Embed Size (px)
Citation preview

Heavy Metals and Plants - a complicated relationship Heavy metal stress and heavy metal resistance
l h l i i h ildHeavy metal-hyperaccumulation in the Wild Westmodified from:
Presented by Hendrik Küpper for the VTK Plant Physiology 2011
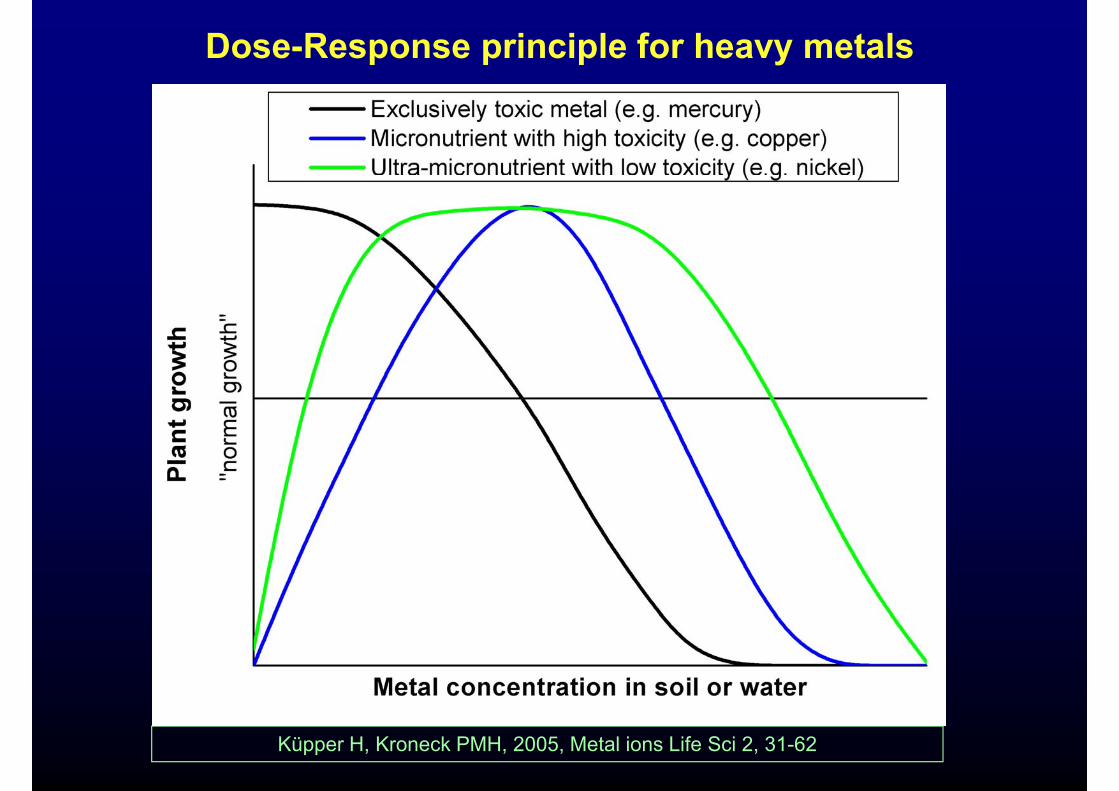
Dose-Response principle for heavy metals
Küpper H, Kroneck PMH, 2005, Metal ions Life Sci 2, 31-62

Environmental relevance of heavy metal toxicity
A seemingly intact, natural creek ...However, the Elodea canadensiso e e , t e odea ca ade s sinside died from zinc stress that converted its chlorophyll to Zn-chlorophyllp y
Zn-Fluosilicate
Küpper H, Küpper F, Spiller M (1996) Journal of Experimental Botany 47 (295), 259-66

Environmental relevance of heavy metal toxicity
Where? How? Why?
• Naturally on heavy metal rich soils(Cu: e.g. in Zaire, Afrika; Zn/Cd: rel. frequent, incl. Europe; Ni: rel frequent serpentine soils e gEurope; Ni: rel. frequent, serpentine soils e.g. in Africa, Australia, North and Middle Amerika): Heavy metal concentrations high enough for being toxic for most organisms.
• Naturally in copper-rich areas of the oceans (e.g. Sargasso sea): Cu-concentrations in the nanomolar range already inhibit some sensitive cyanobacteria.
• Anthropogenically due to the use of heavy metal salts (e.g. CuSO4, z.B. Zn-phosphid, Zn-borate, Zn-fluosilicate): concentrations in themicromolar range are toxic for many plants, mainly water plants in neighbouring ponds g y p , y p g g pand creeks
• Anthropogenically due to ore mining and refining, concentrations in the vicinity of mines, smelters and rubble dumps can be extremely high and toxic for all organisms.
• Anthropogenically due to the activities of other inductries. The longest river in Germany the Rhine contained up to 0 5 µM copper in the 1970‘s which is lethal forGermany, the Rhine, contained up to 0.5 µM copper in the 1970‘s, which is lethal for sensitive water plants like Stratiotes or Elodea.

Heavy-metal induced damage
--> Inhibition of root function ( KK 5th semester)
--> Genotoxicity
--> Direct inhibition of photosynthesis
--> Oxidative stress: direct and as a result of a malfunction of photosynthesis
--> Substitution of active centres in enzymes
--> Inhibition of respiration and other relatively insensitive processes
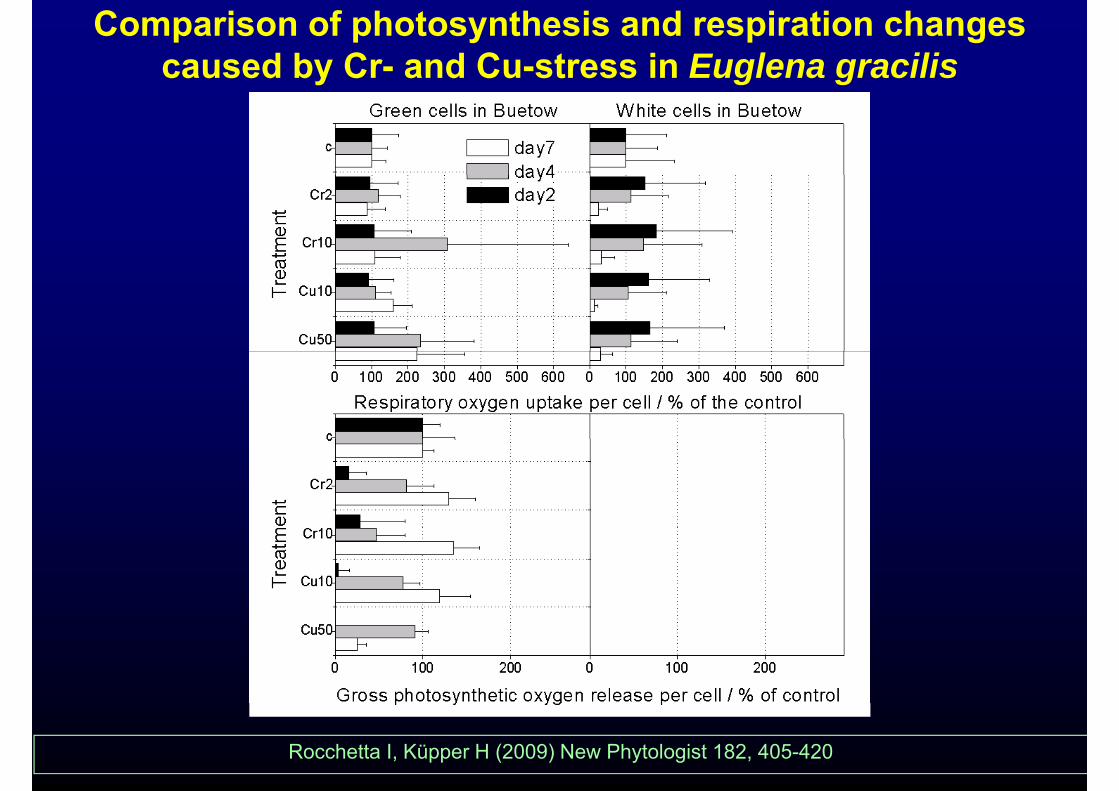
Comparison of photosynthesis and respiration changescaused by Cr- and Cu-stress in Euglena gracilis
Photosynthesis is much more sensitive, ,respiration changes later as a secondary
effect
Rocchetta I, Küpper H (2009) New Phytologist 182, 405-420
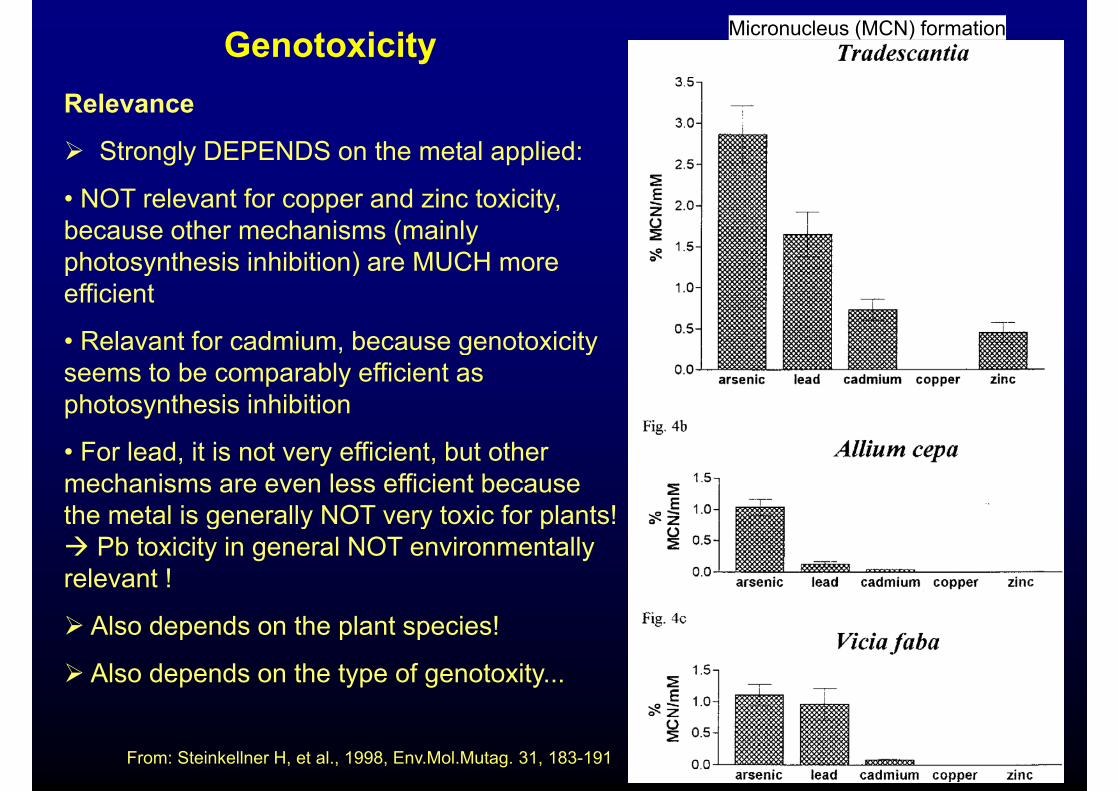
Genotoxicity Micronucleus (MCN) formation
Relevance
Strongly DEPENDS on the metal applied:
• NOT relevant for copper and zinc toxicity, because other mechanisms (mainly photosynthesis inhibition) are MUCH morephotosynthesis inhibition) are MUCH more efficient
• Relavant for cadmium, because genotoxicity , g yseems to be comparably efficient as photosynthesis inhibition
• For lead, it is not very efficient, but other mechanisms are even less efficient because the metal is generally NOT very toxic for plants! g y y p Pb toxicity in general NOT environmentally relevant !
Also depends on the plant species!
Also depends on the type of genotoxity...
From: Steinkellner H, et al., 1998, Env.Mol.Mutag. 31, 183-191
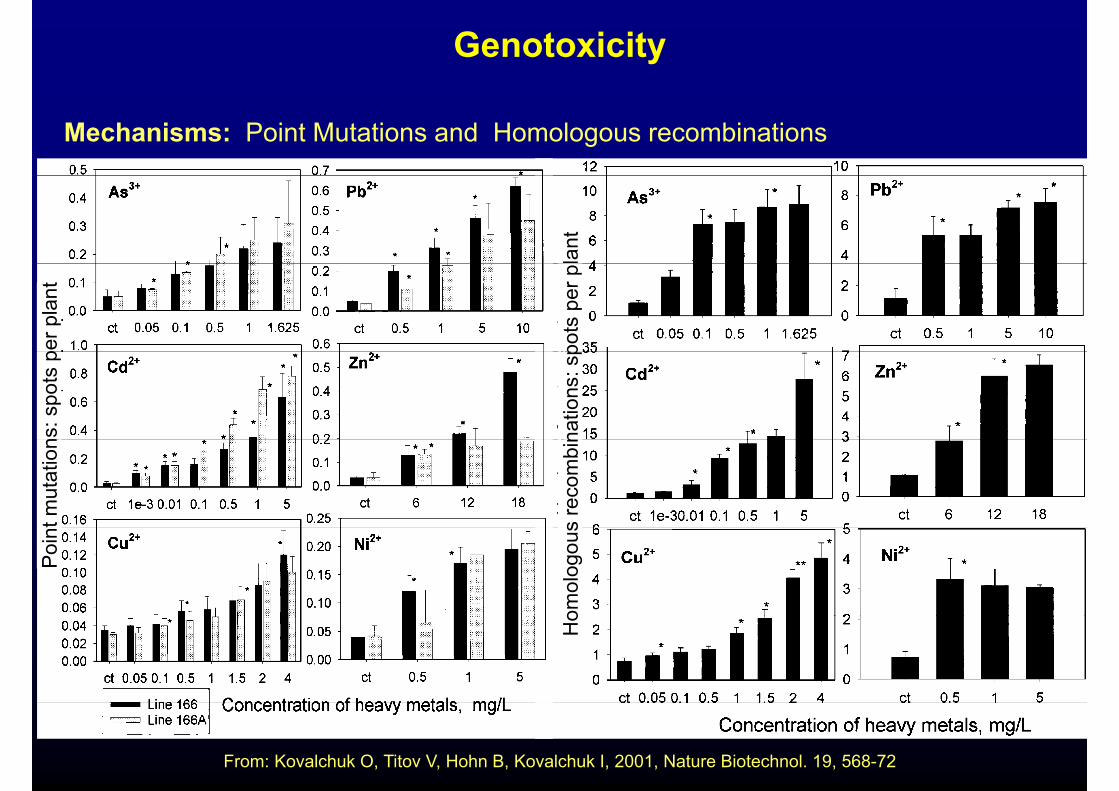
Genotoxicity
Mechanisms: Point Mutations and Homologous recombinations
lant
pots
per
p
per p
lant
natio
ns: s
p
ns: s
pots
p
s re
com
bin
t mut
atio
n
omol
ogou
s
Poi
nt
Ho
From: Kovalchuk O, Titov V, Hohn B, Kovalchuk I, 2001, Nature Biotechnol. 19, 568-72
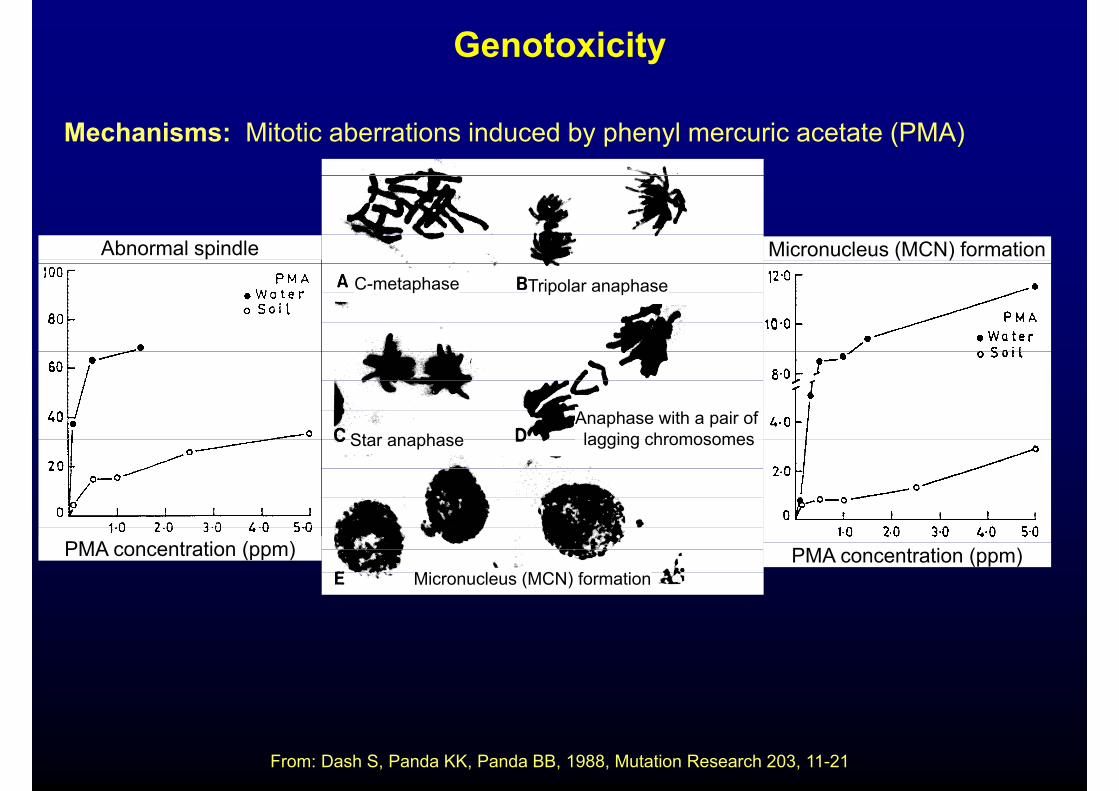
Genotoxicity
Mechanisms: Mitotic aberrations induced by phenyl mercuric acetate (PMA)
Micronucleus (MCN) formationAbnormal spindle
C-metaphase Tripolar anaphase
Star anaphaseAnaphase with a pair of lagging chromosomesStar anaphase lagging chromosomes
Micronucleus (MCN) formationPMA concentration (ppm) PMA concentration (ppm)
From: Dash S, Panda KK, Panda BB, 1988, Mutation Research 203, 11-21

Oxidative StressR lRelevance
• NOT clear: Studies with environmentally relevant realistic but still toxic heavy metal concentrations oft do NOT show oxidative stress! Almost all studies concluding thatconcentrations oft do NOT show oxidative stress! Almost all studies concluding that oxidative stress would be a major factor in heavy metal induced inhibition of plant metabolism were carried out using extremely high metal concentrations.
Mechanisms generating reactive oxygen species during heavy metal stress
• Direct: catalysed by redox-active metal ions (Fe2+, Cu+), hydrogen peroxide is converted to reactive oxygen radicals via the Fenton-Reaction:
• Indirect: malfunktion of photosynthesis and respiration can generate reactive oxygen species Therefore even in vivo redox inert Metal ions like Zn2+ and Cd2+ can causespecies. Therefore, even in vivo redox-inert Metal ions like Zn2+ and Cd2+ can cause oxidative stress.

Oxidative Stress
Mechanisms of damage caused by oxidative stress in plants
•Oxidative stress can lead to oxidation of Lipids in•Oxidative stress can lead to oxidation of Lipids in membranes and thus make them leaky. This is a popular but debated mechanism.
• Oxidation of proteins
From: en.wikipedia.org
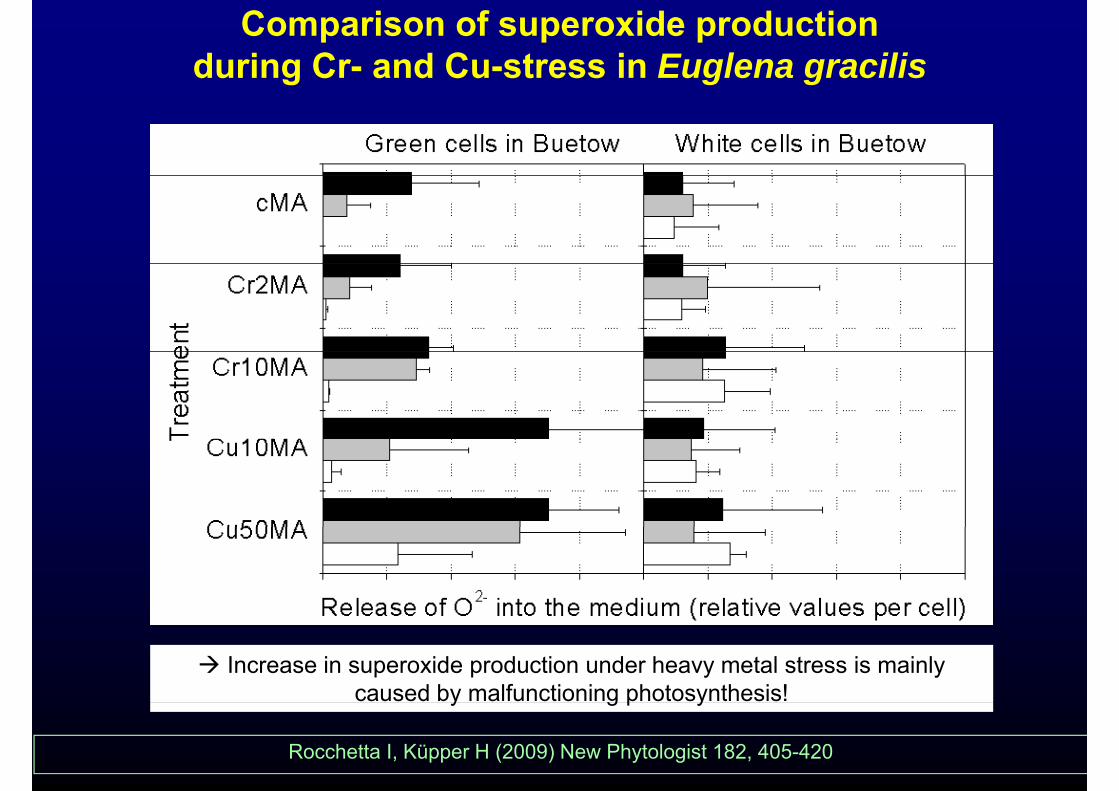
Comparison of superoxide production during Cr- and Cu-stress in Euglena gracilis
Photosynthesis is much more sensitive, ,respiration changes later as a secondary
effect
Increase in superoxide production under heavy metal stress is mainly caused by malfunctioning photosynthesis!
Rocchetta I, Küpper H (2009) New Phytologist 182, 405-420
y g p y
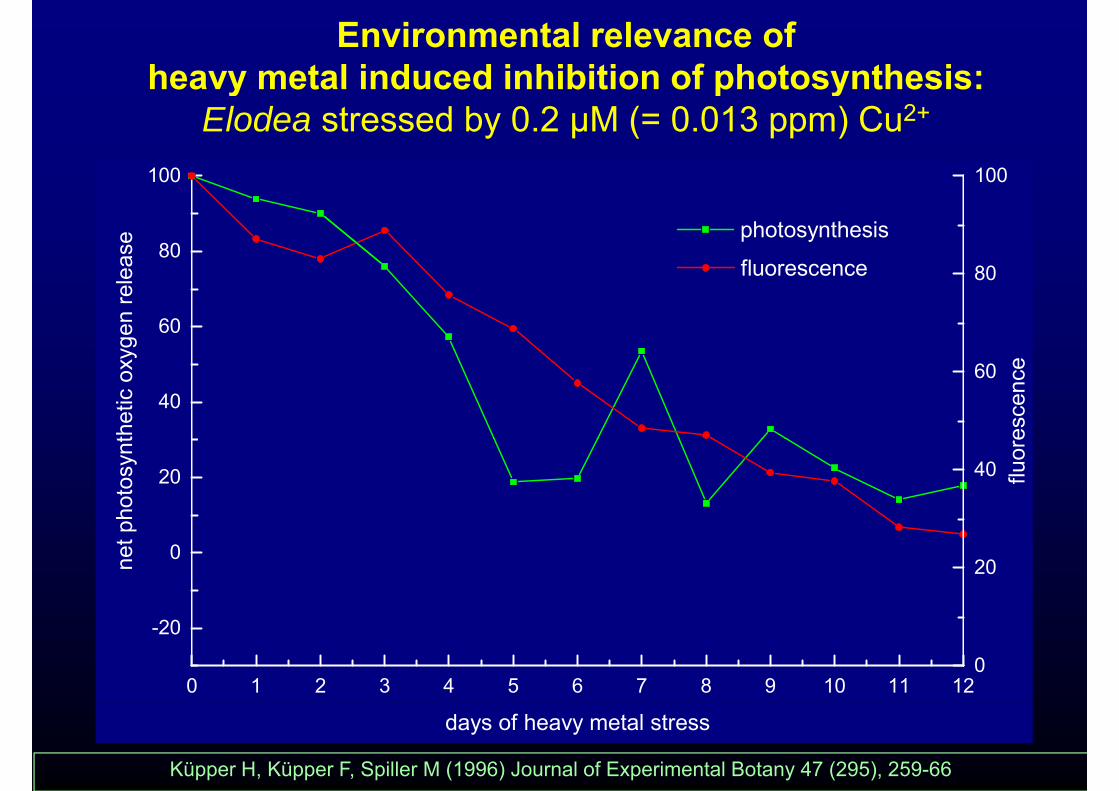
Environmental relevance of heavy metal induced inhibition of photosynthesis:
100 100
heavy metal induced inhibition of photosynthesis: Elodea stressed by 0.2 µM (= 0.013 ppm) Cu2+
80
100
photosynthesis
ase
100
fl
60
ygen
rele
a 80fluorescence
40
thet
ic o
xy 60
esce
nce
20
phot
osyn
t
40 fluor
e
0net p
20
0 1 2 3 4 5 6 7 8 9 10 11 12
-20
0
days of heavy metal stress
Küpper H, Küpper F, Spiller M (1996) Journal of Experimental Botany 47 (295), 259-66
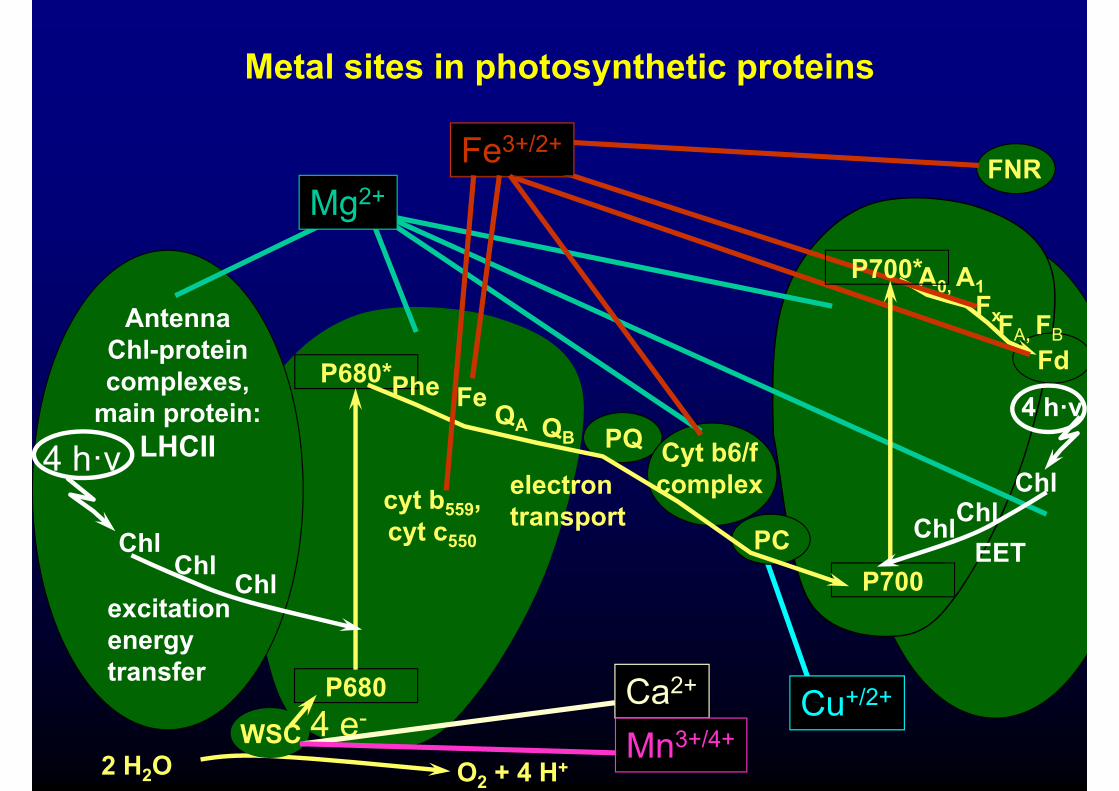
Metal sites in photosynthetic proteins
FNRFe3+/2+FNR
P700*
Mg2+
A0, A1
Antenna Chl-protein Fd
FxFA, FB
P700*
P680*QA
4 h·ν
Chl-protein complexes,
main protein:LHCII
FdPhe
PQFe
QB
ChlChlChl
LHCII4 h·νcyt b559,cyt c
PQ
electron transport
B Cyt b6/fcomplex
EETChl
excitationChl
ChlChl cyt c550 PC
p
P700
P680
excitation energy transfer Ca2+ Cu+/2+
2 H2O O2 + 4 H+WSC 4 e-
Ca Cu /2
Mn3+/4+
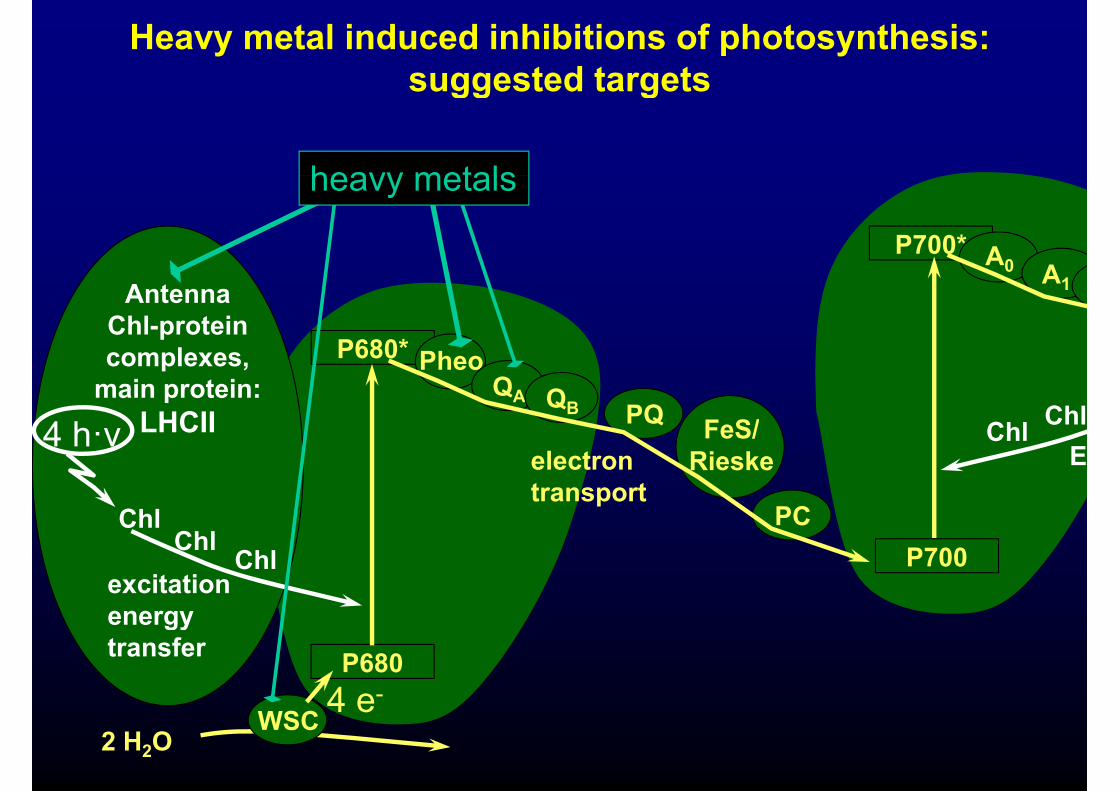
Heavy metal induced inhibitions of photosynthesis:suggested targetssuggested targets
heavy metals
P700* A0
heavy metals
A0 A1
P680* Ph
Antenna Chl-protein
l P680 PheoQA QB ChlChlPQ FeS/
complexes,main protein:
LHCII4 h·νE
ChlRieske
PC
electron transport
4 h ν
ChlP700
excitation energy
ChlChl
Chl
P6804 e-
energy transfer
2 H2OWSC 4 e
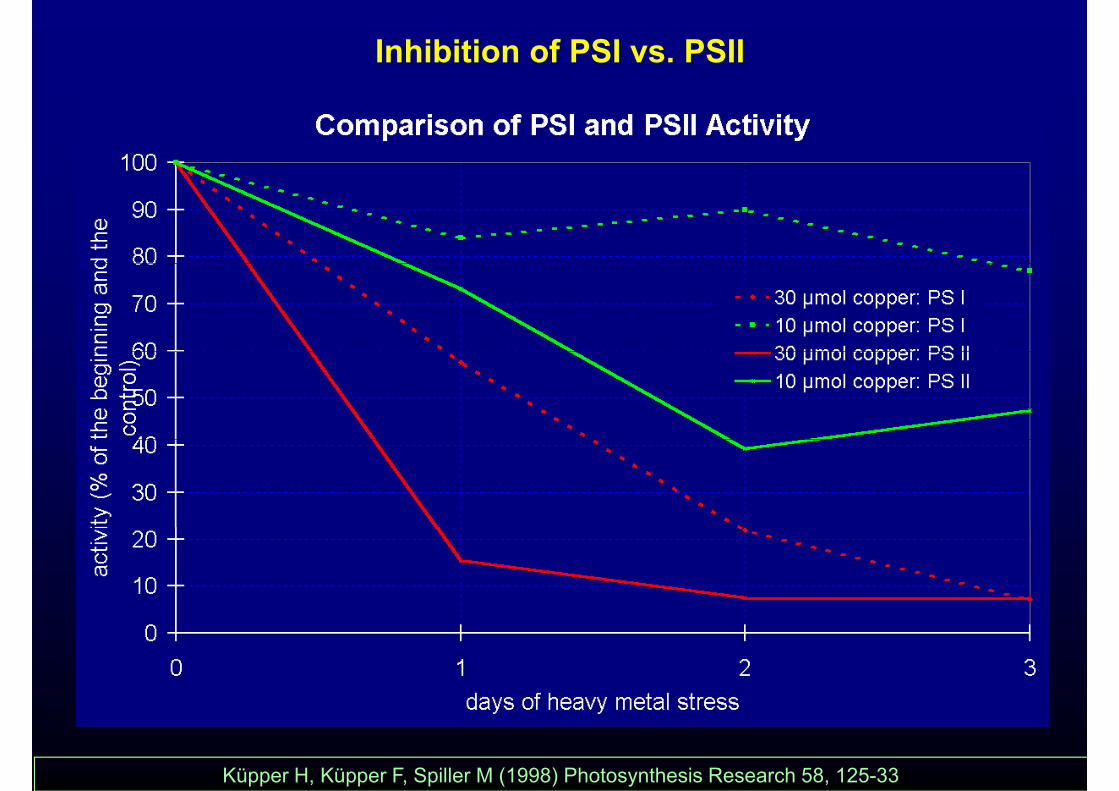
Inhibition of PSI vs. PSII
Küpper H, Küpper F, Spiller M (1998) Photosynthesis Research 58, 125-33
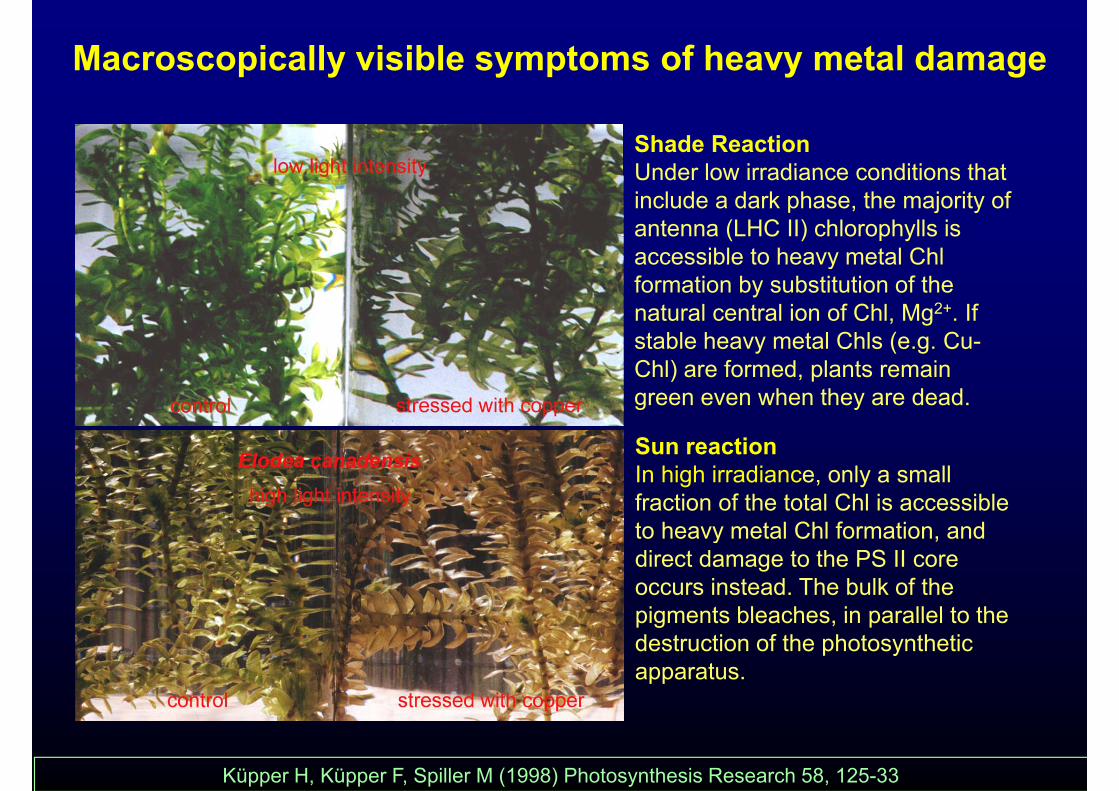
Macroscopically visible symptoms of heavy metal damage
Shade ReactionUnder low irradiance conditions thatUnder low irradiance conditions that include a dark phase, the majority of antenna (LHC II) chlorophylls is accessible to heavy metal Chlaccessible to heavy metal Chl formation by substitution of the natural central ion of Chl, Mg2+. If stable heavy metal Chls (e.g. Cu-
S ti
y ( gChl) are formed, plants remain green even when they are dead.
Sun reactionIn high irradiance, only a small fraction of the total Chl is accessible to heavy metal Chl formation andto heavy metal Chl formation, and direct damage to the PS II core occurs instead. The bulk of the pigments bleaches in parallel to thepigments bleaches, in parallel to the destruction of the photosynthetic apparatus.
Küpper H, Küpper F, Spiller M (1998) Photosynthesis Research 58, 125-33

Why are heavy metal chlorophylls unsuitable for photosynthesis?photosynthesis?
• shift of absorbance/fluorescence bands --> less energy transfer
• unstable singlet excited state --> “black holes“ for excitons
• different structure --> proteins denature
• do not readily perform charge separation when in reaction centre.

Static fluorescence microscopy of metal-stressed Elodeatransmittant light observation
1 µM Cu2+ control 100 µM Zn2+
red (650-700nm) chlorophyll fluorescence( ) p y
Küpper H, Küpper F, Spiller M (1998) Photosynthesis Research 58, 125-33
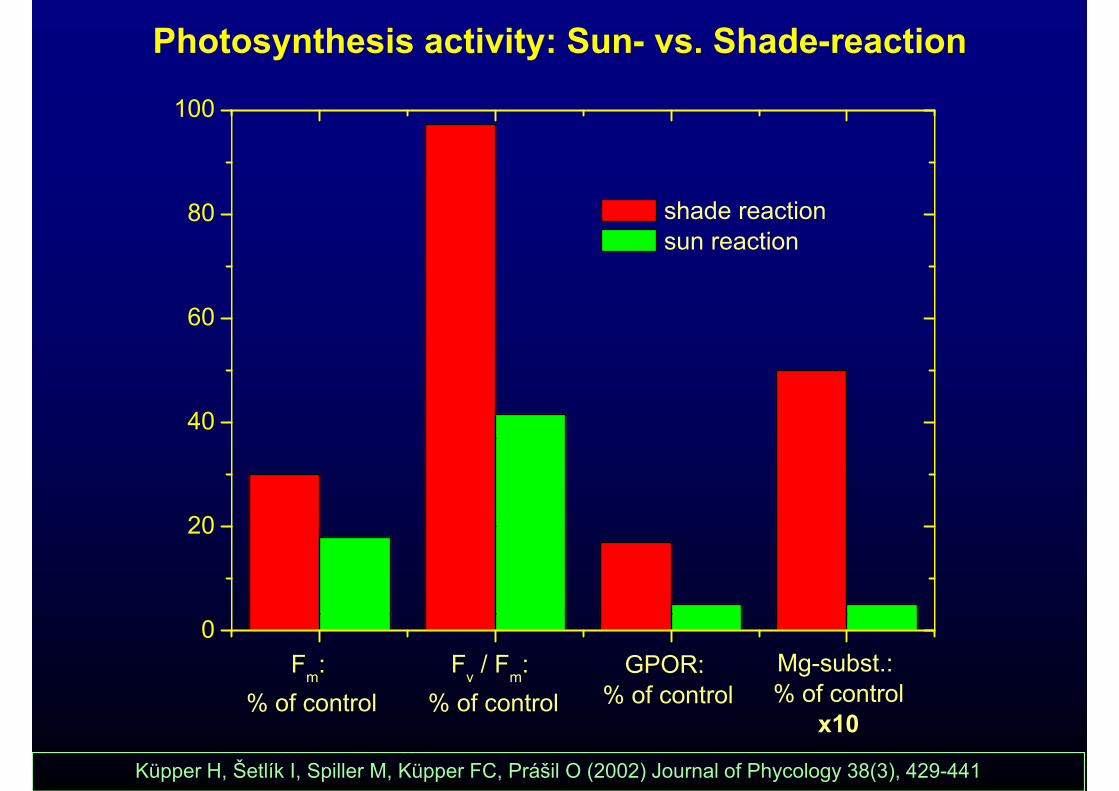
Photosynthesis activity: Sun- vs. Shade-reaction
100
80 shade reaction sun reaction
60
40
2020
0
Mg-subst.: % of control
Fv / Fm: % of control
GPOR: % of control
Fm: % of control
x10% of control % of control% of control
Küpper H, Šetlík I, Spiller M, Küpper FC, Prášil O (2002) Journal of Phycology 38(3), 429-441
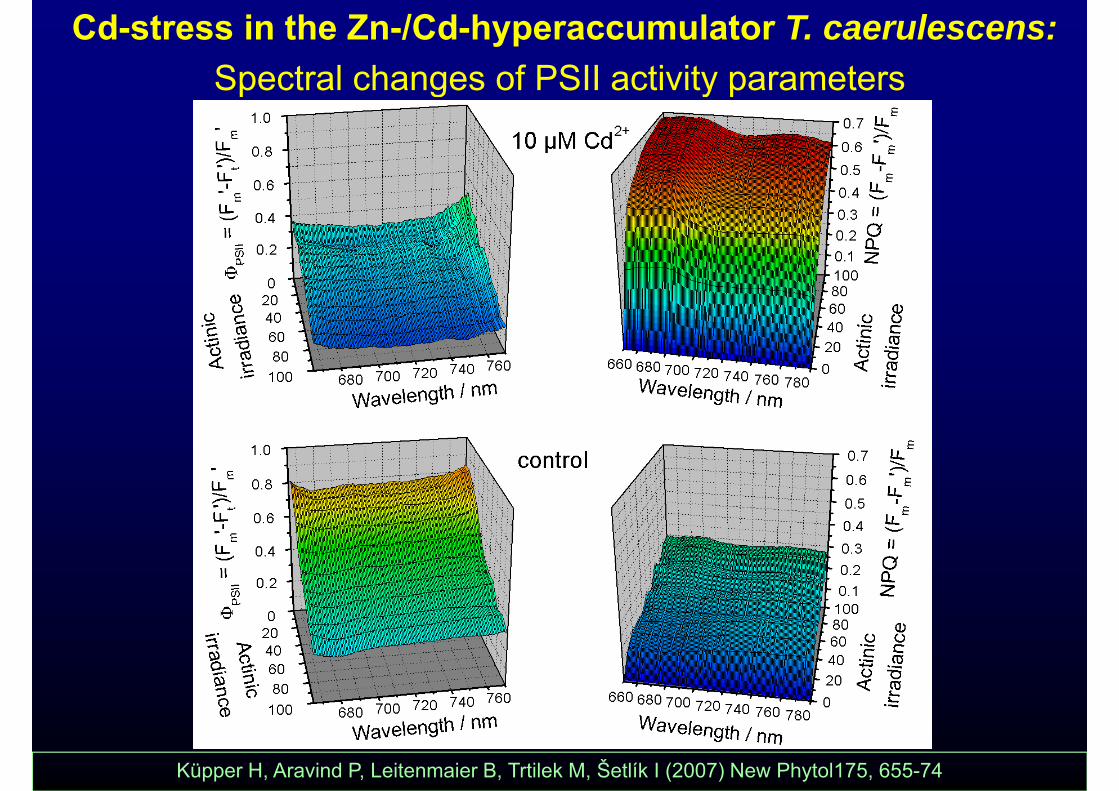
Cd-stress in the Zn-/Cd-hyperaccumulator T. caerulescens: Spectral changes of PSII activity parametersSpectral changes of PSII activity parameters
Küpper H, Aravind P, Leitenmaier B, Trtilek M, Šetlík I (2007) New Phytol175, 655-74
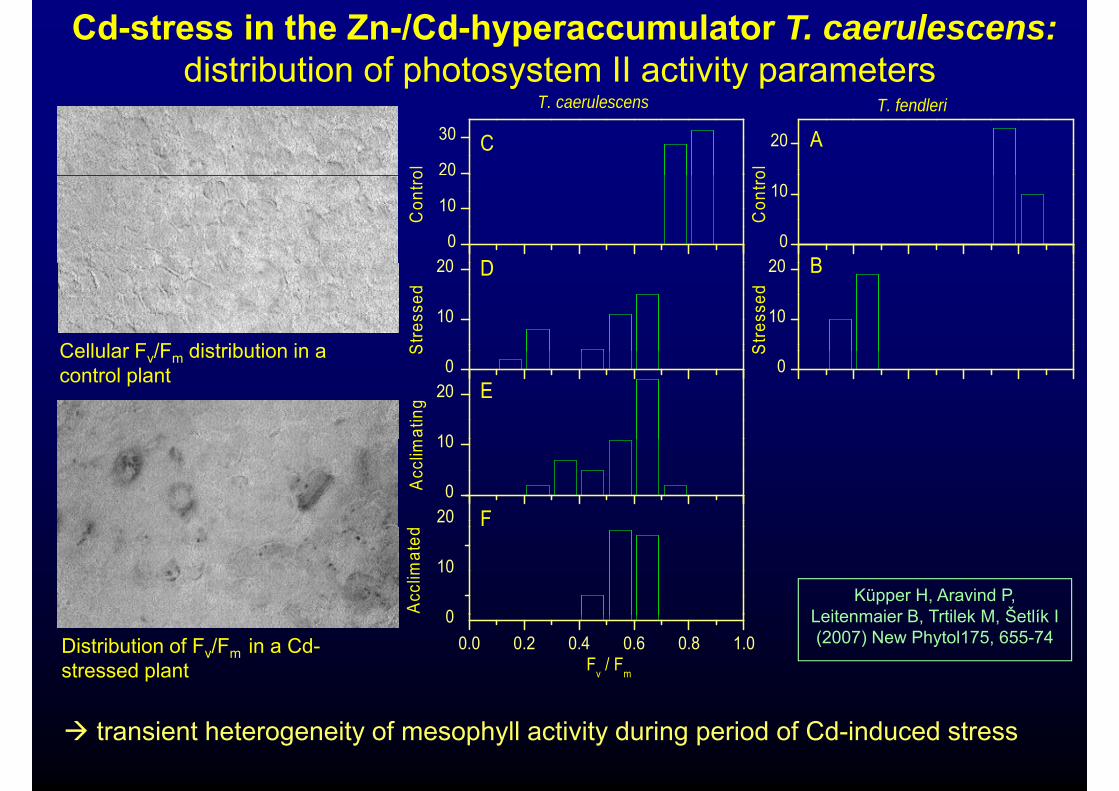
Cd-stress in the Zn-/Cd-hyperaccumulator T. caerulescens: distribution of photosystem II activity parametersp y y p
20
30
T. caerulescens
C
ol
20
T. fendleri
ol
A
0
10
0
Con
tro
20 D0
10
Con
tro
20 B
Cellular F /F distribution in a
10
20 D
Stre
ssed
10
20 B
Stre
ssed
Cellular Fv/Fm distribution in a control plant 0
S10
20 Eat
ing
0
S
0
10Ac
clim
20 F
0
10
Accl
imat
ed
Küpper H, Aravind P, Leitenmaier B Trtilek M Šetlík I
Distribution of Fv/Fm in a Cd-stressed plant
0.0 0.2 0.4 0.6 0.8 1.00
Fv / Fm
Leitenmaier B, Trtilek M, Šetlík I (2007) New Phytol175, 655-74
transient heterogeneity of mesophyll activity during period of Cd-induced stress
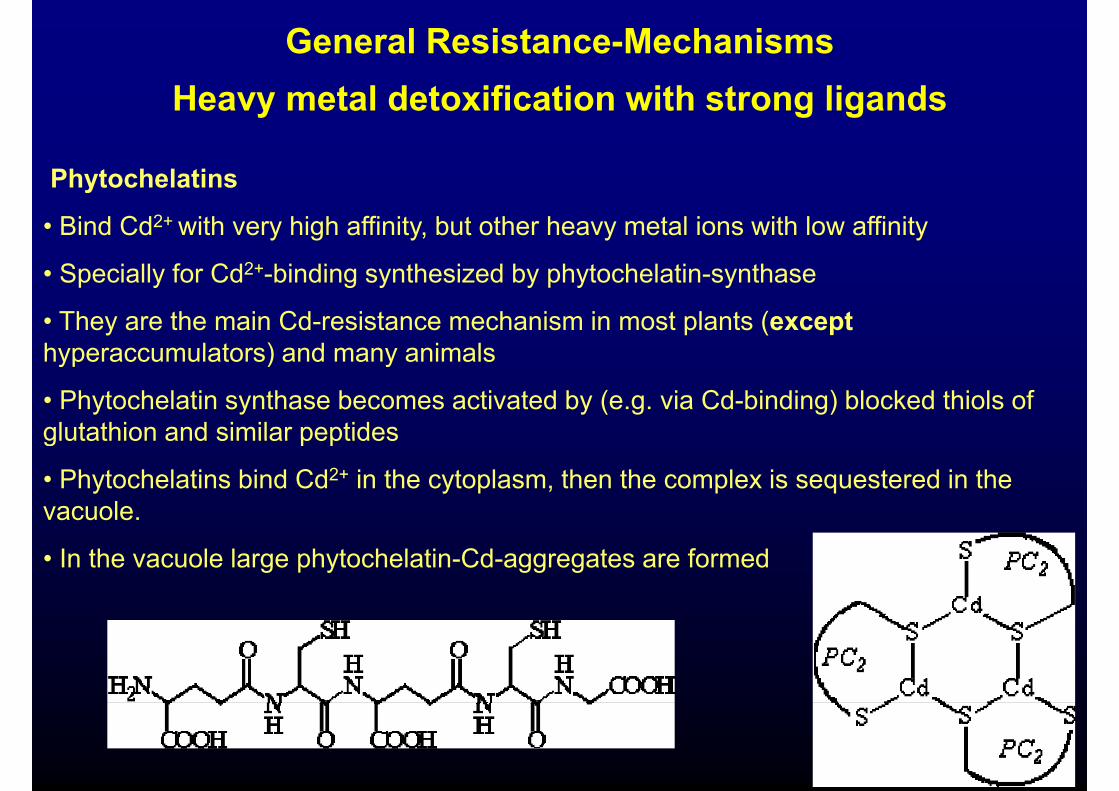
General Resistance-MechanismsH t l d t ifi ti ith t li dHeavy metal detoxification with strong ligands
PhytochelatinsPhytochelatins
• Bind Cd2+ with very high affinity, but other heavy metal ions with low affinity
S f C 2• Specially for Cd2+-binding synthesized by phytochelatin-synthase
• They are the main Cd-resistance mechanism in most plants (excepthyperaccumulators) and many animalshyperaccumulators) and many animals
• Phytochelatin synthase becomes activated by (e.g. via Cd-binding) blocked thiols of glutathion and similar peptidesglutathion and similar peptides
• Phytochelatins bind Cd2+ in the cytoplasm, then the complex is sequestered in the vacuole.
• In the vacuole large phytochelatin-Cd-aggregates are formed

General Resistance-MechanismsH t l d t ifi ti ith t li dHeavy metal detoxification with strong ligands
Glutathion
Also gl tathione itself the b ilding block of ph tochelatins can bind and th s deto if• Also glutathione itself, the building block of phytochelatins, can bind and thus detoxify heavy metals - the in vivo relevance is questionable
Metallothionins
• MTs of type I und II bind Cu+ with high affinity and seem to be involved in its detoxification.
BUT Main role of MTs in plants seems to be• BUT: Main role of MTs in plants seems to be Metal-distribution during the normal (non-stressed) metabolism

General Resistance-MechanismsH t l d t ifi ti ith t li d
Other Ligands
N t i i id i ti i ( l i l d i l t t)
Heavy metal detoxification with strong ligands
• Non-proteogenic amino acid nicotianamine (also involved in normal transport)
• Anthocyanins: seem to be involved in Brassicacae in Molybdemum binding (detoxification or storage?)(detoxification or storage?)
H l t l 2001Hale et al_2001, PlantPhysiol
126, 1391-1402
Cell wall• Cell wall
Carr HP, Lombi E, Küpper H, McGrath SP, Wong MH (2003)
Agronomie 23, 705-10
• Some algae release unidentified thiol-ligands during Cu-stress

Heavy metal detoxification by compartmentationMechanismsMechanisms
• Generelly: aktive transport processes against the concentration gradient transport proteins involved.p p
• Exclusion from cells:- observed in brown algae- in roots
• Sequestration in the vacuole: Kü H t l 2001 J E B t 52 (365) 2291 2300• Sequestration in the vacuole: - plant-specific mechanism (animals+bacteria usually don‘t have vacuoles...)- very efficient, because the vacuole does not contain sensitive enzymes
Küpper H et al., 2001, J Exp Bot 52 (365), 2291-2300
- saves the investment into the synthesis of strong ligands like phytochelatins - main mechanism in hyperaccumulators
• Sequestration in least sensitive tissues e g the epidermis instead of the• Sequestration in least sensitive tissues, e.g. the epidermis instead of the photosynthetically active mesophyll
Küpper H, Zhao F, McGrath SP (1999) Plant Physiol 119, 305-11
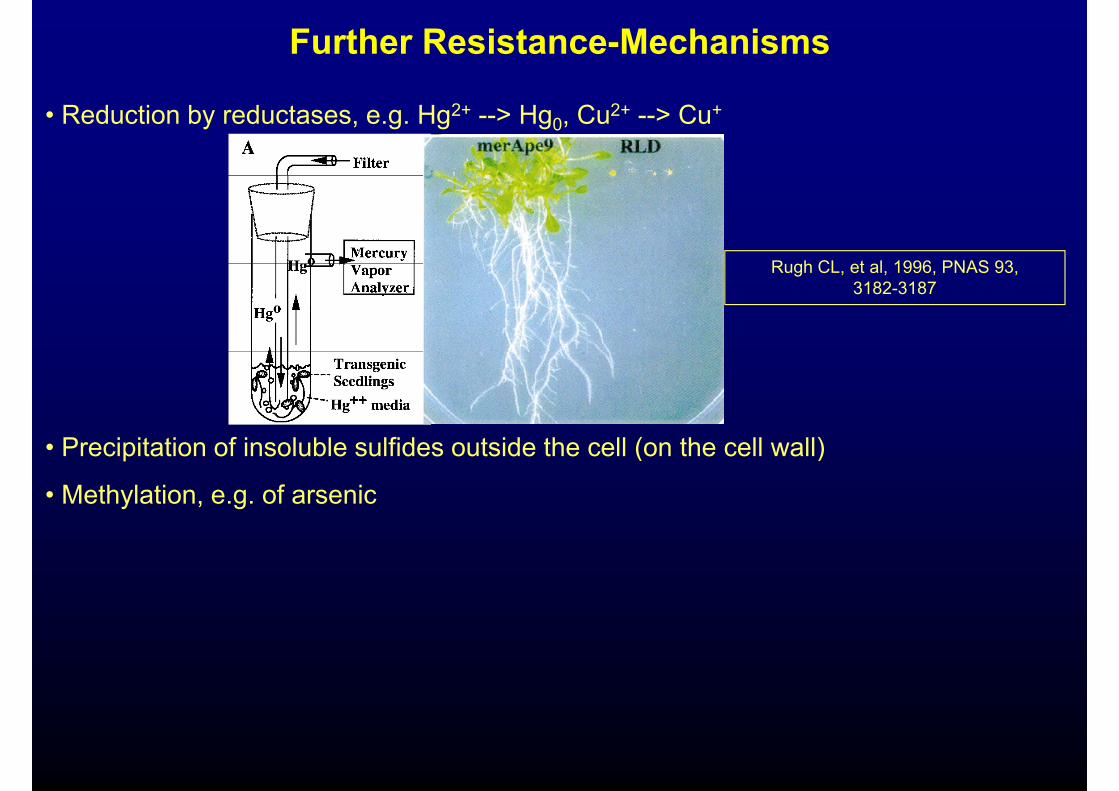
Further Resistance-Mechanisms
• Reduction by reductases, e.g. Hg2+ --> Hg0, Cu2+ --> Cu+
R h CL t l 1996 PNAS 93Rugh CL, et al, 1996, PNAS 93, 3182-3187
P i it ti f i l bl lfid t id th ll ( th ll ll)• Precipitation of insoluble sulfides outside the cell (on the cell wall)
• Methylation, e.g. of arsenic
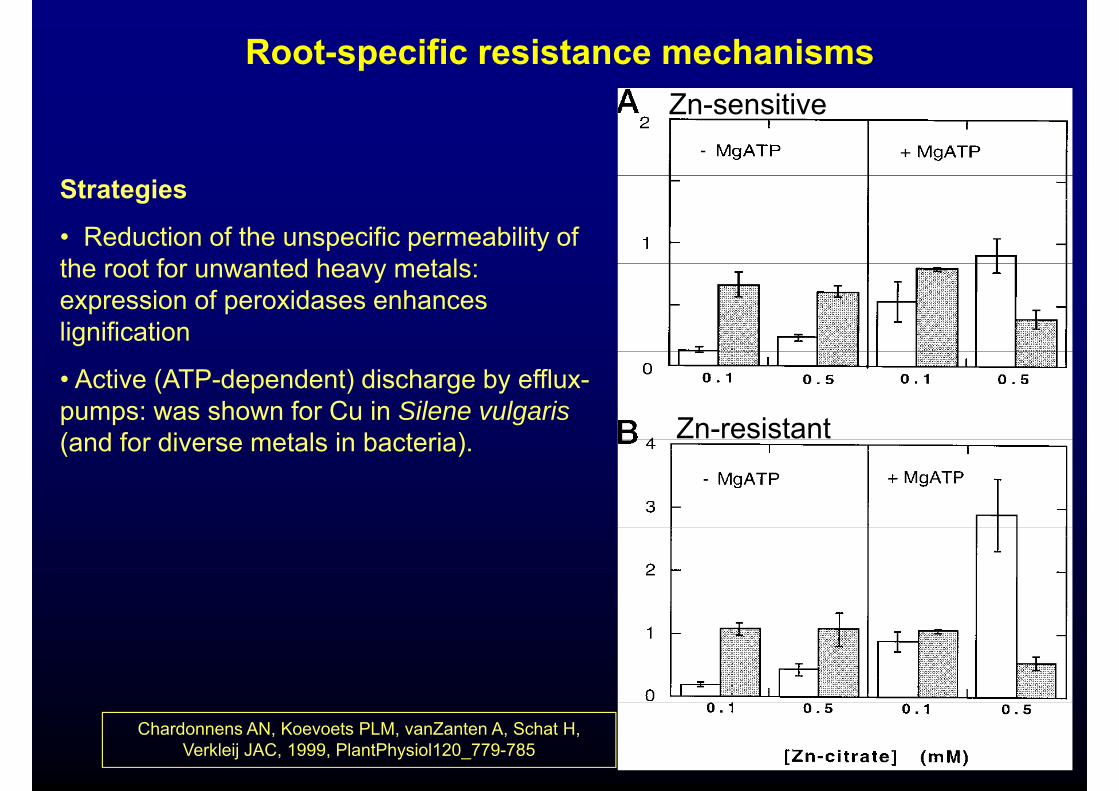
Root-specific resistance mechanismsZn-sensitive
Strategies
• Reduction of the unspecific permeability of the root for n anted hea metalsthe root for unwanted heavy metals: expression of peroxidases enhances lignification
• Active (ATP-dependent) discharge by efflux-pumps: was shown for Cu in Silene vulgaris(and for diverse metals in bacteria) Zn-resistant(and for diverse metals in bacteria).
Chardonnens AN, Koevoets PLM, vanZanten A, Schat H, Verkleij JAC, 1999, PlantPhysiol120_779-785

Resistance mechanisms against oxidative stress
• Enhanced expression of enzymes that detoxify reactive oxygen species (superoxideEnhanced expression of enzymes that detoxify reactive oxygen species (superoxide dismutase+catalase. Problem: inhibition of Zn-uptake (SOD) during Cd-Stress.
• Synthesis of non-enzyme-antioxidants, e.g. ascorbate and glutathione
Ch i th ll b t k th i t t i t th tt k f• Changes in the cell membranes to make them more resistant against the attack of reactive oxygen species: - Lipids with less unsaturated bonds- Exchange of phosphatidyl-choline against phosphytidyl-ethanolamine as lipid-“head“- Diminished proportion of lipids and enhanced proportion of stabilising proteins in the membrane
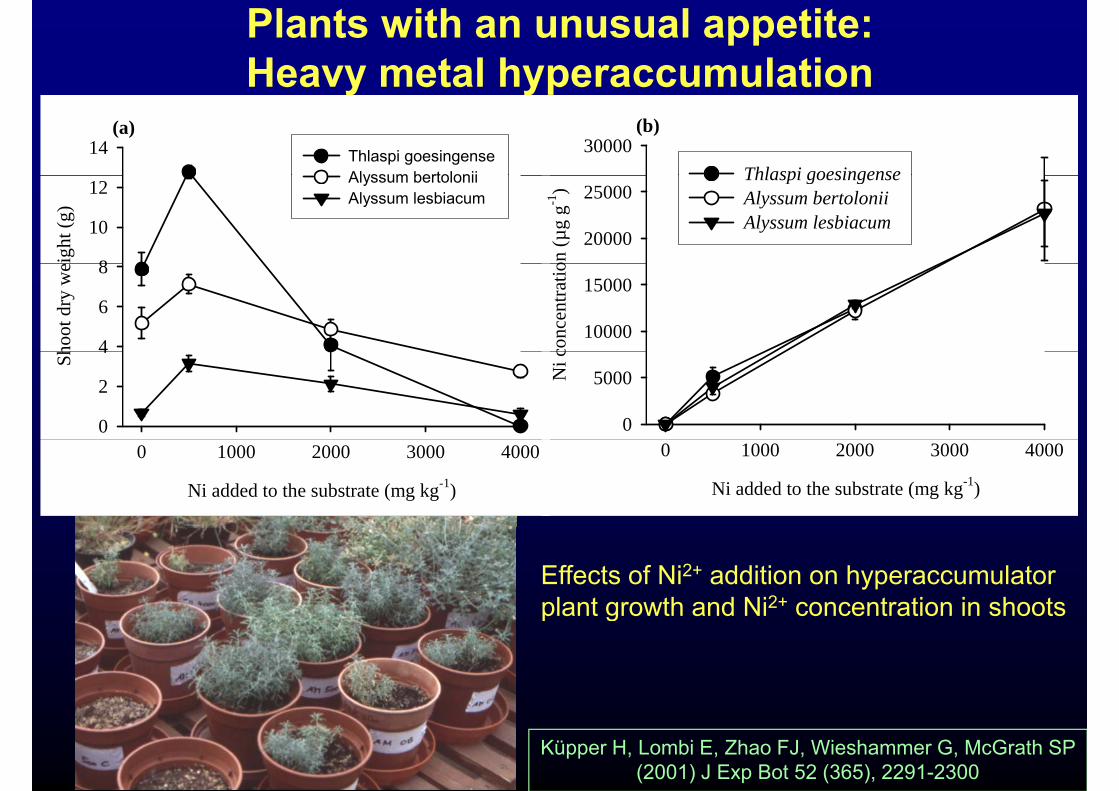
Plants with an unusual appetite:Heavy metal hyperaccumulationy yp
(a)14 Thlaspi goesingense
Alyssum bertolonii
(b)30000
Thlaspi goesingense
ight
(g)
8
10
12 Alyssum bertoloniiAlyssum lesbiacum
on (µ
g g-1
)
20000
25000Thlaspi goesingenseAlyssum bertoloniiAlyssum lesbiacum
hoot
dry
we
4
6
8
conc
entra
tio
10000
15000
Sh
0
2
4
Ni c
0
5000
Ni added to the substrate (mg kg-1)
0 1000 2000 3000 4000
Ni added to the substrate (mg kg-1)
0 1000 2000 3000 4000
Effects of Ni2+ addition on hyperaccumulator plant growth and Ni2+ concentration in shootsplant growth and Ni concentration in shoots
Küpper H, Lombi E, Zhao FJ, Wieshammer G, McGrath SP (2001) J Exp Bot 52 (365), 2291-2300
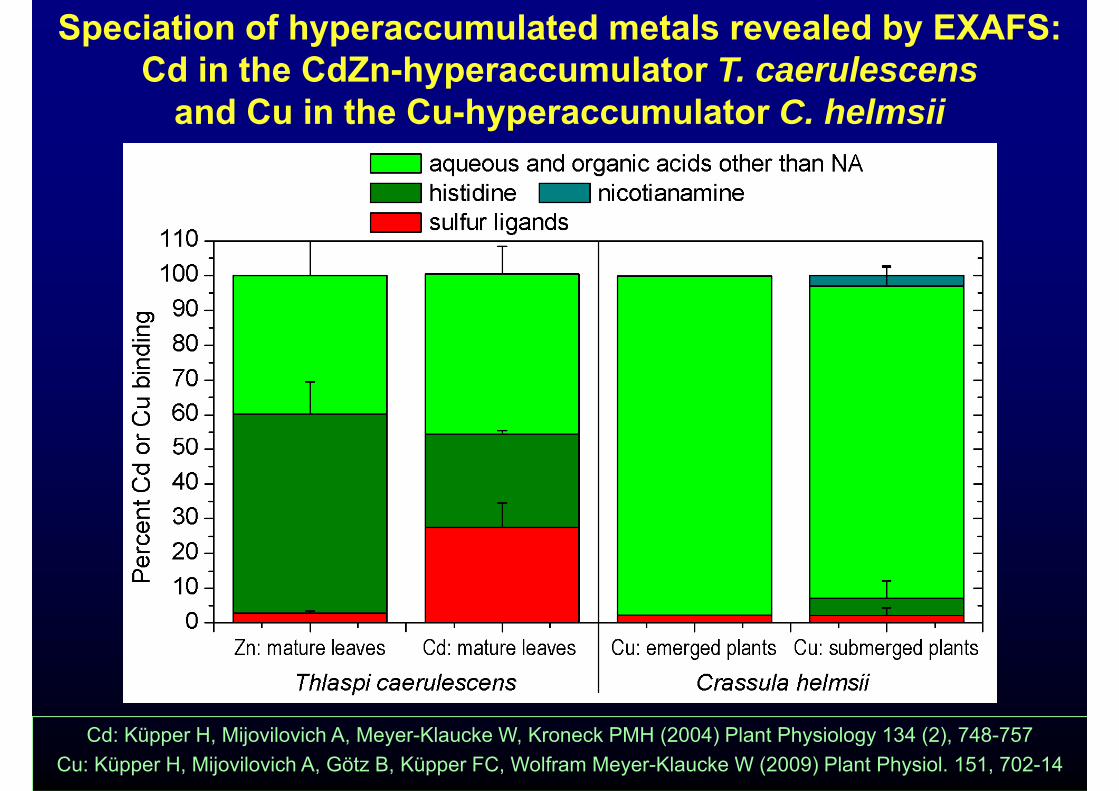
Speciation of hyperaccumulated metals revealed by EXAFS:Cd in the CdZn-hyperaccumulator T. caerulescensyp
and Cu in the Cu-hyperaccumulator C. helmsii
Cd: Küpper H, Mijovilovich A, Meyer-Klaucke W, Kroneck PMH (2004) Plant Physiology 134 (2), 748-757Cu: Küpper H, Mijovilovich A, Götz B, Küpper FC, Wolfram Meyer-Klaucke W (2009) Plant Physiol. 151, 702-14
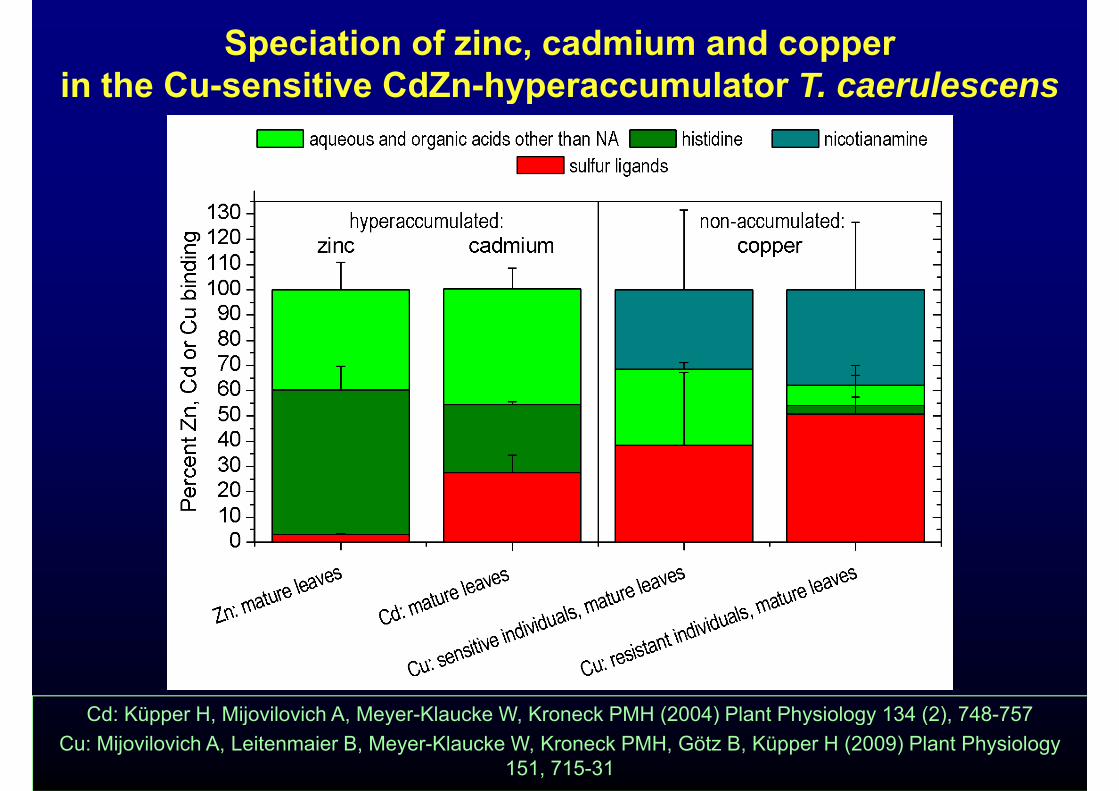
Speciation of zinc, cadmium and copper in the Cu-sensitive CdZn-hyperaccumulator T caerulescensin the Cu-sensitive CdZn-hyperaccumulator T. caerulescens
Cd: Küpper H, Mijovilovich A, Meyer-Klaucke W, Kroneck PMH (2004) Plant Physiology 134 (2), 748-757Cu: Mijovilovich A, Leitenmaier B, Meyer-Klaucke W, Kroneck PMH, Götz B, Küpper H (2009) Plant Physiology
151, 715-31

Compartmentation of metals in leavesZn/Cd/Ni accumulation in epidermal vacuolesZn/Cd/Ni accumulation in epidermal vacuoles
of Thlaspi and Alyssum speciesepidermis
6
8 young leaves
mM
Ca vacuole
p
mesophyll
2
4
mM
/ m
upper lower
8mature leaves
Zn Mg P S Cl K0
4
6
M /
mM
Ca
Zn Mg P S Cl K0
2mM
id i h llupper epidermis upper mesophylllower mesophyll lower epidermis
Concentrations of elements in leaf tissues Thlaspi caerulescens
Zn K α line scan and dot map of a T caerulescens leaf
Ni K α line scan and dot map of a A bertolonii leaftissues Thlaspi caerulescens of a T. caerulescens leaf of a A. bertolonii leaf
Zn: Küpper H, Zhao F, McGrath SP (1999) Plant Physiol 119, 305-11Ni: Küpper H, Lombi E, Zhao FJ, Wieshammer G, McGrath SP (2001) J Exp Bot 52 (365), 2291-2300
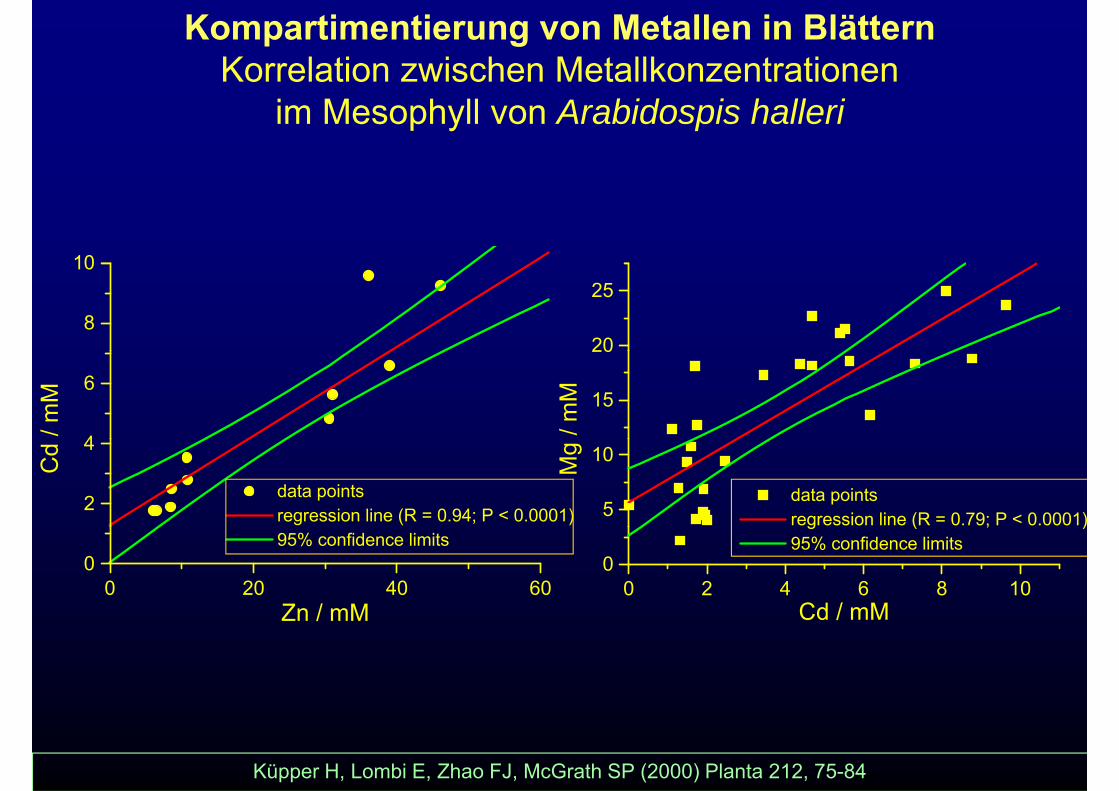
Kompartimentierung von Metallen in BlätternKorrelation zwischen Metallkonzentrationen
im Mesophyll von Arabidospis halleri
10
20
258
10
15
20
/ mM
4
6
/ mM
5
10
data pointsregression line (R = 0.79; P < 0.0001)
Mg
2
4
data points regression line (R = 0.94; P < 0.0001)
Cd
0 2 4 6 8 100
g ( ; ) 95% confidence limits
Cd / mM0 20 40 60
0 95% confidence limits
Zn / mM Cd / mMZn / mM
Küpper H, Lombi E, Zhao FJ, McGrath SP (2000) Planta 212, 75-84
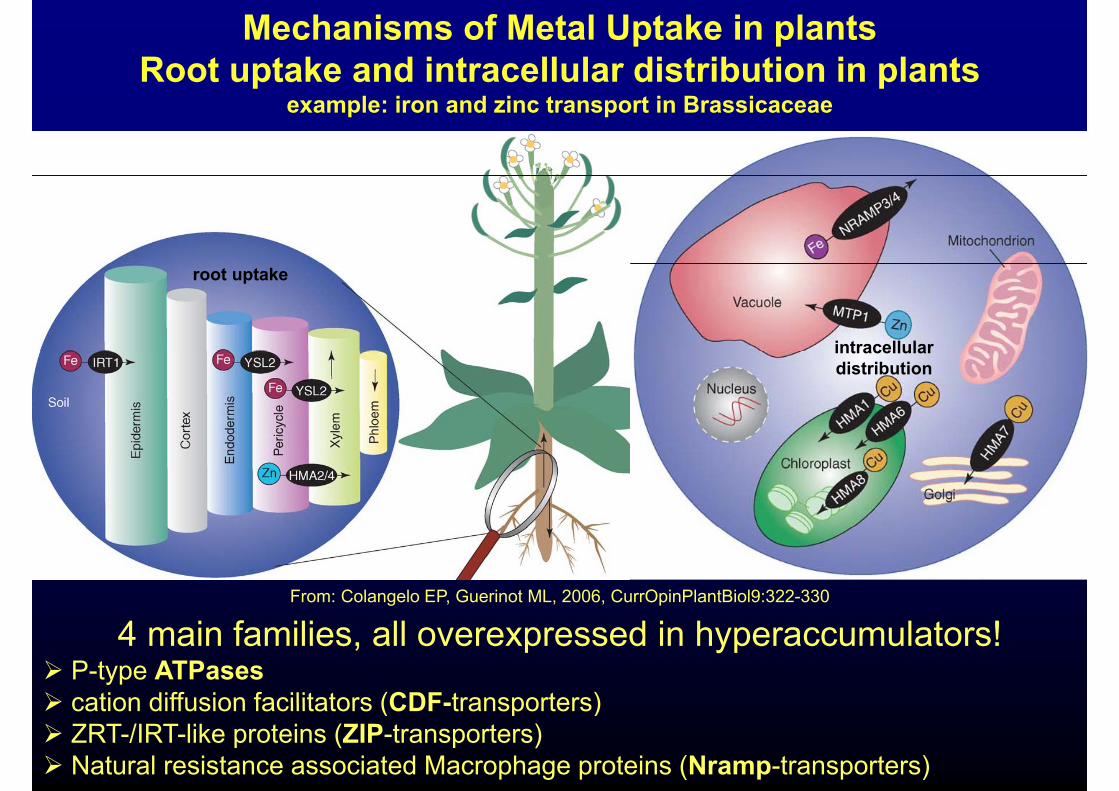
Mechanisms of Metal Uptake in plantsRoot uptake and intracellular distribution in plantsp p
example: iron and zinc transport in Brassicaceae
root uptake
intracellularintracellular distribution
From: Colangelo EP, Guerinot ML, 2006, CurrOpinPlantBiol9:322-330
4 main families, all overexpressed in hyperaccumulators! P-type ATPases cation diffusion facilitators (CDF-transporters) cation diffusion facilitators (CDF-transporters) ZRT-/IRT-like proteins (ZIP-transporters) Natural resistance associated Macrophage proteins (Nramp-transporters)
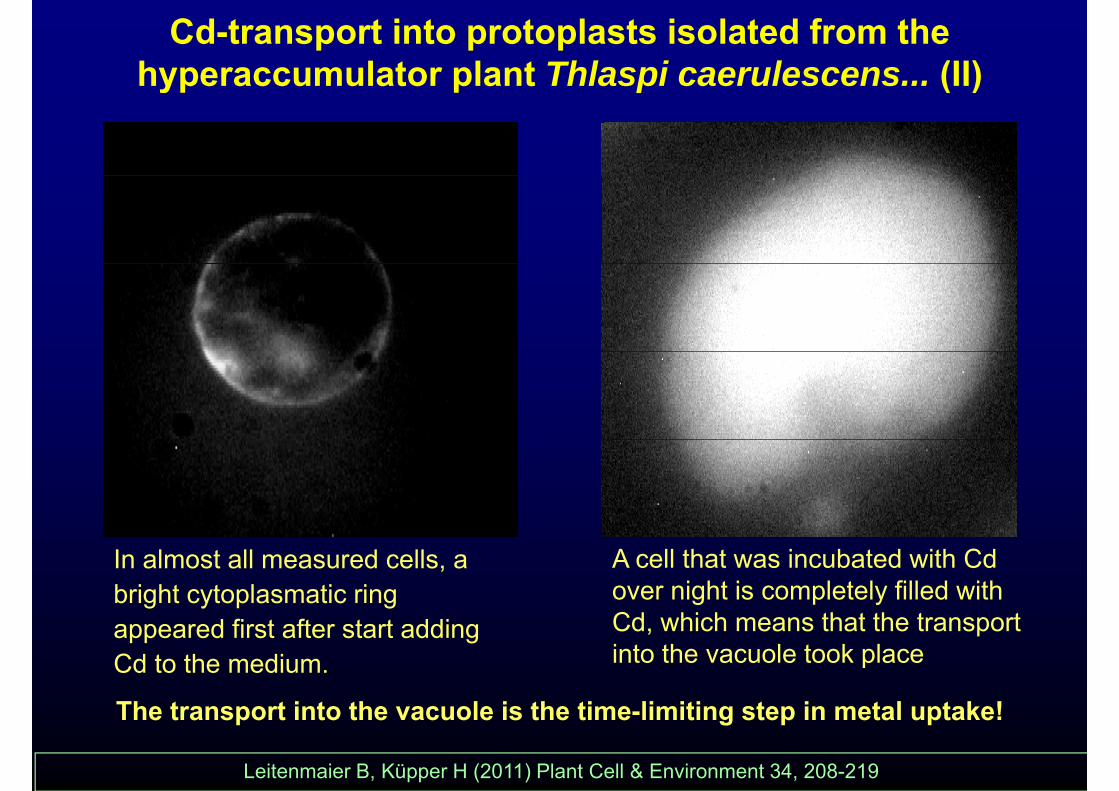
Cd-transport into protoplasts isolated from the hyperaccumulator plant Thlaspi caerulescens... (II)yp p p ( )
In almost all measured cells, a bright cytoplasmatic ring
A cell that was incubated with Cd over night is completely filled with Cd hi h th t th t tappeared first after start adding
Cd to the medium.Cd, which means that the transport into the vacuole took place
Th t t i t th l i th ti li iti t i t l t k !The transport into the vacuole is the time-limiting step in metal uptake!
Leitenmaier B, Küpper H (2011) Plant Cell & Environment 34, 208-219
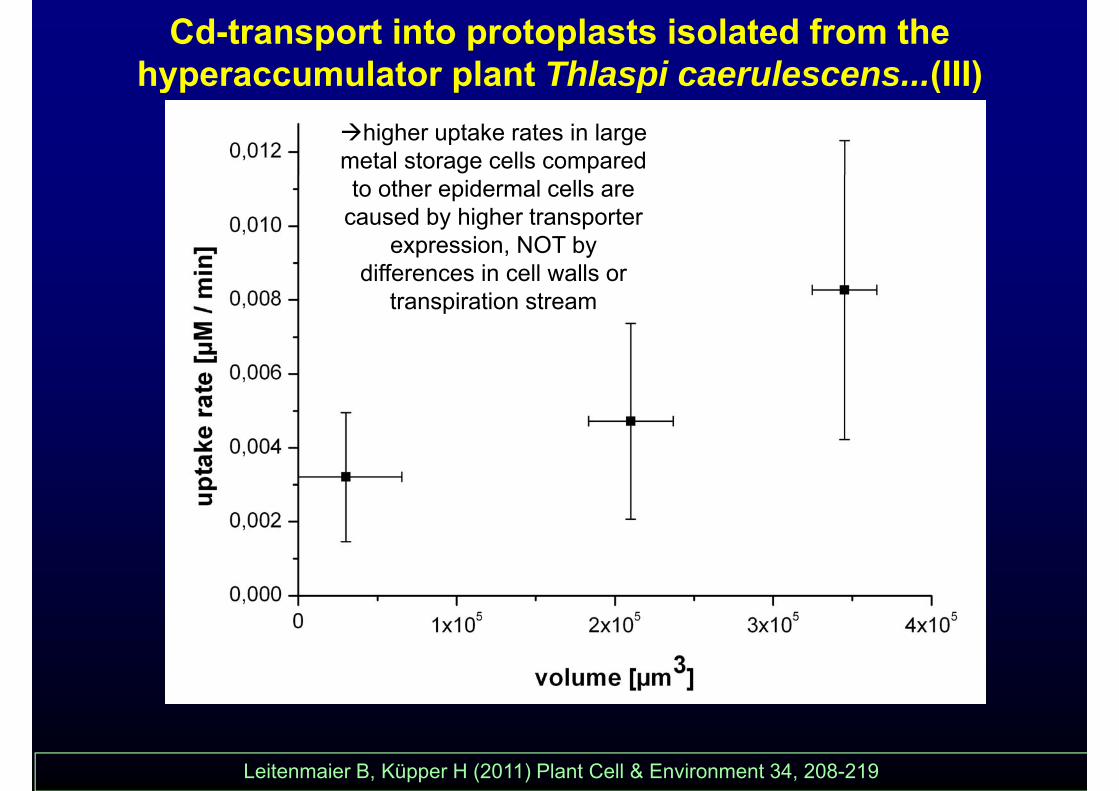
Cd-transport into protoplasts isolated from the hyperaccumulator plant Thlaspi caerulescens...(III)yp p p ( )
higher uptake rates in large metal storage cells compared to other epidermal cells are
caused by higher transporter expression, NOT by
differences in cell walls or transpiration stream
Leitenmaier B, Küpper H (2011) Plant Cell & Environment 34, 208-219
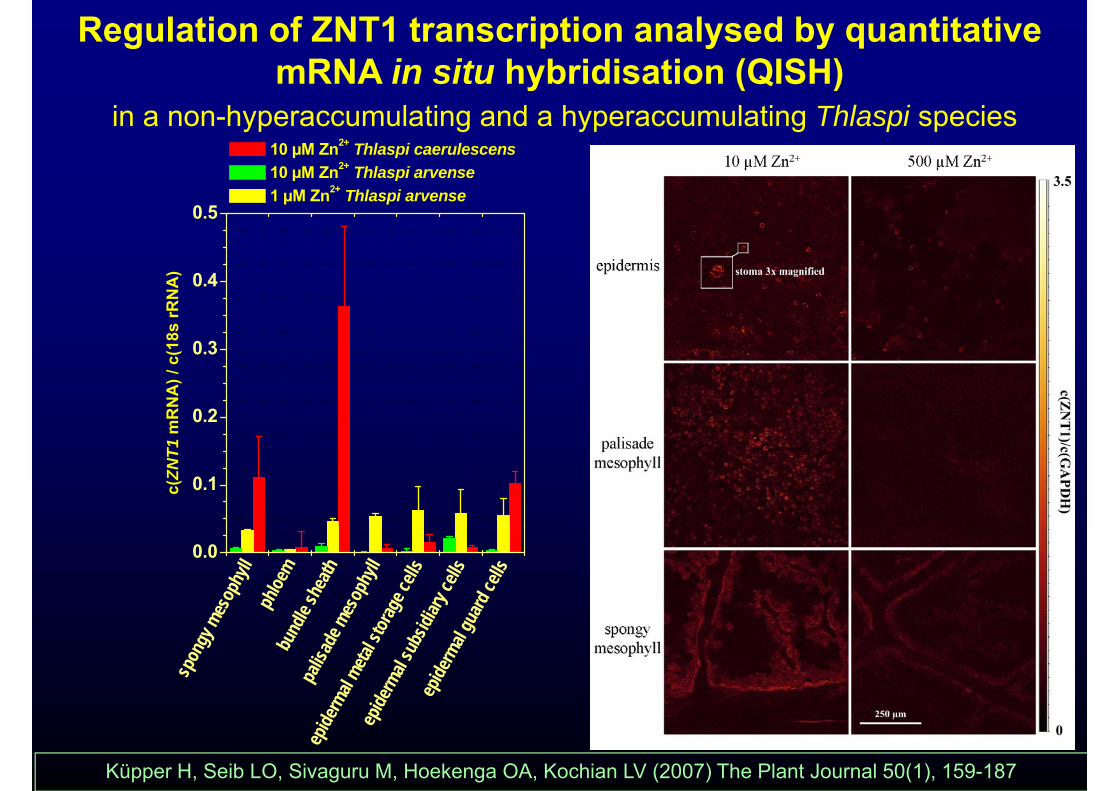
Regulation of ZNT1 transcription analysed by quantitative mRNA in situ hybridisation (QISH)y ( )
in a non-hyperaccumulating and a hyperaccumulating Thlaspi species 10 µM Zn2+ Thlaspi caerulescens10 µM Zn2+ Thlaspi arvense
0.5
10 µM Zn Thlaspi arvense 1 µM Zn2+ Thlaspi arvense
0 3
0.4
(18s
rRN
A)
0.2
0.3
mR
NA
) / c
(
0.1c(ZN
T1
mesop
hyll
phloe
mdle
shea
thmes
ophy
llrag
e cell
sdia
ry ce
llsua
rd ce
lls
0.0
spon
gy m
e
bund
lepa
lisad
e me
rmal
metal s
tora
iderm
al su
bsidi
a ep
iderm
al gu
a
Küpper H, Seib LO, Sivaguru M, Hoekenga OA, Kochian LV (2007) The Plant Journal 50(1), 159-187
epide
rmep
id
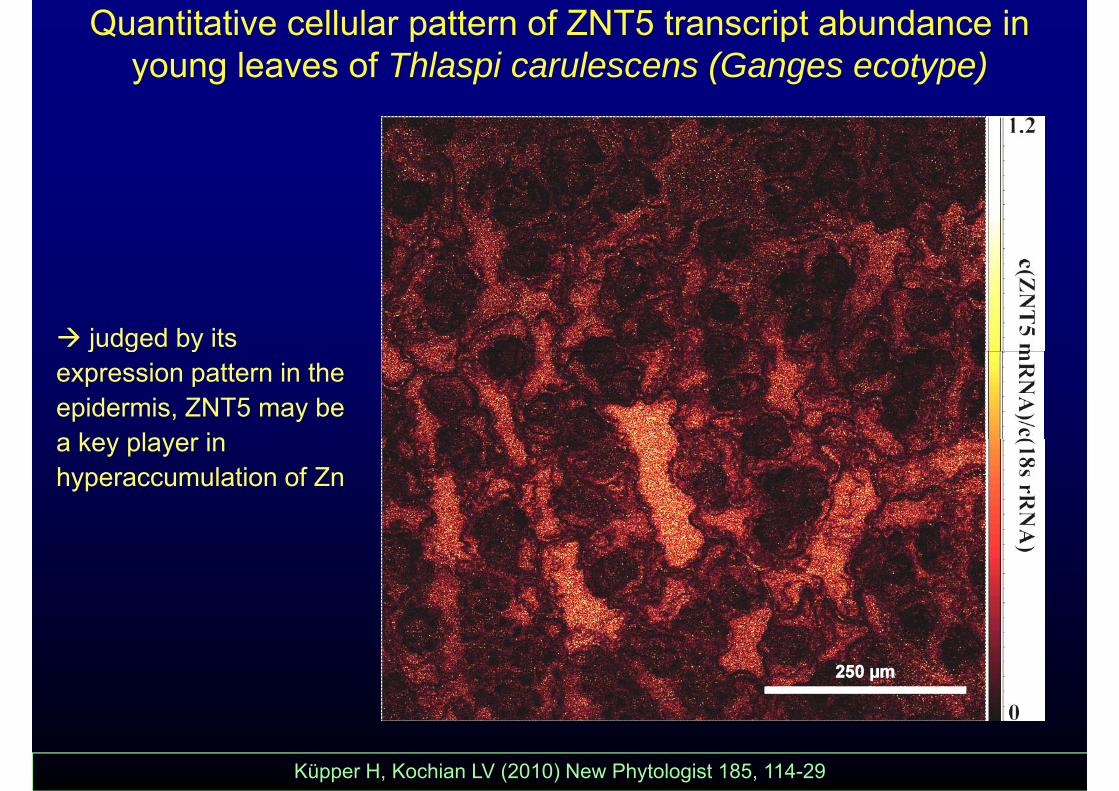
Quantitative cellular pattern of ZNT5 transcript abundance in young leaves of Thlaspi carulescens (Ganges ecotype)
judged by its j g yexpression pattern in the epidermis, ZNT5 may be a key player ina key player in hyperaccumulation of Zn
Küpper H, Kochian LV (2010) New Phytologist 185, 114-29
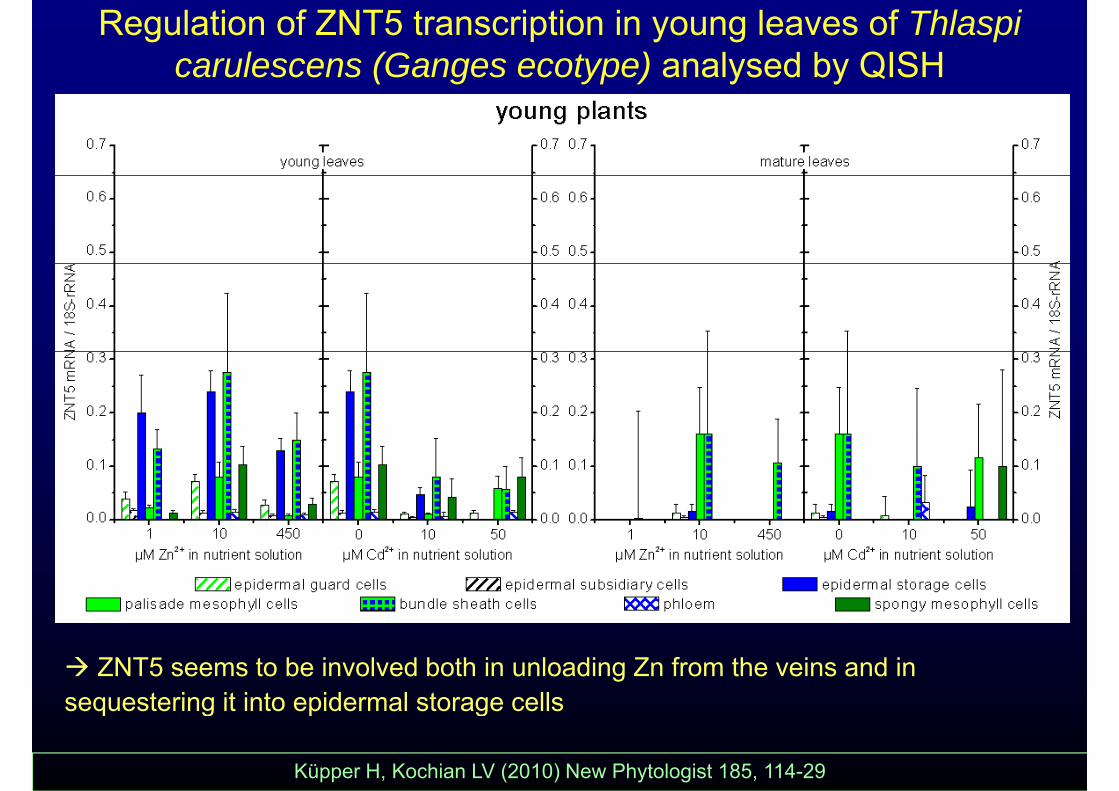
Regulation of ZNT5 transcription in young leaves of Thlaspi carulescens (Ganges ecotype) analysed by QISH
ZNT5 seems to be involved both in unloading Zn from the veins and in sequestering it into epidermal storage cellssequestering it into epidermal storage cells
Küpper H, Kochian LV (2010) New Phytologist 185, 114-29
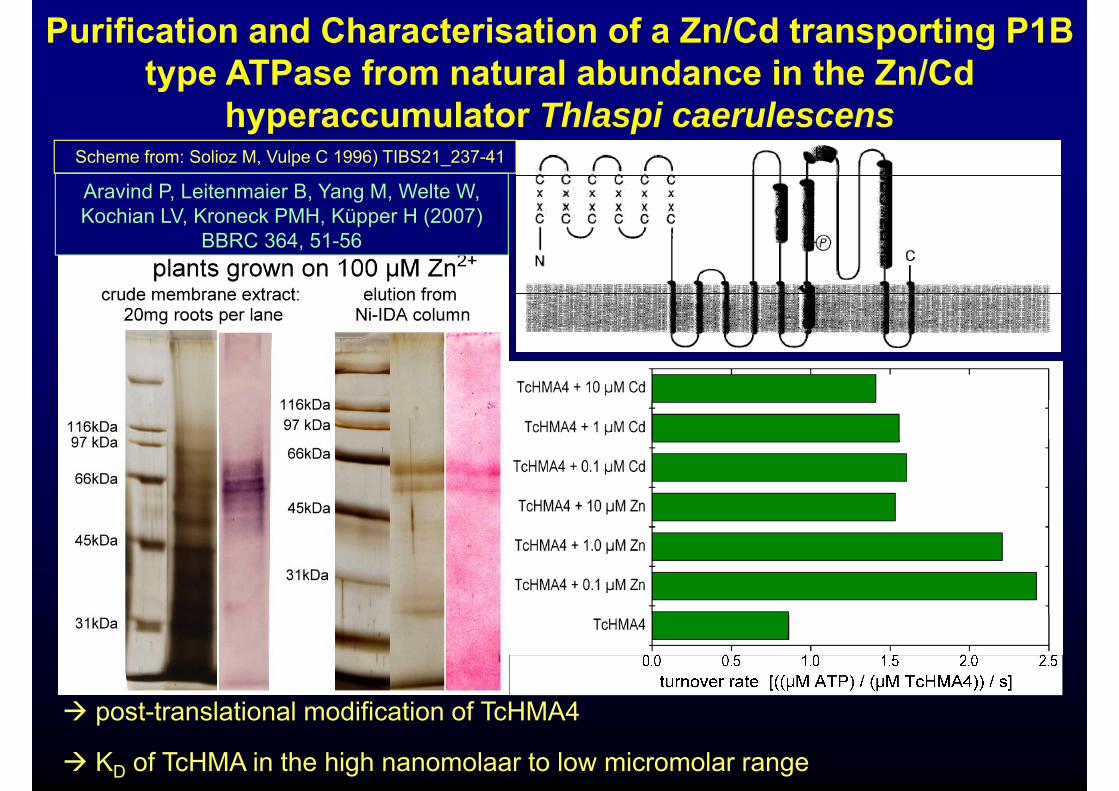
Purification and Characterisation of a Zn/Cd transporting P1B type ATPase from natural abundance in the Zn/Cd
Scheme from: Solioz M, Vulpe C 1996) TIBS21_237-41
yphyperaccumulator Thlaspi caerulescens
Aravind P, Leitenmaier B, Yang M, Welte W, Kochian LV, Kroneck PMH, Küpper H (2007)
BBRC 364, 51-56
t t l ti l difi ti f T HMA4 post-translational modification of TcHMA4
KD of TcHMA in the high nanomolaar to low micromolar range
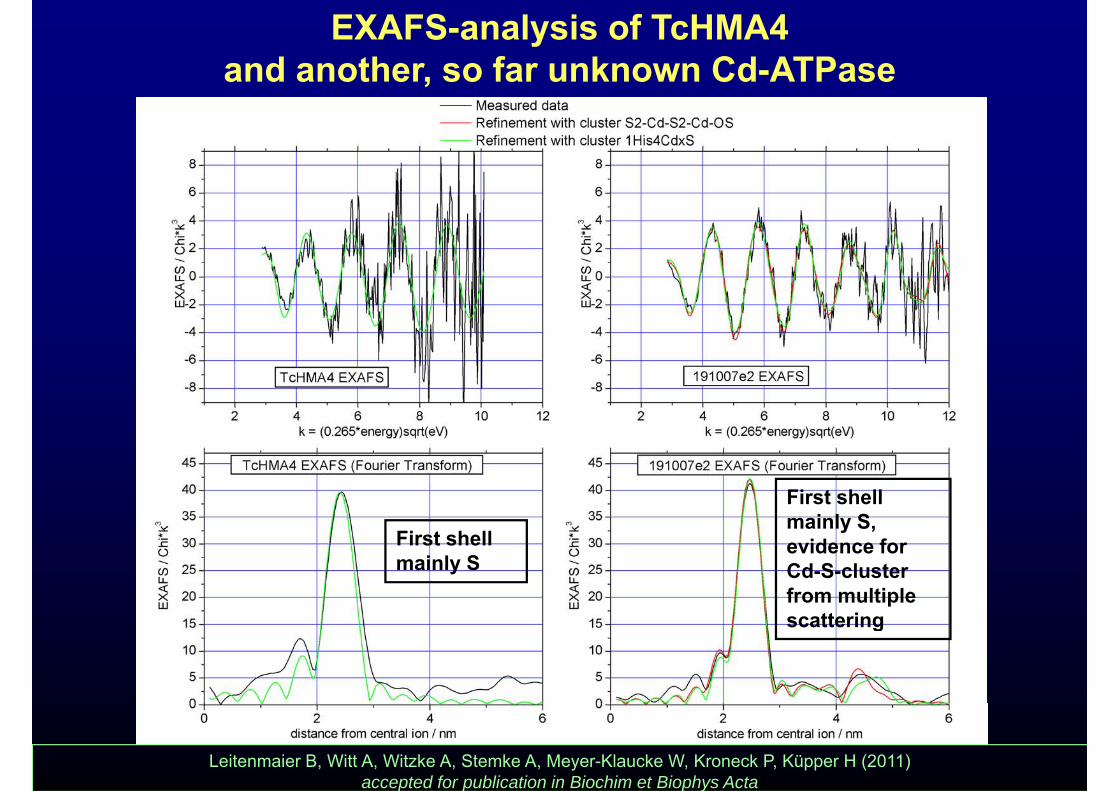
EXAFS-analysis of TcHMA4 and another, so far unknown Cd-ATPase
First shell mainly S,
First shell mainly S
mainly S, evidence for Cd-S-cluster from multiple
tt iscattering
Leitenmaier B, Witt A, Witzke A, Stemke A, Meyer-Klaucke W, Kroneck P, Küpper H (2011) accepted for publication in Biochim et Biophys Acta
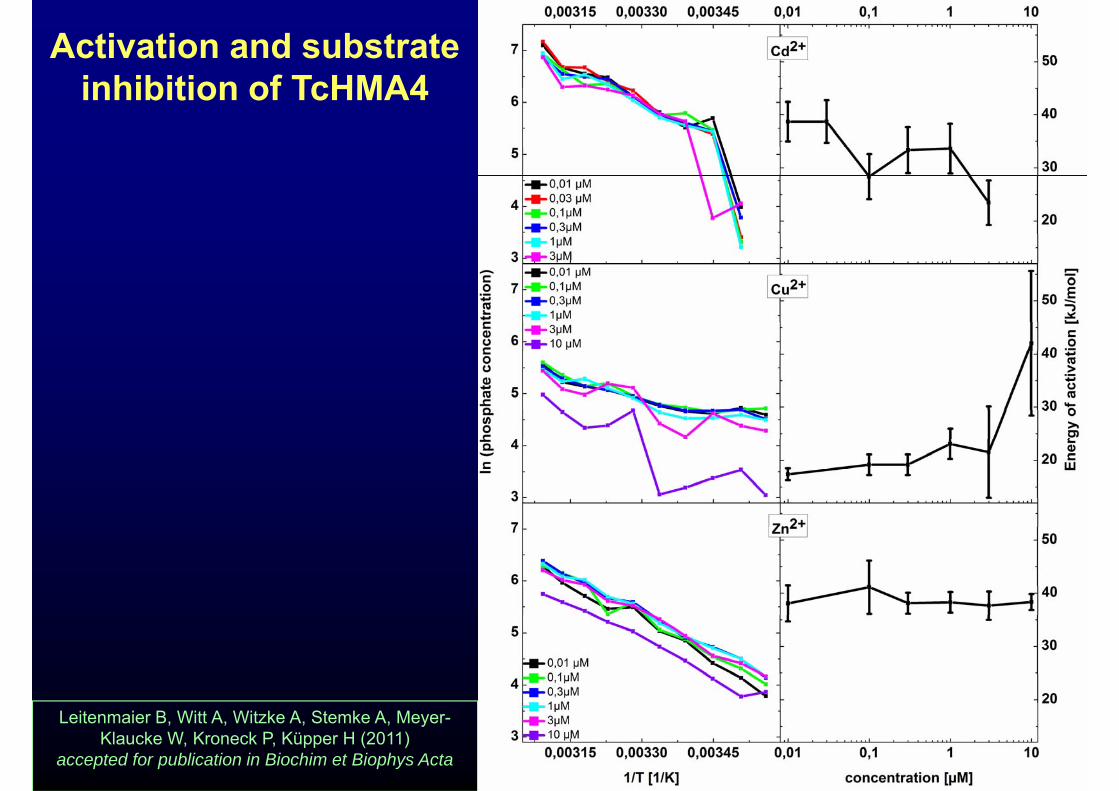
Activation and substrate inhibition of TcHMA4inhibition of TcHMA4
Leitenmaier B, Witt A, Witzke A, Stemke A, Meyer-Klaucke W, Kroneck P, Küpper H (2011)
accepted for publication in Biochim et Biophys Acta
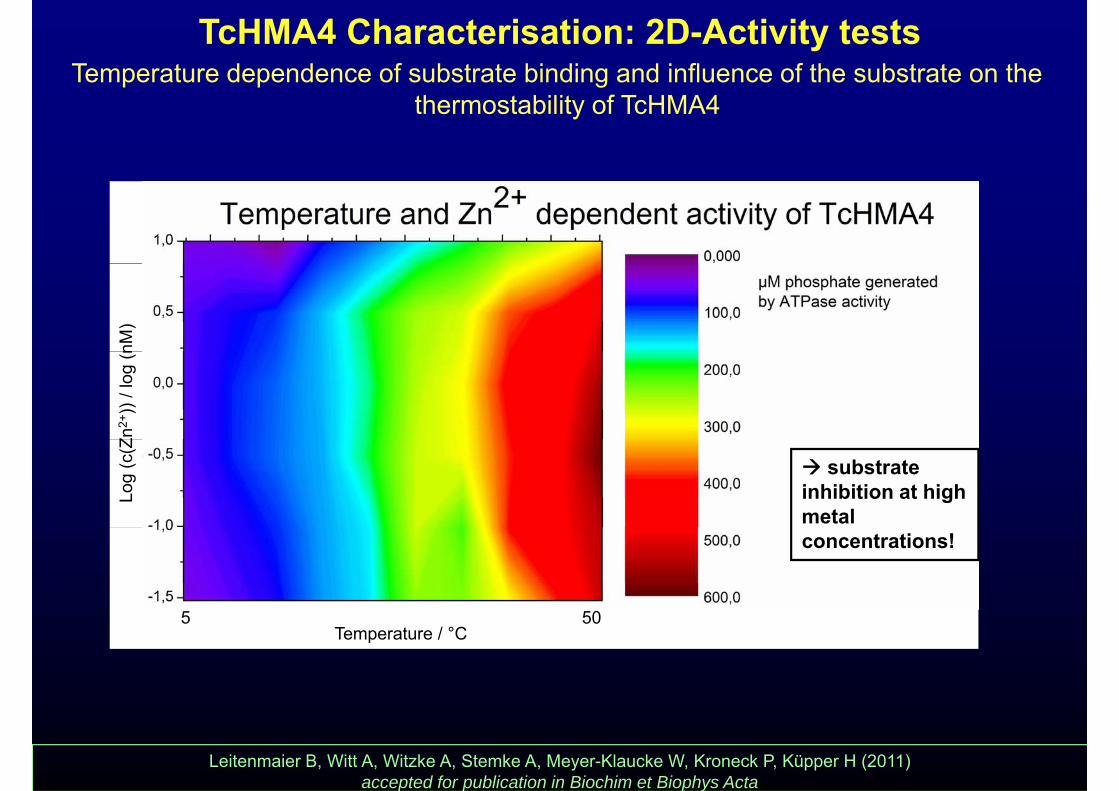
TcHMA4 Characterisation: 2D-Activity testsTemperature dependence of substrate binding and influence of the substrate on the
thermostability of TcHMA4nM
)Zn
2+))
/ log
(nLo
g (c
(Z
substrate inhibition at high metal
5 50
concentrations!
Temperature / °C5 50
Leitenmaier B, Witt A, Witzke A, Stemke A, Meyer-Klaucke W, Kroneck P, Küpper H (2011) accepted for publication in Biochim et Biophys Acta

All slides of my lectures can be downloaded from my k hworkgroup homepage
ni konstan de Department of Biolog Workgro ps Küpper labwww.uni-konstanz.de Department of Biology Workgroups Küpper lab,
or directlyy
http://www.uni-konstanz.de/FuF/Bio/kuepper/Homepage/AG_Kuepper_Homepage.html











![Heavy metal resistance VK2012.ppt [Kompatibilitätsmodus]](https://img.pdfslide.net/doc/110x75/61e3b638aab84b616e27a16f/heavy-metal-resistance-kompatibilittsmodus.jpg)



![[Heavy Metal Music] Heavy Metal Music(BookFi.org)](https://img.pdfslide.net/doc/110x75/577cd2401a28ab9e78954a73/heavy-metal-music-heavy-metal-musicbookfiorg.jpg)
