Embed Size (px)
Citation preview

Update on Weed Control Agents: Path to Modern Agriculture
Herbicides as Weed Control Agents: State of the Art:I. Weed Control Research and Safener Technology:The Path to Modern Agriculture
Hansjoerg Kraehmer*, Bernd Laber, Chris Rosinger, and Arno Schulz
Bayer CropScience AG, D–65926 Frankfurt am Main, Germany
The purpose of modern industrial herbicides is to control weeds. The species of weeds that plague crops today are a consequenceof the historical past, being related to the history of the evolution of crops and farming practices. Chemical weed control beganover a century ago with inorganic compounds and transitioned to the age of organic herbicides. Targeted herbicide research hascreated a steady stream of successful products. However, safeners have proven to be more difficult to find. Once found, themode of action of the safener must be determined, partly to help in the discovery of further compounds within the same class.However, mounting regulatory and economic pressure has changed the industry completely, making it harder to find asuccessful herbicide. Herbicide resistance has also become a major problem, increasing the difficulty of controlling weeds. As aresult, the development of new molecules has become a rare event today.
Modern industrial herbicide research begins with theanalysis and definition of research objectives. A majorpart of this lies in the definition of economically im-portant weeds in major arable crops (Kraehmer, 2012).Weed associations change slowly over time. It is im-portant, therefore, to foresee such changes. Today’sweed associations result from events in the distant past.They are associated with the history of crops and theevolution of farm management. In Europe and theAmericas, some large-acre crops such as winter oilseedrape and spring oilseed rape (canola), both derivedfrom Brassica spp., and soybean (Glycine max) haveattained their current importance only within the last100 years. Other Old World crops, such as cereals, haveexpanded over a very long time span and were alreadyrather widespread in Neolithic times (Zohary et al.,2012). The dominance of crop species in agriculturalhabitats only left room for weed species that couldadapt to cultivation technologies. Changes in cropmanagement and the global weed infestation havehappened in waves. A major early factor in Europewas presumably the grain trade in the Roman period(Erdkamp, 2005). The Romans spread their preferredcrops and, unintentionally, associated weed seedsthroughout Europe, Asia, and Africa. A second wave ofglobal vegetation change started in the 16th centuryafter the discovery of the Americas. Crops and weedswere distributed globally by agronomists and botanists.Alien species started to spread on all continents. A thirdphase can be seen in the 19th century with the indus-trialization of agriculture and the breeding of competi-tive crop varieties. The analysis of weed spectra inarable fields grew from this historical background.
Weeds are plants interfering with the interests of people(Kraehmer and Baur, 2013), which is why they havebeen controlled by farmers for millennia.
Chemical weed control began just about a centuryago with a few inorganic compounds, such as sulfuricacid, copper salts, and sodium chlorate (Cremlyn, 1991).The herbicidal activity of 2,4-dichlorophenoxyaceticacid was detected in the 1940s (Troyer, 2001). Table Iprovides an overview of selected chemical families,selected representatives, and earliest usage reportsaccording to Büchel et al. (1977) and Cremlyn (1991),Worthington and Hance (1991). Targeted herbicide re-search began in the 1950s. In the early days, herbicidecandidates progressed from screens purely on the basisof their having biology that would satisfy farmers’ re-quirements. Mode of action (MoA) studies did not playa major role in the chemical industry prior to the 1970s.Analytical tools were developed and the rapid elucida-tion of plant pathways and in vitro-based screen assayswere used from the 1980s onward. However, in the 1990sand beyond, ever-increasing regulatory and economicpressures have changed the situation of the industrycompletely, and to satisfy the new requirements, selectioncriteria beyond biological activity have needed to be ap-plied. Herbicide resistance in weeds has developed into amore serious problem that now constrains the applicationof certain types of herbicides in somemarkets. Finally, theintroduction of crops resistant to cheap herbicides and ofglyphosate-resistant soybean, in particular, took valueout of the market and resulted in an enormous economicpressure on the herbicide-producing industry. As a resultof this changing and more difficult landscape, the de-velopment of new molecules is now a rare event.
This article is structured into three main topics. First,it provides an historic overview of the development ofweed control history and of screening tools over thepast 100 years. Thereafter, we concentrate on the use of
* Address correspondence to [email protected]/cgi/doi/10.1104/pp.114.241901
Plant Physiology�, November 2014, Vol. 166, pp. 1119–1131, www.plantphysiol.org � 2014 American Society of Plant Biologists. All Rights Reserved. 1119 www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from
Copyright © 2014 American Society of Plant Biologists. All rights reserved.

MoA studies as a tool for optimizing chemical struc-tures based upon knowledge of their receptors. Finally,we review the invention and use of safener technologiesas a tool for improving the crop selectivity of herbicides.In a companion review (Kraehmer, et al., 2014), weaddress the serious challenges that farmers now facebecause of the evolution of herbicide resistance inweeds and the types of innovations that are urgentlyrequired.
AGRICULTURAL CHANGES IN THE PAST ANDTHEIR INFLUENCE ON WEED INFESTATION
Crops Grown on Arable Land within the Last 100 Years,Weed Management, and Changes in Weed Infestation
Crop management practices have had a major impacton weed infestation. Animals such as horses were usedas production tools in the beginning of the last century.
The horses required feeding, and as a result, for exam-ple, in 1920, oats (Avena sativa) were grown on morethan 40 million acres in the United States (area har-vested); today, the acreage amounts to only 1 millionacres. It is not surprising, therefore, that wild oats, whichwere nearly impossible to control in the past, spread soquickly over many continents.
Table II illustrates a few other striking facts: corn (Zeamays) was already planted on 101 million U.S. acres in1920 and on approximately 97 million acres in 2012. Insharp contrast, soybeans were cultivated on less than500,000 acres in 1920 but on 76 million acres in 2012.
One very effective weed management tool is tillage.Mechanical weed management, however, is time con-suming, labor intensive, and leads to a high energyconsumption. Regrettably, it has also resulted in majorerosion problems all over the world (Montgomery,2007). Erosion rates on U.S. arable land declined con-siderably between 1982 and 1987: from 3.06 billion tons
Table I. History of chemical weed control innovations
Post, Postemergence application; Pre, preemergence application, based on data from Cremlyn (1991), Worthington and Hance (1991), Buchelet al. (1977), Herbicide Resistance Action Committee (www.hracglobal.com), and others.
MoA, Target Site Chemical Family Examples Use Earliest Reports
Unspecific Inorganic herbicides H2SO4, Cu2SO4, FeSO4, NaAsO2 Total 1874Uncouplers Dinitrophenoles dinitro-ortho-cresol Post, dicots 1934Auxins Aryloxyalkanoic acid
derivatives2,4-Dichlorophenoxyacetic acid Post, dicots in cereals 1942
Microtubule organization Arylcarbamates Propham, chloropropham Pre, monocots in various crops 1946Lipid synthesis Chloroaliphatic acids TCA, dalapon Pre, monocots in various crops 1947
Thiocarbamates EPTC, triallate Pre, monocots and dicots invarious crops
1954
PSII Arylureas Monuron, diuron, isoproturon,linuron
Pre and Post, monocots anddicots in various crops
1951
1,3,5-Triazines Atrazine, simazine Pre and Post, broad spectrumin corn
1952
Pyridazines Chloridazon Pre, dicots in sugar beet 1962Uracils Bromacil, terbacil, lenacil Soil applied, broad spectrum
in various crops1963
Biscarbamates Phenmedipham Post, dicots in sugar beet 19681,2,4-Triazinones Metribuzin Pre in soybean 1971
Very-long-chain fatty acidbiosynthesis
Chloroacetamides Allidochlor, alachlor Pre, monocots and dicots 1956
PSI Bipyridyliums Diquat, paraquat Nonselective 1958Protoporphyrinogen oxidase Diphenyl ethers Nitrofen, acifluorfen Pre and Post, various crops 1960
Oxadiazoles Oxadiazon Rice, nonselective 1969Microtubule assembly Dinitroanilines Trifluralin, pendimethalin Pre against monocots and dicots 1960Cellulose biosynthesis Nitriles Dichlobenil Plantations 19605-Enolpyruvylshikimate
3-phosphate synthaseGlys Glyphosate Post, nonselective 1971
Phytoene desaturase Pyridazinones Norflurazon Pre and Post in cotton 1973ACCase Aryloxyphenoxy
propanoatesDiclofop, fluazifop Post, grasses 1975
Cyclohexane diones Alloxydim, sethoxydim Post, grasses 1976Gln synthetase Glufosinate Nonselective 1981AHAS or ALS Sulfonylureas Chlorsulfuron, metsulfuron Monocots and dicots in various
crops1982
Imidazolinones Imazapyr, imazethapyr Nonselective or selective insoybean
1983
Pyrimidinyl benzoates Bispyribac sodium Rice 1994HPPD Pyrazolynate, sulcotrione Various crops, monocots and
dicots1984
1120 Plant Physiol. Vol. 166, 2014
Kraehmer et al.
www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

down to 1.73 billion tons, or from 7.3 tons per acre to4.8 tons per acre (National Resources Inventory; http://www.nrcs.usda.gov/Internet/FSE_DOCUMENTS/nrcs143_012269.pdf). Changes in tillage practices werepartly responsible for this achievement.Weed infestation in spring crops and winter crops can
differ considerably (Håkanson, 2003). The profoundreduction of spring crops, and especially the reductionof the oat acreage when tractors replaced horses,resulted in a complete shift from wild oats (Avena fatua)as a major grass weed to blackgrass (Alopecurus myo-suroides) and silky bentgrass (Apera spica-venti) inEurope (Kraehmer and Stuebler, 2012). The hard win-ters in Canada prevent the cultivation of winter crops inmany agricultural areas. This is why winter annualssuch as blackgrass do not play a major role there.Canada has a long tradition of conducting weed sur-veys. Despite major changes in the use of agrochemi-cals, the most dominant weeds have remained the samefor decades (Leeson et al., 2005). The Weed MappingWorking Group of the European Weed Science Societyendeavors to map the most common weeds of Europein all major crops and to document changes inweed infestations (http://www.ewrs.org/weed_mapping.asp). One obvious result is that weed spectra changewith cropping practices and environmental conditionsand that some species are better adapted to the warmerclimates in the Mediterranean area, such as Abutilontheophrasti or Sorghum halepense, whereas others aremore frequent in northern Europe, such as Alopecurusmyosuroides or Apera spica-venti. The weed species Che-nopodium album and Echinochloa crus-galli are character-ized by a wide ecological aptitude (Kraehmer, 2010) andcan be found everywhere in Europe and in many crops.
BIOLOGICAL SCREENING
Glasshouse and Field Screening
In the early days, indicator species or model plantsplayed a major role in herbicide discovery screening.
Weed target species were often not available. Mostherbicide screening in the agrochemical industry (Fig. 1)between the 1950s and 1980s was characterized by aprotocol where solutions of test chemicals were sprayedover sets of plants in pots or over seeded bare soil in thecase of preemergent screens.
The screening compounds were either predissolvedin solvents such as acetone or they were formulated aswettable powders, emulsion concentrates, or suspen-sion concentrates before dilution in water. The sprayingprocess simulated the actual spraying situation in thefield. Every company had its own specially designedspraying equipment. The screening process was di-vided into two to four initial selection steps. The initial,or plus-minus activity, tests started with high doserates, generally between 1 and 10 kg active ingredientha21. Compounds were sprayed in preemergence andpostemergence trials. Later steps with lower dose ratesand varying plant species followed to further refineactivity and the weed spectrum. The results of thesetrials were usually visual phytotoxicity ratings on dif-ferent scales, such as on a 0% to 100% rating scale, 0%meaning no phytotoxicity and 100% equal to completecontrol of plants. Fresh or dry weights were used forcomparisons in special tests only. Symptomology wasan essential and integral part of ratings. Each chemicalclass of compounds usually resulted in typical patternsof symptoms. Observations made in different screeningindications (e.g. insecticide, acaricide, nematicide, orfungicide screens) were also taken into account duringevaluation (Figs. 2 and 3).
Chemists usually prepared sample amounts of 3 to5 g. This amount allowed the early testing of chemicalsin all screening indications. Most midsize and largecompanies tested between 1,000 and 5,000 compoundsper year. Information technology tools suitable to fa-cilitate screening were not available before the 1980s,
Figure 1. Plants in pots. Right, Untreated; left, experimental herbicideat 300 g active ingredient ha21.
Table II. Acreage changes in U.S. crops during the last 100 years
Values shown are million acres harvested. Source: U.S. Departmentof Agriculture Crop Production Historical Track Records, April 2013.
Crop 1920 1970 2012
Barley 7.5 9.7 3.2Canola ,0.1 ,0.1 1.6Corn 101.4 66.1 97.2Cotton 34.4 11.2 9.4Hay 73.1 61.5 56.3Oat 42.8 18.6 1.1Potato 3.3 1.5 1.2Rice 1.3 1.9 2.7Rye 4.9 1.5 0.3Soybean ,0.5 42.3 76.1Sugar beet 0.9 1.5 1.2Wheat 62.4 43.6 49All crops 347 283 309
Plant Physiol. Vol. 166, 2014 1121
Herbicides State of the Art: Overview
www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

and structure-activity relationships had to be derivedmanually in long and time-consuming procedures. Thelast glasshouse step was a profiling procedure in whichrecommendations for field testing were derived. Often,special tests were carried out before or in parallel tofield tests to check the soil-dependent performance ofa compound, the influence of different formulations,potential carryover risks, and crop selectivity ranges.Depending on company size and resources, between10 and 100 compounds were advanced into field testingper year. Compounds were tested in different parts ofthe world on plots between 1 and 10 m2 with three tofour replicates.
Two to 3 years in the field were usually sufficient tomake informed development decisions. In the thirdquarter of the last century, it usually took between4 and 6 years from the first synthesis of a compound untilentry into the market. Later, the development processbecame more involved and took a greater investment oftime to develop a new herbicide, as we will show later.
Early MoA Studies
The number of MoAs was quite limited between 1950and 1970. Most commercial products were character-ized as auxin-type herbicides, PSI or PSII inhibitors, orinhibitors of cell division. During the 1970s, inhibitionof photosynthesis was typically tested at the whole-plant level using infrared gas analyzers in growthchambers. At the biochemical level, the so-called Hillreaction served as a tool for the identification of pho-tosynthesis inhibitors in isolated chloroplasts (Arndtand Kötter, 1968). In the 1980s, Clark electrode mea-surements with isolated leaf cells and fluorescence emis-sion assays with whole leaves provided furthernondestructive tools for the characterization of PSII her-bicides (Voss et al., 1984a, 1984b).
Plant tissue cultures coupled with HPLC analysishave been used since the 1970s to characterize herbicide-induced changes in metabolism. Cultures of aquaticplants such as Lemna minor have also long been used as
an herbicide test system. Perennial weeds sometimesrequired a different approach. Systemic action of her-bicides is required for the control of perennials such ascouch (Elytrigia repens), johnsongrass (Sorghum hale-pense), or field bindweed (Convolvulus arvensis). Specialtranslocation tests were used to check for this property.For example, Phaseolus spp. beans with two leavesprovided some indication of systemic action when oneof the leaves was treated. Translocated compounds thenled to symptoms in the untreated leaf.
From the 1980s onward, special biochemical assayswere developed for the characterization of target sitesand for binding studies. Target enzymes of plant-specificpathways were preferred in order to avoid toxicologicalproblems in mammals. We will come to this period inone of the following paragraphs.
Miniaturized Screening Assays
A perennial problem faced by the agrochemical in-dustry has been the question of where to obtain new
Figure 3. Typical auxin-transport inhibitor symptoms in a nematodescreening assay.
Figure 2. Typical symptoms of HPPD bleachers in an advancedscreening stage.
1122 Plant Physiol. Vol. 166, 2014
Kraehmer et al.
www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

compounds in amounts sufficient for screening. At theend of the last century, an intensive exchange of com-pounds between nonagrochemical and agrochemicalorigins began. Universities and private institutes offeredagrochemical companies their stocks and undertookthe synthesis of additional desired compounds. Naturalproducts were available for testing also. Since mostthird-party compounds were provided in low amounts,it drove the adoption of miniaturized herbicide dis-covery assays. These assays provide a basic initial testfor phytotoxicity but give only limited discriminatoryinformation to guide compound optimization. Seedgermination assays in petri dishes were also quitecommonly used. The first purpose of these assayswas to find hits that would justify the synthesis of an-alogs and of samples with larger substance amounts.Tissue culture tests as described by Gressel et al. (1993)were also employed by a number of companies. Somecompanies have kept and improved such miniaturizedassays until today, as published by Grossmann et al.(2012). Several assays were incorporated into high-throughput screening systems that allow the screeningof one million compounds per year and more (Kraehmer,2012). We will touch upon this screening approach in oneof the following sections.
Glasshouse-to-Field Transfer
One disadvantage of early screening sets of the lastcentury was the use of model plants such as onion(Allium cepa), carrot (Daucus carota), potato (Solanumtuberosum), and others. They were employed becausethe seed of target weeds was not available in the earlyherbicide screens. Today, specialized providers can de-liver high-quality weed seed in guaranteed quantities.Nevertheless, even given the use of real target weeds,laboratory and glasshouse data are still not usually fullypredictive of the performance of compounds in the field.Many parameters contribute to the variable transferfactors from glasshouse to field (Kraehmer and Russell,1994). One of these parameters is the test species.Model plants such as Lemna spp., Arabidopsis (Arabi-dopsis thaliana), and Brachypodium distachyon, differ fromweeds in many ways, including their specific uptakeand translocation properties, metabolism, specific lifecycle, and environmental requirements for growth.Lemna spp. are aquatic weeds in contrast to terrestrialweeds. Arabidopsis and B. distachyon are not seriouscompetitors within arable crops under agriculturalconditions. The same rules apply to many other speciesin miniaturized assays. General phytotoxicity principlescan only rarely be used for the optimization of chemicalstructures or for structure-activity relationships. Toomany positive results in an indicator assay often requirerepeated screening steps to reduce their numbers. Cer-tain types of herbicides may not be detected at all ifthe indicator species is too different from the screenmodel species. For example, acetyl-coenzyme A car-boxylase (ACCase)-inhibiting graminicides would not
have been detected by screening against Arabidopsis.Similarly, the potential of a compound to control pe-rennial weeds will not be evident from screens that onlyinclude annual species, and information about soil-plantinteractions is absent from screens based upon liquidsystems that do not include soil. The potential of acompound to control perennial weeds will often not bedetected with annual species. Plant cells grown in tissuecultures are very different from whole plant cells be-cause they are undifferentiated and can derive nutrients,often present in excess, from the culture medium andare buffered such that the influence of pH values on theuptake of compounds is masked. The actual value of anew compound is always based upon its performanceagainst economically important weed species. It is highlyquestionable if all compounds controlling Lemna spp.,Arabidopsis, and B. distachyon will, for example, auto-matically control herbicide-resistant Amaranthus spp.This is presumably why cells or tissues of target specieshave now found their way into physiological profilingassays (Grossmann et al., 2012).
Environmental Fate of Agrochemicals
Advances in the sensitivity of analytical tools in the1970s and 1980s allowed the detection of trace levelsof agrochemicals in groundwater, and through suchanalyses some chemicals proved to be persistent in theenvironment (Kraehmer, 2012). This led to a tighteningof regulatory requirements and an increased need foragrochemical companies to test and characterize deg-radation rates of compounds in soil and water underaerobic and anaerobic conditions. The physicochemicalparameters such as vapor pressure and the octanol-waterpartition coefficients of new compounds are now mea-sured routinely, and increasingly sophisticated toxicol-ogy tests are carried out before field testing.
How Has Chemical Weed Control Changed AgricultureSo Far?
Herbicide innovations have appeared in waves(Stuebler et al., 2008). The early auxin-type herbicidesprimarily controlled dicot weeds. Later on, monocotscould be controlled with the advent of photosynthesisinhibitors, cell division inhibitors, and very-long-chainfatty acid biosynthesis inhibitors. Yields were consid-erably increased in all crops between 1940 and 2010(Table III). Many factors contributed to this increase;one is definitely breeding, but herbicides also had amajor impact. The value of herbicides was shown byZimdahl (2004), who published data showing the in-fluence of defined weed species on the yield of dif-ferent crops.
Herbicides made it possible to control weeds incrops much more easily than before. Before the adventof herbicides, controlling weeds required hard physi-cal labor such as hoeing. A particular example is weed
Plant Physiol. Vol. 166, 2014 1123
Herbicides State of the Art: Overview
www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

control in sugar beet (Beta vulgaris), where high laborcosts meant that farmers rapidly adopted herbicidetechnology once it became available. Another crop thatcould be cultivated more easily with the invention ofnew herbicides was winter oilseed rape. Broad-spectrumproducts such as paraquat and atrazine accelerated theuse of reduced-tillage measures in agriculture and helpedprevent soil erosion, which had become a major issueespecially in the United States during the first half of thelast century, as mentioned above. Selective postemer-gence grass control in some crops came in the mid-1970s.The ACCase inhibitors selectively controlled grass weedsin postemergence treatments, and the use of safenerseven made their application in cereals possible. Glyphos-ate is a unique molecule with very specific properties. Itallowed the farmer to kill weeds and to plant new seedwithin a few days after its application (similar to para-quat). It killed perennials and had a short soil half-life.Therefore, it was also regarded as one of the most ef-fective products in plantation crops. Glufosinate wasanother nonselective molecule with a new MoA. It wasstrong against a few hard-to-control weeds such as Equi-setum spp., and it appeared to be a bit safer to plan-tation crops than glyphosate. The introduction ofacetolactate synthase (ALS) inhibitors drastically reducedthe total amounts of agrochemicals applied perhectare. Most of them could be applied with a fewgrams per hectare, whereas many older chemicalsrequired amounts in the kilogram range. Finally,4-hydroxphenylpyruvate dioxygenase (HPPD) andphytoene desaturase inhibitors were ideal mixturepartners for existing products and closed some evidentgaps. No major new MoA has been found by the in-dustry since the 1980s. As early as 1990, agrochemicalmarket research indicated that the crop protectionmarket was approaching maturity and that it was be-coming increasingly difficult to discover new agro-chemicals with significant advantages over existingproducts (Kraehmer and Drexler, 2009). The averagetotal costs for the development of an herbicide hadincreased from $50 million to $250 million U.S. be-tween the years 1975 and 1995 (Rüegg et al., 2007).Genetically modified crops entered the market in thesecond half of the 1990s. The high standard of existingproducts in the market and the high registration costscaused many chemical companies to give up theiragrochemical business. Following numerous mergersand acquisitions, only a few companies with herbicideresearch capacity remained. The number of companies
devoted to herbicide discovery was reduced frommore than 40 in the 1970s to five to eight today. It ishighly questionable, therefore, if farmers will experi-ence many new innovations in the years to come, andyet there is an urgent need for new weed-control so-lutions. Several cropping systems in the Americas aswell as in Europe are no longer sustainable withoutfurther herbicide innovations, as we will see in ourfinal sections.
MOAs AND HERBICIDE DIVERSITY
There are several excellent reviews and books on theMoAs of herbicides (Seitz et al., 2003; Liu et al., 2009;Dayan et al., 2010; Krämer at al, 2012); therefore, there isno need to repeat what has been published recently.Instead, it is intended to give a short overview of pre-vious and current trends in MoA research and the fac-tors that affected the search for herbicides with novelMoAs.
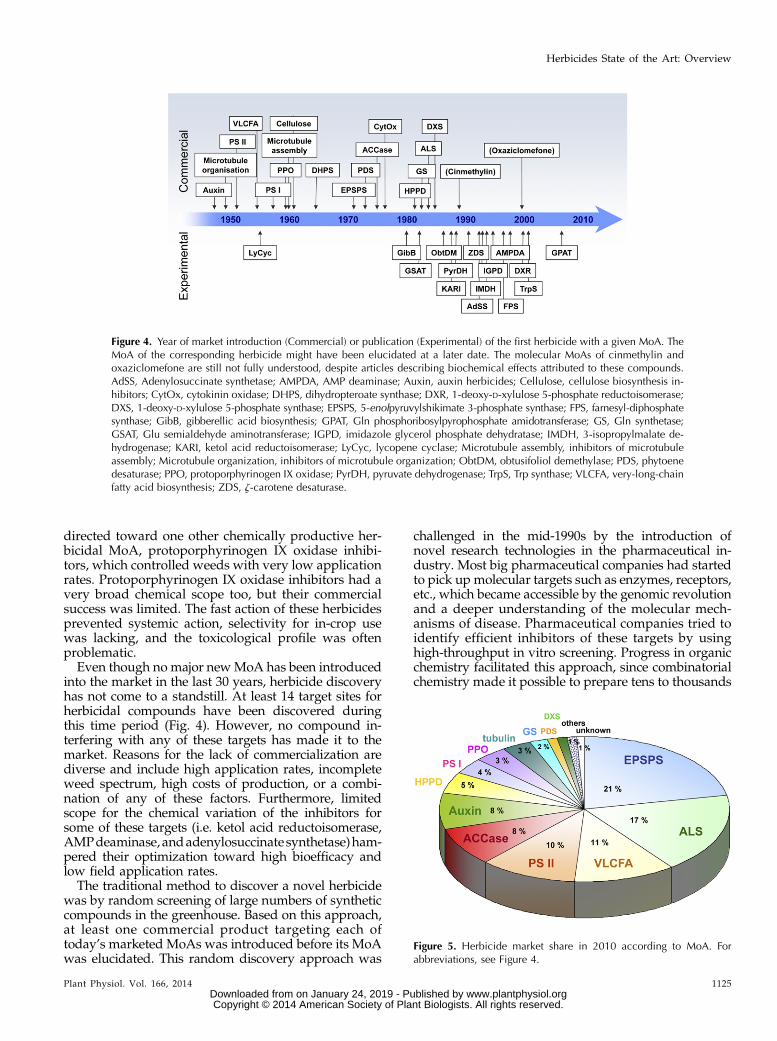
Between 1960 and 1970, only one herbicide with anovel MoA, asulam, an inhibitor of dihydropteroatesynthase, was discovered. In the early 1970s, this periodof low innovation with respect to herbicides with novelMoAs was superseded by what, in retrospect, might becalled the golden years of herbicide discovery. From1971 to 1985, herbicides with eight novel MoAs werediscovered, among them inhibitors of amino acid bio-synthesis (glyphosate, glufosinate, and ALS inhibitors),lipid biosynthesis (ACCase inhibitors), and pigmentbiosynthesis (phytoene desaturase and HPPD inhibi-tors), MoAs that today still dominate the herbicidemarket (Figs. 4 and 5).
These herbicides not only had a profound impact onweed control in agriculture but also played a major rolein expanding the understanding of fundamental plantprocesses through their ingenious use as molecularprobes (Dayan et al., 2010). Many academic groups inthe United States as well as in Europe embarked on thispath, either studying the MoAs of herbicides or usingherbicides to study plant metabolism.
From the mid-1980s until today, more than 140new herbicide active ingredients were commercialized(Gerwick, 2010). Surprisingly, only two of these, thenarrow-spectrum herbicides cinmethylin and oxaziclo-mefone, have an unknown and still not fully understoodmolecular MoA, despite publications describing bio-chemical effects attributed to these compounds. All theother new active ingredients target old (known) MoAs.The reasons for this lack of novelty have been reviewedrecently and have been attributed to a number of mostlyeconomic factors (Kraehmer et al., 2007; Duke, 2012).Two such factors are briefly discussed in the followingparagraphs.
Due to their very low application rates in particular,ALS inhibitors easily outcompeted other chemical clas-ses in the herbicide screening programs and drew sig-nificant synthesis capacity away from them. In additionto ALS inhibitors, significant synthesis capacity was
Table III. Yields per acre of five U.S. crops between 1940 and 2010
Source: U.S. Department of Agriculture Crop Production HistoricalTrack Records, April 2013. b, Bushels; t, tons.
Crop 1940 1950 1960 1970 2010
Barley (b) 23 27.2 31 42.8 73.1Corn (b) 28.9 38.2 54.7 72.4 152.8Soybean (b) 16.2 21.7 23.5 26.7 43.5Sugar beet (t) 13.4 14.6 17.2 18.7 27.7Wheat (b) 15.3 16.5 26.1 31 46.3
1124 Plant Physiol. Vol. 166, 2014
Kraehmer et al.
www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.
directed toward one other chemically productive her-bicidal MoA, protoporphyrinogen IX oxidase inhibi-tors, which controlled weeds with very low applicationrates. Protoporphyrinogen IX oxidase inhibitors had avery broad chemical scope too, but their commercialsuccess was limited. The fast action of these herbicidesprevented systemic action, selectivity for in-crop usewas lacking, and the toxicological profile was oftenproblematic.Even though no major newMoA has been introduced
into the market in the last 30 years, herbicide discoveryhas not come to a standstill. At least 14 target sites forherbicidal compounds have been discovered duringthis time period (Fig. 4). However, no compound in-terfering with any of these targets has made it to themarket. Reasons for the lack of commercialization arediverse and include high application rates, incompleteweed spectrum, high costs of production, or a combi-nation of any of these factors. Furthermore, limitedscope for the chemical variation of the inhibitors forsome of these targets (i.e. ketol acid reductoisomerase,AMPdeaminase, andadenylosuccinate synthetase)ham-pered their optimization toward high bioefficacy andlow field application rates.The traditional method to discover a novel herbicide
was by random screening of large numbers of syntheticcompounds in the greenhouse. Based on this approach,at least one commercial product targeting each oftoday’s marketed MoAs was introduced before its MoAwas elucidated. This random discovery approach was
challenged in the mid-1990s by the introduction ofnovel research technologies in the pharmaceutical in-dustry. Most big pharmaceutical companies had startedto pick up molecular targets such as enzymes, receptors,etc., which became accessible by the genomic revolutionand a deeper understanding of the molecular mech-anisms of disease. Pharmaceutical companies tried toidentify efficient inhibitors of these targets by usinghigh-throughput in vitro screening. Progress in organicchemistry facilitated this approach, since combinatorialchemistry made it possible to prepare tens to thousands
Figure 4. Year of market introduction (Commercial) or publication (Experimental) of the first herbicide with a given MoA. TheMoA of the corresponding herbicide might have been elucidated at a later date. The molecular MoAs of cinmethylin andoxaziclomefone are still not fully understood, despite articles describing biochemical effects attributed to these compounds.AdSS, Adenylosuccinate synthetase; AMPDA, AMP deaminase; Auxin, auxin herbicides; Cellulose, cellulose biosynthesis in-hibitors; CytOx, cytokinin oxidase; DHPS, dihydropteroate synthase; DXR, 1-deoxy-D-xylulose 5-phosphate reductoisomerase;DXS, 1-deoxy-D-xylulose 5-phosphate synthase; EPSPS, 5-enolpyruvylshikimate 3-phosphate synthase; FPS, farnesyl-diphosphatesynthase; GibB, gibberellic acid biosynthesis; GPAT, Gln phosphoribosylpyrophosphate amidotransferase; GS, Gln synthetase;GSAT, Glu semialdehyde aminotransferase; IGPD, imidazole glycerol phosphate dehydratase; IMDH, 3-isopropylmalate de-hydrogenase; KARI, ketol acid reductoisomerase; LyCyc, lycopene cyclase; Microtubule assembly, inhibitors of microtubuleassembly; Microtubule organization, inhibitors of microtubule organization; ObtDM, obtusifoliol demethylase; PDS, phytoenedesaturase; PPO, protoporphyrinogen IX oxidase; PyrDH, pyruvate dehydrogenase; TrpS, Trp synthase; VLCFA, very-long-chainfatty acid biosynthesis; ZDS, z-carotene desaturase.
Figure 5. Herbicide market share in 2010 according to MoA. Forabbreviations, see Figure 4.
Plant Physiol. Vol. 166, 2014 1125
Herbicides State of the Art: Overview
www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

or even millions of compounds within a short timeperiod. Most agrochemical companies also followedthis strategy, especially after the full genome of Arabi-dopsis was published in 2000. Henceforward it becamepossible to validate putative herbicidal targets by ge-netic technologies on a large scale. The analysis of thephenotype of a plant in which a certain gene had beenknocked out completely or the expression had beenreduced to a significant extent by antisense RNA en-abled the identification of so-called lethal targets. Thesetargets, mainly enzymes, were subsequently subjectedto high-throughput in vitro screening of chemical li-braries to identify inhibitors. However, the transmissionof the in vitro activity of compounds discovered thisway into the greenhouse turned out to be an insur-mountable hurdle. Therefore, not only have no herbi-cides with a novel MoA been discovered but also, to theauthors’ knowledge, no compound with broad herbi-cidal activity at low application rates has been identifiedin this way.
In the past few years, the focus of herbicide researchhas once again shifted back toward random screeningof synthetic compounds in the greenhouse and to theuse of hits as starting points for targeted optimizationprocesses. If a phytotoxic compound with interestingbioefficacy is identified, studies are undertaken to findthe underlying MoA as soon as possible and to use thisMoA information in rational chemical design approaches.Technological advances in molecular biology, such asgene expression profiling (transcriptomics), have alsobeen adopted for this task (Eckes and Busch, 2007).When a plant is treated with an herbicide, vital pro-cesses of that plant are affected. This is reflected bydistinctive, MoA-dependent changes in the transcriptome.By comparing the transcriptome of a plant treated with aphytotoxic compound from the research pipeline with alibrary of response profiles to compounds with knownMoAs, it may be possible to classify the compound intoone of the known MoAs. If the compound cannot beclassified into an already existing MoA, it can be assumedthat it has a new MoA, for which the profile might alsoprovide clues.
The lack of herbicide innovation has triggered novelattempts to fight herbicide-resistant weed species.Monsanto is investigating the use of topically appliedRNA molecules to induce a process called RNA in-terference in glyphosate-resistant weeds in order tocounteract resistance to glyphosate. It remains to beseen if this approach will be successful. Countries thatdo not accept genetically modified crops must relysolely on the chemical industry to deliver novel herbi-cides with novel MoAs. To achieve this goal, previouslydiscovered but not commercialized targets might bereconsidered in the light of new technological advancesin drug discovery in the last 20 years, such as fragment(ligand)-based drug design. Alternatively, hits from green-house screening might be subjected to MoA elucidationusing state-of-the-art omics technologies (Grossmann et al.,2012; Tresch, 2013) in order to identify novel startingpoints for herbicide discovery.
HERBICIDE SELECTIVITY VIA SAFENERS
For chemical weed control in fields of crops, theherbicide products that can be used must fulfill twocontradictory objectives: control the weed plants but notinjure the crop plants. Some herbicides provide thesefeatures innately (e.g. atrazine for weed control in corn).However, as a general rule, when herbicides are highlyactive against a wider range of weeds, the chances aremuch lower that they will also be highly crop selective.Crop sensitivity is itself a complex issue being influ-enced by many features such as crop variety, applica-tion timing, soil properties, or weather conditions.Therefore, some herbicides may be selective in somecases but not in others. Other herbicides may be selec-tive in certain crops but not in others. For this reason,methods have been developed to increase crop selec-tivity. The two key selectivity technologies are herbicidetolerance traits (either from mutant selection or geneticmodification) and safeners. Safeners (sometimes calledantidotes or protectants) are chemicals that preventherbicidal injury to crop plants without reducing weedcontrol.
Safener Commercialization
Based on an accidental observation in 1947 by OttoHoffmann of the Gulf Oil Company, the concept thatcertain chemicals could increase the tolerance of plantsto herbicides was born (Hoffmann, 1953). Hoffmannhad seen that tomato (Solanum lycopersicum) plantstreated with 2,4,6-trichlorophenoxyacetic acid appearedto be protected from 2,4-dichlorophenoxyacetic acidvapor injury. Whether this was the first time an herbi-cide researcher had seen such effects is not known.However, in this case, Hoffmann realized that this mayprovide a useful tool to increase crop tolerance to her-bicides. Gulf Oil initiated a research program thateventually led to the first commercial compound, 1,8-naphthalic anhydride (NA). When applied as a seedtreatment to corn, this compound gave protection againstvarious preemergence- and preplant-incorporated her-bicides from the thiocarbamate class (e.g. ethyl dipro-pylthiocarbamate). Subsequent research in variousagrochemical companies has subsequently providednearly 20 commercial safeners (Hatzios and Hoagland,1989; Davies and Caseley, 1999; Davies, 2001; Rosingeret al., 2012; Jablonkai, 2013).
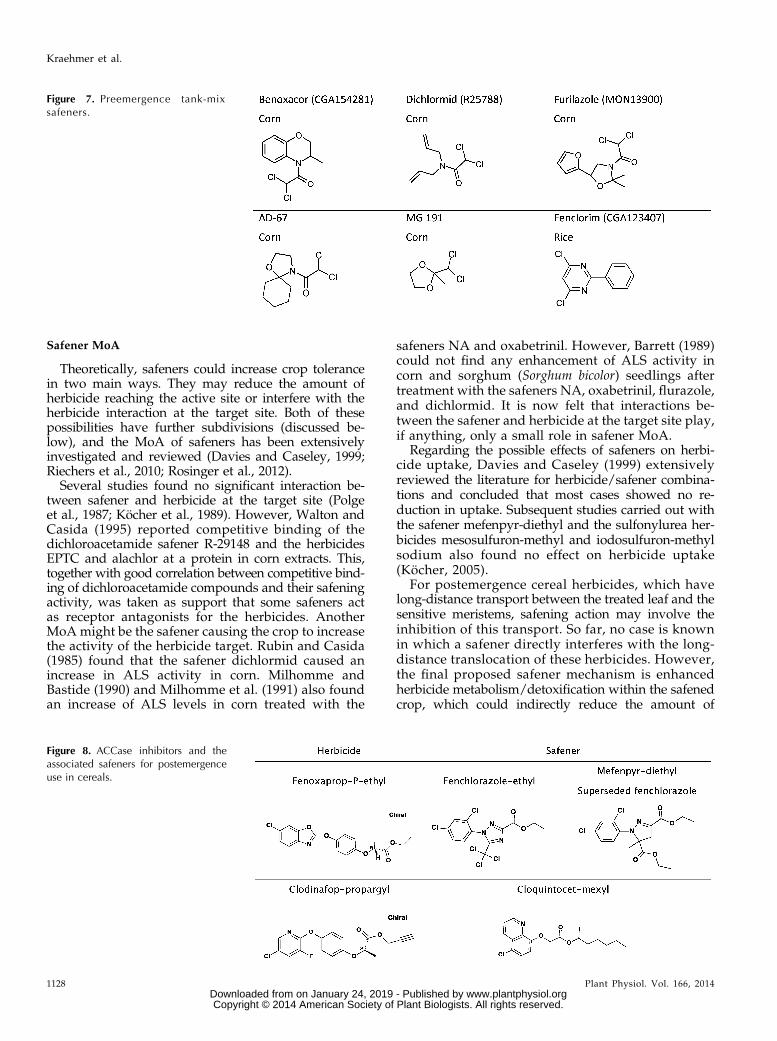
The early phase of safener commercialization duringthe 1970s and early 1980s was dominated by seedtreatment or soil-active (preemergence) safeners (Figs.6 and 7). Seed treatment-applied safeners offer certainadvantages and disadvantages. A key advantage is thatthe safening only influences the crop plant and not theweeds. Therefore, the safener does not need to be cropspecific per se. However, a key disadvantage is that thetreated crop may be sprayed subsequently with herbi-cides from other competitor companies, making valuecapture difficult. Conversely, preemergence tank-mix orcoformulated safeners must be innately crop specific(i.e. with no antagonism on weeds) but offer significantly
1126 Plant Physiol. Vol. 166, 2014
Kraehmer et al.
www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

simplified safener technology for the farmer and manu-facturer. The product could be used just as if the herbi-cide itself were selective. The manufacturer has bettercontrol of the performance of the product containingits herbicides.In the late 1980s, a significant innovation leap oc-
curred whereby “leaf-active” safeners were developedfor coformulation with postemergence herbicides.Fenoxaprop-ethyl was launched in 1984. This moleculecontrolled grasses with postemergence selectively inbroad-leafed crops, but it lacked sufficient selectivity incereals, being moderately damaging to wheat (Triticumaestivum) and more severely damaging to barley (Hor-deum vulgare). Therefore, a research project with the aimof a safener for postemergence use with fenoxaprop-ethyl in cereals was started in the 1980s. A new classof substituted triazole safeners was found from whichfenchlorazole-ethyl was developed and introduced in1992 as a postemergence coformulation product withfenoxaprop-ethyl (Bieringer et al., 1989). This productmeant that fenoxaprop-ethyl could now be used forgrass weed control in wheat and rye (Secale cereale).However, barley was not sufficiently safened byfenchlorazole-ethyl, so testing for further improvedsafeners was continued. Relatively quickly, a new classof safeners was identified with a modified centralchemical scaffold (pyrazoline) from which mefenpyr-diethyl was developed (Hacker et al., 2000). This com-pound had a much stronger safening activity in cerealsthan fenchlorazole-ethyl, making it possible to also usefenoxaprop-ethyl in barley. Among other ACCase in-hibitors was clodinafop-propargyl, which was launchedin 1991. Like fenoxaprop, clodinafop had strong grassweed activity but lacked sufficient crop tolerance incereals. Ciba-Geigy did not launch clodinafop withouta safener. They used the highly effective safenercloquintocet-mexyl, which has a quite different (quino-line) scaffold from fenchlorazole or mefenpyr (Amreinet al., 1989). The structures of the above-mentionedACCase inhibitors and safeners are shown in Figure 8.The safener innovation required to overcome the ce-
real injury meant that by the end of the 1990s, mefenpyr-diethyl and cloquintocet-mexyl were well established in
the cereal market and the two safeners could be testedwith new research compounds. At the end of the 1990s,iodosulfuron and mesosulfuron were introduced for usein cereals (Hacker et al., 1999, 2001). Both had selectivitylimitations that could be overcome by using mefenpyr-diethyl. Cloquintocet-mexyl was also used as a safenerfor pinoxaden, an herbicide from the novel aryl-1,3-dione class of ACCase inhibitors (Hofer et al., 2006).More recently, the safening of mefenpyr-diethyl incereals was used as the selectivity technology for theHPPD inhibitor pyrasulfotole (Schmitt et al., 2008) andthe ALS inhibitor thiencarbazone. Since the basic pat-ents for mefenpyr and cloquintocet have now expired,other companies have used them in their own safenedproducts. Of particular note is a coformulation of theALS inhibitor pyroxsulam with cloquintocet as a safener(Wells, 2008).
During the 1990s and 2000s, safener research wascontinued. One target was the extension of the fenox-aprop market into rice (Oryza sativa), which was nottolerant to this herbicide. Screening efforts identified thesafener isoxadifen-ethyl (Fig. 9), which again retainedthe carboxylic acid ester of fenchlorazole-ethyl andmefenpyr-diethyl, but with a new heterocyclic isoxazolecore. Although fenoxaprop in rice was the originaltarget, early on in the research and development phaseit was found that isoxadifen-ethyl could also safenpostemergence against the sulfonylurea herbicideethoxysulfuron in rice and foramsulfuron and iodo-sulfuron in corn (Collins et al., 2001; Pallett et al., 2001;Hacker et al., 2002). Therefore, isoxadifen now repre-sents the current pinnacle of multicrop and multi-herbicide safening.
The most recently launched safener is cyprosulfa-mide (Fig. 9). Like isoxadifen-ethyl, it is strongly activepostemergence in corn but has the significant advan-tage, for certain herbicides, of also having strong pre-emergence safening activity. Cyprosulfamide comesfrom a class of safeners (acyl sulfonamides) discoveredduring the mid-1990s. It is now used to support asignificant number of different corn herbicides (Phil-brook and Santel, 2008; Santel and Philbrook, 2008;Watteyne et al., 2009).
Figure 6. Seed treatment safeners.
Plant Physiol. Vol. 166, 2014 1127
Herbicides State of the Art: Overview
www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.
Safener MoA
Theoretically, safeners could increase crop tolerancein two main ways. They may reduce the amount ofherbicide reaching the active site or interfere with theherbicide interaction at the target site. Both of thesepossibilities have further subdivisions (discussed be-low), and the MoA of safeners has been extensivelyinvestigated and reviewed (Davies and Caseley, 1999;Riechers et al., 2010; Rosinger et al., 2012).
Several studies found no significant interaction be-tween safener and herbicide at the target site (Polgeet al., 1987; Köcher et al., 1989). However, Walton andCasida (1995) reported competitive binding of thedichloroacetamide safener R-29148 and the herbicidesEPTC and alachlor at a protein in corn extracts. This,together with good correlation between competitive bind-ing of dichloroacetamide compounds and their safeningactivity, was taken as support that some safeners actas receptor antagonists for the herbicides. AnotherMoA might be the safener causing the crop to increasethe activity of the herbicide target. Rubin and Casida(1985) found that the safener dichlormid caused anincrease in ALS activity in corn. Milhomme andBastide (1990) and Milhomme et al. (1991) also foundan increase of ALS levels in corn treated with the
safeners NA and oxabetrinil. However, Barrett (1989)could not find any enhancement of ALS activity incorn and sorghum (Sorghum bicolor) seedlings aftertreatment with the safeners NA, oxabetrinil, flurazole,and dichlormid. It is now felt that interactions be-tween the safener and herbicide at the target site play,if anything, only a small role in safener MoA.
Regarding the possible effects of safeners on herbi-cide uptake, Davies and Caseley (1999) extensivelyreviewed the literature for herbicide/safener combina-tions and concluded that most cases showed no re-duction in uptake. Subsequent studies carried out withthe safener mefenpyr-diethyl and the sulfonylurea her-bicides mesosulfuron-methyl and iodosulfuron-methylsodium also found no effect on herbicide uptake(Köcher, 2005).
For postemergence cereal herbicides, which havelong-distance transport between the treated leaf and thesensitive meristems, safening action may involve theinhibition of this transport. So far, no case is knownin which a safener directly interferes with the long-distance translocation of these herbicides. However,the final proposed safener mechanism is enhancedherbicide metabolism/detoxification within the safenedcrop, which could indirectly reduce the amount of
Figure 7. Preemergence tank-mixsafeners.
Figure 8. ACCase inhibitors and theassociated safeners for postemergenceuse in cereals.
1128 Plant Physiol. Vol. 166, 2014
Kraehmer et al.
www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

active herbicide reaching the target site. It is well un-derstood that differences in herbicide metabolism playan important role in herbicide selectivity (Hatzios andPenner, 1982; Drobny et al., 2012). Therefore, it is notsurprising that this was proposed as a possible safenermechanism as early as the 1970s and has been studiedextensively since that time. Hatzios and Hoagland (1989)reviewed in detail the early studies, especially on theseed treatment safeners and preemergence safeners. Thisshowed a large amount of evidence that safeners wereindeed increasing the rate of herbicide metabolism in thesafened crop plant. For more recent postemergencesafeners, metabolism studies combined with gene ex-pression analysis has strengthened the understand-ing of safener MoA. In wheat (but not the targetgrass weed), the level of fenoxaprop-ethyl and thefree acid fenoxaprop declined (conversion to inactivemetabolites) more rapidly when plants were treated withfenchlorazole-ethyl (Köcher et al., 1989; Yaacoby et al.,1991). For the safener cloquintocet-mexyl, the safening ofwheat against clodinafop-propargyl was also reported tobe based on enhanced detoxification (Kreuz et al., 1991;Kreuz, 1993). Mefenpyr-diethyl also enhanced the rateof conversion of fenoxaprop to nonphytotoxic productsin wheat but not in target weeds (Hacker et al., 2000).Similarly, mefenpyr-diethyl enhanced the rate of meta-bolic degradation of iodosulfuron-methyl sodium andmesosulfuron-methyl in cereals but not in the grassweeds tested (Hacker et al., 1999, 2000, 2001). In corn, thepostemergence safener isoxadifen-ethyl has also beenshown to enhance the metabolism of foramsulfuron(Pallett et al., 2001; Hacker et al., 2002).Herbicide metabolism in plants involves several
enzyme-mediated steps (Hatzios, 1991; Van Eerd et al.,2003). The first step may sometimes be a conversion ofan inactive proherbicide to the active herbicide. How-ever, the metabolic steps relevant to safening are thosethat convert the active herbicide to inactive metabolites.Cytochrome P450 enzymes may catalyze the reduction,oxidation, or hydroxylation of the herbicide and thusprovide functional groups for further metabolic steps.These often involve conjugation reactions between thefirst metabolite and glutathione, Glc, or amino acids thatare catalyzed by multifunctional enzyme, glutathioneS-transferases, or glycosyltransferases (Lamoureux et al.,1991; Armstrong, 1994; Marrs, 1996; Hatzios, 2001). Ad-ditionally, conjugation with malonate via malonyl-CoAhas also been reported (Sandermann et al., 1997). The
next step in herbicide detoxification is the transport ofthe conjugates into the vacuole, which may be catalyzedby ATP-binding cassette transporters (Tommasini et al.,1997). Once in the vacuole, the conjugates may be furtherdegraded by peptidases with the removal of Gly andGlu. The speed of herbicide metabolism/detoxification,therefore, may depend on the level of activity of severalkey enzymes and, thus, the level of expression of genesthat code for these enzymes. Advances in biotechnologyover the past two decades have allowed increasinglydetailed studies into the effect of safeners on geneexpression. It has been shown that genes encoding forenzymes involved in herbicide detoxification are in-duced within a few hours of safener application(Kreuz et al., 1996; Cummins et al., 1997; DeRidderet al., 2002; Scalla and Roulet, 2002; Theodoulou et al.,2003; Zhang et al., 2007).
Gene expression studies have indicated parallels be-tween the oxidative stress-related oxylipin pathway andsafener signaling (Riechers et al., 2010). They have alsoindicated possible overlaps with the plant stress defensesignaling pathway involving salicylic acid. For example,it could be shown that many safener-regulated genesare induced by salicylic acid (Behringer et al., 2011).Therefore, while the primary site of safener reception isstill not known, it seems that several signaling path-ways contribute to the full safener response in plants.
FUTURE PERSPECTIVES
There have been very few new safener structurespatented in the past decade, suggesting that little re-search into new compounds is being conducted in theagrochemical industry. This is probably mainly due tothe emergence of genetically modified herbicide toler-ance as a major selectivity tool (e.g. glyphosate-tolerantRound-Up Ready cotton [Gossypium hirsutum], soybean,and corn) as well as the availability of strong existingsafeners for use with newer herbicides. However, as theproblem of weed resistance increases and the existingsafeners reach the limit of their potential, stronger next-generation safeners may indeed be required for futureselective weed control products. As biotechnologymethods develop quickly, it is possible that the targetsite(s) of safeners will be identified in the not too distantfuture. It may then perhaps be possible to use rationaldesign to identify new safener structures.
Figure 9. Structures and use of iso-xadifen-ethyl and cyprosulfamide.
Plant Physiol. Vol. 166, 2014 1129
Herbicides State of the Art: Overview
www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

ACKNOWLEDGMENTS
We thank Dr. Stephen Lindell for critically reading the article.
Received April 26, 2014; accepted August 3, 2014; published August 7, 2014.
LITERATURE CITED
Amrein J, Nyffeler A, Rufener J (1989) CGA-184927 + S: a new post-emergence grasskiller for use in cereals. In Proceedings of the BrightonCrop Protection Conference: Weeds, Vol 1. British Crop ProtectionCouncil, Farnham, UK, pp 71–76
Armstrong RN (1994) Glutathione S-transferases: structure and mechanismof an archetypical detoxication enzyme. Adv Enzymol Relat Areas MolBiol 69: 1–44
Arndt F, Kötter C (1968) Zur Selektivität von Phenmedipham alsNachauflaufherbizid in Beta-Rüben. Weed Res 8: 259–271
Barrett M (1989) Reduction of imazaquin injury to corn (Zea mays) andsorghum (Sorghum bicolor) with antidotes. Weed Sci 37: 34–41
Behringer C, Bartsch K, Schaller A (2011) Safeners recruit multiple sig-nalling pathways for the orchestrated induction of the cellular xenobi-otic detoxification machinery in Arabidopsis. Plant Cell Environ 34:1970–1985
Bieringer H, Bauer K, Hacker E, Heubach G, Leist K, Ebert E (1989) Hoe70542: a new molecule for use in combination with fenoxaprop-ethylallowing selective post-emergence grass weed control in wheat. InProceedings of the Brighton Crop Protection Conference: Weeds, Vol 1.British Crop Protection Council, Farnham, UK, pp 77–82
Büchel KH, editor (1977) Pflanzenschutz und Schädlingsbekämpfung.Georg Thieme Verlag, Stuttgart, Germany
Collins B, Drexler D, Merkl M, Hacker E, Hagemeister H, Pallett K, Effertz C(2001) Foramsulfuron: a new foliar herbicide for weed control in corn. InProceedings of the Brighton Crop Protection Conference: Weeds, Vol 1.British Crop Protection Council, Farnham, UK, pp 35–42.
Cremlyn RJ (1991) Agrochemicals. John Wiley & Sons, Chichester, UKCummins I, Cole DJ, Edwards R (1997) Purification of multiple glutathione
transferases involved in herbicide detoxification from wheat (Triticumaestivum L.) treated with the safener fenchlorazole-ethyl. Pestic BiochemPhysiol 59: 35–49
Davies J (2001) Herbicide safeners: commercial products and tools foragrochemical research. Pestic Outlook 12: 10–15
Davies J, Caseley JC (1999) Herbicide safeners: a review. Pestic Sci 55:1043–1058
Dayan FE, Duke SO, Grossmann K (2010) Herbicides as probes in plantbiology. Weed Sci 58: 340–350
DeRidder BP, Dixon DP, Beussman DJ, Edwards R, Goldsbrough PB(2002) Induction of glutathione S-transferases in Arabidopsis by herbi-cide safeners. Plant Physiol 130: 1497–1505
Drobny H, Schulte M, Strek H (2012) 25 years of sulfonylurea herbicides: afew grams changed the world of chemical weed control. Julius-Kuhn-Archiv434: 21–33
Duke SO (2012) Why have no new herbicide modes of action appeared inrecent years? Pest Manag Sci 68: 505–512
Eckes P, Busch M (2007) Fast identification of the mode of action of herb-icides by DNA chips. In W Krämer, U Schirmer, eds, Modern Crop Pro-tection Compounds, Vol 3. Wiley-VCH Verlag, Weinheim, Germany, pp1161–1174
Erdkamp P (2005) The Grain Market in the Roman Empire. CambridgeUniversity Press, Cambridge, UK
Gerwick BC (2010) Thirty years of herbicide discovery: surveying the pastand contemplating the future. Agrow VII–IX
Gressel J, Shaaltiel Y, Sharon A, Amsellem Z (1993) Biorational in vitroscreening for herbicide synergists. In JC Streibig, P Kudsk, eds, Herbi-cide Bioassay. CRC Press, Boca Raton, FL, pp 217–252
Grossmann K, Christiansen N, Looser R, Tresch S, Hutzler J, Pollmann S,Ehrhardt T (2012) Physionomics and metabolomics: two key approachesin herbicidal mode of action discovery. Pest Manag Sci 68: 494–504
Hacker E, Bieringer H, Willms L, Lorenz K, Köcher H, Huff H, Borrod G,Brusche R (2001) Mesosulfuron-methyl: a new active ingredient forgrass weed control in cereals. In Proceedings of the Brighton CropProtection Conference: Weeds, Vol 1. British Crop Protection Council,Farnham, UK, pp 43–48
Hacker E, Bieringer H, Willms L, Ort O, Köcher H, Kehne H (1999)Iodosulfuron plus mefenpyr-diethyl: an new foliar herbicide for weedcontrol in cereals In Proceedings of the Brighton Crop Protection Con-ference: Weeds, Vol 1. British Crop Protection Council, Farnham, UK, pp15–22
Hacker E, Bieringer H, Willms L, Rösch W, Köcher H, Wolf R (2000)Mefenpyr-diethyl: a safener for fenoxaprop-P-ethyl and iodosulfuron incereals. Z Pflanz Pflanzenschutz Sonderheft XVII: 493–500
Hacker E, Bieringer H, Willms L, Schnabel G, Köcher H, Hagemeister H,Steinheuer W (2002) Foramsulfuron plus safener: a new technology forweed control in maize. Z Pflanz Pflanzenschutz Sonderheft XVIII: 747–756
Håkanson S (2003) Weeds and Weed Management on Arable Land: AnEcological Approach. CABI Publishing, Wallingford, UK
Hatzios KK (1991) Biotransformations of herbicides in higher plants. In RGrover, AJ Cessna, eds, Environmental Chemistry of Herbicides. CRCPress, Boca Raton, FL, pp 141–185
Hatzios KK (2001) Functions and regulation of plant glutathione-S-transferases. In JC Hall, RE Hoagland, RM Zablotowicz, eds, PesticideBiotransformation in Plants and Microorganisms: Similarities and Di-vergences. American Chemical Society, Washington, DC, pp 218–239
Hatzios KK, Hoagland RE (1989) Crop Safeners for Herbicides. AcademicPress, San Diego
Hatzios KK, Penner D (1982) Metabolism of Herbicides in Higher Plants.Burgess, Minneapolis
Hofer U, Mühlebach UM, Hole S, Zoschke A (2006) Pinoxaden: for broadspectrum grass weed management in cereal crops. J Plant Dis Prot XX:989–995
Hoffmann OL (1953) Inhibition of auxin effects by 2,4,6-trichlorophenoxyaceticacid. Plant Physiol 28: 622–628
Jablonkai I (2013) Herbicide safeners: effective tools to improve herbicideselectivity. In J Price, JA Kelton, eds, Herbicides: Current Research andCase Studies in Use. Intech http://dx.doi.org/10.5772/55168 (August17, 2014)
Köcher H (2005) Mesosulfuron-methyl and combination partner iodosulfuron-methyl sodium: mode of herbicidal action. Pflanzenschutz-Nachr Bayer 58:179–194
Köcher H, Büttner B, Schmidt E, Lötzsch K, Schulz A (1989) Influence ofHOE 70542 on the behavior of fenoxaprop-ethyl in wheat. In Proceed-ings of the Brighton Crop Protection Conference: Weeds, Vol 1. BritishCrop Protection Council, Farnham, UK, pp 495–500
Kraehmer H (2010) Weed surveys and weed mapping in Europe. In LBastiaans, C Bohren, S Christensen, B Gerowitt, P Hatcher, H Krähmer,P Kudsk, B Melander, E Pannacci, B Rubin, J Streibig, F Tei, AThompson, K Torresesn, M Vurro, eds, Proceedings of the 15th EWRSSymposium in Kaposvar. Asszisztencia Congress Bureau Ltd., Buda-pest, p 128
Kraehmer H (2012) Changing trends in herbicide discovery. Outlook onPest Management 23: 115–118
Kraehmer H, Baur P (2013) Weed Anatomy. Wiley-Blackwell, Chichester,UK
Kraehmer H, Drexler D (2009) Global herbicide development: opportuni-ties and constraints. Prairie Soils & Crops Journal 2: 12–16
Kraehmer H, Russell PE (1994) General problems in glasshouse to fieldtransfer of pesticide performance. BCPC Monograph 59: 3–16
Kraehmer H, Schulz A, Laber B (2007) Where are the new herbicides modeof action? FarmTech 2007 Proceedings 88–97 http://farmtechconference.com/wp-content/uploads/2013/01/FarmTech2007-complete-proceedings.pdf(Oct 2, 2014)
Kraehmer H, Stuebler H (2012) Technical demands and political restric-tions for weed control. Julius-Kuhn-Archiv 434: 15–19
Kraehmer H, van Almsick A, Beffa R, Dietrich H, Eckes P, Hacker E, HainR, Strek HJ, Stuebler H, Willms L (2014) Herbicides as weed controlagents: state of the art. II. Recent achievements. Plant Physiol 166: 1132–1148
Krämer W, Schirmer U, Jeschke P, Witschel M, editors (2012) ModernCrop Protection Compounds, Ed 2, Vol 1. Wiley-VCH, Weinheim,Germany
Kreuz K (1993) Herbicide safeners: recent advances and biochemical as-pects of their mode of action In Proceedings of the Brighton Crop Pro-tection Conference: Weeds, Vol 3. British Crop Protection Council,Farnham, UK, pp 1249–1258
1130 Plant Physiol. Vol. 166, 2014
Kraehmer et al.
www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

Kreuz K, Gaudin J, Stingelin J, Ebert E (1991) Metabolism of thearylphenoxypropanoate herbicide CGA 184927 in wheat, barley andmaize: different effects of the safener, CGA 185072. Z Naturforsch 46c:901–905
Kreuz K, Tommasini R, Martinoia E (1996) Old enzymes for a new job(herbicide detoxification in plants). Plant Physiol 111: 349–353
Lamoureux GL, Shimabukuro RH, Frear DS (1991) Glutathione and glu-coside conjugation in herbicide selectivity. In JC Caseley, GW Cussans,RK Atkin, eds, Herbicide Resistance in Weeds and Crops. Butterworth-Heinemann, Oxford, pp 227–261
Leeson JY, Gordon Thomas A, Hall LM, Brenzil CA, Andrews T, BrownKR, Van Acker KR, editors (2005) Prairie Weed Survey: Cereal, Oilseedand Pulse Crops 1970s to 2000s. Agriculture and Agri-Food Canada,Saskatoon, Canada
Liu W, Ye J, Jin M (2009) Enantioselective phytoeffects of chiral pesticides.J Agric Food Chem 57: 2087–2095
Marrs KA (1996) The functions and regulation of glutathione S-transferasesin plants. Annu Rev Plant Physiol Plant Mol Biol 47: 127–158
Milhomme H, Bastide J (1990) Uptake and phytotoxicity of the herbicidemetsulfuron methyl in corn root tissue in the presence of the safener 1,8-naphthalic anhydride. Plant Physiol 93: 730–738
Milhomme H, Roux C, Bastide J (1991) Safeners as corn seedling protec-tants against acetolactate synthase inhibitors. Z Naturforsch 46c: 945–949
Montgomery DR (2007) Soil erosion and agricultural sustainability. PNAS104: 13268–13272
Pallett KE, Veerasekaran P, Crudace M, Köcher H, Collins B (2001)Foramsulfuron and isoxadifen-ethyl: absorption, translocation and me-tabolism. In Proceedings of the NCWSS Conference 56. North CentralWeed Science Society, Champaign, MI, p 77
Philbrook B, Santel HJ (2008) A new formulation of isoxaflutole for pre-emergence weed control in corn (Zea mays). Weed Sci Soc Am Abstr 48:116
Polge ND, Dodge AD, Caseley JC (1987) Biochemical aspects of safeneraction: effects on glutathione, glutathione-S-transferase and acetohydroxyacid synthetase in maize. In Proceedings of the Brighton Crop ProtectionConference: Weeds, Vol 3. British Crop Protection Council, Farnham, UK,pp 1113–1120
Riechers DE, Kreuz K, Zhang Q (2010) Detoxification without intoxication:herbicide safeners activate plant defense gene expression. Plant Physiol153: 3–13
Rosinger C, Bartsch K, Schulte W (2012) Safeners for herbicides. In WKrämer, U Schirmer, P Jeschke, M Witschel, eds, Modern Crop Protec-tion Compounds. Wiley-VCH, Weinheim, Germany, pp 371–398
Rubin B, Casida J (1985) R-25788 effects on chlorosulfuron injury andacetohydroxy acid synthesase activity. Weed Sci 33: 462–468
Rüegg WT, Quadranti M, Zoschke A (2007) Herbicide research and de-velopment: challenges and opportunities. Weed Res 47: 271–275
Sandermann H, Haas M, Messner B, Pflumacher S, Schroder P, Wetzel A(1997) The role of glucosyl and malonyl conjugation in herbicide selec-tivity. In KK Hatzios, ed, Regulation of Enzymatic Systems DetoxifyingXenobiotics in Plants. Kluwer Academic Publishers, Dordrecht, TheNetherlands, pp 211–231
Santel H, Philbrook B (2008) Thiencarbazone-methyl and isoxaflutole: anew herbicide premixture for preemergence weed control in corn (Zeamays). Weed Sci Soc Am Abstr 48: 117
Scalla R, Roulet A (2002) Cloning and characterization of a glutathioneS-transferase induced by a herbicide safener in barley (Hordeum vulgare).Physiol Plant 116: 336–344
Schmitt MH, van Almsick A, Willms L (2008) Discovery and chemistry ofpyrasulfotole, a new dicot herbicide for cereal production.Pflanzenschutz-Nachr Bayer 61: 7–14
Seitz T, Hoffmann M, Krähmer H (2003) Chemical plant protection: her-bicides in agriculture. Chem Unserer Zeit 34: 112–126
Stuebler H, Kraehmer H, Hess M, Schulz A, Rosinger C (2008) Globalchanges in crop production and impact trends on innovation in weedmanagement: an industry view. In Proceedings of the 5th InternationalWeed Science Congress, Vancouver, Canada, pp 309–319
Theodoulou FL, Clark IM, He XL, Pallett KE, Cole DJ, Hallahan DL (2003)Co-induction of glutathione-S-transferases and multidrug resistanceassociated protein by xenobiotics in wheat. Pest Manag Sci 59: 202–214
Tommasini R, Vogt E, Schmid J, Fromentau M, Amrhein N, Martinoia E(1997) Differential expression of genes coding for ABC transporters aftertreatment of Arabidopsis thaliana with xenobiotics. FEBS Lett 411: 206–210
Tresch S (2013) Strategies and future trends to identify the mode of actionof phytotoxic compounds. Plant Sci 212: 60–71
Troyer JR (2001) In the beginning: the multiple discovery of the first hor-mone herbicides. Weed Sci 49: 290–297
Van Eerd L, Hoagland RE, Zablotowicz RM, Hall JC (2003) Pesticidemetabolism in plants and microorganisms. Weed Sci 51: 472–495
Voss M, Renger G, Gräber P (1984a) Measurements of penetration anddetoxification of PSII herbicides in whole leaves by a fluorometricmethod. Z Naturforsch C 39: 359–361
Voss M, Renger G, Kötter C, Gräber P (1984b) Fluorometric detection ofphotosystem II herbicide penetration and detoxification in whole leaves.Weed Sci 32: 675–680
Walton JD, Casida JE (1995) Specific binding of a dichloroacetamide her-bicide safener in maize at a site that also binds thiocarbamate andchloroacetanilide herbicides. Plant Physiol 109: 213–219
Watteyne K, Castillo T, Hinz J, Philbrook B, Bloomberg J (2009) Isoxaflutole +cyprosulfamide, thiencarbazone + isoxaflutole + cyrosulfamide: performancein university corn trials. In Proceedings of the NCWSS Conference 64. NorthCentral Weed Science Society, Kansas City, MO, p 119
Wells G (2008) Pyroxsulam for broad-spectrum weed control in wheat.In RD Van Klinken, VA Osten, FD Panetta, JC Scanlan, eds, 16th Aus-tralian Weeds Conference Proceedings: Weed Management 2008 HotTopics in the Tropics. Queensland Weeds Society, Brisbane, Australia,pp 297-299
Worthington CR, Hance RJ (1991) The Pesticide Manual, Ed 9. British CropProtection Council, Farnham, UK,
Yaacoby T, Hall J, Stephenson G (1991) Influence of fenchlorazole-ethyl onthe metabolism of fenoxaprop-ethyl in wheat barley and crabgrass.Pestic Biochem Physiol 41: 296–304
Zhang Q, Xu F, Lambert KN, Riechers DE (2007) Safeners coordinatelyinduce the expression of multiple proteins and MRP transcripts involvedin herbicide metabolism and detoxification in Triticum tauschii seedlingtissues. Proteomics 7: 1261–1278
Zimdahl R (2004) Weed-Crop Competition: A Review. Blackwell Publish-ing, Oxford
Zohary D, Hopf M, Weiss E (2012) Domestication of Plants in the OldWorld, Ed 4. Oxford University Press, Oxford
Plant Physiol. Vol. 166, 2014 1131
Herbicides State of the Art: Overview
www.plantphysiol.orgon January 24, 2019 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.





























