Embed Size (px)
Citation preview

AI
HABITAT EVALUATION PROCEDURES AT RAY ROBERTS LAKE:
AN ANALYSIS OF THE RELATIONSHIP WITH ECOLOGICAL
INDICATORS AND A STUDY OF OBSERVER AND
TEMPORAL VARIABILITY
THESIS
Presented to the Graduate Council of the
University of North Texas in Partial
Fulfillment of the Requirements
For the Degree of
MASTER OF SCIENCE
By
Jane M. Wattrus, B.S.
Denton, Texas
December, 1993

6rk4
Wattrus, Jane M., Habitat Evaluation Procedures at Ray
Roberts Lake: An Analysis of the Relationship with
Ecological Indicators and a Study of Observer and Temporal
Variability:. Master of Science (Environmental Science),
December 1993, 98 pp., 4 tables, 18 figures, references 41
titles.
Habitat Evaluation Procedure data gathered at Ray
Roberts Lake in 1989 and 1990 were analysed for temporal
variability, observer variability and relationships between
Habitat Units (HUs) and species density/diversity.
observer variability within a group was analysed by
cluster analysis and bootstrapping. Five out of 36 sites
showed significant differences in Habitat Suitability Index
(HSI) values within the group.
A nonparametric Mann-Whitney test was used to analyze
temporal variability. One of 6 sites showed a significant
difference in HSI values between years.
Using Spearman's Rank Correlation Coefficient, a
correlation was found between indicator species density and
HUs. No significant correlation was indicated between
species diversity and HUs.

ACKNOWLEDGMENTS
I wish to thank my major professor, Dr. Sam Atkinson and
the members of my thesis committee, Dr. Ken Dickson and Dr.
Jim Kennedy.
I would like to thank Dr. Ken Steigman for providing some
of the data used in this study and Gini Kennedy for her
assistance with the preparation of some of the figures.
Finally, I would like to thank my husband Dr. Nigel
Wattrus for his guidance and patience in the preparation of
this thesis.
This work was in part supported with funds provided by
the U. S. Army Corp of Engineers.
iii

TABLE OF CONTENTS
ACKNOWLEDGEMENTS.........................................iii
TABLE OF CONTENTS.......................................iv
LIST OF TABLES............................................vi
LIST OF FIGURES..........................................evii
Chapter
1. INTRODUCTION............................... ...... I
Brief backgroundObjectives of the studyScope of the studyBrief description of contents ofremaining chapters
2. LITERATURE REVIEW........................ ...... 5
Outline of chapter contentsTemporal and observer variabilitySpecies density and habitat quality
3. METHODS AND PROCEDURES...........................19
Outline of chapter contentsStudy area and habitat descriptionsSpecies selection and habitat suitability modelsHabitat Evaluation Procedures field evaluationsBird censusesGuild informationStatistical analyses
4. RESULTS.... - -.................. ................... 36
Outline of chapter contentsHypothesis 1: observer variabilityHypothesis 2: temporal variabilityHypothesis 3: density vs. habitat quality
iv

Hypothesis 4: species diversity vs. habitatquality
5. CONCLUSIONS........... ................. 59
Outline of chapter contentsDiscussion of each hypothesis and resultRecommendations for future study
APPENDIX..-...--..------....--.-....................66
A) Habitat suitability index life requisite valuemodels for indicator species
B) Habitat suitability index values for thevegetational transects
C) Habitat suitability index values for the Impactassessment sites
D) Avian census numbers by habitat type and transectsection for 1987 and 1990
E) Bray-Curtis cluster analysis data
REFERENCES. ............................................ 94
V

LIST OF TABLES
Table Page
1. Indicator species and models used for theHEP analysis at Ray Roberts Lake...................24
2. Matrix of variables and species used per HabitatEvaluation Procedures.---------.................26
3. Avian species guilded with HEP indicator speciesas a result of various guild studies.............. 31
4. Habitat units, indicator species density andspecies richness for various impact sites andtransect habitats.--------............... 56
vi

LIST OF FIGURES
Figure Page
1. Location of Ray Roberts Lake in Relationshipto the Dallas-Fort Worth Metroplex................ 2
2. Location of vegetational transects at RayRoberts Lake and habitat types along eachtransect------..------------...-..............2 3
3. Location of the impact assessment sites usedin the HEP----------------...................... 28
4. Bird guilds of the contiguous U.S.A............. 33
5. Bray-Curtis cluster analysis of mean habitatsuitability index values for five observersat impact site water habitat (Green Heronmodel)...----------- ..-................... 38
6. Water habitat Green Heron model variable 2representing habitat suitability index vs.percent of water area < 25 cm deep............... 39
7. Bray-Curtis cluster analysis of mean habitatsuitability index values for five observers attransect EF water habitat (Green Heron model).... 40
8. Bray-Custer cluster analysis of mean habitatsuitability index values for five observers attransect GH water habitat (Green Heron model).... 41
9. Water habitat Green Heron model variable 1representing habitat suitability index vs.aquatic substrate composition in littoral
10. Water habitat Green Heron model variable 4representing habitat suitability index vs.percent of water surface covered by logs,trees, or woody vegetation...................... 44
11. Bray-Curtis cluster analysis of mean habtatsuitability index values for five observers attransect AB shrublandhabitat....................45
vii

12. Shrubland habitat Eastern Woodrat model variable3 representing habitat suitability index vs.number of refuge sites per 0.4 ha................46
13. Bray-Curtis cluster analysis of mean habitatsuitability index values for five observers attransect EF oldfield habitat (E. Cottontailmodel)........................................... 47
14. Oldfield habitat Eastern Cottontail modelvariable 4 representing habitat suitabilityindex vs. percent shrub crown cover.............. 49
15. Oldfield habitat Eastern Cottontail modelvariable 5 representing habitat suitabilityindex vs. number of refuge sites per 0.4 ha.......50
16. Upland forest habitat Fox Squirrel modelvariable 1 representing habitat suitabilityindex vs. percent tree canopy closure ofmast producing trees > 25 cm dbh.................53
17. Upland forest habitat Fox Squirrel modelvariable 3 representing habitat suitabilityindex vs. average diameter at breast height(dbh) of overstorytrees...................... 54
18- Correlation of Habitat Units and AvianIndicator Species Density at Ray Roberts..........57
viii

CHAPTER I
INTRODUCTION
In July of 1987 the construction of Ray Roberts Lake in
northern Denton County, Texas (Fig. 1) was completed by the
U.S. Army Corps of Engineers. A two year pre-impoundment
environmental study of the lake area was started by the
Institute of Applied Sciences at the University of North
Texas in January of 1986. The objective of this study was to
develop baseline descriptions of the wildlife resources in
the area prior to impoundment. In addition to censuses of
vegetation, fish, mammals and birds another component of the
study entailed applying Habitat Evaluation Procedures-(HEP).
HEP was developed by the U.S. Fish and Wildlife Services
(USFWS) to evaluate habitat quality and quantity.
Habitat Evaluation Procedures were initially developed
in 1974, by USFWS, in response to legislation such as the
Fish and Wildlife Coordination act of 1934 and the National
Environmental Policy Act (NEPA) of 1969. NEPA requires that
all federally funded or permitted projects must consider
possible adverse impacts on fish and wildlife. This created
a need for a system of habitat evaluation that would
facilitate assessment of the effects of actions taken in a
federal project. In 1980, HEP was further developed with
1

2
ISL.E O 05E 4
RAY ROBERTS LAKEJAL F..Y. 4 ;.
L / T T
LEWISVLLE'1LAKE
CEN N - -
I NCX EQUALS MILES
CQ 6APYI N E---
L AK E
FT WCATM ALA
Figure 1. Location of Ray Roberts Lake in Relationship tothe Dallas-Fort Worth Metroplex. (IAS, 1992)

3
the use of documented habitat models and analyses of
individual evaluation species rather than by examining cover
types alone (USFWS, 1980a).
The basic unit of HEP is the Habitat Suitability Index
(HSI). Ecological variables (e.g. percent shrub crown cover,
height of mast producing trees, number of refuge sites per
0.4 ha, etc.) are measured in the field and then entered
into the selected evaluation species model. HSIs range from
0 to 1.0 and are determined by this model. Habitat Units
(HUs) are a product of the HSI and the acreage of the
habitat in question.
HEP provides a habitat-based approach for assessing
environmental impacts of proposed land and water resource
developed projects. It can be used to document the quality
and quantity of habitat available for a selected species.
HEP provides information on the relative value of different
areas at a particular point in time (USFWS, 1980a).
A two year post-impoundment study of Ray Roberts Lake
was begun by the Institute of Applied Sciences in January of
1989. HEP was utilized in an attempt to examine the
condition of wildlife habitat in 1990 and to determine if
any changes in the habitat had occurred since the initial
baseline study in 1987.
During data collection and analyses, several questions
arose about HEP which led to the following hypotheses: (1)
there is no significant difference in Habitat Suitability
Index variable values between individuals in a survey party,

4
(2) there is no significant difference in mean Habitat
Suitability Index values at impact assessment sites between
sampling years, (3) for a particular habitat type there is
no correlation between Habitat Units and avian Indicator
Species density, (4) for a particular habitat type there is
no correlation between Habitat Units and avian species
diversity.
In order to address the above four hypotheses, several
statistical methods were utilized to analyze the HEP data
and bird census data which were gathered and tabulated for
both the 1987 pre-impoundment and 1990 post-impoundment
studies.
The following sections of this thesis contain a review
of the literature pertaining to the outlined hypotheses, a
description of the methods and procedures which were used, a
presentation and analysis of the -results and conclusions.

CHAPTER 2
LITERATURE REVIEW
This section reviews the literature which pertains to
the objectives of this study. The information will be
subdivided into two subsections; 1) Temporal and Observer
Variability; and 2) Species Density/Diversity and Habitat
Quality.
The first subsection deals with previous studies
involved in the assessment of observer variability in
estimating/measuring habitat characteristics, and also what
effect making estimates at different time periods has on the
results at a particular habitat. HEP is traditionally used
to evaluate initial conditions in a habitat and then make
projections about the future condition of that habitat. In
this study HSIs were calculated for initial conditions in
1987 and then calculated at a later time (i.e. :1990) for
specific habitats. The results of these HEP studies
conducted at different times, in the same habitat, were then
compared. Upon reviewing the literature in this area, it was
noted that this was an unconventional way of utilizing HEP,
and prior studies involving temporal variability are scarce.
The second subsection provides a review of a variety of
studies which attempted to relate species abundance measures
to habitat characteristics and quality.
5

6
Temporal and Observer Variability
The collection of habitat information is inherent to
the HEP developed by USFWS. Many of the variables that are
integral in developing a HSI are derived from observer
estimates of specific habitat characteristics (i.e.: percent
herbaceous canopy cover). Inevitably, variation in the
measurements of HSI variables will exist among members of a
survey team, despite "group training" in variable
measurement techniques. Gotfryd and Hansell (1985) note that
the problem of variation due to observer differences has in
the past received little attention in the area of field
ecology.
Smith (1944) studied the accuracy and variation between
estimates of vegetation density made by individuals in a
group. He also described what impact a group training period
had on the results. Eight individuals, familiar with
vegetation density measurements, made estimates over a seven
day period. During that time attempts were made to unify the
individuals estimates. An analysis of variance (ANOVA) of
the data from the first, fourth and seventh days of the
survey showed significant differences among the individuals
in the group (p = 0.05). Results also indicated considerable
change in each individual's estimates of vegetation density
during the week. These became more conservative as the
experiment proceeded. The range of estimates made by the

group also decreased as the week progressed. This was not
unexpected due to the group training which was provided.
In a study by Ellis et al. (1979) four different field
methods of habitat evaluation were tested. Several questions
were addressed: (1) are there seasonal (temporal)
differences in scoring by each evaluation method; (2) does
prior experience in habitat evaluation effect scoring; and
(3) how accurate are observers in estimating vegetational
characteristics. Ninety-seven biologists participated in
five field tests each lasting three consecutive days. Each
observer was assigned one of the following habitat
evaluation methods;
1) HEP Form 3-1101 (USFWS, 1976). This method consists
of a form with columns representing species or groups of
species (evaluation elements) and rows representing
evaluation sites. Test participants assign a score on a
scale of 0 - 10 for each evaluation element. HEP Form 3-1101
is a totally subjective method of habitat evaluation, with
no written criteria.
2) HEP Blue Handbook (Flood et al., 1977). Life history
information is provided for each species grouping (e.g.,
Forest Game Species group - white-tailed deer and wild
turkey). Written criteria are given on five-point or ten-
point scales. Resulting scores are then combined using
formula provided.
3) Line Chart (Whitaker et al., 1976). Vegetative
characteristics are objectively estimated by the user with
7

8
this method. Instructions and definitions are provided.
Estimates are entered on a scaled horizontal line and are
later translated into scores ranging from 0 - 10, for
various wildlife species.
4) Matrix Method (Ellis et al., 1978). This method
includes definitions and instructions and is vegetation
orientated. Columns on this form represent habitat
characteristics, while sites with their estimates are
entered in the rows. Estimates of habitat characteristics
from this form are then translated into scores on a scale of
0 - 10 as with the Line Chart method above.
To determine if there were temporal differences in
scoring by each evaluation method, observers habitat scores
for specific species (i.e. deer, quail, rabbit and turkey)
were examined for consistency within and between seasons.
The coefficient of variation (CV) for scores from all
methods, except HEP Form 3-1101, were low (range: 6-39)
inferring consistency in scoring within each season. The CV
for scores from HEP Form 3-1101 method were high (range:28-
90) suggesting consistent scoring within any test period was
improbable with that methodology.
Mean scores derived by the same methods were found to
differ according to season. For all methods, except for two
cases with the Matrix method, mean scores were lowest in
January. Low scores in January were attributed to difficulty
in assessing sites under winter conditions such as snow
cover, no foliage, etc. (Ellis et al.,1979).

9
ANOVA was used to test for significant differences
between seasons (p = 0.05) and pairs of means were then
compared by least significant differences (LSD) analysis (p
< 0.05). Seasonal differences for all four methods were
detected and attributed to the overall low mean scores in
January.
Coefficients of variation (CV) and mean scores of
experienced versus inexperienced participants were analyzed
to determine the effects of prior experience. Results
indicated that participants using all methods, except HEP
Form 3-1101, produced consistent scores regardless of
experience. Significant differences were detected (ANOVA (p
< 0.05)) in mean scores in Oldfields, between experienced
and inexperienced participants, using both HEP Form 3-1101
and HEP Blue Handbook methods. This was attributed to the
subjective and less structured nature of these two methods
giving advantage to experienced biologists.
Detailed measures of site characteristics were made at
each study site by trained and experienced staff to
determine observer accuracy in estimations of vegetational
characteristics. These data were then considered "baseline"
data and were compared to participant's field estimations.
Results indicate that estimations of percent canopy closure
and timber types were generally accurately estimated. Other
parameters were estimated less accurately and this loss of
accuracy was attributed to two factors: (1) some habitat
characteristics are intrinsically difficult to estimate

10
(i.e. percentage of dead trees in overstory); and (2) some
criteria are subjectively worded (i.e: use of the word
"common").
Gotfryd and Hansell (1985) used multivariate vegetation
observations to evaluate the effects of subjectivity on
habitat analyses. Data were collected by four observers who
independently and repeatedly sampled a series of sites in an
oak-maple forest over a five week period. Data were analyzed
through ANOVA, principal component and discriminant function
analyses (p < 0.01). Results indicate observers
significantly differed in their measurements of 18 out of 20
vegetative variables (e.g. percent canopy cover, ground
cover, etc.). Gotfryd and Hansell (1985) suggest that " Low
variability is no guarantee of accuracy, and high
variability does not necessarily imply that the data are, on
average, inaccurate."
Block et al. (1987) studied observer variability when
estimating vegetation characteristics. Three observers
visually estimated vegetation characteristics in 75 plots
and then these same vegetation characteristics were measured
using established quantitative techniques. Prior to ocular
estimates in any plot, observers discussed estimation
techniques in an attempt to standardize the group methods.
Two-way ANOVA was used to compare ocular estimates of the
vegetation by the observers (p < 0.05). Results showed that
the three observers differed significantly for 31 of the 49
variables estimated. In addition, comparisons of observer

11
estimates and actual measurements further demonstrated
differences among observer estimation. For example, all
observers underestimated tree diameters, tree and shrub
numbers and canopy cover and overestimated tree heights and
percent ground cover. Block et al. (1987) concluded that not
only are there significant difference among estimates of
different observers but also significant differences occur
between observer's estimates and actual measurements for
many variables.
Species Density/Richness and Habitat Quality
HSI models are designed for use in natural resource
planning and environmental impact assessment studies. These
models determine habitat quality but as Schamberger dnd
O'Neil (1984) state "They are not models of carrying
capacity because not all factors that influence animal
abundance are included." Habitat is not the sole factor
determining species abundance, other factors such as
disease, mortality, predation, weather and competition are
all a function of carrying capacity and most habitat models
address few of these factors (Schamberger and O'Neil, 1984).
Therefore, in studying HSI models, it must be acknowledged
that they are not designed as carrying capacity models or
comprehensive population level predictors. However, HSIs do
provide a 0 - 1.0 index of habitat suitability and an HSI
value of 0.9 should indicate better habitat than an HSI of

12
0.1 and therefore should represent a greater "potential"
carrying capacity (Schamberger and O'Neil, 1984).
Prior to actual studies involving HSI model validation,
studies were conducted relating species abundance to
specific habitat characteristics. Sturman's (1967) study of
two chickadee species found, using a multiple regression
analysis, that the abundance of the Chestnut-backed
chickadee Parus rufescens and the Black-capped chickadee
Parus atricapillus was highly correlated to fourteen
different vegetative characteristics (R2 values ranged from
0.959 to 0.297). The Chestnut-backed chickadee abundance was
highly correlated with percent upper canopy volume which is
coniferous (R2 = 0.826) and the average height of upper
story conifers (R2 = 0.918). These two variables accounted
for more than 90 percent of the variability in observed
abundances. For the Black-capped chickadee, canopy volume
of: trees (R2 = 0.796), bushes (R2 = 0.974) and middle-story
trees (R2 = 0.921), together accurately predict it's
abundance. Regression of these three variables account for
greater than 90 percent of the variability in this species
abundance.
Anderson and Shugart (1973) used univariate ANOVA to
test for differences in abundance categories (i.e. rare,
common, etc.) of various species of birds with respect to 28
habitat characteristics (i.e. number of saplings, foliage
biomass, etc.). Differences in habitat preferences within
major bird families were apparent from this analysis (a

13
levels = 0.001, 0.01 and 0.05). Discriminant function
analysis was then used to order habitat variables according
to their strength in separating bird abundance categories.
Results indicated that certain bird species were distributed
according to specific habitat characteristics (a = 0.05).
Clawson et al. (1984) trapped various species of
reptile in an attempt to relate their abundance to certain
habitat characteristics (i.e.: % ground cover, litter depth
etc.). Through multiple regression analysis it was
determined that predictive models for abundance of the
following 4 species based upon vegetative characteristics
could be developed:
1) Eastern Yellowbelly Racer - Oldfield habitat, n =
21, R2 = 0.68, p = 0.05, vegetative characteristics and
associated probability a value for F = percent canopy
closure (0.0001), number of woody species > 1 m (0.0009),
multiflora rose clumps/ha (0.0006), herbaceous vegetation
height (0.0243);
2) Prairie Ringneck Snake - Oldfield habitat, n = 21,
R' = 0.30, p = 0.05, vegetative characteristics and
associated probability a value for F = season (summer)
(0.0099), herbaceous vegetation height (0.0092);
3) Five-lined Skink - Upland Forest habitat, n = 18, R2
= 0.49, p = 0.05, vegetative characteristics and associated
a value for F = percent ground cover in forbs (0.0025),
herbaceous vegetation height (0.0036);
4) Broadhead Skink - Upland Forest habitat, n = 18, R2

14
= 0.67, p = 0.05, vegetative characteristics and associated
probability a value for F = season (summer) (0.0004),
understory closure (0.0293).
Van Horne (1983) points out several situations where
habitat quality for a species is not directly correlated
with the density of a species. Reasons cited for the
breakdown in the density-habitat quality relationship are:
(1) "there may be multi-annual variability in local
population densities that reflect small scale variability in
the food source, in predator populations, or in abiotic
environmental factors."; and (2) " .... during years when
species' densities are high, animals may achieve very high
densities in low-quality habitats where survival and
production are low because offspring have been forced into
these habitats by intra-specific competition." Van Horne
(1983) suggests that density may simply reflect conditions
of the recent past or temporary present and not long-term
habitat quality. He adds that a measure of habitat quality
should contain certain components of not only density but
also offspring production and survival.
Larson and Bock (1986) measured habitat variables on
small plots centered on individual birds (bird-centered
analysis or BCA) of four different species. They then
compared these results with data from randomly situated
plots. Bird abundance estimates were determined to be
correlated with several habitat characteristics (e.g.
percent shrub cover, percent bare ground, percent grass

15
cover, maximum height of vegetation, maximum grass/forb
height, maximum shrub height, and percent Sagebrush) for
some of the species studied (p < 0.01). In addition, BCA was
able to identify more significant habitat associations for
specific species than the random plot analysis.
Studies specifically related to HSI models began to
appear in the early 1980's. Many studies were involved with
the development of accurate models, while other research has
been conducted into post facto model verification.
Laymon and Barrett (1986) investigated the pitfalls
that arose in the development and validation of three
different HSI models. The models they tested were the
Spotted Owl, Strix occidentalis (Laymon et al., 1985),
Marten, Martes americana (Spencer et al., 1983) and Douglas'
Squirrel, Tamiasciu.rus douglasii (Laymon and Barrett, 1986).
The Spotted Owl model's ability to predict occupied
Spotted Owl habitat in Sierra Nevada, Ca. was tested by
comparing HSI values from 70 sites where owls were
definitely known to occur (x = 0.53) with HSI values
measured at 70 randomly located sites (x = 0.31), (t = 7.53,
CV of t = 1.96, p < 0.001). In a second test, a field survey
using vocal imitations of Spotted Owl calls was conducted at
56 sites, 14 in each of the following four categories; (1)
unsuitable - (HSI = 0 - 0.23), (2) low - (HSI = 0.24 -
0.47), (3) moderate - (HSI = 0.48 - 0.64), (4) high - (HSI =
0.65 - 1.0). Owls responded at none of the unsuitable sites,
14 percent of the low sites, 36 percent of

16
the moderate sites and 86 percent of the high sites. These
results indicate an increasing trend in owl response with an
increase in HSI value (z = 6.95, CV of z = 1.96, p < 0.001).
For the Marten model, HSIs were calculated for 8 sample
sites and Marten visitation at these sites was calculated by
use of track plates. The results were tested for a
significant trend with a Kruskall-Wallis Trend test for
nonparametric data. The 8 sites were ranked as to
suitability with three of the sites rated "marginal" (HSI =
0.1 - 0.3), one site rated "suitable" (HSI = 0.4 - 0.6), and
four rated "optimal" (HSI = 0.7 - 1.0). Mean Marten
visitation rates increased with increasing habitat rating
but the trend was not found to be statistically significant
(X2 for a linear trend, 1 df = 3.66, p = 0.001). Laymon and
Barrett (1986) suggest a larger sampling effort might--yield
statistically significant findings.
For the Douglas' Squirrel model, HSI values were
computed for each of 210 (4 ha) sample sites. Squirrel
density was calculated from line-transect censuses. Simple
and multiple regression analyses were used to relate
individual variables and total HSI values to squirrel
density. HSI values were transformed into squirrel-density
estimates by assuming that the highest density recorded in
the study for a sample site was equal to an HSI of 1.0.
Results indicated no statistically significant relationship
between HSI variables and squirrel density.
The investigators suggest the time of year they sampled

17
squirrel density (late summer) may have been inappropriate
due to young dispersing into less suitable habitats. Laymon
and Barrett (1986) comment that "correlations between
squirrel density and habitat variables might be more
apparent if sampling occurred during the spring...".
Cook and Irwin (1985) field tested the Pronghorn,
Antilocapra americana HSI model (Allen & Armbruster, 1982)
by correlating model outputs with estimates of wintering
Pronghorn densities which they assumed to reflect habitat
quality. Using simple linear regression they regressed HSI
values, against corresponding Pronghorn density estimates.
Results indicate that the model explains 50 percent of the
variation in Pronghorn density (p < 0.001). They conclude
that despite Van Horne's (1983) caution that density may
sometimes be a misleading indicator of habitat quality, the
Pronghorn HSI model produces ratings of habitat.quality
which correlate well with population indicators of habitat
quality. They point out that the model should not be used to
predict actual Pronghorn numbers, but that habitat potential
for Pronghorns can be described.
Schroeder (1990) examined the relationship between
Black-capped Chickadee HSI model values (Schroeder, 1983)
and Black-capped chickadee population densities through
Least Absolute Deviation Regression analysis. The Null
hypothesis for this test was that regression of HSI values
on estimated Black-capped chickadee population densities
would produce a model with a slope significantly different

18
from zero but not different from the slope of a model
predicting ideal maximum population densities. An ideal
maximum population density model was derived using
historical data gathered in a previous Black-capped
chickadee survey. Results indicated that the HSI model
closely approximated ideal maximum population density when
HSI values are at a maximum. He concludes that the HSI model
considers only a subset of a variety of factors which may
influence population density. He suggests that "The HSI is
best viewed as an index of habitat-imposed limitations on a
population. For example, an HSI of 1.0 would indicate no
habitat-related limitations exist, whereas decreasing HSI
values would indicate increasingly significant habitat
limitations" (Schroeder, 1990).

CHAPTER 3
METHODS AND PROCEDURES
The following section contains descriptions of the
methods and procedures used in this study. The first
subsection describes the study area and the specific
habitats analyzed. The second subsection deals with species
selection and Habitat Suitability models chosen. Subsections
three and four describe the procedures used in the HEP field
evaluations and bird censuses. Species Guilds and their
utility in this study are described in subsection five. The
final subsection discusses statistical analyses used to
address the proposed hypotheses.
Study Area and Habitat Descriptions
Ray Roberts Lake is located largely in Denton County,
Texas, along the Elm Fork of the Trinity River, 30 miles
upstream from Lewisville Dam and approximately 10 miles
north of the City of Denton (Fig.1). The area considered for
both the 1987 and 1990 Habitat Evaluation Procedure (HEP)
studies was within the U.S. Army Corps of Engineers Project
Area boundaries for Ray Roberts Lake totalling 48,347 acres.
A Green Belt area below the dam totalling 1,600 acres was
also included in the evaluation. Six habitat types were
19

20
selected for analysis in the initial HEP study in 1987 and
are defined in the Ray Roberts Lake Pre-impoundment
Environmental Study (IAS, 1988) as follows:
Upland Forest
Areas of sandy soil with abundant rocks noticeable on
slopes. Tree densities range from 200 to 500 trees per acre
with canopy heights of 20 to 26 feet. Three species of tree
are co-dominant; Post Oak, Quercus stellata, Blackjack Oak,
Quercus marilandica and Cedar Elm, Ulmus crassifolia.
Undergrowth is generally low with herbs concentrated in
small clearings between trees.
Bottomland Forest
These areas were characterized by taller trees (canopy
heights range from 25 to 40 feet) and a greater herbaceous
component than that of Upland Forests. Tree species
diversity was high in these areas. Shrubs cover 20 to 50
percent of the ground between trees.
Shrubland
Areas with tree densities intermediate between typical
Oldfields and woodlands are designated as Shrubland.

21
Tree densities range from 20 to 120 per acre. Shrubs provide
2 to 20 percent of ground cover.
Pasture
This habitat is defined as a field either clearly
maintained for cattle or one in which evidence of recent
heavy grazing is apparent. Low plant species diversity and
trampled vegetation are signs of heavy grazing. Pastures are
typically treeless and have few shrubs (average < 2 per
acre).
Oldfield
These areas are distinguished from Shrublands by'lower
shrub and tree densities and from Pasture by their lack of
indications of recent heavy grazing. Tree densities range
from 0 to 16 trees per acre and shrub density from 0 to 300
per acre. Agricultural-Cropland acreage are included under
the Oldfield habitat heading for the HEP.
Riverine/Lacustrine
This group includes all Lotic and Lentic habitats such
as streams, rivers, ponds and lakes.
In the 1990 HEP study all the above habitat types were

22
again considered, along the vegetational transects
established in 1987 (Fig. 2), with the exception of Pasture
habitat which was lost due to inundation by the lake.
Species Selection and Habitat Suitability Models
The habitat types chosen within the study area were
evaluated to determine their ability to fulfill "life
requisites" of select species. Two "indicator species" were
selected for each habitat type based on a species inventory
compiled for the area by the Biology Department, University
of North Texas (Table 1). Certain birds were selected as
"indicator species" because they were considered
representative of generalized communities within the
ecoregion. Although bird, mammal and reptile species were
all considered in the Ray Roberts Lake Environmental Report
HEP studies, only avian species will be considered in this
study due to a lack of sufficient mammal and reptile census
data.
Habitat Suitability Index (HSI) models for all
indicator species, with the exception of the Downy
Woodpecker model, were obtained from the U.S. Fish and
Wildlife Service Terrestrial Habitat Evaluation Criterion
Blue Handbook (Ecoregions 2512 & 2320, 1980). The Downy
Woodpecker model was obtained from the U.S. Fish and
Wildlife Service Habitat Suitability Index Models: Downy
Woodpecker (Schroeder, 1982).

23
CL~
00
'00,l 0
a , 0.
00
00
- 0 ~ 00. U13
6 ~ 0
0> r4zA
- -4%
E>Ee
a -4
c IJ
A -.
S-- -
CL

24
Table 1. Indicator Species and Models used for the HEPAnalysis at Ray Roberts Lake (IAS, 1988).
Habitat Species
Riverine/lacustrine(RL)
Pasture (P)
Oldfield (OF)
Shrubland (SH)
Bottomland Forest(BF)
Upland Forest (UF)
Belted kingfisher Megaceryle alcyonGreen heron Butorides virescens
Eastern meadowlark Sturnella magnaRacer Coluber constrictor
Scissor-tailed flycatcherMuscivora forficata
Eastern cottontailSylvilagus floridanos
Eastern Woodrat Neotoma floridanaDowny woodpecker Picoides pubescens
Barred owl Strix variaRaccoon Procyon lotor
Fox squirrel Sciurus nigerCarolina chickadeeParus carolinensis

25
Table 2 lists the model variables. Appendix A contains the
model equations.
HEP Field Evaluations
Field evaluations for the HEP were conducted in early
summer 1987 and again in late summer of 1990. In 1987 six
habitats were selected for analysis along the vegetational
transects (Fig. 2) with two evaluation species analyzed per
habitat (Table 1). The evaluations in 1987 were performed at
fifteen random sampling sites per species, in each habitat,
for a total of 180 sites. These evaluations were made by one
individual. In 1990 there were only five habitat types along
the vegetational transects due to loss of Pasture habitat by
lake inundation. For each of the five habitat types several
locations were studied, for example, Bottomland Forest was
analyzed along transect GH and also transect EF. At each
site a team of five individuals performed three evaluations
each for a total of 360 measurements during the 1990
analysis. These data were then combined to give an average
Life Requisite HSI value for each indicator species.
Appendix B contains the HSI values measured.
In 1987 several "impact assessment areas" were also
chosen and evaluated as a basis for monitoring successional
changes and indirect effects of impoundment. One 20 acre
assessment area per habitat type was selected from areas
above the Flood Pool level where no plans had been made for

26
4141
04)A.Q,
- (J
N
'-4N>>
AL~ca
uxs
satso
XO
jITXX*1TTOuVnc
uiqSsj~ui
uoIa
N 4
- >>N
--fN
0 0%6w
ifw 6wi~c
40dpI0w dv 1 1to4Pd
ip i2
At m
- I9 , .
L s
1* !u.
% r- >f> c
m0)
04"1
00
4P
C40

27
development. These areas were again evaluated in 1990 to
determine if any indirect effects from impoundment had
occurred (Fig. 3). In 1987 at each "impact area" eight
sample sites were randomly chosen and evaluated by a single
individual. In 1990 a team of five individuals made three
evaluations each per "impact area" for a total of fifteen
sample sites per habitat. Appendix C contains the "impact
area" HSI values measured.
Bird Censuses
During the 1987 pre-impoundment and 1990 post-
impoundment environmental studies seasonal bird censuses
were conducted along the vegetational transects (Fig. 2) and
through the habitat types considered in the HEP analyses.
All censuses were conducted by experienced personnel
familiar with the birds of North Central Texas. The strip
transect method (Merikallio, 1958) was considered most
effective for censusing avian populations along the
transects " because of its seasonal versatility and
appropriateness for both flocking and nonflocking species,
and species such as swifts and swallows that cruise above
vegetation" (Emlen, 1971). More elaborate techniques such as
mist-netting were not utilized due to fiscal and personnel
limitations. The same personnel, with few exceptions, were
used to conduct all surveys. Each census was conducted on
foot beginning at sunrise and continuing for a maximum of 3

28
0000004.0--- .1 t % sC j 0u 0 we 0 er-. . o C
E w tor. %0 4 %~- 0Lfl0s0e0-O~ O 0 O~ O~ O0
1. 0 r. N 4Io C Q o qco aM - t% L \
0- 0 co00000c c
-na, e~n -s,."o- o4~a a,
-ocCCLa * .- 4,-
-f - a4,1a, c
0 IA)
fo- 4 )oco
a 0)
- - r. o o to LL.
-4.- 4 - 0).I- - -0-0..-.-:.a-4J)
-x -L- ) .A c
*0-W --U- -- - -- - - -C -.* 0 4, c:
c
n oen
atv
V0
3 CLL t
yZ _
0)o-
C00
CA,-
-a, -U.

29
hours to coincide with peak avian activity (IAS, 1988).
Transects were censused on consecutive days and/or when
weather conditions were similar to avoid detectability bias
due to weather conditions. For each habitat type along a
transect a strip width of approximately 200 feet was
established. Personnel walked down the center of this 200
foot wide strip, parallel to the vegetational transect, and
identified birds via visual and auditory cues obtained no
further than 100 feet away on either side. Density estimates
of each species per habitat type and transect were
calculated by summing the number of each species counted by
the observers in their respective survey areas (i.e. pooling
all personnel transect-strip data)(IAS, 1988). Appendix D
lists all avian census counts for indicator and guild
species by habitat type and vegetational transect section.
Guild Information
In order to expand the available Ray Roberts Lake bird
census data to be used in this study, the concept of guild
theory was explored. Root (1967) defined a guild as "... a
group of species that exploit the same class of
environmental resources in a similar way." The guild concept
has been recommended for extrapolation of results from
evaluation species to larger groupings (Severinghaus, 1981).
The results of three different approaches to
structuring guilds were incorporated into this study in

30
order to expand the available bird census data beyond single
indicator species. For example, the Carolina Chickadee was
the HEP indicator species for Upland Forest habitat,
however, using results of several guild analyses, the Downy
Woodpecker, Tufted Titmouse and Cedar Waxwing species could
all be grouped with the Carolina Chickadee in a common guild
(their census data were included in the analysis).
Short and Burnham (1982) guild "block concept" results
were used in this study. This approach concentrates on
structural features of vegetation and its' relation to
feeding and breeding criteria for birds. They described a
two-dimensional species-habitat matrix by combining
reproductive and feeding loci as the x and y axes
respectively. A specific feeding/reproductive criteria for a
species is therefore defined within the matrix by an (x,y)
coordinate. This coordinate represents a matrix cell that
best describes the habitat needs of that species. Basic
summary statistics were computed for all species and cluster
analysis routines were run to provide species groupings or
guilds. -This information was then arranged in a phenogram
form. Table 3 contains a list of bird species from the
results of this study which have been guilded with the
chosen HEP indicator species.
Results of Severinghaus (1981) guild analysis were also
used in this study. He defined guilds based on factors
attributable to feeding habits. Initially species were
separated based upon feeding strategies (ie; herbivorous

31
Table 3. Avian Species Guilded with HEP Indicator Species asa Result of Various Guild Studies.
Guild Study Habitat Type/Indicator Species Guild Species
Short & Burnham Pasture/Eastern meadowlark Killdeer(1983)
Dickcissel
Savannahsparrow
Severinghaus Red-winged(1981)
blackbird
Payne & Long Scissor-(1986)
tailedflycatcher
Short & Burnham Upland Forest/Carolina Common(1983) chickadee flicker
Downywoodpecker
Severinghaus Tufted(1981)
titmouse
Payne & Long Cedar waxwing(1986)
Cooper's hawkPayne & Long Bottomland Forest/-Cooper's(1986) Barred Owl hawk
Payne & Long Oldfield/Scissor-tailed Eastern(1986) flycatcher meadowlark
Payne & Long Riverine/lacustrine Wood duck(1986) Green heron, Belted kingfisher

32
versus insectivorous etc.) and then further subdivided
according to feeding method, specific food requirements
etc.. Results were displayed in a dendogram format (Fig. 4).
Table 3 lists species used in this study which were derived
from Severinghaus's (1981) dendogram.
Payne and Long (1986) suggested that analysis of
various HSI models for "shared" variables (i.e.; variables
in common in 2 or more different models) provides insight
into how species should be allocated into guilds. By
examining matrices of "indicator species versus HSI model
variables", guilds of bird species were developed according
to shared model variables. If species share 3 or more
variables (average models contain 5 variables) they were
grouped together in a guild. It was noted in developing
guilds in this manner that "the directness with which
inferences can be drawn from one model to another depends
largely on the number of shared model variables and on the
similarity of the variable curves" (Roberts and O'Neil,
1985). Table 3 contains the species used in this study as a
result of this guilding technique.
Statistical Analyses
To test the first hypothesis, that there is no
significant difference in HSI variable values between
individuals in a survey party, a hierarchical cluster
analysis with subsequent bootstrapping was run using the

wil
A
-J-oW.-
3.Is -
a .,
b. 00I e L0
-- i;-~
.j rI
0 J
44n
+ z00
4 10-
ID a
0 0 0
I IL
0
a w
0-
0~
0-
4440
33
I
iIl10a
I i- 3
7 I~I
I
I01~i0
IL
V)4it
r-4
0
4)
4)
cn
0:)
'4J
c*
0
41
I
F/

34
SIGTREE program (Nemec, 1991). SIGTREE is a nonparametric
method which uses a bootstrap technique to identify
statistically significant clusters (Diaconis & Efron, 1983;
Efron, 1979; Nemec & Brinkhurst, 1988). Bootstrapping infers
the variability in an unknown distribution from which the
data were drawn by resampling data from replicate samples,
with replacement, to create a series of bootstrap samples of
the same size as the original data (Kennedy, et. al., 1993).
Each of these samples is analyzed using the Percent
Similarity (PS) index discussed by Bray and Curtis (1957);
PS, = 2(Z min (Aij,Aik))/(Aij + Aik)
For this thesis; the summations are over all mean HSI
variable values (i). A. and Aik represent the mean HSI
variable values in samples j and k. A Bray-Curtis (PS) value
of 0 would indicate that the mean HSI variable values were
entirely different between two different observers; a value
of one suggests complete similarity. Cluster analysis is
performed on the calculated PS values using the unweighted
pair-group method with arithmetic averages (Sneath & Sokal,
1973).
The bootstrap approach then tests for significant
differences between clusters. The null hypothesis (H") is
that two clusters are sufficiently linked that they can be
considered to represent a single common community.
Accordingly, the percent similarity of each bootstrapped

35
community should produce a linkage level comparable (or
higher) than the linkage level in question. If only a small
proportion of the bootstrapped values (i.e., 5% when the
significance level is at 5%) correspond to the original
similarity level, then the two clusters would be considered
dissimilar and the null hypothesis would be rejected.
The linkage between clusters can be shown in a cluster
dendogram. A stacked bar graph summarizes the mean HSI
variable values for the five observers.
For the second hypothesis, that there is no significant
difference in mean HSI values at impact assessment sites
between sampling years, the data were first checked for
normality and homoscedasticity using the F-test (Zar, 1984).
A nonparametric Mann-Whitney test (Zar, 1984) was then used
to test the hypothesis.
A nonparametric Spearman's rank correlation (Zar, 1984)
was used to test the third hypothesis, that for a particular
habitat type there is no correlation between Habitat Units
and Indicator Species density. The Habitat Unit being the
independent variable (X) and Indicator Species density being
the dependent variable (Y).
The fourth hypothesis, that for a particular habitat
type there is no correlation between Habitat Units (X) and
species diversity (Y) was tested using a nonparametric
Spearman's rank correlation (Zar, 1984). Species diversity
was determined by the Shannon-Wiener and Brillouin's index.

CHAPTER 4
RESULTS
Four hypotheses were proposed in the Introduction to
this thesis. A variety of statistical tests were applied to
the data collected at Ray Roberts Lake in 1987 and 1990 to
evaluate these hypotheses. The results of these analyses are
presented in the following section.
Hypothesis 1:
There is no significant difference in Habitat
Suitability Index variable values between individuals in a
survey party. SIGTREE developed by Nemec & Brinkhursi'(1988)
was a successful method for analyzing observer variability
within a group when comparing HSI values for site variables.
Twenty-four transect habitat sites and twelve impact sites
were assessed by an evaluation team of five individuals in
1990. Results indicate that out of the thirty-six sites
analyzed, only five sites showed significant differences in
HSI variable values between the members in the survey party
(Appendix E).
In interpreting the following Bray-Curtis Cluster
analysis dendograms, it should be noted that the five
observers were listed alphabetically and then numbered
sequentially for convenience in data analysis. The order in
36

37
which the observer number falls out in the dendograms is
determined by the cluster analysis. Observers 1 and 3 were
most "experienced" in HEP techniques at the time of this
study and observer 2 the least experienced. Observers 4 and
5 had moderate experience in HEP. -
Three of the five sites where differences were
indicated were Water habitats (Green Heron model). At the
Impact Site Water habitat (Figure 5, Appendix E) observers 1
and 3 reported average HSI values of 0.81 and 0.71 for
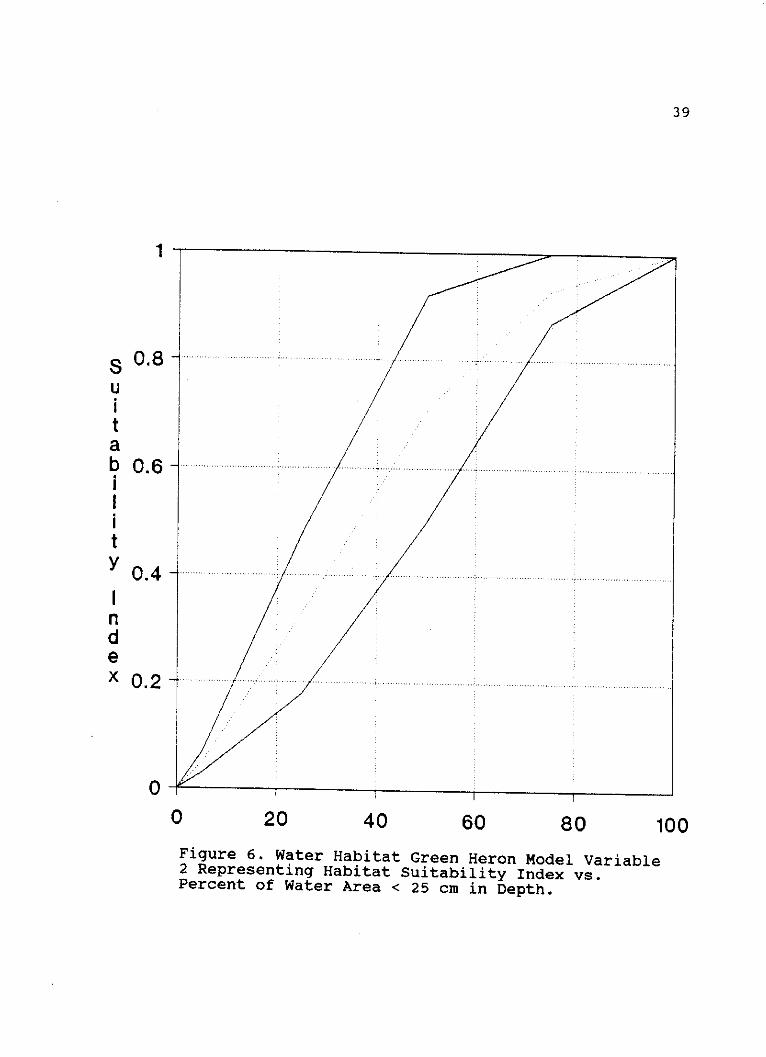
Variable 2 ("Percent of water area less than 25 cm deep").
These HSI values represent estimations of percent water area
less than 25 cm deep to be 62% and 51% respectively.
Observers 2, 4 and 5 had average HSI values of 0.06, 0.27
and 0.10 representing percent water area less than 25 cm
deep to be 6%, 23% and 10% respectively (Figure 6, Appendix
C).
For the Water Habitat on transect EF (Figure 7,
Appendix E) observers 1 and 3 again gave higher average HSI
values than the rest of the team for Variable 2. These
observers scored average HSI values of 0.99 and 0.98
representing 91% and 92% water area less than 25 cm deep.
Observers 2, 4 and 5 had average HSI values of 0.01, 0.08
and 0.01 representing 8% or less water area less than 25 cm
deep (Figure 6, Appendix B).
For the Water Habitat on GH transect (Figure 8,
Appendix E) observer 1 gave an average HSI value of 0.85 for
Variable 1 ("Aquatic substrate composition in littoral

38
Impact Site Water Habitat (Green Heron Model)
5
3
4Observer
ijjSbeve
Variable 1: Aquatic substrate composition in littoral zoneVariable 2: Percent of water area less than 25 cm deepVariable 3: Percent emergent herbaceous canopy cover in littoral zoneVariable 4: Percent water surface covered by trees, logs, woody vegetationVariable 5: Water regimeVariable 6: Water currentVariable 7: Distance to deciduous forested wetland or shrub wetlandFigure 5. Bray-Curtis Cluster Analysis of Mean HabitatSuitability Index Values for Five Observers at ImpactSite Water Habitat (Green Heron model).
2
Uw
E3 vM W*7ol MaWnOo Vftft*3
ClU aWb13M* b
rdty0
0.2
0.4
06
0.8
S
4
.3
0
------------------------------------------------------------
- - - - - - - - - - - - - - - - - - - - - - - - - - -
1
3 I
I
39
1-
So. 8
U
tabO0.6
t y o..4 ..... . ........ .....................
ndex 0 .2 . . . . . .
0
0 20 40 60 80 100Figure 6. Water Habitat Green Heron Model Variable2 Representing Habitat Suitability Index vs.Percent of Water Area < 25 cm in Depth.

Transect EF Water (Green Heron Model)Similar
07
0 2
0 4
0 6
0.8
5
4
0
4Obsever
.1
4 2Observer
Variable 1: Aquatic substrate composition in littoral zoneVariable 2: Percent of water area less than 25 cm deepVariable 3: Percent herbaceous canopy cover in littoral zoneVariable 4: Percent of water surface covered by logs, trees, woody vegetationVariable 5: Water regimeVariable 6: Water currentVariable 7: Distance to deciduous forested wetland or shrub wetland
Figure 7. Bray-Curtis Cluster Analysis of Mean HabitatSuitability Index Values for Five Observers at TransectEF water Habitat (Green Heron Model).
E varmbe 5
WOruWs3o3 VarWabl2a vowi
3
ity
40
I rn
I
I I f 17 T

41
Transect GH Water (Green Heron Model)itySimilar
0
0.2
0. 4
0 6 ?
0.8 t
4
3
2
C
0
5Ob serv er
oa
owI
3
4
U
3
2
2
5Observer
2
Variable 1: Aquatic substrate composition in littoral zoneVariable 2: Percent of water area less than 25 cm deepVariable 3: Percent herbaceous canopy cover in littoral zoneVariable 4: Percent of water surface covered by logs, trees, woody vegetationVariable 5: Water regimeVariable 6: Water currentVariable 7: Distance to deciduous forested wetland or shrub wetland
Figure 8. Bray-Curtis Cluster Analysis of Mean HabitatSuitability Index Values for Five Observers at TransectGH Water Habitat (Green Heron Model).
r __K4 1
-----------------------------------------
ON,
1

42
zone"), whereas the rest of the team gave average HSI values
of 0.20. This higher score represents a difference in
opinion as to what the "aquatic substrate composition" was,
with observer 1 judging the composition to be "muddy", as
opposed to the rest of the team judging the composition to
be "rocky" (Figure 9, Appendix B).
At the Impact Site water habitat (Figure 5, Appendix E)
observer 3 differed from the group in assessment of Variable
4 ("percent of water surface covered by logs, trees, or
woody vegetation"). This observer reported an average HSI
value of 0.48 representing "coverage" to be approximately
15%, whereas, observers 1, 2, 4 and 5 gave average HSI
values of 0.12 or less indicating "coverage" to be 2% or
less (Figure 10, Appendix C).
Another habitat where differences between the team's
HSI values varied was the Shrubland on AB transect (Eastern
Woodrat model) (Figure 11, Appendix E). Observer 2 had an
average HSI value of 0.0 for Variable 3 ("number of refuge
sites per 0.4 ha") indicating an average of one or less
refuges per 0.4 ha. All other observers averaged at least
four refuge sites per 0.4 ha with HSI values of 0.73, 0.65,
0.97 and 0.97 respectively (Figure 12, Appendix B).
The final site where variability in measurements within
the group were detected was the Oldfield habitat on transect
EF (Eastern Cottontail model) (Figure 13, Appendix E).
For Variable 4 ("percent shrub crown cover") observers 1 and
3 had average HSI values of 0.16 and 0.24 representing

1
0.8
0.6K.
1 F~ TYTT~rt~ 9
U........................0.4
0.2
A B
Figure 9. Water Habitat Green Heron Model Variable1 Representing Habitat Suitability Index vs. AquaticSubstrate Composition in the Littoral Zone.A = Muddy, B = Sandy, C = Rocky.
43
SU
b
ta
bd
.I...................... ........ ......
I K x -, K K xTTTTTrr
O
C

44
1
S
t
b
ty
de
- T -
0.8 f
0.2-
o
0 20 40 60 80 100
Figure 10. Water Habitat Green Heron Model Variable 4Representing Habitat Suitability Index vs. Percent of WaterSurface Covered by Logs, Trees, or Woody Vegetation.
I
f-
.......... ....................
................................ ........... ...................................... .................. ..................... .......................
............. ......... . ......... ......................... ............................ .................... .. ............. ........... ...........................
........... ....... ....... ......... ................................. ............................................................. ........... ... . . . . . . . . . . . ...................
............. ........ ................................................ ......................... .............................................................. .......... .. .............
0.6

45
Transect AB Shrubland (E Woodrat Model)S&rndarit y0
0.2
0.4
0.6
0.8
Ob server
2
15
S
0-
--------------------- ---- ------
----- ----
-.....
2.. ......
o 'MWW 3* tim 2
Observ or
Variable 1: Percent shrub crown coverVariable 2: Percent herbaceous canopy coverVariable 3: Number of refuge sites per 0.4 ha
Figure 11. Bray-Curtis Cluster Analysis of Mean HabitatSuitability Index Values for Five Observers atTransect AB Shrubland Habitat (E. Woodrat Model).
I
a

46
1
S
t
b
ty
dex
III1l 1
0.8-
0.6-
0.4
0
0 2 4Figure 12. Shrubland Habitat Eastern Woodrat Model Variable 3Representing Habitat Suitability Index vs. Number of RefugeSites per 0.4 Ha.
I
...................................... .....................................
................ ................. .................. .................. ............. ....................................................
................. ......... ......... .......... ............................... ................................................ ..................
.................................... .......... ....................................... . . . . . . . . . . .. . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . .. . .. . . . . . . . . . . . . .. .. . . . . . . . .
. . . . . . . . .. .. . . . . . . . . ........ . . . . . . .. . . . . . . . . . . . .. . . . . . . . .. . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . .. .. . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. .. ..O.2
6 8 10

Transact EF Oldfield (E CottontaiI)
5 4
Observer3
~Eilllil~L~------~-----
4Observer
Variable 1:Variable 4:Variable 5:
Percent herbaceous canopy coverPercent shrub crown coverNumber of refuge sites per 0.4 ha
Figure 13. Bray-Curtis Cluster Analysis of Mean HabitatSuitability Index Values for Five Observers at TransectEF Oldfield Habitat (E. Cottontail Model).
ity
47
0
0.2
0.4
0.6
0.8
1.4
2
12 -
LI r
---------
c 0.6 7BTFF f0.4 V
0.2
02
r i _
I
I rv
1
1

48
"coverage" of 10% and 15%, whereas, observers 2, 4 and 5 had
average HSI values of 0.0, 0.02 and 0.0 representing
"coverage" of 0%, 1% and 0% (Figure 14, Appendix B). For
Variable 5 ("number of refuge sites per 0.4 ha") observer 4
had an average HSI value of 0.51 representing an average of
2.5 refuges per 0.4 ha. Observer 5 had an average HSI value
of 0.06 representing less than 1 refuge per 0.4 ha. All
other observers estimated between 1 and 2 refuges per 0.4 ha
(Figure 15, Appendix B).
Hypothesis 2:
There is no significant difference in mean Habitat
Suitability Index values at impact assessment sites between
sampling years.
Data from the impact sites in 1987 were collected by
one individual taking many replicate samples (8 data points
per variable). Data collected from these same sites in 1990
were collected by five individuals, each taking three data
points, for a total of fifteen data points per variable. In
order to determine if change or no change in the HSIs at the
impact sites is due to temporal variability, or due to
different sampling methods, it was first necessary to check
these data for normality, inequality of variance and
determine if sample size differences are a problem. Since
the data gathered in 1987 were gathered by one person, it
was assumed that there was consistency in variable
measurements at each impact site by this individual.

49
1-
SU
bO0.6
tyou
d
0
0 20 40 60 80 100Figure 14. Oldfield Habitat (Eastern CottontailModel) Variable 4 Representing Habitat SuitabilityIndex vs. Percent Shrub Crown Cover.

50
1-
SU
t
ty
dexo0.2
0 2 4 6 8
Figure 15. Oldfield Habitat Eastern Cottontail Model Variable5 Representing Habitat Suitability Index vs. Number of RefugeSites per 0.4 Ha.

51
Measurement consistency cannot be assumed for data gathered
by five individuals in 1990. Therefore, these data were
analyzed to check if all measurements taken by the group
could be considered to be of a "common community" by running
SIGTREE (Nemec, 1991). Results indicated group consistency
on all HSI values at all impact sites except Water habitats.
These sites were omitted from further analysis. HSI values
determined for each variable, by the five group members in
1990, at each impact site were then grouped together and
considered as one data set. HSI values for all variables, at
every impact site, in both study years were checked for
normality and then checked for equality of variances
(homoscedasticity between 1987 and 1990 data) using the F-
test (Zar, 1984).
Results indicate nonnormality within the data sets from
both study years. Results of the F-test for all impact
sites, except Shrubland (Downy Woodpecker model), showed
heteroscedasticity. It should be noted that all variance
homogeneity tests are adversely affected by nonnormality.
Although ANOVA is a robust test and can stand some deviation
from underlying assumptions the fact these data sets have
unequal sample sizes in addition to nonnormality and
probable heteroscedasticity suggests ANOVA should not be
used (Zar, 1984). Instead a nonparametric Mann-Whitney test:
U = nin 2 + n1(n1 + 1)/2 - Rl,

52
where; U = Mann-Whitney test statistic,
Rt= sum of the ranks for sample 1
n= number of observations in sample i
(Zar, 1984) was used to analyze mean HSI values for
variables, in both years, at each impact site. Results
indicate that for all impact sites, except Upland Forest
(Fox Squirrel model), mean HSI values between test years
were not significantly different.
The Upland Forest (Fox Squirrel model) result was
significant (a = 0.05, UO.05(2)4 4 = 16, U = 16, p = 0.05).
Mean HSI value of 0.11 for Variable 1 (" Percent tree canopy
closure of mast producing trees greater than 25 cm diameter
breast height (dbh)") indicates a collective judgement by
five individuals in 1990 that the tree canopy closure of
mast producing trees greater than 25 cm dbh was
approximately 10%. In 1987 one individual judged this
"canopy closure" to be approximately 35% (mean HSI value of
0.84) (Figure 16, Appendix B). In 1990 the mean HSI value of
0.04 for variable 3 (" Average dbh of overstory trees")
indicates a collective opinion that "overstory trees" were
on average 5.5 inches dbh. In 1987 the mean HSI value of
0.85 indicates an opinion that "average dbh" was 12 inches
(Figure 17, Appendix B).
Hypothesis 3:
For HEP model habitat types there is no correlation

53
1-
SU
t
b0.6.......
t-
0 .4 .......
d
x 0 .2 -
0
0 20 40 60 80 100Figure 16. Upland Forest Habitat (Fox SquirrelModel) Variable 1 Representing HabitatSuitability Index vs. Percent Tree CanopyClosure of Mast Producing Trees > 25 cm dbh.

54
r
0.8
SU
t
b 0.6-
t
0.4
/
1-/
//
/
/ /
in 0
cm 0
5
12.510
2515
37.5Figure 17. Upland Forest Habitat (Fox Squirrel Model)Variable 3 Representing Habitat Suitability Indexvs. Average Diameter at Breast Height (dbh in in.and cm) of Overstory Trees.
y
ndex
2050
1

55
between Habitat Units (HUs) and Indicator Species density.
Due to the nonnormality of the Habitat Units a
nonparametric Spearman's Rank correlation:
r, = 1 - 6 Z d 2/n - n
where; r. = Spearman Rank correlation coefficient
di = difference between X and Y ranks
(Zar, 1984) was used to test this hypothesis. Results
indicate that there is a correlation between HUs and bird
Indicator Species density (rSO.0 5 (2 ,13 = 0.560, r = 0.643, n =
13, 0.05 < p < 0.02, Nonparametric Spearman's Rank), (Table
4, Figure 18).
Hypothesis 4:
For HEP model habitat types there is no correlation
between HUs and avian species diversity.
Species diversity was determined using both the
Shannon-Weiner index and Brillioun's index:
H' = (Nlog N - Enilogni)/N
and H = (log N! - Zlogni!)/N
where; H' = Shannon-Weiner Index
H = Brillioun's index
n= species abundance
N = sum of abundance

56
Table 4. Habitat Units, Indicator Species Density and SpeciesDiversity* for Various Impact Sites and Transect Habitats.
Habitat Habitat Units Density H' H
Upland Forest
6.053.5
439
0.89 0.801.09 0.90
Bottomland Forest
1.42.8
86
0.84 0.621.11 1.00
Shrubland
0.161.6
AB & EF (1987)Impact Site (1990)Impact Site (1987)
20
9.153.04
11.40
516
65
0.66 1.180.70 0.94
1.410.951.07
1.261.220.98
Oldfield
CDEFWater
EF (BK)**EF (GH)**
30.116.7
6680
22
2.61.3
0.57 1.250.49 1.18
0.84 0.620.84 0.62
* Species diversity indexesand Brillouin Index = H.** (BK) = Belted Kingfisher
used: Shannon-Weaver Index = H'
Model, (GH) = Green Heron Model
CDEF
EFGH
ABCD
Pasture

x
x
\x
x
0C,
-0CMJ
--
C
L(U
Cr--
c0J
4ma
-O ..
LO( I C'l) cm J 0
41> 00
004
CIO
> 44
to 1 -H 4)
H 4): :1
0 *4 ciM04 0Ux
S >,-4
U) 4.)0 41
0 .410
4J) inw0 4J 0*
o- 0 r-*4.CE.O > 4 4~
0 C4 0 0-rq ) 4 4.
o 0 c.$4 M0
0 oc00 0 c
4 $4 4J
41()o~4) o -r - -
C(4 -q
C4J 00t'~-4 ~ 4) :s
-0q 0: a (rz H %-. U
57
C13 -* " D a 0 c 0 --+-#-0

58
Due to the nonnormality of the Habitat Units a
nonparametric Spearman's Rank correlation (Zar, 1984) was
used to test this hypothesis.
Results indicate that there is no correlation between
HUs and species diversity as determined by both diversity
indices (Table 4). (Shannon-Wiener index; r(0.05)2,13, = 0.618,
n = 11, r. = -0.08, p > 0.50, Nonparametric Spearman's
Rank). (Brillouin index; r.(O.OS),, = 0.618, n = 11, r. =
0.384, 0.50 > p < 0.20, Nonparametric Spearman's Rank).

CHAPTER 5
CONCLUSIONS
The final section of this thesis contains a review of
the proposed hypotheses, the statistical tests used to
analyze them, and a discussion of the results along with
recommendations for future study.
The first hypothesis proposed in this thesis was; There
is no significant difference in Habitat Suitability Index
(HSI) variable values between individuals in a survey party.
Results indicate that out of thirty-six sites analyzed only
five sites showed significant differences in HSI values
between group members.
Three of these five sites were Water Habitats (Green
Heron Model). For this model most variability within the
survey party's measurements occurred with variables 1, 2 and
4. The first variable: "Aquatic substrate composition in
littoral zone" was judged by one observer to be "muddy"
whereas all other team members judged the "substrate
composition" to be rocky. Possible explanations for this
difference could be either: a) differing interpretation of
the characteristics of the three substrate types; or b)
misinterpretation of the classification system in that
model. The second variable: "Percent of water area less than
25 cm deep" was judged by Observers 1 and 3 to be 51% or
greater, at two different Water Habitat locations, whereas
59

60
the rest of the survey party judged this to be 23% or less
at these locations. A possible explanation for this
consistent difference in scores could be the fact that,
Observers 1 and 3 were the most experienced team members in
Habitat Evaluation Procedures (HEP) and their prior
experience may have influenced their judgement of this
fairly subjective variable. The fourth variable: "Percent of
water surface covered by logs, trees, or woody vegetation"
was estimated by Observer 3 to be 15%, whereas all other
observers estimated the "coverage" to be 2% or less. This
variable requires observers to estimate the "percentage of
the area covered by material overhanging and projecting from
the water" and so is very subjective and prone to
differences of opinion.
A second habitat where variability between group
measurements occurred was the Oldfield Habitat (Eastern
Cottontail Model). For Variable 4: "Percent shrub crown
cover" Observers 1 and 3 estimated "coverage" to be between
10% and 15%, whereas all other observers estimated
"coverage" to be 1% or less. Once again greater "HEP
experience" may have influenced the judgement of these
individuals, accounting for the differences between them and
the less experienced members of the team. Variable 5:
"Number of refuge sites per 0.4 ha" showed variability
within group measurements at both this habitat (Oldfield)
and the Shrubland Habitat (Eastern Woodrat Model Variable
3).-Observers varied widely in their judgement of how many

61
"refuges" there were at these sites. This variability is
probably due to the subjective nature of this variable as
observers were expected to evaluate the suitability of
burrows, crevices, caves, windthrow and brushpiles in a 0.4
ha area as refuges for these indicator species.
Overall, these results indicate that aquatic habitats
showed the greatest group variability in measurements. This
variability seems to be due in part to the subjective nature
of the variables involved. The amount of prior HEP
experience also seems to have an effect on observer's
measurements of subjective variables.
The second hypothesis proposed in this thesis was:
There is no significant difference in mean HSI values at
impact assessment sites between sampling years. A
nonparametric Mann-Whitney test was used to analyze the mean
HSI values for variables, in both sample years, at each
impact site. Results indicate that for all impact sites,
except Upland Forest (Fox Squirrel model), mean HSI values
between test years were not significantly different. In
future studies the power of this test (1 - 3) would be
greatly increased and the likelihood of committing a Type II
error decreased if a larger sample size were used.
The Upland Forest (Fox Squirrel model) result was
significant (Ut)., 2 , 4 = UO.05(2 ),4 = 16) at the p = 0.05 level.
Variable 1: "Percent tree canopy closure of mast producing
trees greater than 25 cm diameter at breast height (dbh)"
presented variability in mean HSI values between sampling

62
years. "Canopy closure" was judged to be an average of 35%
in 1987 and only an average of 10% in 1990. This difference
may be accounted for, in part, by the fact that the HEP
sampling period in 1987 was done in late Spring/early
Summer, whereas the sampling in 1990 was conducted in late
Summer/early Fall when tree "canopy closure" may indeed be
slightly less. In 1987, variable 3: "Average dbh of
overstory trees" was judged to be on average 12 inches,
whereas in 1990 the "dbh" was estimated at 5.5 inches. The
most likely explanation for this discrepancy in "dbh",
between sampling years, (since there was no evidence of tree
harvesting) is that in reading values off the HSI functional
curve an error was made. This functional curve has an X axis
with two scales on it. One scale in inches and a second in
centimeters. If a "dbh" value of 12.5 cm was estimated in
the field and then erroneously read off the functional curve
in inches, this discrepancy would be accounted for.
The mean HSI values for 5 out of 6 Impact Sites showed
no significant difference between sampling years. The sixth
site (Upland Forest) does show a significant difference in
mean HSI values. Possible explanations for this difference
are: a) seasonal effects on measurements; or b) observer
error in interpreting functional curves in 1987.
The third hypothesis considered in this thesis was: For
HEP model habitat types there is no correlation between
Habitat Units and Indicator Species density. Results of a
nonparametric Spearman's Rank test indicate that the Null

63
hypothesis should be rejected and that there is a positive
correlation between HUs and Indicator Species density at Ray
Roberts Lake (a = 0.05, 0.05 < p < 0.02, r = 0.643, n =
13). Since HUs were designed to indicate habitat quality,
and Indicator Species are supposed to be representative of a
particular habitat, this positive correlation appears to
verify these HEP models. However, density alone may not be a
sufficient indicator of habitat quality (Van Horne, 1983).
Schroeder (1990) suggests "... the HSI, as a measure of key
habitat variables, considers only a subset of all the
factors that may influence individuals and resultant
populations. Thus, habitat measurements may indicate ideal
conditions, but nonhabitat-related factors (such as disease
or severe weather) may cause population declines. The HSI is
best viewed as an index of habitat-imposed limitations on a
population." This implies that a maximum HSI value of 1.0
indicates no habitat-related limitations exist. A low HSI
value such as 0.1 would indicate significant habitat
limitations exist (Schroeder, 1990). Future studies might
include testing other measures of Indicator Species
performance (in addition to just numbers per hectare) such
as number of young fledged or nest success, and increasing
sample size, in order to improve test significance.
Overall, there is a correlation between HUs and avian
indicator species density. However, density alone is not an
adequate indication of habitat quality.
The fourth hypothesis tested in this thesis was: For

64
HEP model habitat types there is no correlation between HUs
and species diversity. Results of a nonparametric Spearman's
Rank test indicate that there is no correlation between HUs
and species diversity as determined by both the Shannon-
Weiner Index and Brillioun's Index (For the Shannon-Weiner
Index, rs(o.o) 2,11 = 0.618, r, = -0.08, p > 0.50: For
Brillioun's Index, r(QQ5 ) 2,1 = 0.618, r, = 0.384, 0.50 > p >
0.2.).
This result seems to indicate that species diversity is
perhaps too general a measurement and therefore not a good
indicator of habitat quality as measured by HUs. An
alternative interpretation of this result is that the data
set used in this analysis was too small to show
significance. A suggestion for future work would be to
increase the sample size.
HEP proved to be a useful tool for examining the
condition of wildlife habitat in both 1987 and 1990. The
results from this thesis can be divided into two categories:
1) observer and temporal variability; and 2) the
relationship of HUs and ecological indicators.
Recommendations for future work within the first
category would be: a) Increase survey team consistency in
variable measurements by a more intensive training period
prior to actual field work. Subjective variables such as
those in the Green Heron model (Water habitat) should be
thoroughly discussed with perhaps example "trial measures"
being taken prior to data collection; b) Insure data is

65
collected at exactly the same time of year and by the same
survey team for each study to increase temporal consistency;
and c) If possible choose models with fewer subjective
variables.
For the second category recommendations are: a) Testing
other measures of Indicator Species performance in addition
to density, such as the number of young fledged or nest
success, mortality, etc.; and b) Increase sample size in
order to improve test significance.

APPENDIX A
HSI LIFE REQUISITE VALUE MODELS FOR INDICATOR SPECIES
66

HSI Life Requisite Value Models for
Species Life Requisite
Belted kingfisher Lotic Food and(R/L) Water Value
Foraging CoverValue
Reproduction
Green heron Food Value(R/L)
Water Value
Eastern Winter Foodmeadowlark Value(P)
Cover andRepro. Value
Racer Food Value(P)
Indicator Species.
Aggregation Function
(vi x v2 x v3 x v4)14
(v6 x v7)1/2
ve
(vi x
(v5 x
vi
v2 x v3)1/3 + v4
vs) 1/2
v2 or (v3 x v4)12 or v,
(2(vl x v2 )12 + v )/2
Scissor-tailedFlycatcher(OF)
EasternCottontail(OF)
Eastern Woodrat(SH)
Downy Woodpecker(SH)
Barred Owl(BF)
Raccoon(BF)
Food Value
Cover andRepro. Value
Winter Food
Cover Type
Cover andRepro. Value
Food Value
Repro. Value
Water Value
Cover andRepro. Value
Water Value
Cover andRepro. Value
(v, x v2)"'2
v3 x v4
v, + v2
v4 + v,
(vI x v2 )1/3 + v3
V,
V2
(v2 x v3 x v4 )1/3
(vI x v2 )1/2
(v3 + v4)/2
67
.

Species-
Fox Squirrel(UF)
Carolinachickadee(UF)
Life Requisite
Food Value
Cover andRepro. Value
Cover Value
Repro. Value
Aggregation Function
vi
(v2 x V 3 X v4)1/3
(v, x v2 x v3 )1/3
V 4
68

APPENDIX B
HABITAT SUITABILITY INDEX VALUES FOR THEVEGETATIONAL TRANSECTS
69

e . C C; -
LO
* *: O O
*: N:- 0---
0i~
* 0000 O
Coc.
' '0000 e
C: _
'0000
re N: v
.. N:
0: :
'0000
Sn SO U'
N
0
U(n t
> >>
4.U)
0
04
4)
0
UIt0'
0
U-J
LU
';
5.I
I-
8
CL
5"UK
ad
C8
. ...*LU
LU ' 0 0 N0 0
'0000 9 e
0:
0000
0I.
000
.n 0. O O
C,,C;
N .,S.
~: :0 0
S0Q.
Qi
'0000 >
. m. OOo'0000OO
N:'N' gg
- - - -
U)- 0000O
SQ.O
'0000OO
vs *
'0000O O
70
4-)
0
041)4.)0
0
000ID
U's0;
LUJ
8
cc
0u
U-J
C;
w00 0 0
* ' u o0o o
:N : o.e ~
wo: :0 0
. . 0 w 0 0 o
N: : '~
9: 8
0 ' % 0 0
( cm'
- .:OO
Co c o'0 0
'00000: :u ~
gro
. ~ 0, *0
* ' II 0u'
* ' 0~
-* w
* I-.
* C . O *
4.
40
0
0
OC;
0
'C:3
I--
oe-
Co
a-w
0u.
.~ ooON0 LM00
*'N 0' 0 C
pn m O N
Ok 0 do
0: :; :C
0-Ow
'000o
U) > >
0: AOO
. ' ' e0'
'0000d
I400
0'0'd
000w
''N'
0' moom,,
. 0'000'
: : ;0w-Ow-g
71
r4)
0
'-4
0
w4)0
4114)
h4
04
0m
-4e
0
0
0
I-U
a-
aof:3A
LACo
0'
-J:3
w

U].
0:e
'
00
* J 00 a 0 Ok & k
* (S * O O O O
; 0 0 0.*
* 00000
N . 'Coo
* 000 0
c 14
-* ' - . 0
C; C; C; O
* 00
a o
4N 0 >
0 0* ONO O* 0
S* 0000O
* m 0
N * M .
'-O
* W
* 0000 O
* 0 00
* - N000 4 g
W . ..
000 O
44 > >
4-)
4)04
r4
0
00
0Q
LUJ
OOk
: It : Z. -.*
-0-
0 00 0
O O N
w . .
. .0000.0000
O. .0 0
O. .0000
'0 '
0 .0000
,a: :
. .000..
(fl .OOOO
0 .0000O
1-
8
72
4.)
0
04v
0zUt
0
0
9
o r-q
C)
0
-0-s000-'
H.;
N *U. i
0~
u
L8
-j
wA0uo

U s
C; C0;0.C0 C;Ln:* * 0 0
I N &A 0
0: 0:
a a 0 O 0 O
* a Oe
coc
* a N N0
* m *
o: :e - a 0- 0r 0
-: : N2 0.0
U) a .e
a .000
H' .000
a. r
N N *' c'O
a . 0 0 0 0. .
C; C; 4O O
LU>' >
a',.
73
r4
2f)
0
t1)
0
41)
04
0F--
LU
LUa>
* a
0:a a
0~U a
0 .a
.U)0
H.
a N
U. N
a:
a -
' 0' N. N 0
a 00
.0 0
a 0 0 00
04 Oh.m
466
. O a 0
OO 0
a 04 0 0
0 0k 0 0m
. O iflO
a .0. .'0000
00.
: 00.0;,'.00 0
'0000O
a. 0 0
a 0a-00
'0 a 0 0
0 N 0 0
. . . 0 .a 0 0 0 0
a. -N .
40
.'-
00".
O'
0 0
(A-
0 -
0 0
LU
0
.a0
a'or.
L"
- 0
0.w

Lu
LU 00 0 0
4'
-
Ok n
F N f n0 00t
0. _
* 0000
N 2 00o
S * 0 0
8 9
' '0a0o0
.N * ' 0 -
* .- *4 '4 N O
* '0000917
* ' N e '4 0 0 '--
0' '
'0 0
0000
(A>4>- 0 >
.0000OO O
LU
I'- **- w- N'4u'i
CA'e> >>
U)
0
'.
U)
0
10
(4.4Var-0
0
0
go
--3Ci
U
m'
U)
0
C'4
(n
0
U)
0
-J
>
LU
0u
U
4 '
r4
'00 0
* 0 0
* 000.-
N 0 0 0 N
000 0
N N 0-
'0000
N 0 0
O O in
=- . *- N 4 t(A I > > > >>
74
'i-q
01
00414J0
4)
v
0
N
N
--J
c3
0Lni
U.
0LU
0u

SU
U -0 00 0
f,*0 0 co 0
S. N .0 . . 0
* *0 C) 0
U).
>- .>0.0. .
* . 0 eo e
* . N * 0 t 0 0en : O NR 9
0 e w -0w-00w
* 0 0 ( 000eo* w-w-0w-
N : O O 0* .O w .0 .
0. eOe0w-ow
w-e . .~ .n 0
N * 0 0 U'. .00 -o O0-
e - * w-N OUe * > - > >
r-
0.9
0 0O
of
LUo
-J
LU
00'
0;
LU
* * u*' LA 0
* * *O * *A
N 0n O 0
e e
.. N e* IA 0C;:
0'l
C;- - -;-8
C O;O
Ok pn 0 0
C; 8
Ln 0
0. *
0 0
e. N .*
cn m
8 8
* * &M 0
> -' >
e N :000w-O
* LU
I-* (0 *
75
0
4
4
f-i
1.
4
0
N 00O0
0
so-J
N
'0
.0)
S
of-JLU
u>0C-)

U
uj C-
pnOn
40't
* O0o
* ' 0 4
U). '> > 00
* '0 0U)0
*' ' 0 o
- - 0 -
** 0 -O O0
.: N:
U)
0
U)
0
4.)
041)41)0
4)U)to~
0 00o 0
0 V.
LU-j
LU
0u-
00o
0
C,
U)
0
U)
0
U)
0
U
(* N 0 -4O
U 0 0 00
S 0
m 0 0 0
* OO
N 00ON
*0 00 0
t O0 00
0000- OO
( A -> > -
'000*
N'8
76
4
(0
4J
-. )
40U)(n
'n
v
0
040
I-
U)
U4
>U

e e . 0
.O *0N
* 0 N O
e e0' * 0* '00
*e N :8'00
8 8
* 0
0s N * 0-
*** ' I 0
* '0O 0
*' N ' 0 0
O: :00
0 0
.0.0
-. eN e
Ue '
t0
C)
0 N
0 00
*>0LUM
*A *
LU)IA. e
0 .00
C; 4
* M * NO
* *0
N 8
.. .:. .
*e .0
* .0
ge . . .
* 88
. .0
. . 0
e .0.0
. .00
* .00-* N * 0" N
* .0
H: :00
N :
* ,* S 8
* * 0 9
* w* I-*
* *, S >
77
ci)
0
ai)
ai)
00
>1
0
C
C
kn
$Me
.j
C
0
w
-
*C
w
w
C'.

LU
0 0 0N
00ON
U, 000
A - -A
00 *0Oa wl ON
' * N IA 0
U. 0000 .
0:' M: 0
O. .l
0 - 0 N 00 0000&
C; C; 0 CN ;
U) *00
0: C
H: CO
N .O:
. . :00> .
0 E' VIO 0
0 0 00 0000 O
0 *U 0. . .0
0 (40
U)
0
0*J
U)
0
'0H
U
0.0
0o 0
4)- '.0
0(l)(0)
U)
0
(1)
0
H
U)0
*.e*eeee.eee*e..
e'-,
*:ee
ee.'|.*eeee
***e
..e*Ieee.e
*e:.|eee,-|,,eee
-*.'e*e
ee*e
*ee
78
U)
U
41 *
* 000* N 0
9: 19.00
Noo
* 0 0- :
C;. c;
* Nu O
000CN
N: r
0 0C03
000
N C
W. O . .
* 0
LU
> >
(I 0
>
a ':uiIx
0%
Q0

LA'
U):
*. N -
-Q.
, w* e -
. .
oN 0 00A 0 t^0
0' 0u'I O .t 0
O o0 0 0 0 0o
o 0 0 0 O 0 0 -
o o 0 d0 0m 0o 0.0 0 0 0 . .
400000C- 0
o o 0 0 0 0 0 C-
40000LOA
o 0 0 009
o o0 0 0 0 0
N01 I G o 0 .
"k do in O M O
C; .; . . . . .:.
p v is 4M O 9 Ic.; . .C .; . ; . .:.
(V Fn t Ln 4
41
0>
U *
a-w09
N
0'
0e
w
I-0-J
U.
a>
8(3
z
.-0-i
LA:
e N
eQ.
4K
* ' N
C'
e e
. uJ* e .
*In
s e e
,, e(A e
o 0 0 0 o 0 o
o 0C; 00C-
0 0
Lon OOto
0000000; o -
00200990
Ck o 4 o O
6n en 0 0 00
0I00 02 0 90 --
In in 0O 0 00O 0 0 o 0 C -
0 0 C;00 C
do mN 4ON
06 n In40 0 0
o o 0 O 0
NM Q " In N o
79
00
"-4
4v.4
4.J
w44-)0J 8
0
LU (Al.
0.
C; C
I--
3
S-0-J
ac0U>200
.-
a0

* LU
* <n
e
. N
* .
* .
-N
e L* I
* I
-eeee
e-ee
eaeeeee- ae-e
-e
ee.eeee,e-ee:*
*- ee
eeee
-eeeae--
eee
ee-eeee
N O 1 I6 %M 00 -t- - 0' 01
0o 0Q 0 00-n '- *n *n *n 0
a O
00~-00
0 0 0 0'0 0 0
0 4 00 0 0
00 000 00
in 0 in 0
000 -- 0000N
In'- 0 0 u *
00-- '- 010 0 0 0 0 0-
0000000 O
0 0 0 0 -0
In 0 0 0 In * 0
0 0 0 0 0
0- 0 0 0 0 0 e
'-Nr t-l e- n0 r
80
, LU
u(3
. I
I-LU
, N
' N
G) u4,)
, Q
, *
.m4.
o iv., ,
'-0 .J. N
a :
Ix,
~eIe n
rw0
"40)
Iz4
,.,
,.,
e.,
e,,
e,,,
e.,,:.
.
e,e-,,e
ee,,
e
e
,e,:e,
0 4 00 0 0
0000N C;-
ry FM C O O
000000
0 N 0 I0
. ..--. .%. O N
NN N0'
0- 0 00o
8 8 8 8OO
00 00
N ?' N e- .
00in00N l000
000 0300
0 NO 0
00 00 0 I 0
0 0 0 0 0 0 0
. .n . . . .o
ONOOM ON N .- '- '-0 00 0 0 0 0 0 0
8 0 0 I 8 88 0
In 0 N e-S 00
.- . . .n .O .
140 0
LU-j
00--
LUc-
C
(I)

0'
0:
U,'0:*
1W *
N
N
F""
N
N
LU
:
0U
0u
M
LU (AIx
0
LUJ
4304
u
0-j
0
4.,
4,
uac0
-
u
0
,.
0
o -0 0 C 0 0
N W i% 0 U
o to 0 0 o 0 0n -
o 0 0 0 0 0 00
o 4^ 0 ion 0U-
C; C qc O
* - . . . .
81
4)r0".4
U)
4
44
14
LU
* 4
LU
.. N -eme
0:
* . N
. N
'-4'
0::
LU
in
: :
000000O U U-
0N9 0 0 0 0 0 -
0 00 0 0
0 0 0 0 0 0 U-
0 0 0 W%00 U00000000w-
W% N in n O
Oh U) 0 Ok
Ok cM o UN In O
0 0 0 o 0 40 0 0 0a
.k . c. . M.0. .hl
N Nin n 0 0 & 0
6Ok O k 40 in C k
0n 0 0U0 0 0 0nU0
N . . . .) .O . .
Ol Ok o n Ok 01.
.^ .%. . . Go.
PO N P.- * 0F' N. N. .Q .
o 000000U) N 0 U) U)U)
&A cm :9 m
WNa W% . .
o N o U -
#0000
0000 0))
N O 04
IF e Ok
CO 40 N m O
0 0 0 40 in
00 014 V- O k1000 W% U00
N 0 - -0-.
0 0 0 0
U,,000 U))e ee0000000o
0000000
00 O n U-' U..
N 0 U - U- r
- - - U- -OO
0 w'- 0 0 0 0 0
co
0 0
LU
-J4
*e
I-
4
aU
4
0

APPENDIX C
HABITAT SUITABILITY INDEX VALUES FOR THEIMPACT ASSESSMENT SITES
82

: , : Sg
U. . 000
:~
, o jooo0. .
- '0000
0. -
o: :000
-.
- U)
U)
0
V.
'0
r13
0
'V
U
00
0N4.
1
L
00
LU
4-:3
U,SQ
0
U)
SQ
0
0
LU
LU>"0('
LU
ugg
- n o-0
N'
0'00
'0E - 0 0
N. I
0 0 0.0
- -0
-;
0''0
(A>N' > 1
83
00
W4UU
4J
0
0
04Ji
4J
0
4I)4.)of-4c41
0
H
N
'A
()0
0
0
U
0
CL
a
09
LU
0
LU
-J
Lu
U
* L 0 0 0 0
C)0
ON* ' N * * 4 uv
* O'0000M
* O'0000O'0
0.
6 6
'0; C;
'000
N 0 CO0..
* ' 0w 00e
* C;O 0 0
'0
> '0000
. . 0*~ '0O N O 0
Vi' N . M
* '0 0
* UO
(n * * * .
00'0(U
U*1-4
U
(U
0
(UU
0(J
U
600
0,
-N0 0
LU.."C
0LU
LU
0
0
U)
,-
* LU
L ' 0 0o0c00400
000
Fn 0r4 m Ck
0000CO
*' N 'C- g
> > ..> >
CA:OO
0a ' V N 100
N * Oe
***r'.. .4.** OOO0
e '0000
0' '
_ * ia
0' ' P. 0 0
* N- 0 O
* LU
U. 'e
84
4J
004
Mr--
4J-U2-
En4J
w
01cfl
0
0
LU
.U
C..
0*
LU
LU
ix
4c"
N
0u

U
* .
on,
on 4200 (Y
* * 0Oa 0
. . .'t .
(A - > 0 v-
'00
* '000'-O
* '09-0'-
* '09-004
* ''- -0
* ' 00O n0
* . .w 0 .
* 'OOON
CD 4 eoO
. ,
C'OON
* '0- 0 0O
*' * * * *
* ' e- e 0 O
* 0 0 O O
e * 00O M
* I - - - -
* * O
P-4
'4
Cu0
041
4J0
44)
41
LUN
.U
0-"C
0>L.L U
LUn
0 O k
40 * U 0 0 0
* 0* ' N * -
.' N ' F" 0 0 F"
0:0
'* * *:* '0000
UJO.~ N:
CD: O
* *' 0 0
*' N
, * ' 0 0 0 4-
' N ' 000 * *
9-' ' * N* 0 *
* '0 0 00N e
* * * *
**0000N
* 0 0* '4 0 0
CD' N OO
- - 0''- 0'
* *'0000
* L
* ".>>>>m
85
-4
0
4!J -H.4 to
4J~'U 0(a
00RC
ON
LU
C0c
N
e-o
N
LU
LU
0u
NUe
0

86
Ln -4 0% 00* LU' 'L
* ' ' ' .* u 0 *LU' 00* >,
* '00O , . O L O
S ' ' 0 0 * * . . ..0 0
000
N OO ,OQeO'00 *
C; C;
' 0 0* 0 0
w:w
' '00' 000
4J* r-39 4J
000000
~ e _ 'LAO ' * 0
o O Oen In 0 N IN
CD 0*
- LA:U
>
e F.1e:'000
* ' 0 0
e N eO O ee N 0 N N
Om- ' 0 0 ' - '- - - 0
N : 2 ' '
S '000 ' -' e eO r
wL O
H' * H'O '
* ' 0 0 A VV' * '
N N'
0: ' 0'NO LUC ..
* ** ' * ' ' ' L
(J. C ' *
* ' "N OLU ' - ' MN ) 0

*
* a
*
* '
' N
* N
* N
0:
* p
U* N
* p.
* (
-.*
e.*'-*
ee*e
.*e*e'
'
'ee*e-.e*e*
**es*
***e' ,***
**e
*
**
'
N OVP N00.
U..
I-
3
CL
a
ac
0*U
2
N
..U
at
0
O O N ON
000
00000
N- O O Ou0 Oh uOhO
OONO it
- 000 O
0 00 0 0
87
0
0
c
4)4J
W
0
$4
4)C.)
01
C.)0
LL)
La (a
at ...
*L '00 0 0
* - 0 0
S . Ni
cm- do
' Nw*P * 00 O -
='kn
* - 0> > > .
* o 00O O
VP o 0 ' O 0
* N * 0 Oh 0
0: : Oh 0
* O e . g .VP ' 0 N O 0
* '0 0
fJ) N : 9 0 A ,-
O * OOO,
* LU
(4 *
M0LU
a
LLA
3
S
(.
r4
..
0
LU
-J
* 000 Oh
NOOOu',
N 0 0 0 Ui
N VP "t u

- L
* L
* I'
0. N
* 94
* N
jO)
' N
* N
* U
e
3
*
.e:**
ee
'eee
-*
''*o*
*
IA iA in IA0 0 0 0WN A co A~
0N 0 0 0 Ok00
0 000 0 0
IA IAIA in 0 0 0 0
O~ IA 0 ONO O00
0h 00N 0 Ok0 0
(V M & t N *4 4
N 4. IA ~O 7% 0
> >
40
-j
:3
I-
I.-0-J
o00N 0 0
in O do
W's A ) 0 00
0 0 0 L0 0 0
0 0 0 W,00%
0 0 0 (m00
0 0 0 0 o OO0
. .. . . . . .0 0 0 0 0 0 0- w-
3 * p
0' 4
N 0:N
IA
IA
.0. *
N N
2 ( N1
* .
0*-*0 W
88
*UC
'*
C
-
-
I,4
4-J2
4J)
.0 do0 0 iAIn0
000 0 IAI# O
4A0 00. I V-
0- Ok
0 0 0 0 0 0 0
0 *-A
0 0 0 0 0 0 0
0 0 0 0 0 0 0
O O AOO IAIAO
0004
0 0- 0 0- 0
0 00 0 n00
ONOOO IO O
0 I N 0 LAIAN 0
NN 14)
N - -0000000OO O
IAOOOIIOO
0 IA N O 00 0 0 0- 0 0
N 94 .4 IAm O:3>>>>>>-
(I) *
0 0
LUj
-j
LU
-J
0
7.
U.
2
U
N
*U
0n 0 i0 0 00
0 00 0 0 - -
in g In In A0 0 00 in O o 0 0ON
C;OO
.Lae
w

APPENDIX D
AVIAN CENSUS NUMBERS BY HABITAT TYPE AND TRANSECTSECTION FOR 1987 AND 1990
89

90
Avian Census Numbers by Habitat Type and Transect Section(1987 and 1990)
e . .an p s Tnsect F- orL )rs(17/1990)
Upland Forest CarolinaChickadee*
Common Flicker EF10
Downy CD7Woodpecker CD9
EF10
Tufted CD7Titmouse CD9
EF10
Cedar Waxwing CD9
Cooper's Hawk CD9
BottomlandForest
Shrubland
Pasture
Barred Owl*
CarolinaChickadee
DownyWoodpecker
E. Meadowlark*
Ki 1 deer
Dickcissel
SavannahSparrow
CD7CD9EF10
9/512/75/5
2/-
2/22/-2/1
16/46/94/3
-/15
-/1
1/-1/-
18/-2/-1/-8/82/2
20/4
1/-2/21/-3/-
12/-11/-62/1526/3
AB3GH3
AB3CD11EF4EF8GH1GH3
AB1AB6CD6EF5
AB2EF3GH2GH4
AB2EF3
AB2
AB2EF3
1/-1/-
1/-10/-
Habitat Tun Aui n" i %g - M- --- qo.%449 or& -C r% I - -3 - t .6 A% 0% Ow s m - - 'L

91
Avian Census Numbers by Habitat Type and Transect Section(1987 and 1990)
Habitat Tvnpp_% es rV46anst9 #-ofr d9 )
Red-wingedBlackbird
EF3
Scissor-tailed AB2Flycatcher EF3
oldfield
Riverine/Lacustrine
Scissor-tailedFlycatcher*
E. Meadowlark
Great Horned
Green Heron*
BeltedKingfisher*
Wood Duck
Pasture2/-
4/-8/-
CD1CD2CD10EF1EF2
AB5CD1CD2CD3CD5CD10EF1EF2EF9
-/3-/7-/1
11/53/-
2/-39/2212/2623/-23/-51/5
123/72-/27/1
CD1
EF
ABEF
1/-
1/-
2/--/1
-/3-/1
ABEF
Au ian Sn t- i TM-- "4r.,M "+ Ji lr% %o . -3 - * 1 0% 0% V-D F .6 A.. 'ok .. 1,

APPENDIX E
BRAY-CURTIS CLUSTER ANALYSIS DATA
92

93
Impact Site Water Habitat (Green Heron Model)
Linkage Clusters Linked Similarity Probability
1 2 4 0.95349 0.624002 2 5 0.93649 0.638003 1 2 0.83546 0.020004 1 3 0.80392 0.00600
Transect EF Water Habitat (Green Heron Model)
Linkage Clusters Linked Similarity Probability
1 1 3 0.98606 0.390002 3 5 0.97641 0.158003 2 4 0.96667 0.324004 1 2 0.87007 0.00400
Transect GH Water Habitat (Green Heron Model)
Linkage Clusters Linked Similarity Probability
1 2 5 0.96791 0.574002 3 4 0.93559 0.432003 2 3 0.90995 0.508004 1 2 0.81755 0.02200
Transect AB Shrubland Habitat (Eastern Woodrat Model)
Linkage Clusters Linked Similarity Probability
1 4 5 0.97448 0.808002 1 3 0.94020 0.904003 1 4 0.71176 0.186004 1 2 0.45396 0.02800
Transect EF Oldfield Habitat (Eastern Cottontail Model)
Linkage Clusters Linked Similarity Probability
1 1 3 0.88099 0.724002 4 5 0.69985 0.018003 1 4 0.67425 0.07600
-4 1 2 0.61704 0.02200

REFERENCES
1. Allen, A. W. and M. J. Armbruster, 1982, "PreliminaryEvaluation of a Habitat Suitability Model for thePronghorn", Proc. Biennial Pronghorn Antelope Workshop,Vol. 19, pp. 93-105.
2. Anderson, S. H. and H. H. Shugart, Jr., 1973, "HabitatSelection of Breeding Birds in an East TennesseeDeciduous Forest", Ecology, vol. 55, pp.828-837.
3. Block, W. M. et al., 1987, "On Measuring Bird Habitat:Influence of Observer Variability and Sample Size",The Condor, Vol. 89, No. 2, pp. 241-251.
4. Bray, J. R., and J. T. Curtis, 1957, "An Ordination ofthe Upland Forest Community of Southern Wisconsin",Ecol. Monogr. Vol. 27, pp. 325-349.
5. Clawson, M. E. et al.., 1984, "An Approach to HabitatModeling for Herpetofauna", Wildlife Society Bulletin,Vol. 12, pp.61-69.
6. Cook, J. G. and L. L. Irwin, 1985, "Validation andModification of a Habitat Suitability Model forPronghorns", WildlifeSociety Bulletin, Vol. 13,pp.440 - 448.
7. Diaconis, P. and B. Efron, 1983, "Computer-intensivemethods in statistics", Scientific American, Vol. 248,pp. 116-130.
8. Efron, B., 1979, "Bootstrap methods: another look at thejackknife", Annals of Statistics, Vol. 7, pp. 1-26.
9. Ellis, J. A. et al.,, 1978, "Results of Testing FourMethods of Habitat Evaluation", Report to the ProjectImpact Evaluation Team, Division of Ecological Services,U. S. Fish and Wildlife Service, Fort Collins, Colorado,pp. 1-92.
94

95
10. Ellis, J. A. et al., 1979, "Appraising Four FieldMethods of Terrestrial Habitat Evaluation",NorthAmericanWildlife Conference, Vol.44, pp. 369-379.
11. Emlen, J. T., 1971, "Population Densities of BirdsDerived from Transect Counts", Auk, Vol. 88, pp. 323-342.
12. Flood, B. S. et al., 1977, A Handbook for HabitatEvaluation, Resource Publication 132, U. S. Fish andWildlife Service, Washington, D. C., pp. 1-77.
13. Gotfryd, A. and R. I. C. Hansell, 1985, "The Impact ofObserver Bias on Multivariate Analyses of VegetationStructure", Qikos, Vol. 45, pp. 223-234.
14. Institute of Applied Sciences, University of NorthTexas, 1988, "Ray Roberts Lake Pre-impoundmentEnvironmental Study", Ft. Worth District U.S. Army Corpsof Engineers, 157 pp.
15. Institute of Applied Sciences, University of NorthTexas, 1992, "Ray Roberts Lake Post-impoundmentEnvironmental Study Year Two", Ft. Worth District U.S.Army Corps of Engineers, 221 pp.
16. Kennedy, J. H. et al.,, 1993,, "Sampling and AnalysisStrategy for Biological Effects in Freshwater FieldTests", EWOFFT, Potsdam, Germany, pp. 31-33.
17. Larson, D. L. and C. E. Bock, 1986, "Determining AvianHabitat Preference by Bird-Centered VegetationSampling", Wildlife 2000 Modelling Wildlife Habitats ofTerrestrial Vertebrates, University of WisconsinPress, Madison, Wisconsin, October 7-11, 1984, pp. 37-43.
18. Laymon, S. A. et al., 1985, "Habitat Suitability IndexModels: Spotted Owl", Biological Report 82(10.113), U.S.Department of Interior, Fish and Wildlife Service,Biological Services Program, Washington, D. C., pp. 1-14.

96
19. Laymon, S. A. and R. H. Barrett, 1986, "Developing andTesting Habitat-Capability Models: Pitfalls andRecommendations", Wildlife 2000 Modeling HabitatRelationships of Terrestrial Vertebrates, University ofWisconsin Press, Madison, Wisconsin, October 7-11, 1984,pp. 87-92.
20. Merikallio, E., 1958, Finnish Birds, Their Distributionand Numbers, Fauna Fennilca V. Helsinofors., Finland.
21. Nemec, A. F. L., 1991, "Documentation for Sigtree",Version 3.4, Canada, pp. 1-11.
22. Nemec, A. F. L. and R. 0. Brinkhurst, 1988, "Using theBootstrap to Assess Statistical Significance in theCluster Analysis of Species Abundance Data", Can. J.Fish. Aguat. Sci., Vol. 45 pp. 965-970.
23. Payne, B. S. and K. S. Long, 1986, "Airborne SensorPotential for Habitat Evaluation Procedures (HEP)',Technical Report EL-86-3, Department of the Army,Vicksburg, Mississippi, pp. 1-45.
24. Root, R. B., 1967, "The Niche Exploitation Pattern ofthe Blue-Gray Gnatcatcher", Ecological Monographs, Vol.37, pp. 317-350.
25. Roberts, T. H. and L. J. O'Neil, 1985, "SpeciesSelection for Habitat Assessments", Misc. Paper EL-85-8,Department of the Army, Vicksburg, Mississippi, pp. 1-16.
26. Schamberger, M. L. and L. J. O'Neil, 1986, "Concepts andConstraints of Habitat-Model Testing", Wildlife 2000Modeling Habitat Relationships of TerrestrialVertebrates, University of Wisconsin Press, Madison,Wisconsin, October 7-11, 1984, pp. 5-10.
27. Schroeder, R. L., 1982, "Habitat Suitability IndexModels: Downy Woodpecker", FWS/OBS-82/10.38, U. S. Dept.Int., Fish and Wildlife Service, Washington, D. C., pp.1-10.

97
28. Schroeder, R. L., 1983, "Habitat suitability IndexModels: Black-capped Chickadee", FWS/OBS-82/10.3 7, U. S.Fish and Wildlife Service, Washington, D. C., pp.1-12.
29. Schroeder, R. L., 1990, "Tests of a Habitat SuitabilityModel for Black-capped Chickadees", FWS BiologicalReport 90(10), U. S. Fish and Wildlife Service,Washington, D.C., pp. 1-8.
30. Severinghaus, W. D., 1981, "Guild Theory Development asa Mechanism for Assessing Environmental Impact",Environmental Management, Vol. 5, No. 3, pp. 187-190.
31. Short, H. L. and K. P. Burnham, 1982, "Technique forStructuring Wildlife Guilds to Evaluate Impacts onWildlife Communities", Spec. Sci. Report No. 244, U. S.Fish and Wildlife Service, Fort Collins, Colorado, pp.1-5.
32. Smith, A. D., 1944, "A Study of the Reliability of RangeVegetation Estimates", Ecology, Vol. 25, pp. 441-448.
33. Sneath P. H. A. and R. R. Sokal, 1973, NumericalTaxonomy:-The Principles and Practice of NumericalClassification, W. H. Freeman, San Francisco, CA. 573 p.
34. Spencer, W. D.,, 1983, "Marten Habitat Preferences in theNorthern Sierra Nevada", Journal of WildlifeManagement, Vol. 47, pp.1181-1186.
35. Sturman, W. A., 1967, "Description and Analysis ofBreeding Habitats of the Chickadees, Parus Atricapillusand P. Rufescens", Ecology, Vol. 49, No. 3, pp. 418-431.
36. U. S. Fish and Wildlife Service, 1976, "HabitatEvaluation Procedures", U. S. Fish and WildlifeServices, Division of Ecological Services, Ft. Collins,Colorado, 77 pp.
37. U. S. Fish and Wildlife Service, 1980a, "The HabitatEvaluation Procedures", U. S. Dept. Int. Fish andWildlife Service Ecological Services Manual 102, Ft.Collins, Colorado, 124 pp.

98
38. U. S. Fish and Wildlife Service, 1980b, "TerrestrialHabitat Evaluation Criteria Handbook (Regions 2512 &2320)", U. S. Dept. Int. Fish and Wildlife Service, Ft.Collins, Colorado.
39. Van Horne, B., 1983, "Density as a Misleading Indicatorof Habitat Quality", Journal of Wildlife Management,Vol. 47, No. 4, pp. 893-901.
40. Whitaker, G. A. et al., 1976, "Inventorying Habitats andRating Their Value for Wildlife Species", Proceedings ofthe Southeastern Association of Fish and WildlifeAgencies, Vol. 30, pp.590-601.
41. Zar, J. H., 1984, Biostatistical Analysis, 2nd ed.,Prentice-Hall, Englewood Cliffs, N.J., 718 pp..





























