Embed Size (px)
Citation preview

Estudios geol., 46: 453-464 (1990)
HIPPIDION SP. (MAMMALIA, PERISSODACTYLA, EQUIDAE)EN SEDIMENTOS DEL PLEISTOCENO SUPERIOR DEL URUGUAY
(EDAD MAMIFERO LUJANENSE)
M. Ubilla * y M. T. Alberdi **
RESUMEN
Se describen los restos más completos de Hippidion sp. reportados hasta el momentopara el Pleistoceno de Uruguay. Corresponden a una forma robusta con extremidades anteriores relativamente cortas respecto del cráneo grande. Se analizan sus caracteres cráneo-mandibulares y postcraneales comparados con otros «hippidiformes» de Sudamérica. El marcosedimentario corresponde a una planicie de inundación de un sistema fluvial meandriformede la Fm. Sopas (Pleistoceno superior; Edad mamífero Lujanense) del Uruguay.
Palabras clave: Perissodactyla, Hippidion, Pleistoceno superior, Paleoambiente, Fm. Sopas, Uruguay.
ABSTRACT
The most complete remains of Hippidion sp. to the Pleistocene of Uruguay are described. They agree with a robust form with fore limbs relatively short to the large skull. Cranial, mandibles and postcranial characters are analized and compared with respect to others «hippidiformes» of South America. The depositional environment belong to floodplainfacies of meandering fluvial systems.
Key words: Perissodactyla, Hippidion, Upper Pleistocene, Paleoenvironment, Sopas Fm.,Uruguay.
Introducción
En estos últimos años los caballos monodáctilos delPleistoceno de Neotrópica son objeto de nuevos estudios. Existe un renovado interés por dilucidar aspectos vinculados a su ordenamiento taxonómico, alas relaciones filogenéticas de los taxones involucrados y a su contexto bioestratigráfico. (Alberdi, 1987;Alberdi el al., 1986; Alberdi el al., 1987; MacFaddeny Azzarolli, 1987, y Prado, 1984 a y b.)
La presencia de équidos en el Pleistoceno de Sudamérica es consecuencia del «Gran IntercambioFaunístico Americano», acontecido en el Plio-pleistoceno y relacionado con la instalación del istmo centroamericano (Bond, 1986; Reig, 1981, y Webb,1978). Se reconocen los «equidiformes», representados por varias especies de Equus (Amerhippus) (verMacFadden y Azzarolli, 1987) y los «hippidiformes»,sobre los cuales existe controversia a nivel genérico.Estos últimos eran considerados formas endémicassudamericanas hasta la publicación de MacFadden y
Skinner (1979), quienes describen Onohippidium galushai para el Plioceno temprano (Hemphilliense tardía) de Arizona, e Hippidion sp. para el Mioceno tardío (Hemphilliense temprano) de Texas, USA. Tales hallazgos descartan el endemismo e implican uncarácter alóctono y de emigrantes primarios (sensuMarshall el al., 1982) a nivel genérico de las formasrepresentadas en el Pleistoceno sudamericano, sinperjuicio del desarrollo de categorías específicaseventualmente autóctonas secundarias (sensu Reig,1981).
Recientemente, Alberdi y Prado (en prensa), dudan de la presencia de este grupo en Norteamérica yconsideran que los restos descritos por MacFadden ySkinner (1979), tanto desde un punto de vista estratigráfico como morfológico, deben considerarsepróximos al grupo Dinohippus-Aslrohippus y podríansituarse en la línea filética de los «hippidiformes».
Respecto del Uruguay, las referencias bibliográficas de restos fósiles de équidos, en especial de «hippidiformes», tienen en general una dudosa asignación
• Dpto. de Paleontología, Facultad de Ciencias. 11200 Tristán Narvaja 1674. Montevideo, Uruguay.•• Museo Nacional de Ciencias Naturales. CSIC. J. Gutiérrez Abascal, 2. 28006 Madrid. España.

454
estratigráfica o carecen de ella y están basados en elementos aislados o fragmentarios: dientes o partes esqueléticas incompletas (d. Burmeister, 1875; Jones,1956; Kraglievich, 1928; Lambert, 1941; Roselli,1938, y Teisseire, 1928).
A partir de esta perspectiva y en relación con unestudio del contenido paleomastofaunístico de depósitos pleistocénicos tardíos del norte del Uruguay,fueron hallados materiales de équidos significativos.Se trata de los restos de «hippidiformes» más completos ttasta ahora exhumados en este país, los quereúnen elementos cráneo-mandibulares y dentarios,con parte del postcráneo pertenecientes a un mismoindividuo. Nos proponemos dar a conocer estos materiales, analizando los caracteres relevantes para su
M. UBILLA, M. T. ALBERDI
determinación y aportando elementos para ulterioresestudios del grupo en general.
Hemos adoptado como hipótesis de trabajo, lacondición monogenérica de los «hippidiformes» sudamericanos (sensu Alberdi, 1987) --esto es- laexistencia de Hippidion compuesto por un conjuntode especies cuyo análisis se encuentra en prensa (AIberdi y Prado).
Estratigrafía
En el norte del Uruguay, especialmente en losDptos. de Artigas, Salto, Paysandú y Tacuarembó,se encuentran sedimentos continentales denominados por Antón (1975) como Fm. Sopas (ver fig. 1 A).
N
f~-------I--"""'ct"~t-;;:::J'I~~-- 30'2.'
cm
50
cm
Fig. l.-A) Ubicación geográfica del material estudiado.B) Distribución espacial de los restos de Hippidion sp.

HIPPlDlON SP. (MAMMALIA, PERISSODACfYLA, EQUIDAE) EN SEDIMENTOS DEL PLEISTOCENO... 455
En esta unidad predominan litologías finas, en especiallimos pardos-claros, enriquecidos ocasionalmente por elementos groseros, con una potencia que puede alcanzar los 6 m, con consolidación pobre y frecuentes inclusiones de materia orgánica. Como expresiones de variación faciológica locales y sedimentologías de tipo limo-arcillosas, limo-arenosas, arcillo-arenosas a franco-limosas; en algunas áreas estosdepósitos desarrollan en el tope niveles calcáreos planares de gran extensión horizontal y de potencia de0,50 a 1 m. En afloramientos del Dpto. de Artigas,se ha registrado un conjunto de taxones de vertebrados -en especial mamíferos- como resultado de relevamientos paleontológicos que uno de los autores(MU) se encuentra realizando. En el cuadro 1 sebrinda la lista de taxones de mamíferos, incluyendoel área con los sedimentos portadores de Hippidionsp. así como otras próximas, pertenecientes a los mismos depósitos. La información en particular de estosmateriales se encuentra en Ubilla (1983,1986). La sedimentología del nivel portador de Hippidion sp. incluida en la columna estratigráfica correspondiente ala localidad Pintado y áreas vecinas (ver fig. 2) que-
da comprendida en la caracterización previa. Los niveles representados por sedimentos limo-arenosos ylimosos comportan a nivel global la mayor parte delos fósiles de vertebrados hasta ahora exhumados. LaFm. Sopas ha sido asumida -en términos prácticos- como unidad litológica de referencia para asignar estratigráficamente restos fósiles (cf. Martínez,1988; Olazarri, 1980; Ubilla, 1986, y Ubilla y Martínez, 1988). Prost (1982) y Bombín (1975, 1976),correlacionaron estos depósitos con sedimentos pleistocénicos tardíos denominados Fm. Touro Passo(Rio Grande do Sul, Brasil). El momento en el cualse produjeron los procesos depositacionales pareceacotarse al Pleistoceno superior: el conjunto de géneros y especies de mamíferos hallados asociados sugieren la Edad-mamífero Lujanense, correspondiente a dicho intervalo de tiempo (Ubilla, 1986). Talesconsideraciones son ratificadas por Perea y Martínez(1989), quienes analizan el contenido paleofaunístico de otra localidad geográfica (Dpto. de Tacuarembó), pero perteneciente a esta formación y proponenel Pleistoceno superior como marco temporal. Estaspropuestas coinciden con la correlación de Bombín
COLUMNA ESTRATIGRAFICAFm. Sopas (Pleistoceno sup.)Localidad: Pintado, Río Cuareim, D. Artigas
m Ar.L.AfAm. Con.
r-.--" Clll:-:
6
-"
~-
4 :..-::;~.
:...-:
o~ /
REFERENCIAS
bTI Arena media
O Arena fina
~ Conglomerado
~ Limo
El Limo arenoso
~ Laminación paralela
~ Est. ondulada
~ Fósil: Hippidion sp.
~ Caliche
Fig. 2.- Columna estratigráfica de la localidad Pintado, Río Cuareim, Dpto. Artigas, Uruguay. Fm. Sopas, Pleistoceno superior. La flecha indica la ubicación de
los restos de Hippidion sp.

456 M. UBILLA, M. T. ALBERDI
Paleoambientes
Cuadro l.-Lista de géneros y especies de mamíferoscorrespondientes a los afloramientos de la Fm. Sopas en la
localidad Pintado, Dpto. Artigas y áreas vecinas.
(1976) mencionada supra, entre la Fm. Touro Passo(Rio Grande do Sul, Pleistoceno sup., C14
: 11.000± 190 años AP) y la Fm. Sopas de Uruguay; por otraparte, Olazarri (1980), le asigna una edad entre11.000 y 15.000 años de acuerdo con dataciones deC14 realizadas en estos depósitos.
Según Antón (1975), habrían actuado en la génesis de los sedimentos de la Fm. Sopas, factores eólicos y regímenes fluviales. Numerosas observacionestafonómicas realizadas en torno a los fósiles incluidos en afloramientos del Opto. de Artigas, permitenconstatar una marcada tendencia a registrarse -además de elementos óseos aislados- esqueletos bastante completos con asociación y articulación de diversas estructuras, sugiriendo procesos de desarticulación incompletos, lo que puede estar vinculado con
breves exposiciones subaéreas y traslado ausente oen pequeña escala (Hill, 1979, Y Beherensmeyer,1988). Tales expresiones tafonómicas son compatibles con los procesos depositacionales vinculados asistemas fluviales (Beherensmeyer, op. cit.) y globalmente pueden corresponder a agregados fosilíferosparautóctonos-alóctonos siguiendo a Kidwell et al.(1986).
En el caso particular de los restos exhumados deHippidion la disposición de los elementos en el sedimento -tal como lo ilustra la figura 1B- indica unaestrecha asociación espacial de estructuras que noconservan la articulación original. Dichas estructurasconforman en el esqueleto unidades mayores (v. gr.:cráneo-mandíbula, miembro anterior derecho) íntimamente asociadas entre sí.
En un análisis primario de los sedimentos portadores de Hippidion sp. observamos una secuenciagrano-decreciente, con depósitos conglomeráticos yarenosos basales y limo-arenosos y limosos terminales (ver fig. 2). Tales expresiones sugieren respectode la base, la existencia de canal fluvial y barras demeandros y para los elementos finos un ambiente deplanicie de inundación. En primera instancia seríanasimilables a un sistema fluvial de tipo meandriforme.
La información paleoambiental que puede inferirse de los taxones hasta ahora analizados pone en evidencia la existencia de cuerpos de agua lénticos y reotópicos así como áreas de planicies (Ubilla, 1986); aesto debe agregarse información concurrente proporcionada por la malacofauna perteneciente a esta formación (Martínez, 1988; Olazarri, op. cit., Y Ubillay Martínez, op. cit.)
Orden Litopterna
Familia Maeraueheniidaecf. Macrauchenia
Orden Notoungulata
Familia ToxodontidaeToxodon p/atensis
Orden Tardigrada
Familia MegatheriidaeMegatherium cf. americanum
Familia MylodontidaeG/ossotherium robustumLestodon cf. armatus
Clase MAMMALIA
Orden CINGULATA
Familia DasypodidaePampatherium humbo/dti
Familia GlyptodontidaePanochthus tubercu/atusG/yptodon ciavipes
Orden CARNIVORA
Familia FelidaeFelis conc%rcf. Smi/odon
Orden RODENTIA
Familia CricetidaeH%chilus sp.
Familia CaviidaeCavia sp.
Familia HydrochoeridaecL HydrochoerusNeochoerus sp.
Familia MyoeastoridaeMyocastor coypus
Orden PROBOSCIDEA
Familia GomphotheriidaeGomphotheriinae indet.
Orden Perissodactyla
Familia EquidaeHippidion sp.
Familia TapiridaeTapirus sp.
Orden ARTIODACTYLA
Familia TayassuidaeCatagonus sp.
Familia CamelidaePa/aeo/ama paradoxa
Familia CervidaeOzotocerus bezoarticusAntifer eL ultraMorene/aphus brachycerosMorene/aphus cf. /ujanensis
Material y métodos
Los restos estudiados provienen del Opto. de Artigas, localidad Pintado, margen uruguaya del RíoCuareim (fig. lA), en sedimentos pleistocénicos tardíos (Fm. Sopas, Antón, 1975). Pertenecen a la colección de Paleontología del Museo Histórico Departamental de Artigas (MHD-P). Las variables incluidas en las tablas 1 a V fueron registradas de acuerdocon Eisenmann et al. (1988) y su nomenclatura respeta puntualmente la propuesta por estos autores. Sebrinda a continuación el significado de las siglas delos materiales comparativos incluidos en las tablas I-V: MLP: Museo de La Plata, Argentina;MCNBR: Museo Argentino de Ciencias Naturales,«Bernardino Rivadavia», Buenos Aires, Argentina;TAR: Tarija, Bolivia, Colecciones del Museo Nacional de Historial Natural de París; MHN-B: Museo deHistoria Natural de La Paz, Bolivia.

HIPPIDION SP. (MAMMALIA, PERISSODACTYLA, EQUIDAE) EN SEDIMENTOS DEL PLEISTOCENO... 457
Sistemática
Orden Perissodactyla Owen, 1848.Familia Equidae Gray, 182l.Género Hippidion Owen, 1869.
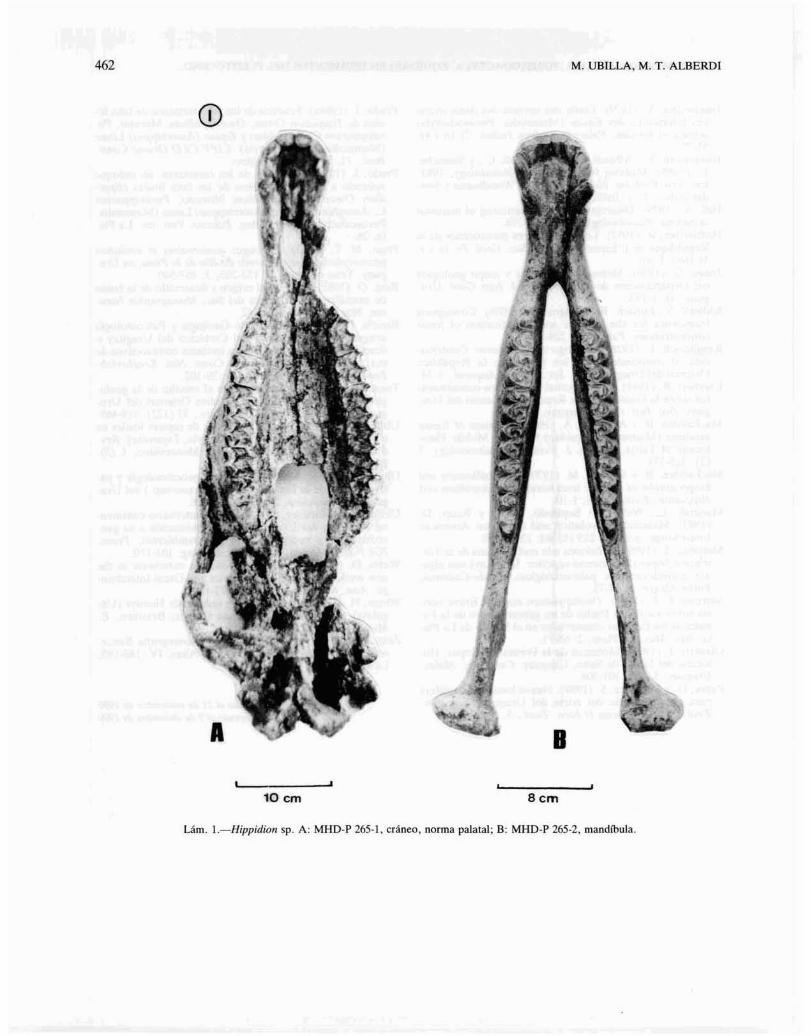
Hippidion sp. (láms. 1-111).
Material: MHD-P 265-1: cráneo (completo en vista palatal), 265-2: mandíbula completa, ambos conincisivos y series de molariformes; 265-3: atlas; 265-4:cúbito-radio derecho; 265-5: Mclll derecho con Mclly MclV fusionados; 265-6: l."FlII anterior; 265-7:3. aFIII anterior; 265-8: lunatum, y 265-9: piramidalderechos.
Medidas: ver tablas I a V.
Procedencia geográfica y estratigráfica: Dpto. Artigas, localidad Pintado, Río Cuareim (fig. lA);Fm. Sopas (Pleistoceno superior) (ver columna estratigráfica, fig. 2). Colector: J. Soloviy.
Descripción del material. Cráneo: en vista palatalestá prácticamente completo; la norma superior fueerosionada por exposición subaérea y por esta razónno conserva la región facial y orbitaria así como laparte superior de la caja craneana. El arco cigomático derecho ha sido preservado; el paladar es bastante amplio; los cóndilos occipitales están divididosy separados por un surco ventral; basioccipital y basiesfenoides completos, mientras que el vómer sóloparcialmente presente; los pterigoides están fragmentados conservando un robusto gancho pterigoideo derecho. Las apófisis yugulares están rotas. Las cavidades glenoideas son someras e internamente a lasmismas se desarrollan profundas depresiones para inserciones musculares. Las bullas auditivas no se hanconservado intactas pero se aprecian los meatos auditivos externos dirigidos hacia afuera y algo haciaatrás; en tamaño relativo son pequeñas' a juzgar porel espacio que ocupan (lam. 1).
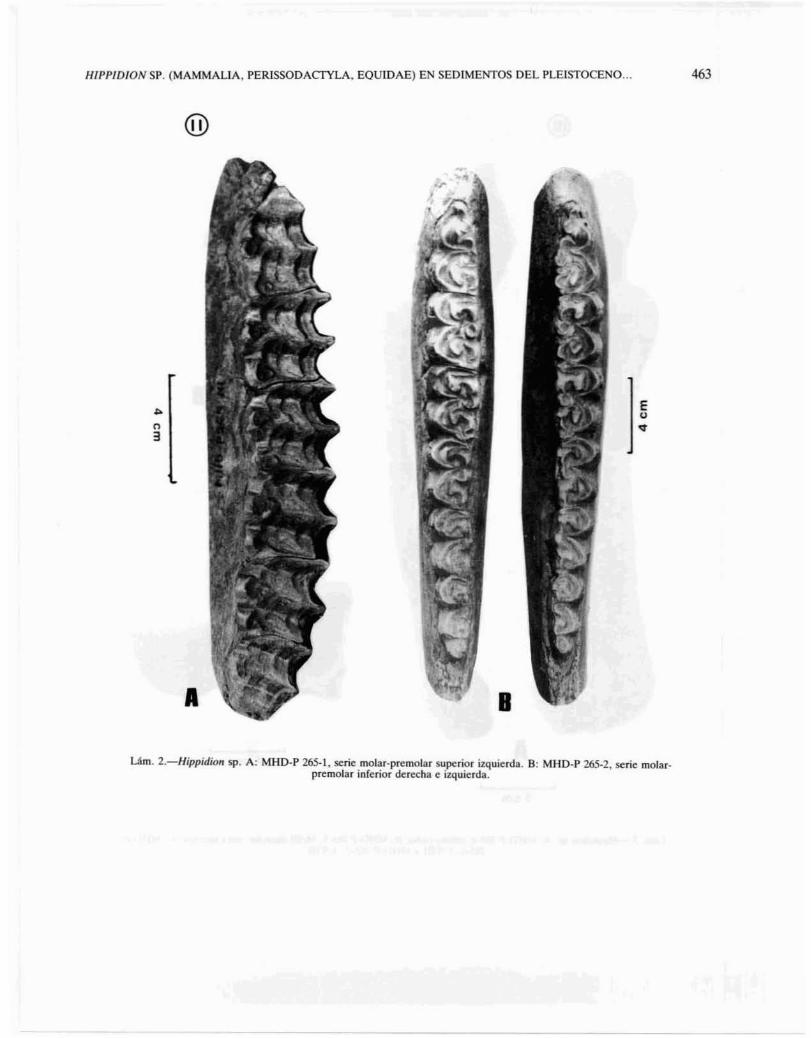
Dentición superior (lam. lA y IIA): los incisivosforman una arcada semicircular y presentan infudíbula. Los caninos son grandes y los p3 Y p4 son loselementos más grandes de la serie de molariformes.Están presentes los pI pequeños y desgastados. El diseño de la superficie masticatoria es simple y con escasos pliegues, con cemento grueso. Los plegamientos existentes se encuentran en el borde posterior dela prefoseta y en el anterior de la postfoseta, en especial en p3
, MI Y M2• Parastilos y mesostilos bien
desarrollados, gruesos y sin pliegue. Los protoconosno tienen desarrollo anterior y, en general, son ovales a elípticos con bordes no angulosos, unidos al protolofo por un istmo ancho. En la mayoría de los molariformes, el surco preprotoconal y el valle postprotoconal están bien marcados. En MI y M2 se observa
un delgado pliegue caballino. En el área hipoconalse observa la constricción homónima bien marcadaen p3 y p4 Ydesarrollo de isleta hipoconal en p2
, p3
Yp4. El hipocono es de menor tamaño que el proto
cono.Mandíbula (lam. 1): ambas ramas mandibulares es
tán prácticamente completas; careciendo la hemimandíbula izquierda de la apófisis coronoides; anteriormente son elongadas. La rama ascendente es profunda y la horizontal se engrosa en el área de los molariformes a expensas de la línea milohioidea. El cóndilo mandibular en vista posterior es asimétrico hacia afuera respecto de la apófisis coronoides.
Dentición inferior (lam. lB y IIB): los incisivosforman una arcada dentaria semicircular y carecende infudíbula. Los caninos se sitúan bien próximosal 13 , La superficie oclusal es simple, con escasos pliegues, con cemento grueso. Los premolares tienenmayor talla que los molares. Protocónido e hipocónido tienen una constante forma de media luna, conbordes externos continuos y ausencia de pticostílidoy pliegue antecaballínido. Ectofléxidos en generalprofundos -especialmente en los P2 , MI, M2 YM3
sin llegar a afectar el istmo pronunciadamente en P3y P4, El «double-knot» en general no tiene bordesmarcadamente angulares, con metacónidos y metastílidos subiguales en talla; linguafléxidos en forma deV poco profundo y subrecto en P3 , P4 YMI' Se observa cierta asimetría en el P2 izquierdo respecto delderecho, en donde el ectofléxido es claramente másprofundo (lam. IIB).
Metacarpianos (lam. IIIB): en su conjunto configuran una pieza corta, robusta y ancha en sus epífisis y en la diáfisis. A nivel de la articulación proximal desarrolla facetas articulares para el hueso grande, unciforme y trapezoide, no observándose para eltrapecio por rotura del material. No observamos faceta articular para el McV. Los Mcll Y MclV pueden visualizarse en vista anterior y sobresalen bastante en vista posterior. El Mcll se extiende distalmente más que el IV. En la faceta articular distal seobserva una cresta que la recorre en sentido anteroposterior de posición submediana ubicándose ligeramente hacia afuera, definiendo dos subfacetas de lasque la interna se ubica algo más proximal (levantada) que la externa.
Discusión sistemática
En este artículo se asume como hipótesis de trabajo la propuesta de Alberdi (op. cit.) sobre la condición monogenérica de los «hippidiformes», determinando este material como Hippidion sp. Su status específico deberá ser evaluado en un análisis generalde las especies de Hippidion existentes en Sudamé-
458 M. UBILLA, M. T. ALBERDI
rica. El patrón oc1usal de la dentición craneal y man- Tabla l.-Dimensiones (mm) craneanas y mandibulares de A:
dibular, así como elementos postcraneales asociados Hippidion sp. (MHD·P 265); B YE: Onohippidium; C y D:
nos permite identificar estos restos como indiscuti- Hippidion principale. Se incluyen en Hippidion sp. las series
blemente pertenecientes al grupo de los «hippidifor- dentarias izquierda y derecha correspondientes con las columnasrespectivas. Odimensiones aproximadas. (Nomenclatura según
mes». Debemos señalar en este sentido y respecto de Eisenmann et al., 1988.)la serie de molariformes superiores la presencia deprotoconos en general ovales a elípticos y no desarro- A B Cllados anteriormente, a diferencia de la típica sub- Craneo: MHD-P 265-1 MLP-62 4102 MCNBRtriangularidad que se observa en Equus (A.), a lo 1 174 162 181,3que se agrega un istmo ancho que lo une al protolo- 2 152,7 102 154
fo y con un marcado desarrollo del surco preproto- 5 249 290 270
conal y valle postprotoconal. Acompañan a este mo- 6 575 556 600
delo fuertes mesostilos y parastilos. En la dentición 7 107,8-108,2 103,5 1048 92,6- 92,5 85 91,2
inferior, cabe señalar la constancia en la disposición 9 194,0-195,0 185 188,5seleniforme de los protocónidos e hipocónidos con 10 102,8 132 102,5bordes continuos exentos de pliegue antecaballínido 11 46 44 (44)
Ycaballínido (pticostílido), usualmente presente en 12 55 43 (40)
Equus (A.). A diferencia de lo que ocurre en éste, 13 94,4 83,5 70
los ectofléxidos son marcadamente profundos en la 14 50,7 56 42,515 64,6 63,7
mayoría de los molariformes --especialmente en P2' 16 82 126 114MI> M2 Y M3- asociados a «double-knot» sin bor- 17 162 192 210,5des marcadamente angulares con metacónidos y me- 21 122,5 136,5 135,5
tastílidos subiguales y linguafléxidos poco profundosA D E
Ysubrectos. En Equus (A.) el metastílido es angulo- Mandíbula: MHD-P 265-2 MLP-6,19 9671 MCNBRso y ellinguafléxido en los molares forma un istmo.
1 490 500 486Globalmente la superficie oclusal de ambas series de 2 124,3 146 135molariformes es simple y con cemento grueso. Res- 3 102,8-103,4 92 106pecto de la presencia-ausencia de infudíbula y en 4 97,2- 98,5 90,5 93,2base a los datos de Eisenmann (1979), nosotros cree- S 199,0-200,0 184 200
mos que éstos suelen estar en relación con la edad 6 140,3 (155) 133
del individuo, llegando a desaparecer en determina- 7 (SOr) 60,58 207 (250) 210
do momento del desgaste. 9 198 (244) 202De acuerdo con la caracterización de Hippidion y 10 112 133 120
Onohippidium de MacFadden y Skinner (1979) 11 79,3 91,2 89-quienes consideran ambos géneros válidos- nues- 12 65 75 67
tro material no queda totalmente comprendido en el 13 111,4 109 109,7
modelo manejado por estos autores. Desde este puno 14 48,3 34,7
to de vista, en nuestro ejemplar los P3 y P4 tienen ec-15 35
Tabla 2.-Dimensiones (mm) dentarias de Hippidion sp. (MHD-P 265-2). Las columnas izquierda (1) y derecha (d) corresponden a lasseries dentarias respectivas en todos los molariformes. FP: forma del protocono. Nomenclatura según Eisenmann et al. (1988).
Dentición superior
ip2d p3 pe MI M2 M3
1 45,5 64 71,5 62,8 66 622 37,9-38,4 32,5-32,6 31,5-31,0 30,0-30,0 29,0-28,2 25,3-25,03 7,0- 7,4 6,3- 6,6 7,0- 7,0 7,5- 7,4 8,0- 8,2 8,0- 8,64 26,6-26,7 30,5-32,0 31,5-31,0 29,4-30,7 27,8-27,4 25,0-24,6
FP 1-2 1 1-2 2 2 2
Dentición inferior
i P2 d P3 P4 MI M2 M3
2 34,8-35,0 31,2-31,0 31,1-31,4 28,5-28,4 28,2-27,4 33,4-33,43 7,0- 7,0 7,8- 7,6 7,1- 7,9 4,0- 5,0 5,8- 5,6 7,0- 7,04 11,6-11,0 12,2-12,0 11,2-11,4 11,9-11,4 10,0-10,4 11,5-11,05 9,4-9,0 8,0- 8,5 7,0- 8,0 7,6- 8,0 5,6- 7,0 5,4- 5,06 19,0-17,6 19,0-19,5 18,4-17,8 15,4-16,0 15,0-14,2 12,4-12,0

HIPPIDION SP. (MAMMALIA, PERISSODAcrYLA, EQUIDAE) EN SEDIMENTOS DEL PLEISTOCENO... 459
tofléxidos que no afectan mayormente el istmo, debiendo descartarse Hippidion, pero no puede ser incluido en Onohippidium dado que, según los autorescitados supra, éste tiene ectofléxidos someros en PPpermanentes y profundos en PP y MM deciduos ynuestro ejemplar posee molariformes permanentes yectofléxidos profundos en P2 y en los molares. Estopodría explicarse si asumimos que las formas norteamericanas son primitivas y con una relación de atarismo lejana respecto de las formas del Pleistocenotardío de Sudamérica, como plantean Alberdi y Pra-
do (en prensa). También hay que tener en cuenta lasmodificaciones de las figuras en las superficies oclusales de équidos hipsodontos producto del desgaste.
El McIlI es robusto y presenta fusionados hasta lamitad de su diáfisis a los McIl y McIV, coincidiendoen general con la morfología de Hippidion.
Alberdi (1987) propuso que los «hippidiformes»se ordenen en tres grupos diferentes entre sí respecto del tamaño. Dicha hipótesis ha sido corroborada mediante análisis comparativos numéricos (AIberdi y Prado, 1989). De acuerdo con la informa-
Tabla 3.-Dimensiones (mm) del radio y u1na de A: HippúJion sp. (MHD-P 265-4); B: HippúJion principale; C: H. neogeus.Nomenclatura según Eisenmann et al. (1988).
Radio 2 3 4 5 6 7 8 9 10 11 12
A MHD-P 265-4 310 296 52 31 86 41 89 71,5 38 86 32 20B Tar 892 334 329,2 57,3 31,1 90,8 44,4 105 74,6 41,3 90,8 31,4 20,8e Tar 946 310 300,5 50 29,2 81,5 39,1 91,9 67,3 39,8 78,2 28 17,9D Tar 940 313 302 51,2 29,5 86,8 39,5 93,9 69,8 41,7 81,5 28,6 18
Ulna 1 2 3 4 5MHD-P 265-4 391 76 89 57,2 65
Tabla 4.-Dimensiones (mm) del Mcm de A: HippúJion sp. (MHD-P 265-5), B: TAR 962, C: TAR 896, D: 401 MCNBR,E: MHN-B-101015, F: MHN.B-001014, G: MLP 6.026. IG: indice de gracilidad. Nomenclatura según Eisenmann et al. (1988).
McHI2 3 4 S 6 7 8 10 11 12 13 14 16 lG
A 198 184 42 30,4 58,9 34,5 51,6 14,4 56,6 52 36,5 28,6 32,2 10 21,21
B 227 220,5 41 32,3 53,7 39,7 46,7 12,6 62,3 51,6 41,6 35,2 37 11 18,06
C 223 215 42,2 31 58,1 37,6 48,5 14 61,8 55,6 40,1 33,6 36,3 12 18,92
D 199 192,5 40 29,8 56,7 36,9 45,8 14,5 59,4 51,4 36,7 30,7 30,1 12 20,10
E 177,2 168,3 31,6 23,7 45,6 29,2 37,2 15 46,6 46 32,6 25,3 28,3 5 17,83
F 189,9 183 32 25 49,9 30,2 39,4 18,4 44,6 43,7 31,7 28,9 16,85
G 199 192 43,2 29,5 53,7 35,5 44,3 17,2 52 50,5 38,3 29,2 31,2 11 21,70
Tabla 5.-Dimensiones (mm) de la l.' F. m y de la 3.' F. m de HippúJion sp. (MBD-P 265-6 Y7) Yotros hippidlformes. Nomenclaturasegún Eisenmann et al. (1988).
1.' F. 111 2 3 4 S 6 7 8 9
MHD·P 265·6 ............................. 76 68,4 45,3 55,8 37,9 51,4 46,9 29 26,2MHN-B 001277 ........................... 85,6 75,2 41,2 57 37,5 48,3 46,7 28,7 44MLP ......................................... 80,4 69,3 36,3 53,4 34,8 44,7 40,8 25,2 41,2TAR-907 ................................... 80,4 72,3 38,7 53,4 35 48,5 47,2 27,7 44MLP 6.342 ................................. 71,2 62 43,3 62,7 37,1 50 43,1 26,7 37,9MLP 6.350 ................................. 73,5 61,6 44,5 62 39,3 50,6 47,4 27,1 36,5401 MCNBR .............................. 70,2 62,1 44,6 59,7 37,1 49,8 46,8 29,9 49
3.' F. 1II
MHD-P 265·7 1 2 3 4 5 6 753,6 56 73 48 32 52 58

460
ción disponible, las dimensiones del cráneo (longitudrostral, longitud basilar, etc.) y de la mandíbula(v. gr.: longitud y alto mandibular, entre otras), asícomo las de las series dentarias superiores e inferiores de nuestro ejemplar, se vinculan con la variaciónde los restos que Moreno (1891) denominó Onohippidium munizi por una parte y, por otra, lo asignadoa Hippidion principale por Boule y Thevenin (1920)de Tarija y Ameghino (1989) de Buenos Aires, ambos de talla muy grande (tabla 1).
Respecto de los elementos postcraneales, el McIlIcorrespondería a un individuo muy grande dentro delgrupo de los de talla media (tabla IV) y la 1.·Fm representa a un ejemplar pequeño entre los de tallagrande conocidos en Sudamérica (tabla V). El McIlIexhibe, por su parte, uno de los valores mayores degracilidad (IG: 21.21) dentro del grupo considerado.Las dimensiones del radio, como la longitud máximay medial, ancho máximo proximal, ancho de la diáfisis, entre otras, lo acercan a una forma bastantegrande dentro de estos animales y que Boule y Thevenin (1920) asignan aH. neogaeus (tabla 111).
De acuerdo con el análisis realizado, un conjuntode caracteres previamente propuestos por autoresprecedentes se han mostrado como criterios válidospara el análisis aquí presentado (cí. Boule y Thevenin, 1920; Hoffstetter, 1952; Winge, 1906, y Zetti,1966).
Conclusiones
El estudio de caracteres morfológicos a nivel cráneo-mandibular de la dentición superior e inferior yde los elementos postcraneales conservados asociados y evaluados, en principio, como pertenecientes aun mismo individuo, permiten determinarlo inequívocamente como Hippidion sp. Se trata de los restosmás completos de este género reportados hasta elmomento para el Pleistoceno de Uruguay y pertenecen a una forma robusta, con extremidades relativamente cortas respecto del cráneo grande. Tanto lasdimensiones del McIlI como las de la l."FIII corresponderían a lo que se puede calificar como un Hippidion de talla grande, con la parte proximal de lasextremidades más larga y la distal proporcionalmente más corta. Por tratarse de los únicos materialesbastante completos procedentes de esta localidad ypor estar llevándose a cabo la revisión del grupo engeneral, creemos conveniente determinarlo comoHippidion sp., de talla grande y «cabezón». Dichogénero queda registrado para sedimentos del nortede Uruguay llamados Fm. Sopas, conteniendo paleomastofauna perteneciente a la Edad-mamífero Lujanense (Pleistoceno superior) y cuyo marco depositacional en el área es un sistema fluvial, estando los res-
M. UBILLA, M. T. ALBERDI
tos analizados en una facie de tipo planicie de inundación.
AGRADECIMIENTOS
El Prof. José Soloviy (MHD Artigas) puso gentilmente el material a nuestra disposición y facilitó las tareas de campo. Los li·cenciados Sergio de Souza y Gerardo Verohslavsky (Dpto. Geología, FHC) nos proporcionaron importante información sedimentológica y valiosos comentarios críticos respectivamente. A todosellos, nuestro sincero reconocimiento.
Referencias
Alberdi, M. T. (1987). La Familia Equidae Gray, 1821(Perissodactyla, Mammalia) en el Pleistoceno de Sudamérica. IV Congr. Latinoamer. Paleont. Bolivia, 1:484-499.
Alberdi, M. T. YPrado, J. L. (1989). Multivariant analysis of remains of the appendicular skeleton oí SouthAmerican equids. Abstracts 5." Intern. TheriologiealCongress, 1: 261-262, Roma.
Alberdi, M. T. YPrado, J. L. (en prensa). A Quantitativestudy of the autopodial remains of South American Hippidion Owen, 1869 (Mammalia, Perissodactyla).
Alberdi, M. T.; Fernández, J.; Menegaz, A., y Prado,J. L. (1986). Hippidion Owen, 1869 (Mammalia, Perissodactyla) en sedimentos del Pleistoceno tardío de la localidad Barrio Negro (Jujuy, Argentina). EstudiosGeol., 42: 487-493.
Alberdi, M. T.; Menegaz, A., y Prado, J. L. (1987). Formas terminales de Hippidion (Mammalia, Perissodactyla) de los yacimientos del Pleistoceno tardío-Holocenode la Patagonia (Argentina, Chile). Estudios Geol., 43:107-115.
Ameghino, F. (1889). Contribución al conocimiento de losmamíferos fósiles de la República Argentina. ActasAead. Nae. Ciene. Córdoba, 6: 1.027 págs.
Antón, D. (1975). Evolución geomorfológíca del norte delUruguay, 1-22. Dir. Suelos y Fertilizantes, MAP, Montevideo.
Beherensmeyer, A. (1988). Vertebrate preservation in fluvial channels. Palaeogeogr. Palaeoclimatol. Palaeoeeol.,63: 183-199.
Bombín, M. (1975). Afinidade paleoecologica, cronologica e estratigrafica do componente de megamamíferos nabiota do Quaternario terminal da Provincia de BuenosAires (Argentina), Uruguai e Rio Grande do Sul (Brasil). Como Mus. Ciene. PUCRGS, Porto Alegre, 9: 1-28.
Bombín, M. (1976). Modelo paleoecologico evolutivo parao neoquaternario da regiao da campanha oeste do RioGrande do Sul (Brasil). A Formacao Tauro Passo, seuconteúdo fossilifero e a pedogenesis posdeposicional.Como Mus. Cien. PUCRGS, Porto Alegre, 15: 1-90.
Bond, M. (1986). Los ungulados fósiles de la Argentina:evolución y paleoambientes. Actas IV Congr. Arg. Paleont., Bioest., 2: 173-185, Mendoza.
Boule, M. y Thevenin, A. (1920). Mammiferes fossiles deTarija. Miss. Scient. Créqui-Montfort et Senechal de laGranje. París, 256 págs.
Burmeister, G. (1875). Los caballos fósiles de la pampa argentina descritos por. .. , 1-88. La Tribuna, Buenos Aires.

HIPPIDlON SP. (MAMMALIA, PERISSODAeIYLA, EQUIDAE) EN SEDIMENTOS DEL PLEISTOCENO... 461
Eisenmann, V. (1979). Etude des cornets des dents incisives inférieures des Equus (Mammalia, Perissodactyla)actuels et fossiles. Palaentographica ltaUca, 71 (n.s.4):55-75.
Eisenmann, V.; Alberdi, M. T.; de Giuli, C., y Staesche,U. (1988). Studying fossil horses. l. Metodology, 1981.En: New York Int. Hipparion Conf., Woodburne y Sondar (eds.), 1-71. Brill.
Hill, A. (1979). Disarticulation and scattering of mammalskeletons. Paleobiology, 5 (3): 261-274.
Hoffstetter, R. (1952). Les mammiferes pléistocenes de laRépublique de L'Equateur. Mem. Soco Geol. Fr. (n.s.),31 (66): 1-391.
Jones, G. (1956). Memoria explicativa y mapa geológicodel Departamento de Canelones. Bol. Inst. Geol. Uruguay, 34: 1-193.
Kidwell, S.; Fursich, F., y Aigner, T. (1986). Conceptualframework for the analysis and c1assification of fossilconcentrations. Palaios, 1: 228-238.
Kraglievich, L. (1928). Catálogo n.O 1 in Berro: Contribución al conocimiento de los fósiles de la RepúblicaOriental del Uruguay. Rev. Soco Amigos Arqueol., 1-74.
Lambert, R. (1941). Estado actual de nuestros conocimientos sobre la Geología de la República Oriental del Uruguay. Bol. Inst. Geol. Uruguay, 29: 1-89.
MacFadden, B. y Azzarolli, A. (1987). Cranium of Equusinsulatus (Mammalia, Equidae) from the Middle Pleistocene of Tarija, Bolivia. J. Vertebrate Paleontology, 7(3): 325-334.
MacFadden, B. y Skinner, M. (1979). Diversification andbiogeography of the one toed horses Onohippidium andHippidion. Postilla, 175: 1-10.
Marshall, L.; Webb, S.; Sepkoski, J. Jr., y Raup, D.(1982). Mammalian evolution and the Great AmericanInterchange. Science, 215 (4538): 1351-1356.
Martínez, S. (1989). Adiciones a la malacofauna de la Formación Sopas (Pleistoceno superior, Uruguay) con algunas consideraciones paleoecológicas. Paula-Coutiana,Porto Alegre, 3: 67-75.
Moreno, F. P. (1891). Onohippidium munizi. Breve noticia sobre los restos fósiles de un género nuevo de la Familia de los Equidae conservados en el Museo de La Plata. Rev. Mus. La Plata, 2: 65-71.
Olazarri, J. (1980). Moluscos de la Formación Sopas, Holoceno del Dpto. de Salto, Uruguay. Como Soco Malac.Uruguay, 5 (39): 301-304.
Perea, D. y Martínez, S. (1989). Nueva localidad fosilíferapara el cuaternario del norte del Uruguay. Bol. SocoZool. Uruguay, Actas II Jom. Zool., 5: 19-20.
Prado, J. (1984a). Fenética de los metatarsianos de taxa fósiles de Hippidion Owen, Onohippidium, Moreno, Parahipparion C. Ameghino y Equus (Amerhippus) Linne(Mammalia, Perissodactyla). CIPF CED Orione Contr.Biol., 11: 11-15, Montevideo.
Prado, J. (1984b). Análisis de los caracteres: un enfoqueaplicado a los metatarsianos de los taxa fósiles Hippidion Owen, Onohippidium Moreno, ParahipparionC. Ameghino y Equus (Amerhippus) Linne (Mammalia,Perissodactyla). 1Jom. Arg. Paleont. Vert. res. La Plata,26.
Prost, M. T. (1982). Heritages quatemaires et evolutiongeomorphologique des bords du Rio de la Plata, en Uruguay. Tesis doctoral, 1: 152-203; 3: 467-540.
Reig, O. (1981). Teoría del origen y desarrollo de la faunade mamíferos de América del Sur. Monographie Naturae, Mar del Plata, 1: 1-162.
Roselli, F. (1938). Apuntes de Geología y Paleontologíauruguayas sobre insectos del Cretácico del Uruguayodescubrimientos de admirables instintos constructivos deesa época. Bol. Soco Amigos Cienc. Nat. KraglievichFontana, Nueva Palmira, 1 (2): 29-102.
Teisseire, A. (1928). Contribución al estudio de la geología y paleontología de la República Oriental del Uruguay. Región de Colonia. An. Univ., 37 (122): 319-469.
Ubilla, M. (1983). Sobre la presencia de tapires fósiles enel Uruguay (Mammalia, Perissodactyla, Tapiridae). Rev.Fac. Hum. Cienc., Ciene. Tierra, Montevideo, 1 (3):85-104.
Ubilla, M. (1986). Mamíferos fósiles, geocronología y paleoecología de la Fm. Sopas (Pleistoceno sup.) del Uruguay. Ameghiniana, 22 (3-4): 185-196.
Ubilla, M. y Martínez, S. (1988). El cuaternario continental del norte del Uruguay, una aproximación a su geocronología y reconstrucción paleoambiental. Proce.IGCP-20I, Mérida, Venezuela meeting: 101-110.
Webb, D. (1978). A history of savanna vertebrate in thenew world. Part 11: South America and Great Interchange. Ann. Rev. Ecol. Syst., 9: 393-426.
Winge, H. (1906). Jordfundne og nulevende Hovdyr (Ungulata) fra Lagoa Santa. Minas Geraes, Brasilien. E.Museo Lundii, 3 (1): 1-239.
Zetti, J. (1966). Perissodaetyla. En: Paleontografía Bon_erense, Borrello (ed.), Prov. Buenos Aires, IV: 188-193,La Plata.
Recibido el 21 de noviembre de 1990Aceptado el 9 de diciembre de 1990'
462 M. UBILLA. M. T. ALBERDI
CD
10 cm
"I
B
8cm
Lám. l.-Hippidion sp. A: MHD·P 265·1. cráneo. norma palatal; B: MHD·P 265·2. mandíbula.
HIPPIDION SP. (MAMMALJA, PERISSODACrYu... EOUIDAE) EN SEDIMENTOS DEL PLEISTOCENO... 463
..n3
I B
l~
Um. 2.-Hippidion sp. A: MHD-P US-!, serie molar-premolar superior i%quierda. B: MUD-P 265-2. serie molarpremolar inferior derecha e izquierda.

464 M. UBILLA, M. T. ALBERDI
. \
5cm
5cm
Um. 3.-Hippidion sp. A: MHD·P 265-4, cubito-radio; B: MHO-P 265-5. McIlI derecho. vista anterior: C: MHD·P265·6, 1,'FIII YMHD-P 265-7, 3.'FIII.





























