Embed Size (px)
Citation preview

EntomoL exp. appl. 52: 191-203, 1989. �9 1989 Kluwer Academic Publishers. Printed in Belgium. 191
Green leaf volatiles enhance aggregation pheromone of boll weevil, Anthonomus grandis
Joseph C. Dickens USDA-ARS, Boll Weevil Research Unit, Mississippi State, MS 39762, USA
Accepted: May 9, 1989
Key words: Green leafvolatiles, green odor, green receptor, pheromone, synergist, plant odor, Anthonomus grandis, boll weevil, electroantennogram, single cell, behavior, host plant
Abstract
Enhancement of an insect pheromone response by green leaf volatiles is reported for the first time in the boll weevil, Anthonomus grandis Boh. (Coleoptera: Curculionidae). Single cell recordings from antennal olfactory receptors revealed a class of cells selectively responsive to six carbon alcohols and aldehydes (i.e., 'green leaf volatiles'). Field tests with released weevils demonstrated enhanced trap captures with trans-2-hexen-l-ol, cis-3-hexen-l-ol, or 1-hexanol paired with grandlure, the boll weevil aggregation phero- mone, when in competition with grandlure alone. Although dose-response curves constructed from electroantennograms were indicative of similar populations of receptor cells for selected six carbon alcohols, one of the compounds tested, cis-2-hexen-l-ol, was inactive in field tests. Trans-2-hexenal was active in single cell recordings, but was also inactive in field tests. In tests in cotton fields with indigenous weevil populations, trans-2-hexen-l-ol not only enhanced pheromone trap captures, but also extended the longevity of attractiveness of pheromone-baited traps. The combined electrophysiological and field data support 'across-fiber coding' of green leaf volatiles by boll weevil olfactory receptors. The results are discussed with regard to the chemistry of the host plant of the boll weevil, cotton (Gossypium hirsutum L.), and potential economic significance for boll weevil eradication/suppression.
Introduction
The importance of olfaction in the orientation of insects to mates and host plants is well-known (Dickens & Payne, 1985). Insect-produced vola- tiles, e.g. pheromones, and host plant odors, e.g. kalromones, may facilitate location of conspe- cifics for mating, and orientation to acceptable host plants for feeding and oviposition. In several insect species, including bark beetles (Borden, 1984) and the boll weevil (Chang et aL, 1986), certain monoterpenes may act in combination
with pheromones, each enhancing the others attraction.
The boll weevil, Anthonomus grandis Boh. (Coleoptera: Curculionidae) is a narrowly oligo- phagous insect which feeds primarily on cotton, Gossypium hirsutum L., and closely related Malvaceae (Cross etal., 1975). Once male boll weevils locate their host plant, feeding ensues, and the weevils release in their frass an aggre- gation pheromone (Tumlinson et aL, 1969). Elec- troantennogram (EAG) recordings revealed populations of receptors responsive to both

192
pheromone and plant odors (Dickens, 1984), including beta-bisabolol, a volatile reported only from cotton and its close relatives (Thompson etaL, 1970; 1971b). More recently, recordings from single olfactory cells on the antenna showed cells specialized for individual pheromone com- ponents (Dickens & Mori, 1989).
Green leaf volatiles are six carbon alcohols and aldehydes which are products of oxidative degradation of plant lipids, e.g. linoleic and linolenic acids (Visser et al., 1979). Commonly reported members of this group of plant volatiles, whose ratios vary among different plant species, are: l-hexanol, trans-2-hexen-l-ol, cis-3-hexen-l-ol and their corresponding aldehydes. Previous reports have identified 1-hexanol, trans-2-hexen- 1-ol, cis-3-hexen-l-ol, hexanal, and trans-2-hexenal from cotton bud extracts (Hedin etal., 1973), while l-hexanol (Hedin etal., 1975) and trans- 2-hexenol (Thompson etaL, 1970) have been identified as volatiles emitted from growing cotton. These compounds elicit electrophysio- logical responses from antennal olfactory recep- tors of many insect species (Visser, 1986), includ- ing the boll weevil (Dickens, 1984) and several other Coleoptera, e.g. the Colorado beetle, Leptinotarsa decemlineata Say (Ma & Visser, 1978; Visser, 1979), the oak flea weevil, Rhynchaenus quercus (Kozlowski & Visser, 1981), and the chrysomelid, Trirhabda bacharides Weber, (Dickens & Boldt, 1985).
In the Colorado beetle, the blend of green leaf volatiles emitted from its host plant is thought to facilitate host location by the beetle over long distances (Visser & Ave, 1978). However in several instances, individual green odor com- ponents have been assigned specific functions, e.g.: 1) hexanol as an attractant for the black fig fly, Silba adipata McAlpine (Katsoyannos & Guerin, 1984), and hexanal and trans-2-hexenal as attractants for the carrot fly, Psila rosae (Guerin et al., 1983); 2) hexanal as a trap capture enhancer when released with another plant vola- tile, trans-asarone, for the carrot fly (Guerin et al., 1983); and 3) trans-2-hexenol as a defensive secre- tion in the plant bug, Hotea gambiae (Westwood) (Hamilton et al., 1985).
The purposes of the experiments reported here were to determine the mechanisms by which green leaf volatiles are detected by the boll weevil, and the possible role of the volatiles in boll weevil behavior. Due to our preliminary behavioral studies, experiments were also performed to deter- mine any practical application for these compounds in trapping of boll weevils in phero- mone traps. An abstract of the current results was previously published (Dickens, 1988).
Methods and materials
Insects
Adult A. grandis used in field releases and electro- physiological experiments were obtained from a small laboratory colony annually infused with feral insects. Upon emergence, insects were sexed and fed cotton squares. Insects were not used until olfactory receptor maturation at four days postemergence (Dickens & Moorman, 1987). For electrophysiological studies, groups of five insects of the same sex were maintained on moist paper (Whatman T M I) in Petri dishes until use at four to six days of age. For field experiments, groups of fifty insects of the same sex were held in paper cartons (ca. 0.5 liter) with screen tops. Insects used in field releases were six to thirteen days postemergence. All insects were held in incuba- tors at 26 ~ under a photoregime of 16 h of light (ca. 700 lux) and 8 h of darkness.
Chemicals
The boll weevil aggregation pheromone, grand- lure, is composed of compounds I [racemic grandisol, ( + )-cis-2-isopropenyl-l-methylcyclo- butaneethanol]; II (cis-3,3-dimethyl-A la-cyclo- hexaneethanol); and III + IV (a 50 : 50 cis: trans mixture of 3,3-dimethyl-AL~-cyclohexaneacet - aldehyde). Each component was > 95 ~ chemi- cally pure as obtained from Albany International, Controlled Release Division, Buckeye, AZ.
Chemicals used (purity given in parentheses)

were: trans- 2-hexen-l-ol (97~0), cis- 3-hex en-l-ol (98~/o), cis-2-hexen-l-ol (92~), l-hexanol (98~o), hexanal (99~o), trans-2-hexenal (99~o), heptanal (95 ~o ), nonanal (98 ~ ), (-)-atpha-pinene (98 ~o), (-)-limonene (97~o), myrcene (85~o), linalool (99~/o), and benzaldehyde (>98~o) (Aldrich Chemical Co., Milwaukee, WI); nerol (> 65~o) and geraniol (>90~o) (Pfaltz & Bauer, Inc., Stamford, CN); beta-caryophyUene (92~o) (ICN-K&K Laboratories, Plainview, NY); alpha- bisabolol (92~o) and beta-bisabolol (83~o) (obtained from P.A. Hedin, USDA-ARS, Crop Science Research Laboratory, Mississippi State, MS); gossonerol ( > 9 8 ~ ) (obtained from H.J. Williams, Department of Entomology, Texas A&M University). Serial dilutions of experi- mental odorants were prepared in hexane. 1-Hexanol used as a standard was diluted in pen- tane as in previously reported experiments on boll weevil olfaction (Dickens, 1984; Dickens & Mori, 1989; Dickens & Prestwich, 1989).
Electrophysiology
Single cell recordings. Single cell recording tech- niques are described in detail elsewhere (Dickens, 1979; Dickens & Mori, 1989). In brief, micro- electrodes were constructed from 50.8 #m diame- ter tungsten wire electrolytically sharpened to a tip of 1-2 gm. The recording electrode was posi- tioned near either the proximal edge of one of the three sensory bands encircling the club, or the base of an individual sensillum just distal to the sensory band region. The ground electrode was inserted in the distal end of the scape. Action potentials were amplified by a Grass P-15 pre- amplifier, stored on cassette tapes using a Teac R51-D data recorder, and afterwards photo- graphed from the oscilloscope with a Tektronix C-5C camera.
Upon obtaining a single cell preparation, the first two stimuli tested were volatiles emanating from 1 #g of grandlure; followed by volatiles emanating from a crushed cotton square. These two stimuli served to determine whether the cell was activated by the aggregation pheromone of
193
the boll weevil and/or volatiles emanating from its host and food source.
If the cell were activated by volatiles from the crushed cotton square, then presentation of I #g doses of other selected odorants followed. Stimuli were selected based on their presence in the cotton plant (Hedin etal., 1973) and their activity as determined in a previous EAG study (Dickens, 1984). Odorous stimuli were delivered as 1 #1 ali- quots placed on filter paper (8 mm x 18 ram) inserted into glass cartridges (80 mm x 5 mrn ID) oriented toward the preparation from a distance of 1 cm. Molecules evaporating from the filter paper were carried over the preparation by hydro- carbon-free air which also had been filtered and dried. Stimulus duration was 0.5 sec with an air flow of ca. 1 liter/rain. Two or three minutes were allowed between each stimulation.
Electroantennograms. EAG techniques were de- scribed in detail elsewhere (Dickens, 1979; 1984). Briefly, two glass microcapillaries filled with physiological saline (Pantin, 1948) were inserted into the antenna of an intact animal: the ground electrode was placed into the dista/ end of the scape and the recording electrode into the distal end of the club. Electrical contact was provided by a Ag-AgC1 wire placed into the shank of each capillary. Electrical activity was amplified by a Grass P-16 DC preamplifier. EAGs were recorded using a stripchart recorder.
Serial dilutions of trans-2-hexen-l-ol, cis- 3-hexen-l-ol, cis-2-hexen-l-ol, and l-hexanol were delivered from the lowest to the highest concen- tration. Apparatus for stimulus delivery and air flow were the same as described for single cell stimulations. Stimulus duration was ca. 1 sec. Three minutes were allowed between stimuli at lower dosages; while five to seven minutes were allowed at dosages greater than 1 gg. These times were adequate for complete recovery of the EAG (Dickens, 1984).
Three replicates were recorded for serial dilu- tions of each six carbon alcohol for both male and female A. grandis. 1-Hexanol (100/lg; 10 #1 of 10 #g//ll dissolved in pentane) served as a stand- ard to: 1) normalize responses; 2) to control for

194
viability and constancy of the preparation; 3)to allow relative comparisons with previously recorded species (Dickens, 1984; Dickens & Boldt, 1985). Stimulation with the standard both preceded and followed each serial dilution level. Although preparations were stimulated with a sol- vent control, seldom was a response noted. EAGs to test chemicals were expressed as a percent of the mean of the two nearest responses to the standard (Dickens, 1981; 1984). The threshold of response was considered to be the dosage at which the standard error of the mean was not equal to or less than zero (Dickens, 1984).
Behavior
Traps used for all field experiments were those currently used by the Boll Weevil Eradication Program which were a modification of a previ- ously described trap (Mitchell et al., 1976).
Competitive field tests with released weevils. Two series of field tests involving released boll weevils in which grandlure was placed in competition with grandlure + a green leaf volatile, and in one instance, a green leaf volatile alone, were per- formed. In order to avoid competition with cotton or pheromone-producing weevils, these initial tests were conducted in open fields located at least 1 mile from cotton fields. In the first series of experiments, one trap was placed at each corner of a 20 m equilateral triangle. The three treat- ments released were: 1) grandlure 3 mg; 2) grand- lure 3 mg + trans-2-hexen-l-ol 100 mg; and 3) trans-2-hexen-l-ol 100 mg. Cigarette filters were used as release substrates for the experi- mental odorants which had been diluted in 0.5 ml of the following mixture by weight: polyethylene glycol 1000, 2 0 ~ ; methanol, 42.5~o; glycerol, 25~o; and distilled water, 12.5~o (McKibben etal., 1980). The ratio of the pheromone com- ponents used in the cigarette filters was [1(3) : 11(4) : III + IV(3)] that currently used in the Boll Weevil Eradication Program (BWEP). This ratio was chosen for use in the BWEP based on economic considerations even though the ratio of
components in the natural pheromone was more attractive in competitive tests (Hardee, 1974). Grandlure and trans-2-hexen-l-ol were released from separate filters. One hundred insects (50 males and 50 females) were released along a 20 m line parallel to each side of the triangle at a dis- tance of 20 m (300 insects total) at ca. 1000 h prior to baiting of the traps at 1200 h. Traps were checked and insects removed at 1400 h, 1600 h, and 1800 h, the time at which the test was termi- nated. This experiment was replicated three times.
The second series of experiments involved an array of four traps. One trap was placed at each corner (four traps total) of a 20 m square. The two treatments being tested simultaneously in this competitive arrangement were placed alternately in the traps, i.e. the two traps baited with the same treatment were located diagonally from each other. Odorants diluted in hexane were released from 2.2 ml vials as 10/~g/#l dilutions using glass capillaries (Dickens & Mori, 1989; Dickens & Prestwich, 1989). One vial placed in each trap contained ( + )-I (0.4 mg); a second vial contained II (0.3 mg) and III + IV (0.1 mg). The ratio of the pheromone components used in these studies [I(4) : II(3) : III + IV(I)] approximated that of the natural pheromone as identified from the frass produced by male boll weevils (Tumlinson et al., 1969). A third vial contained the green leaf volatile being tested. Following transport to the field in a cooler, the teflon-lined screw caps were removed from the vials and both caps and vials were placed in the top of an aluminum film container (3 cm dia. • 4.3 cm height) that had 5 holes (0.23 mm) drilled near its outer edge. The bottom of the can was then screwed onto the top and the inverted film container was placed in the trap below the screen funnel.
One hundred insects (50 males + 50 females) were released along a 20 m line parallel to each side of the square at a distance of 20 m (400 insects total) at 1400 h. Traps were then immedi- ately baited. In each test, traps were checked and insects removed at 1530 h and 1700 h on the day of release, and before 0900 h the next day (the time at which the test was terminated). Treat-

ments tested in this competitive arrangement were: grandlure 0.8 mg vs a.) grandlure 0.8 mg + trans-2-hexen-l-ol 0.1 mg; b.) grandlure 0.8 mg + trans-2-hexen-l-ol 1.0 mg; c.) grandlure 0.8 mg + cis-3-hexen-l-ol 1.0 mg; d.) grandlure 0.8 mg + l-hexanol 1.0 mg; e.) grandlure 0.8 mg + trans-2- hexenal 1.0 mg; f.) grandlure 0.8 mg + cis-2- hexen-l-ol 1.0 mg; and g.) grand-lure 0.8 mg + trans-2-hexen-l-ol 1.0 mg vs grand-lure 0.8 mg + l-hexanol 1.0 mg. Each of these tests was repli- cated five times with the exception of 'g.' which was replicated three times.
Competitive field tests to cotton fields with natural boll weevilpopulations. Tests in three cotton fields located in Oktibbeha Co., MS (ca. 0.4 hectare each) were used to determine the effectiveness of trans-2-hexen-l-ol in enhancing the attractiveness of grandlure for operational use. The tests were conducted in September and October, 1987 when the cotton was mature. Cigarette filters were used to release the experimental odorants which had been diluted in 0.5 ml of the previously defined solvent. The ratio of pheromone used in the cigarette filters was [1(3) : II(4) : III + IV(3)] the economical formula currently used in the BWEP. Grandlure and trans-2-hexen-l-ol were released from separate filters.
The experimental design consisted of placing one trap at each comer of a 20 m square within a cotton field. Four additional traps were placed ca. 2 meters outside the cotton field at a distance of ca. 40-60 m from the traps in the field, and located near imaginary diagonal lines extending through the comers of the square within the field. Each trap contained a cigarette filter with 1 mg of grandlure. Cigarette filters containing trans- 2-hexen-l-ol were placed alternately in the traps. The result resembled an 'X' array of traps placed across each field with each arm of the 'X' consist- ing of four traps, two within and two outside of the field. Traps within one arm contained grandlure (1 mg) alone; those within the other arm were baited with grandlure (1 mg) + trans-2-hexen-l-ol at a particular dosage. Only one dosage of trans- 2-hexen-l-ol occurred in each field. Dosages of trans-2-hexen-l-ol tested were: 1 mg, 10 mg, and
195
100 mg. Traps were freshly baited each week, and checked daily and weevils removed at 1500 h for the initial three days and at the end of the week. This experiment was replicated four times over a four week period.
An additional experiment using the same experimental design as above was performed to determine the longevity of attractiveness of traps baited with grandlure at 1 mg alone and grandlure at 1 mg + trans-2-hexen-l-ol at 1 mg, 10 mg, and 100 mg. In this experiment, baits remained in the traps for two weeks, and weevils were collected daily at 1500 h for the f'trst three days of each week and at the end of each week.
Statistical analyses
EAGs were evaluated by analysis of variance for two factors, namely treatment and sex (Ostle, 1963). Where differences were noted, compari- sons were made using Duncan's new multiple range test (Duncan, 1955). Results of the first field test with the triangular array of traps were also compared by analysis of variance and Duncan's new multiple range test. Trap captures of released weevils using the four trap array were analysed for significant differences by a paired t-test (Ostle, 1963).
Competitive tests in cotton fields with natural boll weevil populations were evaluated by a paired t-test (Ostle, 1963). For the first test in which baits were changed weekly, total trap captures for each test for each week were compared (Table 4). For the second experiment in which baits remained in the traps for a two week period, numbers of insects captured for each check for each competi- tive treatment were compared (Table 5).
Results
Electrophysiology
Single cell recordings. Recordings from 45 indi- vidual olfactory cells from 10males and 14 females revealed 18 cells which were responsive to
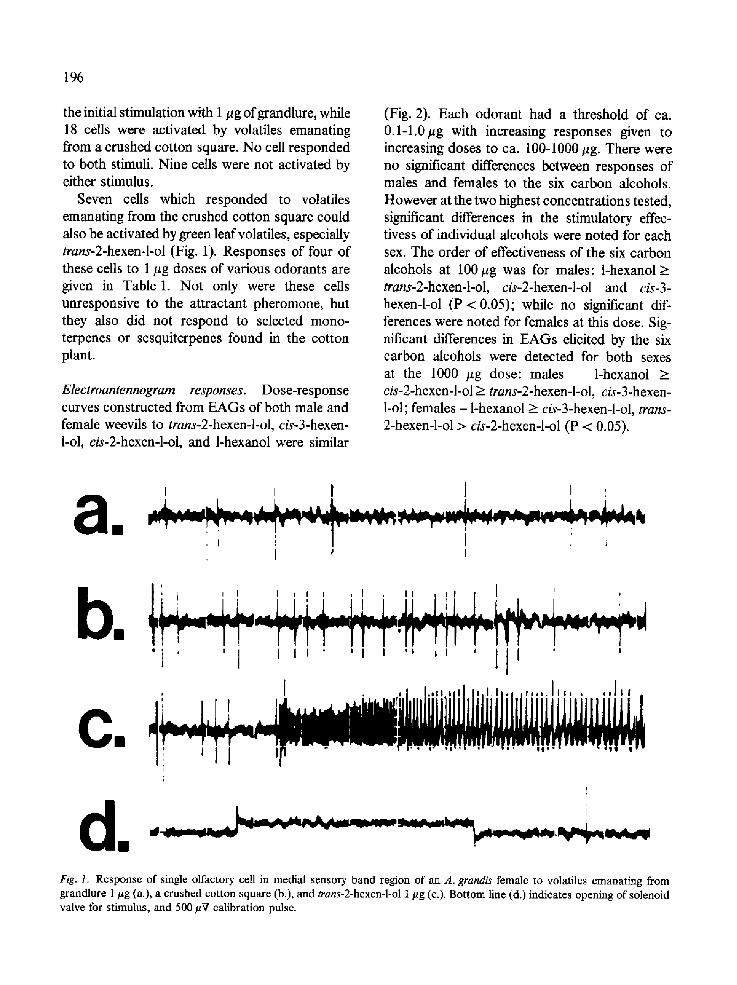
196
the initial stimulation with i #g ofgrandlure, while 18 cells were activated by volatiles emanating from a crushed cotton square. No cell responded to both stimuli. Nine cells were not activated by either stimulus.
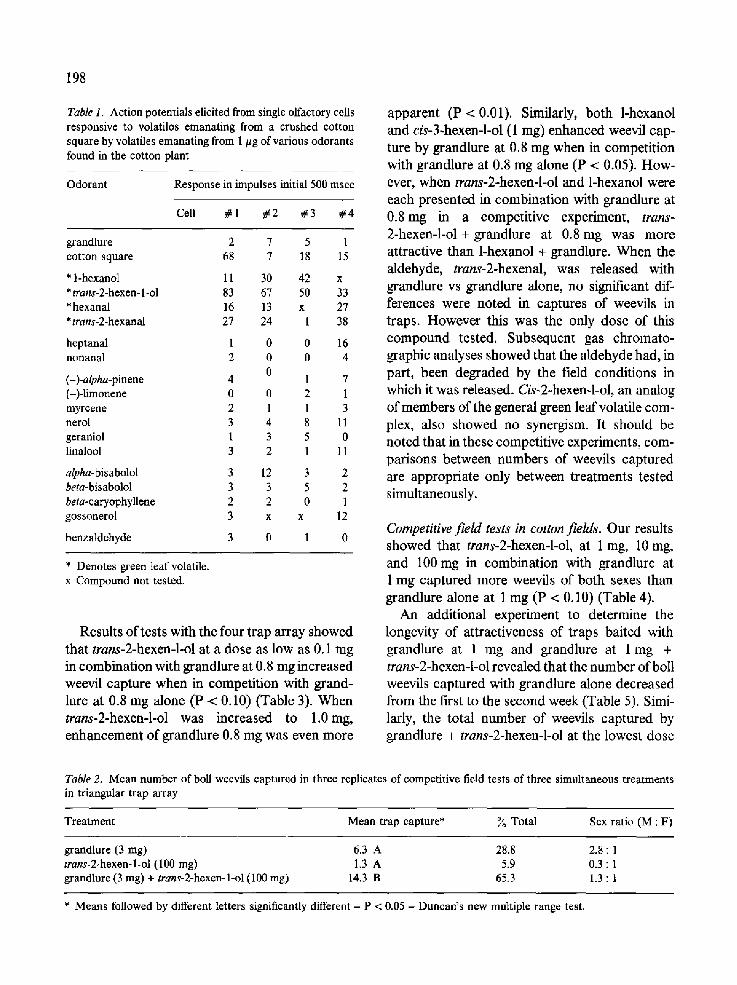
Seven cells which responded to volatiles emanating from the crushed cotton square could also be activated by green leaf volatiles, especially trans-2-hexen-l-ol (Fig. 1). Responses of four of these cells to 1 #g doses of various odorants are given in Table 1. Not only were these cells unresponsive to the attractant pheromone, but they also did not respond to selected mono- terpenes or sesquiterpenes found in the cotton plant.
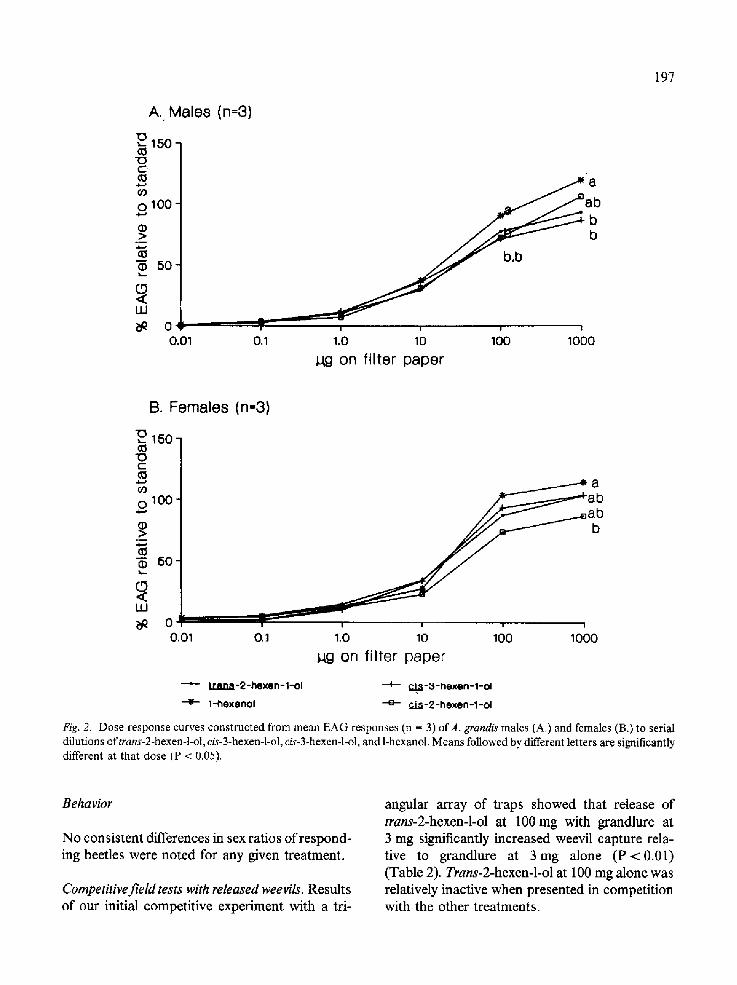
Electroantennogram responses. Dose-response curves constructed from EAGs of both male and female weevils to trans-2-hexen-l-ol, cis-3-hexen- 1-ol, cis-2-hexen-l-ol, and l-hexanol were similar
(Fig. 2). Each odorant had a threshold of ca. 0.1-1.0/~g with increasing responses given to increasing doses to ca. 100-1000 #g. There were no significant differences between responses of males and females to the six carbon alcohols. However at the two highest concentrations tested, significant differences in the stimulatory effec- tivess of individual alcohols were noted for each sex. The order of effectiveness of the six carbon alcohols at 100#g was for males: 1-hexanol > trans-2-hexen-l-ol, cis-2-hexen-l-ol and cis-3- hexen-l-ol (P < 0.05); while no significant dif- ferences were noted for females at this dose. Sig- nificant differences in EAGs elicited by the six carbon alcohols were detected for both sexes at the 1000 #g dose: males - 1-hexanol > cis-2-hexen-l-ol > trans-2-hexen-l-ol, cis-3-hexen- 1-ol; females - 1-hexanol > cis-3-hexen-l-ol, trans- 2-hexen-l-ol > cis-2-hexen-l-ol (P < 0.05).
I I
b ' [ I , i , , I . I I I I i , I ' i ~ r ' J~LL .J . , i a_J.J, - L y._. LI . [ i__J_L_ L. JJ L,,I, . . ,L.- . ,_._~J �9 r r r - ] ] r-rrT �9
C l l
'1 ' ' I I I I ' '1 , " , , ' 1] I
' 11 it, . . . . . " . . . . . . . ' . . . . . " " -
I
d k . - - - . - - . . . . . . . . ~L. ::
Fig. 1. Response of single olfactory cell in medial sensory band region of an A. grandis female to volatiles emanating from grandlure 1 #g (a.), a crushed cotton square (b.), and trans-2-hexen-l-ol 1/zg (c.). Bottom line (d.) indicates opening of solenoid valve for stimulus, and 500 #V calibration pulse.
A, Males (n=3)
"0 1 5 0 -
r
o l O 0 -
>o
-~ 50-
UJ
O~ 0,01 0.1 1.0 10 100 1000
lag on fi l ter paper
197
B. Females (n-3)
150 r "0
0 100
60
W
0 0.01
T f T 1
o,1 1.o io 1oo 1ooo l.Lg on fi l ter paper
�9 trAns-2-hexen-l-oI ! Gj~-3-hexen-l-ol
--IK-- 1-hexsnol o c is-2-hexen- l -o l
Fig. 2. Dose-response curves constructed from mean EAG responses (n = 3) ofA. grandis males (A.) and females (B.) to serial dilutions oftrans-2-hexen-l-ol, c/s-3-hexen-l-ol, cis-3-hexen-l-ol, and 1-hexanol. Means followed by different letters are significantly different at that dose (P < 0.05).
Behavior
No consistent differences in sex ratios of respond- ing beetles were noted for any given treatment.
Competitive field tests with released weevils. Results of our initial competitive experiment with a tri-
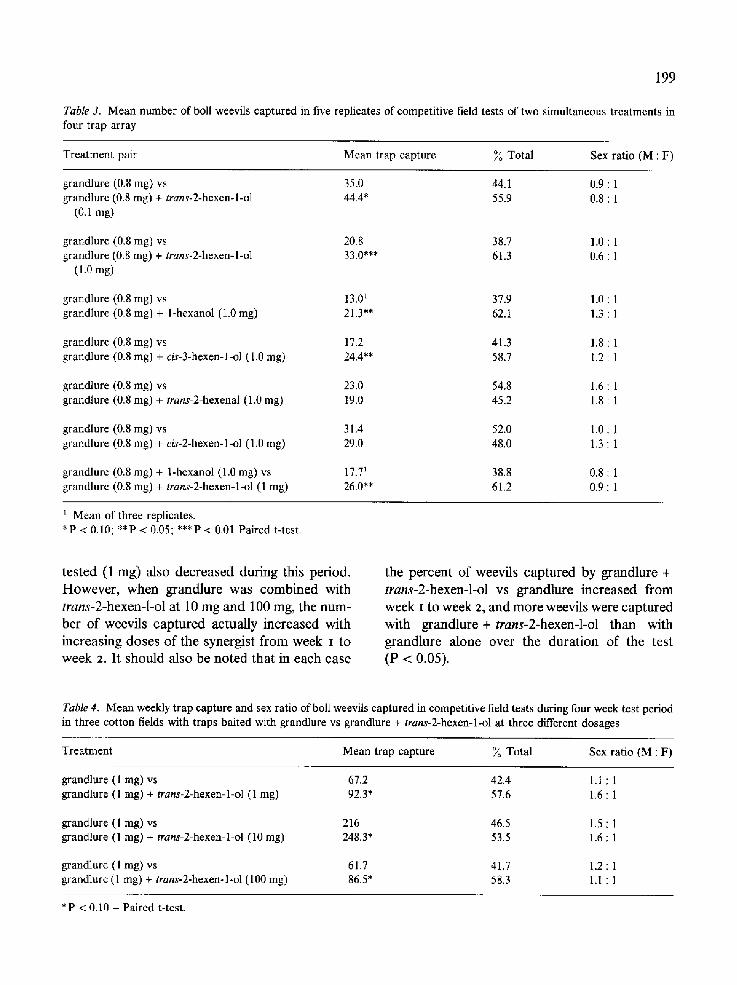
angular array of traps showed that release of trans-2-hexen-l-ol at 100 mg with grandlure at 3 mg significantly increased weevil capture rela- tive to grandlure at 3mg alone (P<0 .01 ) (Table 2). Trans-2-hexen-l-ol at 100 mg alone was relatively inactive when presented in competition with the other treatments.
198
Table 1. Action potentials elicited from single olfactory cells responsive to volatilos emanating from a crushed cotton square by volatiles emanating from 1/~g of various odorants found in the cotton plant
Odorant Response in impulses initial 500 msec
Cell # 1 #2 #3 #4
grandlure 2 7 5 1 cotton square 68 7 18 15
* l-hexanol 11 30 42 x *trans-2-hexen-l-ol 83 67 50 33 *hexanal 16 13 x 27 *trans-2-hexanal 27 24 1 38
heptanal 1 0 0 16 nonanal 2 0 0 4
0 (-)-alpha-pinene 4 1 7 (-)-limonene 0 0 2 1 myrcene 2 1 1 3 nerol 3 4 8 11 geraniol 1 3 5 0 linalool 3 2 1 11
alpha-bisabolol 3 12 3 2 beta-bisabolol 3 3 5 2 beta-caryophyllene 2 2 0 1 gossonerol 3 x x 12
benzaldehyde 3 0 1 0
* Denotes green leaf volatile. x Compound not tested.
Results of tests with the four trap array showed that trans-2-hexen-l-ol at a dose as low as 0.1 mg in combination with grandlure at 0.8 mg increased weevil capture when in competi t ion with grand- lure at 0.8 mg alone (P < 0.10) (Table 3). When trans-2-hexen-l-ol was increased to 1.0rag, enhancement of grandlure 0.8 mg was even more
apparent (P < 0.01). Similarly, both l-hexanol and cis-3-hexen-l-ol (1 mg) enhanced weevil cap- ture by grandlure at 0.8 mg when in competit ion with grandlure at 0.8 mg alone (P < 0.05). How- ever, when trans-2-hexen-l-ol and 1-hexanol were each presented in combinat ion with grandlure at 0 . 8mg in a competitive experiment, trans-
2-hexen-l-ol + grandlure at 0 . 8mg was more attractive than 1-hexanol + grandlure. When the aldehyde, trans-2-hexenal, was released with grandlure vs grandlure alone, no significant dif- ferences were noted in captures of weevils in traps. However this was the only dose of this compound tested. Subsequent gas chromato- graphic analyses showed that the aldehyde had, in part, been degraded by the field conditions in which it was released. Cis-2-hexen-l-ol, an analog of members of the general green leaf volatile com- plex, also showed no synergism. It should be noted that in these competitive experiments, com- parisons between numbers of weevils captured are appropriate only between treatments tested simultaneously.
Competitive f ield tests in cotton fields. Our results showed that trans-2-hexen-l-ol, at i mg, 10 mg, and 100 mg in combination with grandlure at 1 mg captured more weevils of both sexes than grandlure alone at i mg (P < 0.10) (Table 4).
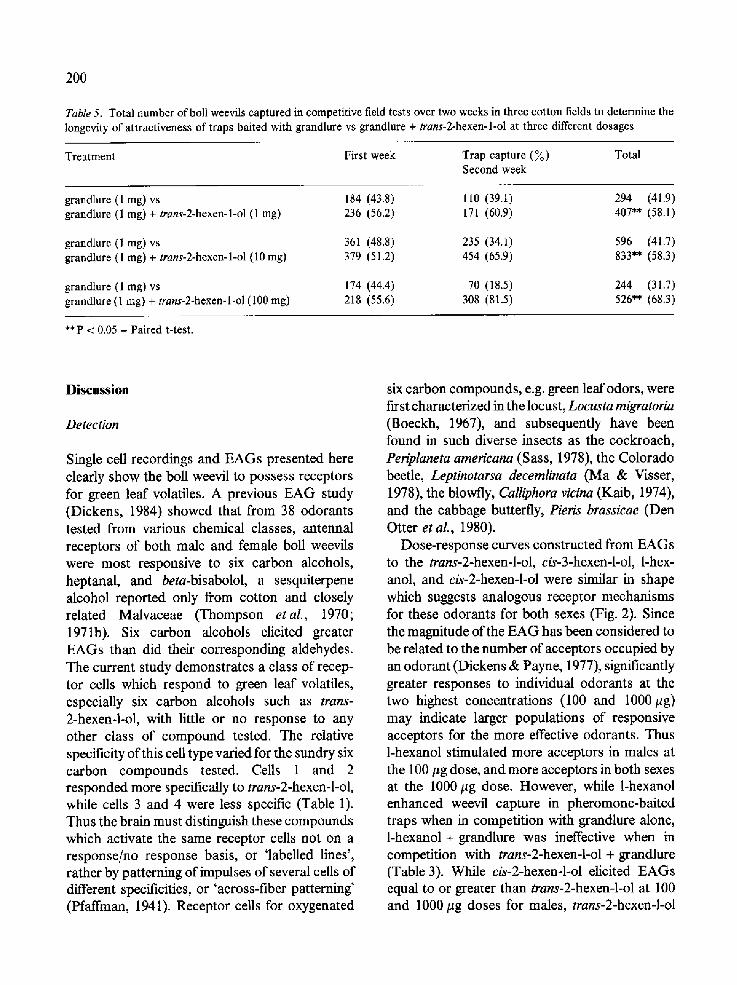
An additional experiment to determine the longevity of attractiveness of traps baited with grandlure at 1 mg and grandlure at 1 mg + trans-2-hexen-l-ol revealed that the number of boll weevils captured with grandlure alone decreased from the first to the second week (Table 5). Simi- larly, the total number of weevils captured by grandlure + trans-2-hexen-l-ol at the lowest dose
Table 2. Mean number of boll weevils captured in three replicates of competitive field tests of three simultaneous treatments in triangular trap array
Treatment Mean trap capture* % Total Sex ratio (M : F)
grandlure (3 mg) 6.3 A 28.8 2.8 : 1 trans-2-hexen-l-ol (100 mg) 1.3 A 5.9 0.3 : 1 grandlure (3 mg) + trans-2-hexen-l-ol (100 mg) 14.3 B 65.3 1.3 : 1
* Means followed by different letters significantly different - P < 0.05 - Duncan's new multiple range test.
199
Table 3. Mean number of boll weevils captured in five replicates of competitive field tests of two simultaneous treatments in four trap array
Treatment pair Mean trap capture % Total Sex ratio (M : F)
grandlure (0.8 mg) vs 35.0 44.1 0.9 : 1 grandlure (0.8 mg) + trans-2-hexen-l-ol 44.4* 55.9 0.8 : 1
(0.1 mg)
grandlure (0.8 mg) vs 20.8 38.7 1.0 : 1 grandlure (0.8 mg) + trans-2-hexen-l-ol 33.0*** 61.3 0.6 : 1
(1.0 mg)
grandlure (0.8 mg) vs 13.01 37.9 1.0 : 1 grandlure (0.8 mg) + 1-hexanol (1.0 mg) 21.3"* 62.1 1.3:1
grandlure (0.8 mg) vs 17.2 41.3 1.8 : 1 grandlure (0.8 mg) + cis-3-hexen-l-ol (1.0 mg) 24.4** 58.7 1.2:1
grandlure (0.8 mg) vs 23.0 54.8 1.6 : 1 grandlure (0.8 mg) + trans-2-hexenal (1.0 mg) 19.0 45.2 1.8 : 1
grandlure (0.8 rag) vs 31.4 52.0 1.0 : 1 grandlure (0.8 mg) + cis-2-hexen-l-ol (1.0 mg) 29.0 48.0 1.3 : 1
grandlure (0.8 mg) + 1-hexanol (1.0 mg) vs 17.71 38.8 0.8 : 1 grandlure (0.8 mg) + trans-2-hexen-l-ol (1 mg) 26.0** 61.2 0.9 : 1
1 Mean of three replicates. *P < 0.10; **P < 0.05; ***P < 0.01 Paired t-test.
tested (1 mg) also decreased during this period. However, when grandlure was combined with trans-2-hexen-l-ol at 10 mg and 100 mg, the num- ber of weevils captured actually increased with increasing doses of the synergist from week I to week 2. It should also be noted that in each case
the percent of weevils captured by grandlure + trans-2-hexen-l-ol vs grandlure increased from week I to week 2, and more weevils were captured with grandlure + trans-2-hexen-l-ol than with grandlure alone over the duration of the test (P < 0.05).
Table 4. Mean weekly trap capture and sex ratio of boll weevils captured in competitive field tests during four week test period in three cotton fields with traps baited with grandlure vs grandlure + trans-2-hexen-l-ol at three different dosages
Treatment Mean trap capture % Total Sex ratio (M : F)
grandlure (1 mg) vs grandlure (1 mg) + trans-2-hexen-l-ol (1 mg)
grandlure (1 mg) vs grandlure (1 mg) + trans-2-hexen-l-ol (10 mg)
grandlure (1 mg) vs grandlure (1 mg) + trans-2-hexen-l-ol (100 mg)
67.2 42.4 1.1 : 1 92.3* 57.6 1.6 : 1
216 46.5 1.5 : 1 248.3* 53.5 1.6 : 1
61.7 41.7 1.2 : 1 86.5* 58.3 1.1 : 1
*P < 0.10 - Paired t-test.
2O0
Table 5. Total number of boll weevils captured in competitive field tests over two weeks in three cotton fields to determine the longevity of attractiveness of traps baited with grandlure vs grandlure + trans-2-hexen-l-ol at three different dosages
Treatment First week Trap capture (~o) Total Second week
grandlure (1 mg) vs 184 (43.8) 110 (39.1) 294 (41.9) grandlure (1 mg) + trans-2-hexen-l-ol (1 mg) 236 (56.2) 171 (60.9) 407** (58.1)
grandlure (1 mg) vs 361 (48.8) 235 (34.1) grandlure (1 mg) + trans-2-hexen-l-ol (10 mg) 379 (51.2) 454 (65.9)
grandlure (1 mg) vs 174 (44.4) 70 (18.5) grandlure (1 mg) + trans-2-hexen-l-ol (100 mg) 218 (55.6) 308 (81.5)
596 (41.7) 833** (58.3)
244 (31.7) 526** (68.3)
** P < 0.05 - Paired t-test.
Discussion
Detection
Single cell recordings and EAGs presented here clearly show the boll weevil to possess receptors for green leaf volatiles. A previous EAG study (Dickens, 1984) showed that from 38 odorants tested from various chemical classes, antennal receptors of both male and female boll weevils were most responsive to six carbon alcohols, heptanal, and beta-bisabolol, a sesquiterpene alcohol reported only from cotton and closely related Malvaceae (Thompson etal., 1970; 1971b). Six carbon alcohols elicited greater EAGs than did their corresponding aldehydes. The current study demonstrates a class of recep- tor cells which respond to green leaf volatiles, especially six carbon alcohols such as trans- 2-hexen-l-ol, with little or no response to any other class of compound tested. The relative specificity of this cell type varied for the sundry six carbon compounds tested. Cells 1 and 2 responded more specifically to trans-2-hexen-l-ol, while cells 3 and 4 were less specific (Table 1). Thus the brain must distinguish these compounds which activate the same receptor cells not on a response/no response basis, or 'labelled lines', rather by patterning of impulses of several ceils of different specificities, or 'across-fiber patterning' (Pfaffman, 1941). Receptor cells for oxygenated
six carbon compounds, e.g. green leaf odors, were first characterized in the locust, Locusta migratoria (Boeckh, 1967), and subsequently have been found in such diverse insects as the cockroach, Periplaneta americana (Sass, 1978), the Colorado beetle, Leptinotarsa decemlinata (Ma & Visser, 1978), the blowfly, Calliphora vicina (Kaib, 1974), and the cabbage butterfly, Pieris brassicae (Den Otter et aL, 1980).
Dose-response curves constructed from EAGs to the trans-2-hexen-l-ol, cis-3-hexen-l-ol, 1-hex- anol, and cis-2-hexen-l-ol were similar in shape which suggests analogous receptor mechanisms for these odorants for both sexes (Fig. 2). Since the magnitude of the EAG has been considered to be related to the number of acceptors occupied by an odorant (Dickens & Payne, 1977), significantly greater responses to individual odorants at the two highest concentrations (100 and 1000 #g) may indicate larger populations of responsive acceptors for the more effective odorants. Thus 1-hexanol stimulated more acceptors in males at the 100/~g dose, and more acceptors in both sexes at the 1000 #g dose. However, while l-hexanol enhanced weevil capture in pheromone-baited traps when in competition with grandlure alone, 1-hexanol + grandlure was ineffective when in competition with trans-2-hexen-l-ol + grandlure (Table 3). While cis-2-hexen-l-ol elicited EAGs equal to or greater than trans-2-hexen-l-ol at 100 and 1000 #g doses for males, trans-2-hexen-l-ol

effectively enhanced weevil capture in phero- mone-baited traps, but cis-2-hexen-l-ol was ineffective. Single cell studies revealed that trans- 2-hexenal also stimulated green odor receptors (Table 1), but did not enhance capture of weevils in traps (Table 3). Thus activation of the green odor receptor, at least in this context, does not necessarily result in an observed response based on the behavioral test used. This fact is indicative of 'across-fiber' coding for this class of compounds as previously mentioned herein, and suggested heretofore for the Colorado beetle (Ma & Visser, 1978).
Behavior
The enhancement of an insect pheromone response by green leaf volatiles is reported here for the first time. Results of competitive field tests with released boll weevils outside of cotton fields, and natural populations of weevils within cotton fields clearly demonstrate that certain green leaf volatiles enhance the boll weevil pheromone response (Tables 2-5). The effectiveness of the alcohols, trans-2-hexen-l-ol, cis-3-hexen-l-ol, and 1-hexanol correlates well with previous chemical studies demonstrating the presence of these compounds in essential oil of the cotton plant (Hedin et al., 1973).
The fact that individual green odor components were effective in enhancing response to phero- mone is not surprising, since two previous reports of volatiles collected from growing cotton each mention only one six carbon alcohol (Thompson etaL, 1971a; Hedin etal. , 1975). In a detailed study of growing cotton, Thompson and his coworkers (1971 a) found that as the cotton plant matured trans-2-hexenol, which was absent in volatile collections in early morning, was released in concentrations of 7 to 27 ~o during the after- noon. In another study, 1-hexanol was reported as a component of the air space above growing cot- ton (Hedin et al., 1975).
The data presented here also indicate a hier- archy in behavioral response to individual green leaf odors when paired with the pheromone (Table 3). Thus, while both 1-hexanol and trans-
201
2-hexen-l-ol enhanced capture of weevils by grandlure when in competition with grandlure alone, trans-2-hexen-l-ol + grandlure attracted more weevils when placed in competition with 1-hexanol + grandlure.
The activity of the green leaf volatiles in enhancing captures and extending the activity of grandlure-baited traps could have practical sig- nificance. Presently more than $900 000 are spent on the aggregation pheromone alone for use in survey traps for the BWEP in cotton growing regions in the United States. Each pheromone trap is baited with 10 mg of grandlure which is replaced biweekly. Green leaf volatiles, e.g. trans- 2-hexen-l-ol and 1-hexanol, are considerably less expensive than components of the boll weevil aggregation pheromone. The results presented here, and tests in Arizona and Texas (Dickens etal. , unpublished) show that activity of traps baited with as little as 1 mg of grandlure is not only enhanced, but also may be extended for as long as two weeks with the addition of trans- 2-hexen-l-ol (Table 5). Furthermore, recent results show that green leaf volatiles may enhance capture of boll weevils in pheromone traps throughout the entire growing season (Dickens et al., unpublished). If the quantity of pheromone used in each trap could be decreased with the addition of green leaf volatiles, then thousands of dollars currently spent on pheromone could be saved annually.
As an additional note, current experiments already underway with other insects, including the Mediterranean fruit fly, Ceratitis capitata Wied., and the bark beetle, Scolytus multistriatus, indicate that enhancement of pheromone activity by green leaf volatiles may be widespread among insects (Dickens et al., in preparation).
Acknowledgements
The author is grateful for technical assistance pro- vided by D. Wu, E. Mattix, W. Harris and S. Matthews. Critical reviews of the manuscript were kindly provided by Prof. Dr. T.L. Payne, Virginia Polytechnic Institute and State Univer-

202
sity, Blacksburg, VA; Dr. W.L. Parrot, USDA-ARS, Crop Science Research Labora- tory, Mississippi State, MS; and Dr. D.M. Light, USDA-ARS Western Regional Research Center, Albany, CA.
R~sum~
Augmentation de l'efficacit~ de la ph&omone agrd- gative de Anthonomus grandis sous l'influence de l 'odeur verte
Pour la premi6re fois, l 'augmentation de la puis- sance de la ph6romone d'agr6gation sous l'action de l'odeur verte est signal6e chez A. grandis. Les enregistrements unitaires de cellules des r6cep- teurs olfactifs des antennes a mis en 6vidence une cat6gorie de cellules r6agissant s~lectivement aux alcools et aux ald6hydes/t 6 carbones (c'est-/t-dire aux substances volatiles des feuilles vertes). Des essais dans la nature ont montr6 une augmenta- tion des captures par les pi6ges de grandlure (ph6romone d'agr6gation du charan~on) ren- forc6s par du trans-2-hexen-l-ol, du cis-3-hexen- 1-ol, ou du l-hexanol.
Bien que les courbes des r6ponses en fonction des doses construites/t partir des 61ectroantenno- grammes indiquent des cat6gories de cellules r6ceptrices identiques pour les alcools ~t 6 car- bones examin6s, l'une de ces substances, cis- 2-hexen-l-ol, s'est r6v616e inactive dans la nature. Le trans-2-hexenal qui 6tait actif avec des enregistrements unitaires de cellules, 6tait lui aussi inactif dans la nature. Dans des essais dans des champs de coton avec des populations indig6nes de charangons, le trans-2-hexen-l-ol a, non seule- ment augment6 l'efficacit6 des pi6ges ~t ph6ro- mone, mais a prolong6 leur attractivit6. Les r6sul- tats d'61ectrophysiologie combin6s aux donn6es de la nature appuient le modelage transversal des substances vertes volatiles des feuilles par les r6cepteurs olfactifs du charan~on. Ces r6sultats sont interpr6t6s en relation avec la chimie de la plante-h6te de A. grandis, le coton, Gossypium hirsutum L., et leur possibilit6 d'utilisation 6cono- mique pour l'6radication du charan~on.
References
Boeckh, J., 1967. Reaktionsschwelle, Arbeitsbereich und Spezifitat eines Geruchsrezeptors auf der Heuschrecken- antenne. Z. Vergl. Physiol. 55: 378-406.
Borden, J.H., 1984. Semiochemical-mediated aggregation and dispersion in Coleoptera, pp. 123-149. In, T. Lewis (ed.) Insect Communication. Academic Press, NY.
Chang, J. F., J. H. Benedict, T. L. Payne, B. J. Camp, 1986. Methods for collection and identification of volatile ter- penes from cotton and evaluation of their attractiveness to boll weevils. Southwest. Entomol. 11: 233-241.
Cross, W. H., M. J. Lukefahr, P. A. Fryxell & H. R. Burke, 1975. Host plants of the boll weevil. Environ. Entomol. 5: 565-571.
Den Otter, C. J., M. Behan & F. W. Maes, 1980. Single cell responses in female Pieris brassicae (Lepidoptera: Pieridae) to plant volatiles and conspecific egg odours. J. Insect PhysioL 26: 465-472.
Dickens, J.C., 1979. Electrophysiological investigations of olfaction in bark beetles. Mitt. Schweiz. EntomoL Ges. 52: 203-216.
Dickens, J.C., 1981. Behavioural and electrophysiological responses of the bark beetle, Ips typographus, to potential pheromone components. Physiol. Entomol. 6: 251-261.
Dickens, J. C., 1984. Olfaction in the boll weevil, Anthonomus grandis Boh. (Coleoptera: Curculionidae): Electroantenno- gram studies. J. Chem. Ecol. 10: 1759-1785.
Dickens, J. C., 1988. Neural detection and behavioral role of green leaf volatiles in the boll weevil, Anthonomus grandis Boh. (Coleoptera: Curculionidae). Proc. XVIII Internat. Congr. Entomol. Abstr.: 213.
Dickens, J.C. & P.E. Boldt, 1985. Electroantennogram responses of Trirhabda bacharides (Weber) (Coleoptera: Chrysomelidae) to plant volatiles. J. Chem. Ecol. 11: 767-779.
Dickens, J. C. & E. E. Moorman, 1987. Role of host plant and sensory environment in maturation of olfactory recep- tor system of the boll weevil, Anthonomus grandis. Neuro- science Abstr. 13: 82.
Dickens, J.C. & K. Mori, 1989. Receptor chirality and behavioral specificity of the boll weevil, Anthonomus grandis Boh. (Coleoptera: Curculionidae) for its phero- mone, (+)-grandisol. J. Chem. EcoL 15: 517-528.
Dickens, J. C. & T. L. Payne, 1977. Bark beetle olfaction: Pheromone receptor system in Dendroctonus frontalis. J. Insect Physiol. 23: 481-489.
Dickens, J. C. & T.L. Payne, 1985. Chemical messengers and insect behavior, pp. 201-230. In, N. B. Mandava (ed.) Handbook of Natural Pesticides: Methods, Volume 1: Theory, Practice and Detection. CRC Press, Boca Raton.
Dickens, J. C. & G. D. Prestwich, 1989. Differential recog- nition of geometric isomers by the boll weevil, Anthonomus grandis Boh. (Coleoptera: Curculionidae): Evidence for only three essential components in the aggregation phero- mone. J. Chem. Ecol. 15: 529-540.

Duncan, D.B., 1955. Multiple range and multiple F tests. Biometrics 11 : 1-42.
Guerin, P. M., E. Stadler & H. R. Buser, 1983. Identification of host plant attractants for the carrot fly, Psilae rosae. J. Chem. Ecol. 9: 843-861.
Hamilton, J. G. C., A. J. E. Gough, B. W. Staddon & D. E. Games, 1985. Multichemical defense of plant bug Hotea gambiae (Westwood) (Heteroptera: Scutelleridae): (E)-2- Hexenol from abdominal gland in adults. J. Chem. Ecol. 11 : 1399-1409.
Hardee, D.D., G.H. McKibben, D.R. Rummel, P.M. Huddleston & J.R. Coppedge, 1974. Response of boll weevils to component ratios and doses of the pheromone, grandlure. Environ. Entomol. 3: 135-138.
Hedin, P. A., A. C. Thompson & R. C. Gueldner, 1973. The boll weevil-cotton plant complex. Toxicol. Environ. Chem. Rev. 1: 291-351.
Hedin, P.A., A.C. Thompson & R.C. Gueldner, 1975. Survey of air space volatiles of the cotton plant. Phyto- chem. 14: 2088-2090.
Kaib, M., 1974. Die Fleisch- und Blumensrezeptoren aufden Antenne der Schmeissfliege Calliphora vicina. J. Comp. Physiol. 95: 105-121.
Katsoyannos, B. I. & P. M. Guerin, 1984. Hexanol: A potent attractant for the black fig fly, Silba adipata. Entomol. Exp. Appl. 35: 71-74.
Kozlowski, M. W. & J. H. Visser, 1981. Host plant-related properties of the antennal olfactory system in the oak flea weevil, Rhychaenus quercus. Electroantennogram study. Entomol. Exp. AppL 30: 169-175.
Ma, W.-C. & J. H. Visser, 1978. Single unit analysis ofodour quality coding by the olfactory antennal receptor system of the Colorado beetle. Entomol. Exp. Appl. 24: 320-333.
McKibben, G. H., W. L. Johnson, R. Edwards, E. Kotter, J. F. Kearny, T. B. Davich, E. P. LLoyd & M. C. Ganyard, 1980. A polyester-wrapped cigarette filter for dispensing grandlure. J. Econ. Entomol. 73: 250-251.
Mitchell, E. B., D. D. Hardee & T. B. Davich, 1976. In-field boll weevil trap. U. S. Patent No. 3,949,515.
203
Ostle, B., 1963. Statistics in Research. The Iowa State Univer- sity Press, Ames, Iowa. xv + 585 pp.
Pantin, C. F. A., 1948. Notes on Microscopical Techniques for Zoologists. Cambridge University Press, Cambridge.
Pfaffmann, C., 1941. Gustatory afferent impulses. J. Cell Comp. Physiol. 17: 243-258.
Sass, H., 1978. Olfactory receptors on the antenna of Periplaneta: Response constellations that encode food odors. J. Comp. Physiol. 128: 227-233.
Thompson, A.C., B.J. Wright, D.D. Hardee, R.C. Gueldner & P. A. Hedin, 1970. Constituents of the cotton bud XVI. The attractancy response of the boll weevil to the essential oils of a group of host and nonhost plants. J. Econ. Entomol. 63: 751-753.
Thompson, A.C., D.N. Baker, R.C. Gueldner & P.A. Hedin, 1971a. Identification and quantitative analysis of the volatile substances emitted by maturing cotton in the field. Plant Physiol. 48: 50-52.
Thompson, A.C., B.W. Hanny, P.A. Hedin & R.C. Gueldner 1971b. Phytochemical studies in the family Malvaceae. I. Comparison of essential oils of six species by gas-liquid chromatography. Amer. J. Bot. 58: 803-807.
Tumlinson, J.H., D.D. Hardee, R.C. Gueldner, A.C. Thompson, P. A. Hedin & J. P. Minyard, 1969. Sex phero- mones produced by male boll weevils: Isolation, identifi- cation, and synthesis. Science 166: 1010-1012.
Visser, J.H., 1979. Electroantennogram responses of the Colorado beetle, Leptinotarsa decemlineata, to plant vola- tiles. Entomol. Exp. Appl. 25: 86-97.
Visser, J. H., 1986. Host odor perception in phytophagous insects. Ann. Rev Entomol. 31: 121-144.
Visser, J. H. & D. A. Ave, 1978. General green leaf volatiles in the olfactory orientation of the Colorado beetle, Leptinotarsa decemlineata. Entomol. Exp. Appl. 24: 538-549.
Visser, J. H., S. Van Straten & H. Maarse, 1979. Isolation and identification of volatiles in the foliage of potato, Solanum tubersum, a host plant of the Colorado beetle, Leptinotarsa decemlineata. J. Chem. Ecol. 5: 13-25.
























![[ pheromone ] 01](https://img.pdfslide.net/doc/110x75/568caab71a28ab186da2ad9b/-pheromone-01.jpg)



