Embed Size (px)
Citation preview

gy 63 (2007) 1–18www.elsevier.com/locate/marmicro
Marine Micropaleontolo
Guilds among epiphytal foraminifera on fibroussubstrates, Nevis, West Indies
Brent Wilson ⁎
Petroleum Geoscience Programme, Department of Chemical Engineering, The University of the West Indies,St. Augustine, Trinidad and Tobago, West Indies
Received 20 July 2006; received in revised form 6 October 2006; accepted 9 October 2006
Abstract
The nearshore foraminifera of the oligotrophic Caribbean region are predominantly epiphytal, living on calcareous algae andseagrasses. However, their community ecology (interactions between and associations among species) is little understood. Clusteranalysis is used here to identify guilds among the epiphytal foraminifera, a guild being a group of species that forms an arena ofintense competition within which species' niches overlap considerably in habitat, time and food.
Samples were collected monthly between September 1994 and August 1995 from (a) calcareous algae and (b) seagrass rhizomesand leaf bosses from two bays around Nevis, northern Lesser Antilles. One bay was impacted by organic enrichment from a leakingseptic tank. Data from both bays were combined to form a metapopulation, analysis of which was conducted using common species(i.e., those that formed N1% of the recovery from any substrate in either bay). Q-mode cluster analysis using populations on allsubstrates of common species clearly separated the two bays, despite sampling of the same substrate in both, indicating that the baysoccupy different biofacies. However,Q-mode analysis did not separate the different substrates in the individual bays. R-mode clusteranalysis of the two-bay metapopulation based on common species revealed three clusters that are interpreted as guilds:
Guild I A higher-energy guild intolerant of nutrient enrichment.Guild II A guild of rapidly-reproducing opportunists on short-lived substrates.Guild III A lower-energy guild tolerant of nutrient enrichment.
These guilds are not limited to any one bay, but vary in proportion between the two, Guild I being most prolific in the virtuallypristine back-reef bay on the windward side of the island, and Guilds II and III forming a greater proportion of the epiphytalforaminiferal fauna in the bay impacted by lower-energy waves and a flux of organic matter. Guild II shows a preference for the bushycapitulae of the alga Penicillus capitatus. In cross-correlation of 12-month time-series for the three guilds, the virtually pristine bayshowed that they bloomed sequentially, at intervals of 4 months.
The recognition of guilds clears the way for improved understanding of the foraminiferal community ecology of nearshore areas.Nearshore sediments in the Caribbean yield primarily foraminifera washed from phytal substrates. The palaeoecology of theseassemblages can now be interpreted with greater confidence.© 2006 Elsevier B.V. All rights reserved.
Keywords: Guild; Epiphytic foraminifera; Eastern Caribbean; Seagrass; Penicillus; Halimeda
⁎ Tel.: +1 868 662 2002x3676; fax: +1 868 662 4414.E-mail address: [email protected].
0377-8398/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.marmicro.2006.10.001

2 B. Wilson / Marine Micropaleontology 63 (2007) 1–18
1. Introduction
It is axiomatic in ecology that neither individualorganisms nor species populations exist in isolation, butinteract with others, as an integral part of a community.Benthonic foraminifera in nearshore waters of the oligo-trophic Caribbean Sea, of which many hundreds of spe-cies have been recorded (Culver and Buzas, 1982), areprimarily epiphytal (Cushman, 1922; Wilson, 1998),living in densely-packed communities on the calcareousalgae Penicillus capitatus, Halimeda spp. and Dasycla-dus vermicularis, and the seagrasses Thalassia testinu-dum and Syringodium filiforme (Steinker and Clem,1984). However, details of the community ecology ofthese Caribbean foraminifera remain little known.
Brasier (1975) suggested that the standing crop andfaunal composition of these epiphytal communities arerelated to the host's structure, sediment content, and thephysical conditions of the habitat, especially salinity andtemperature (see also Semenuik, 2000, 2001). Hedivided foraminifera living on West Indian plants intoprimary (obligate) and secondary (facultative) weed-dwelling faunas, the latter being capable of living bothon plants and in the associated seafloor sediments.However, Langer (1993) noted that there are difficultiesin assigning all motile species to Brasier's primary orsecondary groups, and that Brasier himself occasionallyplaced species in both.
A guild, wrote Root (1967, p. 335), is “a group ofspecies that exploit the same class of environmentalresources in a similar way,” and a guild “groups togetherspecies, without regard to taxonomic position, that over-lap significantly in their niche requirements.” Simberloffand Dayan (1991) wrote that the term “niche” has beenused in two senses: for the functional role of a species in acommunity and as the set of conditions that permits aspecies to exist in a particular biotope. The introductionof the guild concept by Root (1967) effectively limitedthe term niche to the latter definition. Thus, a guild is agroup of species having very similar ecological roleswithin a community, not species occupying precisely thesame niche. Nevertheless, Pianka (1980) describedguilds as “arenas of intense competition.”Niche overlap,a major component of Root's (1967) definition, is mea-sured in three separate dimensions: habitat, food andtime (Pianka, 1974, 1975).
Guilds may be recognised subjectively: Putnam(1994, p. 83) suggested that among insectivorous birdsone might regard foliage gleaners, flycatchers andground feeders as guilds. Brosset et al. (2001) subjec-tively used foraging habitat and feeding habits to placebats in South American rainforests into such guilds as
(a) aerial insectivores in uncluttered space, (b) aerialinsectivores in background cluttered space, and (c)gleaning insectivores in highly cluttered space—although Putnam (1994) had previously noted wrylythat “studies of the resource relationships of aerial,vertebrate insectivores might produce mystifying resultsif bats are not considered alongwith birds.”Many studiesassume that co-generic species compete most strongly(e.g., Walter and Ikonen, 1989; Palmer, 2003—see alsoSimberloff and Dayan, 1991). However, Putnam's(1994) comment reiterates Root's (1967) intention thatguilds may contain unrelated taxa.
Knowlton and Jackson (2001) suggested that sub-jectively defined guilds can be used to overcomeinadequate knowledge of taxonomy. They state, forexample, that studies of the replacement of corals bymacroalgae will capture the essence of this ecologicalphenomenon “even if the species compositions of thetwo groups are ignored.” However, such a broad brushapproach is probably unwise, as it ignores the possibilitythat guilds exist within the coral and macroalgalcommunities. Regarding foraminifera, it might betempting to regard Brasier's (1975) primary andsecondary weed dwellers as guilds. However, as thesecategories were derived from subjective observationsand consider habitat only, this too is unwise.
Among Recent, live organisms, species in guilds willhave similar spatial and temporal distributions. (Notethat this need not be the case with fossils, which can beredistributed in both space and time by post mortemprocesses.) However, not all co-occurring organismswill comprise guilds; predators and their prey, forexample, might be similarly distributed in time andspace (Berryman, 1999, 2002) but, because one uses theother as a food resource, they do not constitute a guild.Following Sugihara's (1980) observation that dendro-grams of population census figures sequentially subdi-vide communities into smaller and more tightly relatedfunctional groups of species, Joern and Lawlor (1981)used cluster analysis to determine guilds among grass-hoppers, and suggested that, while clusters containspecies whose competitive interactions are strongestwithin the guild, interactions between guilds are weaker(see also Crome, 1978; Landres and MacMahon, 1980).Authors have used a range of other multivariatetechniques (Principal Component Analysis, FactorAnalysis) to derive guilds—see, for example, reviewsin Simberloff and Dayan (1991), Holmes et al. (1979),and Baldoa and Drake (2002).
Langer (1993) analysed epiphytal foraminiferalassemblages in twenty-two phytal samples, collected onthree occasions in 1986–1987 from sites in the

3B. Wilson / Marine Micropaleontology 63 (2007) 1–18
Mediterranean Sea, using both Q- and R-mode analyses.Q-mode cluster analysis grouped together assemblageson structurally similar plants such as (a) those with flatleaves, (b) arborescent algae and (c) sediment-trappingalgae and seagrass rhizomes. This implies that weed type,not location, is the main factor affecting epiphytal fora-miniferal community composition. Interpreting R-modeanalysis of 62 epiphytal foraminiferal species on allsubstrates, Langer (1993) presented a simple scheme offour epiphytic morphotypes or bauplans (see alsoKitazato, 1984): Morphotype A, permanently attachedforms (e.g., planorbulinids and Sorites); Morphotype B,temporarily attached forms with a flat attachment surface(e.g., Rosalina, Discorbis, Asterigerina); Morphotype C,suspension feeding motile species (e.g., elphidiids with acarinate or acute periphery); and Morphotype D, per-manently motile, grazing epiphytes (Peneroplis, Textu-laria, Quinqueloculina). Members of Morphotypes Aand B tend to be low and domal; those ofMorphotype Dstand upright. Langer's (1993) language and his Fig. 3indicate that the four morphotypes he recognised do notform clusters, and so must not be regarded as guilds:instead he suggests (Langer, 1993, p. 239) that the spe-cies in each cluster derived from R-mode analysis onlyappear to share common morphological characters.While he does not explicitly draw attention to the wayhis clusters group different morphotypes, his Fig. 3clearly shows that his cluster 3 comprises four species ofElphidium (Morphotype C) and one each of Spiroplecti-nella and Eggerelloides (bothMorphotype D). Converse-ly, two species of Elphidium are clustered in his cluster 2alongside Cibicidella variabilis (Morphotype A), andTextularia bocki and Siphonaperta sp. 1 (Morphotype D),among others. Evidently the niches of species in theseclusters overlap sufficiently for them to operate as guilds,despite their differing bauplans.
Berryman (2002) asserted that the ecology of livingorganisms should be viewed through the lens of popu-lation dynamics over ecological (i.e., short-term) time.(This will not be possible with most fossil assemblages.)Members of a guild will show similar population dynam-ics to one another, guild population size as a whole fluc-tuating as the resources utilised become more or lessavailable. For example, Rappoldt and Hogeweg (1980)noted that populations of all lizards in each of theKalahari, North American and Australian deserts increasewith rainfall. Tedesco and Thunell (2003) inferred that theflux of planktonic foraminifera to the seafloor off northernVenezuela increases during times of upwelling ofnutrient-rich water, and Wilson and Dawe (2006) usedcross-correlation with rainfall data to suggest that this wasprobably a seasonal phenomenon.
Langer's (1993) work notwithstanding, there havenot yet been any explicit studies of guilds among epi-phytal foraminifera. Studies of the population dynamicsof epiphytal foraminifera are also rare (e.g., Wilson,1998). Given that Caribbean foraminifera dwell uponplants, they might be regarded as part of a single, globalmegaguild of epiphytal foraminifera. However, confin-ing all Caribbean species to a single, local guild—Caribbean epiphytal foraminifera—would be overly res-trictive, grouping species with different diets (detriti-vores, herbivores, suspension feeders) and differentmodes of life (permanently attached, temporarily at-tached, etc.) and possibly also different populationdynamics (i.e., blooming at different times of year).Here I examine the guild structure in live foraminifera intwo bays around the NE Caribbean island of Nevis:Mosquito Bay (MB)—also known as Oualie Beach—and Long Haul Bay (LHB). The data set comprisesepiphytal foraminifera that were monitored monthlyover a 12 month period on the calcareous algae Hali-meda opuntia and P. capitatus, and on the exposedrhizomes and leaf bosses (leaves removed) of the sea-grasses S. filiforme and T. testinudum. Cluster analysis isused to discern the guilds in epiphytal foraminiferalcommunities on these plants, and cross-correlations areused to compare the population dynamics of the differentguilds.
2. Study area
The 50 km2 island of Nevis (17°10′N, 62°35′W,Fig. 1) is approximately conical and lies near thenorthern end of the Lesser Antillean island arc (Martin-Kaye, 1969). It is dominated by the centrally-locatedNevis Peak, but comprises nine volcanic centres sur-rounded by lahars (Hutton and Nockolds, 1978; Wilson,1992) and volcanigenic sediments (Wilson, 2005). Theislands' climate is tropical: temperatures vary little overthe year, but there are pronounced wet and dry seasons.The former are marked by hurricanes during which rain-fall can be torrential. Nevis was impacted by N60 hurri-canes and tropical storms between 1871 and 1986 alone(Lugo et al., 2000). The most recent one prior to the timeof sampling (September 1994–August 1995) was Hurri-cane Hugo (September 1989).
The two bays examined here lie in the northern part ofthe island. LongHaul Bay (LHB) is a back-reef bay lyingon the NE (windward) coast, andMosquito Bay (MB) anopen-mouthed embayment on the NW (leeward) coast.The author's unpublished observations following hurri-canes Luis and Marilyn (September 1995) indicate that,except during storms, the leeward MB is exposed to

Fig. 1. A. Sketch-map of the Leeward Islands, NE Caribbean Sea,showing the location of Nevis. B. Nevis, showing the locations ofMosquito Bay (MB) and Long Haul Bay (LHB).
4 B. Wilson / Marine Micropaleontology 63 (2007) 1–18
wave energies lower than those in the windward LHB.During hurricanes, however, shoreline damage is moresevere on the leeward coast, where hurricanes either stripseagrass meadows bare or bury them under shifting sand.The lesser damage along the eastern coast can beascribed to the baffling of storm waves by the fringingreefs ∼200 m offshore along the windward coast.
At the time of sampling both bays contained seagrassmeadows and calcareous algae, although the vegetation inLHB was lusher than that in MB. The seagrass in LHBwas a climax community that comprised primarilyT. testinudum, while that in MB comprised a pioneeringcommunity of primarily S. filiforme (cf. ecological details
in Zieman and Zieman, 1989). Of the four major phytalhabitats identified by Steinker and Clem (1984),D. vermiculariswas found only in LHB,where it was rare.
Overgrazing by sheep and feral donkeys on thewindward side of the island has in recent decadesinduced soil erosion (Wilson, 2005) that is especiallymarked during hurricanes, storms and lesser events(tropical waves). Mosquito Bay was subject to organicenrichment from at least one leaking septic tank(Leonard Huggins, Nevis Island Administration, oralcommunication, 1995), although it was unknown howsevere that impact of the pollution was, or how long ithad been going on. Thus, both bays are undergoing somenutrification, although that in MB is more focused andappears more severe.
3. Materials and methods
Samples were collected during the third week of eachcalendar month. On those occasions when rough seassuspended sediments and rendered the water cloudy,sampling was restricted to substrates that could be lo-cated and identified by touch. All the samples comprisedsediment-trapping algae and seagrass rhizomes sensuLanger (1993).
Three substrates were sampled in MB: H. opuntiafrom a gabion at 30 cm depth at the head of the bay, andP. capitatus heads (capitulae) and exposed S. filiformerhizomes and basal leaf bosses (leaves removed) from anarea ∼200 m2 and 1-m deep immediately seaward of thegabion. The gabion was destroyed during Hurricane Luis.
From LHB P. capitatus capitulae were collected∼10 m from shore within a seagrass meadow adjacent toa patch reef. T. testinudum rhizomes and basal leafbosses (leaves removed) were collected along the steep,shoreward edge of the meadow, all at ∼1 m depth.
Sampling involved combining material from severalindividual plants (i.e., replicates taken were and com-bined in the field) to form “pseudoreplicates” sensuHurbert (1984). All samples of P. capitatus comprisedten capitulae.
The five samples collected each month (three fromMB, two from LHB) were immediately stained bysoaking for 24 h in a solution of 70% isopropyl alcohol(the only preservative readily available on Nevis) androse Bengal. The latter is used to distinguish tests con-taining protoplasm (Walton, 1952) and thought to be liveat the time of sampling. Buzas et al. (1977) had difficultydistinguishing stained and unstained Discorbis rosea,the tests of which are red. This was not found to be aproblem providing a strong solution of rose Bengal wasapplied. Specimens were washed from the plants under

5B. Wilson / Marine Micropaleontology 63 (2007) 1–18
running water over a 63 μm mesh, and the plantsexamined for evidence of permanently attached forami-nifera (Morphotype A) still in place. Few were found.The detached foraminifera were dried at ∼100 °C, andsieved over a 125 μm mesh. Stained specimens werepicked from the N125 μm fraction, sorted into speciesthat were identified using d'Orbigny (1839), Cushman(1921, 1922, 1918–1931) and Schnitker (1971), and thenumber of specimens in each counted. The populationdiversity was measured using the Information Function(H′) and dominance using the Equitability Function E(for details see Magurran, 1988). The entire populationrecovered is referred to as the two-bay metapopulation(cf. Hanski and Gilpin, 1999).
Those species forming N1% of total recovery from anyone substrate, hereafter called common species, weretabulated. Q- (sample-wise) and R-mode (species-wise)agglomerative hierarchical clustering (Pearson's prod-uct–moment correlation coefficient, weighted-averagepair-groupmethod) was conducted across all substrates inboth bays using these common species.R-mode clusteringwas also conducted for the individual bays, using for eachbay only those species common within that bay. To mini-mise terminological complexity, R-mode clusters basedon the two-bay metapopulation are numbered usingroman numerals, those in LHB using uppercase letters,and those in MB using Arabic numerals. Interactionsbetween clusters were examined using cross-correlations(seeWilson and Dawe, 2006), which show correlations atdifferent time lags ô (in this case, months). The statisticalpackage used, STATISTIX version 2.0 (AnalyticalSoftware, 1998), can only compute cross-correlationsfor time-series comprising N12 observations.
4. Results
4.1. Comparison of the two bays using totalforaminiferal recovery
Abundances of the common species recovered aregiven in Tables 1–5. Table 6 gives the monthly andmean values of H′ and E based on all foraminiferarecovered. Several lines of evidence suggest that the twobays lie in different epiphytal foraminiferal biofacies.
In Long Haul Bay total recovery from Thalassiarhizomes and basal leaf bosses (12 samples, 2307 speci-mens, 91 species) was dominated by D. rosea (43.7%)with lesser D. mira (13.0%) and Tretomphalus bulloides(9.6%). Recovery from LHB P. capitatus capitulae (12samples, 3633 specimens, 75 species) was likewise domi-nated overwhelmingly by D. rosea (54.0%): the nextmost abundant species,T. bulloides, comprised only 7.6%
of total recovery. The fauna on Thalassia rhizomes wasboth more diverse and showed less species dominancethan that on P. capitatus capitulae, the mean values of H′and E being higher on the former.
The fauna in Mosquito Bay (MB) was markedlydifferent from that in LHB. Total recovery from MB P.capitatus capitulae (10 samples, 1803 specimens, 78species), was dominated by the porcelaneous Triloculinarotunda (29.9%), with subdominant calcareous-perfo-rate T. bulloides (12.0%). D. rosea formed only 5.5% oftotal recovery from this plant. Recovery from MBH. opuntia (11 samples, 1896 specimens, 90 species)was dominated by T. rotunda (20.9%), with subdominantD. rosea (7.7%), Massilina gualtieriana (6.0%) andTriloculina trigonula (5.7%). On MB Syringodiumrhizomes and basal leaf bosses the fauna (8 samples,964 specimens, 93 species) were dominated by T. rotunda(12.9%), with lesser T. trigonula (7.8%), T. bulloides(5.8%) and Miliolinella circularis (5.6%). In MB theP. capitatus capitulae fauna was the least diverse and hadthe least equitable species distribution, while that onSyringodium rhizomes was the most diverse and had themost equitable distribution of species abundances. Forcomparable substrates in the two bays (P. capitatus,seagrass rhizomes), the fauna in MB was both morediverse and had more equitable species distributions.
4.2. Comparison of the two bays using commonspecies only
A total of 34 species were common (i.e., formed N1%of recovery from at least one substrate) when both baysare considered (for species names and morphotypes,see Fig. 3). Of these, one (Sorites orbiculus) has aMorphotype A morphology and was common only inLHB. Eight belong toMorphotype B, one toMorphotypeC (Elphidium advenum), and 24 toMorphotype D. Whenindividual bays are considered, 27 species (3908 speci-mens comprising common species) were common inMB,while in LHB 17 (5315 specimens) were common. Only 9species were common in both bays: Dicorbis mira,D. rosea, Hauerina bradyi, M. circularis, Peneroplispertusus, Quinqueloculina seminula, T. bulloides, Trilo-culina bermudezi, and T. rotunda.
4.3. Q-mode cluster analysis, two-bay metapopulation
Q-mode agglomerative hierarchical clustering of thetwo-bay metapopulation (i.e., common species fromeither bay on all samples from both bays) produced fiveclusters (Fig. 2), but Clusters II to IV collectively com-prised only six samples. Clusters containing the

Table1
Recoveryof
common
species(i.e.speciesform
ingN1%
oftotalrecovery
from
anyon
esubstrate)
from
Thalassia
rhizom
es,Lon
gHaulBay
Sam
pleno
.(W
ilson
,19
98)
LBHE-C1
LBHE-D
2LBHE-C3
LBHE-A
4LBHE-B5
LBHE-C6
LBHE-E7
LBHE-D
8LBHE-A
9LBHE-B10
LBHE-C11
LBHE-B12
Bay/sub
strate/m
onth
LHBRhiz1
LHBRhiz2
LHBRhiz3
LHBRhiz4
LHBRhiz5
LHBRhiz6
LHBRhiz7
LHBRhiz8
LHBRhiz9
LHBRhiz10
LHBRhiz11
LHBRhiz12
Ammon
iatepida
02
00
00
01
00
00
Articulinamucrona
ta3
10
00
00
00
00
0Articulinasagra
31
00
00
00
00
00
Cym
baloporetta
atlantica
06
33
23
08
41
08
Discorbismira
2921
1620
4013
2353
4415
232
Discorbisrosea
2744
7516
208
6221
788
106
7061
35Elphidium
advenum
70
21
10
23
70
10
Hau
erinabradyi
35
41
00
12
00
00
Hau
erinaornatissima
12
20
00
00
00
00
Miliolasp.
96
01
00
00
10
00
Miliolnella
circularis
46
84
20
13
124
137
Miliolinella
labiosa
20
00
00
00
10
01
Milioliella
vigu
erasi
17
75
12
31
00
00
Penerop
liscarina
tus
34
41
42
21
50
24
Penerop
lispertusus
34
20
31
102
80
30
Quinq
ueloculin
abo
sciana
12
12
01
00
11
14
Quinq
ueloculin
afuna
futiensis
00
30
10
10
01
31
Quinq
ueloculin
apo
lygo
na0
02
00
00
42
11
1
Quinq
ueloculin
aseminula
112
54
13
13
11
23
Rosalinaflo
ridana
02
00
00
00
00
00
Rosalinaorientalis
100
02
21
010
32
42
Soritesorbiculus
03
20
10
10
30
00
Spiroloculinaan
gulata
03
00
00
13
00
21
Tretom
phalus
bullo
ides
44
2720
3014
3333
1212
1221
Trilo
culin
aberm
udezi
180
13
01
00
31
41
Trilo
culin
arotunda
41
621
100
34
14
31
6 B. Wilson / Marine Micropaleontology 63 (2007) 1–18
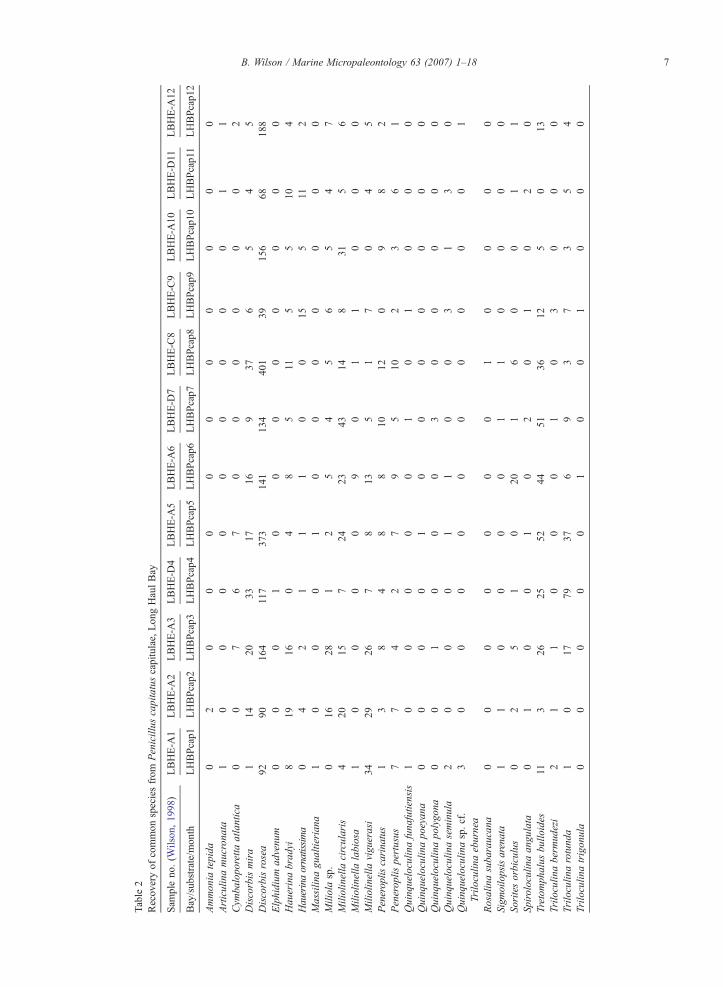
Table2
Recoveryof
common
speciesfrom
Penicillus
capitatuscapitulae,Lon
gHaulBay
Sam
pleno
.(W
ilson
,19
98)
LBHE-A
1LBHE-A
2LBHE-A
3LBHE-D
4LBHE-A
5LBHE-A
6LBHE-D
7LBHE-C8
LBHE-C9
LBHE-A
10LBHE-D
11LBHE-A
12
Bay/substrate/m
onth
LHBPcap1
LHBPcap2
LHBPcap3
LHBPcap4
LHBPcap5
LHBPcap6
LHBPcap7
LHBPcap8
LHBPcap9
LHBPcap10
LHBPcap11
LHBPcap12
Ammon
iatepida
02
00
00
00
00
00
Articulinamucrona
ta1
00
00
00
00
01
1Cym
balopo
retta
atlantica
00
76
70
00
00
02
Discorbismira
114
2033
1716
937
65
45
Discorbisrosea
9290
164
117
373
141
134
401
3915
668
188
Elphidium
advenu
m0
00
10
00
00
00
0Hau
erinabrad
yi8
1916
04
85
115
510
4Hauerinaornatissima
04
21
11
00
155
112
Massilin
agu
altieriana
10
00
10
00
00
00
Miliolasp.
016
281
25
45
65
47
Miliolinella
circularis
420
157
2423
4314
831
56
Miliolinella
labiosa
10
00
09
01
10
00
Miliolinella
viguerasi
3429
267
813
51
70
45
Peneroplis
carinatus
13
84
88
1012
09
82
Penerop
lispertusus
77
42
79
510
23
61
Quinqueloculin
afunafutiensis
10
00
00
10
10
00
Quinqueloculin
apoeyana
00
00
10
00
00
00
Quinqueloculin
apolygona
00
10
00
30
00
00
Quinqueloculin
aseminula
20
00
11
00
31
30
Quinqueloculin
asp.cf.
Trilo
culin
aeburnea
30
00
00
00
00
01
Rosalinasubaraucan
a0
00
00
00
10
00
0Sigm
oilopsisarenata
11
00
00
11
00
00
Soritesorbiculus
02
51
020
16
00
11
Spiroloculinaan
gulata
01
00
10
20
10
20
Tretom
phalus
bullo
ides
113
2625
5244
5136
125
013
Trilo
culin
aberm
udezi
21
10
00
10
30
00
Trilo
culin
arotund
a1
017
7937
69
37
35
4Trilo
culin
atrigonula
00
00
01
00
10
00
7B. Wilson / Marine Micropaleontology 63 (2007) 1–18

Table 3Recovery of common species from Syringodium filiforme rhizomes, Mosquito Bay
Sample no. (Wilson, 1998) NMBA2 NMBD3 NMBD6 NMBF7 NMBA8 NMBC9 NMBC10 NMBB11
Bay/substrate/month MBRhiz2 MBRhiz3 MBRhiz6 MBRhiz7 MBRhiz8 MBRhiz9 MBRhiz10 MBRhiz11
Ammonia sobrina 0 0 0 2 3 4 2 1Ammonia tepida 0 0 2 5 1 1 3 5Articulina mucronata 2 0 0 4 2 6 6 9Articulina sagra 0 0 0 1 1 6 0 0Discorbis mira 0 2 0 0 0 4 5 2Discorbis rosea 0 0 14 0 19 6 0 0Fursenkoina pontoni 0 1 4 1 0 1 2 0Hauerina bradyi 1 1 0 5 3 2 9 10Massilina gualtieriana 0 0 0 2 0 0 0 0Miliolinella circularis 2 5 0 0 5 27 7 8Miliolinella labiosa 1 0 4 5 0 0 0 0Mliolinella viguerasi 4 1 6 6 1 2 3 11Peneroplis pertusus 1 0 0 1 1 1 2 9Quinqueloculina bosciana 4 1 2 2 2 6 1 6Quinqueloculina funafutiensis 3 0 2 0 0 0 1 2Quinqueloculina poeyana 1 1 5 10 2 0 2 4Quinqueloculina polygona 0 4 0 5 8 4 2 4Quinqueloculina seminula 5 1 3 3 0 0 0 11Quinqueloculina sp. cf. Triloculina eburnea 0 2 4 1 0 0 2 3Rosalina floridana 1 0 2 7 0 2 5 15Rosalina subaraucana 2 0 6 3 3 0 2 4Sigmoilopsis arenata 1 0 6 6 4 1 1 7Spiroloculina angulata 2 0 2 4 0 0 1 2Tretomphalus bulloides 3 9 20 0 6 8 4 6Triloculina bermudezi 5 1 3 0 3 4 1 24Triloculina rotunda 9 7 29 15 16 17 4 27Triloculina trigonula 2 3 3 20 4 8 14 21
8 B. Wilson / Marine Micropaleontology 63 (2007) 1–18
remaining 47 samples clearly differentiated the twobays: Cluster I contained 22 samples from LHB only,while Cluster V comprised 23 samples from MB and 2from LHB. Clustering neither grouped similar substrates(P. capitatus capitulae, seagrass rhizomes) in differentbays, nor clearly differentiated between plant substrateswithin individual bays.
4.4. R-mode cluster analysis, two-bay metapopulation
Clustering of the 34 common species in the two-baymetapopulation yielded three clusters (Fig. 3). Cluster Iincluded 19.6% of the foraminiferal fauna in MB, but86.1% of that in LHB. Clusters II and III respectivelyincluded 44.9% and 36.4% of recovery from MB, butonly 12.2% and 1.7% respectively of foraminiferalrecovery from LHB. Of eleven species in Cluster I,which include D. rosea and D. mira, one belonged to theMorphotype A group, one to Morphotype C, four toMorphotype B and a small majority (five species, 45% ofspecies) to Morphotype D. All species in Cluster IIcomprisedMorphotype Dmiliolids. Species in Cluster IIIinclude four in Morphotype B, but the majority (13species, 76% of species) are ofMorphotype D.
4.5. Long Haul Bay
Clustering of the 17 common species in LHB (Fig. 4)did not replicate the clusters for the two-bay metapo-pulation precisely, but produced four clusters. However,the differences between the LHB Clusters A–D and thetwo-bay metapopulation Clusters I–III were not large.
Cluster A (0.8% of LHB common specimens)contained Hauerina ornatissima only, and Cluster D(5.4% of common specimens) only Cymbaloporettaatlantica and T. rotunda. Of the two species in Cluster D,C. atlantica belongs toMorphotype B, and T. rotunda toMorphotype D.
Cluster C (11.6% of LHB common specimens, 5species) contained members ofMorphotypes B,C, andD,and was dominated by D. mira (75.8% of this cluster).Three species in this cluster (D. mira, E. advenum andRosalina orientalis) occur in Cluster I as determinedfrom the two-bay metapopulation, whereas Q. seminulaand T. bermudezi occur in Cluster III.
Cluster B was the largest, both in terms of speciesrichness (9 species) and recovery of specimens in com-mon species (82.1% of LHB common specimens). It wasdominated by D. rosea (68.1% of this cluster), with

Table 4Recovery of common species from Halimeda opuntia, Mosquito Bay
Sample no. (Wilson, 1998) MBD2 MBA3 MBD4 MBA5 MBC6 MBf7 MBC8 MBB9 MBB10 MBC11 MBA12
Bay/substrate/month MBHal2 MBHal3 MBHal4 MBHal5 MBHal6 MBHal7 MBHal8 MBHal9 MBHal10 MBHal11 MBHal12
Ammonia sobrina 2 1 0 0 0 0 0 0 0 1 0Ammonia tepida 3 6 0 4 0 1 2 4 0 1 1Articulina mucronata 9 4 0 3 3 2 3 3 0 1 5Articulina sagra 0 3 0 0 1 2 1 0 4 0 0Cymbaloporetta atlatnica 1 2 0 0 0 0 0 0 0 0 0Discorbis mira 5 3 0 1 6 9 8 7 6 3 2Discorbis rosea 16 4 8 23 34 13 12 8 10 17 2Elphidium advenum 0 0 1 0 0 0 0 0 0 0 0Fursenkoina pontoni 1 5 1 5 2 5 3 1 2 0 1Hauerina bradyi 2 3 2 0 0 0 1 0 1 1 0Hauerina ornatissima 2 0 0 1 0 0 0 1 0 3 2Massilina gualtieriana 2 15 4 1 5 31 7 3 14 28 3Miliola sp. 2 6 0 0 0 0 1 1 0 2 6Miliolinella circularis 3 13 6 11 1 0 1 0 6 10 6Miliolinella labiosa 0 0 0 3 6 11 0 0 0 0 0Miliolinella viguerasi 12 13 20 17 6 5 9 1 4 0 2Peneroplis carinatus 7 0 1 1 0 0 0 0 0 0 0Peneroplis pertusus 1 1 0 0 0 0 0 0 0 0 0Quinqueloculina bosciana 0 5 1 5 0 2 1 5 2 10 7Quinqueloculina
funafutiensis1 2 1 2 1 2 4 8 0 3 0
Quinqueloculina poeyana 3 14 5 4 3 10 0 3 0 3 5Quinqueloculina polygona 17 15 3 3 1 11 8 6 8 6 8Quinqueloculina seminula 0 17 1 4 2 6 6 6 0 5 7Quinqueloculina sp. cf.
Triloculina eburnea0 5 1 1 3 10 0 0 0 2 0
Rosalina floridana 1 0 7 3 6 1 2 1 1 1 2Rosalina orientalis 0 0 0 0 0 3 0 0 1 0 0Rosalina subaraucana 13 6 5 7 4 1 2 2 0 0 0Sigmoilopsis arenata 1 9 0 3 0 1 1 0 0 0 0Sorites orbiculus 1 4 1 1 0 1 0 0 0 0 1Spiroloculina angulata 4 8 4 4 1 2 0 3 0 0 3Tretomphalus bulloides 8 9 2 10 9 11 7 9 10 6 13Triloculina bermudezi 1 3 0 3 0 1 1 8 0 1 8Triloculina rotunda 6 49 30 65 37 59 35 18 19 45 33Triloculina trigonula 6 8 21 14 10 17 4 1 12 7 8
9B. Wilson / Marine Micropaleontology 63 (2007) 1–18
subdominant T. bulloides (11.5%), both of which have aMorphotype B bauplan. The majority of species (67%) inthis cluster belong to Morphotype D, although thismorphotype formed only 19.4% of Cluster B recovery.This cluster most closely resembles Cluster I as derivedfrom the two-bay metapopulation: of the nine species inCluster B, six occurred in metapopulation Cluster I, twoin metapopulation Cluster II, and one in metapopulationCluster III.
4.6. Mosquito Bay
Only species of morphotypes B and D were common(i.e., formed N1% of total recovery from any plant) in thisbay. Clustering of the 27 common species across all threesubstrates sampled in MB produced 4 clusters (Clusters
1–4: Fig. 5). As was the case in LHB, these clusters didnot replicate the two-bay metapopulation Clusters I–IIIprecisely, although the differences were not large. MBCluster 3 contained only Ammonia sobrina, and com-prised only 0.4% of recovery of common specimens fromthis bay.
Cluster 1 (2085 specimens, 53.3% of common speci-mens, 7 species) was dominated by T. rotunda, whichformed 50.8% of this cluster: the subdominantT. bulloides formed 17.6%. Of the seven species inCluster 1, six have aMorphotype D bauplan. This clustermost closely matched Cluster I established using the two-bay metapopulation, but also contained T. bulloides.
Cluster 2 (743 specimens, 19.0% of common speci-mens, 7 species) was dominated by D. rosea, whichcomprised 38.5% of specimens in this cluster.

Table 5Recovery of common species from Penicillus capitatus capitulae, Mosquito Bay
Sample no.(Wilson, 1998)
N:MB-A1 N:MB-A2 N:MB-C3 N:MB-B5 N:MB-B6 N:MB-C7 N:MB-B8 N:MB-A9 N:MB-A10 N:MB-A11
Bay/substrate/month MBPCap1 MBPcap2 MBPcap3 MBPcap5 MBPcap6 MBPcap7 MBPcap8 MBPcap9 MBPcap10 MBPcap11
Ammonia tepida 2 5 1 0 0 0 0 0 1 0Articulina mucronata 17 0 0 0 1 18 14 7 38 12Articulina sagra 0 0 0 0 1 1 6 1 16 11Cymbaloporetta atlantica 0 2 1 2 1 0 0 0 0 0Discorbis mira 1 0 1 2 1 5 3 0 9 2Discorbis rosea 7 12 4 26 16 14 9 2 5 5Elphidium advenum 10 0 0 0 0 0 0 0 0 0Hauerina bradyi 32 5 2 1 1 7 24 5 16 12Hauerina ornatissima 10 1 0 0 1 2 4 3 10 12Massilina gualtieriana 0 0 0 0 0 0 0 0 1 0Miliola sp. 0 0 3 0 2 1 4 6 6 2Miliolinella circularis 28 17 8 13 36 23 10 3 27 12Miliolinella labiosa 0 0 0 1 0 1 1 0 0 0Miliolinella viguerasi 2 2 0 1 1 0 0 1 1 2Peneroplis carinatus 0 2 0 0 0 1 1 2 1 2Quinqueloculinabosciana
1 3 2 1 3 1 1 1 4 3
Quinqueloculinafunafutiensis
0 1 0 1 0 0 0 0 0 0
Quinqueloculina poeyana 1 0 5 1 3 4 0 2 0 0Quinqueloculinapolygona
0 4 3 1 1 7 1 2 3 0
Quinqueloculinaseminula
7 7 0 0 1 2 3 0 0 5
Quinqueloculina sp. cf.Triloculina eburnea
0 1 0 0 0 1 0 1 3 11
Rosalina floridana 0 0 0 1 4 0 1 0 0 0Rosalina orientalis 0 0 0 0 0 0 1 0 0 0Rosalina subaraucana 2 1 2 0 0 1 2 0 0 2Sigmoilopsis arenata 1 5 2 1 0 2 5 2 2 1Sorites orbiculus 1 2 0 1 0 10 0 1 0 0Spiroloculina angulata 0 0 0 2 2 2 0 0 0 0Tretomphalus bulloides 5 24 31 24 34 51 16 1 7 24Triloculina bermudezi 4 3 4 0 0 3 3 1 2 8Triloculina rotunda 43 7 26 37 119 124 26 19 80 59Triloculina trigonula 4 5 0 0 3 2 1 2 12 0
10 B. Wilson / Marine Micropaleontology 63 (2007) 1–18
Quinqueloculina polygona was subdominant (18.2%),and this cluster also contained 11.7% D. mira. Only twospecies (28.6% of species in this cluster) belonged toMorphotype B, the remainder being Morphotype D. Thiscluster most closely resembled Cluster III establishedusing the two-bay metapopulation.
In terms of species richness, Cluster 4 was the larg-est, comprising 13 species, of which 76.9% were ofMorphotype D. It was not, however, the most abundant:Cluster 4 comprised only 1064 specimens (27.2% ofcommon specimens), and thus had a smaller populationthan Cluster 1. Cluster 4 was dominated by T. trigonula,which formed 19.9% of this cluster. All but one of thespecies (P. pertusus) in this cluster occurred in the two-bay metapopulation Cluster III.
5. Discussion
Cluster analysis of the two-bay metapopulationproduced three clusters (Fig. 3), of which none waslimited to a single bay. These are here regarded as guilds,which in turn form part of an epiphytal foraminiferalmegaguild. Cluster III, which was common (36.4% ofrecovery) in MB but rare (1.7% or recovery) in LHB,contains several species known to favour high availabilityof organic carbon (A. sobrina and A. tepida (Sen Guptaet al., 1996; Osterman et al., 2005), Fursenkoina pontoni(Seiglie, 1971) andQuinqueloculina bosciana (Donnelly,1994)), and is interpreted as a “lower-energy guildtolerant of nutrient enrichment.” Conversely Cluster I,which dominated in LHB (86.1% of recovery) but was
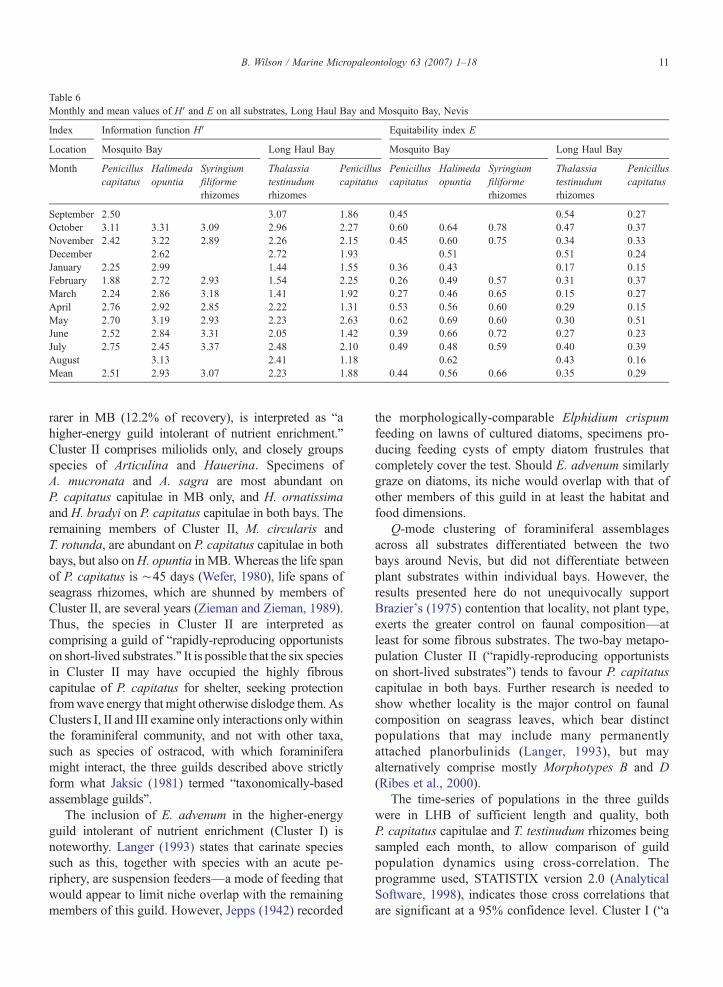
Table 6Monthly and mean values of H′ and E on all substrates, Long Haul Bay and Mosquito Bay, Nevis
Index Information function H′ Equitability index E
Location Mosquito Bay Long Haul Bay Mosquito Bay Long Haul Bay
Month Penicilluscapitatus
Halimedaopuntia
Syringiumfiliformerhizomes
Thalassiatestinudumrhizomes
Penicilluscapitatus
Penicilluscapitatus
Halimedaopuntia
Syringiumfiliformerhizomes
Thalassiatestinudumrhizomes
Penicilluscapitatus
September 2.50 3.07 1.86 0.45 0.54 0.27October 3.11 3.31 3.09 2.96 2.27 0.60 0.64 0.78 0.47 0.37November 2.42 3.22 2.89 2.26 2.15 0.45 0.60 0.75 0.34 0.33December 2.62 2.72 1.93 0.51 0.51 0.24January 2.25 2.99 1.44 1.55 0.36 0.43 0.17 0.15February 1.88 2.72 2.93 1.54 2.25 0.26 0.49 0.57 0.31 0.37March 2.24 2.86 3.18 1.41 1.92 0.27 0.46 0.65 0.15 0.27April 2.76 2.92 2.85 2.22 1.31 0.53 0.56 0.60 0.29 0.15May 2.70 3.19 2.93 2.23 2.63 0.62 0.69 0.60 0.30 0.51June 2.52 2.84 3.31 2.05 1.42 0.39 0.66 0.72 0.27 0.23July 2.75 2.45 3.37 2.48 2.10 0.49 0.48 0.59 0.40 0.39August 3.13 2.41 1.18 0.62 0.43 0.16Mean 2.51 2.93 3.07 2.23 1.88 0.44 0.56 0.66 0.35 0.29
11B. Wilson / Marine Micropaleontology 63 (2007) 1–18
rarer in MB (12.2% of recovery), is interpreted as “ahigher-energy guild intolerant of nutrient enrichment.”Cluster II comprises miliolids only, and closely groupsspecies of Articulina and Hauerina. Specimens ofA. mucronata and A. sagra are most abundant onP. capitatus capitulae in MB only, and H. ornatissimaand H. bradyi on P. capitatus capitulae in both bays. Theremaining members of Cluster II, M. circularis andT. rotunda, are abundant on P. capitatus capitulae in bothbays, but also onH. opuntia in MB.Whereas the life spanof P. capitatus is ∼45 days (Wefer, 1980), life spans ofseagrass rhizomes, which are shunned by members ofCluster II, are several years (Zieman and Zieman, 1989).Thus, the species in Cluster II are interpreted ascomprising a guild of “rapidly-reproducing opportunistson short-lived substrates.” It is possible that the six speciesin Cluster II may have occupied the highly fibrouscapitulae of P. capitatus for shelter, seeking protectionfromwave energy that might otherwise dislodge them. AsClusters I, II and III examine only interactions only withinthe foraminiferal community, and not with other taxa,such as species of ostracod, with which foraminiferamight interact, the three guilds described above strictlyform what Jaksic (1981) termed “taxonomically-basedassemblage guilds”.
The inclusion of E. advenum in the higher-energyguild intolerant of nutrient enrichment (Cluster I) isnoteworthy. Langer (1993) states that carinate speciessuch as this, together with species with an acute pe-riphery, are suspension feeders—a mode of feeding thatwould appear to limit niche overlap with the remainingmembers of this guild. However, Jepps (1942) recorded
the morphologically-comparable Elphidium crispumfeeding on lawns of cultured diatoms, specimens pro-ducing feeding cysts of empty diatom frustrules thatcompletely cover the test. Should E. advenum similarlygraze on diatoms, its niche would overlap with that ofother members of this guild in at least the habitat andfood dimensions.
Q-mode clustering of foraminiferal assemblagesacross all substrates differentiated between the twobays around Nevis, but did not differentiate betweenplant substrates within individual bays. However, theresults presented here do not unequivocally supportBrazier's (1975) contention that locality, not plant type,exerts the greater control on faunal composition—atleast for some fibrous substrates. The two-bay metapo-pulation Cluster II (“rapidly-reproducing opportunistson short-lived substrates”) tends to favour P. capitatuscapitulae in both bays. Further research is needed toshow whether locality is the major control on faunalcomposition on seagrass leaves, which bear distinctpopulations that may include many permanentlyattached planorbulinids (Langer, 1993), but mayalternatively comprise mostly Morphotypes B and D(Ribes et al., 2000).
The time-series of populations in the three guildswere in LHB of sufficient length and quality, bothP. capitatus capitulae and T. testinudum rhizomes beingsampled each month, to allow comparison of guildpopulation dynamics using cross-correlation. Theprogramme used, STATISTIX version 2.0 (AnalyticalSoftware, 1998), indicates those cross correlations thatare significant at a 95% confidence level. Cluster I (“a
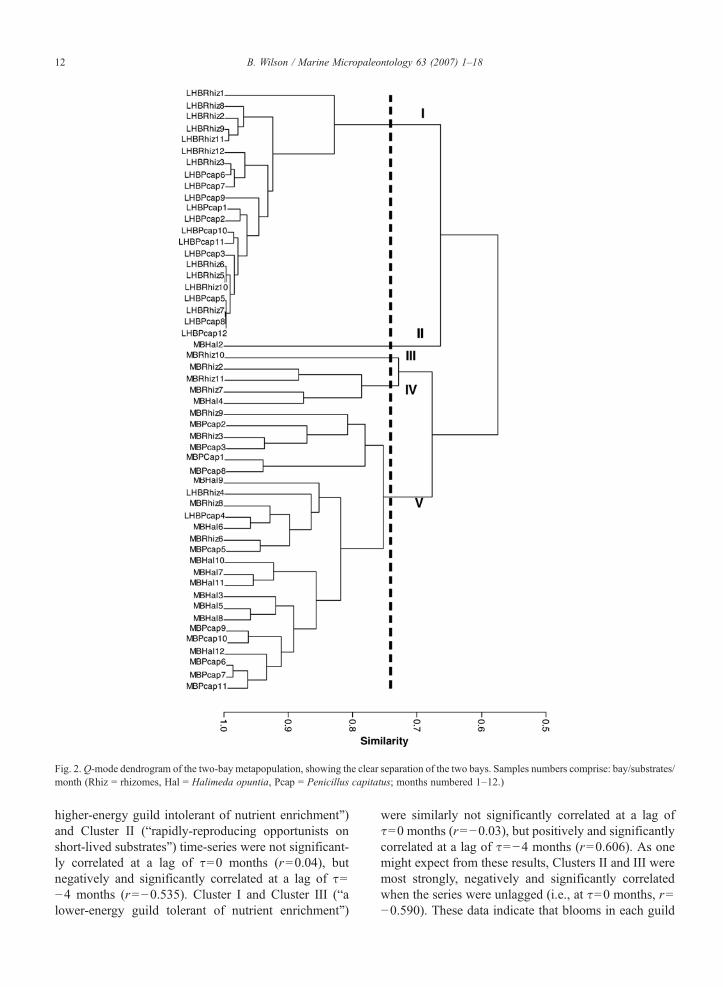
Fig. 2.Q-mode dendrogram of the two-bay metapopulation, showing the clear separation of the two bays. Samples numbers comprise: bay/substrates/month (Rhiz = rhizomes, Hal = Halimeda opuntia, Pcap = Penicillus capitatus; months numbered 1–12.)
12 B. Wilson / Marine Micropaleontology 63 (2007) 1–18
higher-energy guild intolerant of nutrient enrichment”)and Cluster II (“rapidly-reproducing opportunists onshort-lived substrates”) time-series were not significant-ly correlated at a lag of τ=0 months (r=0.04), butnegatively and significantly correlated at a lag of τ=−4 months (r=−0.535). Cluster I and Cluster III (“alower-energy guild tolerant of nutrient enrichment”)
were similarly not significantly correlated at a lag ofτ=0 months (r=−0.03), but positively and significantlycorrelated at a lag of τ=−4 months (r=0.606). As onemight expect from these results, Clusters II and III weremost strongly, negatively and significantly correlatedwhen the series were unlagged (i.e., at τ=0 months, r=−0.590). These data indicate that blooms in each guild

Fig. 3. R-mode dendrogram including, for all substrates, all species common (i.e., forming N1% or total recovery) on any one substrate. Guilds(clusters) I–III separated by horizontal, dashed lines.
13B. Wilson / Marine Micropaleontology 63 (2007) 1–18
are timed such that they do not coincide with oneanother, and support a model of pulsating patches(Buzas et al., 2002).
Clustering of the two-bay metapopulation groupedtogether species belonging to different morphotypes ofepiphytal foraminifera sensu Langer (1993), confirmingthat the morphotypes are not guilds. The two bays eachcontained different percentages of specimens in the fourmorphotypes. Of 17 species common in Long Haul Bay(LHB), one belonged to Morphotype A (permanentlyattached), five to Morphotype B (low, domal, temporar-ily attached), one to Morphotype C (suspensionfeeding), and ten (59% of species), to Morphotype D
(upright, permanently motile). Specimens inMorphotypeB comprised 76% of the total LHB recovery, and Mor-photype D 23%. Of the 27 species common in MosquitoBay (MB), 7 belonged to Morphotype B, and the re-maining 22 (76% of species) to Morphotype D. Speci-mens in Morphotype B comprised 24% of the total MBrecovery, and Morphotype D 76%.
The abundance of Ammonia spp. and Q. bosciana inMB indicates that differences in the communitycomposition between the two bays is related to theflux of organic matter into MB. The greater relativeabundance and species diversity of Morphotype D inMB may, like that of the guild of “rapidly-reproducing
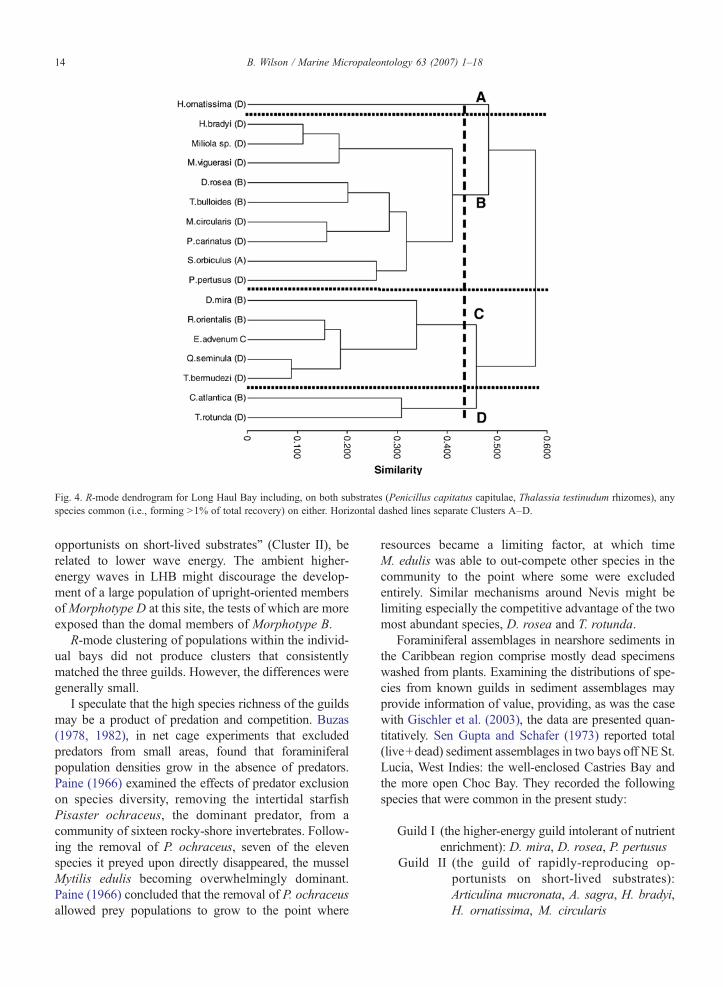
Fig. 4. R-mode dendrogram for Long Haul Bay including, on both substrates (Penicillus capitatus capitulae, Thalassia testinudum rhizomes), anyspecies common (i.e., forming N1% of total recovery) on either. Horizontal dashed lines separate Clusters A–D.
14 B. Wilson / Marine Micropaleontology 63 (2007) 1–18
opportunists on short-lived substrates” (Cluster II), berelated to lower wave energy. The ambient higher-energy waves in LHB might discourage the develop-ment of a large population of upright-oriented membersofMorphotype D at this site, the tests of which are moreexposed than the domal members of Morphotype B.
R-mode clustering of populations within the individ-ual bays did not produce clusters that consistentlymatched the three guilds. However, the differences weregenerally small.
I speculate that the high species richness of the guildsmay be a product of predation and competition. Buzas(1978, 1982), in net cage experiments that excludedpredators from small areas, found that foraminiferalpopulation densities grow in the absence of predators.Paine (1966) examined the effects of predator exclusionon species diversity, removing the intertidal starfishPisaster ochraceus, the dominant predator, from acommunity of sixteen rocky-shore invertebrates. Follow-ing the removal of P. ochraceus, seven of the elevenspecies it preyed upon directly disappeared, the musselMytilis edulis becoming overwhelmingly dominant.Paine (1966) concluded that the removal of P. ochraceusallowed prey populations to grow to the point where
resources became a limiting factor, at which timeM. edulis was able to out-compete other species in thecommunity to the point where some were excludedentirely. Similar mechanisms around Nevis might belimiting especially the competitive advantage of the twomost abundant species, D. rosea and T. rotunda.
Foraminiferal assemblages in nearshore sediments inthe Caribbean region comprise mostly dead specimenswashed from plants. Examining the distributions of spe-cies from known guilds in sediment assemblages mayprovide information of value, providing, as was the casewith Gischler et al. (2003), the data are presented quan-titatively. Sen Gupta and Schafer (1973) reported total(live+dead) sediment assemblages in two bays off NE St.Lucia, West Indies: the well-enclosed Castries Bay andthe more open Choc Bay. They recorded the followingspecies that were common in the present study:
Guild I (the higher-energy guild intolerant of nutrientenrichment): D. mira, D. rosea, P. pertusus
Guild II (the guild of rapidly-reproducing op-portunists on short-lived substrates):Articulina mucronata, A. sagra, H. bradyi,H. ornatissima, M. circularis

Fig. 5. R-mode dendrogram for Mosquito Bay including, on all three substrates (Penicillus capitatus capitulae, Syringodium filiforme rhizomes,Halimeda opuntia), any species common (i.e., forming N1% of total recovery) on any of the three substrates. Horizontal dashed lines separateClusters 1–4.
15B. Wilson / Marine Micropaleontology 63 (2007) 1–18
Guild III (the lower-energy guild tolerant of nutrientenrichment): Ammonia tepida, Miliolinellalabiosa, Quinqueloculina poeyana, Q.polygona, Rosalina floridana, T. trigonula.
Guild I was most abundant in the more exposed ChocBay, where D. rosea formed up to 36.8% of assem-blages, and D. mira up to 11.3%, and was absent to rareat stations in Castries Bay. Guild III was most abundantin the more-enclosed Castries Bay, where A. tepida
formed 13.6–35.7% of assemblages. In Choc BayA. tepida usually formed b3% of assemblages, althoughit formed 14.2% of recovery from one of eleven samplescollected. Guild II was most abundant in the more openChoc Bay. That Guild II did not co-occur with Guild IIIin Castries Bay, but with Guild I in Choc Bay, confirmsthat the factor encouraging the development of GuildII is not raised nutrient levels, as might be inferred fromits distribution around Nevis, but the protection affordedby the substrate on which it lives.

16 B. Wilson / Marine Micropaleontology 63 (2007) 1–18
6. Conclusions
Foraminiferal communities in Caribbean bays arerenowned for their species richness and ecological com-plexity. Buzas et al. (1977, p. 61) lamented, followingcomparisons of the population dynamics of individualspecies in back-reef bays off northern Jamaica, that thecomplexity of the nearshore environment in the Caribbeanmay be so fantastic that “we may never be able to unravelit”. However, examination of a metapopulation from twobays around Nevis has clarified the ecology of the nears-hore epiphytal community on fibrous, sediment-trappingsubstrates.
Q-mode cluster analysis of the foraminiferal metapo-pulation in the two bays clearly distinguished betweenthem, but did not distinguish between fibrous substrates(calcareous algae, seagrass rhizomes) within either bay.R-mode cluster analysis of the same metapopulation in-dicated the presence of three foraminiferal guilds:
Guild I A higher-energy guild intolerant of nutrientenrichment.
Guild II A guild of rapidly-reproducing opportunistson short-lived substrates.
Guild III A lower-energy guild tolerant of nutrientenrichment.
These were not limited to any one bay, but varied inproportion between the two, Guild I being most prolific ina virtually pristine back-reef bay on the windward side ofthe island, and Guilds II and III forming a greater pro-portion of the epiphytal foraminiferal fauna in the bayimpacted by lower-energy waves. Guild II shows a pre-ference for the bushy capitulae of the alga P. capitatus.Cross-correlation of 12-month time-series for the threeguilds in one bay showed that they bloomed sequentially,at intervals of 4 months.
The recognition of guilds—associations of specieswith overlapping niches that interact more strongly witheach other than with other guilds—greatly simplifies thejob of interpreting community ecology in epiphytal fora-minifera, and provides insights useful in the interpretationof sediment assemblages comprising dead epiphytalforaminifera. Guilds recognised in other environmentsmight similarly simplify ecological and palaeoecologicalinterpretations.
Acknowledgements
Thanks are due to the British charity Voluntary ServiceOverseas (VSO), which posted BW to Nevis as aschoolteacher. This afforded him an opportunity to take
samples from such a remote location over a protractedperiod. Thanks are also due to Marty Buzas (SmithsonianInstitution), Robin Whatley (University of Wales, Aber-ystwyth), David Rollinson (Nevis) and JacquelineWilson(The University of the West Indies), each of whom aidedconsiderably, albeit in different ways. This work wassupported by the Research and Publications Fund of theUniversity of the West Indies.
Appendix A
Original citations for the common species recorded.Most of these species were illustrated using SEMphotomicrographs by Radford (1998)
Ammonia sobrina (Shupack) = Rotalia beccarii(Linné) var. sobrina Shupack, 1934
Ammonia tepida(Cushman) = Rotalia beccarii(Linné) var. tepida Cushman, 1926
Articulina mucronata (d'Orbigny) = Vertabralinamucronata d'Orbigny, 1839
Articulina sagra d'Orbigny, 1839Cymbaloporetta atlantica (Cushman) = Tretompha-
lus atlantica Cushman, 1934Discorbis mira Cushman, 1922Discorbis rosea (d'Orbigny) = Rosalina rosea
d'Orbigny, 1839Elphidium advenum (Cushman) = Polystomella
advenaCushman, 1922Fursenkoina pontoni (Cushman) = Virgulina pontoni
Cushman, 1932Hauerina bradyi Cushman, 1917Hauerina ornatissima (Karrer) = Quinqueloculina
ornatissima Karrer, 1868Massilina gualtieriana (d'Orbigny) = Quinquelocu-
lina gualtieriana d'Orbigny, 1839Miliola sp.Miliolinella circularis (Bornemann) = Triloculina
circularis Bornemann, 1855Miliolinella labiosa (d'Orbigny) = Triloculina
labiosa d'Orbigny, 1839Miliolinella viguerasi (Bermudez) = Triloculina
viguerasi Bermudez, 1935Peneroplis carinatus d'Orbigny, 1839Peneroplis pertusus (Forskål) = Nautilus pertusus
Forskål, 1775Quinqueloculina bosciana d'Orbigny, 1839Quinqueloculina funafutiensis (Chapman) = Milio-
lina funafutiensis Chapman, 1901Quinqueloculina poeyana d'Orbigny, 1839Quinqueloculina polygona d'Orbigny, 1839Quinqueloculina seminula (Linné) = Serpula semi-
nulum Linné, 1758

17B. Wilson / Marine Micropaleontology 63 (2007) 1–18
Quinqueloculina sp. cf. Triloculina eburnea d'Or-bigny, 1839
Rosalina floridana Cushman, 1922Rosalina orientalis (Cushman) =Discorbis orientalis
Cushman, 1925Rosalina subaraucana (Cushman) = Discrobis sub-
araucana Cushman, 1922Sigmoilopsis arenata (Cushman) = Spiroloculina
arenata Cushman, 1921Sorites orbiculus (Forskål) = Nautilus orbiculus
Forskål, 1775Spiroloculina angulata Cushman, 1921Tretomphalus bulloides (d'Orbigny) = Rosalina
bulloides d'Orbigny, 1839Triloculina bermudezi Acosta, 1940Triloculina rotunda d'Orbigny, 1839Triloculina trigonula (Lamarck) = Miliola trigonula
Lamarck, 1804
References
Acosta, J.T., 1940. Triloculina bermudezi, un nuevo foraminifera delas Islas Bahamas. Memorias de la Sociedad Cubana de HistoriaNatural 14, 37–38.
Analytical Software, 1998. STATISTIX for Windows, User's Manual,333 p.
Baldoa, F., Drake, P., 2002. A multivariate approach to the feedinghabits of small fishes in the Guadalquivir Estuary. Journal ofFish Biology 61, 21–32.
Bermudez, P.J., 1935. Foraminiferos de la Costa Norte de Cuba.Memorias de la Sociedad Cubana de Historia Natural “FelipePoey” 9, 129–244.
Berryman, A.A., 1999. Principles of Population Dynamics and theirApplication. Stanley Thornes Publishes, Cheltenham, England.243 pp.
Berryman, A.A., 2002. Population: a central concept for ecology.Oikos 97, 439–442.
Bornemann, J.G., 1855. Die mikroskopische Fauna des Septar-ienthones von Hermsdorf bei Berlin. Zeitschrift der DeutschenGeologischen Gesellschaft 7, 307–371.
Brasier, M.D., 1975. Ecology of Recent sediment-dwelling and phytalforaminifera from the lagoons of Barbuda, West Indies. Journal ofForaminiferal Research 5, 42–62.
Brosset, A., Charles-Dominique, P., Cockle, A., 2001. The batcommunity. In: Bongers, F., Charles-Dominique, P., Forget, P.-M.,Théry, M. (Eds.), Nouragues: Dynamics and Plant–Animal Interac-tions in a Neotropical Rainforest. Kluwer Academic, Dordrecht,Holland, pp. 115–120.
Buzas, M.A., 1978. Foraminifera as prey for benthic deposit feeders:results of predator exclusion experiments. Journal of MarineResearch 36, 617–625.
Buzas, M.A., 1982. Regulation of foraminiferal densities by predationin the Indian River, Florida. Journal of Foraminiferal Research 12,66–71.
Buzas, M.A., Smith, R.K., Beem, K., 1977. Ecology andsystematics of foraminifera in two Thalassia habitats, Jamaica,West Indies. Smithsonian Contributions to Paleobiology, vol. 31.138 pp.
Buzas, M.A., Hayak, L.-A., Reed, C., Jett, S.A., 2002. Foraminiferaldensities over five years in the Indian River Lagoon, Florida: amodel of pulsating patches. Journal of Foraminiferal Research32, 68–92.
Chapman, F., 1901. Foraminifera from the lagoon at Funafuti. Journalof the Linnean Society of London, Zoology 28, 161–210.
Crome, F.H.J., 1978. Foraging ecology of an assemblage of birds inlowland rainforest in northern Queensland. Australian Journal ofEcology 3, 195–212.
Culver, S.J., Buzas, M.A., 1982. Distribution of Recent BenthicForaminifera in the Caribbean Region. Smithsonian Contributionsto Marine Science, vol. 14. 382 pp.
Cushman, J.A., 1917. A monograph of the foraminifera of the NorthPacific Ocean. Part 6. Miliolidae. Bulletin of the United StatesNational Museum 71 (6), 1–108.
Cushman, J.A., 1918–1931. The Foraminifera of the Atlantic Ocean,Parts 1–8. Bulletin of the US National Museum, vol. 104.
Cushman, J.A., 1921. Foraminifera from the north coast of Jamaica.Preceedings of the US National Museum, vol. 59, pp. 47–82.
Cushman, J.A., 1922. Shallow-water foraminifera of the Tortugasregion, vol. 17. Carnegie Institution of Washington, pp. 3–85.
Cushman, J.A., 1925. Foraminifera of the tropical central Pacific.Bernice P. Bishop Museum, Honolulu, Bulletin 27, 121–144.
Cushman, J.A., 1926. Recent foraminifera from Porto Rico. Publica-tions of the Carnegie Institution of Washington no. 344,Department of Marine Biology Papers 23, 73–84.
Cushman, J.A., 1932. Notes on the genus Virgulina. Contributionsfrom the Cushman Laboratory for Foraminiferal Research 8, 7–23.
Cushman, J.A., 1934. Notes on the genus Tretomphalus, withdescriptions of some new species and a new genus, Pyropilus.Contributions from the Cushman Laboratory for ForaminiferalResearch 10, 79–101.
d'Orbigny, A.C., 1839. Foraminifères. In: de la Sagra, M.R. (Ed.),Histoire Physique, Politique et Naturelle de l'Ile de Cuba. 224 pp.
Donnelly, K.B., 1994. Characterization of the foraminiferal assem-blage off the southwest coast of Puerto Rico at La Parguera.Unpublished M. Sc. Thesis, University of South Florida, Tampa,Florida, 136 pp.
Forskål, P., 1775. Descriptiones animalium. Haunie, Copenhagen.164 p.
Gischler, E., Hauser, I., Heinrich, K., Scheitel, U., 2003. Character-ization of Depositional Environments in Isolated CarbonatePlatforms Based on Benthic Foraminifera, Belize, CentralAmerica. Palaios 18, 236–255.
Hanski, I., Gilpin, M., 1999. Metapopulation Biology: Ecology,Genetics and Evolution. Academic Press, London. 512 pp.
Holmes, R.T., Bonney, R.E., Paccala, S.W., 1979. Guild structure ofthe Hubbard Brook bird community. Ecology 60, 512–520.
Hurbert, S.J., 1984. Pseudoreplication and the design of ecologicalexperiments. Ecological Monographs 54, 187–211.
Hutton, C.O., Nockolds, S., 1978. The petrology of Nevis, LeewardIslands, West Indies. Institute of Geological Sciences, OverseasGeology and Mineral Resources, vol. 52. 31 pp.
Jaksic, M.F., 1981. Use and abuse of the term “guild” in ecologicalstudies. Oikos 37, 397–400.
Jepps, M.W., 1942. Studies on Polystomella Lamarck (Foraminifera).Journal of the Marine Biological Association of the UnitedKingdom 25, 607–666.
Joern, A., Lawlor, L.R., 1981. Guild structure in grasshopper assem-blages based on food and microhabitat resources. Oikos 37, 93–104.
Karrer, F., 1868. Die Miocene Foraminiferenfauna von Kostejim Banat. Sitzungberichte der Kaiserlichen Akademie der

18 B. Wilson / Marine Micropaleontology 63 (2007) 1–18
Wissenschaften in Wien, Mathematisch-NaturwissenschaftlicheKlasse 58, 121–193.
Kitazato, H., 1984. Microhabitats of benthic foraminifera and theirapplication to fossil assemblages. Benthos '83; Second Interna-tional Symposium on Benthic Foraminifera (Pau, April 1983),pp. 339–344.
Knowlton, N., Jackson, J.B.C., 2001. The ecology of coral reefs.In: Bertness, M.D., Gaines, S.D., Hay, M.E. (Eds.), MarineCommunity Ecology. Sinauer Associates, Sunderland, Massa-chusetts, pp. 395–422.
Lamarck, J.B., 1804. Suite des mêmoires sur les fossils des environsde Paris. Annales Muséum National d’Histoire Naturelle 5,349–357.
Landres, P.B., MacMahon, J.A., 1980. Guilds and communityorganization: analysis of an oak woodland avifauna in Sonora,Mexico. Auk 97, 351–365.
Langer, M.R., 1993. Epiphytic foraminifera. Marine Micropaleontol-ogy 20, 235–265.
Linné, C., 1758. Sytema Naturae, 10th edition. Holmiae, Stockholm,Sweden. 824 p.
Lugo, A.E., Rogers, C.S., Nixon, S.W., 2000. Hurricanes, coral reefsand rainforests: resistance, ruin and recovery in the Caribbean.Ambio 29, 106–114.
Magurran, A.E., 1988. Ecological Diversity and its Measurement.Chapman and Hall, London, England.
Martin-Kaye, P.H.A., 1969. A summary of the geology of the LesserAntilles. Institute of Geological Sciences, Overseas Geology andMineral Resources, vol. 10, pp. 172–205.
Osterman, L.E., Poore, R.Z., Swarzenski, P.W., Turner, R.E., 2005.Reconstructing a 180 yr record of natural and anthropogenicinduced low-oxygen conditions from Louisiana continental shelfsediments. Geology 33, 329–332.
Paine, R.T., 1966. Food web complexity and species diversity.American Naturalist 100, 65–75.
Palmer, T.M., 2003. Spatial habitat heterogeneity influences compe-tition and coexistence in an African acacia ant guild. Ecology 84,2843–2855.
Pianka, E.R., 1974. Niche overlap and diffuse competition. Proceed-ings of the National Academy of Sciences of the United States ofAmerica 71, 2141–2145.
Pianka, E.R., 1975. Niche relations of desert lizards. In: Cody, M.L.,Diamond, J. (Eds.), Ecology and Evolution of Communities.Harvard University Press, Cambridge, USA, pp. 292–314.
Pianka, E.R., 1980. Guild structure in desert lizards. Oikos 35,194–201.
Putnam, R.J., 1994. Community Ecology. Chapman and Hall, London,England. 178 pp.
Radford, S., 1998. Foraminifera from the Southern Caribbean–Atlantic Province. Transactions of the 3rd Geological Conferenceof the Geological Society of Trinidad and Tobago, Port-of-Spain,Trinidad, July 16–21, 1995, pp. 163–178.
Rappoldt, C., Hogeweg, P., 1980. Niche packing and number ofspecies. American Naturalist 116, 480–492.
Ribes, T., Salvadó, H.J.R., del Pilar Gracia, A., 2000. Foraminiferalcolonization on artificial seagrass leaves. Journal of ForaminiferalResearch 30, 192–201.
Root, R.B., 1967. The niche exploitation pattern of the blue-grey gnat-catcher. Ecological Monographs 37, 317–350.
Schnitker, D., 1971. Distribution of foraminifera on the North Carolinacontinental shelf. Tulane Studies in Geology and Paleontology 8,169–215.
Seiglie, G.A., 1971. A preliminary note on the relationships betweenforaminifers and pollution in two Puerto Rican bays. CaribbeanJournal of Science 11, 93–98.
Semenuik, T.A., 2000. Spatial variability in epiphytic foraminifera frommicro- to regional scale. Journal of Foraminiferal Research 30,99–109.
Semenuik, T.A., 2001. Epiphytic foraminifera along a climaticgradient, Western Australia. Journal of Foraminiferal Research31, 191–200.
Sen Gupta, B.K., Schafer, C.T., 1973. Holocene benthonic foraminif-era in leeward bays of St. Lucia, West Indies. Micropaleontology19, 341–365.
Sen Gupta, B.K., Turner, R.E., Rabalais, N.N., 1996. Seasonal oxygendepletion in continental-shelf waters of Louisiana: historical recordof benthic foraminifers. Geology 24, 227–230.
Shupack, B., 1934. Some Foraminifera from western Long Island andNew York Harbor. American Museum Novitates 737. 12 p.
Simberloff, D., Dayan, T., 1991. The guild concept and the structure ofecological communities. Annual Review of Ecology and System-atics 22, 115–143.
Steinker, D.C., Clem, K.V., 1984. Some near-shore foraminiferalassemblages from phytal and bottom sediments, Bermuda. TheCompass 61, 98–115.
Sugihara, G., 1980. Minimal community structure: an explanation ofspecies abundance patterns. American Naturalist 116, 770–787.
Tedesco, K.A., Thunell, R.C., 2003. Seasonal and interannual variationsin planktonic foraminiferal flux and assemblage composition in theCariacou Basin, Venezuela. Journal of Foraminiferal Research 33,192–210.
Walter, D.E., Ikonen, E.K., 1989. Species, guilds, and functionalgroups: taxonomy and behavior in nematophagous arthropods.Journal of Nematology 21, 315–327.
Walton, W.R., 1952. Techniques for recognition of living foraminifera.Contributions to the Cushman Foundation for ForaminiferalResearch 3, 56–60.
Wefer, G., 1980. Carbonate production by algae Halimeda, Penicillusand Padina. Nature 285, 323–324.
Wilson, B., 1992. The Saddle Hill Eruptive Centre, Nevis, West Indies.Geology Today 8, 28–29.
Wilson, B., 1998. The ecology and population dynamics of epiphytalforaminifera, Nevis, West Indies. Unpublished PhD theses,University of Wales, Aberystwyth, UK.
Wilson, B., 2005. A relict alluvial fan at Coconut Walk Estate, Nevis,West Indies. Caribbean Journal of Science 41, 14–21.
Wilson, B., Dawe, R.A., 2006. Detecting seasonality using timeseries analysis: comparing foraminiferal population dynamicswith rainfall data. Journal of Foraminiferal Research 36,108–115.
Zieman, J.C, Zieman, R.T., 1989. The ecology of the seagrass meadowsof the west coast of Florida: a community profile. Biological Reportof the US Department of the Interior, vol. 85. 155 pp.





























