Embed Size (px)
Citation preview

J. Cell Sci. 5a, 71-84 (1981) 71Printed in Great Britain © Company of Biologists Limited 1981
GROWTH AND DIFFERENTIATION OF THE
GOLGI APPARATUS AND WALL FORMATION
DURING CARPOSPOROGENESIS IN THE
RED ALGA, GIGARTINA TEEDII
(ROTH) LAMOUR.
IOANNES TSEKOS*Zellenlehre, University of Heidelberg, Federal Republic of Germanyand Botanical Institute, University of Thessaloniki, Greece
SUMMARY
The fine structure of the Golgi apparatus during development of carposporangia of Gigartinateedii has been studied. Dictyosomes in differentiating carposporangia undergo extraordinarychanges in size and morphology.
Dictyosomes of gonimoblast cells and the auxiliary cell are composed of a stack of 5-7straight cisternae and are approximately o-i8/tm thick; from the structural and functionalviewpoint, these are similar to dictyosomes in most other plants.
Hypertrophied Golgi cisternae with fibrillar contents occur in immature and nearly maturecarposporangia. Secretory vesicles originate from the ends of cisternae and by completevesiculation of terminal cisternae; vesicles with a diameter of O-2-O-6 fitn and fibrillar contentsare abundant throughout the cytoplasm of immature and nearly mature carposporangia. Somevesicles secrete their contents into the carpospore wall. At this stage the dictyosomes arestraight in profile and only some Golgi cisternae and Golgi vesicles show acid phosphataseactivity.
Mucilage sacs appearing to be derived from either dilating endoplasmic reticulum or mem-braneous formations with a lysosomal character exist in both the mature gonimoblast cells andthe immature carposporangia. These mucilage sacs constitute a considerable portion of thecytoplasmic volume and ultimately migrate to the outside of the cytoplasm. The mucilage sacsdisappear completely in the mature carposporangia.
Throughout the development of the wall of immature and nearly mature carposporangia,multi-vesicular lomasome-like bodies, showing acid phosphatase activity, are released throughthe plasmalemma. Apart from any other possible activities, these bodies seem to contribute tothe plasticity of the expanding cell wall.
The older carposporangia are recognized by having very active dictyosomes with a semi-circular profile, producing vesicles with an electron-opaque centre (cored vesicles). Thesevesicles are likely to contain glycoproteins and to contribute to the formation of vacuoles withfibrous contents, as they are dominant in the carpospores before their liberation.
The dictyosomes of older carposporangia are composed of a polarized stack usually com-posed of 7 to 11 cisternae. They exhibit anomalous, densely staining, laminated mid-regions,which are formed due to close apposition of 6—7 adjacent cisternae from the forming face.The mid-regions of the rest of the cisternae towards the maturing face have large intercisternalspaces, which usually contain darkly staining small vesicles probably originating from theendoplasmic reticulum. The cisternae towards the maturing face contain dense granules.
Observations suggest that materials polymerize into fibrils within the Golgi system and theinflated Golgi-derived vesicles participate directly in the formation of the fibrillar network ofthe carpospore wall in the Gigartina teedii.
• Present address: Botanical Institute, University of Thessaloniki, Thessaloniki, Greece.

72 /. Tsekos
INTRODUCTION
The role of the Golgi apparatus in the formation and concentration of plant cell-wall materials is known and well established (Mollenhauer, Morre & Kelley, 1966;Miihlethaler, 1967; Northcote & Pickett-Heaps, 1966; Pickett-Heaps, 1967; Beams& Kessel, 1968; Whaley, 1975). Materials synthesized in a regulated sequence aretransported within Golgi-derived vesicles to the cell wall.
In different categories of plant cells certain components of the cell wall are releasedthrough exocytosis (Miihlethaler, 1967). Golgi vesicles have the same stainingcharacteristics as the ground substance (matrix) of the cell plate (Miihlethaler, 1967).The polysaccharides synthesized in the dictyosomes and transported in Golgi vesiclesto the cell wall must be pectic substances (Northcote & Pickett-Heaps, 1966; Miihle-thaler, 1967) or hemicelluloses (Northcote & Wooding, 1966; Wooding, 1968),forming the matrix of the cell wall.
During the carposporogenesis of red algae, parallel to the other changes that theimmature carpospores undergo after having been cut off from the gonimoblast, theyalso comprise large amounts of cell wall material (Chamberlain & Evans, 1973;Kugrens & West, 1973; Konrad Hawkins, 1974a, b).
Carpospore and tetraspore formation in red algae are nearly identical (Fritsch,1945). In addition, most of the ultrastructural features accompanying sporogenesisare remarkably similar to those of the red algae studied so far, which are mostlymembers of the Ceramiales (Peyriere, 1970; Tripodi, 1971, 1974; Kugrens & West,1972, 1973, 1974; Chamberlain & Evans, 1973; Scott & Dixon, 1973; Peel et al. 1973;Konrad Hawkins, 1974a, b; Wetherbee & West, 1977; Alley & Scott, 1977; Pueschel,1979; Gori, 1980). As a result of this taxonomic bias, the extent of variation incarposporogenesis and in the role the organelles play in the composition of the cellwall among the orders of red algae remains unknown.
Several papers on algae (Manton, 1967; Brown et al. 1970; Konrad Hawkins,1974 a, b) indicate a role for the Golgi apparatus in the formation of the frameworkas well as the matrix material of algal cell walls.
Despite the observations made on red algae concerning the release of membrane-bound droplets into the wall on the one hand, and the role of Golgi vesicles in wallformation on the other hand (Bouck, 1962; Bisalputra, Rusanowski & Walker, 1967;Ramus, 1972; Rawlence & Taylor, 1972; Kugrens & West, 1973; Konrad Hawkins,1974a, b), very little is known about the fine-structural mechanisms involved.
Generally the cell walls of the red algae consist of 2 components, a fibrillar oneand a mucilaginous one (Mackie & Preston, 1974). The latter is usually regarded asforming a non- or para-crystalline matrix in which the former is embedded as sets ofmicrofibrils (Hanic & Craigie, 1969). Polysaccharides in which some of the mono-saccharide units carry sulphate half-ester groups appear to have been found in thegenus Gigartina (Parker & Diboll, 1966; Mackie & Preston, 1974).

Carposporogenesis in Gigartina teedii 73
MATERIALS AND METHODS
Cystocarpic plants of Gigartina teedii were collected at Micron Emvolon (Gulf of Thessa-loniki) and were immediately fixed ' in situ' or were carried to the laboratory where they werefixed within 1 h of collection.
Material for light microscopy was fixed in 10 % acrolein in tap water at pH 7-0, for 24-48 hon ice. Dehydration and embedding in glycol methacrylate were carried out as described byFeder & O'Brien (1968) and Evans & Holligan (1972). Sections of 0-5-1-5 /(m were cut on aReichert OmU-J ultramicrotome using glass knives.
Material for electron microscopy was treated according to the method published by Galatis,Katsaros & Mitrakos (1977). Semi-thin sections were cut and stained with 1 % toluidine blue Oin 1 % borax solution for light microscopy also. Ultra-thin sections were cut and collected oncopper grids and stained with uranyl acetate and lead citrate (Reynolds, 1963). These sectionswere examined and photographed either on a Philips EM400 or on a Jeol 100-B electronmicroscope.
The following cytochemical investigations were carried out. Specimens were treated forspecific staining of neutral polysaccharides with periodic acid-Schiff (PAS) (Jensen, 1962;McCully, 1966) or with periodic acid-silver methenamine (Rambourg, 1967) and of proteinswith fast green F.C.F. (Ruthman, 1966), with mercury-bromophenol blue (Ruthman, 1966)or with acid fuchsin (Robinow & Marak, 1966).
For detection of acid phosphatase, specimens of reproductive thalli were cut into smallpieces in fixative solution (5 % glutaraldehyde in o-i M-sodium cacodylate buffer, 0-25 M-sucrose). Fixation in glutaraldehyde was carried out for 5 h at room temperature. Fixationwas followed by a series of washes in cold o-i M-cacodylate buffer plus sucrose. The original0-25 M-sucrose was reduced by 50 % in each successive wash.
The thalli, fixed in glutaraldehyde and washed in sodium cacodylate buffer were againplaced for washing for 2 h in o-i M-sodium acetate buffer (pH 5-5) in the refrigerator (abouto °C). Incubation followed at 37 °C for 1 h in Gomori medium (Gomori, 1952) modified tocontain half the original concentration of PbNO,, and in which the pH was adjusted to 5-5instead of 5-0. The substrate employed was /?-glycerophosphate. Following incubation speci-mens were rinsed for 1-5 h in 3 changes of 01 M-sodium acetate (pH 5-5) at room temperature.Post-fixation was done at o °C in 1 % OsO4 cacodylate buffer. The steps to be taken after-wards follow the same procedure as in the usual fixation until the uptake of blocks (see Galatiset al. 1977). Sections were observed either unstained, or following staining with 4% aqueousuranyl acetate for 5 min. Control specimens were treated identically to reactive specimens,except that the Gomori medium contained o-oi M-NaF as inhibitor.
RESULTS AND DISCUSSION
Auxiliary cell and gonimoblast cells
The cystocarpic plants of Gigartina have a thallus 10-20 cm high and compressedflatly; the branching is irregularly pinnate. The cystocarps are almost spherical(0-7-0-8 mm diameter) and are set sideways on the small branches.
According to Sjostedt (1926), in the red alga, Gigartina teedii, the fertilized carpo-gonium fuses with the auxiliary cell from which the gonimoblast cells form bycytokinesis. The auxiliary cell is irregular in shape and multinucleate. The nucleihave a typical morphology with a distinct nucleolus in the centre and the membraneis interrupted by numerous well-sized pores of complex structure (see also Kugrens& West, 1973). The auxiliary cell has numerous starch grains and plastids (in theform of proplastids).
The morphology of the Golgi apparatus, as well as the type and quantity of Golgi-derived vesicles, changes considerably during carpospore differentiation. One con-
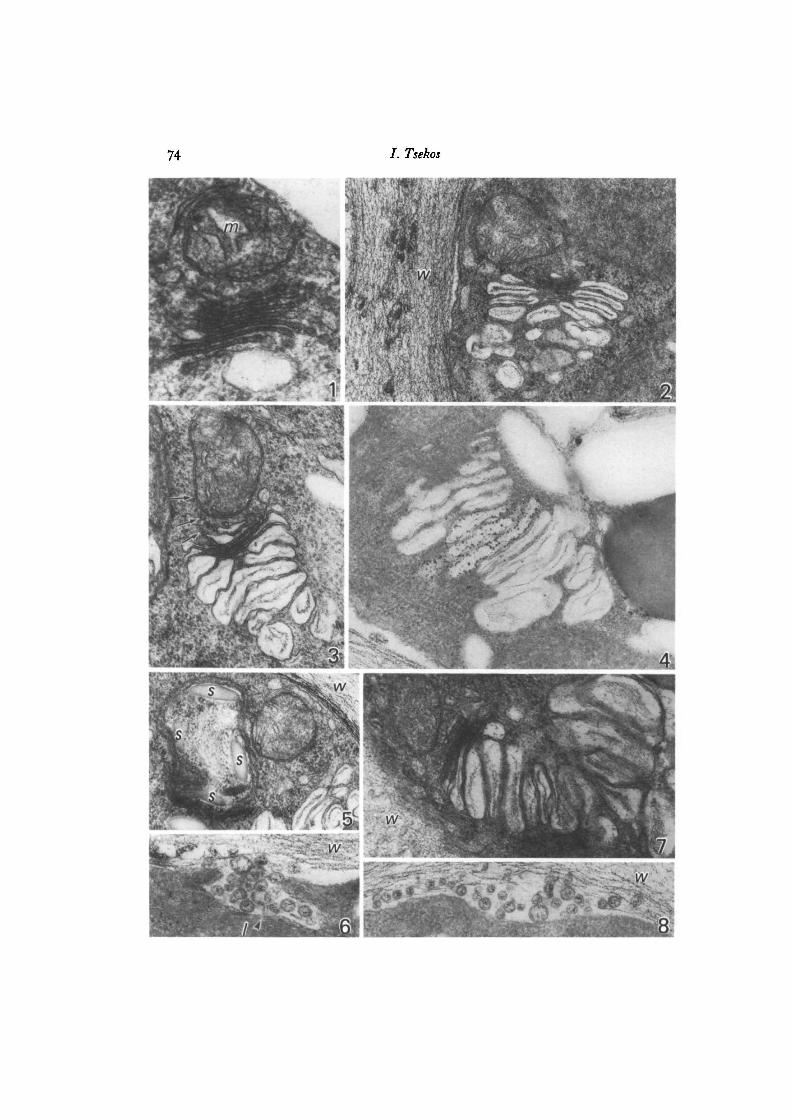

Carposporogenesis in Gigartina teedii 75
stant feature that has been frequently reported in the red algae, is the presence of amitochondrion at the base of the Golgi forming face (Figs, i, 2, 3, 7, 14) (KonradHawkins, 1974a, b; Kugrens & West, 1972, 1973; Wetherbee & West, 1977; Alley &Scott, 1977).
In the auxiliary cell and the young gonimoblast cells the number of dictyosomes issmall and the Golgi-mitochondrion complex is surrounded by elements of the endo-plasmic reticulum (ER). In those 2 cell categories, dictyosomes are approximately0-23-0-60 /im long, 0-15-0-21/tm wide and usually consist of 5-7 flat, straightcisternae (Fig. 1). The morphology, size and manner of the formation of secretoryvesicles and dictyosomes at this stage are quite comparable to those found in activelygrowing vegetative cells of G. teedii and in most other algae, fungi and higher plants(see also Alley & Scott, 1977). Fig. 1 shows a single dictyosome at high magnification.Note the degree to which the cisternae are separated by a consistent intercisternalspace.
Immature sporangia
The mature gonimoblast is accordingly surrounded on the innermost side bymedullary pericarp, which at a later stage of carposporogenesis becomes more andmore impoverished, and outside that by the thick cortical tissue, which as usual formsthe true pericarp (cortical pericarp) (Sjostedt, 1926).
The plasmalemma profile of gonimoblast cells appears highly irregular. When agonimoblast cell begins to differentiate into a carpospore, the cytoplasmic constituentsare similar to gonimoblast cells, but the plasmalemma profile becomes more regular.Pit plugs between young carpospores and gonimoblast cells are also present initially,
Figs. 1-23. fv, fibrous vacuole; /, lomasome-like body; ly, lysosome; m, mitochon-drion; ms, mucilage sac; s, starch grain; iv, cell wall.Fig. 1. Typical form of dictyosome and its associated mitochondrion in an auxiliarycell, x 54000.Fig. 2. Part of an immature carpospore with hypertrophied dictyosome producingvesicles with fibrillar contents, x 40000.Fig. 3. Dictyosome with inflated cisternae of an immature carpospore, possiblyER-derived small vesicles (arrows) at the forming face, x 46000.Fig. 4. Dictyosome of an immature carpospore showing acid phosphatase activity onlyin certain cisternae. x 60000.Fig. 5. Part of an immature carpospore with a formation, which seems to be thefunctional equivalent of a lysosome, inside which starch grains are found, x 27000.Fig. 6. Part of an immature carpospore with a lomasome-like body (/) releasing itscontents into the wall and showing acid phosphatase activity, x 77000.Fig. 7. Hypertrophied dictyosome producing considerably inflated cisternae of anearly mature carpospore. Note that the fibrous material within Golgi vesicles issimilar to that of the cell wall, x 30000.Fig. 8. Part of an immature carpospore with a lomasome-like body which has releasedits contents into the cell wall showing acid phosphatase activity, x 60000.
/. Tsekos
14

Carposporogenesis in Gigartina teedii 77
but these pit plugs presumably degenerate and disappear in later development (seealso Kugrens & West, 1973).
A young spore during the first stage in carpospore differentiation is shown inFig. 9 (compare with an older spore in Fig. 13). Chloroplasts increase in size andnumber, and develop their membrane system. Mitochondria and ER become moreabundant. At this time, the formation of starch grains commences and they pro-gressively increase in size and number, and are infested with mature carpospores.
An increase in the size of the Golgi apparatus occurs in immature carposporangia.Most apparent increases are recorded in the length (0-61-0-75 /<m)> m t n e number(6-8) of cisternae per stack and presumably in the number of dictyosomes (Figs. 2, 3).This increase of the Golgi apparatus is connected with an increased secretory activityproducing considerably inflated cisternae beneath the plasma membrane, withfibrous material that is similar to that of the cell wall (Fig. 2). Small vesicles (30 nmdiameter), appear at the forming face (Figs. 2, 3). Dilation, removal and variousstages of incomplete as well as complete vesiculation of mature cisternae are observedat the distal poles of dictyosomes (Figs. 2, 3, 5).
Dilation of cisternae and expansion of vesicles were the first changes seen in theGolgi apparatus of immature carpospores. These alterations were accompanied bythe thickening of the cell wall of immature carpospores and the presence of denseER-associated bodies, 0-40 /<m wide. Similar bodies have been observed in Griffithsiaflosculosa (Peyriere, 1970), Bryopsis hypnoides (Burr & West, 1970), and in Calli-thamnion roseum (Konrad Hawkins, 19746).
The young sporangial wall of G. teedii, as Chamberlain & Evans (1973) describedfor Ceramium rubrum, consists of a closely packed reticulum of microfibrils (Figs. 2, 5).The microfibrils are orientated parallel to the plasmalemma. In certain positions onthe cell surface the microfibrils are found to be in close contact with the plasmalemmaand appear more electron-dense (Fig. 5). All of the carposporangia are surrounded bya matrix of amorphous material containing fibrils (Figs. 9, 10).
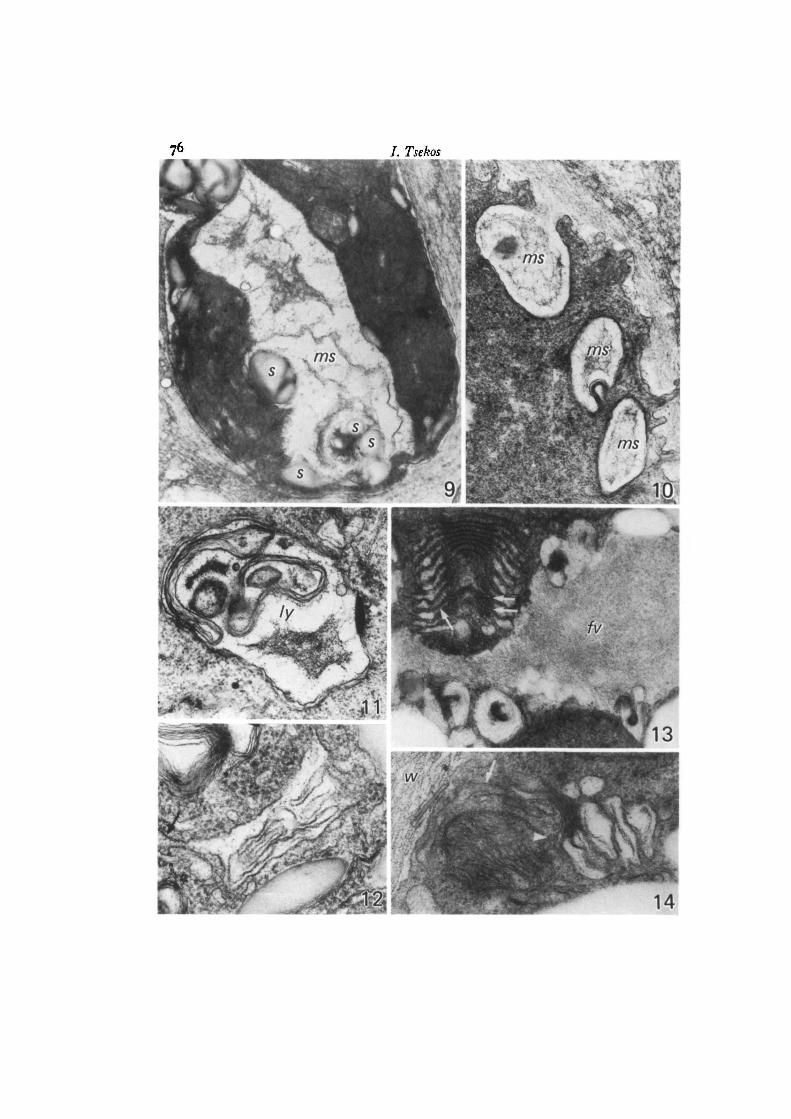
Fig. 9. Part of an immature carpospore with a central mucilage sac inside which anumber of starch grains are found, x 16000.Fig. 10. Mucilage sacs of an immature carpospore. One of them is discharging its con-tents into the wall, x 21000.Fig. 11. Lysosome of an immature carpospore inside which mucilage is found,x 31000.
Fig. 12. Probable commencement of formation of a mucilage sac from dilating ER ofan immature carpospore. Note cisterna attached to the initial mucilage sac (arrow),x 60000.
Fig. 13. Dictyosome of an older carpospore prior to spore liberation demonstratingdilated peripheral regions and densely staining laminated mid-regions. Several coredvesicles and a portion of a large vacuole (Jv) with fibrous contents are visible. Notethat the cisternae towards the maturing face contain darkly staining granules (arrows),x 46000.
Fig. 14. Part of a nearly mature carpospore with hypertrophied dictyosome. At theforming face there is association with membranes of the endoplasmic reticulum(arrow) and possibly ER-derived small vesicles (arrowhead), x 60 000.

/ . Tsekos

Carposporogenesu in Gigartina teedii 79
The initial gonimoblast wall will eventually become the carposporangial wall.Carpospore wall formation is usually initiated during the early stage of carposporedifferentiation and ends with the commencement of the production of cored vesicles.
The mature gonimoblast cells and the carpospores at the first stages of theirdifferentiation exhibit formations, i.e. the mucilage sacs (Figs. 9—11); the latter con-stitute a considerable portion of the cytoplasmic volume (Fig. 9) and ultimatelymigrate to the outside of the cytoplasm (Fig. 10).
It is probable that there are 2 ways for the formation of the mucilage sacs, asfollows: (1) by membrane formations, the structure of which can be seen in Fig. 5,surrounding protoplasmic material (such as the ground plasma, mitochondria,ribosomes, starch grains, etc.). These formations seem to be the functional equivalentsof lysosomes (Figs. 5, 9, 11) involved in the intracellular breakdown of starch and inthe synthesis of a mucilaginous material prior to the secretion of their content intothe wall (Fig. 10). (2) The mucilage sacs seem to be derived from dilating ER inFig. 12.
The mucilage sacs disappear gradually with the differentiation of carpospores andhave completely disappeared in the mature condition. The mucilage sacs of thegonimoblast cells and the immature carposporangia show morphological similarityto the mucilage sacs occurring during tetrasporogenesis in the red alga Palmariapalmata (Pueschel, 1979).
Fig. 15. Dictyosome of an older carpospore prior to spore liberation demonstratingdilated peripheral regions and densely staining, laminated mid-regions. Note that thecisternae towards the maturing face contain darkly staining granules (arrows),x 53000.Fig. 16. Part of a nearly mature carpospore with hypertrophied dictyosomes. Arrowsindicate fibrous vesicles discharging their contents into the wall, x 24000.Fig. 17. Part of a nearly mature carpospore with a lomasome-like body (/), which hasreleased its contents into the cell wall. Hypertrophied Golgi cisternae and vesicles(arrows) continuous with starch grains (*).Fig. 18. Part of a nearly mature carpospore with hypertrophied dictyosome. Arrowindicates Golgi vesicle discharging its contents into the wall, x 27000.Fig. 19. Dictyosome of an older carpospore prior to spore liberation. Note the forma-tion of the cored vesicles by the curling of one inflated Golgi cisterna around somecytoplasmic material, x 50000.Fig. 20. Part of an older carpospore prior to spore liberation with several coredvesicles. Periodic acid—silver methenamine. The starch grains and the central core ofthe Golgi vesicles react strongly with silver methenamine, depositing silver grains.Arrow indicates cored vesicle discharging its contents into the wall, x 12500.Fig. 21. Dictyosome of an older carpospore. The intercisternal spaces towards thematuring face contain darkly staining small vesicles (arrows). At the maturing facethe formation of a cored vesicle seems to be taking place (arrowhead), x 45000.Fig. 22. Part of an older carpospore prior to spore liberation. Arrow indicates fibrousvacuole discharging its content into the wall, x 21 000.Fig. 23. Dictyosome of an older carpospore prior to spore liberation. Note that asingle cored vesicle is formed from the direct transformation of a single Golgicisterna. x 62000.

80 I. Tsekos
It must also be emphasized that concentric bodies of double membranes arelocated at the wall of differentiating carpospores. It can also be seen that at somepositions the plasmalemma profile of immature carpospores appears irregular. Onemay also discern plasmalemmal extensions (plasmalemmavilli) (Cottier, 1971)throughout all the stages of differentiation of the carpospores and in the relativelymature ones.
Nearly mature and older sporangia
The nearly mature stage of the carpospores may be recognized by the followingcharacteristics. The mucilage sacs disappear from the cytoplasm. The dictyosomesat this time exhibit extreme hypertrophy and are also straight in profile (Figs. 7, 14,18) and in close contact with the ER (Fig. 14), producing vesicles with fibrous con-tents. These vesicles migrate to the outside of the cytoplasm and are responsible forthe bulk of the wall deposition comprising the carpospore wall (Figs. 16, 18), as inthe case of Polysiphonia sp. (Konrad Hawkins, 1974a). Fibrils of the wall in theimmature and nearly mature carposporangia are morphologically identical to thosewithin Golgi vesicles (in addition to variably sized fibrils) (Figs. 2, 7, 14, 16, 18).
The morphology of the Golgi apparatus and the Golgi vesicles in nearly maturecarpospores presents similarities to that of the immature carposporangia.
Starch grains are abundant and they are frequently in direct contact with Golgicisternae and vesicles (Fig. 17). Flow of membrane extending the new plasma mem-brane presumably increased with continuing hypertrophy of the Golgi apparatus andexpansion of carpospores in G. teedii. Hypertrophy is a particular trend of differen-tiating dictyosomes (Goodman & Porter, i960; Mollenhauer, Whaley & Leech, 1961;Whaley, Kephart & Mollenhauer, 1964; Flickinger, 1969). It is not known how thishypertrophied condition is brought about. Juniper & Roberts (1966) suggested thatthe hypertrophy of dictyosomes is linked with a rich supply of carbohydrates derivingfrom the breakdown of starch grains. From this point of view, the close relationshipbetween inflated Golgi cisternae and vesicles with swollen starch grains in G. teediiappears significant. This is supported by the fact that the dictyosomes of G. teediicontain acid phosphatase (Fig. 4).
Throughout the development of the wall of immature and nearly mature carpo-sporangia many multivesicular lomasome-like bodies (Fig. 17), identical to thoseshown by Chamberlain & Evans (1973) in the young tetrasporangium of Ceramiumrubrum, are released through the plasmalemma. These exhibit acid phosphataseactivity (Figs. 6, 8). Many functions have been ascribed to lomasomes (Bracker, 1967).In accord with Marchant & Robards (1968), an involvement with enzyme transportrelated to subsequent modifications of the developing cell wall must be attributed tothe multivesicular lomasome-like bodies of the carposporangia in G. teedii. Becauseof the acid phosphatase activity of the lomasome-like bodies, these must, apart fromany other possible activities, contribute to the plasticity of the cell wall. At this stage,activity of acid phosphatase was found in only some of the Golgi cisternae (Fig. 4),that is, not all cisternae necessarily contain the same enzymes (Novikoff, Essner,

Carposporogenesis in Gigartina teedii 81
Goldfischer & Heus, 1962; Dauwalder, Whaley & Kephart, 1969; Wise & Flickinger,1970).
The above observations support the view that materials polymerize into fibrilswithin the Golgi system and become discharged by Golgi-derived vesicles, which fusewith the plasma membrane.
The older carposporangia are recognized by having Golgi vesicles with electron-opaque centres (Figs. 13, 19, 20), called cored vesicles. The cytoplasm now containslarge quantities of starch and ER, and large fibrillar vacuoles (fv) (Figs. 13, 22),which are derived from the Golgi cored vesicles (Fig. 13) and appear to containcomplex polysaccharides (positive PAS-reaction). Starch grains are frequentlyassociated with fibrillar vacuoles (Fig. 13).
Using light (PAS-reaction) and electron microscopic (periodic acid-silver methen-amine; Rambourg, 1967) techniques it is shown that the mucilaginous component ofcell wall in G. teedii gives a weak positive reaction for polysaccharides, while thefibrillar one gives a negative reaction.
The central core of the cored vesicles yields a positive reaction in both protein(staining with fast green) and polysaccharides as well (PAS-reaction and silvermethenamine; Rambourg, 1967) (Fig. 20). It is likely that the cored vesicles containcompounds of a glycoprotein type (see also Chamberlain & Evans, 1973). Accordingto Peyriere (1970) the cored vesicles are mucopolysaccharides. The cored vesicles andthe fibrillar vacuoles, following exocytosis (Figs. 20, 22), may function in sporeprotection or adhesion subsequent to spore liberation.
In the cored vesicles and the cisternae of the semicircular dictyosomes of the oldercarposporangia, acid phosphatase has not been detected. In my opinion, the coredvesicles, of G. teedii at least, must not function in a similar way to lysosomes, asKonrad Hawkins (1974 a) also believes for the red alga Polysiphonia. Fig. 13 showsseveral cored vesicles and a portion of one of the many large fibrous vacuoles that arepresent in older carposporangia prior to spore liberation. At this stage the microfibrilsof the cell wall are loose and an electron-transparent zone is formed between theplasmalemma and microfibrils of the cell wall.
The dictyosomes are semicircular in profile. Usually 9-10 cisternae are observed,but occasionally 11 or more can be present (Figs. 13, 15, 19). The most strikingfeatures seen at this stage, however, are the changes in appearance of the middle andperipheral regions of each dictyosome. The peripheral portions are dilated while themid-regions of the first 6-7 cisternae from the forming face are compressed anddensely laminated. The mid-regions of the rest of the cisternae towards the maturingface have large intercisternal spaces, which usually contain darkly staining smallvesicles (30 nm diameter) (Fig. 21); these latter vesicles appear to be derived fromthe ER. The cisternae towards the maturing face contain darkly staining granules(Figs. 13, 15).
This unusual modification of the dictyosomes in G. teedii has been noticed insporangia of other species of red algae (Kugrens & West, 1972, 1973, 1974; Wetherbee& West, 1976, 1977; Alley & Scott, 1977).
It is not sufficiently clear how the cored vesicles develop. Some appear to originate

82 7. Tsekos
by inflation of distal cisternae (Figs. 21, 23) (see also Konrad Hawkins, 1974a).Cisternae at the maturing face of the dictyosome assume a spherical configuration.As a result, only a single, large secretory vesicle is formed from the direct transforma-tion of a single cisterna. The densely staining central core of material appears toresult from a progressive centripetal accumulation of granular intracisternal substanceas the cisternae advance from the forming to the maturing face of the dictyosome.Others may form by the curling of 1 or 2 inflated cisternae around some cytoplasmicmaterial (Figs. 15, 19). Fusion of distal ends of such cisternae would produce vesiclesthat have entrapped parts of the cytoplasm.
I wish to thank Professor Dr E. Schnepf for offering me the opportunity to work at hisInstitute and for critically reading the manuscript. I would also like to thank Miss G. Deich-graber for her valuable assistance and Miss A. Kiratzidou for assisting in the preparation ofthe manuscript in the English language. Special thanks are given to the Stiftung Volkswagen-werk for providing the necessary research media and to Alexander von Humbolt-Stiftung forhaving granted a scholarship.
REFERENCES
ALLEY, C. D. & SCOTT, J. L. (1977). Unusual dictyosome morphology and vesicle formation intetrasporangia of the marine red alga Polysiphonia denudata. J. Ultrastruct. Res. 58, 289-298.
BEAMS, H. W. & KESSEL, R. G. (1968). The Golgi apparatus: structure and function. Int. Rev.Cytol. 23, 209-276.
BISALPUTRA, T., RUSANOWSKI, P. C. & WALKER, W. S. (1967). Surface activity, cell wall andfine structure of pit connection in the red alga Laurencia spcctabilis. J. Ultrastruct. Res. 20,277-289.
BOUCK, G. B. (1962). Chromatophore development, pits, and other fine structure in the redalga Lomentaria baileyana (Han1.) Farlow. J. Cell Biol. 12, 553-569.
BRACKER, C. E. (1967). Ultrastructure of fungi. Rev. Phytopathol. 5, 343-374.BROWN, R. M., KLEINIG, H., FALK, H., SITTE, P. & FRANKE, W. (1970). Scale formation in
Chrysophycean algae. J. Cell Biol. 45, 246-271.BURR, F. A. & WEST, J. A. (1970). Light and electron microscope observations on the vegetative
and reproductive structures of Bryopsis hypnoides. Phycologia 9, 17-37.CHAMBERLAIN, A. H. L. & EVANS, L. V. (1973). Aspects of spore production in the red alga
Ceramium. Protoplasma 76, 139—159.COTTLER, M. H. (1971). Plasmalemmal extensions in Chondrus Crispin (L.) Stackh. J. Ultra-
struct. Res. 37, 31-36.DAUWALDER, M., WHALEY, W. G. & KEPHART, J. E. (1969). Phosphatases and differentiation
of the Golgi apparatus. J. Cell Set. 4, 455-497.EVANS, L. V. & HOLLIGAN, M. S. (1972). Correlated light and electron microscope studies on
brown algae. I. Localization of alginic acid and sulphated polysaccharides in Dictyota. NewPhytol. 71, 1161-1172.
FEDER, N. & O'BRIEN, T. P. (1968). Plant microtechnique: some principles and new methods.Am.J. Bot. 55, 123-142.
FRITSCH, F. E. (1945). Structure and reproduction of the algae, vol. 2. Cambridge UniversityPress.
FLICKINGER, C. J. (1969). The pattern of growth of the Golgi complex during the fetal andpostnatal development of the rat epididymis. J. Ultrastruct Res. 27, 344-360.
GALATIS, B., KATSAROS, CHR. & MITRAKOS, K. (1977). Fine structure of vegetative cells ofSphacelaria tribuloides Menegh. (Phaeophyceae, Sphacelariales) with special reference tosome unusual proliferations of the plasmalemma. Phycologia 16, 139-151.
GOMORI, G. (1952). Microscopic Histochemistry. Principles and Practice. University of ChicagoPress.

Carposporogenesis in Gigartina teedii 83
GOODMAN, G. C. & PORTER, K. R. (i960). Chondrogenesis, studied with the electron micro-scope. J. biophys. biochem. Cytol. 8, 719-760.
GORI, P. (1980). Ultrastructure of carpospores in Gastroclonium clavatitm (Roth) Ardissone(Rhodymeniales). Protoplasma 103, Z63-271.
HANIC, L. A. & CRAIGIE, J. S. (1969). Studies on the algal cuticle. J. Phycol. 5, 89-102.JENSEN, W. A. (1962). Botanical Histochemistry. San Francisco: Freeman.JUNIPER, B. E. & ROBERTS, R. M. (1966). Polysaccharide synthesis and the fine structure of
root cap cells. jfl R. microsc. Soc. 85, 63-72.KONRAD HAWKINS, E. (1974a). Golgi vesicles of uncommon morphology and wall formation
in the red alga, Polysiphonia. Protoplasma 8o, 1-14.KONRAD HAWKINS, E. (19746). Growth and differentiation of the Golgi apparatus in the red
alga, Callithamnion rosenm.J. Cell Sci. 14, 633-655.KUGRENS, P. & WEST, J. A. (1972). Ultrastructure of tetrasporogenesis in the parasitic red alga
Levringiella gardneri (Setchell) Kylin. J. Phycol. 8, 370-383.KUGRENS, P. & WEST, J. A. (1973). The ultrastructure of carpospore differentiation in the
parasitic red alga Levringiella gardneri (Setch.) Kylin. Phycologia 12, 163-173.KUGRENS, P. & WEST, J. A. (1974). The ultrastructure of carposporogenesis in the marine
hemiparasitic red alga EryUirocystis saccata.J. Phycol. 10, 139-147.MCCULLY, M. E. (1966). Histological studies on the genus Fucus. I. Light microscopy of
mature vegetative plant. Protoplasma 62, 287-305.MACKIE, W. & PRESTON, R. D. (1974). Cell wall and intercellular region polysaccharides. In
Algal physiology and biochemistry, vol. 10 (Botanical Monographs) (ed. W. D. P. Stewart).Oxford, London, Edinburgh, Melbourne: Blackwell Scientific Publications.
MANTON, I. (1967). Further observations on the fine structure of Chrysochromulina chiton withspecial reference to the haptonema, a 'peculiar' Golgi structure and scale productionJ. Cell Sci. 2, 265-272.
MARCHANT, R. & ROBARDS, A. W. (1968). Membrane systems associated with the plasmalemmaof plant cells. Ann. Bot. 32, 457-471.
MOLLENHAUER, H. H., WHALEY, W. G. & LEECH, J. H. (1961). A function of the Golgi appara-tus in outer rootcap cells. J . Ultrastruct Res. 5, 193-200.
MOLLENHAUER, H. H., MORRE, D. J. & KELLEY, A. G. (1966). The widespread occurrence ofplant cytosomes resembling animal microbodies. Protoplasma 62, 44-52.
MUHLETHALER, K. (1967). Ultrastructure and formation of plant cell walls. A. Rev. PI. Physiol.18, 1-24.
NORTHCOTE, D. H. & PICKETT-HEAPS, J. D. (1966). A function of the Golgi apparatus inpolysaccharide synthesis and transport in the root-cap cells of wheat. Biochem. J. 98, 159-167.
NORTHCOTE, D. H. & WOODING, F. B. P. (1966). Development of sieve tubes in Acer pseudo-platanus. Proc. R. Soc. B, 163, 524-537.
NOVIKOFF, A. B., ESSNER, E., GOLDFISCHER, S. & HEUS, M. (1962). Nucleoside phosphataseactivities of cytomembranes. In The Interpretation of Ultrastructure, vol. 1, Symp. Int. Soc.Cell. Biol. (ed. R. J. C. Harris), pp. 149-192. New York, London: Academic Press.
PARKER, B. C. & DIBOLL, A. G. (1966). AJcian stains for histochemical localization of acid andsulphated polysaccharides in algae. Phycologia 6, 37-46.
PEEL, M. C , LUCAS, I. A. N., DUCKETT, J. G. & GREENWOOD, A. D. (1973). Studies of sporo-genesis in the Rhodophyta. I. An association of the nuclei with endoplasmic reticulum inpostmeiotic tetraspore mother cells of Corallina offidnalis L. Z. Zellforsch. mikrosk. Anat.147. 59-74-
PEYRIERE, M. (1970). Evolution de l'appareil de Golgi au cours de la te'trasporogenese deGriffithsia flosculosa (Rhodophyc6e). C.r. Seanc. Acad. Set., Paris 270, 2071-2074.
PICKETT-HEAPS, J. D. (1967). The use radioautography for investigating wall secretion in plantcells. Protoplasma 64, 50-66.
PUESCHEL, C. M. (1979). Ultrastructure of tetrasporogenesis in Palmaria palmata (Rhodophyta).J. Phycol. 15, 409-424.
RAMBOURG, A. (1967). An improved silver methenamine technique for the detection of periodicacid-reactive complex carbohydrates with the electron microscope. J. Histochem. Cytochem.15, 409-412.
RAMUS, J. (1972). The production of extracellular polysaccharide by the unicellular red alg8Porphyridium aerugineum. J. Phyccl. 8, 97-111.

84 /• Tsekos
RAWLENCE, D. J. & TAYLOR, A. R. A. (1972). A light and electron microscopic study of rhizoiddevelopment in Polysiphonia lanosa (L.) Tanty. J. Phycol. 8, 15-24.
REYNOLDS, E. S. (1963). The use of lead citrate at high pH as an electron-opaque stain inelectron microscopy. J. Cell Biol. 17, 208-213.
ROBARDS, A. W. (1969). Particles associated with developing plant cell walls. Planta 88,376-379-
ROBINOW, C. F. & MARAK, J. (1966). A fibre apparatus in the nucleus of the yeast cell. J. CellBiol. 29, 131-151.
RUTHMANN, A. (1966). Methoden der Zellforschung. Franckh'sche Verlagshandlwtg. Stuttgart:W. Keller und Co.
SCOTT, J. L. & DIXON, P. S. (1973). Ultrastructure of tetrasporogenesis in the marine red algaPtilota hypnoides. J. Phycol. 9, 29-46.
SJOSTEDT, L. G. (1926). Floridean studies. Lunds Univ. Arsskrift, N. F. Avd. 2, Bd 22, Lund.TRIPODI, G. (1971). The fine structure of the cystocarp in the red alga Polysiphonia sertulacea
(Gmel.) Born, et Thur. J. Submicrosc. Cytol. 3, 71-79.TRIPODI, G. (1974). Ultrastructure changes during carpospore formation in the red alga
Polysiphonia. J. Submicrosc. Cytol. 6, 275-286.WETHERBEE, R. & WEST, J. A. (1976). Unique Golgi apparatus and vesicle formation in a red
alga. Nature, Lond. 259, 566-567.WETHERBEE, R. & WEST, J. A. (1977). Golgi apparatus of unique morphology during early
carposporogenesis in a red alga. J. Ultrastruct. Res. 58, 119-133.WHALEY, W. G. (1975). The Golgi apparatus. New York: Springer-Verlag.WHALEY, W. G., KEPHART, J. E. & MOLLENHAUER, H. H. (1964). The dynamics of cytoplasmic
membranes during development. In Cellular Membranes during Development (ed. M. Locke),PP- I35~I73- New York: Academic Press.
WISE, G. E. & FLICKINGER, C. J. (1970). Cytochemical staining of the Golgi apparatus inAmoeba proteus. J. Cell Biol. 46, 620-626.
WOODING, F. B. P. (1968). Radioautographic and chemical studies of incorporation intosycamore tissue walls. J. Cell Sci. 3, 71-80.
{Received 27 April 1981)





























