Embed Size (px)
Citation preview

Histochemical Investigation of the Modal Specificity of Taste
BRUCE TREFZSchool of Dentistry, University of Michigan, Ann Arbor, Michigan 48104, USA
The taste mechanism was investigated in aprimate (Macaca mulatta). Based on thehypothesis that intracellular enzymes con-tribute to the transduction of tastes to elec-tric impulses by taste cells, a histochemicalsurvey of the activity of several enzymeswas made on taste buds from regions ofthe mouth associated with sweet, salt, sour,and bitter tastes. Considerable differenceswere noted among the modalities, whichconfirmed the hypothesis. An exclusivelybitter enzyme was identified.
The sense of taste is unique among the"chemical" senses, in that it is associatedwith discrete receptor organs of non-neuralderivation, the taste buds. Their existencein humans has been recognized for morethan a hundred years,' but their exact modeof function remains obscure.
Classically, taste has been subdivided intofour modalities, sweet, salt, bitter, and sour(acid). Each of these modalities has beenlocalized to a discrete region on the surfaceof the tongue: sweet, to the fungiform papil-lae of the tip of the dorsum; salt, to thefungiform papillae of the lateral aspects ofthe anterior two thirds of the dorsum;bitter, to the circumvallate papillae of theposterior third of the dorsum; and sour,to the foliate papillae of the lateral aspectsof the posetrior third of the dorsum (Fig).2In 1951, Baradi and Bourne3 proposed
a theoretical taste mechanism wherein thesubstance being tasted either inhibited oraccelerated the rate of an on-going bio-chemical reaction in a taste region by alter-
This study was supported by funds from under-graduate research fellowships awarded in 1970 and 1971by the School of Dentistry, University of Michigan andby USPHS Research Grant DE-01604 from the NationalInstitute of Dental Research, National Institutes ofHealth, Bethesda, Md.Hatton award: 1st place, predoctoral division.
ing the efficacy of the enzyme that catalyzedthe reaction. A net change in a reaction,then, was the source of the taste impulse,and the fact that enzymes are highly spe-cific accounted for the specificity of thetaste impulse. If this hypothesis is valid, thetaste buds from a region of predominantlyone modality should display an enzymecomplement different from that of a regionof another modality.More recent research4 has shown that
taste cells have differing enzyme comple-ments at different stages in their fetal devel-opment; perhaps this reflects the developing,and therefore changing, level of function.
It is not unlikely that taste buds of dif-fering modalities also have differing enzy-matic compositions, although currently thisis a topic of conjecture.The enzyme complements of the taste
buds of the four classical modalities wereassayed histochemically, and their speci-ficities, vocalizations, and intensities weredetermined.
Materials and MethodsAdult rhesus monkeys (Macaca mulatta)
were used. All monkeys were killed by vas-cular perfusion of physiologic isosmoticsaline while they were under anesthesia.Tongues were removed by incision posteriorto the circumvallate papillae. Immediatelyafter excision, regions containing taste budswere removed from well within the bound-aries of the areas classically assigned toeach of the modalities (Fig 2). The tissuethen was frozen in liquid nitrogen-isopen-tane, sectioned at 12 micrometers on acryostat,* and mounted on microslides.With each modality, the tissue blocks wereoriented so that the microtome cut the taste
* American Optical "Cryo-Cut," American OpticalCo., Instrument Division, Buffalo, NY.
1203

Sour
10 - 7 7 Salt
h ~ ~~~I __ n\ ~~~I- -
I ,Qo ,
9 s ;vI
Sweet
FIG 1.-Classical taste modality localizationon the tongue. FIG 2.-Excision site for modality assays.
FIG 3.-Alkaline phosphatase. Sweet (SW) and salt (SA) displayed activity throughout tastebuds (arrows), but sour (SO) and bitter (BI) stained only lightly and principally in the regionof the taste pore (square) where the oral fluids contact the taste cells (orig mag X200, repro-duced at 56%).
1204

HISTOCHEMISTRY OF TASTE 1205
TABLEENZYMATIC ACTIVITY IN TASTE BUDS
Enzyme Sweet Salt Sour Bitter
Succinic dehydrogenaseCholine oxidaseCytochrome oxidaseNonspecific esteraseNAD diaphoraseNADP diaphoraseu-Glycerophosphate
dehydrogenaseP-Hydroxybutyric
dehydrogenaseD-amino acid oxidaseAlkaline phosphataseGlucose-6-phosphate
dehydrogenaseAcid phosphataseAdenosine triphosphataseNucleotide diphosphatase
+++±±+
+
++
A+
-4-_- --4-++
+ + +_ + A-+±±± +±±± +++±+ ±+± ++++
11 A-± A--++++ ++++±++
+± ±±±±++ ~+
±nd
buds in the long axis to facilitate compari-son.The following are the enzyme systems
examined and the pathways they represent:succinic dehydrogenase5 (Krebs cycle ac-
tivity); choline oxidasej) cytochrome oxi-dase,7 ct-glycerophosphate dehydrogenase 8
3-hydroxybutyric dehydrogenase,9 and D-amino acid oxidase"i (electron transportchain activity); nonspecific esterase&" (mem-
FIG 4.-Acid phosphatase. Sour (SO) and bitter (B!) taste buds stained darkly (great activity)and sweet (SW) and salt (SA) buds showed only minimal activity, also in the region of the pore(orig mag X200, reproduced at 58%).
Vol 51 1972
i.
I
J TDent Res Sitpplemnent to No. S
:S-tW., A SA
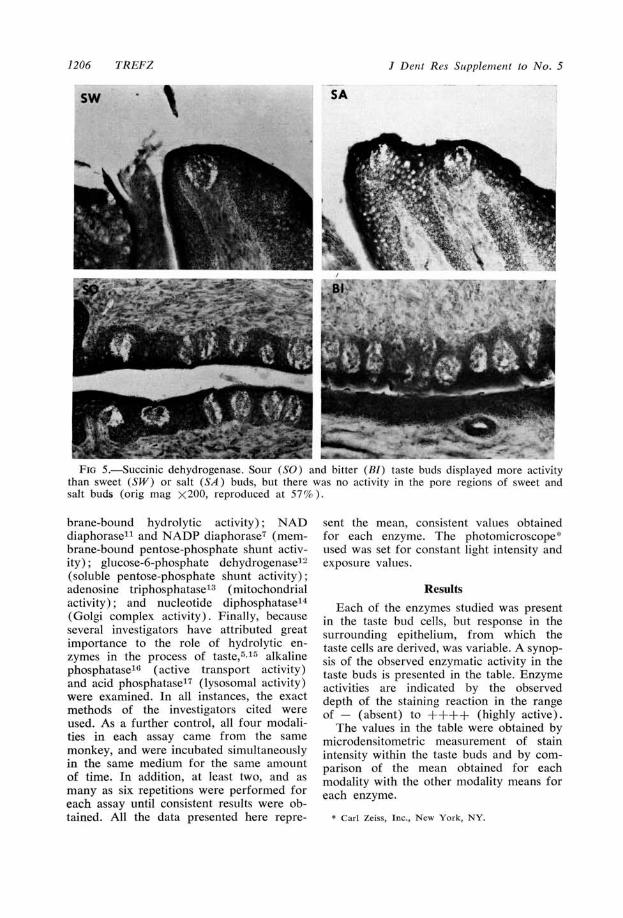
FIG 5. Succinic dehydrogenase. Sour (SO) and bitter (Bi) taste buds displayed more activitythan sweet (SW) or salt (SA) buds, but there was no activity in the pore regions of sweet andsalt buds (ofig mag X200, reproduced at 57% ).
brane-bound hydrolytic activity); NADdiaphorase11 and NADP diaphoraseT (mem-brane-bound pentose-phosphate shunt activ-ity); glucose-6-phosphate dehydrogenase12(soluble pentose-phosphate shunt activity);adenosine triphosphatase"1 (mitochondrialactivity); and nucleotide diphosphatasel1(Golgi complex activity). Finally, becauseseveral investigators have attributed greatimportance to the role of hydrolytic en-zymes in the process of taster5s15 alkalinephosphatasel6 (active transport activity)and acid phosphatase17 (lysosomal activity)were examined. In all instances, the exactmethods of the investigators cited wereused. As a further control, all four modali-ties in each assay came from the samemonkey, and were incubated simultaneouslyin the same medium for the same amountof time. In addition, at least two, and asmany as six repetitions were performed foreach assay until consistent results were ob-tained. All the data presented here repre-
sent the mean, consistent values obtainedfor each enzyme. The photomicroscope-Iused was set for constant light intensity andexposure values.
Results
Each of the enzymes studied was presentin the taste bud cells, but response in thesurrounding epithelium, from which thetaste cells are derived, was variable. A. synop-sis of the observed enzymatic activity in thetaste buds is presented in the table. Enzymeactivities are indicated by the observeddepth of the staining reaction in the rangeof (absent) to ++++ (highly active).The values in the table were obtained by
microdensitometric measurement of stainintensity within the taste buds and by com-parison of the mean obtained for eachmodality with the other modality means foreach enzyme.
* Carl Zeiss, Inc., Ncw York, NY.
1206 TREFZ

HISTOCHEMISTRY OF TASTE 1207
SW SA
B':
FIG 6.-Beta-hydroxybutyric dehydrogenase. Sweet (SW), salt (SA), and sour (SO) tastebuds displayed little activity, but the bitter taste buds (Bi) showed considerable activity, espe-cially in the surrounding epithelium (orig mag x 200, reproduced at 56% ).
Alkaline phosphatase (Fig 3) reactedstrongly in the sweet and salt modalities,and reacted hardly at all in the bitter andsour modalities. All activity seemed to beconcentrated in the region of the taste porein the sour and bitter buds, whereas thesweet and salt buds stained rather generallyand very darkly. This situation is reversedin the acid phosphatase reaction (Fig 4);sour and bitter stained heavily throughout,and sweet and salt showed only slight ac-tivity, which is concentrated in the regionof the pore. The characteristic staining ofthe capillary endothelium by the alkalinephosphatase reaction is evident and uni-from through the different modalities.The activity of succinic dehydrogenase
(Fig 5) seemed to parallel that of acidphosphatase, with the exception of the poreregions of the sweet and salt buds, whichdisplayed no special activity.
Beta-hydroxybuItyric dehydrogenase (Fig
6) appeared to be specific for the bittermodality; all the other modalities displayedno activity. The reaction in the surroundingepithelium was especially notable.NADP diaphorase (Fig 7) displayed a
spectrum of activity that can be graded asleast in sweet, progressively more in saltand sour, and most in bitter.Alpha- glycerophosphate dehydrogenase
(Fig 8) showed its strongest activity in thesour and bitter modalities, with heavy epi-thelial activity around the taste buds. Thisis especially apparent in the bitter (circum-vallate papilla) region, where the heavilystained gustatory epithelium opposed a non-neural epithelium that showed essentiallyno activity.
Other enzymes studied, but not photo-graphed, showed measurable but somewhatless striking results (Table): Cytochromeoxidase, D-amino acid oxidase, and glucose-6-phosphate dehydrogenase displayed fairly
Vol 51 1972

J Dent Res Supplement to No. 5
...SA
foFIG 7. NAPD diaphorase. Taste bud enzyme activity was graded from least to most in theblowing order: sweet (SW), salt (SA), sour (SO), and bitter (BI) (orig mag X200, repro-
duced at 56%o).
even staining from modality to modality.The same is true for adenosine triphosphateand nucleotide diphosphatase, but sharpintracellular differences were apparent.Adenosine triphosphatase stained the cellmembranes of all four modalities sharply,but the pore regions in the sour modality,and especially the bitter modality, displayedintense activity. Nucleotide diphosphatase,however, darkly stained the nuclei of onlysome of the cells of the buds of all fourmodalities and the pore regions in all mo-dalities except salt. NAD diaphorase had agraded spectrum of activity that was simi-lar, but not identical to that of NADPdiaphorase. Choline oxidase and nonspecificesterase showed activity in all the modalitiesexcept salt; greater differences were notedwith choline oxidase.
DiscussionSuccinic dehydrogenase has long been
used as a marker for the activity of the
Krebs cycle,5 especially in the taste buds.'8Its localization in significant amounts insour and bitter taste buds, but not in sweetor salt buds, infers that in the former twomodalities the missing electrochemical im-pulse conversion system of Baradi andBourneA3'19 is in some way linked to theKrebs tricarboxylic acid cycle.
High acid phosphatase activity in sourand bitter taste buds and low activity insweet and salt buds is to be expected, be-cause most sour and bitter substances areacidic to some degree,20 and lysosomes,which contain the enzyme, have been ob-served in ultrastructural studies of foliateand circUmvallate papillae in lower spe-cies.21t22 In addition, it is a lytic phosphate-cleaving enzyme that lends itself to energymediation.2' This fact also is true of alka-line phosphatase, which is sweet- and salt-specific; but, in addition, alkaline phospha-tase is an energy-mediating enzyme oftenassociated with active transport at the cell-
1208 TREFNZ

HISTOCHEMISTRY OF TASTE 1209
.SW SA
B!'
FIG 8. Alpha-glycerophosphate dehydrogenase. More activity was evident in sour (SO) andbitter (BI) taste buds than in sweet (SW) or salt (SA) buds. Notable activity in epithelium sur-rounding sour and bitter taste buds was present (orig mag X200, reproduced at 56%).
membrane level,24 as evidenced by its pre-viously noted activity in capillary endothe-liums. Alkaline phosphatase may be theelectrochemical converter for sweet andsalty tastes.
Choline oxidase, ux-glycerophosphate de-hydrogenase, 3-hydroxybutyric dehydroge-nase, and D-amino acid oxidase are indi-cators of the so called electron-transportchain,25- another cellular energy path. Thevarying results observed in the four mo-dalities for these four enzymes points con-vincingly toward enzymatic specificity asthe determining factor in taste specificity.In addition, P-hydroxybutyric dehydrogenasemay well be a true "bitter enzyme."The other enzymes assayed all showed
varying differences both among themselvesand among the four modalities. All werelinked to either a metabolic pathway (NADand NADP diaphorase, membrane-boundpentose phosphate shunt26 and glucose-6-
phosphate dehydrogenase, free cytoplasmicpentose phosphate shunt27), or to a biologicfunction associated with a discrete' ultra-structural entity (adenosine triphosphatase,mitochondria"; nucleotide diphosphatase,Golgi apparatus,28 and nonspecific esterase,ribosomes and rough-surfaced endoplasmicreticulum29).
This wide variation of enzymatic activity,and wide variation of kinds and types ofenzymes- involved, easily explains why weare capable of more than 16 tastes (themaximum nUmber of combinations and per-mutations of the four basic taste modalities).This study only considered a fraction of theenzymes that occur in cells, but variationswere found from modality to modality inalmost every enzyme studied. One notableexception is cytochrome oxidase, which isan indicator of anaerobic metabolic activ-ity.30 As expected, the activity is relativelyconstant from cell to cell; this may imply
[70 1 51 1972

1210 TREFZ
that the ultimate transmission (or transduc-tion or both) of the taste signal is by an
aerobic pathway, rather than by an anaero-
bic, pathway.An overall survey of the data in the table
indicates the following generalized sub-grouping, on the basis of enzyme activities;sweet with salt and sour with bitter. Thismay be of developmental significance, sincesweet and salt taste buds receive their in-nervation from the VII nerve, and sour andbitter taste buds from the IX and X nerves
(Fig 1), which supply structures derivedfrom different bronchial arches.
This fact is of medical interest, becausediminutions in taste acuity of these modalsubgroups that occur in certain human dis-eases have been noted and are well-docu-mented.3' Sweet functions with salt, andsour with bitter, and the two subunits oper-
ate quite independently of each other dur-ing the course of therapy.
ConclusionsThe mechanism of taste was studied his-
tochemically in a primate. Notable differ-ences in enzymatic activity were demon-strated among taste buds from tongue re-
gions associated with sweet, salt, sour, andbitter taste. The hypothesis of Baradi andBourne32 that the specificity and transduc-tion of the taste impulse is due to specificenzymatic involvement was confirmed.Functional association was noted betweensweet and salt enzymatic activity, and be-tween sour and bitter enzymatic activity;this agrees with clinical findings. A bitter-specific enzyme, 3-hydroxybutyric dehydro-genase, was identified.
The author thanks his sponsor, J. K. Avery, ofthe Laboratory of Oral Histology, School of Dentistry,University of Michigan for encouragement and adviceand C. Cox, of the same institution, for his aid in thetechnical aspects of this study.
References1. LOVEN, O.C.: Bidrag till Kannendomenom tungans smakpapiller, Med Arch(Stockholm) 3(9):1, 1867.
2. ADEY, W.R.: The Sense of Smell, inFIELD, J.; MAGOUN, H.W.; and HALL, V.E.(eds), Handbook of Physiology, vol I,
Washington: American Physiological So-ciety, 1959, p 481.
3. BARADI, A.F., and BOURNE, G.H.: Locali-zation of Gustatory and Olfactory En-
J Dent Res Supplement to No. 5
zymes in the Rabbit, and the Problem ofTaste and Smell, Nature (Lond) 168:977, 1951.
4. NADA, O., and IWAYAMA, T.: Histochemi-cal Observation on the Phosphatase Ac-tivities of the Differentiating Taste Bud,Anat Rec 165(1):61, 1969.
5. DEFENDI, V., and PEARSON, B.: Quantita-tive Estimation of Succinic DehydrogenaseActivity in a Single Microscopic Tissue Sec-tion, J Histochem Cytochem 3:61, 1955.
6. FARBER, E.; STERNBERG, W.H.; and PEARCE,N.A.M.: Histochemical Localization ofCholine Oxidase and D-Amino Acid Oxi-dase with Tetrazolium Salts and PhenozineMethosulfate, J Histochem Cytochem 6:389, 1958.
7. ALTMANN, F.P.: The Cellular Chemistryof Certain Cytoplasmic Oxidative Enzymesand Their Relevance to the Metabolism ofCancers, PhD thesis, University of London,1968.
8. Biochemists' Handbook, Long, C. (ed),London: Spon, 1961, p 368.
9. HESS, R.: Beta-Hydroxybutyric Dehydro-genase Activity in Tumors, Am J Pathol37:583, 1960.
10. BURSTONE, M.S.: The Relationship BetweenFixation and Techniques for the Histo-chemical Localization of Hydrolytic en-zymes, J Histochem Cytochem 6:322, 1958.
11. DIXON, M., and WEBB, E.C.: Enzymes, 2nded, London: Longmans, 1964, p 233.
12. CHIQUOINE, A.D.: The Distribution of Glu-cose-6-Phosphatase in the Liver and Kidneyof the Mouse, J Histochem Cytochem 1:429, 1953.
13. WACHSTEIN, M., and MEISEL, E.: Histo-chemistry of Substrate-Specific Phospha-tases at a Physiological pH, J HistochemCytochem 4:424, 1956.
14. NOVIKOFF, A., and GOLDFISCHER, S.: Nu-cleotide Diphosphatase Activity in theGolgi Apparatus, Proc Natl Acad Sci 47:802, 1961.
15. BOURNE, G.H.: Alkaline Phosphatase inTaste Buds and Nasal Mucosa, Nature(Lond) 161:444, 1948.
16. BURSTONE, M.S.: Histochemical Demon-stration of Acid Phosphatases with Naph-thol AS-Phosphates, J Natl Cancer Inst 19:523, 1958.
17. GOMORI, G.: Microscopic Histochemistry,Principles and Practice, Chicago: Univer-sity of Chicago Press, 1952, p 193.
18. PEVZNER, R.A.: Determination of the Suc-cinic Dehydrogenase Activity in Taste Budsof Some Vertebrate Mammals, Dokl AkadNauk SSSR 155:191, 1964.
19. BARADI, A.F., and BOURNE, G.H.: NewObservations on the Alkaline Phosphatase

HISTOCHEMISTRY OF TASTE 1211
Reaction in the Papilla Foliata, J BiophysBiochem Cytol 5:173, 1959.
20. VON BEKESY, G.: The Duplexity Theoryof Taste, Science 145:834, 1964.
21. FARBMAN, A.I.: Fine Structure of the TasteBud, J Ultrastruct Res 12:328, 1965.
22. NEMETSCHEK-GANSLER, H., and FERNER,H.: On the Ultrastructure of the TasteBuds, Z Zellforsch Mikrosk Anat 63:155,1964.
23. DEDUVE, C.: Lysosomes, a New Group ofCytoplasmic Particles, Hayashi, T. (ed):Subcellular Particles, New York: Ronald,1959.
24. DANIELLI, J.F.: Cytochemistry: A CriticalApproach, London: Chapman and Hall,1953, p 81.
25. FARBER, E.: Principles of Oxidative EnzymeHistochemistry, in Histochemistry, KansasCity: University of Kansas Press, 1964,p 11.
26. BURSTONE, M.S.: Histochemical Demon-stration of Cytochrome Oxidase with New
Amine Reagents, J Histochem Cytochem8:63, 1960.
27. ALTMANN, F.P.: The Retention and Esti-mation of "Soluble" Dehydrogenases in Un-fixed Tissue Sections, Biochem J 96:53,1965.
28. NOVIKOFF, A.; GOLDFISCHER, S.; and Ess-NER, E.: The Importance of Fixation in aCytochemical Method for the Golgi Ap-paratus, J Histochem Cytochem 9:459,1961.
29. UNDERHAY, E.; HOLT, S.J.; BEAUFAY, H.;and DEDUGE, C.: Intracellular Localizationof Esterase in Rat Liver, J Biophys Bio-chem Cytol 2:635, 1956.
30. WEST, E.S., and TODD, W.R.: Textbook ofBiochemistry, New York: Macmillan, 1961,p 853.
31. HENKIN, R.I.; GRAZIADEI, P.P.; and BRAD-LEY, B.F.: NIH Clinical Staff Conference:The Molecular Basis of Taste and Its Dis-orders, Ann Intern Med 71:791, 1969.
32. BARADI, A.F., and BOURNE, G.H.: Theoryof Taste and Odors, Science 113:660, 1951.
Vol 51 1972





























