Embed Size (px)
Citation preview

ISSUE No. 49 October 2007
AEPThe magazine of the European Association for Grain Legume Research
Le magazine de l’Association Européenne de recherche sur les Protéagineux
GRAIN LEGUMES

No.33
Degradability of grain legumesin ruminants
No.35
Grain legumes andhuman health
3rd
quar
ter 2
001
1st q
uarte
r 200
2
No.36
Environmental benefitsof grain legumes
2nd
quar
ter 2
002
No.37
Genetic resourcesof grain legumes
3rd
quar
ter 2
002
No.34
Highlights from the4th AEP Conference
4th
quar
ter 2
001
Back issuesBack issues
No.38
Use of syntenyfor genetic progress
4th
quar
ter 2
002
ISSUE No. 392nd quarter 2004
AEPThe magazine of the European Association for Grain Legume Research
Le magazine de l’Association Européenne de recherche sur les Protéagineux
GRAIN LEGUMES
No.39
EU projects on
grain legumes
2nd
quar
ter 2
004
No.40
Highlights from Dijon 2004(AEP-5 and ICLGG-2)
3rd
quar
ter 2
004
No.41
The GRAIN LEGUMESIntegrated Project
Janu
ary
2005
ISSUE No. 42
June 2005
AEPThe magazine of the European Association for Grain Legume Research
Le magazine de l’Association Européenne de recherche sur les Protéagineux
GRAIN LEGUMES
No.42
Drought and saline stress in legumes
June
200
5
No.43
Food uses and healthbenefits of lupins
Sept
embe
r 200
5
No.44
Seed protein in grain legumes
4th
quar
ter 2
005
No.45
Economic and environmentalvalue of grain legumes
1st q
uarte
r 200
6
No.46
Integrated activities in the EU Grain Legumes Integrated Project
2nd
quar
ter 2
006
No.47
Vetches fromFeed to Food?
3rd
quar
ter 2
006
No.48
Fababeans
Janu
ary
2007

3 GRAIN LEGUMES No. 49 – October 2007
CONTENTS
Carte blanche4 Available markets should be supplied – we need to encourage
farmers to enlarge areas (P. Cuypers)
News5 Changes and challenges for AEP (The AEP Executive Committee)
5 Lisbon 2007 – Integrating legume biology for sustainableagriculture – a coming milestone
5 Insert: First GL-TTP workshop in April 2007
6 5th Phaseomics meeting in Varenna, Italy, May 2007 (F. Sparvoli, N. Terryn, W. Broughton)
6 Towards sustainability: eleventh Serbian forage crops symposium(A. Mikic)
Research7 Manipulating flowering time
(B. Wenden, C. Rameau I. Lejeune-Hénaut)
8 Insert: Digestibility of Hr winter peas (K. Crépon)
Special reportAbiotic stress in legumes9 Introduction10 Growth and functioning of legumes under drought: a whole
plant perspective (B. Davies)12 Plant traits and crop management to limit the effects of water stress
in Mediterranean agronomic situations (F. Lelièvre) 14 How genomics may help to understand tolerance to abiotic
stress in legumes? (V. Gruber, M. Crespi)16 Integrating molecular and genetic data in legumes (T. Huguet)17 Benefits of inoculation under abiotic stress in the Mediterranean
region (C. Vargas, M. E. Aouani)18 Abiotic stress breeding at ICARDA (M. Baum)19 Where we go from here – Concluding remarks of the abiotic stress
in legumes workshop(M. Crespi, A. Schneider, T. Huguet, M. E. Aouani)
Crops, uses & markets20 Dynamics and prospects for grain legumes in the European feed
market (A. Schneider, F. Pressenda, K. Crépon)22 Decision support system for efficient disease control
(T. Volk, J.-S. von Richthofen)24 A jump in price for French faba beans exported to Egypt
(J.-P. Lacampagne)
25 Events & new books
Around the world26 Pigeonpea development in China (S. Yang and colleagues)
The magazine of the European Association for Grain Legume Research
Le magazine de l’Association Européenne deRecherche sur les Légumineuses à graines
AEP12, avenue George V, 75008 Paris, France
Grain Legumes magazine is available on subscription to anyone interested
in grain legumes.
Subscription prices and ordering informationare available from the AEP secretariat or the
AEP web site(http://www.grainlegumes.com)
Publishing Director/Directeur de la publication:Álvaro RAMOS MONREAL
(Consejeria de Agricultura Ganadería, Spain)
Managing Editor/Rédacteur en chef: Anne SCHNEIDER (AEP, FR)
Email: [email protected]
Editorial Secretary/Secrétaire derédaction:
Jill CRAIG (AEP, UK)Email: [email protected]
Editorial Board/Comité de rédaction:Jill CRAIG (AEP, UK)
Gaëtan DUBOIS (UNIP, FR)Simon KIGHTLEY (NIAB, UK)
Pascal LETERME (Prairie Swine Centre,Saskatoon, Canada)
Wolfgang LINK (IPP, Göttingen, DE)Diego RUBIALES (CSIC, Córdoba, SP)
Anne SCHNEIDER (AEP, FR)
Overseas correspondents/Correspondants à l’étranger
Brian CLANCEY (Stat Publishing, USA)David MCNEIL (Victorian Department of
Primary Industries, AU)George HILL (Lincoln University, NZ)
Electronic prepress/InfographiePrinting/Impression:
Herald Graphics, Reading (UK)
Subscriptions/Abonnements:AEP Executive SecretariatEmail: [email protected]
Quarterly magazine/Revue trimestrielle
ISSN 245-4710
Commission paritaire n° 74 616
GG RR AA II NN LL EE GG UMEUME SS

4GRAIN LEGUMES No. 49 – October 2007
*UNIP President, Paris, France.([email protected])
Available markets should besupplied – we need to encouragefarmers to enlarge areas
Pierre Cuypers*
In the past three years, grain legume areas in France have decreasedsignificantly: 2007 areas are 50% less for peas and 30% less
for faba beans compared with 2004 areas. In contrast to peas whoseyield also decreased, faba beans benefited from a quite valuable averageyield this 2007 harvest.
There has been a general trend in recent seasons for enhanced damageto arable crops caused by adverse climatic conditions during the cropcycle. However the decrease in grain legume area in France and inother EU countries is especially alarming for farming systems.
Indeed, as highlighted by Mr Christophe Terrain, President ofArvalis1 at a French press conference this summer, it is vital to managecropping decisions at the rotation level, and grain legumes bring aclear benefit in rotations. The Arvalis Acting Director, Gérard Moricepointed out that they enhance the yield of the following cereal by0.7 to 1 t/ha, and have costs €50 to €70 lower than a wheat oroil crop.
We need to respect the cycle of crops on one field, by avoiding thereturn of similar crops too soon, and maintaining the diversity ofcultivated species.
On the market side, UNIP was frustrated to see that there wasa low offer of faba beans and peas for the attractive export outletswhich have reached very high prices this commercial campaign!
Grain legumes compete indirectly with some other crops that benefitfrom a more positive image and from lucrative contracts, but we alsoidentified some logistical difficulties at collection, storage and marketlevels that are linked to minor volumes. That is why we believe indemonstrating actions towards farmers and farmers’ unions and ininnovative partnerships at regional levels to reinforce stability of thecrop chain from the grower to the market users.
In addition, we believe in collaboration with scientists to achievegenetic enhancement or agronomic solutions to overcome diseases andpests or soil and climatic variations, while maintaining high seed quality.
Grain legumes have benefits especially at agronomic andenvironmental levels, so we need to be creative to make themeconomically valuable! �
1The technical institute for cereals and several other crops in France.

5 GRAIN LEGUMES No. 49 – October 2007
AEP NEWS
Changes and challenges for AEPSince autumn 2006, there have been some changes at AEP
headquarters.
A lighter structure for AEP in ParisOne of AEP’s main sponsors, UNIP, which hosts the AEP
headquarters, has experienced difficulties associated with the strongdecline in grain legume areas in recent years in France (as inother EU countries). UNIP has always supported AEP activitiesat financial and political levels and appreciates the successfulnetworking and dissemination efforts that have been carried outby AEP in the past years. However, with the new situation, UNIPcannot continue to support AEP without reductions in the activitiesand finances that go through the AEP office in Paris hosted byUNIP. The consequence is that the Paris headquarters of AEPnow have a lighter structure with only one half-time person andno more secretarial assistance.
In fact, UNIP proposes to take a greater part in AEP activitiesand decisions while ensuring the balance of AEP budgets. AEPhas always wished to involve more representatives from the industrysector. It is expected that this involvement will be followed bysimilar involvement of other market-related organisations oreconomic partners.
Therefore today the AEP network needs to find the best way(i) to operate with a lighter permanent structure for its day-to-day running as well as (ii) to meet the needs and demands of allthe different types of members within AEP.
New challenges for AEP after 15 years ofsuccessful development
Up to now AEP’s priority has been to establish an active networkin the legume scientific community, and AEP has been successfulin raising interest and funds for legume research at EU andinternational levels.
Now a new challenge is to develop additional activities of greaterinterest to economic players who face short and medium termproblems associated with the production and use of grain legumes;this means, identifying the technical issues to translate into questionsfor research for the short and medium terms, but also to preparelong term for the agriculture of the future.
At the same time, the activities of interest to scientists themselvesshould be maintained in AEP and the Executive Secretariat shouldfind a new way to function with smaller resources. This shouldbe discussed with any interested parties who wish to sustain suchan informal and neutral platform for collaboration, a joint voiceand the easiest dissemination of information. The joint activitieswill also be facilitated by the communication tools that havebeen developed by AEP in the recent years (web site, event cycles,GL magazine, etc).
Therefore facing the positive trends in R&D activities, thefuture AEP mandate will be an exciting and challenging periodfor legume enthusiasts!
Join AEP and apply for candidature to the Scientific Committeeto contribute to these challenges, facilitated by the previous effortsand success. �
Source: The AEP Executive Committee.
Lisbon-2007 – Integrating legume biology
for sustainable agriculture – a coming milestone for legumes
We are delighted to announce that our Lisbon-event to be heldfrom 12 to 16 November 2007 has received the patronage
of Mr José Manuel Barroso, President of the European Commission.Mr Timothy Hall, the Acting Director of Biotechnologies,
Agriculture and Food of the European Commission Directorate-General for Research will officially open the conference and thePortuguese Minister of Agriculture and Rural Development, JaimeSilva, will give a welcome address on behalf of the PortugueseGovernment.
This event, the 6th European Conference on Grain Legumesfeatures the final dissemination event of the EU FP6 Grain LegumesIntegrated Project and promises to be a truly stimulating, scientificallyrewarding and sociably enjoyable experience for the 400 participants.
The programme is composed of plenary sessions, workshopsand poster displays, with presentations given by invited speakersand experts from a wide range of different fields, as well as plentyof opportunity for discussions.
This milestone will be the occasion to assess progress in grainlegume research and to prepare future collaborative work on legumes.
More up-to-date information about the programme is availableat: http://www.grainlegumes.com/index.php/aep/events/lisbon_2007/welcome_to_lisbon_2007 �
First GL-TTP workshop in April 2007The Grain Legumes Transfer Platform organised an exciting workshop entitled ‘TargetingScience to Real Needs” which attracted 63 participants from 17 countries (five continents) togather in Paris on 23–25 April 2007.
Reflecting the needs of grain legume breeders, the workshop focused on the exploitation ofgenetic resources, and on the concrete use and integration of molecular technologies inbreeding. The main themes addressed were genetic diversity, disease resistance, abiotic stresstolerance and quality traits.
As a follow-up to the event, GL-TTP is currently working with GL-TTP members to complete thetransfer of information and ready-to-use material presented at the workshop.
Finally, GL-TTP is building on the ideas that resulted from the brainstorming across professionalsectors to set up Research & Development and Technology Transfer projects in partnership withresearch scientists and plant breeders.
More information at http://www.gl-ttp.com

6GRAIN LEGUMES No. 49 – October 2007
EURO AND WORLD NEWS
5th Phaseomics meeting in Varenna, Italy, May 2007
Towards sustainability: eleventh Serbian forage crops symposium
The 5th Phaseomics meeting was held in the beautiful settingof Villa Monasteria, Varenna, Lake Como, Italy. ‘Phaseomics’
is a worldwide network of committed scientists that have ongoingresearch on beans. For more details visit www.phaseolus.net.
The highlights of the meeting were very diverse. There weresessions ranging from seed quality, to evolutionary genetics, genomics,genetic tools and resources, biotic and abiotic stresses, as well asthe interaction with beneficial organisms. Abstracts from themeeting, and interesting links can be found at www.phaseomicsv.net.
Of particular interest were talks from Matthew Blair (CIAT,Cali, Colombia) describing work on the use of wild accessionsto improve common bean varieties (1). This follows a trend setby work on rice (2) that uses wild accessions to improve crops.Despite plant breeders’ initial reluctance, the data shows clearlythat this is a process that also ‘yields’ in grain legumes!
Other interesting talks covered the food and feed qualities ofbean (Francesca Sparvoli and Bruno Campion, Milano, Italy).They have developed and are now evaluating mutants with lowphytic acid contents as well as beans that are lectin-free. Bothcould help improve the digestibility of beans. Nutritional workis in progress to verify this hypothesis.
As for genomics tools, a TILLING population in the varietyBAT93 is being set up by collaboration between three teams(CIAT, University Geneva and USDA/ARS/TARS Puerto Rico).Part of the bean genome will be sequenced and different groupshave contributed to the growing number of bean ESTs in Genbank.
In general the meeting was very successful, and the feelingwas that the future for beans, as the most important food legume,looks very positive. �
Source: Francesca Sparvoli ([email protected]), Nancy Terryn([email protected]) and William Broughton([email protected])
(1) Blair, M. W. et al. (2006). Theoretical Applied Genetics 112, 1149–1163.
(2) McCouch, S. R. et al (2007). Euphytica 154 (3), 317–339.
The eleventh symposium on forage crops of the Republic ofSerbia was held from 30 May to 1 June 2007 at the Faculty
of Agriculture in Novi Sad under the auspices of the SerbianMinistries of Science and of Agriculture, Forestry and WaterManagement. The symposium was organised by the Forage CropsSociety of Serbia, the Institute of Field and Vegetable Crops(IFVCNS) in Novi Sad and the Faculty of Agriculture of theUniversity of Novi Sad (FANS).
With the title ‘Systems of sustainable production and utilisationof forage crops’, the meeting brought together more than 100researchers, with about 70 from Serbia and more than 30 from14 other countries. The three main sessions focused on (1) breeding,seed production and genetic resources, (2) field forage cropsproduction and utilisation and (3) grassland production.
Half the papers were on legumesAbout half of the one hundred papers presented were related
to legumes, confirming the essential role that these crops, togetherwith grasses and other forage crops, play in modern systems offeed production. Some papers focused on single species, and dealtmostly with lucerne, followed by vetches and red clover, whileothers focused on breeding and agronomy.
Following plenary talks on recent achievements and the futuredirection of breeding perennial and annual legumes in Serbia(Dragan -Dukic , FANS, Serbia and Vojislav Mihailovic, IFVCNS,
Serbia) and also a review of the current status and prospects forannual forage and grain legumes in contemporary Serbian agriculture(Branko Cupina, FANS, Serbia), other authors from Serbia presentednumerous interesting contributions on the diversity of legumespecies and their utilisation. Among the highlights of the first twosessions were talks by foreign delegates: the genetics of agronomiccharacteristics in pea (Noel Ellis, JIC, UK), improving forageand seed yield in temperate legumes (Athole Marshall, IGER,UK), common vetch breeding (Rade Matic, SARDI, Australia),breeding sunn hemp (Crotalaria juncea) and sericea lespedeza(Lespedeza cuneata) (Jorge Mosjidis, Auburn University, USA),various cultivar evaluations (Fred Eickmeyer, Saatzucht Steinach,Germany and Piotr Stypinski, Warsaw Agricultural University,Poland), forage potential of a grain legume collection (MargaritaVishnyakova, VIR, Russia), lucerne management in irrigated areas(Jaume Lloveras, University of Lleida, Spain), the tripartite symbioticsystem in legumes (Alexey Borisov, ARRIRAM, Russia), the roleof legumes in sward management (Miluse Svobodová, CzechUniversity of Agriculture) and red clover and other crops asalternatives to grasses for ensilage (Pádraig O’Kiely (Teagasc, Ireland).
All the symposium papers were published as Volume 44 of thejournal, A Periodical of Scientific Research on Field and Vegetable Crops,issued by the Institute of Field and Vegetable Crops and freelyavailable on a CD in pdf. format from Aleksandar Mikic. �
Source: Aleksandar Mikic ([email protected]).
(Pho
to F.
Spa
rvol
i, IB
BA-C
NR,
Mila
n, It
aly)

GRAIN LEGUMES No. 49 – October 20077
RESEARCH
*SGAP INRA, Versailles, France.([email protected],[email protected])
**UMR INRA/USTL SADV, Estrées-Mons,Péronne, France. ([email protected])
Manipulating flowering timeChanger la date de floraison du poisby Bénédicte WENDEN*, Catherine RAMEAU* and Isabelle LEJEUNE-HÉNAUT**
Pea, the most representative Europeangrain legume has limited development
prospects mainly due to unstable yieldswithin years or sites. Stabilising andimproving the productivity of the dry peacrop could be achieved through the releaseof winter cultivars, as has been done, forexample, for wheat and canola.
Towards an ideal winter pea Escape from drought and heat stresses in
the spring and through the earlier floweringperiod, together with a longer developmentcycle leading to higher global biomassproduction, would make the productivityof such winter grown cultivars more reliable(Figure 1).
For an autumn-sown pea however, it isessential to overcome frost which is anotherimportant abiotic stress. For the last 30years, winter dry pea cultivars have beenbred progressively for better frost tolerance,but until now they have never reachedthe level of tolerance expressed by someforage lines, like Champagne or AustrianWinter. Given that frost sensitivity risesafter floral initiation, previous observationshave shown that such frost tolerant peas areable to escape the main winter freezingperiods by delaying their floral initiationunder short days (3).
For the winter pea crop, themanipulation of the flowering genes appearsto be a promising way to control the keydevelopmental stages which are the datesof floral initiation and flowering. The idealwinter pea should initiate its flowerprimordia late enough to avoid winter frostsand should flower early enough to escapedrought and heat stresses in late spring.
Regulating flowering time Since the early 1950s, pea has been used
as a model species for the study of thegenetic control of flowering time throughthe physiological characterisation of mutantsobserved in different environmentalconditions (long days, short days) and theuse of grafting experiments. These studiesresulted in a classical model which iscurrently being reconciled with themolecular genetic models developed inArabidopsis by J. Weller’s group at theUniversity of Tasmania. The fact that mostof the flowering genes identified inArabidopsis are present in legume genomesequence databases (2) suggests that similarmechanisms are likely to occur betweenpea and Arabidopsis, and that already wehave the tools to understand and to controlflowering time in pea. In particular, theresponse to photoperiod in pea could, asfor Arabidopsis, also involve the circadianclock and the precise cyclic regulation ofphotoperiodic genes (CONSTANS) (5).
The winter pea breeding strategydeveloped at INRA relies partly on themanipulation of two major genescontrolling the natural variation forflowering time, namely LF and HR. Thesegenes are promising targets because theyare known to control, respectively, theintrinsic earliness for the beginning offlowering and the strength of the response
to the photoperiod. More precisely, thedominant allele HR is known to delay thefloral initiation of autumn-sown peas untila longer daylength is reached (approximately13 h 30 min) in the following spring. Thishelps to escape the main winter freezingperiods. The knowledge of the sequenceof these genes will allow the developmentof molecular markers suitable for buildingoptimal associations of the HR and LFalleles. A candidate gene approach hasalready led to the cloning of the LF geneas the homologue of the TFL1 Arabidopsisgene (1), and molecular markers are beingdeveloped to follow the different allelesin crosses. For HR the same approach isbeing used, thanks to the microsyntenywith Medicago truncatula, in order to clonethe gene and/or develop molecular markersto backcross the high photoperiod responsedominant HR allele in agronomic geneticbackgrounds.
Developing integrative toolsTo understand the biology of complex
systems it is necessary to integrate atdifferent scales the existing models,experimental data, and main hypothesesinto new models that are able to describe,explain and predict biological phenomena.A PhD research project is ongoing at INRAVersailles to develop an integrative toolcombining molecular, physiological and
Spring pea
Traditional winter pea
Winter pea ideotype
Autumnrains
Wintercold stress
Sowing Floralinitiation
Yield maturity
Heat and droughtstresses
HR :Photoperiod
sensitivity
LF :Endogenous
precocity
Figure 1. The winter pea strategy: manipulating the dates of floral initiation and flowering in order to escapewinter frosts as well as drought and heat stresses in late spring.

RESEARCH
8GRAIN LEGUMES No. 49 – October 2007
ecophysiological data into a computer modelof flowering time in pea. This model willintegrate the hypotheses that best representour current understanding of the geneticand physiological regulation of floweringtime from the genetic to the crop level.
Presently, two categories of floweringmodels can be described for pea. First, anagroecophysiological model has beenestablished, primarily for two pea varieties,in which the time of flowering has beenbroken down into two componentvariables: the leaf appearance rate and thenode of the first open flower (4). However,the response of flowering to theenvironment also depends on genotype andtherefore extrapolation of these results toother environmental conditions orgenotypes is limited. Secondly, geneticmodels of the control of flowering in peahave been based on the analyses of floweringmutants (see above) describing theinteractions of signals that lead to theproduction of a flowering signal and thus
to floral initiation (6), but these geneticmodels have no prediction capacity.
The aim of the PhD project is to gatherall the sources of knowledge in the formof mathematical equations that will togetherallow the simulation of as many differentenvironmental and genetic conditions aspossible. In particular, this work shouldenable the effects of the different genes,and their interaction with environmentalconditions, on flowering time to bequantified. The modelling approach is basedon two sets of data (Figure 2): first thegenotype of the key flowering genes (LF,HR, SN, DNE, GI) and secondenvironmental data (photoperiod and meantemperature). For this model, we assume,as it was shown in Arabidopsis, that theproduction of the flowering signal isquantitatively controlled by an integratednetwork of pathways. The response offlowering to photoperiod is modelledthrough the effect of cycling genescontrolled by the circadian clock and the
coincidence between their level ofexpression and the daylength. Theflowering signal accumulates over time untilit reaches a flowering threshold, determinedby the LF allele, and then triggers floralinitiation at the apex. The main outputs ofthis integrative model are the node andtime of floral initiation as well as the nodeand time of flowering. These variables couldin turn be integrated as a prediction offlowering into a crop growth model.
How can this model be used?Knowledge of the genetic and
physiological basis of the interactionbetween flowering genes and theenvironment is essential to the selectionof varieties closely adapted to a specificenvironment. This work will lead first toa better comprehension of the effects ofthe different flowering genes in pea, andtheir interaction with the environment.
Then, with a better understanding andprediction of flowering time, new ‘virtual’pea genotypes could be constructed andtheir flowering phenotypes predicted underdifferent environmental conditions. Bytesting different LF/HR allele associations,under virtual winter conditions, we will beable to predict the best winter pea floweringgenotype for a given environment and toprovide precise targets to the breeders. �
(1) Foucher, F. et al. (2003). Plant Cell 15,2742–2754.
(2) Hecht, V. et al. (2005). Plant Physiol 137,1420–1434.
(3) Lejeune-Henaut, I. et al. (1999). Euphytica109 (3), 201–211.
(4) Truong, H. H. and Duthion, C. (1993). AnnBot. 72(2), 133–142.
(5) Weller, J. (2005). Flowering Newsletter 40,39–42
(6) Weller, J. L. et al. (1997). Trends in PlantScience 2 (11), 412–418.
Data Integrative tools Predictions
GenotypeFlowering keygenes
•LF•HR
Environment
Photoperiod
Temperature
ModellingGenetic network
Circadianclock
Otherpathways
SignalsCycling genes
Production of a flowering signal
Flowering threshold (LF)
Time or Node(developmental rate)
Node offloral
initiation
Floralinitiation
time
Floweringnode
Floweringtime
Figure 2. An integrative model to predict the flowering time in pea.
Insert
Digestibility of Hr winter peasPea breeders have started to breed varieties sensitive to photoperiod(length of the daylight) since they show an advantageous longerperiod of frost tolerance. This photoperiod sensitivity is controlledby the Hr gene.
In parallel, the nutritional value of these types of peas for pigshas been tested by UNIP and Arvalis Institut du Végétal in France,by comparing one classical spring pea (SP) to both one linesensitive to the photoperiod (HR1) and one line with a better
agronomic value (HR2) issued from the cross between HR1 and aspring variety.
The two winter type varieties (HR1 and HR2) have a lower trypsininhibitor activity compared with the one of spring type. The aminoacid profiles of HR1 and HR2 are similar and slightly different fromthe one of SP but are very close to the usual value for peas.
The protein digestibility is 82% in HR2, slightly higher than for HR1 (80%) and SP (78%) but without significant difference.
The amino acid digestibility is high and similar in the three types.
Therefore the genetic character which modifies the timing ofthe floral initiation does not modify the chemical compositionor digestibility of the proteins and amino acids in pigs.
Source: Katell Crépon, UNIP, [email protected]

GRAIN LEGUMES No. 49 – October 20079
Legumes are sensitive to abiotic stresses such as waterdeficit and soil salinity. Tolerance and susceptibility to thesestresses are very complex traits and plants show complex
cell signalling pathways activating metabolic functions anddevelopmental switches to cope with environmental stresses.
Hence, international experts working on the analysis ofplant tolerance were gathered at a scientific workshop ‘Abioticstresses in legumes’ (Tunis, 22–24 March 2007, supported byGLIP1). Basic scientists, molecular biologists, genomicists,physiologists and agronomists involved in several ongoingEU projects were present together with representatives ofinternational agricultural organisations. Legume experts andtheir colleagues working on non-legume plants came togetherso as to learn from a variety of stress response models:agronomic, eco-physiological and genomic models. In thelatter case, the well known Arabidopsis model has allowedtremendous progress in the genomic analysis of abiotic stresseseven though several physiological characteristics of legumesare not found in this plant. More recently Medicago truncatula,an annual, diploid and autogamous legume, has been adoptedas a model legume in various European and USA laboratoriesand diverse genetic, genomic and reverse genetic tools arebeing developed for the analysis of physiological responsesat genomic level.
This special report2 brings together some highlights of thisworkshop and presents current perspectives of the way plantscope with abiotic stresses with particular emphasis on theimplications for grain legume crops. The data discussed dealwith whole-plant approaches and crop management as well asgenetic and genomic analysis of abiotic stress responses. �1the EU Grain Legumes Integrated Project www.eugrainlegumes.org 2a previous dossier dealt with the same topic: Special report (2005)Drought and saline stress in legumes, Grain Legumes 42, 13–22.
Les légumineuses sont sensibles aux stresses abiotiques commele déficit hydrique et la salinité du sol. La tolérance et lasusceptibilité à ce type de contraintes sont des caractères
complexes et les plantes présentent des processus d’échangescellulaires élaborées, activant des fonctions métaboliques et deschangements de développement pour gérer les stressesenvironnementaux.
Il a été décidé de rassembler plusieurs experts internationauxtravaillant sur l’analyse de la tolérance végétale à un séminairescientifique ‘Les stress abiotiques et les légumineuses’ (Tunis,22–24 mars 2007, co-organisé par GLIP1), avec des scientifiquesplus fondamentaux, des génomiciens, des physiologistes etdes agronomes. Les experts légumineuses et leurs collèguesspécialistes des plantes modèles ont partagé leurs différentsschémas d’analyse de la réponse végétale au stress :agronomiques, éco-physiologiques ou génomiques. La plantemodèle Arabidopsis a notamment permis des avancéesconsidérables sur la génomique de la réponse au stress mêmesi cette plante n’a pas certaines fonctionnalités des légumineuses.Plus récemment, Medicago truncatula, une légumineuse annuelle,diploïde et autogame, a été adoptée comme modèle deslégumineuses par différents laboratoires européens et américains,et des outils génétiques et de génomie fonctionnelle ont étédéveloppés pour l’analyse des réponses physiologiques.
Le dossier de ce numéro2 rapporte quelques informations saillantesdu séminaire de Tunis, analysant les différentes façons dont lesplantes gèrent les stress abiotiques, en insistant sur les conséquencessur les cultures de légumineuses. Sont traités à la fois les approchesintégrant la gestion de la culture, la compréhension de laphysiologie de la plante dans son ensemble et les analysesgénomiques des réponses de la plante au stress. �
1le projet intégré européen sur les légumineuses à graines www.eugrainlegumes.org 2un dossier précédent traitait aussi de ce sujet: Special report (2005) Drought andsaline stress in legumes, Grain Legumes 42, 13–22.
Salt effect on the growth of Medicago truncatula F83005.5 line. Left: no salt, right: 45 mM NaCl. Abiotic stress in field trial of faba beans at El-Sharkiain Egypt.
(Pho
to T
. Hug
uet,
INP-
ENSA
T, F
ranc
e)
SPECIAL REPORT
Abiotic stress in legumesLes stress abiotiques et les légumineuses
(Pho
to A
. Sch
neid
er A
EP)
Abiotic stress in legumesLes stress abiotiques et les légumineuses

GRAIN LEGUMES No. 49 – October 200710
SPEC IAL REPORT
ABIOTIC STRESS
fixation, vegetative growth and developmentand aspects of reproductive developmentare most sensitive to soil drying.
A high leaf area index is vital to maximisethe interception of radiation forphotosynthate production. Restrictions incanopy development under drought, andlimitation in vegetative growth before a plantcommunity has covered the ground can bevery damaging for carbon (C) accumulationand yielding. Furthermore, covering thesoil early in the growing season can restrictsoil water loss significantly and therebyenhance the transpiration (T) componentof evapotranspiration (ET), resulting inmore productive use of water (16). Onlywater taken up by the plant can contributeto C gain (W in Equation 1 below).
TurgorCellular water status (turgor) provides
the driving force for organ growth andusually leaf growth becomes restricted asthe soil dries because leaf turgor falls belowa threshold value for leaf expansion. Somegenotypes accumulate solute in cells as shootwater potential falls, thereby maintainingturgor and the potential for growth.However, the maintenance of leaf turgordoes not usually result in maintenance ofshoot growth as the soil dries. A goodcorrelation between restricted shoot growthand high solute accumulation in shootsindicates that some variable other than cellturgor can restrict cell growth under drought.In this situation solutes accumulate as a resultof restricted leaf growth, rather thanproviding the driving force for cell expansionas the soil dries (7). Solute accumulationcan prolong the period before cells die andin some dryland legumes, postponing celldeath by this method is adaptive (5).
All crop plant species are highlysensitive to soil drying, withreductions in productivity
occurring before the amount of wateravailable to the plant has changedsignificantly. Sensing involves both reduceduptake of water from the soil and/or themodified production and transport of a rangeof chemical signals in roots in contact withdrying soil. Both of these responses canprovide information to the shoots on soilwater availability. These changes in chemicaland hydraulic signalling allow plants toregulate growth and development as afunction of resource availability to the roots.
Plant breeders believe that characteristicsimportant for yield in resource-poorenvironments have more to do with growthunder favourable conditions than withresistance to stress per se. As there is littlegrowth by plants once stress develops, overallproductivity is best improved by maximisinggrowth in favourable times (10). Processesthat help the plant survive severe cellulardehydration may be important in perennialcrops but to sustain yields of annuals weneed to understand the regulation of growthand development under moderate droughtstress, rather than focussing on survival andtolerance of low plant water content.
Drought regulates a varietyof plant processes
The sensitivity of different aspects of theplant’s physiology and biochemistry todrought can vary substantially; lesions inmost processes will contribute to reducedyields and/or crop quality. Nitrogen (N)
There is often some root growth in dryingsoil, even when shoot growth is entirelyrestricted. As a result the plant root:shootratio is increased. The development of waterand nutrient depletion zones in soilsurrounding slowly growing roots(particularly when roots clump in compactedsoil) restricts resource uptake from dryingsoil. In these circumstances sustained rootgrowth sustains water and nutrient uptake.
Selection for solute regulation capacityin roots may be a way of developing cropsfor dryland regions (11) and many haveshown the positive impact of root turgormaintenance on growth (3). Sustained rootgrowth in drying soil may also requireaccumulation of the plant hormone abscisicacid (ABA) which prevents run-awayethylene production at low water potentials
Growth and functioning of legumes under drought: a whole plant perspectiveCroissance et fonctionnement des légumineuses sous déficithydrique : une approche à l’échelle de la planteby Bill DAVIES*
*Lancaster Environment Centre, LancasterUniversity, UK. ([email protected])
What is water stress?Successful growth and development of landplants requires maintenance of a favourablecellular water status in even the mostchallenging of climates. The dry airsurrounding the above-ground parts of theplant promotes rates of water loss(transpiration) from the shoots that potentiallycan be very high. Plants must protectthemselves against excessive water loss (usingwater-proofing of aerial plant surfaces andstomatal pores of variable aperture). This willhelp avoid the damaging effects of stresswhich will inevitably limit all aspects of growthand development and ultimately result in celldeath. These stresses may be low cell watercontents (low water potential) and also thebuild-up of growth-inhibiting chemicals.Sustained water uptake from soil and transportof this water to leaves is also required toreplace water lost via transpiration andavoid the development of damaging deficits.

GRAIN LEGUMES No. 49 – October 200711
SPEC IAL REPORTABIOTIC STRESS
(15). In many circumstances ethylene is apowerful growth inhibitor (13) and, as thesoil dries, increased soil strength may alsolimit shoot growth through up-regulationof ethylene synthesis. Ethylene accumulationcan also restrict nodulation significantly.Low ethylene transgenics show reducedleaf growth sensitivity to compacted soiland to soil at low water potential (14).
Three key cropping variablesAn analysis by Passioura (9) highlights
three key cropping variables that willcontribute to sustained crop yield (Y) underdrought:
Y = WUE x W x HI (Equation 1)
WUE = plant biomass produced per unitof water used, W = water available, and HI= harvest index (proportion of biomassproduced as grain). We have seen abovehow W can be manipulated to sustain Y.WUE can be enhanced, for example, byselecting for or manipulating particularaspects of photsynthesis or stomatalbehaviour. Crop yield can also be sustainedby ensuring that water is available at keydevelopmental periods so that a greaterproportion of crop biomass is yield (HI).
Recent work shows the importance ofsustaining C supply to developing maizegrains immediately after anthesis. These Callocation processes are extremely sensitiveto water availability and if water relationsare adverse during this critical period ofcrop development then grain yield fails (1),in spite of favourable water relations anddevelopment of the vegetative crop.
N fixation and droughtSoil drying can reduce nodule mass
substantially, having a negative impact on Naccumulation through fixation. Interestingly,Rhizobia are quite resistant to soil dryingand their survival is probably not a limitingstep for N fixation under drought, eventhough symbiosis establishment is extremelysensitive to drought. Despite the apparentrobustness of Rhizobia in water-limitedenvironments, data indicating superiorperformance of some, often indigenous,strains in dryland environments are limited.
In many legumes, N fixation per unitmass of nodules is highly sensitive to drought,often more sensitive than other plantphysiological and developmental processes(12). Carbon and oxygen limitation in thenodules and/or feedback regulation of N
fixation by N accumulation are possibleexplanations for this sensitivity, linked directlyto changes in the water status of the plant.However, nodules obtain most of their waterfrom the phloem rather than the xylem.Variation in phloem flow can be highlysensitive to small changes in plant water statusand can impact severely on nodule activity.
Variation in phloem and xylemfunctioning could also affect chemicalregulation of nodule activity. For example,if an increase in the concentration of phloemcontents occurs at reduced fluxes, assimilatesdelivery to nodules may be sustained.However, export of N products from nodulesvia the xylem may be decreased by stress,resulting in N accumulation and N feedbackinhibition of nodule activity. Suppressionof N fixation of this kind can result fromlocal signalling (8) or a more systemic signalimported by the phloem (12).
Manipulation of chemicalsignalling
There is significant genetic variationbetween cultivars in N-fixation sensitivity tosoil drying, but there is still some uncertaintyover the basis of this variation. Until thereis some agreement over the mechanismby which soil drying reduces N-fixationactivity, the opportunities for manipulatingthis activity will remain restricted, and wewill not be able to benefit from thedevelopment of novel biotechnologicaltechniques for plant improvement.
Increased understanding of chemicalsignalling mechanisms in drought-affectedplants raises the possibility that artificialmanipulation of signal synthesis andaccumulation may enhance plantperformance when water is restricted. Thismay be achieved via plant improvement orvia a change in management practice, forexample, via deficit irrigation. One of thesetechniques, partial root drying, has beendesigned so that some plant roots are alwaysin contact with drying soil and will thereforecontinue to produce chemical signals. Usingdeficit irrigation, signal production can beenhanced or decreased at critical periodsof plant development resulting in restrictedshoot growth and water use with very littlelimitation in reproductive yield (increasedHI – Equation 1) (4). Other manipulations,such as addition of specific bacteria to therhizosphere (2) or fertiliser treatments willaugment or restrict other chemical signals
in plants. These could, for example, ensurerapid canopy development in drying soil(16) or ultimately promote plant senescencethereby enhancing partitioning of C andN to increase HI of grain crops (17).
Opportunities for plantbiologists
A range of plant stress-sensing mechanismshas evolved so that, during mild soil drying,plants start to shut down growth andfunctioning to save scarce water for criticalphases of their development, even whenthere is still water available in the soil. Thissuggests that there is some scope for plantbiologists to over-ride these defensiveresponses with the benefit of sustained plantdevelopment and yielding as the soil dries.Further elucidation of the drought-sensingsystem to increase the efficiency of wateruse in agriculture (‘more crop per drop’)is most relevant as rainfall patterns aredisrupted by climatic change. �
(1) Boyle, M. G. et al. (1991). Crop Science 31,1246–1252.(2) Belimov, A. A. et al. (2007). Journal ofExperimental Botany 58, 1485–1495.(3) Davies, W. J. (2007). In: Advances inmolecular breeding towards salinity and droughttolerance, 55–72 (Eds M. A. Jenks et al.).Springer, Heidelberg, Germany.(4) Davies, W. J. et al. (2002). New Phytologist153, 449.(5) Flower, D. J. and Ludlow, M. M. (1986).Plant Cell and Environment 9, 33–40.(6) Kirda, C. et al. (1989). Plant and Soil 120,49–55.(7) Kuang, J. B. et al. (1990). Journal ofExperimental Botany 41, 217–221.(8) Marino, D. et al. (2007). Plant Physiology143, 1968–1974(9) Passioura, J. B. (1977). Journal of theAustralian Institute of Agricultural Science 43,117–121.(10) Richards, R. J. (1993). In: Plant responsesto cellular dehydration, 211–223. (Eds T. J.Close and E. A. Bray). American Society ofPlant Physiologists, Rockville, USA.(11) Serraj, R. and Sinclair, T. R. (2002). PlantCell and Environment 25, 333–334.(12) Serraj, R. et al. (1999). Journal ofExperimental Botany 50, 143–155.(13) Sharp, R. E. (2002). Plant Cell andEnvironment 25, 211–222. (14) Sobeih, W. et al. (2004). Journal ofExperimental Botany 55, 2353–2364.(15) Spollen, W. G. et al. (1993). In: Waterdeficits: plant responses from cell to community,37-52. (Eds J. A. C. Smith and H. Griffiths)BIOS Scientific, Oxford, UK.(16) Turner, N. C. (2004). Journal ofExperimental Botany 55, 2413–2425.(17) Yang, J. (2001). Agronomy Journal 93,196–206.

GRAIN LEGUMES No. 49 – October 200712
SPEC IAL REPORT
ABIOTIC STRESS
Plant traits and crop managment to limit the effectsof water stress in Mediterranean agronomicsituationsCaractères des plantes et conduite des cultures pour limiterl’impact de la sécheresse en agriculture méditerranéenneby François LELIÈVRE*
*INRA, UMR SYSTEM, Montpellier, France.([email protected])
Water deficit is the main constraintfor agricultural production inMediterranean environments.
Genetic improvement and agronomicoptimisation of harvested yield, HY(g DM/m2) depend on three termsaccording to the relation (Equation 1)proposed by Passioura (3, 4):
Equation 1: HY = T x WUE x HI (with DMt = T x WUE)
where T (g H2O/g DM per m2) is the croptranspiration, WUE is average water useefficiency (g DM/kg H2O), HI is the harvestindex, and DMt (g DM/m2) is the totaldry matter elaborated. If E (g H2O/m2) isthe soil evaporation, ET = E+T is the actualevapo-transpiration of the crop during itscycle. WUE is often related to aerial drymatter, DMa, instead of DMt (aerial +underground). All these variables are sumsor means for the whole cycle of the crop,representing several months or even yearswhen pluri-annual yields are considered.The improvement of each term T, WUEand HI has limits which are discussed.
Increasing and stabilisingharvest index
In annual grain crops in all environments,yields have progressed in the last half centurythrough continuous genetic HI increase(from 0.1–0.2 to around 0.5 in manyspecies). In rainfed semi-arid environments,subsequent improvement of HI stability has
been obtained in winter cereals by plantbreeding through: (i) genetic control ofvegetative growth and stem length; (ii)drought resistance of pollination-fertilisationphase; (iii) drought escape of the grain fillingperiod through earliness and deep rooting;(iv) increased resistance of late photosynthesisand late translocation during grain filling;(v) improved diseases resistance. Much lesssignificant are the results obtained in HIstability of grain legumes, but progress shouldbe obtained by following the same objectives.
Optimising crop transpirationCrop transpiration (T) can be increased
through plant cycle duration (earliness)within limits in one year. In temperate areas,annual rainfall (Pa) generally exceeds annualpotential evapo-transpiration (ETpa); actualtranspiration (Ta) is limited by ETpa
(Ta≤ETpa≤Pa). In Mediterranean areas ETpa
exceeds Pa which is the upper limit of Ta
(Ta<Pa<ETpa). To optimise yield, the croptranspiration, Ta must be as close as possibleto Pa. This requires the unproductivefractions of Pa (run-off, drainage,evaporation) to be minimised through plantmaterial traits and agronomical conditions:crop cycle matching the rainy period; earlyautumn sowing; vigorous establishment ofthe cultivated plants at the onset of the wetseason (rapid extension of canopy to coverthe soil; high root growth rate); continuousweeding; rainfall well distributed throughthe growth period; high soil permeability;important final root depth; high soil waterreserve; drought resistance at the end ofthe wet season. Even when all theseconditions are fulfilled, Ta tends to have an
upper limit between 60% and 85% of Pa
because soil evaporation, Ea, cannot beavoided during the non-growing period(hot dry summer) and at early stages ofautumn growth when the wet soil surfaceis not covered by the canopy. InMediterranean cultivated fields, Ea valuesoften range between 50 and 150 mm (5).The unproductive fraction of annual rainfallis at least 15–35%, increasing with aridity.It is much higher than in temperate climates,
Creeping self-reseeding annual legumes tested tointercrop vineyards on stony shallow soils insouthern France.
(Pho
to F
ranç
ois
Leliè
vre
INRA
Mon
tpel
lier,
Fran
ce)

GRAIN LEGUMES No. 49 – October 200713
SPEC IAL REPORTABIOTIC STRESS
where the non-growing period is winter. Recent work on wheat in semi-arid
conditions has demonstrated that earlyautumn sowing combined with genotypeswith high leaf and root extension rates at low temperatures (autumn, winter, early spring) are very efficient to increaseTa if the balance between reproductive and vegetative growth is controlled bygenetic factors (6). A similar approach is valuable for winter grain legumes in the Mediterranean.
Optimising water useefficiency
Water use efficiency (WUE) is theeffectiveness of a crop in H2O/CO2
exchange, the value representing a verylong and complicated process ofdevelopment and growth in real conditions(1). Values of 3–5 g DM/kg H2O representhigh utilisation of water by agriculture,from which approximately a half can beallocated to seeds in grain crops. Such levelsare not possible everywhere at any time,because WUE follows biophysical laws: itdecreases under high temperatures and highclimatic demands (2–3 times lower in hotcompared with cool seasons/climates). Inhot periods or climates, it is higher in C4than in C3 plants. High nutrient availabilityincreases WUE, especially nitrogen forcereals and grasses and phosphorus for
legumes. In North Africa, WUE ofirrigated alfalfa is 3.0–4.0 g DM/kg H2Oin spring but falls to 0.5–1.0 in summer.In annual legumes, adaptation to earlyautumn sowings with efficient nitrogenfixation and growth at low temperaturesis important. Conventional selection forDM yield in rainfed field plots selectsefficiently for WUE because high yieldingmaterial of similar earliness useapproximately the same quantity of water(2). However, conventional breeding forDMt and WUE increase is slow becauseof low variability in high yielding materialin a species. Molecular engineering shouldprovide opportunities to modify H2O/CO2
exchange (1).
The case of a perennialforage crop
Harvested yield for one year (HYa b =Ta x WUEa) is generally the sum of severalcuts (growth cycles at different developmentperiods having different daily Tj and WUEj.For pluri-annual forage crops like alfalfa,yield is the sum during n years: HYn = Sn
(HYa).In temperate areas, forage yield HYa is
maximised when Ta tends to ETPa (dailyTj = ETPj throughout the year). It requires:(i) high plant density; (ii) early start ofgrowth in spring and late end in autumn;(iii) drought resistance in summer. Plants
must have the ability to maintain optimalor significant T, growth and yield duringmoderate dry periods that do not exceedtwo months with cumulated climaticdeficits (ETp-P) not lower than –250 mm.It is obtained through depletion of deepsoil water reserves, mainly conditionedby soil and root depth.
In rainfed Mediterranean areas, adaptedplants must alternate: (i) a productivestrategy with high T and high WUE duringthe cool rainy season (5–9 months, P:300–700 mm, ETP: 300–600 mm); (ii) aconservative strategy (survival) withoutgrowth, with partial senescence and verylow T, during the long hot dry season (3–7months, P: 0–100 mm, ETP: 400–1000mm). During this period, surviving organslose water which is compensated for byextraction from the soil to maintainhydration above the lethal level for as longas possible. This conservative transpiration(Ts), not used for direct production, isthe ‘water cost’ of summer survival andperenniality, in addition to soil evaporation.Adaptation in perennials depends on acombination of plant traits like deep rooting,different levels of summer dormancy, andrelative desiccation tolerance of survivingorgans (7, 8). Interest and limits of perennialvs. annual (reseeded or self-reseeding) foragelegumes and grasses must be evaluated atthe farming systems level. In this area, thegap from plant physiology and moleculargenetics studies to the availability ofinnovative cultivars as commercial seedmust be reduced. �
(1) Bacon, M. A. (Editor) (2004). Water useefficiency in plant biology. Blackwell PublishingCRC Press, Oxford, UK. 327 p.
(2) Lelièvre, F. (2006). In: Proc. of Eucarpiaconf., fodder crops and amenity grasses section,Peruggia, Italy, 3–7 Sept 2006, 279–283 (Eds D.Rossellini and F. Veronesi). Univ. di PerugggiaPubl., Peruggia, Italy.
(3) Passioura, J. B. (1977). J. Austr. Inst. ofAgric. Sc. 43, 117–121.
(4) Passioura, J. B. (2002). Functionnal PlantBiology 29, 537–546.
(5) Passioura, J. B. (2004). In:Water useefficiency in plant biology, 302–321 (Ed. M. A.Bacon). Blackwell Publishing, Oxford, UK.
(6) Richards, R. A. et al. (2002). Crop Science42, 111–121.
(7) Volaire, F. et al. (2005). Annals of Botany 95,981–990.
(8) Volaire, F. and Norton, M. (2006). Annals ofBotany 98, 927–933.
Trial to intercrop vineyards with perennial legumes and grasses on shallow soils in French Catalogne (southern France).
(Pho
to F
ranç
ois
Leliè
vre
INRA
Mon
tpel
lier,
Fran
ce)

GRAIN LEGUMES No. 49 – October 200714
SPEC IAL REPORT
ABIOTIC STRESS
How genomics may help to understand tolerance toabiotic stress in legumes? Comment la génomique contribue t-elle à comprendre latolérance aux stress abiotiques chez les légumineuses?by Véronique GRUBER* and Martin CRESPI*
soil generate a low water potential zonemaking it increasingly difficult for the plantroots to acquire both water and nutrients.Both hyperionic and hyperosmotic stressinduce drastic metabolic changes that canlead to plant death. In legumes, salt stressalso inhibits nitrogen fixation, affectingtotal nitrogen uptake in the legume host aswell as its contribution to the soil combinednitrogen content.
Abiotic stress activatescomplex signalling pathways
To cope with these stresses, plants haveevolved complex cell signalling pathwaysactivating metabolic functions anddevelopmental switches to permit theiradaptation to these conditions. The plantroot system, which is the primary site ofperception of environmental changes, isable to adapt its growth and architecture.This adaptation is a result of integratedevents occurring at all levels of organisation,from anatomical to morphological, andto cellular, biochemical, and molecular.Cellular responses to stress include changesin the cell cycle and cell division,modifications in the endomembrane systemand vacuolisation of cells, and changes incell wall architecture, all leading to enhancedstress tolerance of cells. At the biochemicallevel, plants alter their metabolism in variousways to accomodate environmental stresses,including the production of osmoregulatorycompounds.
The molecular events linking theperception of a stress signal with the genomicresponses leading to tolerance have beeninvestigated intensively in recent years,mainly in Arabidopsis. However, the universalnature of these mechanisms is still debated
because different strategies may be used bydifferent plants to cope with environmentalstresses. Insights into gene functions andregulatory control of biological processesthat are associated with stress responses inlegumes have been facilitated with thedevelopment of genomic resources andinformation for the two model legumespecies, Medicago truncatula and Lotusjaponicus (7, 8). For each of these tworeference legumes, extensive investmentsare being made to generate a full range ofgenomics resources, including sequencingthe entire genome and thus giving accessto a complete description of the responseat gene expression level. The increasingavailability of genetic and genomic data aswell as the high degree of synteny betweenlegume genomes has resulted in these twospecies becoming valuable models for themolecular genetic study of abiotic constraintshampering yield, since they are very closeto legume crops. Indeed, combininggenomic and biological knowledge aboutreference legumes that has a bearing onother food and feed legumes of majoreconomic importance, presents a majorscientific opportunity that can impact uponother less-easily handled crops.
Transcriptome analysis givespromising results
A critical step of the cell signallingpathways controlling stress responses involvestranscriptional regulation, generally mediatedby transcription factors that may governand coordinate the expression of large groupsof genes. In legumes, extensive sequencinghighlighted around 2,000 transcriptionfactors per genome, less than 1% of themgenetically characterised (9) and only six
Legumes provide more than one-thirdof mankind’s nutritional nitrogenrequirement and represent about
25% of the world’s major crop production(6). Legumes can interact symbioticallywith nitrogen-fixing soil bacteria to fixatmospheric nitrogen, reducing productioncosts and environmental damage becausethey do not require soil fertilisers. Inaddition, the roles of legumes in the wholecrop rotation (for N cycle, soil structureand activities, disease and weed break, etc)benefit other subsequent non-nitrogenfixing crops. Therefore legumes play anessential role in sustainable agriculture.
Only 10% of arable land is consideredto have non-stressed soils, implying thatcrop growth on the remaining 90% of arableland is submitted to diverse environmentalstresses (3). Indeed, various abiotic stressesincluding salinity, drought, extremetemperatures (heat, freezing, chilling),waterlogging and mineral toxicities, reducethe yields of most major crops by morethan 50% (1). Abiotic stresses affect legumegrowth, yield and productivity as well asthe efficiency and function of the symbioticnitrogen fixation process. In general,exposure of plants to dehydration or osmoticstress causes a decline in photosyntheticactivity due to stomatal closure and reducedactivity of photosynthetic enzymes limitingcarbohydrate metabolism and plant growth.On a world scale, soil salinity is also aprimary cause of crop yield losses on morethan 20% of agricultural land and 50% ofcropland (4). High salt depositions in the
*Institut des Sciences Végétales – CNRS, Gif-sur-Yvette and Université Paris Diderot, France.([email protected], [email protected])

GRAIN LEGUMES No. 49 – October 200715
SPEC IAL REPORTABIOTIC STRESS
involved in abiotic stress tolerance (drought,salinity, extreme temperature). Thedevelopment of novel powerful tools fortranscriptome analysis increased considerablythe access to gene functions on a globalgenome-wide scale for the identificationof differentially expressed genes in responseto stress in legumes. These tools includesuppression subtractive hybridisation (SSH)library, super serial analysis of gene expression(SuperSAGE) for genome-wide quantitativegene expression profiling, array-basedtranscript profiling technologies (measuringexpression levels of tens of thousands of genesin parallel), and massive quantitative real timePCR (qRT-PCR) with specific in silicodesigned primers against known genes, suchas transcription factors (9). Recently, aMt16K microarray covering 16,086 tentativeconsensus sequences derived mainly fromapproximately 164,000 M. truncatula ESTscollected in the TIGR M. truncatula GeneIndex 5 (http://www.tigr.org/tdb/mtgi),was obtained. This Mt16K microarray wasused to monitor, for example, changes inthe transcriptome of dessication-sensitiveradicles of M. truncatula seeds at differentpoints in time, and therefore to investigateregulatory processes and protectivemechanisms leading to dessication tolerancein seeds (2). Another example is theapplication of SuperSAGE technology toprofile transcripts of drought- and salt-stressed roots and nodules from chickpeagiven access to a series of genes exclusivelyexpressed under both stresses, but not innon-stressed controls.
Novel regulatory genesrevealed by genomics
Thus, genomics and transcriptomicsprovide powerful tools to fully understandthe molecular basis of responses to abioticstresses in legumes as well as to the capacityof the plant to cope with environmentalstresses. This may lead consequently to theidentification of regulatory genes able tomodify crop behaviour under adverseconditions (as schematised in Figure 1). Asa recent example, expression analysis inmodel Medicago plants using a ‘dedicated’macroarray (containing selected genes fromSSH libraries), revealed novel regulatorygenes, including transcription factors,associated with reacquisition of root growthafter a salt stress (5). These selected genes
were classified into different functionalcategories depending on homologies toother species. Several stress-related genesinclude specific legume ESTs suggesting,therefore, the presence of novel pathwayslinked to abiotic stress responses in this family.Over-expression of one of the transcriptionfactor genes (MtZpt2-1) linked to recoveryprocesses in transgenic Medicago roots allowedthem to grow under salt stress conditionsand affected the expression of three putativetargets in a predicted manner: a cold-regulated A homolog, a flower-promotingfactor homolog and an auxin-inducedproline-rich protein gene. This establishesa role for this gene in the activation of aspecific genetic programme in theadaptation of legume roots to salt stress.
Combining genomics andgenotypic diversity
A large diversity of genotypes adapted toabiotic stresses was found in the glycophyteM. truncatula. This resource of diversity isused by breeders to manage abiotic stressesin their cultivation areas. Complex geneticsystems subject to interactions betweengenotype and environment controlled theplant traits for adaptation to environmentalstresses. Therefore, by using comparativegenomic approaches between thesegenotypes, the right complements of genesand alleles in a breeding programme could
be proposed. The understanding of thegenetic mechanisms involved in theseadaptive processes is particularly importantin cases such as drought or salt tolerancewhere few genes may drastically affect thebehaviour of related genotypes in a specificenvironment. Advances in genomic toolscombined with the exploitation of geneticdiversity from model legumes are allowingthe dissection of the genetic control ofcomplex traits. Determination ofphysiological tolerance mechanisms indiverse genotypes to various abiotic stressesin legumes may direct agronomicimprovement of legumes increasingproduction in a sustainable way. �
(1) Bray, E. A. et al. (2000). In: The biochemistryand molecular biology of plants, 1158–1249(Eds B. Buchanan et al.) The American Societyof Plant Physiologists, Rockville, USA.
(2) Buitink, J. et al. (2006). Plant J. 47, 735–750.
(3) Dita, M. A. et al. (2006). Euphytica 147, 1–24.
(4) Flowers, T. J. and Yeo, A. R. (1995). Aust. J.Plant Physiol. 22, 875–884.
(5) Merchan, F. et al. (2007). Plant J. 51, 1–17.
(6) O’Brian, M. R. and Vance, C. P. (2007).Plant Physiol. 144, 537.
(7) Pedrosa, A. N. et al. (2002). Genetics 161,1661–1672.
(8) Thoquet, P. et al. (2002). BMC Plant Biol. 2, 1.
(9) Udvardi, M. K. et al. (2007). Plant Physiol.144, 538–549.
Drought Salinity
Heat Freezing Chilling
Mineral toxicities
Waterlogging
ABIOTIC STRESSES
ADAPTATION
ARCHITECTUREGROWTH
SYMBIOTIC NODULES
REGULATORY GENESTRANSCRIPTION FACTORS
GENOMICTRANSCRIPTOMIC
APPROACHES
Mt16KMICROARRAYS
qRT-PCR
Figure 1. General view of the different steps linking genomics to the identification of key regulatory genesinvolved in abiotic stress responses in legumes. Exploiting genetic diversity in model legumes combined withtransgenic approaches will reveal regulatory mechanisms involved in legume adaptation to abiotic stress.

GRAIN LEGUMES No. 49 – October 200716
SPEC IAL REPORT
ABIOTIC STRESS
Abiotic stresses, salinity, cold anddrought have the largest impact,causing yield losses of up to 50%
in legumes. Evidence from various speciessuggests that abiotic stress tolerance is aquantitative, developmentally regulated,stage-specific phenomenon, and thattolerance at one stage of development maynot be correlated with tolerance at othersuch stages. This suggests that specific stagesshould be evaluated separately to developcultivars with stress-tolerance characteristics(1). Altogether, this implies that abiotic stresseshave an impact on a large number ofinteracting genes. Plant breeding thus facestwo questions: a) how to identify all the genes,and their allelic variations, that each confera better tolerance to stress and b) how tocombine all these genes in elite cultivars (3)?
A number of genes involved in abioticstress tolerance have been identified usingmolecular approaches (4, 6). However, itis very difficult to reveal their contributionto the plant phenotype in a givenenvironment, especially since these genesare often involved in more than one stress.Only the combination of molecular geneticapproaches such as Transmission genetics,Association genetics or Ecotilling allows abridge between molecular and genetic data.
The need for a model plantThe large number of gene x environment
x cultivated species combinations requiredfor legume breeding justifies the use oftaxonomically related model plants. Specialinterest in M. truncatula as a model plant forlegumes relies on its capacity to supportmolecular genetics programmes and itstaxonomic proximity with crops such as
alfalfa, pea, chickpea, lentil and fababean.Many molecular and genetic tools (cDNAand BAC banks, software, genetic maps)have been developed in conjuction withM. truncatula (5) and the sequence of itsexpressed genome is expected to be finishedin 2008.
One factor of major interest is thatM. truncatula grows spontaneously all aroundthe Mediterranean basin in very diverse,and sometimes hostile environments.Therefore, allelic variations responsible foradaptation to most abiotic stress could beexpected within M. truncatula naturalvariations. In addition to the existingcollections of M. truncatula (8, 2), a dedicatedcollection has been created based onTunisian natural populations growing indiverse eco-environmental conditions (7).
Molecular genetics ofM. truncatula salt tolerance
We first selected the genotypes Jemalongand F83005.5 which show contrastingphenotypes when submitted to salt stress(Figure 1). Using the M. truncatula geneticmap and the population of RecombinantInbred Lines (RILs) already developed, weidentified a number of Quantitative TraitLoci QTLs) involved in salt tolerance (Figure1). These QTLs underly genes whose allelicvariations are responsible for the observedphenotypic differences between these twogenotypes. But how are QTLs and the actualgenes connected? Transcriptomic approachesprovide important information about putativecandidate genes but only the cloning of the
alleles from one parent and its transfer tothe other will give the final proof. This map-based cloning of these QTLs is in progress.
Since the genes/alleles responsible forsalt-tolerant behaviour are not always thesame as in the plants analysed, this geneticapproach should be repeated using othercontrasting genotypes in order to identifymore QTLs/genes and evaluate their rolein the plant response to salt stress.
From models to cropsDue to the conservation of genome
organisation between related species, thelocation of a gene in a crop species couldbe deduced from model species studies andcould thus be used for marker- or genome-assisted selection of crops. However, itstill has to be established whether thefunction of a gene, and its allelic variations,are similar in different, even closely related,species. This research should be developedin the future. �
(1)Bayuelo-Jiménez, J. S. et al. (2002). CropScience 42, 2184–2192.(2) Ellwood, S. R. et al. (2006). Theor. Applied.Genet. 112, 977–983.(3) Flowers, T. J. et al. (1999). Journal ofExperimental Botany 51, 99–106.(4) Mahajan, S. and Tuteja, N. (2005). Arch.Biochem. Biophys. 444(2),139–158.(5) Medicago truncatula handbook:http://www.noble.org/MedicagoHandbook/(6) Merchan, F. et al. (2007). The Plant Journal51, 1–17.(7) Lazrek, F., Huguet, T. and Aouani, M. E. Inpreparation.8) Ronfort, J. et al. (2006). BMC Plant Biology6, 28.
*Laboratoire Symbioses et Pathologies desPlantes (SP2), INP-ENSAT, Castanet Tolosan,France. ([email protected])
Integrating molecular and genetic data in legumesIntégration des approaches moléculaires et génétiques chez leslégumineusesby Thierry HUGUET*
Figure 1. Contrasting tolerance to NaCl (45mM) of M. truncatula Jemalong (left) and F83005.5 (right);M.truncatula genetic map based on LR5 population of RILs. Vertical lines indicate QTL locations involved in thereduction of stem growth (1), reduction in the number of leaves (2), reduction of root growth (3).
(Pho
to T
. Hug
uet,
INP-
ENSA
T,C
asta
net T
olos
an, F
ranc
e)

GRAIN LEGUMES No. 49 – October 200717
SPEC IAL REPORTABIOTIC STRESS
Benefits of inoculation under abioticstress in the Mediterranean regionL’intérêt de l’inoculation sous stress abiotique en région méditerranéenneby Carmen VARGAS* and Mohamed Elarbi AOUANI**
*University of Seville, Spain. ([email protected])
**Centre de Biotechnologie, Technopole deBorj Cedria, Tunisia and NEPAD/North Africa Biosciences Network (NABNet), National Research Centre, Cairo, Egypt.([email protected])
If appropriately managed, and especiallyunder abiotic stress (drought, salinity),microbial biofertilisers can improve
food legume yield and soil fertility andreduce pollution by inorganic fertilisers. Ingeneral, three types of inoculants can helplegumes overcome stressful environments:
• Efficient and stress-tolerant symbioticrhizobial strains, used when native populationsare scarce and/or inefficient. The Aquarhizproject1 targeted four south Mediterraneancountries (Algeria, Egypt, Morocco,Tunisia) to study the inoculation of fababean, chickpea and common bean underdrought and/or saline conditions. Severaltrials showed a positive effect of inoculation,with variation according to countries andcrops but also according to strains. At someexperimental sites the inoculation had noeffect or the increase in yield was notsignificant. In the case of faba beans inMorocco, soils are naturally hosting efficientlocal strains. However, in several other cases,sowing seeds with addition of these stress-tolerant rhizobium strains enhanced nitrogenfixation activity and improved yield of grainlegumes. The symbiotic systems which areinhibited by abiotic stress are well known:oxygen permeability, shortage of watersupply to nodules from the phloem,overproduction of reactive-oxygen species,feedback inhibition involving shoot Nstatus. However the mechanisms underlyingthe stress tolerance of symbiosis remainunclear. Candidate systems from the plant
(aquoporins, carbonic anhydrase) or thebacterial (osmoprotectants, bacteroidalrespiration) side should be investigated inorder to select successful combinations oflegume and microsymbionts better adaptedto stress.
• Arbuscular mycorrhizal fungi (AMF)colonise root systems and form symbiosescalled mycorrhiza with almost all legumes.By exploring a greater volume of soilthrough the fungal hyphae, mycorrhizaenhance nutrient (particularly phosphorus)and water uptake from the soil, therebyenhancing plant growth under drought stress.
• Plant Growth Promoting Rhizobacteria(PGPR), include a diverse group of free-living soil bacteria that can stimulate plantgrowth by a number of direct or indirectmechanisms. Interestingly, many strainsisolated in the Aquarhiz project are free-living non-rhizobial species with promisingapplications as inoculants.
Inoculation is limited anddecreasing
Although inoculation is a well developedagricultural practice in Australia, Canada,USA and Latin America, it is not commonin Europe and Africa. The status of legumeinoculation in the Mediterranean region andthe related research strategies were discussedat a FABAMED2 workshop organised atRabat in February 2005, following the firstGeneral Meeting of the Aquarhiz project.It was shown that legume inoculation withrhizobia was limited and decreasing in theregion, whereas the use of N fertilisers wasincreasing, especially at Egypt.
The workshop participants concludedthat the symbiosis should be taken intoaccount in research activities and in the
technical messages disseminated to farmersto improve legume production in theMediterranean region, but some key pointsshould be considered:
- The inoculation is not the uniquesolution as shown in past experience, anda systematic diagnosis is required torationalise where and when inoculantapplication can have an impact.
- Active rhisophere and an efficientrelationship between enhanced cultivarsand the best adapted rhizobia, either presentin the soil or added as inoculant, is a keycomponent to boost N fixation and rootgrowth leading to better tolerance of abioticstress.
- Inoculation should be associated withappropriate crop and soil management.Plant genotype and nutrient conditions arekey factors influencing inoculation success.
- Progress in inoculation technology isrequired, i.e. second generation inoculants(rhizobial plus mycorrhiza or PGPR).
- Inoculant production is an easy and cheaptechnology that could be scaled-up withinstitutional support.
- Farmers’ adoption of the technology willrequire participatory field research andappropriate technological packages. Theseactivities have been initiated within theframework of the Aquarhiz project. �
1FP6 project www.grainlegumes.com/aquarhizand Grain Legumes 39, 23.2FABAMED is a network of scientists interestedin N fixation in the Mediterranean Region. Setup in 1994, it meets on a regular basis forinformation exchange, workshops and thepreparation of joint projects.
Faba beans with inoculation (left of photo) andwithout inoculation (right of photo), grown in sandysoils in Beheira in Egypt.
(Pho
to Y
. G. Y
anni
, Sak
ha A
gric
ultu
ral R
esea
rch
Stat
ion,
Egy
pt)

GRAIN LEGUMES No. 49 – October 200718
SPEC IAL REPORT
ABIOTIC STRESS
*ICARDA, Aleppo, Syria. ([email protected])
Abiotic stress breeding at ICARDAAmélioration variétale et stress abiotique à l’ICARDAby Michael BAUM*
The mission of the InternationalCenter for Agricultural Researchin the Dry Areas (ICARDA) is to
meet the challenges of dry-areaenvironments, which are harsh, stressfuland variable. ICARDA has both global andregional mandates. It has a globalresponsibility for the improvement of threeimportant food crops – barley, lentil, andfaba bean. ICARDA's regional responsibilitywithin Countries of West Asia and NorthAfrica (CWANA) focuses on theimprovement of wheat, chickpea, forageand pasture crops.
ICARDA holds the largest gene bankin the Mediterranean region, with about133,000 accessions, which representsapproximately 20% of the germplasm inCGIAR centres. Particularly important arethe landraces and wild relatives that haveevolved under harsh conditions overmillennia. About 70% of ICARDA’scollections are now geo-referenced. Thiscollection-site information combined withclimatic layers in Geographical InformationSystem (GIS) allows targeted collection anda rapid exploitation of accessions withtolerance to drought and heat to meet theanticipated effects of climate change.ICARDA has been sharing these resourcesfreely with partners all over the world.On average, the Center distributes 35,000samples per year. Overall, the shift fromcollection and ex situ conservation of plantgermplasm to its characterisation, evaluationand documentation will help to utilise thebiodiversity held at ICARDA.
Using modern technolgiesICARDA collaborates with advanced
research institutes (ARIs) and the NationalAgricultural Research Systems (NARS) in
using modern technologies for its cropimprovement programmes. Emphasis isgiven to the identification and exploitationof genetic resources for improved stressresistance and water-use efficiency. DNAmolecular marker techniques allowconstruction of linkage maps for crops.Together with statistical techniques theselinkage maps can be used to locate andestimate phenotypic effects of quantitativetrait loci (QTL) and the genes responsiblefor the expression of agronomic traits. Fora homozygous population derived from across with parents contrasting in responseto, for example, water, QTL analysis revealsthe approximate map location of lociassociated with performance under drylandconditions. This is then amenable tomarker-assisted selection using DNAmarkers flanking the identified QTLs.
Lentil and chickpeaimprovement
Radiation–frost injury is an importantabiotic constraint to lentil production inWest Asia. If cold tolerance of lentil couldbe improved, its yield in low rainfall areascould be increased by early planting.Recombinant inbred lines of a lentil crosswere tested in northern Syria for plant frostinjury levels and genotyped with 254 DNAmarkers. DNA markers were identifiedlinked to the locus for radiation–frosttolerance trait and Fusarium wilt resistance.These are proving useful in lentilimprovement. Instead of selecting forradiation–frost tolerance and Fusarium wilt,breeders can select for the resistance markeralleles.
Chickpea is an important dryland cropsown in spring in WANA. Its yield andwater-use efficiency can be nearly doubledby advancing its date of sowing to winter.Ascochyta blight, a fungal disease attackingchickpea, has developed into a major threat
for winter-grown chickpea. DNA markershave been used to characterise the availablepathotypes of the fungus. This allows thedevelopment of geographical distributionmaps of the pathogen and the deploymentof effective host-plant resistance genes. Fortagging host-plant resistance genes inchickpea, microsatellite based markers havebeen developed in collaboration with theUniversity of Frankfurt. Host-plantresistance for Ascochyta blight is beingmapped in several populations and geneticbackgrounds.
BiosafetyICARDA is also exploring the possibility
of using genetic transformation to achieveimproved tolerance to drought and otherabiotic stress resistance. ICARDA alsoactively promotes the development andestablishment of national and regionalbiosafety regulations. So far, Egypt andSyria are the countries in the region thathave established biosafety regulations. Inseveral other countries the preparations forthe development of biosafety regulationsare underway. ICARDA’s policy is guidedby international biosafety practices and theregulations of the Syrian Arab Republicfor the development and deployment ofGMOs. �
Field of chickpeas at Ain Temouchet, Algeria inApril 2007.
(Pho
to H
ouci
ne Ir
ekti,
INAT
, Alg
eria
)

GRAIN LEGUMES No. 49 – October 200719
SPEC IAL REPORTABIOTIC STRESS
Where we go from here – concluding remarks of theabiotic stress in legumes workshop Quelles perspectives pour la suite – conclusion du séminaire sur lestress abiotique chez les légumineusesby Martin CRESPI*, Anne SCHNEIDER**, Thierry HUGUET*** and Mohamed Elarbi AOUANI****
Maintaining or improving cropproductivity under conditions ofabiotic constraint in the field is
one major concern in many areas in theworld where legumes are grown. Abioticstresses refer to different environmentalfactors: water deficit, high temperatures,saline stress, mineral deficiency, frost,chilling, and others, which can occur atdifferent levels and development stages. Asshown in the Tunis workshop1, it is veryuseful to integrate different disciplinaryexpertise to apprehend the complexity oftolerance and susceptibility traits.
From whole plant to genomicapproaches, a strategy toexploit genetic diversity
The whole plant approach allows keyvariables of the plant/genotype interactionthat maintains yield under stress to be assessed.Environmental signals and plant physiologicalmechanisms activated at different growthstages can be useful to determine theirhierarchy and relevance in the physiologicalprocesses involved. Adapted agronomicmanagement techniques are needed to copewith abiotic stresses in the field. However,modelling the eco-physiological interactionsbetween plants and the environment,considering the particular aspects of legumephysiology, will enhance the basicunderstanding of mechanisms for furtheragronomic improvement.
In legumes, the symbiotic bacterial partner,the rhizobium, and other components ofthe rhizosphere have key roles in the
agronomic performance of these crops, andshould be taken into account in the analysisof plant response. Improving symbiosisunder stress conditions will contribute tothe development and use of legumes aspioneering crops since dry or saline soilsare also poor in combined nitrogen.
Parallel development of genomicapproaches will reveal molecularmechanisms involved in the regulation ofthe plant physiological responses. Exploitingthe data obtained for different plant species(particularly different legumes) willhighlight key molecules and genes involvedin stress responses. These approaches shouldevolve from the initial description levelunder specific conditions to more elaboratescreenings defining traits and genes involvedin crop adaptation strategies. Targeted cropstrategies should focus on the enhancementof tolerance to abiotic stress during grainfilling or on rapid soil coverage in specificsystems by controlling germinationpotential and initial root growth.
In addition, exploiting genetic diversityis crucial to obtain both contrasted typesand sources of tolerances. Genetic resourcesare in fact mutants already selected byspecific environmental conditions, possiblymaking them more suitable for fieldconditions than laboratory mutants.
Interdisciplinary approachesshould be developed
Integrating molecular and genetic datainto eco-physiological models is requiredin order to define the regulatorymechanisms involved in the control of plantgrowth and development under abioticstress constraints and to create novel elitecultivars. This will bring new perspectives
to the development of novel cropmanagement techniques and to theidentification of crucial breeding targets.
The general approaches favoured in orderto make progress in this area were discussedamong Tunis workshop participants. Theneed for interdisciplinary approaches toenhance knowledge in abiotic stress responsesand tolerance was agreed upon. Plantphysiology expertise should be reinforcedparticularly to establish common referencesand protocols in case studies: how to applystress, what sort of stress and its level ofintensity, what type of plant response istargeted (e.g. survival or adaptation). Geneticmodels will provide information on plantresponses at different levels that can be appliedin crop species although the specificity ofthese responses according to species or evenlines should also be considered. Two differentresearch strategies were debated: one focussingon the search for few key genes whosemodulation would provide a breakthrougheffect on crops or a more integrative approachaiming to dissect the complex mechanismsinvolved in stress responses. Both willcontribute to the development of improvedvarieties adapted to cropping systems.
In spite of strong debate, specific pointswere unanimously agreed: the kindhospitality of the Tunisian hosts, the interestin participating in interdisciplinaryworkshops and the need to structurecollaborations between experts fromdifferent fields to increase the relevance ofthe strategies in crop management.
1‘Abiotic stresses ain legumes’ (Tunis, 22–24March 2007), a scientific workshop organised atthe initiative of the Grain Legumes IntegratedProject (GLIP) which is supported by theEuropean Commission from 2004 to 2008.
*CNRS-ISV, Gif sur Yvette, France**AEP, Paris, France***INRA-ENSAT, Toulouse, France****CBBC-LILM, Hammam Lif, Tunisia
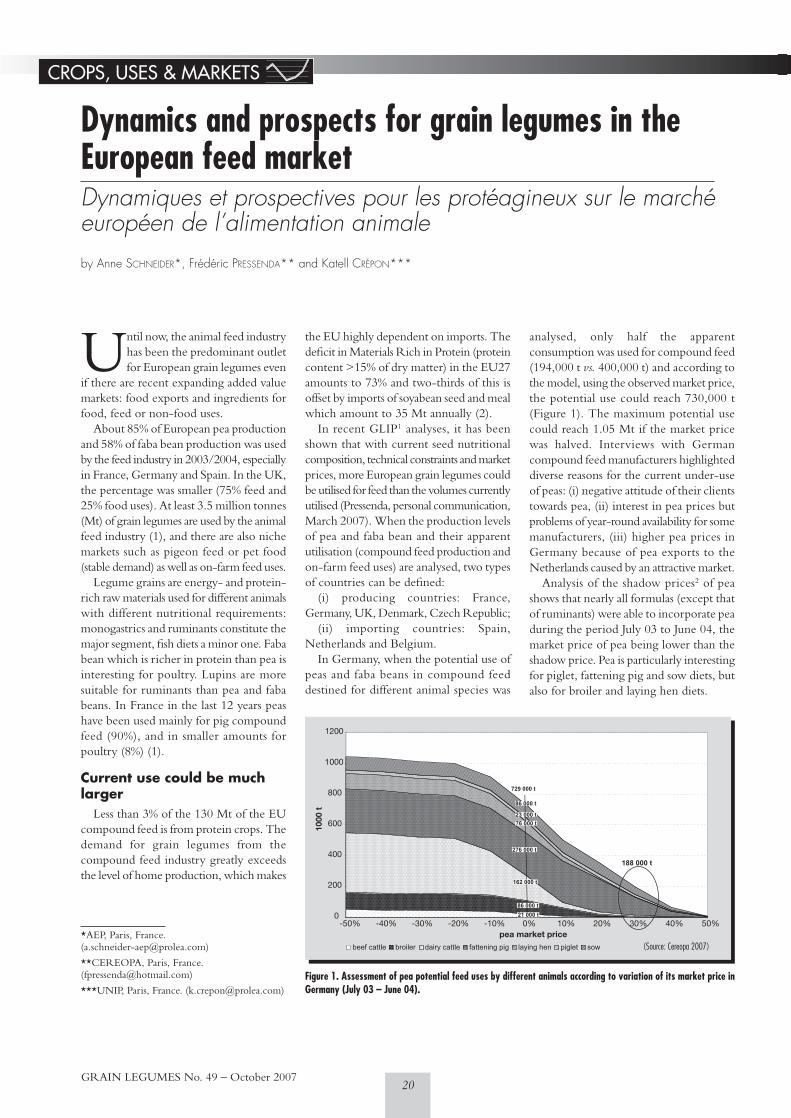
GRAIN LEGUMES No. 49 – October 200720
Dynamics and prospects for grain legumes in theEuropean feed marketDynamiques et prospectives pour les protéagineux sur le marchéeuropéen de l’alimentation animaleby Anne SCHNEIDER*, Frédéric PRESSENDA** and Katell CRÉPON***
analysed, only half the apparentconsumption was used for compound feed(194,000 t vs. 400,000 t) and according tothe model, using the observed market price,the potential use could reach 730,000 t(Figure 1). The maximum potential usecould reach 1.05 Mt if the market pricewas halved. Interviews with Germancompound feed manufacturers highlighteddiverse reasons for the current under-useof peas: (i) negative attitude of their clientstowards pea, (ii) interest in pea prices butproblems of year-round availability for somemanufacturers, (iii) higher pea prices inGermany because of pea exports to theNetherlands caused by an attractive market.
Analysis of the shadow prices2 of peashows that nearly all formulas (except thatof ruminants) were able to incorporate peaduring the period July 03 to June 04, themarket price of pea being lower than theshadow price. Pea is particularly interestingfor piglet, fattening pig and sow diets, butalso for broiler and laying hen diets.
the EU highly dependent on imports. Thedeficit in Materials Rich in Protein (proteincontent >15% of dry matter) in the EU27amounts to 73% and two-thirds of this isoffset by imports of soyabean seed and mealwhich amount to 35 Mt annually (2).
In recent GLIP1 analyses, it has beenshown that with current seed nutritionalcomposition, technical constraints and marketprices, more European grain legumes couldbe utilised for feed than the volumes currentlyutilised (Pressenda, personal communication,March 2007). When the production levelsof pea and faba bean and their apparentutilisation (compound feed production andon-farm feed uses) are analysed, two typesof countries can be defined:
(i) producing countries: France,Germany, UK, Denmark, Czech Republic;
(ii) importing countries: Spain,Netherlands and Belgium.
In Germany, when the potential use ofpeas and faba beans in compound feeddestined for different animal species was
*AEP, Paris, France. ([email protected])
**CEREOPA, Paris, France.([email protected])
***UNIP, Paris, France. ([email protected])
Until now, the animal feed industryhas been the predominant outletfor European grain legumes even
if there are recent expanding added valuemarkets: food exports and ingredients forfood, feed or non-food uses.
About 85% of European pea productionand 58% of faba bean production was usedby the feed industry in 2003/2004, especiallyin France, Germany and Spain. In the UK,the percentage was smaller (75% feed and25% food uses). At least 3.5 million tonnes(Mt) of grain legumes are used by the animalfeed industry (1), and there are also nichemarkets such as pigeon feed or pet food(stable demand) as well as on-farm feed uses.
Legume grains are energy- and protein-rich raw materials used for different animalswith different nutritional requirements:monogastrics and ruminants constitute themajor segment, fish diets a minor one. Fababean which is richer in protein than pea isinteresting for poultry. Lupins are moresuitable for ruminants than pea and fababeans. In France in the last 12 years peashave been used mainly for pig compoundfeed (90%), and in smaller amounts forpoultry (8%) (1).
Current use could be muchlarger
Less than 3% of the 130 Mt of the EUcompound feed is from protein crops. Thedemand for grain legumes from thecompound feed industry greatly exceedsthe level of home production, which makes
0
200
400
600
800
1000
1200
-50% -40% -30% -20% -10% 0% 10% 20% 30% 40% 50%pea market price
1000
t
beef cattle broiler dairy cattle fattening pig laying hen piglet sow
729 000 t
21 000 t
86 000 t
276 000 t
76 000 t
23 000 t
86 000 t
188 000 t
162 000 t
Figure 1. Assessment of pea potential feed uses by different animals according to variation of its market price inGermany (July 03 – June 04).
CROPS, USES & MARKETS
(Source: Cereopa 2007)

GRAIN LEGUMES No. 49 – October 200721
Similar analyses in the Netherlandsshowed that the potential use of pea reached1.6 Mt when its price was 25% lower thanthe observed market price. According tostatistical data, the feed uses of pea amountto 195,000 t, showing that all importedpeas are used by compound feedmanufacturers. Most available peas are usedin pig diets, and their use is highly sensitive:if the market price decreases by 3%, theuses of pea would be 200,000 t (or 450,000 tfor the market price observed during theperiod July 03–June 04).
The discrepancy between the actual andpotential feed uses of grain legumes inseveral EU countries is due mainly to thesmall quantities available on the market inrecent years. Sufficient volumes availableon the market and regular availability arerequired for the compound feed industryto establish a purchasing policy.
When volumes are low, grain legumesare used mainly in pig diets; with increasedvolumes, dairy cows could become apotential important user (Cereopaexpertise).
Competition with other rawmaterials?
The clear correlation between peamarket price and its uses shows that pea isa raw material that can be easily substitutedin the feed industry. The price of feed peais linked directly to the prices of the twomain raw materials used in animal feed:wheat (or barley) and soyabeans.
Imported soyabeans (seeds and especiallymeals) are the major competitor for EUgrain legumes. The nutritional compositionof soya meal fits especially well with thenutritional requirement of intensive poultryand dairy cows. Another key advantage isits relatively low price due to large volumesof soyabean production and its two outlets(oil and meal).
However seeds of other grain legumesgrown in the EU are richer in energy(starch) and have potential for less intensiveanimal production. In addition they ensureregional autonomy and security byproducing raw materials close to their areasof use, and providing an environmentalbenefit to feed chains. When the uses ofBrazilian soya meal and German peas in German pig production are comparedusing Life Cycle Analysis, the results forthe criteria ‘energy consumption’ and‘global warming potential’ show a clear
environmental benefit for local pea(Nemecek, personal communication, GLIP,2006). The benefits are even clearer whenthe grain legume is produced and usedfor animal feed on the same farm.
In addition to soyabeans, several otherEU raw materials are of interest for animalfeed (meals from rapeseeds and sunflowers,corn gluten and coprah and cotton,dehydrated forage and others) but grainlegumes can complement any of these. Peais characterised by a high starch content,with a good energy value, and by protein,especially rich in lysine, which complementcereal proteins that have a low lysinecontent. Rape seed meal is richer in proteinthan pea and its proteins are rich in sulphuramino acids which can complement thepea proteins. Therefore the priority usesof these two raw materials rich in proteinare different and are often not for the sameformula: pea is suitable for pig diets, oilseedmeal is especially suitable for ruminantsand poultry.
Future competition?The development of biofuel and
bioethanol production could lead to asignificant increase in the amount ofrapeseed meal and cereal distillers’ residuesin the EU, and these materials could beavailable for the animal feed industry.Rapeseed meal and cereal distillers’ residuesare used mainly in ruminant feed, whereaspeas are used mainly in pig feed, but theycan also compete with peas in pig formulas.Simulations using a French formula showthat an increased use of rapeseed meal inthe feed industry only slightly decreases theuse of peas, since rapeseed meal is usedprimarily in ruminant formula (Figure 2).
Peas are still useful in feed rations becausethey are a good complement for rapeseedmeal in pig formulas.
Therefore the likely future increase inprotein-rich by-products from biofuelproduction may not be a major threat. InFrance, for example, an increase in biofuelsproduction could add about 2 Mt rapeseedmeal and 700,000 t wheat distillers’ residueson the market, but an analysis by Cereopain 2007 (Figure 2) shows that two-thirdsof oilseed meal will be used in ruminantformulas, 25% in pig formulas and 10% inpoultry formulas. Pea is used mainly in pigformulas so will not compete directly withother protein-rich materials. An adjustmentof only 5% decrease in pea price wouldbe enough to maintain the current volumeof peas used in the pig feed industry (2005-06 basis). In addition, the energy rich rawmaterials may become more expensive withthe development of bio-fuel so protein-rich materials could be interesting.
In conclusion, prospects for the feedmarket are difficult to forecast because ofthe changing economic and politicalcontext which influence market dynamics,but for grain legumes the offer appears tobe the major factor limiting utilisation sincethis falls far short of the level of the potentialutilisation. �
(1) Pressenda, F. (2006). The place of peas in thefeed industry and ways to improve pea uses.GLIP Deliverable D 2.2.1b.
(2) More at: http://www.grainlegumes.com/aep/production/economic_integrated_chain/current_state_of_the_art1Grain Legume Integrated Project (2004-2008)Food-CT-2004-506223 –www.eugrainlegumes.org2the price that a raw material must reach to beincluded in the compound feed formula
Dairycattle T
Dairycattle Pr
Beefcattle T
Beefcattle Pr
Pig T Pig Pr Poultry T Poultry Pr0
500
1000
1500
2000
2500
3000
1000
ton
nes
Wheat distillers Sunflower meal Rapeseed meal Soya meal Pea
T: today Pr: Prospects
Figure 2. Case study in France: current use of protein-rich raw materials in compound feed compared withprojections of their likely uses with the availability of by-products from bio-fuel.
CROPS, USES & MARKETS
(Source: Cereopa 2007)

GRAIN LEGUMES No. 49 – October 200722
Decision support system for efficient disease controlSystème d’aide à la décision pour une lutte efficace contre lesmaladiesby Thomas VOLK and Julia-Sophie VON RICHTHOFEN*
Farmers decide to grow grain legumesprimarily because of their good breakcrop effects – faba beans, field peas
and lupins improve soil fertility and lead tohigh additional yields of the following crop,in most cases winter wheat.
However, the yield of the grain legumecrop itself should also be high and stable.Diseases can – depending on the year andthe region – reduce grain yields significantly(Table 1). In these cases the use of fungicidesseems to be advantageous economically.
In Germany in many cropping seasonsand in many fields the level of infestationis low because
– the acreage cultivated with grainlegumes is only small in most regions,
– very few winter types are grown,– field peas are grown mainly in the
eastern parts of Germany, characterised bya continental climate and low precipitation.
Experience shows that if the infestationlevel is high, fungicide treatments can leadto significant additional grain yield (forexample >10 t/ha for faba bean rust) and ahigher gross margin. If the weather conditionsare favourable for pathogen developmentfarmers and advisers need to decide on theoptimal products, application rates and datesto control the disease effectively.
In Germany the number of fungicidesregistered for grain legumes is very limited.
Currently only two foliar fungicides areallowed for faba beans, namely Folicur(tebuconazole) and Amistar (azoxystrobin).For field peas farmers can use Amistar andVerisan (iprodione), for lupins Amistar,Folicur and Switch (fludioxonil, cyprodinil).
So in which cases does a fungicidetreatment pay off in grain legumes? Thisis shown for faba beans:
Weather for infection On the chosen application date a farmer
should have diagnosed disease infestation inhis field, and the weather conditions shouldfavour an increase in the existing infestation.Faba bean diseases have different climaticrequirements. Whereas Ascochyta blightspreads during long humid periods even ifthe temperatures are relatively low, faba beanrust requires only a short spell of rain oreven dew, but combined with much highertemperatures. Consequently the first rustsymptoms are usually found some weekslater than Ascochyta spots. Fungicidesshould be applied between the beginningand end of flowering. Because of the limitedpreventive efficacy of the registeredfungicides, earlier treatments bear a higherrisk of late infections. The mixture of0.5 l/ha Folicur and 0.5 l/ha Amistar is analternative solution to the single use of1.0 l/ha Folicur or 1.0 l/ha Amistar becauseit combines the curative properties of Folicurwith the preventive properties of Amistar.
Furthermore the susceptibility of thevariety plays an important role in thetreatment decision. In varieties susceptible
to infection the infestation appears earlierthan in varieties with low susceptibility. Forexample, a higher yield increase regularlyresults from fungicide treatment againstfaba bean rust of the highly susceptiblevariety Scirocco than from treatment ofGloria, which is not so susceptible to rust.
Table 2 gives the results of a field trialimplemented by FH SWF1in 2004 withinthe GL-Pro2 trial network. Compared withthe untreated control the yield increase forScirocco was 16% when treated with0.75 l/ha Folicur at full flowering and only9% for Gloria.
The increase in yield is due primarily toa higher thousand seed weight (TSW) andat high yield levels (> 6 t/ha) the yieldincrease with a fungicide treatment is higherthan at low yield levels. Therefore the useof fungicides is more efficient if high yieldsare expected.
proPlant expert.classicFor many years the computer-based
decision support system ‘proPlantexpert.classic’ has proved of value for cereals,rapeseed, potatoes and sugar beet. Since2005 the system has been used to assistgrowers of field peas and faba beans to makefield specific fungicide treatment decisions.
Each consultation is based on both fieldspecific data and climatic data from officialmeteorological services. Together withinformation on the infestation level, thesystem processes the data to develop arecommendation for the application of aspecific fungicide, if necessary.
*proPlant Ltd, Muenster, Germany ([email protected])
Table 1. Important grain legume diseases in Germany.
Crop English name Latin name
Field pea Ascochyta blight Ascochyta pisiDowny mildew Peronospora pisiBotrytis grey mould Botrytis cinereaPea rust Uromyces pisi
Faba bean Chocolate spot Botrytis fabaeFaba bean rust Uromyces vicia-fabaeDowny mildew Peronospora viciaeAscochyta blight Ascochyta fabae
Lupin Anthracnose of lupin Colletotrichum lupini
Table 2. Fungicide treatment of faba beans: difference between varieties.
Variety Susceptibility Yield (untreated) Yield (treated)b Relative yield to rusta t/ha (86% dry matter) t/ha (86% DM) (treated)
Gloria 4 4.33 4.71 109
Scirocco 6 6.88 8.00 116aGerman National List of plant varieties 2004: 1 (not or very low) – 9 (very high)bFungicide treatment: 0.75 l/ha Folicur on 21 June 2004 (full flowering)
Source: GL-Pro trial network. Trial results 2004 at the experimental station Merklingsen of FH SWF near Dortmund in Northwest Germany (750 mm average annual rainfall, 9°C average annual temperature, fertile soils with 80–85% silt).
CROPS, USES & MARKETS

GRAIN LEGUMES No. 49 – October 200723
CROPS, USES & MARKETS
Infection probability At the beginning of a consultation the
user gets a quick overview of the situationin his region. The so called ‘warning service’shows the weather conditions during thepast 3–4 weeks and a weather forecast forthe following three days. The climaticoverview gives at least daily maximum andminimum temperatures, hours of sunshineand rainfall levels. As rainfall may varybetween the farm and the location of themeteo station, the user can changeprecipitation levels according to his ownrecordings. This is important because rainfallhas a significant impact on the spreadingof fungal diseases.
Taking into account the previous days andthe weather forecast, the system determineswhether recent weather conditions wereconducive, or will become conducive, topathogen infection in the next three days.Black or light grey dots3 mark good weatherconditions for an infection, with light greydots indicating a situation that is conduciveto pathogen development and black dotsindicating a situation that is optimal forpathogen development. The dark grey
background marks days on which curativepathogen management is still possible.
For peas, the system specifies conditionsfor grey mould, Ascochyta blight and pearust infections. For faba beans the systemdetermines conditions for faba bean rust,chocolate spot, Ascochyta blight and downymildew infections. In addition, days withheat stress are indicated for both crops.
For example Figure 1 reflects thesituation in the GL-Pro trial mentionedabove in June 20044. As the infection dots
indicate, the weather conditions were goodfor disease infection, especially for rust andchocolate spot.
Specific recommendationTo get a field-specific recommendation
the user also needs to enter appropriatedata into the system, for example, variety,growth stage and soil drying. Furthermorea field check-up is required to determinethe current infestation level. Figure 2 showsthe parameters which influence theconsultation results. For example, the systemsuggests a certain treatment for faba beanrust during the flowering stage if recentweather conditions have been optimumfor infection and if the variety’s susceptibilityto infection is average or high.
proPlant expert.classic creates a list offungicides that are able to manage thespecific infestation in the field. Among otherfactors, the date of infection plays a majorrole in determining the application rate.If the infection is recent, a low applicationrate will have a good (curative) effect onfungal diseases. If the infection has existeda long time, higher rates will be necessaryto provide effective control. The list ofrecommended fungicides also provides theoptimal application date or period as wellas the approximate costs of an application.
The efficiency of a treatment can varydepending on the growth stage, applicationrate and properties of the chemical used.To ensure an unbiased recommendation fora specific fungicide, the products providedin the database of the system are assessedregarding their curative, eradicative andpreventive properties by results that wereobtained in official field trials. The preventiveeffect of a product is indicated by thenumber of days during which the productprovides protection at average temperatures.
The effect of applying 0.7 l/ha Folicurat full flowering on 21 June 2004 in theGL-Pro trial mentioned above is shownin Figure 1. In addition to the infectionprobabilities the curative, eradicative andpreventive properties of the treatment tocontrol the diseases in the specific climaticconditions are given. Because of its curativeefficacy on rust, infections since 15 June2004 could be controlled by the treatment,but not infections which took place earlier.
The dark grey background above showsthat it would have even been possible tocontrol rust infections which establishedon 13 June. But since the conditions on 13
Figure 1. proPlant expert.classic warning service: conditions for faba bean diseases in June 2004 and efficacy ofa treatment with 0.7 l/ha Folicur at full flowering on 21 June 2004.
Legend:
Current day. Optimum conditions for spreading the disease. Good conditions for spreading the disease.
Crop: heat stress.
Potential curative effect of the best fungicide against a disease in a current treatment.Infections controlled by a treatment (dimmed infection spots).
Curative and protective effect of a fungicide treatment.
Number of days with protective fungicide effect (at average temperatures).
Partly eradicative effect of a fungicide.
Figure 2. Parameters that influence the results of aconsultation.
GRAIN LEGUMES No. 49 – October 200724
CROPS, USES & MARKETS
and 14 June 2004 were not conducive torust infections, a higher application rate(for example, 1.0 l/ha Folicur) would havehad no effect on rust, but would haveresulted in higher production costs.
Besides the curative effect, Folicur hasa partly eradicative effect on faba bean rust.The preventive effect of the treatment lasteduntil 7 July 2004 (17 days).
The great variation in fungal diseaseinfestation levels between years becomesobvious when the situation in 2004 is
compared with conditions in 2006 (Figure3). In 2006 a fungicide treatment was notprofitable for faba beans in this north-western region. As proPlant expert.classicshows, fewer days with good and optimalweather conditions for disease infestationoccurred during the flowering period inmid June and rust and chocolate spotappeared very late in the fields.
In 2007, because of the warm springtemperatures faba beans were alreadyflowering by the end of May in north-west
Germany, about three weeks earlier thanusual. Because of the long period with high-pressure and dry weather from the end ofMarch until the beginning of May the diseaseinfestation potential was at a very low level.
Control may not be profitableUsually it is not profitable to control
foliar diseases in peas and faba beans inGermany. But in certain years and in certainregions high infestation pressure can causesevere yield losses. The computer-basedconsultation system proPlant expert.classicprovides support for farmers and consultantswho need to make field-specific treatmentdecisions while taking into accountagronomic data, weather reports andinfestation levels. Against the backroud ofsmall margins in standard feed grain legumeproduction, the decision support systemenables input costs to be saved and diseaseto be controlled efficiently. �
1South Westfalia University of Applied Sciences,Soest, Germany.2European extension network for thedevelopment of grain legume production in theEU (QLK-CT-2002-02418).3Normally the graphs are coloured and the blackand grey dots are red and yellow respectively.4Weather station Werl close to the experimentalstation Merklingen of South WestfaliaUniversity of Applied Sciences.
Figure 3. proPlant expert.classic warning service: conditions for faba bean diseases in June 2006.
A jump in price for French faba beans exported to EgyptFlambée du prix de la féverole française exportée vers l’Egypte by Jean-Paul LACAMPAGNE*
*UNIP, Paris, France. ([email protected])
With annual exports amountingto between 130,000 t and180,000 t since 2002–03. France
is rapidly becoming the foremost exporterto Egypt, the main country importing fababeans for human consumption.
Previously, Egypt imported faba beansprimarily from Australia, but the volumeof supplies was irregular, with low yieldsevery two years since 2002–03. In 2006,Australian production was only 108,000 tcompared with 329,000 t the previous year.
In 2006 and 2007, in spite of attractiveprices, the European supply has alsodecreased, because of a sharp decline in the
production area in France(102,000 ha in 2005 to55,000 ha in 2007) andproblems of quality in 2006(bruchid attack), and alsodisappointing yields in 2007in the UK (the third mostimportant exporting country).
In the context of reducedavailability on the worldmarket and notably in theprincipal exporting countries, togetherwith a steady ongoing demand from Egypt,the price of French faba beans destined forEgypt has been rising for more than a year(Figure 1), reaching a record level of €320/tin French ports in September 2007, in
the context of a very nervous world marketfor wheat and other cereals.
Very attractive prices and high 2007 yieldsin the northern regions of France (theprincipal areas of production) could providea boost for the faba bean crop in France. �
Figure 1. Prices of French faba bean (human consumption quality –trade prices at Rouen, Normandy).
80
110
140
170
200
230
260
290
320
350
Jul-02 Jul-03 Jul-04 Jul-05 Jul-06 Jul-07
€/t
Faba beanfor Egypt
Wheat

25 GRAIN LEGUMES No. 49 – October 2007
Ascochyta blights ofgrain legumes
Bernard Tivoli, Alain Baranger,Fred J. Muehlbauer and B. M.
Cooke (Eds)
November 2007, vi + 142 pages
Springer, Heidelberg, Germany
ISBN: 978-1-4020-6064-9Hardcover
This special issue of the EuropeanJournal of Plant Pathology, 2007 119,
No. 1, reflects some major contributions made at the 1st InternationalAscochyta Workshop on Grain Legumes held on 2–6 July 2006at Le Tronchet, France (see Grain Legumes 47, 6–7.
Ascochyta blights consistently affect large areas of grain legumeproduction (pea, lentil, chickpea and faba bean) in all countrieswhere they are cultivated. These diseases are capable of causinglarge yield losses under conducive environmental conditions. Thisbook considers the state of the art by taking a comparative approachof Ascochyta blight diseases of cool season food and feed legumes.Topics considered are pathogen diversity, legume genetics andbreeding, and integrated disease management.
This special issue, written for scientists, university teachers,students and extension specialists, has a foreword by the editors,followed by thirteen chapters all written by experts in differentareas of Ascochyta research.
Price: (excluding postage): Europe: €69.95 Rest of World:$US99.00 Orders: Springer Distribution Centre GmbH, Haberstr. 7, 69126 Heidelberg, Germany. Tel: +49 (0)6221-345-4301, Fax: +49 (0)6221-345-4229, Email: [email protected] or visit www.springer.com
Pests, diseases and disorders of peas and beans:
a colour handbookA. J. Biddle and N. D. Cattlin
2007, English, 268 x 190 mm, 128 pages
267 colour photographs, ISBN 978 1 84076 018 7
In his Foreword Dr John Kraft,formerly of the USDA Agricultural
Research Service, Washington, USA describes this book as “athorough compilation of information on the diagnosis and controlof pests, diseases and disorders of peas, faba beans and commonbeans in one fully illustrated, easy to read publication.”
The Introduction covers peas and beans in agriculture, peaand bean production and includes a Quick guide to diagnosis.There follow further sections on diseases and pests of seedlingsand young plants, fungal and bacterial diseases, viral diseases, pestsof stem, foliage and produce and seedling and crop disorders. Inall of these sections there high quality photographs of the problemsand the disease or disorder is listed along with the host crop,symptoms, economic importance, disease cycle and control. Thereis a glossary, an index and a list of further reading, and the handbookis described as a must-have publication for all advisory plantspecialists, growers, seedsmen, production specialists, diagnosticclinicians and agribusiness reps who have an interest in peas andbeans.
Price: £35.00Order: Manson Publishing Ltd, 73 Corringham Road, LondonNW11 7DL, UK. Tel: +44 (0) 20 8905 5150, Web: www.mansonpublishing.com
7–9 November, 2007Third International Aphanomyces Workshop on LegumesRennes, FranceEmail: [email protected]: http://www.grainlegumes.com/default.asp?id_biblio=453
November 12–16, 2007Lisbon-2007 Dissemination Conference of GLIP and other legume-related activities6th European Conference on Grain LegumesLisbon, PortugalEmail: [email protected]
July 18–22, 20083rd Euroscience Open ForumBarcelona, SpainEmail: [email protected]: http://www.esof2008.org/#|
September 14–18, 200812th International Lupin ConferenceFreemantle, Western AustraliaEmail: conference @lupins.orgWeb: http://www.lupins.org
EVENTS
NEW BOOKS
Chickpea breeding andmanagement
S. S. Yadav, R. Redden, W. Chenand B. Sharma (Eds)
April 2007, English, 448 pages
ISBN Hardback 978 184593 213 8
Indian Agricultural ResearchInstitute New Delhi, India,
Victoria Institute for DrylandAgriculture Horsham, Australia
and USDA-ARS, Washington State University, USA
This authoritative account by international experts covers allaspects of chickpea breeding and management, and the integratedpest management and biotechnology applications that are importantto its improvement. With topics covered including origin andtaxonomy, ecology, distribution and genetics, this book combinesthe many and varied research issues impacting on productionand utilisation of the chickpea crop on its journey from paddockto plate.
Price: £99.50/US$198.00/€160.00Order: CABI, Wallingford, UK. Tel.+44 (0) 1491 832111, Email: [email protected], Web: www.cabi.org/bookshop

GRAIN LEGUMES No. 49 – October 200726
AROUND THE WORLD
Pigeonpea (Cajanus cajan (L.) Millsp.,2n = 22), also called red gram, is oneof the major grain legumes in the
world. Pigeonpea is important due to itsability to grow under diverse cropping systemsand environments and its ability to recoverfrom the losses caused by various biotic andabiotic stresses. Pigeonpea is about 5% ofthe total world production of grain legumes.
Pigeonpea is grown in more than 90countries with a total cultivated area ofapproximately 4.6 million ha, and a yield ofapproximately 3.3 million tonnes worldwidein 2005. The major pigeonpea growing areais in Asia (88.8%), Africa (10.6%) and LatinAmerica (0.6%). In 2005, more than 76% ofthe pigeonpea area was in India. The othermajor pigeonpea growing countries areMyanmar, Kenya, Malawi, Uganda, Tanzania,Nepal, Dominican, Congo and Haiti (2).
China started to plant pigeonpea in the1950s. It has been used in the forestry industryand as a host for the lac insect that secretesthe resinous substance, ‘lac’. There were only4,000ha of pigeonpea being grown as a cropin China before 1997. In 2000, the sowingarea was over 15,000ha (9, 11), with a stableincrease to 60,000 ha in 2006 (12, 3).
A very versatile cropTraditionally, in the major areas of
production, pigeonpea has been used forhuman food, vegetable matter, animal feedand fuel, but it is a very versatile crop. Dhal(decorticated split seeds of pigeonpea) isthe major food for human consumptionprimarily in south Asian countries, but thisis not the case in China where eating habitsare very different. Some pigeonpea seedis used for animal feed.
At the same time pigeonpea is used as aherbal medicine in India, Indonesia, China,West Africa, Madagascar and the West Indiesfor healing wounds, destroying internalworms and curing lung diseases (5). It cancause reversion of sickled cells in patientssuffering from sickle-cell anaemia (1), andit is a resource for honeybees since itsflowering period can last a few months (9).
In recent years, however, pigeonpea hasbeen developed for use in south China insoil conservation, for forest recovery andpoverty alleviation. It is also used as a foddercrop for feeding goats , buffalo and rabbitsin the Guangxi province of China, andfor water conservation and ecological re-construction in Yunnan province of China.The reason for the unbelievably fast increasein the use of this crop is that pigeonpea helps in forest regeneration and poverty reduction being grown both as a shrub and a food legume crop. It suits the steep (>25 degrees) slopes of the karst mountainous landscapes that are dominant in the south-west part of China, aregion where different ethnicpopulations are concentratedand living conditions are poor.This area was considered a keyarea for a poverty reductionstrategy by the centralChinese Government. After
pigeonpea was proved to be superior toother crops as a ‘gap-coverage’ shrub forboth forest recovery and poverty reductionin reforestation practices, local farmersgradually developed their own methods ofpigeonpea-based agro-forestry using thenew pigeonpea varieties/cultivars.
ICM packagesIn 1997, a Chinese pigeonpea team
began to introduce these new varieties fromICRISAT for farm testing (11). Variouspigeonpea variety/farming and eco-systemcombinations have been evaluated andscreened in different targeted counties inChina from 2001 to 2006. Farmer initiativeIntegrated Crop Management (ICM)packages have been optimised with scientists’participation; and scientist designed ICMpackages have been optimised with farmers’
Pigeonpea development in ChinaDéveloppement du pois caján en Chineby Shiying YANG* and colleagues**
*Diversity Arrays Technology, Canberra, H.Graham Centre for Agricultural Innovation,Wagga Wagga, Australia and Guangxi Academyof Agricultural Sciences, P. R. China([email protected])
**As* also ICRISAT, Patancheru, India andChinese Academy of Agricultural Sciences,Beijing, P. R. China.
Fresh branches and leaves of pigeonpea provide good fodder for goatsin Longan county, Guangxi province, China.
Pigeonpea for soil conservation in Duan county, Guangxi province, China.
(Pho
to S
. Yan
g, G
uang
xi A
cade
my
of A
gric
ultu
ral S
cien
ces,
P. R
. Chi
na)
(Pho
to S
. Yan
g)

GRAIN LEGUMES No. 49 – October 200727
AROUND THE WORLD
COVER PHOTO:Top left: Field of chickpeas at Ain Temouchet, Algeria in April 2007. (Photo Houcine Irekti, INAT, Algeria)
Top right: Salt accumulation in a faba bean field in Al-Sharkia, Egypt. (Photo Y. G. Yanni, Sakha Agricultural Research Station, Egypt)Bottom: Microarray approach to identify salt stress-responsive genes from legumes (Photo ISV CNRS, France)
PHOTO DE COUVERTURE : En haut à gauche: Champ de pois chiches à Ain Temouchet, Algérie, en avril 2007. (Photo Houcine Irekti, INAT, Algeria)
En haut à droite : Accumulation de sel dans un champ de féveroles à Al-Sharkia dans le delta du Nil en Egypte. (Photo Y. Yanni, Sakha Agricultural Research Station, Egypt)
En bas : Analyse de puces d’ADN pour identifier les gènes répondant au stress salin chez les légumineuses. (Photo ISV CNRS, France)
participation. A farmers’ association basedon pigeonpea commercial grain productionand vegetable production systems has been established in the major pigeonpeaproduction area in Yunnan province (12,3). ICM packages aimed for ruminantlivestock were developed with farmerparticipation in Guangxi province at the same time. Pigeonpea intercroppingsystems for soil conservation, pigeonpeaintercropped with neem trees (Azadirachtaindica) and other perennial plants likewalnuts (Juglans spp.), are the most commonpractices for degraded soils on steep slopesalong the Nujiang and Lancang river banksin Yunnan province (12, 3). Furthermore,pigeonpea intercropping systems for moreeconomic benefits, pigeonpea intercropwith groundnuts and other crops like chilli, are the common practices in dry andhot areas like Yunmu county in Yunnanprovinces (12, 3).
Pigeonpea is also being used as a legumevegetable source in China (6), and thisuse became well developed in most of thepigeonpea sowing areas in China between2001 and 2006. The three best pigeonpeavarieties, ICPL 87091, ICP 7035 and ICP12746, used for both vegetable and dry
grain production are recommendedaccording to their performance in termsof high yield and superior quality ofgreen peas.
Genetic diversity mainly inwild relatives
A genetic diversity study using DiversityArrays Technology (DArT) for pigeonpeaand its wild relatives revealed that most ofthe genetic diversity was among the wildrelatives of pigeonpea or between the wildspecies and the cultivated pigeonpea (10).There is limited polymorphism among the cultivated accessions. Therefore, theChinese team are introducing wider ranginggenetic materials from ICRISAT whichwere progenies crossed between differentspecies as a resource to establish newbreeding systems (7, 8). Currently, the aimof pigeonpea genetic improvement activitiesin China is to select for better yield andquality, as well as better disease resistance.This work is undertaken by the GuangxiAcademy of Agricultural Sciences, theYunnan Academy of Agricultural Sciences,the Chinese Academy of AgriculturalSciences and the Chinese Academy ofForestry (12, 3, 4). �
Acknowledgments: We would like toacknowledge the Department of GuangxiScience and Technology of China, the StateAdministration of Foreign Experts Affairs ofChina and the International Fund forAgricultural Development for supporting thepigeonpea development research in the GuangxiAcademy of Agricultural Science.
(1) Ekeke, G. F. (1985). Planta Medica, p.504.
(2) FAOSTAT (2006). FAO, Rome.
(3) ICRISAT (2006a). ICRISAT Happenings(2006). 1228, 4–5.
(4) ICRISAT (2006b). ICRISAT Happenings(2006). 1228, 6.
(5) Morton, J. F. (1976). Hort. Sci. 11, 11.
(6) Saxena, K. B. and Yang, S. (2000).International Chickpea and Pigeonpea Newsletter,p.2–3.
(7) Saxena, K. B. et al. (2000). In: Proc. Int.Conf. on Engineering and TechnologicalSciences (ICETS 2000). Beijing, 499–500.
(8) Saxena, K. B. et al. (2003). In: Proc. Int.Symposium, 21–24 May 2001, Kathmandu,Nepal, 115–117.
(9) Yang, S. et al. (2001). International Chickpeaand Pigeonpea Newsletter, p.54
(10) Yang, S. et al. (2006). Theor. Appl. Genet.113, 585–595.
(11) Zong, X. X. et al. (2002). In: Pigeonpea,p.4–6. Dalian Press, Dalian, China.
(12) Zong, X. X. et al. (2006). In: Descriptorsand data standards for pigeonpea (Cajanus spp.),p.1–3. China Agriculture Press, Beijing, China.
ISSUE No. 50 NUMÉRO 50November 2007 Novembre 2007
Special report Dossier
Economic and envirEconomic and environmentalonmental Analyses économiques et Analyses économiques et analyses of grain legumesanalyses of grain legumes envirenvironnementales sur lesonnementales sur les
légumineuses à graineslégumineuses à graines
GRAINGRAIN LEGUMELEGUME SS

GRAIN LEGUMESGRAIN LEGUMES
The AEP is an associative network of persons with interests in grain legumeresearch (peas, faba beans, lupins, chickpeas, lentils, dry beans, etc.) to favour
the exchange of information and multidisciplinary collaborations (Conferences,publications, workshops, joint projects). It aims both to strengthen the research worksand to enhance the application of research into the integrated chain of grain legumes.
Technical Institutefor Cereals and Forage
Arvalis – Institut du végétal
3 rue Joseph et Marie Hackin – 75116 PARISTel: 33 (0)1 44 31 10 00 • Fax: 33 (0)1 44 31 10 10
Email: [email protected] • http://www.arvalisinstitutduvegetal.fr
APPOBelgian Association for Oilseeds and Protein Crops
Association pour la promotion des protéagineux et des oléagineux
Faculté universitaire des sciences agronomiquesPassage des déportés, 2 – 530 Gembloux – Belgium
Tel: +32 81 62 21 37 • Fax: +32 81 62 24 07 • Email: [email protected]
UFOPUnion zu Förderung von Oel- und Proteinpflanzen
AEPEuropean Association for Grain Legume Research
Association Européenne de recherche sur les Protéagineux
12 Avenue George V – 75008 Paris – FranceTel: +33 1 40 69 49 09 • Fax: +33 1 47 23 58 72
Email: [email protected] • http://www.grainlegumes.com
The UNIP is the representative organisation of all the French professionalbranches of the economic integrated chain of grain legumes. It provides
information about pulse production, utilisation, and the market and it coordinatesresearch works related to grain legumes in France, especially peas, faba beans andlupins for animal feeding.
The PGRO provides technical support for producers and users of all types ofpeas and beans. Advice is based on data from trials sited from Scotland to the
South West of England and passed to growers and processors through technicalbulletins and articles in the farming press.
Run and financed by French farmers, Arvalis-Institut du végétal carries out anddisseminates applied research on the production, storage and utilisation of
cereals, grain legumes, potatoes, and forage.
The APPO is the representative organisation of Belgian growers of oilseeds andprotein crops, especially rapeseed, peas and faba beans. The main tasks are
experimentation, giving advice to producers, providing technical and economicinformation through meetings and mailings and encouraging non-food uses ofvegetable oil.
UNIPFrench Interprofessional Organisation of Protein Crops
Union Nationale Interprofessionnelle des plantes riches en Protéines
12 Avenue George V – 75008 Paris – FranceTel: +33 1 40 69 49 14 • Fax: +33 1 47 23 58 72
Email: [email protected] • http://www.prolea.com/unip
Processors and Growers Research Organisation
Research Station – ThornhaughPE8 6HJ Peterborough – United Kingdom
Tel: +44 1780 78 25 85 • Fax: +44 1780 78 39 93Email: [email protected] • http://www.pgro.co.uk
UFOP is the representative organisation for German producers of oil and proteincrops. It encourages professional communication, supports the dissemination of
technical information on these crops and also supports research programmes to improvetheir production and use.
Pulse Canada is a national industry association. This organisation representsprovincial pulse grower groups from Alberta, Saskatchewan, Manitoba, Ontario
and the pulse trade from across Canada who are members of the Canadian SpecialCrops Association. Pulse crops include peas, lentils, beans and chickpeas.
1212–220 Portage Avenue – WinnipegManitoba – Canada R3C 0A5
Tel: +306 651 0858 • Fax: +306 668 5557Email: [email protected] • http://www.pulsecanada.com
Abroad research topic of the Animal Production and Nutrition Department dealswith the utilisation of lupin and pea seeds in animal feeding (ruminant,
monogastric and poultry) in terms of nutritional value, environmental benefits, proteinutilisation and economic aspects. The research is also concerned with the development oflegume silages, seed treatments prior to feeding and seed processing for non-food uses.
CRA-W
Animal Production and Nutrition DepartmentRue de Liroux 8 – 5030 Gembloux (Belgique)
Tel: +32 81 62 67 70 • Fax: +32 81 61 58 68 Email: [email protected] • http://cra.wallonie.be
Andreas Hermes Haus – Godesberger Allee 142-148 – 53175 Bonn – GermanyTel: +49 22 88 19 82 27 • Fax: +49 22 88 19 82 03Email: [email protected] • http: //www.ufop.de





























