Embed Size (px)
Citation preview

Vol. 56, No. 11
Effects of Hg2", CH3-Hg', and Temperature on the Expression ofMercury Resistance Genes in Environmental Bacteria
YU-LI TSAI AND BETTY H. OLSON*
Program in Social Ecology, University of California, Irvine, California 92717
Received 29 May 1990/Accepted 14 August 1990
Twenty different bacterial isolates obtained from a mercury-contaminated site in Oak Ridge, Tenn., weregrown on plate count agar amended with 25 ,ug of Hg2+ or 3 ,ug of CH3-Hg' (R-Hg+) per ml. The total cellularRNA was extracted from each isolate by an acid-guanidine-thiocyanate-phenol-chloroform method. Thetranscripts of merA and merB were detected and quantitated by Northern (RNA) hybridization. A qualitativeassay of mercuric reductase was used to confirm the enzyme activity. Low temperature (4°C) with the presenceof Hg2+ (25 ,ug/ml) significantly increased the net merA transcripts of mid-log-phase cells of six environmentalisolates. The net merA transcript production by 18 of the isolates increased when they were grown on 50% platecount broth with 15 ,ug of Hg2+ per ml, but only 8 isolates showed increased production of merB transcripts.The MICs of Hg2' and R-Hg+ for 10 methyl mercury-resistant isolates ranged from 45 to 110 ,ug of Hg2' and0.6 to 4.5 ,ug of R-Hg+ per ml. R-Hg+ was able to induce the expression of merB in 70% of methyl mercury-resistant strains.
Mercury has received considerable attention because ofits high toxicity and has become an excellent model chemicalfor the study of biotransformation of metal salts. The com-plete detoxification of mercury involves the reduction of theinorganic (Hg2+) or organomercurial (CH3-Hg' [R-Hg+])form to the less toxic elemental mercury (Hg°) (9, 31).Mercuric reductase, the enzyme encoded by the merA gene,is involved in the conversion of Hg2" to Hg°, and theorganomercurial lyase enzyme encoded by the merB gene isinvolved in the cleavage of H3C-Hg bond to release Hg2"(28, 30). Mercury-resistant bacteria isolated from mercury-polluted sediments have been found to transform organicand inorganic mercury to Hg0 (19, 29). Strong positivecorrelations between mercury concentration and the fre-quency of mercury resistance genes (mer) in bacteria iso-lated from mercury-contaminated sediments have been dem-onstrated by Barkay and Olson (4). In addition, Barkay et al.(3) used mer (Tn2J)-mediated reduction of Hg2+ to show thatmicrobial communities of fresh water volatilized mercury atsignificantly different rates than those of saline water.A number of studies have been instrumental in character-
izing the enzymes mercuric reductase and organomercuriallyase and how they function in intact cells and cell extracts(10, 22, 25, 27, 32, 33). None of these investigations wereconcerned with how these enzymes function in environmen-tal isolates or in the variability within or among genera.Since the main purposes of these studies were to describeenzyme function and kinetics, many were performed withconstructed plasmids and model hosts (10, 22, 27, 33). Thedevelopment of mRNA assay methods for pure culture (7)made it possible to examine larger numbers of organisms fortranscript production under conditions more representativeof the environment. An additional advantage of measuringmRNA is that the technique is rapid and provides greatersensitivity than do standard enzyme assays (7, 15, 35). In thepresent work, we report the first data on the actual function-ing unit (mRNA) of the mer genes from environmental
* Corresponding author.
isolates belonging to the genus Pseudomonas under stressconditions. The two variables examined wtih respect tomRNA production were mercury and temperature.
MATERIALS AND METHODS
Organisms and culture conditions. Bacteria in sedimentsamples from East Fork Poplar Creek, Oak Ridge, Tenn., asite heavily contaminated with mercury, were serially di-luted and isolated on plate count agar (Difco Laboratories,Detroit, Mich.) amended with 25 ,ug of Hg2+ as HgCl2 per mlor with 3 ,ug of R-Hg+ as CH3(Hg) OH per ml. The plateswere incubated at 23°C for 3 days. Ten morphologicallydifferent colonies were randomly chosen from each type ofmercury-amended plates and subcultured in 50% plate countbroth (Difco) with the respective mercury species. All envi-ronmental isolates were identified with an API Rapid NFTkit (API Analytab Products, Plainview, N.Y.).
Isolation of total RNA from environmental isolates. Purecultures (10 ml) of each isolate grown to late exponentialphase were pelleted by centrifugation at 3,000 x g for 10 minat 4°C. The pellets were vigorously mixed with 1.0 ml ofdenaturing solution containing 4 M guanidinium thiocyanate,25 mM sodium citrate, 0.5% sarcosyl, and 0.1 M 2-mercap-toethanol. The resulting cell lysate was extracted with acid-phenol-chloroform as described previously by Chomczynskiand Sacchi (7). The purity of isolated total cellular RNA wasmonitored by measuring A26JA280 ratios (26) and by gelelectrophoresis after RNase and DNase treatments. In addi-tion, the total cellular RNA was quantitated by spectropho-tometry at 260 nm (26). A laboratory strain, Pseudomonasaeruginosa PU21, containing the plasmid Rip64 encodingmercury resistance was used as a positive control for RNAextraction (O. A. Ogunseitan, Y.-L. Tsai, and B. H. Olson,submitted for publication).
Radiolabeling of DNA probes and hybridization conditions.Both mercuric reductase (merA) and organomercurial lyase(merB) mercury resistance genes originating from plasmidpDU1358 were kindly provided by S. Silver and were
3266
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Nov. 1990, p. 3266-32720099-2240/90/113266-07$02.00/0Copyright C) 1990, American Society for Microbiology
on February 7, 2020 by guest
http://aem.asm
.org/D
ownloaded from
EFFECTS OF MERCURY AND TEMPERATURE ON mRNA 3267
subcloned into Escherichia coli DHSx cells. Purified DNAfragments (0.1 ,ug) of merA (1 kilobase) and merB (457 basepairs) were labeled with [at-32P]dCTP by random priming,following the protocol of the manufacturer (Bethesda Re-search Laboratories, Gaithersburg, Md.). Labeled probeswere purified from unincorporated nucleotides with Elutip-dcolumns (Schleicher & Schuell, Keene, N.H.). The specificactivities of the labeled probes were about 108 dpm/jig ofDNA.
Total cellular RNA (5 jLg) from each strain was filteredonto a nitrocellulose membrane (0.2 jim; Schleicher &Schuell), using a multifiltration apparatus (Minifold II Slot-Blotter; Schleicher & Schuell). The membranes were thenwashed with 0.1x SSC (0.15 M NaCl, 0.015 M trisodiumcitrate, pH 7.0) for 1 h at 65°C. For high-stringency hybrid-ization, the filters were first prehybridized in 50% formam-ide-5x SSC-50 mM Na2HPO4-100 jig of salmon spermDNA per ml-5x Denhardt solution for 2 h at 42°C beforehybridization was conducted for 18 to 24 h at 42°C in anidentical solution but with 32P-labeled probe DNA (finalcounts, 3 x 106 cpm/ml). The membranes were washed fourtimes with 2x SSC-0.1% sodium dodecyl sulfate at roomtemperature and then finally twice with 0.1x SSC-0.1%sodium dodecyl sulfate at 50°C. For low-stringency hybrid-ization, the filters were hybridized in 25% formamide hybrid-ization solution and washed only with 2 x SSC-0.1% sodiumdodecyl sulfate four times at room temperature.
Quantitation of mRNA. Autoradiography was used todetect the presence of DNA-mRNA hybridization frommerA or merB. The AMBIS Radioanalytic Imaging System(AMBIS systems, San Diego, Calif.) was used to quantifythe radioactive signals from the hybridized transcripts. Thesignal counts were normalized to total radioactivity countsper 107 cells or per 5 ,ug of total RNA. The value of totalradioactivity counts was obtained from averaged radioactiv-ity counts of internally duplicate RNA samples.
Effects of temperature on merA transcription. One Hg2+-resistant environmental isolate identified as P. stutzeri OR2was subcultured twice in 50% plate count broth at 23°C.Cells (600 ml) at mid-log phase were evenly distributed intotwo sets of three flasks. One set was spiked with 25 jig ofHg2+ per ml and the other set served as the unamendedcontrol. The flasks from each set were incubated individuallyat three different temperatures (4, 23, and 37°C) with shakingfor 45 h. Cells from each experiment were collected at sixtime intervals (3, 5, 7, 9, 20, and 45 h) for merA transcriptquantification and viable cell enumeration. Five other Hg2+-resistant isolates from the same mercury-polluted site werealso tested for the production of merA transcripts at 4 and230C.
Effects of inorganic mercury and methyl mercury. TheMICs of Hg2+ and R-Hg+ for mercury-resistant isolateswere determined in 50% plate count broth amended withdifferent concentrations of tested mercury species. The totalRNA of late exponential-phase isolates grown with 15 pug ofHg2+ or 2.5 ,ug of R-Hg+ per ml was extracted for merA ormerB transcript analyses. Pellets from 10 ml of cells fromeach isolate were washed with equal volumes of phosphatebuffer (pH 7.0) containing 70 mM sodium phosphate, 0.5 mMEDTA, and 0.2 mM magnesium acetate, followed by cen-trifugation at 3,000 x g for 10 min, and suspended in 100 pAlof phosphate buffer for the following mercuric reductase andorganomercurial lyase assays.Mercury volatilization test. Fifty microliters of washed
resting-stage cells was transferred into each well of a micro-dilution plate (Falcon 3872; Becton Dickinson & Co., Ox-
TRC/10E7 Cells
Time (hours)
0 10 20 30 40 50Time (hours)
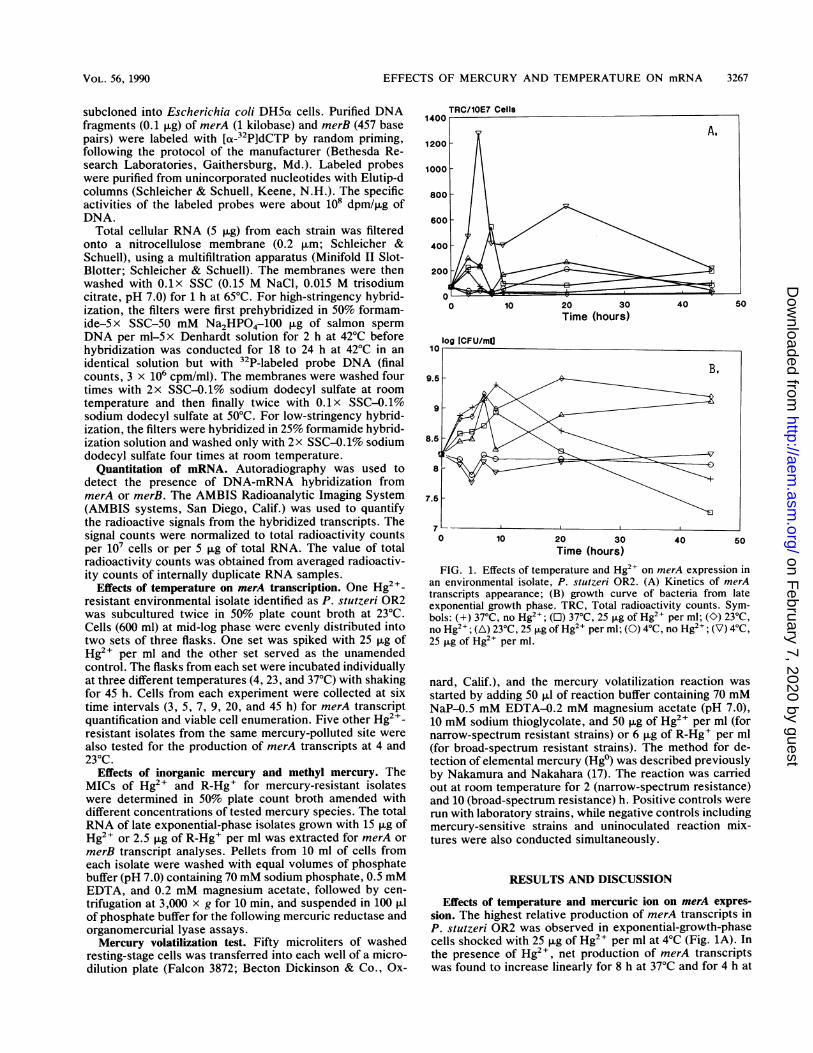
FIG. 1. Effects of temperature and Hg2+ on merA expression inan environmental isolate, P. stutzeri OR2. (A) Kinetics of merAtranscripts appearance; (B) growth curve of bacteria from lateexponential growth phase. TRC, Total radioactivity counts. Sym-bols: (+) 370C, no Hg2+; (L) 37°C, 25 ,ug of Hg2" per ml; (O) 230C,no Hg2+; (A) 23°C, 25 ,ug of Hg2+ per ml; (0) 4°C, no Hg2+; (V) 4°C,25 ,ug of Hg2+ per ml.
nard, Calif.), and the mercury volatilization reaction was
started by adding 50 ,ul of reaction buffer containing 70 mMNaP-0.5 mM EDTA-0.2 mM magnesium acetate (pH 7.0),10 mM sodium thioglycolate, and 50 jig of Hg2+ per ml (fornarrow-spectrum resistant strains) or 6 jig of R-Hg+ per ml(for broad-spectrum resistant strains). The method for de-tection of elemental mercury (Hg°) was described previouslyby Nakamura and Nakahara (17). The reaction was carriedout at room temperature for 2 (narrow-spectrum resistance)and 10 (broad-spectrum resistance) h. Positive controls were
run with laboratory strains, while negative controls includingmercury-sensitive strains and uninoculated reaction mix-tures were also conducted simultaneously.
RESULTS AND DISCUSSION
Effects of temperature and mercuric ion on merA expres-sion. The highest relative production of merA transcripts inP. stutzeri OR2 was observed in exponential-growth-phasecells shocked with 25 jig of Hg2+ per ml at 4°C (Fig. 1A). Inthe presence of Hg2+, net production of merA transcriptswas found to increase linearly for 8 h at 37°C and for 4 h at
VOL. 56, 1990
on February 7, 2020 by guest
http://aem.asm
.org/D
ownloaded from
3268 TSAI AND OLSON
TRC/10E7 cells250
B.200
150 no Hg
100 -° 25 wg
50--
0 ,0 1 2 3 4
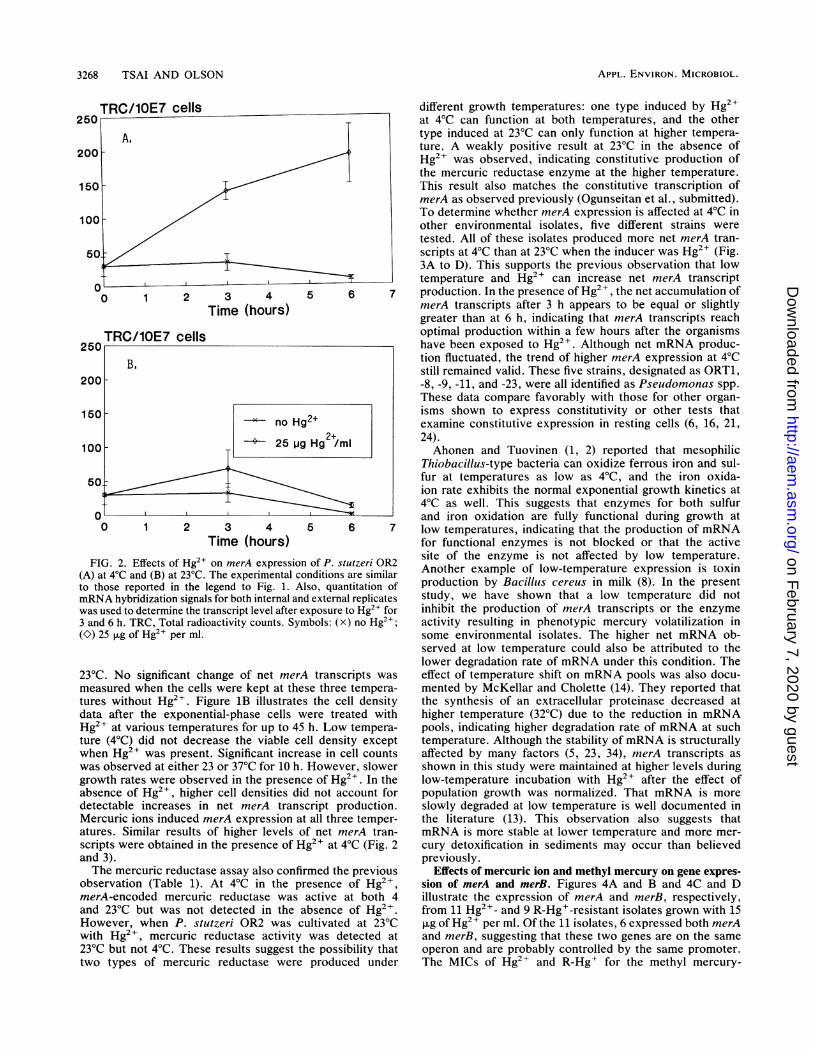
Time (hours)FIG. 2. Effects of Hg2+ on merA expression
(A) at 4°C and (B) at 23°C. The experimental ccto those reported in the legend to Fig. 1. A]mRNA hybridization signals for both internal an(was used to determine the transcript level after e3 and 6 h. TRC, Total radioactivity counts. Syn(O) 25 pLg of Hg2" per ml.
23°C. No significant change of net merAmeasured when the cells were kept at the:tures without Hg2I. Figure 1B illustratesdata after the exponential-phase cells w
Hg2+ at various temperatures for up to 45ture (4°C) did not decrease the viable cewhen Hg2+ was present. Significant incre,was observed at either 23 or 37°C for 10 h.growth rates were observed in the presencabsence of Hg2+, higher cell densities di(detectable increases in net merA transiMercuric ions induced merA expression atatures. Similar results of higher levels oscripts were obtained in the presence of Hand 3).The mercuric reductase assay also confir
observation (Table 1). At 4°C in the prmerA-encoded mercuric reductase wasand 23°C but was not detected in the a
However, when P. stutzeri OR2 was clwith Hg2+, mercuric reductase activity23°C but not 4°C. These results suggest titwo types of mercuric reductase were
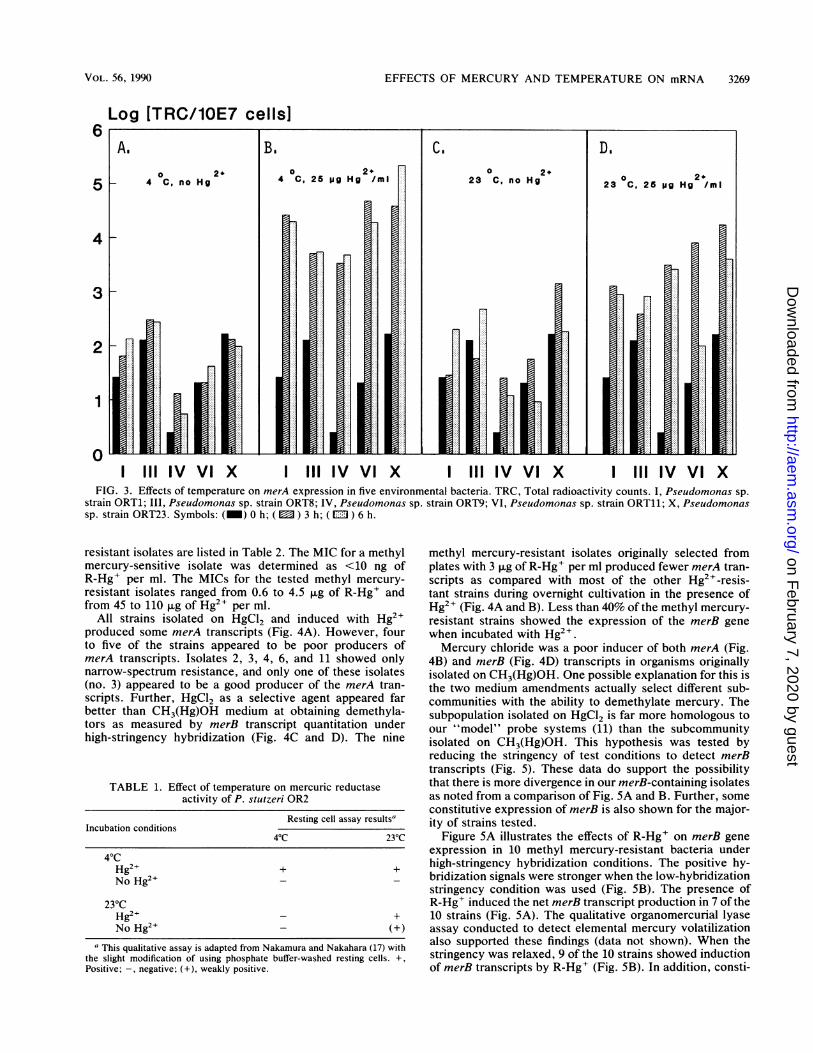
different growth temperatures: one type induced by Hg2"at 4°C can function at both temperatures, and the othertype induced at 23°C can only function at higher tempera-ture. A weakly positive result at 23°C in the absence ofHg2+ was observed, indicating constitutive production ofthe mercuric reductase enzyme at the higher temperature.This result also matches the constitutive transcription ofmerA as observed previously (Ogunseitan et al., submitted).To determine whether merA expression is affected at 4°C inother environmental isolates, five different strains weretested. All of these isolates produced more net merA tran-scripts at 4°C than at 23°C when the inducer was Hg2+ (Fig.3A to D). This supports the previous observation that lowtemperature and Hg2+ can increase net merA transcript
5 6 7 production. In the presence of Hg2+, the net accumulation ofmerA transcripts after 3 h appears to be equal or slightlygreater than at 6 h, indicating that merA transcripts reachoptimal production within a few hours after the organismshave been exposed to Hg2+. Although net mRNA produc-tion fluctuated, the trend of higher merA expression at 4°Cstill remained valid. These five strains, designated as ORT1,-8, -9, -11, and -23, were all identified as Pseudomonas spp.These data compare favorably with those for other organ-isms shown to express constitutivity or other tests thatexamine constitutive expression in resting cells (6, 16, 21,
2+ 2)Hg2+/ml Ahonen and Tuovinen (1, 2) reported that mesophilicThiobacillus-type bacteria can oxidize ferrous iron and sul-fur at temperatures as low as 4°C, and the iron oxida-ion rate exhibits the normal exponential growth kinetics at4°C as well. This suggests that enzymes for both sulfurand iron oxidation are fully functional during growth at
5 6 7 low temperatures, indicating that the production of mRNAfor functional enzymes is not blocked or that the active
i of P. stutzeri OR2 site of the enzyme is not affected by low temperature.nditions are similar Another example of low-temperature expression is toxinIso, quantitation of production by Bacillus cereus in milk (8). In the presentd external replicates study, we have shown that a low temperature did notxposure to Hg2 for inhibit the production of merA transcripts or the enzymenbols: (x) no Hg2+; activity resulting in phenotypic mercury volatilization in
some environmental isolates. The higher net mRNA ob-served at low temperature could also be attributed to thelower degradation rate of mRNA under this condition. The
4 transcripts was effect of temperature shift on mRNA pools was also docu-se three tempera- mented by McKellar and Cholette (14). They reported thats the cell density the synthesis of an extracellular proteinase decreased atiere treated with higher temperature (32°C) due to the reduction in mRNAh. Low tempera- pools, indicating higher degradation rate of mRNA at such11 density except temperature. Although the stability of mRNA is structurallyase in cell counts affected by many factors (5, 23, 34), merA transcripts asHowever, slower shown in this study were maintained at higher levels duringSe of Hg2+. In the low-temperature incubation with Hg2+ after the effect ofd not account for population growth was normalized. That mRNA is morecript production. slowly degraded at low temperature is well documented int all three temper- the literature (13). This observation also suggests thatof net merA tran- mRNA is more stable at lower temperature and more mer-[g2+ at 4°C (Fig. 2 cury detoxification in sediments may occur than believed
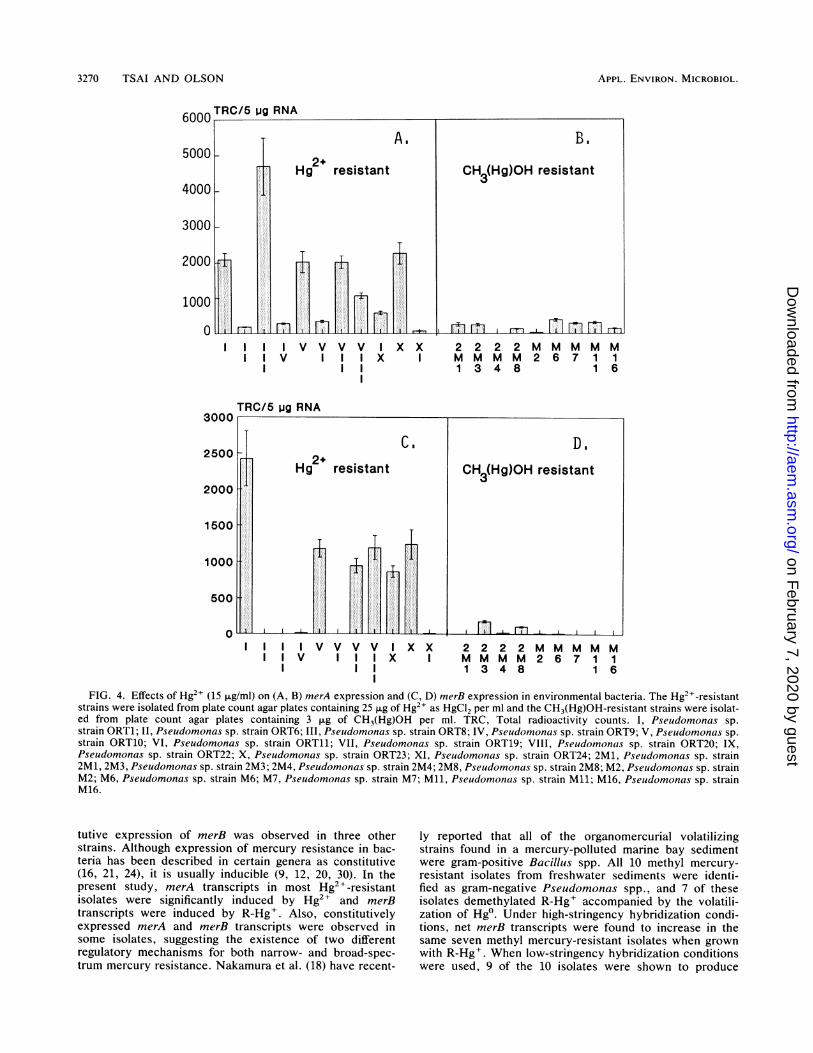
previously.rmed the previous Effects of mercuric ion and methyl mercury on gene expres-resence of Hg2+, sion of merA and merB. Figures 4A and B and 4C and Dactive at both 4 illustrate the expression of merA and merB, respectively,ibsence of Hg2+. from 11 Hg2+- and 9 R-Hg+-resistant isolates grown with 15ultivated at 230C ,ug of Hg2+ per ml. Of the 11 isolates, 6 expressed both merAwas detected at and merB, suggesting that these two genes are on the samehe possibility that operon and are probably controlled by the same promoter.produced under The MICs of Hg2+ and R-Hg+ for the methyl mercury-
APPL. ENVIRON. MICROBIOL.
on February 7, 2020 by guest
http://aem.asm
.org/D
ownloaded from
EFFECTS OF MERCURY AND TEMPERATURE ON mRNA 3269
Log [TRC/10E7 cells]6 1
5
4
3
2
1
0I III IV VI X I IIIIV VIX I IIIIV VIX I IIIIV VI X
FIG. 3. Effects of temperature on merA expression in five environmental bacteria. TRC, Total radioactivity counts. I, Pseudomonas sp.strain ORT1; III, Pseudomonas sp. strain ORT8; IV, Pseudomonas sp. strain ORT9; VI, Pseudomonas sp. strain ORT11; X, Pseudomonassp. strain ORT23. Symbols: (_) 0 h; ( ) 3 h; (m ) 6 h.
resistant isolates are listed in Table 2. The MIC for a methylmercury-sensitive isolate was determined as <10 ng ofR-Hg+ per ml. The MICs for the tested methyl mercury-resistant isolates ranged from 0.6 to 4.5 ,ug of R-Hg+ andfrom 45 to 110 ,ug of Hg2+ per ml.
All strains isolated on HgCl2 and induced with Hg2+produced some merA transcripts (Fig. 4A). However, fourto five of the strains appeared to be poor producers ofmerA transcripts. Isolates 2, 3, 4, 6, and 11 showed onlynarrow-spectrum resistance, and only one of these isolates(no. 3) appeared to be a good producer of the merA tran-scripts. Further, HgCl2 as a selective agent appeared farbetter than CH3(Hg)OH medium at obtaining demethyla-tors as measured by merB transcript quantitation underhigh-stringency hybridization (Fig. 4C and D). The nine
TABLE 1. Effect of temperature on mercuric reductaseactivity of P. stutzeri OR2
Resting cell assay resultsaIncubation conditions
4°C 23°C
40CHg2+ + +No Hg2+ _
230CHg2+ _ +No Hg24 _ (+)
a This qualitative assay is adapted from Nakamura and Nakahara (17) withthe slight modification of using phosphate buffer-washed resting cells. +,Positive; -, negative; (+), weakly positive.
methyl mercury-resistant isolates originally selected fromplates with 3 ,ug of R-Hg+ per ml produced fewer merA tran-scripts as compared with most of the other Hg2+-resis-tant strains during overnight cultivation in the presence ofHg2+ (Fig. 4A and B). Less than 40% of the methyl mercury-resistant strains showed the expression of the merB genewhen incubated with Hg2+.Mercury chloride was a poor inducer of both merA (Fig.
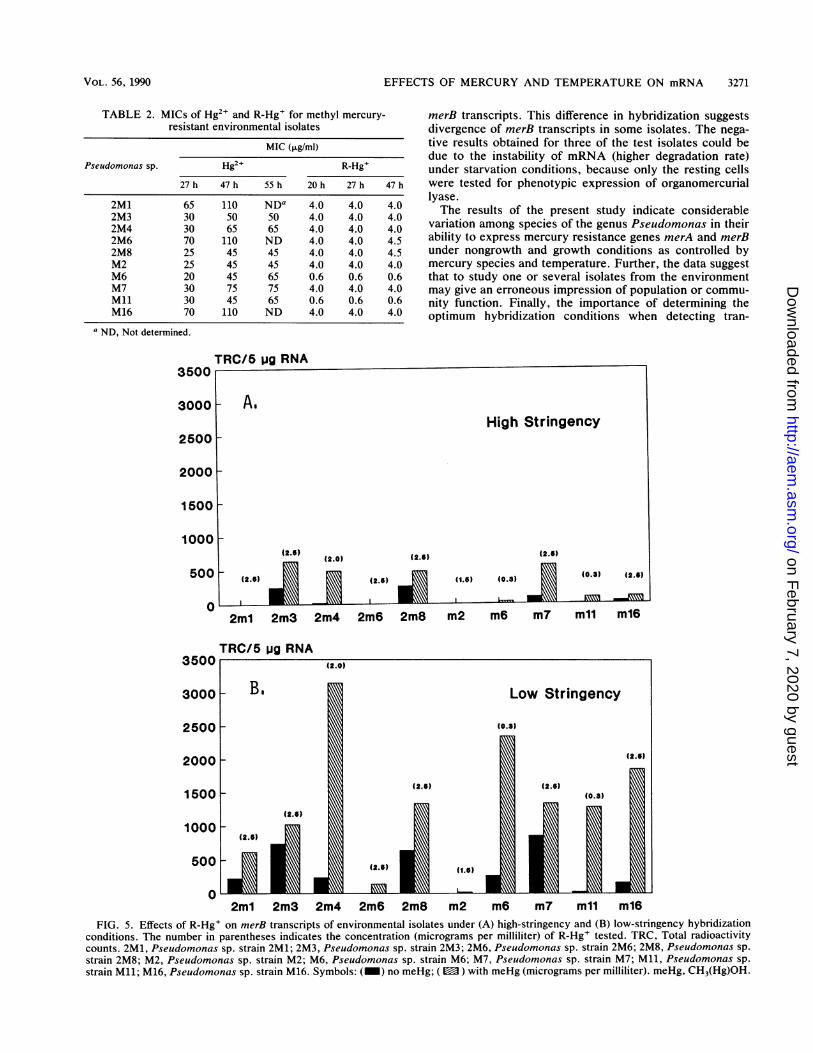
4B) and merB (Fig. 4D) transcripts in organisms originallyisolated on CH3(Hg)OH. One possible explanation for this isthe two medium amendments actually select different sub-communities with the ability to demethylate mercury. Thesubpopulation isolated on HgCl2 is far more homologous toour "model" probe systems (11) than the subcommunityisolated on CH3(Hg)OH. This hypothesis was tested byreducing the stringency of test conditions to detect merBtranscripts (Fig. 5). These data do support the possibilitythat there is more divergence in our merB-containing isolatesas noted from a comparison of Fig. 5A and B. Further, someconstitutive expression of merB is also shown for the major-ity of strains tested.
Figure 5A illustrates the effects of R-Hg+ on merB geneexpression in 10 methyl mercury-resistant bacteria underhigh-stringency hybridization conditions. The positive hy-bridization signals were stronger when the low-hybridizationstringency condition was used (Fig. 5B). The presence ofR-Hg+ induced the net merB transcript production in 7 of the10 strains (Fig. 5A). The qualitative organomercurial lyaseassay conducted to detect elemental mercury volatilizationalso supported these findings (data not shown). When thestringency was relaxed, 9 of the 10 strains showed inductionof merB transcripts by R-Hg+ (Fig. SB). In addition, consti-
VOL. 56, 1990
on February 7, 2020 by guest
http://aem.asm
.org/D
ownloaded from
3270 TSAI AND OLSON
I I I IV V V V I Xx 2 2 2 2 M M M M MI I v I I I X I M M M M 2 6 7 1 1
I I 1 1 3 4 8 1 6l
3000TRC/5 pg RNA
I I I V V V V I Xx 2 2 2 2 M M M M MI I v I I I x I M M M M 2 6 7 1 1II 1 1 3 4 8 1 6
FIG. 4. Effects of Hg2" (15 ,ug/ml) on (A, B) merA expression and (C, D) merB expression in environmental bacteria. The Hg2+-resistantstrains were isolated from plate count agar plates containing 25 p.g of Hg2+ as HgCl2 per ml and the CH3(Hg)OH-resistant strains were isolat-ed from plate count agar plates containing 3 [ig of CH3(Hg)OH per ml. TRC, Total radioactivity counts. I, Pseudomonas sp.strain ORT1; II, Pseudomonas sp. strain ORT6; III, Pseudomonas sp. strain ORT8; IV, Pseudomonas sp. strain ORT9; V, Pseudomonas sp.strain ORT10; VI, Pseudomonas sp. strain ORT11; VII, Pseudomonas sp. strain ORT19; VIII, Pseudomonas sp. strain ORT20; IX,Pseudomonas sp. strain ORT22; X, Pseudomonas sp. strain ORT23; XI, Pseudomonas sp. strain ORT24; 2M1, Pseudomonas sp. strain2M1, 2M3, Pseudomonas sp. strain 2M3; 2M4, Pseudomonas sp. strain 2M4; 2M8, Pseudomonas sp. strain 2M8; M2, Pseudomonas sp. strainM2; M6, Pseudomonas sp. strain M6; M7, Pseudomonas sp. strain M7; Mll, Pseudomonas sp. strain Mll; M16, Pseudomonas sp. strainM16.
tutive expression of merB was observed in three otherstrains. Although expression of mercury resistance in bac-teria has been described in certain genera as constitutive(16, 21, 24), it is usually inducible (9, 12, 20, 30). In thepresent study, merA transcripts in most Hg2+-resistantisolates were significantly induced by Hg2+ and merBtranscripts were induced by R-Hg+. Also, constitutivelyexpressed merA and merB transcripts were observed insome isolates, suggesting the existence of two differentregulatory mechanisms for both narrow- and broad-spec-trum mercury resistance. Nakamura et al. (18) have recent-
ly reported that all of the organomercurial volatilizingstrains found in a mercury-polluted marine bay sedimentwere gram-positive Bacillus spp. All 10 methyl mercury-resistant isolates from freshwater sediments were identi-fied as gram-negative Pseudomonas spp., and 7 of theseisolates demethylated R-Hg+ accompanied by the volatili-zation of Hg°. Under high-stringency hybridization condi-tions, net merB transcripts were found to increase in thesame seven methyl mercury-resistant isolates when grownwith R-Hg+. When low-stringency hybridization conditionswere used, 9 of the 10 isolates were shown to produce
APPL. ENVIRON. MICROBIOL.
on February 7, 2020 by guest
http://aem.asm
.org/D
ownloaded from
EFFECTS OF MERCURY AND TEMPERATURE ON mRNA 3271
TABLE 2. MICs of Hg2' and R-Hg+ for methyl mercury-resistant environmental isolates
MIC (pg/ml)Pseudomonas sp. Hg2+ R-Hg+
27 h 47 h 55 h 20 h 27 h 47 h
2M1 65 110 NDa 4.0 4.0 4.02M3 30 50 50 4.0 4.0 4.02M4 30 65 65 4.0 4.0 4.02M6 70 110 ND 4.0 4.0 4.52M8 25 45 45 4.0 4.0 4.5M2 25 45 45 4.0 4.0 4.0M6 20 45 65 0.6 0.6 0.6M7 30 75 75 4.0 4.0 4.0M11 30 45 65 0.6 0.6 0.6M16 70 110 ND 4.0 4.0 4.0
a ND, Not determined.
merB transcripts. This difference in hybridization suggestsdivergence of merB transcripts in some isolates. The nega-tive results obtained for three of the test isolates could bedue to the instability of mRNA (higher degradation rate)under starvation conditions, because only the resting cellswere tested for phenotypic expression of organomercuriallyase.The results of the present study indicate considerable
variation among species of the genus Pseudomonas in theirability to express mercury resistance genes merA and merBunder nongrowth and growth conditions as controlled bymercury species and temperature. Further, the data suggestthat to study one or several isolates from the environmentmay give an erroneous impression of population or commu-nity function. Finally, the importance of determining theoptimum hybridization conditions when detecting tran-
TRC/5 ug RNA
0
3500
WJ (2. f)E.j
0F
2m1 2m3 2m4 2m6 2m8 m2 m6 m7 mll m16FIG. 5. Effects of R-Hg+ on merB transcripts of environmental isolates under (A) high-stringency and (B) low-stringency hybridization
conditions. The number in parentheses indicates the concentration (micrograms per milliliter) of R-Hg' tested. TRC, Total radioactivitycounts. 2M1, Pseudomonas sp. strain 2M1; 2M3, Pseudomonas sp. strain 2M3; 2M6, Pseudomonas sp. strain 2M6; 2M8, Pseudomonas sp.strain 2M8; M2, Pseudomonas sp. strain M2; M6, Pseudomonas sp. strain M6; M7, Pseudomonas sp. strain M7; Ml, Pseudomonas sp.strain Ml; M16, Pseudomonas sp. strain M16. Symbols: (_) no meHg; (E ) with meHg (micrograms per milliliter). meHg, CH3(Hg)OH.
2m1 2m3 2m4 2m6 2m8 m2 m6 m7 mll m16
TRC/5 pg RNA
VOL. 56, 1990
on February 7, 2020 by guest
http://aem.asm
.org/D
ownloaded from

3272 TSAI AND OLSON
scripts in environmental isolates is obvious from the datapresented.
ACKNOWLEDGMENTS
This work was supported by a grant (8000-25) from the ElectricPower Research Institute.We thank Marie Park for the preparative work for the mRNA
analyses and 0. A. Ogunseitan for reviewing the manuscript.
LITERATURE CITED1. Ahonen, L., and 0. H. Tuovinen. 1989. Microbiological oxida-
tion of ferrous iron at low temperatures. Appl. Environ. Micro-biol. 55:312-316.
2. Ahonen, L., and 0. H. Tuovinen. 1990. Kinetics of sulfuroxidation at suboptimal temperatures. Appl. Environ. Micro-biol. 56:560-562.
3. Barkay, T., C. Liebert, and M. Gillman. 1989. Environmentalsignificance of the potential for mer(Tn2l)-mediated reductionof Hg2" to Hg° in natural waters. Appl. Environ. Microbiol.55:1196-1202.
4. Barkay, T., and B. H. Olson. 1986. Phenotypic and genotypicadaptation of aerobic heterotrophic sediment bacterial commu-nities to mercury stress. Appl. Environ. Microbiol. 52:403-406.
5. Belasco, J. G., G. Nilsson, A. von Gabain, and S. N. Cohen. 1986.The stability of E. coli gene transcripts is dependent on deter-minants localized to specific mRNA segments. Cell 46:245-251.
6. Booth, J. E., and J. W. Williams. 1984. The isolation of amercuric ion-reducing flavoprotein from Thiobacillus ferrooxi-dans. J. Gen. Microbiol. 130:725-730.
7. Chomczynski, P., and N. Sacchi. 1987. Single-step method ofRNA isolation by acid guanidinium thiocyanate-phenol-chloro-form extraction. Anal. Biochem. 162:156-159.
8. Christiansson, A., A. S. Naidu, I. Nilsson, T. Wadstrom, andH.-E. Pettersson. 1989. Toxin production by Bacillus cereusdairy isolates in milk at low temperatures. Appl. Environ.Microbiol. 55:2595-2600.
9. Foster, T. J. 1987. The genetics and biochemistry of mercuryresistance. Crit. Rev. Microbiol. 15:117-140.
10. Fox, B., and C. T. Walsh. 1982. Mercuric reductase. J. Biol.Chem. 257:2498-2503.
11. Griffin, H. G., T. J. Foster, S. Silver, and T. K. Misra. 1987.Cloning and DNA sequence of the mercuric- and organomercu-rial-resistance determinants of plasmid pDU1358. Proc. Natl.Acad. Sci. USA 84:3112-3116.
12. Helmann, J. D., Y. Wang, I. Mahler, and C. T. Walsh. 1989.Homologous metalloregulatory proteins from both gram-posi-tive and gram-negative bacteria control transcription of mercuryresistance operons. J. Bacteriol. 171:222-229.
13. Kennell, D., and I. Bicknell. 1973. Decay of messenger ribonu-cleic acid from the lactose operon of Escherichia coli as afunction of growth temperature. J. Mol. Biol. 74:21-31.
14. McKellar, R. C., and H. Cholette. 1987. Effect of temperatureshifts on extracellular proteinase-specific mRNA pools in Pseu-domonas fluorescens B52. Appl. Environ. Microbiol. 53:1973-1976.
15. Munjaal, R. P., 0. M. Conneely, and R. W. O'Malley. 1989. Insitu detection of progesterone receptor mRNA in the chickenoviduct using probe-on slides. BioTechniques 7:1104-1108.
16. Nakahara, H., J. L. Schottel, T. Yamada, Y. Miyakawa, M.Asakawa, J. Harville, and S. Silver. 1985. Mercuric reductaseenzymes from Streptomyces species and group B Streptococ-cus. J. Gen. Microbiol. 131:1053-1059.
17. Nakamura, K., and H. Nakahara. 1988. Simplified X-ray filmmethod for detection of bacterial volatilization of mercurychloride by Escherichia coli. Appl. Environ. Microbiol. 54:2871-2873.
18. Nakamura, K., M. Sakamoto, H. Uchiyama, and 0. Yagi. 1990.Organomercurial-volatilizing bacteria in the mercury-pollutedsediments of Minamata Bay, Japan. Appl. Environ. Microbiol.56:304-305.
19. Nelson, J. D., and R. R. Colwell. 1975. The ecology of mercury-resistant bacteria in the Chesapeake Bay. Microb. Ecol. 1:191-218.
20. O'Halloran, T. V., B. Frantz, M. K. Shin, D. M. Ralston, andJ. G. Wright. 1989. The MerR heavy metal receptor mediatespositive activation in a topologically novel transcription com-plex. Cell 56:119-129.
21. Olson, G. J., F. D. Porter, J. Rubinstein, and S. Silver. 1982.Mercuric reductase enzyme from a mercury volatilizing strain ofThiobacillus ferrooxidans. J. Bacteriol. 151:1230-1236.
22. Rinderle, S. J., J. E. Booth, and J. W. Williams. 1983. Mercuricreductase from R-plasmid NR1: characterization and mechanis-tic study. Biochemistry 22:869-876.
23. Ross, J. 1989. The turnover of messenger RNA. Sci. Am.260:48-55.
24. Rudrik, J. T., R. E. Bawdon, and S. P. Guss. 1985. Determina-tion of mercury and organomercurial resistance in obligateanaerobic bacteria. Can. J. Microbiol. 31:276-281.
25. Sahlman, L., A.-M. Lambeir, S. Lindskog, and H. B. Dunford.1984. The reaction between NADPH and mercuric reductasefrom Pseudomonas aeruginosa. J. Biol. Chem. 259:12403-12408.
26. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecularcloning: a laboratory manual, 2nd ed. Cold Spring HarborLaboratory Press, Cold Spring Harbor, N.Y.
27. Schottel, J. L. 1978. The mercuric and organomercurial detox-ifying enzymes from a plasmid-bearing strain of Escherichiacoli. J. Biol. Chem. 253:4341-4349.
28. Silver, S., and T. K. Misra. 1988. Plasmid-mediated heavy metalresistances. Annu. Rev. Microbiol. 42:717-743.
29. Spangler, W. J., J. L. Spigarelli, J. M. Rose, and H. M. Miller.1973. Methylmercury: bacterial degradation in lake sediments.Science 180:192-193.
30. Summers, A. 0. 1986. Organization, expression, and evolutionof genes for mercury resistance. Annu. Rev. Microbiol. 40:607-634.
31. Summers, A. O., and E. Lewis. 1973. Volatilization of mercuricchloride by mercury-resistant plasmid-bearing strains of Esch-erichia coli, Staphylococcus aureus, and Pseudomonas aerug-inosa. J. Bacteriol. 113:1070-1072.
32. Tezuka, T., and K. Tonomura. 1976. Purification and propertiesof an enzyme catalyzing the splitting of carbon-mercury link-ages from mercury-resistant Pseudomonas K-62 strain. J. Bio-chem. 80:79-87.
33. Weiss, A. A., S. D. Murphy, and S. Silver. 1977. Mercury andorganomercurial resistances determined by plasmids in Staph-ylococcus aureus. J. Bacteriol. 132:197-208.
34. Wong, H. C., and S. Chang. 1986. Identification of a positiveretroregulator that stabilizes mRNAs in bacteria. Proc. Natl.Sci. USA 83:3233-3237.
35. Zhu, Y. S., and S. Kaplan. 1985. Effects of light, oxygen, andsubstrates on steady-state levels of mRNA coding for ribulose-1,5-bisphosphate carboxylase and light-harvesting and reactioncenter polypeptides in Rhodopseudomonas sphaeroides. Appl.Environ. Microbiol. 162:925-932.
APPL. ENVIRON. MICROBIOL.
on February 7, 2020 by guest
http://aem.asm
.org/D
ownloaded from






![e'e - wsrc.mnis.wsrc.mn › uploads › files › ... › sanhuu...sudalgaa.pdf · 3 ®a¯¯ewel¯¯^ Owf`bo gw]` hg hg hg hg hg hg »k»el [mmjZel - Gbcllmk]Zca»\r»»j»e wawfrb]q](https://img.pdfslide.net/doc/110x75/5f122ea398fae574504c501e/ee-wsrcmniswsrcmn-a-uploads-a-files-a-a-sanhuu-3-aewel.jpg)
![SOIL ANNUAL REPORT - SOIL Haiti - SOIL Haiti · 2015. 9. 4. · *-%))) `Zeehgl h_ pZlm^ _khf hnk mhbe^ml Zkhng] AZbmb' Hg\^ lZ_^er ]^ebo^k^] mh hg^ h_ hnk mk^Zmf^gm lbm^l% ma^ pZlm^](https://img.pdfslide.net/doc/110x75/6123b4d5d307f120895da0ed/soil-annual-report-soil-haitiaaa-a-soil-haitiaaa-2015-9-4.jpg)





















