of Indiana Lakes and Streams, vol. 2(1942). No.6, pp. 99-110. figs. 1-3.
LAGI.ER. KARL F., and W. E. RICKER.1943.' Biological fisheries investigations of I<'oots
Pond. Gibson Count~·. Indinna. Indiana Dept. Conserv.• Dir. I<'ish amI GalUe. Illvestil,.ratiolls of IutliannLakes and Streams, vol. 2 (1942 I. No.3. pp. 47-72,figs. 1-13.

SYSTEMATICS AND BIOLOGY OF GIZZARD SHAD 391
LAGLER, KARL FRAriK, and WALTER CHARLES KRAATZ.1!"I45. PIIIIl'~'ngeal pockets in the gizzal'd shad, Doro
sOl/l.a· ccpf1dill/lI/.//I. {LeSueUl·). Papel's MichiganA(:ad. Sci., Al·tS, amI Letters. vol. 30 (1944). pp.311-3~O, pIs. 1-2, fig. 1.
LANGLOIS, THOMAS F.1954. 'fhe western end of Lake El'ie and its ecology.
.J. W. Edwal'ds, Inc., Ann Arbor, Mich. 479 pp.,HhIS.
LEWIS, WIT.LIAM M.1953. Analysis of the gizzard shad population of
Crab OI'char(1 Lake. Illinois. Tl·ans. Illinois Acad.Sci., vol. 46, pp. ~31-234, fig. 1.
MADDEN. K. M.1951. Gizzard shad suppl'esses Black Hawk Lake
fish populations. Iowa Conser\'.. vol. 10, No. 12,pp. 185, Un-192, 2 figs.
MARTIN. ROBERT G., Hnd ROBERT S. CA~[PBELL.
1953. The small fishes of Black River and ClearwaterLake, Missoul"i. III The Black River' ~tlldies, byJohn L. Funk et al. Univ. Missouri Stud.• vol. 26,No.2, pp. 45-06, fig. 1.
MASSMANN, WILT.IAIII H. ,1953. Relative' abundance of ~'oung fishes in Virginia
estuaries. '1'runs. l~tll No. Amer. Wildlife Coni.,1953, pp. 439-449, figs. 1-4.
MILLER, ROBERT RUSH..If150: A review of the American clupeid fishes of the
genus Dorosot/loll. Proc. U.S. Natl. Mus., vol. 100,pp. 387-410. . .
1957. Ol'igin and dispersal of thp alewife. AJoSGpS61tdoha.rell!1l1S, IIml the gizzard shad, Dorosol/l.lI·cepedillnulII-. in the Great Lakes. 'frans. Amer.Fish. SlIC., vol. 80 (1956), pp. 97-111.
MOODY, HAROT.D L.1954. Adult fish populations by haul seine in seven
FIOI'irla lakes. Quart. Jour. FIOl"irla Acael. Sci., vol.17, No.3. pp. 147-167.
1957. A fisheries study of Lake Panasoffkee. Florida.Ibid., vol. 20, No. 1, pp: ~1-88.
NASH, CAIlROI.L BI.UE.]!)!'i0. Associations between fish species in tI'ibutaries
and shol'e waters (If westel'n Lake El·ie. Ecology.vol. 31. No. ,I, pp. 5131-566.
PARSONS, .TORN W.. 1111(1 .J. BRUCE KIMSEY.1954. A I'epol't on the Mississippi threadfin shad.
U.S. ~'ish and Wildlife Sel'viee, PI·og. F'ish-Cult..vol. 16, No.4, pp. 179-181, 1 fig.
PATRIARCHF.:, ~b;RCER H.1953. The fishel'y in Lake Wappapello, a flood-con
tI'ol reservoir on the St. Fl'ancis Rivel', Missouri.Tran. Amel'. Fish. Soc., vol. 82 (1952), pp. 24~-254.
PATRIARCHE, ~IERCER H., and EDWARD 1\:1. LOWRY.1953. Age Ilnd rate of gl'owth of five species of fish
in Black River, Missouri. III The Black RiverStudies, b~' John L. Funk et al. Univ. MissouriStud., vol. ~(I, Nn. 2. pp. 85-109, figs. 1-11.
POW~;RS, EIlWIN B.1938. Factol'S in\"olvel.l in the snrlrlen mortalit:\" of
fishes. Tl"Ilns. Amel·. Fish. Soc., \"0J. 67 (1937),pp.271-281.
REGAN, CHARLES TATE.1917. A re\"ision of the dupeoid fishes of the geneI'll
l'oll/.olobus, Bl'cvoortia and Dorosollla.,· and theil'allies. Ann. and Mag. Nat. Hist., ser. 8. vol. 19,pp. 297-316.
ROSE, EARL T.1957. Results of til$h mllnagement at Blackhawk Lake.
Iowa Conserv. Comm., Quart. BioI. Repts., vol. 9,No.1. pp. 6-11.
ROSTLUND, ERHARD.195~. Freshwater fish and fishing in nath'e NOI·th
America. Univ. California Publ. Geog., vol. 9, pp. x+ 313, fig. I, maps 1-!7.
SCOTT, DONALD C.1951. SlImpling fish populations in the Coosa River,
Alabama. 'frans. Amer. Fish. Soc.. vol. 80 (1950),pp. 28--40, figs. 1-~.
SCOTT, W. B., and E. J. CROSSMAN.1959. The freshwater fishes of New Brunswick: A
checklist with dish'ibutional notes. Contrib. RoyalOntario Museum, Div. Zool. and Paleon.. No. 51,pp. 1-..'l7, figs. 1-2.
SHIELDS, J AlIlES T.1956. Repol't of' fisheries investigations during the
third ~'ear of impoundment of Fort Randall Reser\"oir, SO~lth Dakota, 1955. South Dakota Dept.Game, Fish, and Parks. Ding-el-Johnson Proj. F-lR-5;195~, 91 pp., apllend. 1-2.
STROUD, RICHARD H.1949. Growth' of NOl'ris Reservoir walleye during the
first twelve ~-ears of impoundment. Jour. WUdlife. Mgmt., '-01. 13. No.2. pp. 151'::'177. figs. 1-5.
SWANSON, GUSTAV.1932. A mid-winter migl'ation of gizzard shad. Co
peia, No. I, p. 34.SWINGLE. H. S.
1949. Experiments with combinations of largemouthblack bass. bluegills, and minnows in ponds. Trans.ArneI'. Fish. Soc.. \"01. 7(; (1946), pp. 46-62.
THOMPSON, DAVID H.1941. The fish l)l'oduction (If inland streams and
lakes. III A srmposium on hydrobiology. Univ.Wisconsin Pl'ess, 1941, pp. 206-217, figs. 1-2.
TIFFANY. L. H.1921. The gizzal'd shad in relation to plants and
game fishes. TI·ans. Amer. Fish. Soc., vol. 50 (1920.1.pp. 381-380.
1922. Some algal statistics gleaned fl'om the gizzaJ'dshad. Science, \"0J. 5(j, pp. 285-286.
TRAUDIAri. MILTON B.1940. The birds of Blll:ke~'e Lake. Univ. Michigan,
Mus. Zool. ~lisc. Pnb\. No. 44, pp. 1--466, pIs. 1-15,maps 1-~. frontis.
1057. The fishes of Ohio. Ohio Stnte Un i\". Press,1957. pp. xvii + 083, maps I-XI and 1-172, figs.I-II lind 1-Ii2, pIs. I-VII (col. I.
UrilTEIl STATES FISH AND WILDLIFE SERVICE.1056. Feeding fl'esh-water fish to fur animals. Comm.
Fish. Re,-.• vol. 18, No.1, p. H.

392 FISHERY BULLETIN OF FISH AND WILDLIFE SERVICE
V.:r"MlQUJo:7.. GR.:GORIO 1.'.10HO. On the viahilit~' of algae ohtained from the
lligestive tl'ad of the gizzlll'd shad. Dorosoma(:n/lcdiullllln (LeSueur). Amel'. Milll. Nat., vol. 22.No.2. pp. 37(;--405, figs. 1-3.
VI.AUYKOV. VAunl DllnTRI,T.104fi. TJ"Ois poissons nouveaux pour lit Province de
Quebec. Cana,lian Nat.• vol. 72, pp. 27-30, figs. 1-5.WALSH, BIU..
lo.iH. The "big sUnk" at Erie. Pennsylvania Angler.vol. 22. Nil. 5. pp. 2-4, figs. 1-5.
WARNER, E. N.19-H. Studies on thE- emlwyology and eal'ly life his
tlll'y of the gizzal"l shad. Dorosotllu. ccpedi.umtf1l.
LeSuE-ur. Abst!'act Dodol'al Dissertation, OhioState Dniv., vol. 34, pp. 637-643.
WIl:KLIn', E. L.1933. Are newly impounded watel's in Ohio suitable
for fish life? Trans. Amel'. Fish. Soc., vol. 62(1932). pp. 275-277.
1945. 1<'ood of the gizzard shad. Ohio Div. Conserv.and Nat. Res.• Fish Mgmt. Leaflet No. 184, pp. 1-4(mimeographed) .
1953. Gizzal'd shad rides again. Ohio Conserv, Bull..vol. 17, pp. 23, 25.
WICKLIn', E. L., and M. B. TRAUTMAN.19..'11. Some food and game fishes of Ohio. Ohio Dept.
Agr., Bull. 7, pp. 1-40, illus.'VIER, H. C., and E. P. CHURCHILL.
1946. The anatom~' and histology of the digestive system of the gizZlll'd shad. Dorosoma oepedianum(LeSueur). Proc. South Dakota Acad. ScL, vol.25, pp. 34-43, 8 figs.
WRIGHT, ALRERT HAZEN.1918, Fish succession in some Lake Ontario tribu
taries. Scientific Monthly, vol. 7, pp. 535-544, maps1-3.
.u. S. GOVERNMENT PRINTING OFFICE: 1960 0 ·528919










![ELECTRONIC MEDICAL RECORDS (EMR) STREAMLINE …chapter.aapm.org/pennohio/2013FallSympPresentations/SI7... · 2013. 11. 12. · Microsoft PowerPoint - Ppt0000033.ppt [Read-Only] Author:](https://img.pdfslide.net/doc/110x75/5fe0a80f4c1efe2f7a2f5c72/electronic-medical-records-emr-streamline-2013-11-12-microsoft-powerpoint.jpg)


















