Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1989 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 264, No. 22, Issue of August 5, pp. 13074-13079.1989 Printed in U.S.A.
Glucocorticoids Induce the Accumulation of Novel Angiotensinogen Gene Transcripts*
(Received for publication, January 10, 1989)
Elia T. Ben-AriS, Kevin R. LynchQV, and James C. Garrison11 From the Departments of Pharmaco1oE.y and $Biochemistry, University of Virginia School of Medicine, Charlottesuilk, Virginia 22908
”
Previous work from this laboratory has shown that glucocorticoids and (Sp)-CAMPS, a cyclic AMP ana- logue, stimulate the accumulation of angiotensinogen mRNA in isolated hepatocytes. The present study dem- onstrated that glucocorticoids stimulate the accumu- lation of a novel, 2.25-kilobase transcript of the angi- otensinogen gene, both in isolated hepatocytes and in the intact liver. (Sp)-CAMPS was synergistic with dex- amethasone in causing the accumulation of the larger species of RNA in hepatocytes, but had no effect by itself. Primer extension analysis and S1 mapping ex- periments proved that the 2.25-kilobase angiotensin- ogen RNA consisted of two larger forms of angioten- sinogen RNA extended at their 5’ ends. The novel transcripts are generated by the use of two new t ran- scription initiation sites in the angiotensinogen gene, located at nucleotide positions -328 and -386 relative to the start site at position +1 used in the absence of hormone. A consensus TATA box is found at the ex- pected position 25-30 nucleotides upstream from the start site at position -328, and another TATA-like sequence is found 25-30 nucleotides upstream from the start site at -386. In addition, two consensus glu- cocorticoid response elements occur upstream from the initiation sites. The larger angiotensinogen RNAs do not appear to code for a novel form of the angiotensin- ogen protein. It appears that glucocorticoids can direct transcription from a second promoter in the angioten- sinogen gene and that this promoter is absolutely de- pendent on the hormone. To date, this situation appears to be unique to the angiotensinogen gene.
Angiotensinogen (renin substrate) is the glycoprotein pre- cursor of angiotensin 11, the octapeptide hormone that plays an important role in the regulation of blood pressure and fluid and electrolyte balance. Angiotensin I1 is formed from angi- otensinogen by successive processing steps catalyzed by renin and angiotensin-converting enzyme (29). The major source of plasma angiotensinogen is the liver and the rate of its synthe- sis is regulated in vivo and in vitro by glucocorticoids and a
Health Grants DK 37494 (to K. R. L.) and DK 19552 (to J. G. C. and * These studies were supported in part by National Institutes of
E. T. B.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Section, Bldg 560, NCI-FCRF, Frederick, MD 21701. 4 Current address: Laboratory of Viral Carcinogenesis, Cell Biology
n Established Investigator of the American Heart Association with partial funding provided by their Virginia Affiliate.
11 To whom correspondence should be addressed Dept. of Phar- macology, Box 448, University of Virginia Medical Ctr., Charlottes- ville, VA 22908.
number of other hormones (7, 8, 10, 18, 19, 24, 25, 36, 40). Therefore, it is not surprising that the 5’ region of the angiotensinogen gene contains two glucocorticoid responsive elements (GRE)’ (27), or that dexamethasone stimulates the accumulation of the mRNA for angiotensinogen in hepato- cytes (2). In the course of the studies on hepatocyte angioten- sinogen mRNA, dexamethasone was also found to stimulate the accumulation of a novel, 2.25-kb species of RNA that hybridized with the 1.65-kb angiotensinogen cDNA probe (2). The molecular nature of this RNA was particularly intriguing, since the rat genome contains only a single angiotensinogen gene (27). An interesting possibility was that the large species of angiotensinogen RNA could code for a different form of the protein, as previous reports have described molecular weight heterogeneity of angiotensinogen (4, 5, 15, 22, 34, 39).
The present study explores possible mechanisms whereby glucocorticoids stimulate accumulation of the 2.25-kb angio- tensinogen transcript in isolated hepatocytes. Analysis of the structure of this RNA using a variety of techniques revealed that there are actually two large forms of angiotensinogen RNA that are generated by the use of two novel transcription initiation sites in the gene, approximately 328 and 386 nucle- otides upstream from the primary initiation site. The absence of a long open reading frame in the 5’ sequence unique to the larger species of RNA suggests that the RNA does not encode a novel protein. To date, the use of a different initiation site in response to glucorticoid hormones appears to be unique to the angiotensinogen gene.
EXPERIMENTAL PROCEDURES AND RESULTS~
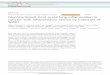
Dexamethasone and a Cyclic AMP Analogue Stimulate Ac- cumulation of a Novel Form of Angiotensinogen RNA-Pre- vious studies using isolated hepatocytes (2) have shown that dexamethasone and other glucocorticoids increase angioten- sinogen mRNA levels. A surprising finding was that dexa- methasone stimulates the accumulation of a novel species of RNA that hybridizes to the angiotensinogen cDNA and is about 300-350 nucleotides longer than the primary angioten- sinogen mRNA species of approximately 1900 nucleotides (2). The Northern blot in Fig. 1 shows the accumulation of this larger RNA (indicated by arrow) after incubation of rat he- patocytes for 3 h with 10 I.LM dexamethasone (lanes 2 and 5), as compared to controls (lanes 1 and 4 ) . (S,)-CAMPS, an
The abbreviations used are: GRE, glucocorticoid responsive ele- ment; (S,,)-CAMPS, cyclic adenosine 3’,5’-phosphorothioate; kb, kil- obase; bp, base pair.
Portions of this paper (including “Experimental Procedures,” part of “Results,” part of “Discussion,” and Figs. 2-4) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are included in the microfilm edition of the Journal that is available from Waverly Press.
13074

Glucocorticoids Induce Novel Angiotensimgen Transcripts 13075
Total Poly A+ in vivo nn n
origin - 1 2 3 4 5 6
2 8 s -
1 8 s - - - 7.5 kb - 4.4 kb
- 2.4kb
- 1.4 kb
- 0.24 kb
FIG. 1. Dexamethasone and (SJ-CAMPS stimulate accu- mulation of a novel form of angiotensinogen RNA. Lanes 1 3 (Total): 5 pg of total RNA extracted from hepatocytes incubated for 3 h at 37 ‘C with vehicle ( l a n e I ) , 1 p~ dexamethasone ( l a n e 2), or 1 p~ dexamethasone and 100 p~ (&)-CAMPS ( l a n e 3). Lanes 4-6 (Poly(A+)): 1 pg of poly(A+) RNA prepared from hepatocytes treated with vehicle ( l a n e 4) , 10 p~ dexamethasone ( l a n e 5), or 10 p~ dexamethasone and 100 p~ (S,)-CAMPS ( l a n e 6). Lanes 7 and 8 (in vivo): 5 pg of total RNA isolated from rats treated for 6 h with vehicle ( l a n e 8 ) or 7 mg/kg dexamethasone ( l a n e 7) as described under “Experimental Procedures.’’ These RNA samples were used for Northern blot analysis with a 32P-labeled 1.65-kb angiotensinogen cDNA probe, and the blot was exposed to x-ray film at room temper- ature as described under “Experimental Procedures.” The origin of electrophoresis and the positions of 18 S and 28 S ribosomal RNA species and of RNA size markers are indicated at the sides of the autoradiograph. The arrow indicates the position of the novel 2.25- kb species of angiotensinogen mRNA.
active CAMP analogue, also stimulates accumulation of the primary form of angiotensinogen mRNA (2) and is synergistic with dexamethasone in causing accumulation of the large transcript (lanes 3 and 6). However, (S,)-CAMPS by itself does not cause accumulation of the larger RNA (not shown). The 2.25-kb angiotensinogen RNA is present in both total and poly(A+) RNA samples from induced cells (Fig. 1, lanes 2 and 3 or 5 and 6, respectively). In addition, the novel transcript accumulates in livers of intact rats injected with dexamethasone but not in the livers of control animals (Fig. 1, lanes 7 and 8, respectively). Cell fractionation experiments revealed that the large form of angiotensinogen RNA is found in the cytoplasm, arguing against its being a RNA precursor (not shown).
Experiments were designed to investigate the possible sources of the additional sequence in the 2.25-kb angiotensin- ogen RNA species. Use of an alternate polyadenylation site, addition of a longer poly(A) tail and alternative processing of precursor RNA were all considered as potential mechanisms for generating the larger transcript. The data presented in the Miniprint Section do not support these possibilities. The availability of a genomic clone of the angiotensinogen gene made it possible to demonstrate that a novel transcription start site in the 5‘ end of the gene was responsible for the longer transcript. These experiments are described below.
Primer Extension Analysis and SI Nuclease Mapping of the Large Form of Angiotensimgen RNA-The data shown in Fig. 4 in the Miniprint Section demonstrate that the additional sequence in the large angiotensinogen RNA was derived from the 5’ region of the gene. Therefore, more precise information was sought concerning the structure and sequence of this
novel transcript. To determine the size of the novel sequence, primer extension analysis was performed with the 40-residue oligonucleotide complementary to nucleotides -154 to -115 of the angiotensinogen gene. This primer, which was labeled at the 5’ end as described under “Experimental Procedures,’’ was expected to hybridize only to the larger angiotensinogen RNA. Ideally, extension of this hybridized primer with reverse transcriptase should proceed until the transcriptase reaches the 5’ end of the transcript, yielding an end-labeled species of DNA that can be visualized by electrophoresis and auto- radiography. The location of the 40-mer in the angiotensino- gen sequence is represented schematically in Fig. 5 (bottom).
The results of the primer extension analysis are shown in Fig. 5 (lanes 1-4) and are diagrammed below the autoradi- ograph. Somewhat surprisingly, two extended species were seen reproducibly in these experiments in samples from dex- amethasone- or dexamethasone and (&.)-CAMPS treated cells (lanes 2 and 3, respectively). As expected, no extended species were observed when the primer was hybridized with RNA from uninduced cells (lane 1 ) or with yeast RNA (lane 4). The size of the extended species (approximately 214 and 272 nucleotides) indicated that there were two larger forms of the RNA, with approximately 328 and 386 additional nucleotides at their 5’ ends, re~pectively.~
While the primer extension analysis yielded information on the length of the additional sequence, it did not definitively prove from what region(s) of the gene this sequence was derived. The possibility still existed that there was another exon (and thus another intron) in the 5‘ region of the angio- tensinogen gene, such that the 5’ end of one or both of the larger transcripts was derived from this additional exon. To address this possibility, S1 nuclease mapping analysis of the larger angiotensinogen RNAs was performed. The 650-bp BamHI-XhoI fragment from the 5’ end of the angiotensinogen gene was isolated, end-labeled, and hybridized to poly(A+) RNA samples from control and hormone-induced cells (see Fig. 5, bottom, and under “Experimental Procedures”). Hy- bridized samples were treated with S1 nuclease, which digests single-stranded regions of DNA and RNA. Thus, the size of the protected, end-labeled fragment(s) obtained from the 650- bp probe, when compared to the sizes of the primer-extended species, will indicate whether there is another intron in the 5’ region of the gene.
The results of the S1 mapping analysis are shown in Fig. 5 (lanes 5-7), and the experiment is represented schematically below the autoradiograph. As the optimal amount of S1 nu- clease needed differs for each message, it was titrated and found that addition of 300 units of S1 nuclease/sample yielded results that were consistent with those of the primer extension experiments. All of the samples contained some undigested 650-bp double-stranded probe (Fig. 5, lanes 5-7). Samples from hormone-treated cells (lanes 6 and 7), but not from uninduced cells (lane 5 ) contained two predominant, S1 nu- clease-resistant species which were approximately 285 and 340 nucleotides long. Thus, the S1 nuclease digested the end- labeled 650-nucleotide probe at positions approximately 321 and 376 nucleotides upstream from the primary transcription initiation site (i.e. nucleotides -321 and -376; see Fig. 5, bottom). These positions corresponded very closely to the positions of the 5’ ends of the two transcripts predicted from the results of the primer extension experiments, at nucleotides -328 and -386, respectively. Thus, dexamethasone stimulates
The low intensity of the extended species in lane 3 is most likely due to poor recovery. These two species were considerably more prominent in a second, identical experiment that used the same RNA samples.

13076
Primer Extension
-
- -
272 -
214 -
B.m HI
Glucocorticoids Induce Novel Angiotensinogen Transcripts
S1 Mapping
650- -
340-
285-
xkol 5’ 3’ gum
609 -3w -328 + I “ (AIn
--(A),, mRNA -- (A), L I * 650ntproa
I SIN- -38 -340nt , - 285 nt I I *
-272nt r-* - 214 nt r*
40-mer
-115
FIG. 5. Primer extension and S1 nuclease mapping analysis of angiotensinogen RNAs. Top, lanes 1-4: 5 pg of poly(A+) RNA isolated from hepatocytes incubated for 3 h with vehicle (lane 1 ), 10 p~ dexamethasone (lane 2 ) , or 10 pM dexamethasone and 100 p~ (S,)-CAMPS (lane 3), or 20 pg of yeast total RNA (lane 4 ) were used for primer extension analysis as described under “Experimental Pro- cedures.” The sizes of the extended species are indicated at the left of the autoradiograph. A 123-bp DNA ladder and a DNA of known sequence were used as size markers. Top, lanes 5-7: 2.5 pg of the same poly(A+) hepatocyte RNA samples used for the primer extension were used for S1 nuclease mapping as described under “Experimental Procedures.“ Lanes 5-7 are from cells treated with vehicle, dexameth- asone, or dexamethasone and (S,)-CAMPS, respectively. The 650-bp probe and sizes of protected fragments are indicated on the left. Size standards were the same as for the primer extension. Samples from both experiments were run together on a urea-polyacrylamide gel, and the fixed and dried gel was exposed to Kodak X-AR x-ray film at -20 “C with an intensifying screen (primer extension), or a t room temperature (S1 mapping). Bottom: schematic representation of primer extension and S1 mapping experiments. The relevant region of the angiotensinogen gene is diagrammed at the top, and the novel transcription initiation sites identified by these experiments are indicated at -386 and -328. The line represents untranscribed re- gions of DNA. The box (open and dark regions) represents the new boundaries of exon 1, and the dark section of the box represents exon 1 as previously defined, with +1 indicating the primary transcription initiation site. The upper two wavy lines represent the two large forms of angiotensinogen RNA, and the lower wavy line represents the predominant form of angiotensinogen mRNA. The 650-nucleotide ( n t ) probe used for S1 mapping, and the protected products of this experiment are represented as lines below the mRNA. The two products of the primer extension are represented as outlined boxes at the bottom of the figure; the 40-bp primer is indicated by the hatched region. Asterisks denote the 3zP label at the 5‘ ends.
transcription of two novel species of angiotensinogen RNA from transcription initiation sites located approximately 328 and 386 nucleotides upstream from the previously identified initiation site. These two new transcription start sites are located approximately 25-30 nucleotides downstream from two putative “TATA” boxes, which corresponds very well with the position of the TATA homology in most eukaryotic genes. There does not appear to be a sixth exon in the 5’ region of the angiotensinogen gene.
DISCUSSION
Previous work in isolated hepatocytes showed that dexa- methasone causes the accumulation of a novel, 2.25-kb RNA that hybridizes with a 1.65-kb angiotensinogen cDNA probe (2). The 2.25-kb transcript also appears in rat liver after treatment of the animal with dexamethasone for approxi- mately 6 h. Interestingly, the cyclic AMP analogue, (Sp)- CAMPS, is synergistic with dexamethasone in causing the accumulation of the 2.25-kb RNA, while having little or no effect on the accumulation of this transcript by itself. The experiments reported here demonstrate that the 2.25-kb RNA band seen on Northern blots probed with the angiotensinogen cDNA represents two novel angiotensinogen transcripts. S1 nuclease mapping and primer extension analysis proved that these novel RNAs are generated via the use of two new transcription initiation sites, located approximately 328 and 386 nucleotides upstream from the primary initiation site in the angiotensinogen gene. The two larger angiotensinogen transcripts probably are not processing intermediates, as they appear in the cytosolic fraction of hepatocytes and contain no intervening sequences.
The findings of the present study are summarized in Fig. 6, which shows a diagram of the 5’ region of the angiotensinogen gene between nucleotides -689 and +31. The nucleotide se- quences shown are as previously reported (27, 38). The pre- dominant angiotensinogen transcript, with its start site at nucleotide +1, is represented by the solid arrow. The TATA box consensus sequence, which is found approximately 25 nucleotides upstream from most mRNA transcription start sites (13), appears 27 nucleotides upstream from the angio- tensinogen mRNA start site, as reported previously (38). The two novel dexamethasone- and (S,)-CAMPS-induced tran- scripts, with start sites at nucleotides -328 and -386, are represented by the dashed arrows. The first nucleotide of these new transcripts would be an adenine and a guanine residue, respectively. A TATA box is located 27 nucleotides upstream from the initiation site at nucleotide -328. The complete AT-rich sequence from nucleotides -359 to -354 is TTATAA. In the case of the most 5’ transcription initiation site, located at nucleotide -386, there is a TTAT sequence located 25 nucleotides upstream, which may represent another
Barn HI ‘GREs’ -386 -328 *1
5‘ ~AGAACA-AGAACA- TClTGT TClTGT r--- r-- I I I I
lTAT”( TATA TATA 3’
-586 -4n -411 -3s -27
FIG. 6. Diagram of the 5’ region of the angiotensinogen gene. Thc two GREs containing the consensus hexanucleotide se- quence, 5’-TGTTCT-3’, are indicated. Both GREs shown are part of longer imperfect GRE palindromes in the gene with the following 5’- 3‘ sequences, a t -586, AGAACATTTTGTTTC and at -477, AGAA- CAGACAACTAA. However, the importance of the entire palindrome to glucocorticoid responsiveness is not clear. The predominant tran- script, initiating a t the +1 position, is indicated by the solid arrow, while the two novel, glucocorticoid-induced transcripts, initiating at positions -328 and -386, are denoted by dashed arrows. The TATA boxes located approximately 25-30 nucleotides upstream from each initiation site are also represented.

Glucocorticoids Induce Novel Angiotensinogen Transcripts 13077
TATA box-like region. Fig. 6 also shows the two GREs that have been previously noted to occur in the 5‘ region of the coding strand of the angiotensinogen gene (27). The GREs, which contain the conserved core sequence identified as the binding sites for the transformed glucocorticoid receptor, 5’- TGTTCT-3’ (28, 33), are located upstream from the three angiotensinogen transcription start sites in this gene.
The accumulation of the two novel angiotensinogen tran- scripts in response to glucocorticoids may result from stimu- lation of transcription or stabilization of these RNAs by the hormone, or may be due to a combination of these two mechanisms. The synergistic effect of (S,)-CAMPS on accu- mulation of the large forms of angiotensinogen RNA, despite the fact that this CAMP analogue has little, if any, effect on accumulation of these RNAs by itself, suggests that this agent acts at least in part by RNA stabilization.
There is only one molecular weight form of angiotensinogen immunoprecipitated by an anti-angiotensinogen anti-sera in the in vitro translation products of mRNA isolated from- dexamethasone-treated hepatocytes (see Miniprint). This re- sult is completely consistent with computer analysis of the sequence of the 2.25-kb form of the angiotensinogen tran- script showing that there is no long open reading frame in the novel sequence of the steroid-induced transcript. Although we have not formally proven that the novel angiotensinogen transcripts are mRNAs, they have very similar structure to the predominant angiotensinogen mRNA at their 3‘ and 5’ ends, are polyadenylated, and are exported to the cytoplasm. Therefore, all available evidence suggests that they encode a normal angiotensinogen protein. The functional significance of the longer angiotensinogen transcripts is not known.
A number of instances are known where hormones play a role in the use of multiple transcription initiation sites. The expression of both forms of chicken very low-density apoli- poprotein I1 mRNA is absolutely dependent on estrogens (35). In contrast, the two forms of rat thyrotropin &subunit mRNA are initiated from two different promoters, one of which is constitutively expressed, while the other, more 3‘ promoter is negatively regulated by thyroid hormone. Both forms of the thyrotropin message apparently code for the same protein (6). The latter case is similar to that described in the present study, except that with the angiotensinogen gene hormonal stimulation leads to accumulation of transcripts with a longer 5’ untranslated sequence. In addition, in the angiotensinogen gene, accumulation of the larger transcripts appears to be absolutely dependent on the presence of hormone. Given these considerations, the hepatic angiotensinogen gene appears to provide the first example of the induction of a novel form of mRNA generated by use of a different initiation site in response to glucocorticoid hormones.
Acknowledgments-We would like to thank Gwendolyn Harris for performing the two-dimensional polyacrylamide gel electrophoresis and Rob Figler for his generous help in running the acrylamide/urea sequencing gels.
REFERENCES 1. Aviv, H., and Leder, P. (1972) Proc. Natl. Acad. Sci. U. S. A. 6 9 ,
2. Ben-Ari, E. T., and Garrison, J. C. (1988) Am. J. Physiol. 2 5 5 ,
3. Berk, A. J., and Sharp, P. A. (1977) Cell 12 , 721-732 4. Campbell, D. J., Bouhnik, J., Coezy, E., Pinet, F., Clauser, E.,
Menard, J., and Corvol, P. (1984) Endocrinology 114 , 776-785
1408-1412
E70-E79
5. Campbell, D. J., Bouhnik, J., Menard, J., and Corvol, P. (1984)
6. Carr. F. E.. Need. L. R.. and Chin, W. W. (1987) J . Biol. Chem. Nature 308 , 206-208
8. 9.
10.
11.
12.
13.
14.
15.
16.
17. 18.
19.
20.
21.
22. 23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39. 40.
262,981-987 7. Clauser, E., Bouhnik, J., Coezy, E., Corvol, P., and Menard, J.
11983) Endocrinology 112,1188-1193 Dzau, V. J., and Herimann, H . C. (1982) Life Sci. 30 , 577-584 Field, L. J., Philbrick, W. M., Howles, P. N., Dickinson, D. P.,
McGowan, R. A., and Gross, K. W. (1984) Mol. Cell. Biol. 4 ,
Freeman, R. H., and Rostorfer, H. H. (1972) Am. J. Physiol. 2 2 3 ,
Garrison, J. C., and Wagner, J. D. (1982) J. Biol. Chem. 2 5 7 ,
Garrison, J. C., Johnsen, D. E., and Campanile, C. P. (1984) J.
Goldberg, M. (1979) Ph.D. Thesis, Stanford University, Stanford,
Hart, R. P., McDevitt, M. A., Ali, H., and Nevins, J. R. (1985)
Hilgenfeldt, U., and Hackenthal, E. (1982) Biochim. Biophys.
Ish-Horowicz. D., and Burke, J. F. (1981) Nucleic Acids Res. 9 ,
2321-2331
364-370
13135-13143
Biol. Chem. 259,3283-3292
CA
Mol. Cell. Biol. 5, 2975-2983
Acta 708,335-342
2989-2998 . .
Kessler. S. W. (1975) J. Zmmunol. 116.1617-1624 ~~~~~~~~
Khayyall, M., Macdregor, J., Brown,.J. J., Lever, A. F., and
Krakoff, L. R., and Eisenfeld, A. J. (1977) Circ. Res. 4 1 , Suppl.
Lynch, K. R., Simnad, V. I., Ben-Ari, E. T., and Garrison, J. C. (1986) Hypertension 8,540-543
Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
, -
Robertson, J. I. S. (1973) Clin. Sci. Lond. 44,87-90
2,43-46
Moffett, R. B. (1987) Biochim. Biophys. Acta 9 1 2 , 1-8 Moore, D. (1987) in Current Protocols in Molecular Biology (Au-
subel, F. M. Brent, R., Kingston, R. E., Moore, D. D., Smith, J. E., Seidman, J. G., and Struhl, K., eds) pp. 1.7.1-1.7.7, Greene Publishing Associates and Wiley-Interscience, New York
Nasjletti, A., and Masson, G. M. C. (1972) Circ. Res. 30-31,
Nasjletti, A., and Masson, G. M. C. (1973) Proc. SOC. Exp. Biol. Med. 142,307-310
Ohkubo, H., Kageyama, R., Ujihara, M., Hirose, T., Inayama, S., and Nakanishi, S. (1983) Proc. Natl. Acad. Sci. U. S. A. 80,
Ohkubo, H., Nakayama, K., Tanaka, T., and Nakanishi, S. (1986) J. Biol. Chem. 261,319-323
Pfahl, M. (1986) in Biochemical Actions of Hormones (Litwack, G., ed) Vol. XIII, pp. 325-357, Academic Press, Orlando, FL
Reid, I. A., Morris, B. J., and Ganong, W. F. (1978) Annu. Reu. Physiol. 40,377-410
Rigby, P. W. J., Dieckmann, M., Rhodes, C., and Berg, P. (1977) J . Mol. Biol. 113 , 237-251
Robinson, B. G., Frim, D. M., Schwartz, W. J., and Majzoub, J. A. (1988) Science 2 4 1 , 342-344
Sargent, T. D., Wu, J., Sala-Trepat, J. M., Wallace, R. B., and Bonner, J. (1979) Proc. Natl. Acad. Sci. U. S. A. 76,3256-3260
Scheidereit, C., Geisse, S., Westphal, H. M., and Beato, M. (1983) Nature 304 , 749-752
Skeggs, L. T., Jr., Lenz, K. E., Hochstrasser, H., and Kahn, J. R. (1963) J. Exp. Med. 118 , 73-98
Strijker, R., van Assendelft, G. B., Dikkeschei, B. D., Gruber, M., and AB, G. (1986) Gene (Amst.) 45,27-35
Stuzmann, M., Radziwill, R., Komischke, K., Klett, C., and Hackenthal, E. (1986) Biochim. Biophys. Acta 886,48-56
Subramani, S., Mulligan, R., and Berg, P. (1981) Mol. Cell. Biol.
Tanaka, T., Ohkubo, H., and Nakanishi, S. (1984) J. Biol. Chem.
Tewksbury, D. A. (1983) Fed. Proc. 42,2724-2728 Weigand, K., Wernze, H., and Falge, C. (1977) Biochem. Biophys.
Suppl. 2,187-202
2196-2200
1,854-864
259,8063-8065
Res. Commun. 75, 102-110
Continued on next page.

13078 Glucocorticoids Induce Novel Angiotensimgen Transcripts SUPPLEMENTARY MPITERIAL TO
Glucoconroids bdum me lONmvlalion of Novel Angolensmogen Tranmptr
by Elia T. BBn.Ari. KwIn R. L r n and Jmas C. Canhcn
probe: 15.2 E 2.2 E 3.7 E 4.3
28s - 1 8 s -

Glucocorticoids Induce Novel Angiotensinogen Transcripts
uncut Bgl I Nco I Rsa I
3.0 kb - ~~
2.0 kb - 1.6 kb - I 1500- m a b -1375
1.0 kb - 1200- ern
oskb -
C H c n C H c n
probe: 1650 b.p. cDNA 70 b.p. (-115 to -184)
origin - 1 2 3 origin - 1 2 3 4 5
28s - 1 8 s -
- 28s - c - 18s -
hepatocytes in vivo hepatocytes in vivo
13079
- - 2.25 kb