Embed Size (px)
Citation preview

In Situ Compositional Analysis of Acidocalcisomes inTrypanosoma cruzi*
(Received for publication, April 17, 1997, and in revised form, July 11, 1997)
David A. Scott and Roberto Docampo‡
From the Laboratory of Molecular Parasitology, Department of Pathobiology, University of Illinois, Urbana, Illinois 61802
James A. Dvorak§, Shanling Shi¶, and Richard D. Leapman¶
From the §Laboratory of Parasitic Diseases, NIAID and ¶Biomedical Engineering and Instrumentation Program,National Center for Research Resources, National Institutes of Health, Bethesda, Maryland 20892
We measured the elemental content of different com-partments in Trypanosoma cruzi epimastigotes usingquick freezing, ultracryomicrotomy, and electron probemicroanalysis. Vacuoles identified by high electron den-sity contained (in units of mmol/kg dry weight 6 S.E.)large amounts of phosphorus (1390 6 13), magnesium(646 6 19), calcium (171 6 5), sodium (161 6 18), and zinc(148 6 6). No other compartment had appreciable cal-cium or zinc content. Iron (128 6 16 mmol/kg) was de-tected only in vacuoles distinct from the electron-densevacuoles and other organelles. Incubation of cells for 70min in culture medium in the presence of ionomycinplus nigericin led to a very significant 3- or 2-fold in-crease in potassium in the electron-dense vacuoles andthe iron-rich vacuoles, respectively, with no significantchange in the other elements investigated. This indi-cated the acidic nature of the vacuoles and demon-strated that the electron-dense vacuoles correspond towhat were described previously as acidocalcisomes,i.e. acidic compartments rich in Ca21. The acidocalci-somes were investigated by separation of epimastigotefractions on Percoll gradients in combination with Tri-ton WR-1339 treatment. This detergent caused a rapidvacuolation; these vacuoles were shown by electron mi-croscopy to be largely transparent, with a diffuse ma-trix. Percoll gradient fractionation demonstrated de-creases in the density of various organelle markers indetergent-treated cells compared with controls. Largedecreases in the density of the acidocalcisome and themitochondrion were seen, as well as smaller decreasesin the density of the other markers. Conventional elec-tron microscopy of epimastigotes loaded with gold-la-beled transferrin indicated that the endosomal systemwas separate from vacuoles that probably correspondedto the calcium-containing organelles detected by elec-tron probe microanalysis. The combined results provideevidence that acidocalcisomes are organelles differentfrom lysosomes or other organelles previously describedin these parasites.
Invasion of host cells by Trypanosoma cruzi, the etiologic
agent of Chagas’ disease, is dependent upon an elevation in theconcentration of cytosolic free calcium in the invading trypo-mastigote (1). Unlike mammalian cells, T. cruzi possesses mostof its intracellular Ca21 in an acidic compartment named theacidocalcisome (2). The biochemical characterization of thisorganelle has provided evidence that it is acidified by a vacu-olar-type proton-translocating (V-H1)-ATPase and that it has aCa21/H1 countertransporting ATPase for Ca21 uptake (2). Aci-docalcisomes have also been found in other trypanosomatidssuch as Trypanosoma brucei (3–5) and Leishmania mexicanaamazonensis (6) and in Toxoplasma gondii (7). This organelle isin various aspects similar to the vacuole present in fungi andplant cells (8) but apparently has no counterpart in animalcells.
Using fluorescence microscopy it was possible to detectbafilomycin A1-, nigericin-, and NH4Cl-sensitive acridine or-ange accumulation in round vacuoles of varying size locatedmainly to the posterior and anterior of T. cruzi epimastigotes(2) and T. brucei procyclic trypomastigotes (3), and these vacu-oles were suggested to correspond to the acidocalcisomes (2, 3).
Several authors (9–11) have reported the presence of calci-um-rich inclusion vacuoles in different trypanosomatids follow-ing energy-dispersive x-ray microanalysis. Vacuoles observedin Trypanosoma cyclops were tentatively termed polyphos-phate bodies on the basis of a high phosphorus content, anappearance similar to algal polyphosphate bodies and the priorreport of polyphosphates in other trypanosomatids. They hadan appreciable calcium content and, possibly, some zinc, al-though their composition was not quantified (9). T. cruzi epi-mastigotes were shown to possess electron-dense organellesidentified by scanning transmission electron microscopy ofwhole cells. These contained large amounts of magnesium,potassium, calcium, phosphorus, and zinc (10). Iron was alsofound in these studies although its co-localization with theother elements detected was not so evident. Electron probex-ray microanalysis of Leishmania major promastigotes dem-onstrated that their electron-dense vacuoles are also rich inphosphorus, magnesium, calcium, sodium, and zinc, with con-centrations of chloride and potassium below that of the cyto-plasm (11). Incubation of promastigotes in the presence orabsence of glucose did not cause significant changes in thevacuolar content of phosphorus, calcium, magnesium, or zinc,but changes in potassium and chloride were observed in bothelectron-dense vacuoles and in the cytoplasm (11).
In this work we used quick freezing, ultracryomicrotomy,and electron probe x-ray microanalysis (12–15) to study theelemental composition of electron-dense vacuoles in T. cruzi,with or without prior treatment with ionophores. Our resultssuggest that the electron-dense organelles previously identified
* This work was supported in part by Grant AI-23259 from theNational Institutes of Health (to R. D.). The costs of publication of thisarticle were defrayed in part by the payment of page charges. Thisarticle must therefore be hereby marked “advertisement” in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
‡ To whom correspondence should be addressed: Laboratory of Mo-lecular Parasitology, Dept. of Pathobiology, College of Veterinary Med-icine, University of Illinois, 2001 S. Lincoln Ave., Urbana, IL 61802.Tel.: 217-333-3845; Fax: 217-244-7421; E-mail: [email protected].
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 272, No. 44, Issue of October 31, pp. 28020–28029, 1997Printed in U.S.A.
This paper is available on line at http://www.jbc.org28020
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

in T. cruzi epimastigotes (10) correspond to acidocalcisomes (2).In addition, subcellular fractionation and gold-labeled trans-ferrin studies of T. cruzi epimastigotes provide evidence thatthese organelles are different from lysosomes or the prelysoso-mal vacuoles previously described as reservosomes (16).
EXPERIMENTAL PROCEDURES
Culture Methods—T. cruzi clone Silvio X10/4 (17) epimastigotes weremaintained at 26 °C in LIT medium (18) supplemented with 10% heat-inactivated fetal bovine serum, 100 units/ml penicillin, 100 mg/ml strep-tomycin, and 20 mg/ml hemin and harvested for electron probe x-raymicroanalysis in late exponential growth phase. T. cruzi Y strain (19)epimastigotes were grown at 26 °C in a liquid medium consisting ofbrain-heart infusion (37 g/liter), 100 units/ml penicillin, 100 mg/mlstreptomycin, hemin chlorohydrate (20 mg/liter dissolved in 50% tri-ethanolamine), and 10% heat-inactivated newborn calf serum (2). Fivedays after inoculation, Y strain epimastigote cells were collected bycentrifugation, washed twice with Dulbecco’s phosphate-buffered saline(138 mM NaCl, 2.7 mM KCl, 1.2 mM KH2PO4, 8.1 mM Na2HPO4, pH 7.2),and used for the fractionation studies.
Chemicals—Arsenazo III, fetal and newborn calf serum, nigericin,EGTA, sodium dithionite, leupeptin, RNase, DNase, glucose-6-phos-phate dehydrogenase (type XXIII), succinate, benzoyl-Pro-Phe-Arg-p-nitroanilide, p-nitrophenyl phosphate, gold chloride, polyethyleneglycol compound, and Triton WR-1339 were purchased from Sigma.5(6)-Carboxyfluorescein-N-hydroxysuccinimide ester and transferrin(bovine “holoenzyme”) were bought from Boehringer Mannheim; iono-mycin was from Calbiochem; Percoll was from Pharmacia Biotech Inc.,and silicon carbide was from Aldrich. All other reagents were analyticalgrade.
Elemental Microanalysis—Epimastigotes were harvested and resus-pended at 2 3 108 cells/ml in fresh growth medium (LIT) to maintaintheir morphology. Portions of the suspension were incubated in theabsence (control) or presence of 2 mM ionomycin plus 4 mg/ml nigericinfor 70 min at room temperature (22 °C). After incubation the cells wereconcentrated by centrifugation, washed once in LIT plus 0.5% fetal calfserum, and centrifuged for 1 min at 14,000 3 g in an Eppendorfcentrifuge to compact the pellet. The supernatant liquid was removed,and the cells were transferred by Pipetman to the end of wooden sticksfor freezing in liquid ethane. The cells were quick frozen by plungingthem into liquid nitrogen-cooled liquid ethane at approximately2185 °C in a Leica KF80 plunge freezing device (Leica, Vienna, Aus-tria); frozen blocks were subsequently stored under liquid nitrogen.Cryosectioning was performed on a Leica Ultracut/E ultramicrotomeequipped with an FC-4E cryosectioning attachment and an ionizationantistatic device (Hauf Static Line, Diatome). Blocks were first trimmedto ;200 3 300 mm using a diamond trimming tool, and then frozen thinsections were cut at a specimen temperature of 2160 to 2165 °C usinga 35° diamond knife kept at a temperature of 2155 °C. Sections fromeach ribbon cut by the microtome were collected by means of an eyelashprobe and placed onto a hexagonal 400-mesh Formvar/carbon-coatedgrid. This was covered with a second coated grid, sandwiched betweenthe leaves of a small indium foil envelope, and pressed to attach thesections to the support film (20). Specimens were stored in grid boxesunder liquid nitrogen. Sections were analyzed in a VG MicroscopesHB501 scanning transmission electron microscope (VG Scientific, Bev-erly, MA) as described previously (21). Grid pairs were removed fromstorage and were separated under liquid nitrogen. A single frozenhydrated grid was cryotransferred, freeze-dried at about 2110 °C, andsubsequently re-cooled to approximately 2160 °C for analysis. Darkfield images were recorded digitally at low electron dose (,103 e/nm2)by means of a Gatan Digiscan system (Gatan Inc., Pleasanton, CA)interfaced to an Apple Macintosh Quadra 950 computer. Images wereanalyzed with the Gatan Digital Micrograph program. Energy-disper-sive x-ray spectra were acquired with a Tracor Northern TN 5500multichannel analyzer and Micro-ZHV ultrathin window detector (No-ran, Middleton, WI) having a collection solid angle of 0.18 sterad. Theacquisition time for each spectrum was 100 s and the probe current ;2nA. Square regions were analyzed with dimensions of ;200 nm fornuclei, cytoplasm, iron-rich vacuoles, and kinetoplast and ;100 nm formitochondria and calcium-rich vacuoles. The electron probe was ras-tered at high magnification during acquisition to ensure that there wasno specimen drift. Spectra were transferred to a Quadra 950 computerand were processed using the program Desktop Spectrum Analyzer(22). A Simplex non-linear least squares fitting routine and the Hallpeak/continuum method were used to quantify the elemental composi-tion in terms of millimoles per kg dry weight (14). Standards were
prepared by analyzing dried cryosections of rapidly frozen solutionscontaining known concentrations of salts and polyvinylpyrrolidone asan organic matrix. Iron and zinc were quantified by calculating sensi-tivity factors relative to potassium with the Desktop Spectrum Ana-lyzer program. Electron energy loss spectra (EELS)1 were recorded witha Gatan model 666 parallel detection spectrometer (23) and the GatanEL/P acquisition program running on a Quadra 950 computer (24).EELS spectra were acquired with an integration time of 0.3 s and 50read-outs of the photodiode array; the electron dose was approximately105 e/nm2. Estimated ratios of nitrogen, oxygen, phosphorus, and cal-cium in the calcium-rich vacuoles were obtained by means of the EELSquantitative analysis program in the EL/P software. Spectra were col-lected from granules with mass thickness below one inelastic mean freepath at 100 keV beam energy to avoid background subtraction errorsdue to plural inelastic scattering (25, 26).
Cell Loading with Triton WR-1339 and Surface Labeling—Epimas-tigotes collected from culture by centrifugation were resuspended atabout 3 3 concentration (approximately 108 cells/ml) in Dulbecco’sphosphate-buffered saline, pH 7.2, containing 10 mg/ml Triton WR-1339 and incubated at 30 °C for 50 min. This resulted in, on average,84% vacuolation in treated cells versus less than 5% in untreated cells,as observed by light microscopy (n 5 8). Cells were recovered by cen-trifugation, resuspended in 3 ml of 20 mM Hepes, 5 mM KCl, 125 mM
NaCl, pH 8.0, with 0.53 mM 5(6)-carboxyfluorescein-N-hydroxysuccin-imide ester and left on ice for 20 min to label surface proteins. Thereaction was stopped by dilution in the same buffer containing 5 mg/mlbovine serum albumin.
Preparation of Cell Fractions—Epimastigotes (about 1 3 1010 foreach gradient) were centrifuged and washed once in lysis buffer (20 mM
Hepes, 40 mM KCl, 125 mM sucrose, 1 mg/ml bovine serum albumin, 5mM sodium dithionite, 0.5 mM EDTA, 50 mg/ml DNase, 50 mg/ml RNase,and 2 mM leupeptin). The cell pellet was mixed with 1.5 3 wet weightsilicon carbide and ground with a mortar and pestle until lysis wasgreater than 90% (generally 40 s; Ref. 27). Silicon carbide and unlysedcells were removed by centrifugation twice at 150 3 g for 5 min. Thesupernatant fraction was centrifuged for 40 min at 105,000 3 g. Thepellet was recovered in resuspension buffer (lysis buffer without DNase,RNase, or leupeptin) with passage several times through a 23-gaugeneedle. This fraction was applied to an 18-ml density gradient contain-ing 34% Percoll, 34% 0.5 M sucrose, and 32% resuspension buffer andcentrifuged for 50 min at 69,500 3 g (max) in a Beckman 70Ti rotor.Gradients were fractionated by upward displacement with 2.25 M
sucrose.Assay for Acidic Calcium Storage—Calcium accumulation in the
presence of ATP was assayed by an end point assay based on previousmethods (2–4). Fractions were incubated in a mixture of 10 mM Hepes,65 mM KCl, 125 mM sucrose, 2 mM MgCl2, 50 mM EGTA, 90 mM EDTA,80 mM CaCl2 (free Ca21 4 mM; Ref. 2), 1 mM ATP, 2 mg/ml oligomycin, 1mg/ml antimycin A, 1 mM ionomycin, 40 mM arsenazo III, pH 7.2. Theoligomycin and antimycin A prevented uptake of calcium into mitochon-dria, and the ionomycin released calcium from other non-acidic com-partments (2–4). After 15 min incubation at 30 °C in a water bath,mixtures were transferred into an SLM-Aminco DW2000 dual wave-length spectrophotometer, and the calcium accumulated in acidic com-partments was released by the addition of 5 mM nigericin (measuredwith arsenazo III at 675–685 nm; Ref. 2).
Subcellular Compartment Marker Assays—The following marker as-says were used: for lysosomes, acid phosphatase, a-mannosidase, andcysteine proteinase; for mitochondria, NADH-succinate-cytochrome creductase; for glycosomes, hexokinase; for cytosol, glucose-6-phosphatedehydrogenase; for cell surface, measurement of surface fluorescentlabeling. Acid phosphatase, a-mannosidase, and acid protease activitieswere assayed by modifications of previous methods using 96-well mi-crotiter plates (28). For acid phosphatase, the sample (20 ml) wasincubated for different periods at 30 °C with 50 ml of 10 mM p-nitrophe-nyl phosphate in 0.1 M sodium acetate, pH 5.5, before the reaction wasstopped by the addition of 100 ml of 150 mM sodium hydroxide. a-Mannosidase was assayed similarly with 5 mM p-nitrophenyl a-D-man-nopyranoside in 0.1 M potassium Mes, pH 5.5, with 60 mM sodiumhydroxide as the stop solution. Samples for protease assay were mixedwith 0.1 M potassium Mes, pH 5.5, spun briefly in a microcentrifuge,and the supernatant fraction used in the assay, with the addition of 5mM dithiothreitol and 0.1 mM benzoyl-Pro-Phe-Arg-p-nitroanilide. Theplate was incubated at 30 °C and read intermittently. Under these
1 The abbreviations used are: EELS, electron energy loss spectra;Mes, 4-morpholineethanesulfonic acid.
Analysis of Acidocalcisomes in T. cruzi 28021
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

assay conditions, it was confirmed that the protease activity was sen-sitive to 40 mM trans-epoxysuccinyl-L-leucylamido-(4-guanidino)-butane(E64; results not shown) and therefore corresponded to the lysosomalcysteine proteinase cruzipain (29, 30) and not the cytosolic alkalineprotease that is insensitive to this inhibitor (31). The release of p-nitrophenol or p-nitroaniline from substrates was measured using aDynatech MR5000 reader at 410–450 nm. Possible surface localizationof these enzymes was tested by assaying intact cells in reaction mix-tures as above, followed by removal of cells, re-incubation of the assaymixture to determine released activity, and lysis of cells (by freeze-thawin dry ice/ethanol) and assay of lysates to determine total activity. Thecell surface was labeled with a fluorescein reagent as described above,and fractions were assayed by 20-fold dilution in water followed bymeasurement of fluorescence (lex 490 nm; lem 520–460 nm) in a Hita-chi F-2000 fluorimeter. NADH-succinate-cytochrome c reductase activ-ity was determined by the method used in Ref. 32, using succinate asthe substrate and following the reaction at 550–540 nm in the dualwavelength spectrophotometer. This enzyme has previously been de-tected in T. cruzi epimastigote mitochondrial fractions, using NADH assubstrate (33). Hexokinase (34) was assayed at 30 °C in a mixture of 10mM D-glucose, 0.6 mM ATP, 0.6 mM NADH, 10 mM magnesium chloride,2.5 units/ml glucose-6-phosphate dehydrogenase, 50 mM potassiumHepes, pH 7.8. The oxidation of NADH was monitored in a dual wave-length spectrophotometer at 340–430 nm. Glucose-6-phosphate dehy-drogenase was assayed as per hexokinase, with the omission of glucose,ATP, and added glucose-6-phosphate dehydrogenase, and with the ad-dition of 0.1 mM glucose 6-phosphate. All marker enzymes were assayedfollowing freezing of fractions (220 °C, at least overnight).
Labeling of Epimastigotes with Gold-Transferrin—Colloidal gold (15nm diameter) was prepared by the method of Frens (35). Gold suspen-
sion (0.1 mg/ml, pH 6.0) was added to transferrin to give a final trans-ferrin concentration of 25 mg/ml (optimal concentration determined asper Ref. 36). Colloidal gold-transferrin was recovered and washed bycentrifugation (64,000 3 gmax; 30 min) in 0.6 mg/ml polyethylene glycolcompound, 20 mM Mes, pH 6.0. Cells were washed 3 times in Dulbecco’sphosphate-buffered saline, resuspended in 116 mM NaCl, 5.4 mM KCl,0.8 mM MgSO4, 50 mM Hepes, 5.5 mM glucose, pH 7.2 at 3 3 107 cells/ml,and loaded with the gold complex for 1–8 h at 30 °C, with the amountof gold added being equivalent to a final A520 of 0.2–0.3, before fixationfor electron microscopy.
Conventional Electron Microscopy—Control, Triton WR-1339-treated, and gold-transferrin-loaded epimastigotes were fixed in 2.5%glutaraldehyde, 0.1 M K1-Hepes buffer, pH 7.5, for 40 min at roomtemperature and resuspended in 0.1 M K1-Hepes buffer, pH 7.5, 50 mM
sucrose, before post-fixation in 2% OsO4 in water using a microwaveoven (Ted Pella 3440): two treatments at 42 °C (max), each 8 s on, 20 soff, and 8 s on, with a 15-min incubation at room temperature after eachtreatment. Specimens were then stained en bloc with 3% potassiumferricyanide for 10 min, rinsed in water, and incubated in saturateduranyl acetate for 30 min (sample in uranyl acetate was microwaved atthe beginning of the incubation), dehydrated in ethanol/propylene ox-ide, and embedded in Lx112 Epon substitute. Ultrathin sections weremade in a Reichert Ultracut E ultramicrotome, stained with uranylacetate and lead citrate, and observed in a Jeol 100CX electron micro-scope operating at 80 kV. For imaging whole epimastigotes, these weresuspended in Dulbecco’s phosphate-buffered saline, pH 7.4. Drops wereapplied to Formvar-coated grids, and cells were allowed to adhere for 10min and then were carefully blotted dry and observed directly with theJEOL electron microscope.
Statistical Analysis—Elemental concentrations are expressed as
FIG. 1. Transmission electron mi-crograph of epimastigotes. Unstainedcells were prepared by quick freezing andultracryomicrotomy as described under“Experimental Procedures.” The cellularcompartments that were analyzed by en-ergy-dispersive x-ray microanalysis in-cluded the nucleus (N), mitochondria (M),kinetoplast-DNA (K), electron-densevacuoles (arrows), iron-rich vacuoles (Fe),and cytoplasm (C). Note that a singlemembrane surrounds some of the elec-tron-dense vacuoles (arrowheads). Insetsshow 2 3 magnifications of a calcium-richvacuole showing a limiting membrane(upper left) and an iron-containing vacu-ole (lower left). Bar 5 1 mm.
Analysis of Acidocalcisomes in T. cruzi28022
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

means 6 S.E. for n measurements. Statistical significance betweendifferent treatments was determined by Student’s t test. Significancewas considered at the p , 0.001 level for potassium changes in thecalcium-rich vacuoles and at the p , 0.01 level for potassium changes inthe iron-rich vacuoles.
RESULTS
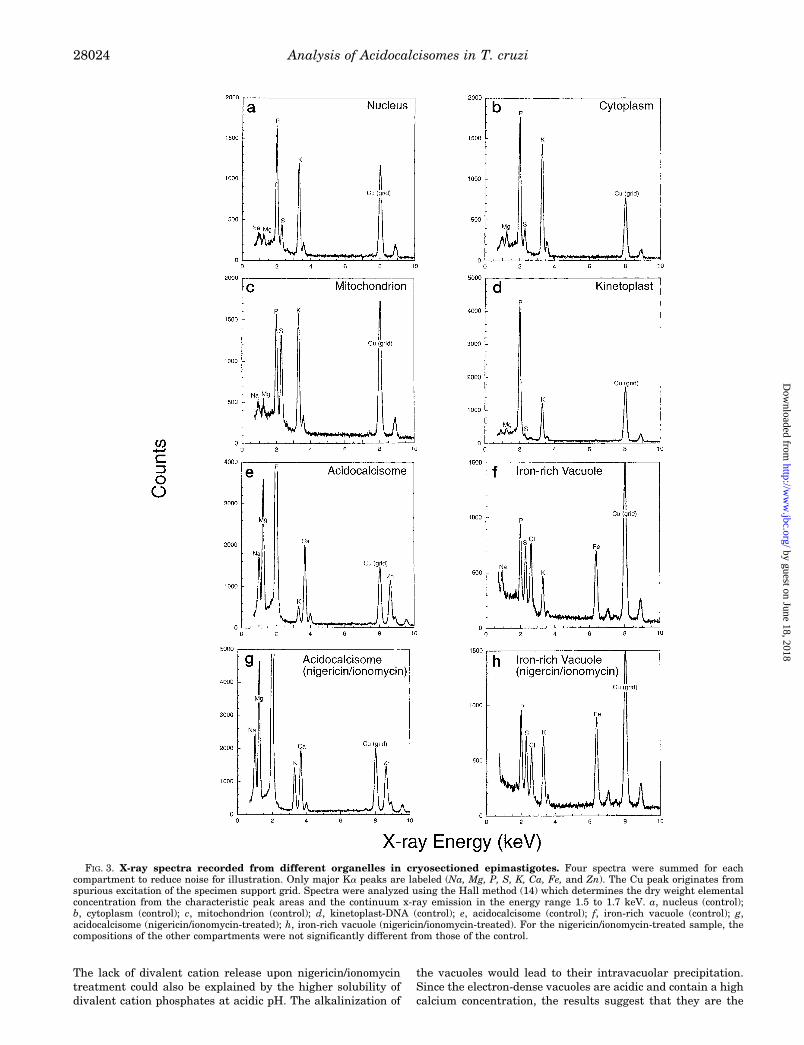
Elemental Analysis of T. cruzi Epimastigotes—Scanningtransmission electron micrographs of rapidly frozen, cryosec-tioned, and freeze-dried epimastigotes showed very good ultra-structural preservation (Fig. 1). Easily identified structuresinclude the mitochondrion with the kinetoplast-DNA, subpel-licular microtubules, flagellar microtubules, the nucleus, and(not shown in this figure) endoplasmic reticulum and the Golgi.The contrast of a given structure in the image arises solely fromits mass density since these preparations were not stained. Inaddition, there were various organelles unidentifiable by theirappearance, in comparison with conventional (chemicallyfixed) electron microscopy preparations (see below). The mostprominent of these were spherical vacuoles of high mass den-sity, as reflected by their marked electron opacity comparedwith other compartments (Fig. 1, arrows). These electron-densevacuoles have an average diameter of 200 6 90 nm (6 S.D.). Asingle membrane surrounding the electron-dense material wasclearly seen in some of these vacuoles (Fig. 1, arrowheads, andupper left inset). In each longitudinal cryosection 1–4 of thesevacuoles were found per cell. Since the average cross-section ofan epimastigote is about 3–4 mm, and the section thicknesswas approximately 100 nm, there are 30–40 longitudinal sec-tions per cell. Therefore a 200-nm vacuole would be visible in2–3 sections; the range in vacuole number per cell is 10–60,with an average of 30. Similar electron-dense vacuoles wereseen when whole epimastigotes were observed by transmissionelectron microscopy without fixation (Fig. 2). More than 40electron-dense vacuoles of varying sizes are observed in each ofthe epimastigotes shown in Fig. 2. X-ray spectra were acquiredfrom nuclei, mitochondria, kinetoplast-DNA (dense region),electron-dense vacuoles, iron-rich vacuoles, and cytoplasmicspaces of cryosections. Typical x-ray spectra from these com-partments are shown in Fig. 3, a–f, for the control and Fig. 3, gand h, for the nigericin/ionomycin-treated sample (see below).Quantification of spectra from nuclei (Fig. 3a), cytoplasm (Fig.3b), and mitochondria (Fig. 3c) gave similar ion concentrationsas in many other cell types (e.g. Refs. 12–15). Quantitativeanalysis of the electron-dense vacuoles indicated that theirmass density was due to very high amounts of phosphorus,magnesium, calcium, sodium, and zinc (Fig. 3, e and g). Theelectron-dense vacuoles were the only compartment withmarked calcium and zinc content, whereas iron was only de-tected in larger vacuoles that were less electron-dense (Fig. 1,
lower left inset, and Fig. 3, f and h). It is important to mentionthat x-ray microanalysis measures total calcium rather thanfree calcium.
The elemental content of the various compartments exam-ined in the control cells was determined in units of millimole/kgdry weight, and the results of one experiment are shown inTable I, although analysis was performed on samples from twoindependent experiments. Results were comparable exceptthat the sodium content in most organelles, and the calciumcontent of the calcium-rich vacuoles, was about 2-fold higher inone experiment (data not shown). Generally, in the electron-dense vacuoles, potassium and chloride levels were lower thanin the cytoplasm. The low sulfur level in these vacuoles indi-cates that they do not contain much cysteine or methionine orproteins containing these amino acids. Analysis of EELS (Fig.4) from 11 electron-dense vacuoles in nigericin/ionomycin-treated cells (see below) yielded a phosphorus-to-oxygen ratioof 0.11:1 (615% S.E.) and a nitrogen-to-oxygen ratio of 0.065:1(613% S.E.). Because spectra were acquired at a relativelyhigh dose of 105 e/nm2, it is likely that significant oxygen waslost due to beam damage, so these measured P:O and N:Oratios represent upper limits. As discussed later, the resultssuggest high levels of carbohydrate in the vacuolar matrix.
Changes in the Elemental Composition of Epimastigotes afterIncubation in the Presence of Ionomycin and Nigericin—Previ-ous studies (2) demonstrated that acidic calcium storage or-ganelles in T. cruzi (acidocalcisomes) are the most importantCa21-containing compartments in these parasites. In the pres-ence of EGTA to prevent refilling of internal stores, Ca21 can bereleased from acidocalcisomes by combined treatment with acalcium ionophore, such as ionomycin, and a K1/H1 exchangersuch as nigericin. This is because ionomycin binds essentiallyno calcium below pH 7.0 and cannot transport Ca21 out ofacidic compartments because of competition from protons atthe inside face of the membrane (37). In the absence of nigeri-cin, ionomycin releases a relatively small amount of Ca21 onlyfrom neutral or alkaline compartments in T. cruzi but releasesmuch more Ca21 after nigericin has elevated the pH of acidiccompartments. To determine whether treatment with ionomy-cin plus nigericin could alter the elemental composition of theelectron-dense vacuoles and demonstrate that they correspondto what has been described before as acidocalcisomes (2), weincubated epimastigotes in the presence of these ionophores.However, to preserve their morphology we performed theseincubations in culture medium, which contains an appreciableamount of Ca21 (about 1.2 mM), and therefore, we looked forchanges in other elements that could indicate the acidic natureof these vacuoles. After 70 min incubation in the presence ofionomycin plus nigericin, there was a marked 3-fold increase inpotassium in the electron-dense vacuoles, with no change in theother elements investigated (Fig. 3g and Table II; the potas-sium content of the calcium-rich vacuoles was about 50%higher in a second experiment, after ionophore treatment;other elements were at similar concentrations in the two ex-periments, except as noted above for control cells). Since nigeri-cin is a K1/H1 exchanger this increase in potassium could bedue to the vacuoles being acidic. An increase in potassium alsooccurred in the iron-rich vacuoles (Fig. 3h and Table II) but notin the mitochondria or nuclei. These data suggest that theelectron-dense vacuoles and the iron-rich vacuoles were theonly acidic compartments of those investigated. The lack ofchange in calcium concentration in the electron-dense vacuolescould be attributed to refilling by extracellular calcium. Thelevel of cytoplasmic calcium did not change but remained low,implying the operation of calcium pumps (in the vacuoles andin the plasma membrane) even in the presence of ionomycin.
FIG. 2. Transmission electron micrograph of whole epimasti-gotes. Whole, unstained cells were prepared as described under “Ex-perimental Procedures.” Note the numerous electron-dense vacuoles ofdifferent size. Bar 5 500 nm.
Analysis of Acidocalcisomes in T. cruzi 28023
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from
The lack of divalent cation release upon nigericin/ionomycintreatment could also be explained by the higher solubility ofdivalent cation phosphates at acidic pH. The alkalinization of
the vacuoles would lead to their intravacuolar precipitation.Since the electron-dense vacuoles are acidic and contain a highcalcium concentration, the results suggest that they are the
FIG. 3. X-ray spectra recorded from different organelles in cryosectioned epimastigotes. Four spectra were summed for eachcompartment to reduce noise for illustration. Only major Ka peaks are labeled (Na, Mg, P, S, K, Ca, Fe, and Zn). The Cu peak originates fromspurious excitation of the specimen support grid. Spectra were analyzed using the Hall method (14) which determines the dry weight elementalconcentration from the characteristic peak areas and the continuum x-ray emission in the energy range 1.5 to 1.7 keV. a, nucleus (control);b, cytoplasm (control); c, mitochondrion (control); d, kinetoplast-DNA (control); e, acidocalcisome (control); f, iron-rich vacuole (control); g,acidocalcisome (nigericin/ionomycin-treated); h, iron-rich vacuole (nigericin/ionomycin-treated). For the nigericin/ionomycin-treated sample, thecompositions of the other compartments were not significantly different from those of the control.
Analysis of Acidocalcisomes in T. cruzi28024
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

acidocalcisomes.Separation of T. cruzi Organelles: Effects of Density Modifi-
cation Treatments—The nature of the acidocalcisomes was in-vestigated by separation of cell fractions on Percoll gradients.Fractions obtained from untreated T. cruzi epimastigotes didnot show much separation in density, the exception being theglycosomal marker hexokinase, which was recovered fromdenser fractions, compared with other markers (Fig. 5, shadedbars; the cytosolic marker glucose-6-phosphate dehydrogenaseis not shown. Where detectable, it was found only in fractions1–3.)
Treatment of T. cruzi epimastigotes with 10 mg/ml TritonWR-1339 caused rapid vacuolation of the cells. After 50 minincubation at 30 °C, typically 75–100% of cells were visiblyvacuolated as observed by phase contrast microscopy (notshown). Electron microscopy (Fig. 6b) revealed the vacuoles tobe largely transparent, with a diffuse matrix. The kinetoplast-DNA was typically seen in a vacuole. Some of the vacuoles werebounded by a double membrane (indicative of a mitochondrion)but in other cases it was not possible to see a delimiting mem-brane. The morphology of reservosomes (multivesicular bodies)(16) appeared unaffected by WR-1339 treatment (Fig. 6, a andb). Control cells (Fig. 6a, arrows) showed vacuoles of similarsize and appearance to the electron-dense vacuoles observed incryosections (Fig. 1) but devoid of electron-dense material or
with only some electron-dense material attached to their mem-branes. After Triton WR-1339 treatment similar vacuoles wereobserved (Fig. 6b, arrows). However, it is possible that some ofthe bigger vacuoles not bound by a double membrane mightcorrespond to the electron-dense vacuoles. Separation of ex-tracts of treated cells on Percoll gradients demonstrated vary-ing shifts (decreases) in the density of organelle markers (Fig.5, stippled bars). Large decreases in the density of the acido-calcisome and the mitochondrion (indicated by succinate cyto-chrome c reductase activity) were consistently seen in the ex-periments performed, as were smaller decreases in the densityof the other markers. The minor influence on the density of thelysosomal markers, a-mannosidase, acid phosphatase, and cys-teine proteinase was surprising, given the original use of TritonWR-1339 as an agent to isolate lysosomes via density reduction(38). The recovery of these enzyme activities was similar fromTriton-treated and control cells (average relative recovery ofenzymes, treated versus untreated, ranged from 104 to 127%,except for succinate cytochrome c reductase, 82%; this com-pared with 108% for calcium release from acidic compart-ments). It was possible that hydrolytic enzymes might alsooccur in other subcellular locations, particularly the cell sur-face. Surface location of acid phosphatase and a-mannosidasewas tested by comparative assays of whole cells and lysates. Anaverage of only 9% acid phosphatase activity (three experi-ments, range 4–16%) and 1% a-mannosidase activity (two ex-periments, both 1%) was on the cell surface; consequently,surface enzyme was not contributing greatly to the observeddensity profiles of these enzymes.
Labeling of Epimastigotes with Transferrin-Gold—Epimas-tigotes of T. cruzi bound and internalized gold-labeled trans-ferrin (Fig. 7). Bound transferrin accumulated in an invagina-tion of the plasma membrane, the cytostome, involved inendocytosis (Fig. 7a, C) (39). Internalized transferrin was ob-served in vacuoles of varying size, some with the heterogeneouscontent and multivesicular appearance typical of the pre-lyso-somal compartment described as the reservosome (Fig. 7b, R),and others with the typical appearance of lysosomes (Fig. 7b,L), but was not observed in the nearly empty vacuoles thatcorresponded in size to the calcium-containing vacuoles (arrow-heads in Fig. 7b; compare with Fig. 6a and Fig. 1). The micro-graphs shown in Fig. 7 were taken from cells incubated for 1 hwith transferrin-gold, but similar results were obtained withcells incubated for 8 h.
DISCUSSION
Elemental Composition of T. cruzi Epimastigotes—The aimof this study was to characterize morphologically the acidocal-cisomes and establish their distinct nature with respect tolysosomes and other intracellular organelles. By using electron
TABLE IElemental analysis of different compartments in T. cruzi control
Numbers indicate concentration (mean 6 S.E.).
Compartment Sodium Magnesium Phosphorus Sulfur Chloride Potassium Calcium Iron Zinc
mmol/kg dry wtNucleus 59 6 15 63 6 5 503 6 33 95 6 6 12 6 2 308 6 17 0.8 6 0.7
(n 5 17)Cytoplasm 28 6 18 74 6 9 522 6 35 95 6 6 22 6 5 346 6 23 0.3 6 2.0
(n 5 19)Mitochondrion 48 6 9 40 6 3 291 6 30 170 6 10 21 6 5 225 6 13 1.0 6 0.6
(n 5 21)Kinetoplast 29 6 3 52 6 4 796 6 33 21 6 6 11 6 3 206 6 12 0.4 6 0.5
(n 5 14)Calcium-rich vacuole 161 6 18 646 6 19 1390 6 13 10 6 1 2 6 1 37 6 2 171 6 6 148 6 6
(n 5 58)Iron-rich vacuole 57 6 10 6 6 2 199 6 9 135 6 11 183 6 17 72 6 6 20.5 6 0.6 128 6 16
(n 5 11)
FIG. 4. Electron energy loss spectrum from electron-dense vac-uole (acidocalcisome). Ionization edges of phosphorus (L23), carbon(K), calcium (L23), nitrogen (K), and oxygen (K) are indicated. Spectrawere analyzed according to Leapman and Ornberg (26).
Analysis of Acidocalcisomes in T. cruzi 28025
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

probe x-ray microanalysis we found that the only organellescontaining a measurable amount of calcium were electron-dense vacuoles. A few previous publications report the use ofthis technique with trypanosomatids. The earliest study, on T.cyclops (9), was not quantitative, but the results are neverthe-less similar to those found here; calcium and possibly zinc weredetected in the “polyphosphate” granules. The prior work on T.cruzi (10), although done on whole cells rather than sections,matches the present data. The only previous analysis of un-fixed sections of a trypanosomatid (11) provided data on the
elemental composition of calcium-rich vacuoles and cytoplasmin L. major. Their results are similar to ours, except that wefound chloride levels 4–10-fold lower in both compartments,potassium about 10-fold lower in the calcium vacuoles (untreat-ed cells), and calcium about 10-fold lower in the cytoplasm, and6–8-fold higher in the vacuoles. The apparent enrichment ofcalcium in these compartments is therefore much more markedin our study. Vacuoles similar in both appearance and elemen-tal composition occur in the slime mold Dictyostelium discoi-deum (40). The vacuolar content of phosphorus, magnesium,
TABLE IIElemental analysis of different compartments in T. cruzi treated with nigericin/ionomycin
Numbers indicate concentration (mean 6 S.E.).
Compartment Sodium Magnesium Phosphorus Sulfur Chloride Potassium Calcium Iron Zinc
mmol/kg dry wt
Nucleus 63 6 15 68 6 4 593 6 43 86 6 10 12 6 3 292 6 9 1.5 6 0.5(n 5 17)
Cytoplasm 30 6 14 99 6 7 579 6 18 87 6 4 24 6 3 346 6 17 1.2 6 0.8(n 5 20)
Mitochondrion 40 6 12 37 6 4 258 6 15 168 6 5 24 6 4 228 6 14 1.7 6 0.5(n 5 21)
Kinetoplast 45 6 15 61 6 5 818 6 30 20 6 5 12 6 3 179 6 10 1.3 6 0.2(n 5 15)
Calcium-rich vacuole 170 6 12 738 6 20 1430 6 14 9 6 1 2 6 1 92 6 7 167 6 7 131 6 6(n 5 63)
Iron-rich vacuole 35 6 11 4 6 3 202 6 24 145 6 6 161 6 33 147 6 26 20.9 6 1.3 182 6 14(n 5 10)
FIG. 5. Distribution on Percoll gra-dients of organellar markers fromepimastigotes with (stippled bars)or without (shaded bars) TritonWR-1339 treatment. y axis (except fordensity graph) indicates relative distribu-tion (total 5 500 for each gradient); x axisindicates fraction number (1 5 lightest).Bars show means from five experiments(four for surface labeling). Density meas-ured in one experiment, using Percolldensity marker beads.
Analysis of Acidocalcisomes in T. cruzi28026
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

and calcium increased substantially during aggregation ofslime mold amoebae, passing through the range of concentra-tions observed here. Zinc was not detected in the slime mold.The sulfur content in these organelles was 10-fold higher thanreported here, suggesting a higher protein content. The au-thors proposed that the phosphate in the vacuoles might be in
the form of inositol phosphates, rather than polyphosphate.EELS measurements on the electron-dense vacuoles indi-
cated the presence of a major organic component. If the vacu-olar matrix was composed only of simple phosphates orpolyphosphates the expected phosphorus-to-oxygen ratio wouldbe 0.25:1 or 0.33:1, respectively, compared with the measuredratio of 0.11:1 (615% S.E.). Therefore, at least half of theoxygen comes from organic species. The low measured nitro-gen-to-oxygen ratio of 0.065:1 (613% S.E.) suggests that themajor organic component is carbohydrate rather than protein,which is also consistent with the very low sulfur in the electron-dense vacuoles. An additional clue about the composition comesfrom the fact that the vacuolar matrix “bubbled” under electronirradiation; such bubbling is well-known to occur in sugars(41). It therefore appears likely that a substantial fraction ofthe vacuolar mass is due to sugar molecules. The small amountof nitrogen detected in the electron-dense vacuoles could orig-inate from membrane proteins included in the analyzed re-gions. Our x-ray measurements of monovalent (Na1 and K1)and divalent (Mg21, Ca21, and Zn21) cation concentrationsallow us to determine the ratio of cation charges per phosphate;the proton concentration is ,1mM and can therefore be ignored.From Tables I and II and the requirement of charge neutrality,we find that there are 1.6 6 0.1 (6 S.E.) negative charges perphosphate. This value falls between a polyphosphate composi-tion that would give 1.0 negative charges per phosphate and apyrophosphate composition that would give 2.0 negativecharges per phosphate. Our measurements would be consist-ent, for example, with sugar diphosphates that have 1.5 nega-tive charges per phosphate. Interestingly, a-glucose 1,2,3,4,6-pentakis(diphosphate) has recently been identified as a novelmetabolite present in phosphorus-, magnesium-, and potassi-um-containing electron-dense bodies of the sulfate-reducingbacterium Desulfovibrio gigas (42). The nature of the phospho-rus-containing metabolite(s) present in T. cruzi epimastigotesis currently being investigated. The presence of calcium-, mag-nesium-, and phosphorus-containing vacuoles in yeast (43) andpossibly also Chlorella (44) further illustrates the importanceof these organelles in phylogenetically diverse organisms.
T. cruzi is an obligate intracellular parasite, living in hostcell cytoplasm. The high concentration of calcium in an intra-cellular store could indicate either an adaptation to the cyto-plasmic environment where the free Ca21 concentration is verylow (of the order of 1027 M) or its requirement for signalingduring host cell invasion (1).
Although over 300 enzymes have been identified represent-ing more than 50 different types that are known to require zincfor their function (45), little is known about zinc cell contentand free concentration (46). In addition to its accumulation inall trypanosomatids so far examined (see above), zinc is alsopresent in granules in pancreatic beta cells (47) and in certainneurons in the brain, especially in areas of the hippocampus(48), although the reason for its accumulation in any of thesecompartments is unknown.
A distinctive organelle having an average diameter of 420 670 nm (6 S.D.) and much less electron density than the calci-um-rich vacuole was found in many cell cross-sections (Fig. 1and Table I); this organelle was notable for its high iron andchloride content (Fig. 3, f and h). Previously, elemental map-ping of whole epimastigotes (10) indicated substantial differ-ences in the iron content of different strains of T. cruzi, whichcould be reproducibly observed by eye as differences in the colorof pellets of cells from the different strains (little iron, white;more iron, brown). T. cyclops contains a prominent iron-con-taining vacuole (9), visible even with the light microscope al-though not when it is grown in the absence of hemoglobin. The
FIG. 6. Effects of Triton WR-1339 on the morphology of epimas-tigotes. a, control cells chemically fixed as described under “Experi-mental Procedures.” Arrows show vacuoles that may correspond to theelectron-dense vacuoles. Note electron-dense material attached to themembrane in some of them; N, nucleus; M, mitochondrion; C, cytos-tome; K, kinetoplast-DNA; R, reservosomes. Bar 5 1 mm. b, TritonWR-1339-treated cells. Note the kinetoplast-DNA (K) inside a vacuole,vacuoles similar to those present in control cells (arrows), and reservo-somes (R) with unchanged morphology. Bar 5 1 mm.
FIG. 7. Labeling of epimastigotes with transferrin-gold. Epi-mastigotes were incubated for 1 h with gold-labeled transferrin asindicated under “Experimental Procedures.” Bound transferrin accu-mulated in the cytostome (C in a) and in vacuoles of varying size, somewith appearance typical of reservosomes and lysosomes (R and L in b),but was not observed in the nearly empty vacuoles which correspondedin size to the calcium-containing vacuoles (arrowheads in B). N, nu-cleus. Bars 5 0.5 mm.
Analysis of Acidocalcisomes in T. cruzi 28027
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

vacuoles in T. cruzi may represent smaller versions of thesame, and the iron may be present as degradation products ofhemin, an essential component of the epimastigote growthmedium. Therefore, these organelles may represent a compo-nent of the endo-/lysosomal system in this organism and couldcorrespond to reservosomes (16) that appear as multivesicularbodies using chemical fixation methods (see Figs. 6 and 7).Reservosomes are of similar size to the iron-rich vacuoles andare known to be acidic (49) and to contain cysteine protease(50). Use of energy-dispersive x-ray microanalysis associatedwith transmission electron microscopy showed that anothertrypanosomatid, Herpetomonas samuelpessoai, possessed elec-tron-dense granules containing iron (51), similar to those in T.cyclops (9).
Treatment of epimastigotes with ionomycin and nigericinmarkedly increased the potassium content of the electron-dense vacuoles thus indicating their acidic nature. This makesthem fulfill the two conditions for the definition of acidocalci-somes, acidity with a high calcium content.
A small but measurable amount of sulfur was found associ-ated with the kinetoplast-DNA (Fig. 3d and Table I). The ratioof P:S in the kinetoplast was approximately 40:1 comparedwith about 5:1 in the nucleus. This provides evidence that thereis a small amount of protein bound to the kinetoplast-DNA.Although initial cytochemical studies (52) suggested that, likemitochondrial DNA, the kinetoplast-DNA was not associatedwith proteins, later ultrastructural studies demonstrated theirpresence (53), and some kinetoplast-DNA-associated basic pro-teins have recently being identified in Crithidia fasciculata(54).
Effects of Triton WR-1339 Treatment on T. cruzi Epimasti-gotes—Triton WR-1339 was the original reagent used to modifythe density of lysosomes, and its effects on rat liver wereextensively characterized (38). It was also injected into ratsinfected with T. brucei (55), where it produced large vacuoles inthe trypanosomes. Parasite proteinase activity was largely re-covered from density gradients in the position of cytosolic pro-teins, suggesting the lysosomal membrane had been weakenedby the treatment, although there was only a small decrease inthe density of recovered a-mannosidase activity. The position ofnon-lysosomal enzymes in density gradients was not reported(55). We found, in T. cruzi epimastigotes, no change in thedensity of lysosomal markers (and no reduction in the yields ofany marker enzymes) following Triton WR-1339 treatment.These data imply that the lysosomal effects are dependentupon uptake of the detergent by endocytosis. Bloodstreamforms of T. brucei, as well as having well-characterized recep-tor-mediated uptake mechanisms for transferrin and low den-sity lipoprotein, also readily take up markers for fluid-phaseendocytosis (56, 57). Epimastigotes will take up gold-labeledproteins including low density lipoproteins. These results havebeen used as evidence that receptor-mediated endocytosis isoccurring (49), but the uptake of catalase was shown to be anon-saturable process, indicating that membrane receptorswere not involved (39). Receptor-mediated endocytosis (oftransferrin) has been demonstrated biochemically only in theamastigote stage of T. cruzi (58). We confirmed that epimas-tigotes take up gold-labeled transferrin (Fig. 7), but the extentof uptake was insufficient to modify the lysosome density, asachieved with lymphocytes (59). Similarly, we were unable toincrease the density of lysosomes by dextran loading (28, 60).We conclude, therefore, that T. cruzi epimastigotes are not veryefficient at endocytosis and that the observed effects of TritonWR-1339 may arise via a route other than endocytosis; itseffects are therefore not restricted to lysosomes/endosomes.Electron microscopy showed that vacuolation in epimastigotes
was most prominent in mitochondrial elements (shown by thepresence of the kinetoplast-DNA in vacuoles or vacuoles withdouble limiting membranes). However, the detergent’s density-altering effects were not necessarily limited to the mitochon-drion, and the separation of mitochondria from acidocalcisomeswas shown by the following: 1) the discrepancy between Percollgradient profiles of “acidic calcium” and succinate cytochrome creductase activity recovered from Triton-treated cells; 2) theassays for acidic calcium were performed in the presence of 1mg/ml antimycin A and 2 mg/ml oligomycin, which will depletemitochondria of calcium (61); and 3) mitochondria are notacidic (62), so any calcium therein should be releasable byionomycin, which was preincubated with fractions prior to as-say for acidic calcium storage.
Distinguishing Acidocalcisomes from Lysosomes—The aboveresults suggest that the acidocalcisome is an organelle distinctfrom the lysosome in T. cruzi epimastigotes. In mammaliancells Ca21 has also been reported to be present in organelleshaving an acidic milieu, such as endosomes, lysosomes, thetrans-Golgi network, and secretory granules including chro-maffin, pancreatic zymogen, and atrial-specific granules (62–67), but the functional significance of the high Ca21 content ofthese organelles is unknown (62). In addition, a Ca21-ATPasefrom rat liver lysosomes has been characterized (68), and aCa21-ATPase gene that was cloned from rat stomach (69) ex-hibits 50% amino acid identity with the Golgi-located PMR1gene of Saccharomyces cerevisiae (70). Calcium storage has alsobeen reported in lysosomes in Trypanosoma rhodesiense (71).To establish further the distinction between acidocalcisomesand lysosomes, we loaded epimastigotes with gold-labeledtransferrin to identify the endosomal/lysosomal system in theparasites (Fig. 7). Internal compartments were clearly labeled,but these were distinct from the vacuoles that corresponded tothe electron-dense organelles (Fig. 7 and Fig. 6a). The report onT. rhodesiense (71) was based on the apparent coincidence ofcalcium storage with acid phosphatase location. Electron-densevacuoles were identified as “lysosomal vesicles,” but the basisfor this designation was not established. A possible acid phos-phatase reaction was shown in one vacuole, but adequate con-trols were not presented. Additionally, the presence of acidphosphatase in an organelle does not mean it is a lysosome.Williamson and McLaren (71) themselves showed activity inthe flagellar pocket of T. rhodesiense, whereas previous subcel-lular fractionation of T. cruzi (Tulahuen strain) indicated alikely non-lysosomal location of acid phosphatase (30). Theactivity has been also detected cytochemically in the Golgi andon the cell surface of T. cruzi (39). We found a minor component(9%) of the activity associated with the cell surface in theepimastigotes that we used. As indicated above, the elementalcomposition of L. major electron-dense vacuoles (11) is quanti-tatively different from that of the T. cruzi acidocalcisomesreported here, and we cannot rule out a different biochemicalcomposition in acidocalcisomes of different stages of T. cruzi orin the different parasites in which they have been described(2–7).
In conclusion, we found evidence to suggest that the electron-dense vacuoles previously detected in T. cruzi epimastigotes(10) correspond to what has been described biochemically asacidocalcisomes (2), as indicated by their high calcium contentand their acidic character revealed by their sensitivity to ni-gericin treatment. In addition, cell fractionation studies andgold-labeled transferrin experiments indicated that these or-ganelles are different from lysosomes or other endocytic vacu-oles. The presence in trypanosomatids of a distinct organellewith apparently no counterpart in mammalian cells makes it apotential target for chemotherapy.
Analysis of Acidocalcisomes in T. cruzi28028
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

Acknowledgments—We acknowledge Carol Cunnick for technical as-sistance, Lou Ann Miller for help with the conventional electron mi-croscopy techniques, and Dr. M. Segre for use of the enzyme-linkedimmunosorbent assay reader.
REFERENCES
1. Moreno, S. N. J., Silva, J., Vercesi, A. E., and Docampo, R. (1994) J. Exp. Med.180, 1535–1540
2. Docampo, R., Scott, D. A., Vercesi, A. E., and Moreno, S. N. J. (1995) Biochem.J. 310, 1005–1012
3. Vercesi, A. E., Moreno, S. N. J., and Docampo, R. (1994) Biochem. J. 304,227–233
4. Scott, D. A., Moreno, S. N. J., and Docampo, R. (1995) Biochem. J. 310,789–794
5. Vercesi, A. E., and Docampo, R. (1996) Biochem. J. 315, 265–2706. Lu, H.-G., Zhong, L., Chang, K.-P., and Docampo, R. (1997) J. Biol. Chem. 272,
9464–94737. Moreno, S. N. J., and Zhong, L. (1996) Biochem. J. 313, 655–6598. Wada, Y., and Anraku, Y. (1994) J. Bioenerg. Biomembr. 26, 631–6379. Vickerman, K., and Tetley, L. (1977) Ann. Soc. Belge Med. Trop. 57, 441–455
10. Dvorak, J. A., Engel, J. C., Leapman, R. D., Swyt, C. R., and Pella, P. A. (1988)Mol. Biochem. Parasitol. 31, 19–26
11. LeFurgey, A., Ingram, P., and Blum, J. J. (1990) Mol. Biochem. Parasitol. 40,77–86
12. Shuman, H., Somlyo, A. V., and Somlyo, A. P. (1976) Ultramicroscopy 1,317–339
13. Somlyo, A. V., Shuman, H., and Somlyo, A. P. (1977) J. Cell Biol. 74, 828–85714. Hall, T. A., and Gupta, B. L. (1983) Q. Rev. Biophys. 16, 279–33915. Ornberg, R. L., Kuijpers, G. A. J., and Leapman, R. D. (1988) J. Biol. Chem.
263, 1488–149316. Soares, M. J., Souto-Padron, T., and de Souza, W. (1992) J. Cell Sci. 102,
157–16717. Silveira, F. T., Viana-Dias, M. G., Pereira-Pardal, P., Lobao, A., and Britto-
Melo, G. (1979) Hileia Med. 1, 61–6218. Bone, G. J., and Steinert, M. (1956) Nature 178, 308–30919. Silva, L. H. P., and Nussenzweig, V. (1953) Folia Clin. Biol. 20, 191–20720. Shi, S., Sun, S., Andrews, S. B., and Leapman, R. D. (1996) Microsc. Res. Tech.
33, 241–25021. Leapman, R. D., and Andrews, S. B. (1991) J. Microsc. (Oxf.) 161, 3–1922. Fiori, C. E., and Swyt, C. R. (1992) NIST/NIH Desktop Spectrum Analyzer,
SRD-38, Version 2.0 National Institute of Standards and Technology,Gaithersburg, MD
23. Krivanek, O. L., Ahn, C. C., and Keeney, R. B. (1987) Ultramicroscopy 22,103–116
24. Kundmann, M., Chabert, X., Truong, K., and Krivanek, O. (1990) EL/PSoftware for Macintosh II Computer, Version 3.0 Gatan Inc., Pleasanton,CA.
25. Egerton, R. F. (1996) Electron Energy-Loss Spectroscopy in the ElectronMicroscope, 2nd Ed., 245–299, Plenum Publishing Corp., New York
26. Leapman, R. D., and Ornberg, R. L. (1988) Ultramicroscopy 24, 251–26827. Toner, J. J., and Weber, M. M. (1972) Biochem. Biophys. Res. Commun. 46,
652–66028. Scott, D. A., North, M. J., and Coombs, G. H. (1995) Int. J. Parasitol. 25,
657–66629. Cazzulo, J. J., Cazzulo Franke, M. C., Martinez, J., and Franke de Cazzulo,
B. M. (1990) Biochim. Biophys. Acta 1037, 186–19130. Bontempi, E., Martinez, J., and Cazzulo, J. J. (1989) Mol. Biochem. Parasitol.
33, 43–4831. Healy, N., Greig, S., Enahoro, H., Roberts, H., Drake, L., Shaw, E., and Ashall,
F. (1992) Parasitology 104, 315–32232. Sottocasa, G. L., Kuylenstierna, B., Ernster, L., and Bergstrand, A. (1967)
J. Cell Biol. 32, 415–43833. Boveris, A., Hertig, C. M., and Turrens, J. F. (1986) Mol. Biochem. Parasitol.
19, 163–16934. Cannata, J. J. B., Valle, E., Docampo, R., and Cazzulo, J. J. (1982) Mol.
Biochem. Parasitol. 6, 151–16035. Frens, G. (1973) Nature Phys. Sci. 241, 20–2236. Horisberger, M., and Clerc, M. F. (1985) Histochemistry 82, 219–22337. Liu, C., and Hermann, T. E. (1978) J. Biol. Chem. 253, 5892–589438. Leighton, F., Poole, B., Beaufay, H., Baudhuin, P., Coffey, J. W., Fowler, S.,
and deDuve, C. (1968) J. Cell Biol. 37, 482–51339. Avila, J. L. (1992) in Subcellular Biochemistry: Intracellular Parasites (Avila,
J. L., and Harris, J. R., eds) Vol. 18, pp. 189–234, Plenum Publishing Corp.,New York
40. Schlatterer, C., Buravkov, S., Zierold, K., and Knoll, G. (1994) Cell Calcium 16,101–111
41. Dubochet, J., Adrian, M., Chang, J. J., Homo, J.-C., Lepault, J., McDowell,A. W., and Schultz, P. (1988) Q. Rev. Biophys. 21, 129–228
42. Hensgens, C. M. H., Santos, H., Zhang, C., Kruizinga, W. H., and Hansen, T. A.(1996) Eur. J. Biochem. 242, 327–331
43. Zierold, K. (1988) Methods Microbiol. 20, 91–11144. Atkinson, A. W., John, P. C. L., and Gunning, B. E. S. (1974) Protoplasma 81,
77–10945. Vallee, B. L., and Falchuk, K. H. (1993) Physiol. Rev. 73, 79–11846. Reyes, J. G. (1996) Am. J. Physiol. 270, C401–C41047. Foster, M. C., Leapman, R. D., Li, M. X., and Atwater, I. (1993) Biophys. J. 64,
525–53248. Perez-Clausell, J., and Danscher, G. (1985) Brain Res. 337, 91–9849. Soares, M. J., and de Souza, W. (1991) Parasitol. Res. 77, 461–46950. Souto-Padron, T., Campetella, O. E., Cazzulo, J. J., and De Souza, W. (1990)
J. Cell Sci. 96, 485–49051. Carvalho, T. U., and De Souza, W. (1977) J. Parasitol. 63, 1116–111752. Steinert, M. (1965) Exp. Cell Res. 39, 69–7253. de Souza, W., Souto-Padron, T., Docampo, R., and Leon, W. (1979) Acta Trop.
36, 253–25554. Xu, C. W., Hines, J. C., Engel, M. L., Russell, D. G., and Ray, D. S. (1996) Mol.
Cell. Biol. 16, 564–57655. Opperdoes, F. R., and van Roy, J. (1982) Mol. Biochem. Parasitol. 6, 181–19056. Webster, P., and Fish, W. R. (1989) Eur. J. Cell Biol. 49, 303–31057. Hager, K. M., Pierce, M. A., Moore, D. R., Tytler, E. M., Esko, J. D., and
Hajduk, S. L. (1994) J. Cell Biol. 126, 155–16758. Lima, M. F., and Villalta, F. (1990) Mol. Biochem. Parasitol. 38, 245–25259. Beaumelle, B. D., Gibson, A., and Hopkins, C. R. (1990) J. Cell Biol. 111,
1811–182360. Arai, K., Kanaseki, T., and Ohkuma, S. (1991) J. Biochem. (Tokyo) 110,
541–54761. Docampo, R., and Vercesi, A. E. (1989) J. Biol. Chem. 264, 108–11162. Pozzan, T., Rizzuto, R., Volpe, P., and Meldolesi, J. (1994) Physiol. Rev. 74,
595–63663. Virk, S. S., Kirk, C. J., and Shears, S. B. (1985) Biochem. J. 226, 741–74864. Somlyo, A. V., Broderick, R., Shuman, H., Buhle, E. L., and Somlyo, A. P.
(1988) Proc. Natl. Acad. Sci. U. S. A. 85, 6222–622665. Gerasimenko, O. V., Gerasimenko, J. V., Belan, P. V., and Petersen, O. H.
(1996) Cell 84, 473–48066. Haller, T., Dietl, P., Deetjen, P., and Volkl, H. (1996) Cell Calcium 19, 157–16567. Bode, H.-P., Himmen, A., and Goke, B. (1996) Pfluegers Arch. Eur. J. Physiol.
432, 97–10468. Adachi, T., Arai, K., and Ohkuma, S. (1996) Biol. Pharm. Bull. 19, 1291–129769. Gunteski-Hamblin, A. M., Clarke, D. M., and Shull, G. E. (1992) Biochemistry
31, 7600–760870. Antebi, Y., and Fink, G. R. (1992) Mol. Biol. Cell 3, 633–65471. Williamson, J., and McLaren, D. J. (1981) J. Protozool. 28, 460–467
Analysis of Acidocalcisomes in T. cruzi 28029
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

LeapmanDavid A. Scott, Roberto Docampo, James A. Dvorak, Shanling Shi and Richard D.
Trypanosoma cruzi Compositional Analysis of Acidocalcisomes in In Situ
doi: 10.1074/jbc.272.44.280201997, 272:28020-28029.J. Biol. Chem.
http://www.jbc.org/content/272/44/28020Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/272/44/28020.full.html#ref-list-1
This article cites 68 references, 21 of which can be accessed free at
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from





























