Embed Size (px)
Citation preview

HOST MECHANISMS WHICH ACT TO REMOVE BACTERIA FROM THEBLOOD STREAM
DAVID E. ROGERS
Department of Medicine, Vanderbilt University School of Medicine, Nashville, Tennessee
The body possesses remarkably efficient mech-anisms for sterilizing the blood stream. Contraryto popular belief, most microorganisms are lesscapable of provoking disease when injected in-travenouslty than when admi'nistered by an'y otherroute (1, 2). With rare exceptions, living bacteriawhich enter the blood stream of animals or mandisappear swiftly from the circulation.
In the 73 years which have elapsed since theinitial observations of Wyssokowitsch (3), theevents which follow intravascular injections ofbacteria have received extensive investigation.Studies on the intravascular behavior of differentmicroorganisms or particulate substances whichmay mimic bacteria in their initial host manage-ment in the circulation have aided in clarifyingcertain aspects of the blood stream clearanceprocess. Although bacteremia can be profoundlymodified by humoral immunity, initial hostmechanisms acting to clear the blood stream areprobably not dependent on prior experience withthe invading microorganism. It is thus proper thatthe ways in which animals and man handle bac-teria which enter the blood stream be included inthis symposium on nonspecific host resistance.
It is the purpose of this paper to review what isknown of the dynamics of blood stream clearance,the host tissues which participate in the removalof circulating microorganisms, the fate of bacterialodged in organs which retain them, and the cir-cumstances which may augment, or interferewith, host mechanisms dealing with bacteriawhich invade the blood stream.
CHARACTERISTICS OF BLOOD STREAM CLEARANCE
AS DETERMINED BY PERIPHERAL BLOOD
CULTURES
Although the pattern of clearance varies withthe particular animal species and the particularmicroorganism under study, certain general char-acteristics of the removal process may serve toorient a more detailed commentary on hostmechanisms at work during bacteremia.When large numbers of living bacteria are rap-
idly injected into the veins of an experimentalanimal, quantitative blood cultures reveal a pre-
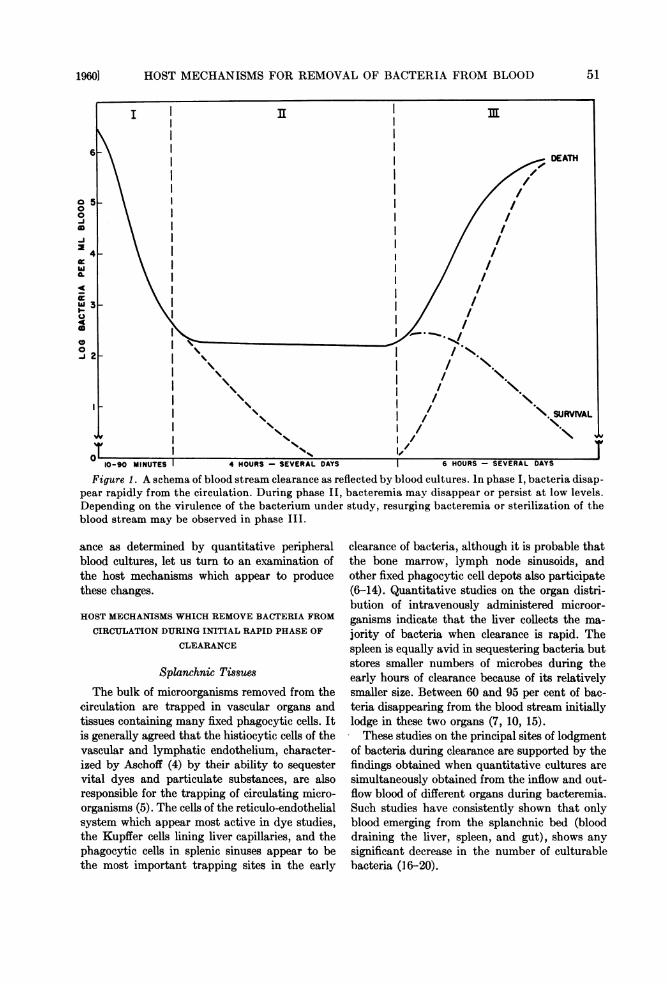
dictable series of events which can be arbitrarilyconsidered in three phases (figure 1).
In the first phase, clearance is rapid. Cultur-able bacteria'.disappear swiftly from the bloodstream during the first 10 min to 5 hr. When theresults of quantitative blood cultures are graphedlogarithmically against time, initial clearancerates are a straight line function and quite con-stant for the microbial parasite under study. Arather constant fraction of circulating bacteriapresent at any sampling interval is removed intransit through the circulatory system. Ninety to99.9 per cent of circulating bacteria disappearduring this period. This initial removal processappears essentially independent of the nature ofthe microbial parasite, the animal under study, orthe subsequent outcome of infection.
This phase of dramatic disappearance of cir-culating bacteria is followed by a phase in whichmicroorganisms persist in the circulation at lowerconcentrations, or by their more gradual removalat slower rates. There may be an abrupt cessationof clearance, resulting in a relatively static lowgrade bacteremia, or clearance rates may slowprogressively, depending on the animal speciesand particular microorganism under study. Per-sisting or slowly declining bacteremia may con-tinue for several hours or several days.
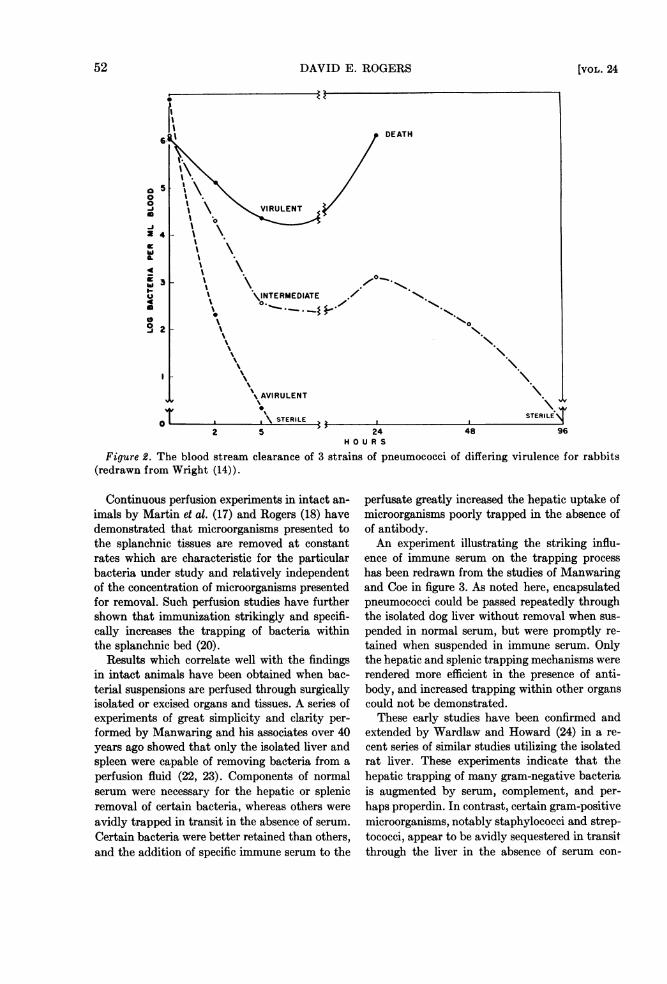
Subsequent events appear to relate to the even-tual outcome of infection. (The word "determine"will be avoided because it seems probable that theoutcome of the bacteremic state is settled prior tothis period). During this late phase, there may bea temporary rise in circulating bacteria, but themajority of microorganisms disappear completelyfrom the blood stream. Bacteria producing fatalinfections reappear in the peripheral blood inincreasing numbers, and bacteremia rises to highlevels which persist until the death of the animal.That the virulence of the microbe governs thelater blood stream findings was beautifully il-lustrated in the early studies of Wright (14). Thebacteremias produced by three strains of pneumo-cocci of varying pathogenicity for rabbits are re-drawn from Wright's studies in figure 2.With this simplified characterization of clear-
50
HOST MECHANISMS FOR REMOVAL OF BACTERIA FROM BLOOD
a00-Jo
-1
ma4L
ma
4ca
0I
I,~~ RIVAL{
10-90 MINUTES 4 HOR -
1090MIUTS HUR SEVERAL DAYS 6 HOURS -SEVERAL DAYS
Figure 1. A schema of blood stream clearance as reflected by blood cultures. In phase I, bacteria disap-pear rapidly from the circulation. During phase II, bacteremia may disappear or persist at low levels.Depending on the virulence of the bacterium under study, resurging bacteremia or sterilization of theblood stream may be observed in phase III.
ance as determined by quantitative peripheralblood cultures, let us turn to an examination ofthe host mechanisms which appear to producethese changes.
HOST MECHANISMS WHICH REMOVE BACTERIA FROM
CIRCULATION DURING INITIAL RAPID PHASE OF
CLEARANCE
Splanchnic TissuesThe bulk of microorganisms removed from the
circulation are trapped in vascular organs andtissues containing many fixed phagocytic cells. Itis generally agreed that the histiocytic cells of thevascular and lymphatic endothelium, character-ized by Aschoff (4) by their ability to sequestervital dyes and particulate substances, are alsoresponsible for the trapping of circulating micro-organisms (5). The cells of the reticulo-endothelialsystem which appear most active in dye studies,the Kupffer cells lining liver capillaries, and thephagocytic cells in splenic sinuses appear to bethe most important trapping sites in the early
clearance of bacteria, although it is probable thatthe bone marrow, lymph node sinusoids, andother fixed phagocytic cell depots also participate(6-14). Quantitative studies on the organ distri-bution of intravenously administered microor-ganisms indicate that the liver collects the ma-jority of bacteria when clearance is rapid. Thespleen is equally avid in sequestering bacteria butstores smaller numbers of microbes during theearly hours of clearance because of its relativelysmaller size. Between 60 and 95 per cent of bac-teria disappearing from the blood stream initiallylodge in these two organs (7, 10, 15).
These studies on the principal sites of lodgmentof bacteria during clearance are supported by thefindings obtained when quantitative cultures aresimultaneously obtained from the inflow and out-flow blood of different organs during bacteremia.Such studies have consistently shown that onlyblood emerging from the splanchnic bed (blooddraining the liver, spleen, and gut), shows anysignificant decrease in the number of culturablebacteria (16-20).
1960] 51
DAVID E. ROGERS
i
I6I'
aZ 000-IS
-ie 4
hiaI-
14
92
DEATH
VIRULENT
\INTERMEDIATE X/ N.\ _ A I,-0
vAV
O L
X AVIRULENT
, \ STERILE >
2 5 24HOU R S
Figure 2. The blood stream clearance of 3 strains of pneumococci of(redrawn from Wright (14)).
Continuous perfusion experiments in intact an-imals by Martin et al. (17) and Rogers (18) havedemonstrated that microorganisms presented tothe splanchnic tissues are removed at constantrates which are characteristic for the particularbacteria under study and relatively independentof the concentration of microorganisms presentedfor removal. Such perfusion studies have furthershown that immunization strikingly and specifi-cally increases the trapping of bacteria withinthe splanchnic bed (20).
Results which correlate well with the findingsin intact animals have been obtained when bac-terial suspensions are perfused through surgicallyisolated or excised organs and tissues. A series ofexperiments of great simplicity and clarity per-formed by Manwaring and his associates over 40years ago showed that only the isolated liver andspleen were capable of removing bacteria from aperfusion fluid (22, 23). Components of normalserum were necessary for the hepatic or splenicremoval of certain bacteria, whereas others wereavidly trapped in transit in the absence of serum.Certain bacteria were better retained than others,and the addition of specific immune serum to the
48
STERILE
96
differing virulence for rabbits
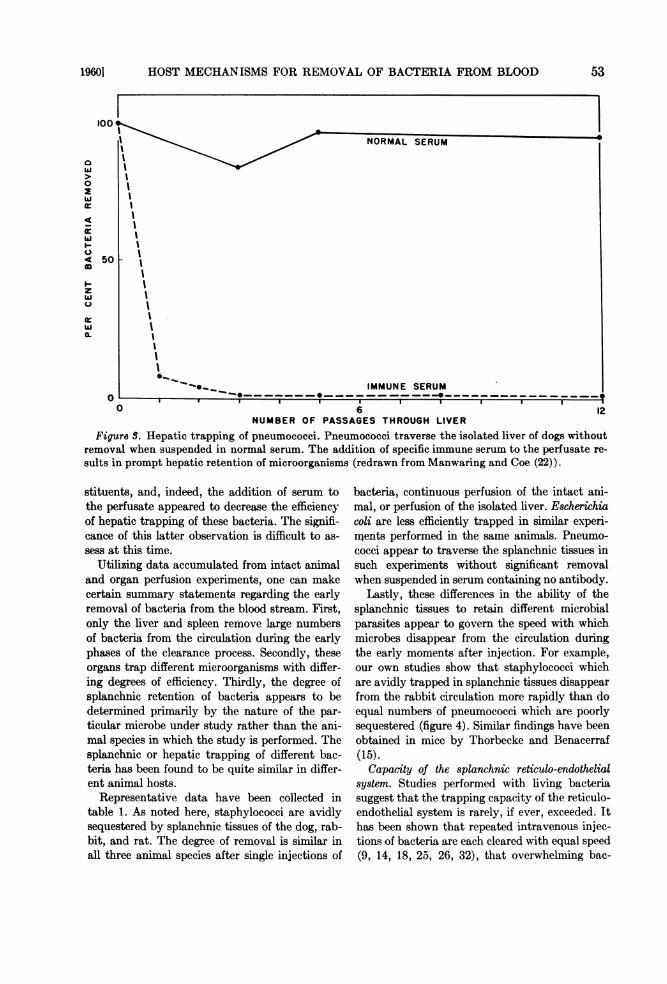
perfusate greatly increased the hepatic uptake ofmicroorganisms poorly trapped in the absence ofof antibody.An experiment illustrating the striking influ-
ence of immune serum on the trapping processhas been redrawn from the studies of Manwaringand Coe in figure 3. As noted here, encapsulatedpneumococci could be passed repeatedly throughthe isolated dog liver without removal when sus-pended in normal serum, but were promptly re-tained when suspended in immune serum. Onlythe hepatic and splenic trapping mechanisms wererendered more efficient in the presence of anti-body, and increased trapping within other organscould not be demonstrated.These early studies have been confirmed and
extended by Wardlaw and Howard (24) in a re-cent series of similar studies utilizing the isolatedrat liver. These experiments indicate that thehepatic trapping of many gram-negative bacteriais augmented by serum, complement, and per-haps properdin. In contrast, certain gram-positivemicroorganisms, notably staphylococci and strep-tococci, appear to be avidly sequestered in transitthrough the liver in the absence of serum con-
52 [VOL. 24
hr
HOST MECHANISMS FOR REMOVAL OF BACTERIA FROM BLOOD
I00
0
0
49
I.-
z
C)
50
0
0 6 12
NUMBER OF PASSAGES THROUGH LIVER
Figure S. Hepatic trapping of pneumococci. Pneumococci traverse the isolated liver of dogs withoutremoval when suspended in normal serum. The addition of specific immune serum to the perfusate re-
sults in prompt hepatic retention of microorganisms (redrawn from Manwaring and Coe (22)).
stituents, and, indeed, the addition of serum tothe perfusate appeared to decrease the efficiencyof hepatic trapping of these bacteria. The signifi-cance of this latter observation is difficult to as-
sess at this time.Utilizing data accumulated from intact animal
and organ perfusion experiments, one can makecertain summary statements regarding the earlyremoval of bacteria from the blood stream. First,only the liver and spleen remove large numbersof bacteria from the circulation during the earlyphases of the clearance process. Secondly, theseorgans trap different microorganisms with differ-ing degrees of efficiency. Thirdly, the degree ofsplanchnic retention of bacteria appears to bedetermined primarily by the nature of the par-
ticular microbe under study rather than the ani-mal species in which the study is performed. Thesplanchnic or hepatic trapping of different bac-teria has been found to be quite similar in differ-ent animal hosts.
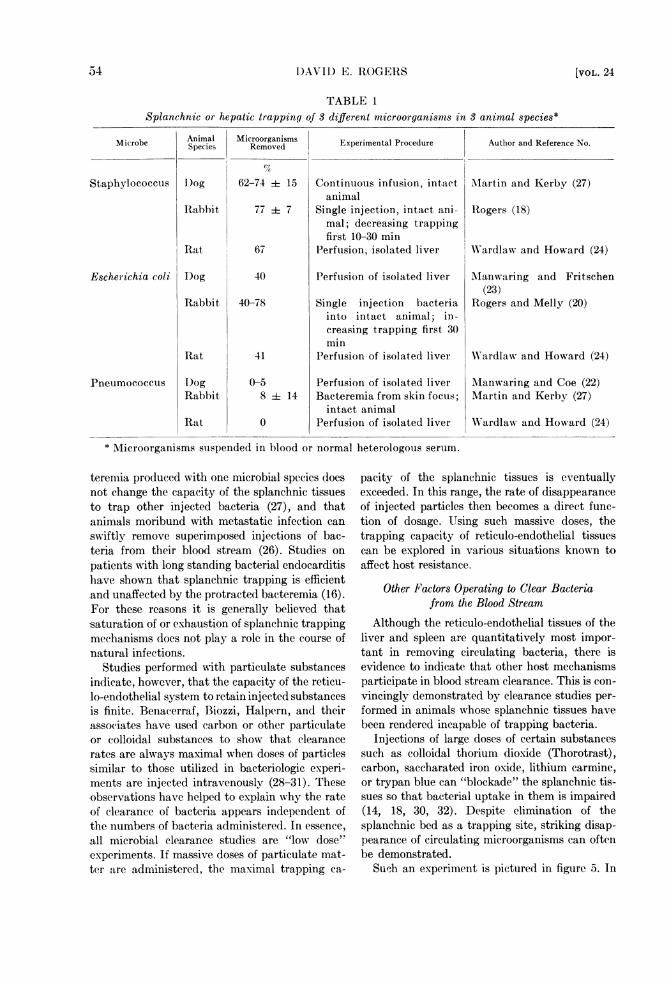
Representative data have been collected intable 1. As noted here, staphylococci are avidlysequestered by splanchnic tissues of the dog, rab-bit, and rat. The degree of removal is similar inall three animal species after single injections of
bacteria, continuous perfusion of the intact ani-mal, or perfusion of the isolated liver. Escherichiacoli are less efficiently trapped in similar experi-ments performed in the same animals. Pneumo-cocci appear to traverse the splanchnic tissues insuch experiments without significant removalwhen suspended in serum containing no antibody.
Lastly, these differences in the ability of thesplanchnic tissues to retain different microbialparasites appear to govern the speed with whichmicrobes disappear from the circulation duringthe early moments after injection. For example,our own studies show that staphylococci whichare avidly trapped in splanchnic tissues disappearfrom the rabbit circulation more rapidly than doequal numbers of pneumococci which are poorlysequestered (figure 4). Similar findings have beenobtained in mice by Thorbecke and Benacerraf(15).
Capacity of the splanchnic reticulo-endothelialsystem. Studies performed with living bacteriasuggest that the trapping capacity of the reticulo-endothelial system is rarely, if ever, exceeded. Ithas been shown that repeated intravenous injec-tions of bacteria are each cleared with equal speed(9, 14, 18, 25, 26, 32), that overwhelming bac-
I~~~~~~~~~~~~~~~~~~~~~~~~~
NORMAL SERUM
IIII
II
--a__ ~~~~IMMUNE SERUM.I__ _ _ _ _ _ _ __.
II . . . . .
19601 53
I
II
1
l)AVID E. ROGERS
TABLE 1Splanchnic or hepatic trapping of 3 different microorganisms in 3 animal species*
Microbe Animal Microorgandsms Experimental Procedure Author and Reference No.
Staphylococcus I)og 62-74 + 15 Continuous infusion, intact Martin and Kerby (27)animal
Rabbit, 77 i 7 Single injection, intact ani- Rogers (18)mal; decreasing trappingfirst 10-30 min
Rat 67 Perfusion, isolated liver Wardlaw and Howard (24)
Escherichia coli log 40 Perfusion of isolated liver MIanwaring and Fritschen(23)
Rabbit 40-78 Single injection bacteria Rogers and Melly (20)into intact animal; in-creasing trapping first 30min
Rat 41 Perfusion of isolated liver Wardlaw and Howard (24)
Pneumococcus D)og 0-5 Perfusion of isolated liver Mlanwaring and Coe (22)Rabbit 8 i 14 Bacteremia from skin focus; Martin and Kerby (27)
intact animalRat 0 Perfusion of isolated liver Wardlaw and Howard (2-4)
* Microorganisms suspended in blood or normal heterologous serum.
teremia produced with one microbial species doesnot change the capacity of the splanchnic tissuesto trap other injected bacteria (27), and thatanimals moribund with metastatic infection canswiftly remove superimposed injections of bac-teria from their blood stream (26). Studies onpatients with long standing bacterial endocarditishave shown that splanchnic trapping is efficientand unaffected by the protracted bacteremia (16).For these reasons it is generally believed thatsaturation of or exhaustion of splanchnic trappingmechanisms does not play a role in the course ofnatural infections.
Studies performed with particulate substancesindicate, however, that the capacity of the reticu-lo-endothelial system to retain injected substancesis finite. Benacerraf, Biozzi, Halpern, and theirassociates have used carbon or other particulateor colloidal substances to show that clearancerates are always maximal when doses of particlessimilar to those utilized in bacteriologic experi-ments are injected intravenously (28-31). Theseobservations have helped to explain why the rateof clearance of bacteria appears independent ofthe numbers of bacteria administered. In essence,all microbial clearance studies are "low dose"experiments. If massive doses of particulate mat-ter are administered, the maximal trapping ca-
pacity of the splanchnic tissues is eventuallyexceeded. In this range, the rate of disappearanceof injected particles then becomes a direct func-tion of dosage. Using such massive doses, thetrapping capacity of reticulo-endothelial tissuescan be explored in various situations known toaffect host resistance.
Other Factors Operating to Clear Bacteriafrom the Blood Stream
Although the reticulo-endothelial tissues of theliver and spleen are quantitatively most impor-tant in removing circulating bacteria, there isevidence to indicate that other host mechanismsparticipate in blood stream clearance. This is con-vincingly demonstrated by clearance studies per-formed in animals whose splanchnic tissues havebeen rendered incapable of trapping bacteria.
Injections of large doses of certain substancessuch as colloidal thorium dioxide (Thorotrast),carbon, saccharated iron oxide, lithium carmine,or trypan blue can "blockade" the splanchnic tis-sues so that bacterial uptake in them is impaired(14, 18, 30, 32). Despite elimination of thesplanchnic bed as a trapping site, striking disap-pearance of circulating microorganisms can oftenbe demonstrated.
Such an experiment is pictured in figure 5. In
54 [VOL. 24
HOST MECHANISMS FOR REMOVAL OF BACTERIA FROM BLOOD
t: a~~~~~~TAPHYLOCOCC Ico
0 o
1 5 10M INUTES
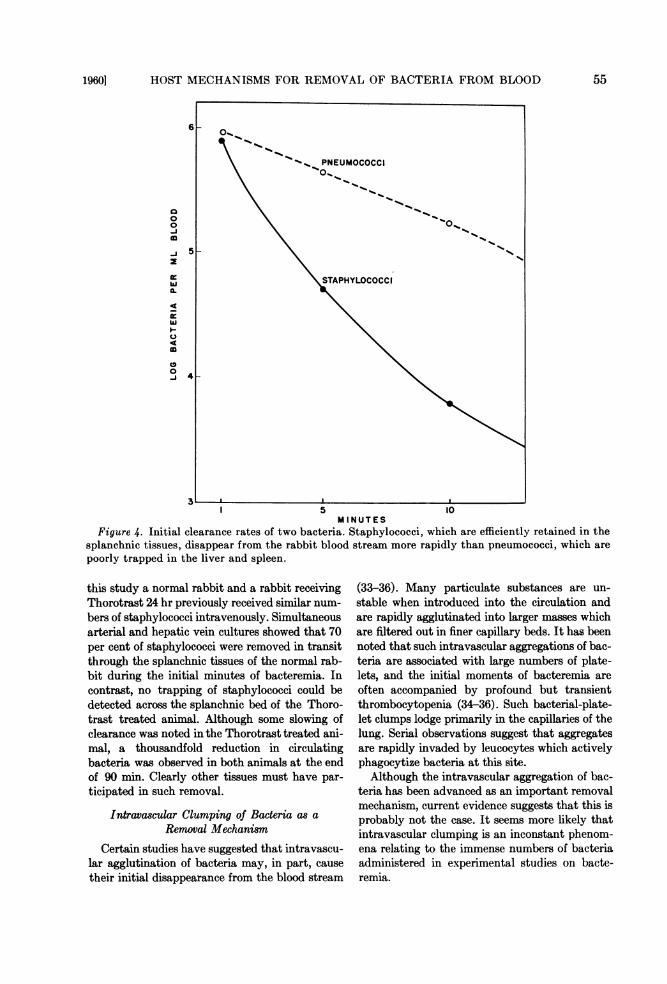
Figure 4. Initial clearance rates of two bacteria. Staphylococci, which are efficiently retained in thesplanchnic tissues, disappear from the rabbit blood stream more rapidly than pneumococci, which arepoorly trapped in the liver and spleen.
this study a normal rabbit and a rabbit receivingThorotrast 24 hr previously received similar num-bers of staphylococci intravenously. Simultaneousarterial and hepatic vein cultures showed that 70per cent of staphylococci were removed in transitthrough the splanchnic tissues of the normal rab-bit during the initial minutes of bacteremia. Incontrast, no trapping of staphylococci could bedetected across the splanchnic bed of the Thoro-trast treated animal. Although some slowing ofclearance was noted in the Thorotrast treated ani-mal, a thousandfold reduction in circulatingbacteria was observed in both animals at the endof 90 min. Clearly other tissues must have par-ticipated in such removal.
Intravascular Clumping of Bacteria as aRemoval Mechanism
Certain studies have suggested that intravascu-lar agglutination of bacteria may, in part, causetheir initial disappearance from the blood stream
(33-36). Many particulate substances are un-stable when introduced into the circulation andare rapidly agglutinated into larger masses whichare filtered out in finer capillary beds. It has beennoted that such intravascular aggregations of bac-teria are associated with large numbers of plate-lets, and the initial moments of bacteremia areoften accompanied by profound but transientthrombocytopenia (34-36). Such bacterial-plate-let clumps lodge primarily in the capillaries of thelung. Serial observations suggest that aggregatesare rapidly invaded by leucocytes which activelyphagocytize bacteria at this site.Although the intravascular aggregation of bac-
teria has been advanced as an important removalmechanism, current evidence suggests that this isprobably not the case. It seems more likely thatintravascular clumping is an inconstant phenom-ena relating to the immense numbers of bacteriaadministered in experimental studies on bacte-remia.
19601 55
DAVID E. ROGERS
a00-j
co
-J
C.,4
(9)
0-i
10 20 M I N U T E S
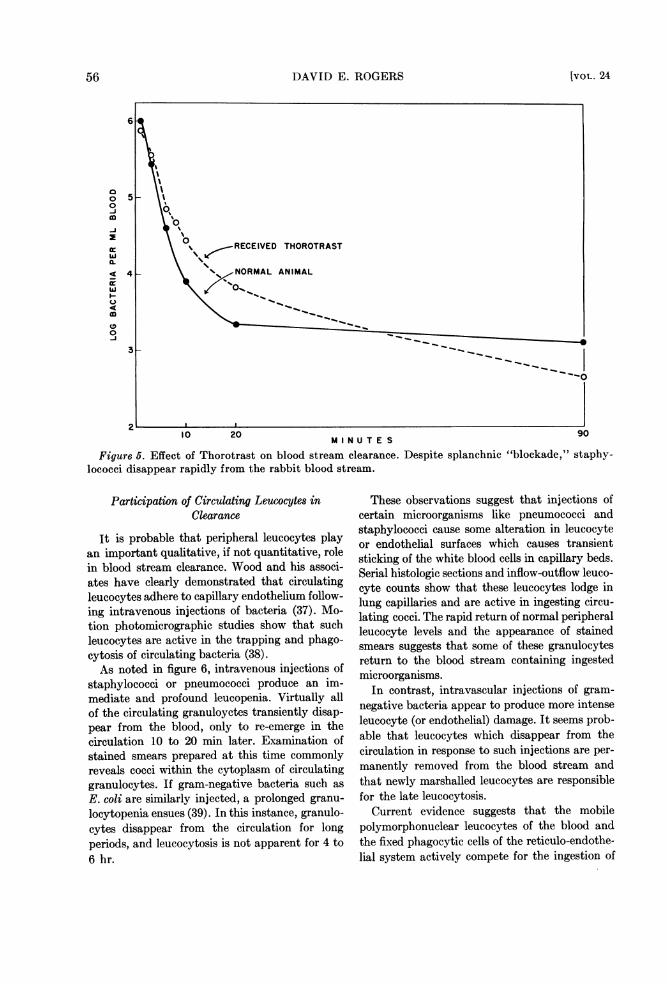
Figure 6. Effect of Thorotrast on blood stream clearance. Despite splanchnic "blockade," staphy-lococci disappear rapidly from the rabbit blood stream.
Participation of Circulating Leucocytes inClearance
It is probable that peripheral leucocytes playan important qualitative, if not quantitative, rolein blood stream clearance. Wood and his associ-ates have clearly demonstrated that circulatingleucocytes adhere to capillary endothelium follow-ing intravenous injections of bacteria (37). Mo-tion photomicrographic studies show that suchleucocytes are active in the trapping and phago-cytosis of circulating bacteria (38).As noted in figure 6, intravenous injections of
staphylococci or pneumococci produce an im-mediate and profound leucopenia. Virtually allof the circulating granuloyctes transiently disap-pear from the blood, only to re-emerge in thecirculation 10 to 20 min later. Examination ofstained smears prepared at this time commonlyreveals cocci within the cytoplasm of circulatinggranulocytes. If gram-negative bacteria such as
E. coli are similarly injected, a prolonged granu-
locytopenia ensues (39). In this instance, granulo-cytes disappear from the circulation for longperiods, and leucocytosis is not apparent for 4 to6 hr.
These observations suggest that injections ofcertain microorganisms like pneumococci andstaphylococci cause some alteration in leucocyteor endothelial surfaces which causes transientsticking of the white blood cells in capillary beds.Serial histologic sections and inflow-outflow leuco-cyte counts show that these leucocytes lodge inlung capillaries and are active in ingesting circu-lating cocci. The rapid return of normal peripheralleucocyte levels and the appearance of stainedsmears suggests that some of these granulocytesreturn to the blood stream containing ingestedmicroorganisms.
In contrast, intravascular injections of gram-negative bacteria appear to produce more intenseleucocyte (or endothelial) damage. It seems prob-able that leucocytes which disappear from thecirculation in response to such injections are per-manently removed from the blood stream andthat newly marshalled leucocytes are responsiblefor the late leucocytosis.
Current evidence suggests that the mobilepolymorphonuclear leucocytes of the blood andthe fixed phagocytic cells of the reticulo-endothe-lial system actively compete for the ingestion of
56 [VOL. 24

HOST MECHANISMS FOR REMOVAL OF BACTERIA FROM BLOOD
O .E.COLI*-100 -
o ST~~APHYLOCOCC
00 | 2 3 4 5 6
H 0 U R SFigure 6. Changes in circulating polymorphonuclear leucocytes following intravenous injection of
bacteria. Staphylococci and pneumococci produce a transient disappearance of circulating granulocytes.Injections of Escherichia coli produce a prolonged granulocytopenia (from D. E. Rogers, Cellular Man-agement of Bacterial Parasites, Pasteur Fermentation Centennial, 1857-1957, pp. 61-73, 1958, reprinted bypermission of Chas. Pfizer and Co., Inc.).
circulating bacteria. When splanchnic trappingis avid, polymorphonuclear leucocytes probablyplay a minor quantitative role in clearance. Whenbacteria are poorly retained in the splanchnictissues, granulocytes may be more important incontrol of bacteremia.
Evidence can also be marshalled to suggestthat the intracellular behavior of bacteria in-gested by polymorphonuclear leucocytes in theblood stream may, in part, determine whetherbacteremia will persist or disappear during thesecond phase of clearance. This can be illustratedby the intravascular behavior of a strain of co-
agulase-positive staphylococcus, an organismwhich survives in significant numbers withinleucocytes in our studies (18), and a strain oftype III pneumococcus, a microbe promptly de-stroyed by phagocytosis (40).When large numbers of staphylococci are ad-
ministered to rabbits, splanchnic trapping is ini-tially avid, then rapidly declines, and virtuallyceases within 15 to 20 min. A low grade bacte-remia then persists for many hours. Extensivestudies have shown that this cessation of splanch-nic trapping is not due to saturation of splanchnicclearance mechanisms, or decreasing bacterial-reticulo-endothelial cell contact (18). Differentialcultural studies indicate that virtually all of the
viable staphylococci are associated with the cir-culating leucocytes at the time that granulocytesreappear in the circulation. It is at this time thatsplanchnic trapping ceases, suggesting that theincorporation of cocci within leucocytes protectsthem from trapping within the reticulo-endothe-lial bed.
This thesis is supported by the observationthat rabbits rendered granulocytopenic can re-move more staphylococci from their circulationthan can normal animals. Furthermore, if viablestaphylococci are incorporated within rabbit leu-cocytes in vitro and then reinjected into rabbits,the staphylococci remain in the circulation forlonger periods than do injected extracellular cocci(18). Thus in this instance the intravascular phag-ocytosis of a microbe which can survive withinthe cytoplasm of leucocytes may permit its per-sistence in the rabbit circulation.
Conversely, while pneumococci are inefficientlyremoved in the splanchnic circuit, these micro-organisms disappear completely from the rabbitblood stream during the first 60 to 90 min afterinjection. Examination of peripheral blood smearsperformed on animals receiving intravenouspneumococci also revealed visible cocci withincirculating granulocytes, yet in this situation via-ble pneumococci are culturable only from the
19601 57
DAVID E. ROGERS
plasma layer. It would appear that in this in-stance leucocytes have rendered the ingestedpneumococci nonculturable. Thus in pneumococ-cal bacteremia it would appear that destructionby polymorphonuclear leucocytes may play animportant role in blood stream sterilization.These observations are supported by earlier
workers. Many years ago Wright showed that theblood stream clearance of a particular strain ofpneumococcus could be directly related to thebactericidal power of the peripheral blood in vitro(14). This finding has been confirmed by others(8).There is recent evidence to suggest that the
-leucocytes which disappear from the circulationmay still be capable of clearing bacteria from theblood stream. Hollingsworth and Beeson (41)have shown that rabbits rendered leucopenic byirradiation maintain higher bacteremias followingintravenous injections of E. coli than do normalanimals. However, if such leucopenic animals aretransfused with healthy rabbit leucocytes, subse-quent injections of E. coli are cleared normally,despite the fact that the transfused leucocyteshave disappeared from the peripheral blood (42).These observations strongly suggest that leuco-cytes sequestered in capillary beds can, in certaininstances, remain active in removing bacteriafrom the circulation.
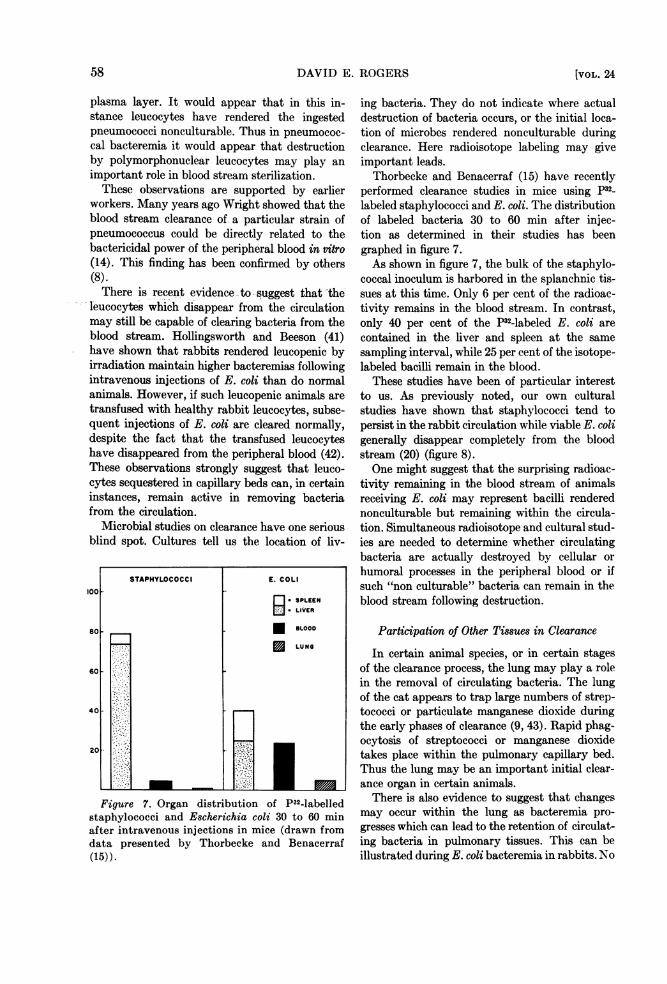
Microbial studies on clearance have one seriousblind spot. Cultures tell us the location of liv-
100
80o
60O
40
20
Figure 7. Organ distribution of P32-labelledstaphylococci and Escherichia coli 30 to 60 minafter intravenous injections in mice (drawn fromdata presented by Thorbecke and Benacerraf(15)).
ing bacteria. They do not indicate where actualdestruction of bacteria occurs, or the initial loca-tion of microbes rendered nonculturable duringclearance. Here radioisotope labeling may giveimportant leads.Thorbecke and Benacerraf (15) have recently
performed clearance studies in mice using p32_labeled staphylococci and E. coli. The distributionof labeled bacteria 30 to 60 min after injec-tion as determined in their studies has beengraphed in figure 7.As shown in figure 7, the bulk of the staphylo-
coccal inoculum is harbored in the splanchnic tis-sues at this time. Only 6 per cent of the radioac-tivity remains in the blood stream. In contrast,only 40 per cent of the P32-labeled E. coli arecontained in the liver and spleen at the samesampling interval, while 25 per cent of the isotope-labeled bacilli remain in the blood.
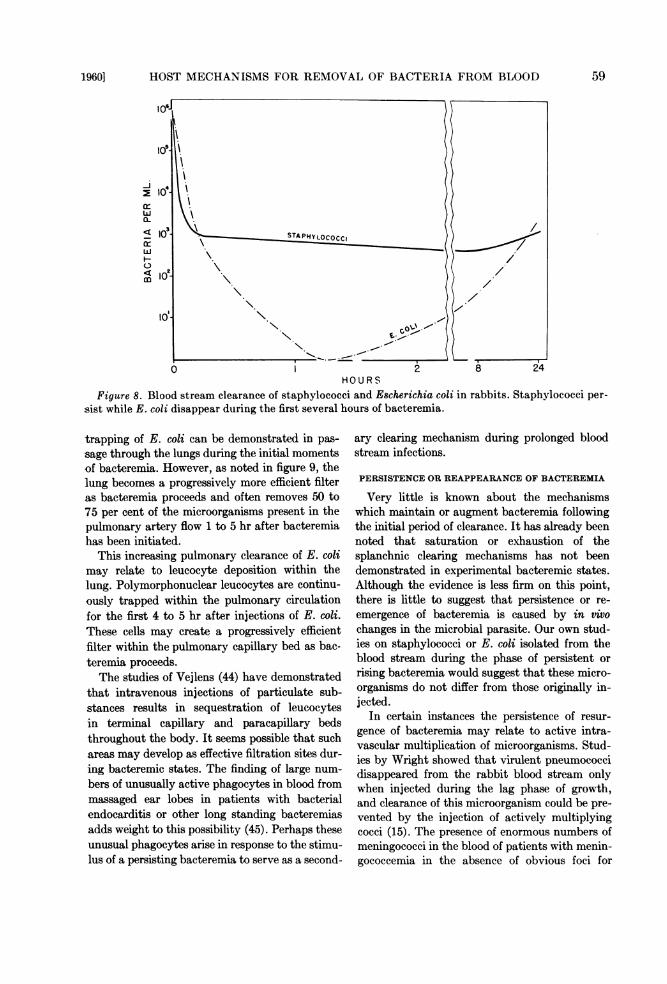
These studies have been of particular interestto us. As previously noted, our own culturalstudies have shown that staphylococci tend topersist in the rabbit circulation while viable E. coligenerally disappear completely from the bloodstream (20) (figure 8).One might suggest that the surprising radioac-
tivity remaining in the blood stream of animalsreceiving E. coli may represent bacilli renderednonculturable but remaining within the circula-tion. Simultaneous radioisotope and cultural stud-ies are needed to determine whether circulatingbacteria are actually destroyed by cellular orhumoral processes in the peripheral blood or ifsuch "non culturable" bacteria can remain in theblood stream following destruction.
Participation of Other Tissues in Clearance
In certain animal species, or in certain stagesof the clearance process, the lung may play a rolein the removal of circulating bacteria. The lungof the cat appears to trap large numbers of strep-tococci or particulate manganese dioxide duringthe early phases of clearance (9, 43). Rapid phag-ocytosis of streptococci or manganese dioxidetakes place within the pulmonary capillary bed.Thus the lung may be an important initial clear-ance organ in certain animals.There is also evidence to suggest that changes
may occur within the lung as bacteremia pro-gresses which can lead to the retention of circulat-ing bacteria in pulmonary tissues. This can beillustrated during E. coli bacteremia in rabbits. No
STAPHYLOCOCCI E. COUl
aI* SPLEEN
.I* LIVER
* BLOOD
U LUNG
;. 101 | ~~WI
58 [VOL. 24
HOST MECHANISMS FOR REMOVAL OF BACTERIA FROM BLOOD
G 0 STA PHY LOCOCCI 7 /
I0 /82
cr~~~ ~ ~ ~~~~~~~.
0 \
H 2U R SFigure 8. Blood stream clearance of staphylococci and Escherichia coli in rabbits. Staphylococci per-
sist while E. coli disappear during the first several hours of bacteremia.
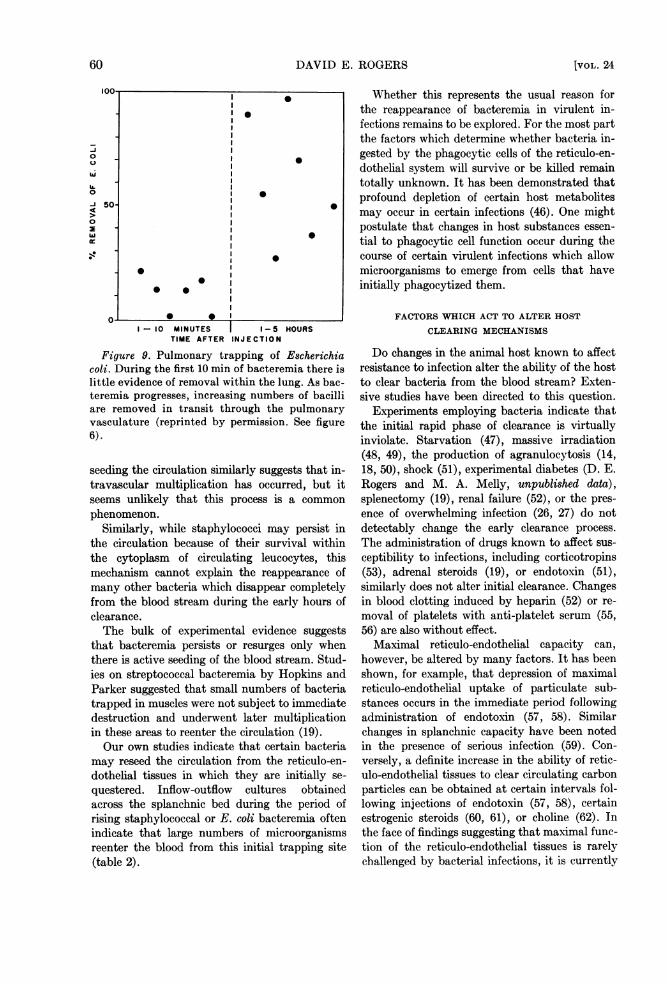
trapping of E. coli can be demonstrated in pas-sage through the lungs during the initial momentsof bacteremia. However, as noted in figure 9, thelung becomes a progressively more efficient filteras bacteremia proceeds and often removes 50 to75 per cent of the microorganisms present in thepulmonary artery flow 1 to 5 hr after bacteremiahas been initiated.
This increasing pulmonary clearance of E. colimay relate to leucocyte deposition within thelung. Polymorphonuclear leucocytes are continu-ously trapped within the pulmonary circulationfor the first 4 to 5 hr after injections of E. coli.These cells may create a progressively efficientfilter within the pulmonary capillary bed as bac-teremia proceeds.The studies of Vejlens (44) have demonstrated
that intravenous injections of particulate sub-stances results in sequestration of leucocytesin terminal capillary and paracapillary bedsthroughout the body. It seems possible that suchareas may develop as effective filtration sites dur-ing bacteremic states. The finding of large num-bers of unusually active phagocytes in blood frommassaged ear lobes in patients with bacterialendocarditis or other long standing bacteremiasadds weight to this possibility (45). Perhaps theseunusual phagocytes arise in response to the stimu-lus of a persisting bacteremia to serve as a second-
ary clearing mechanism during prolonged bloodstream infections.
PERSISTENCE OR REAPPEARANCE OF BACTEREMIA
Very little is known about the mechanismswhich maintain or augment bacteremia followingthe initial period of clearance. It has already beennoted that saturation or exhaustion of thesplanchnic clearing mechanisms has not beendemonstrated in experimental bacteremic states.Although the evidence is less firm on this point,there is little to suggest that persistence or re-emergence of bacteremia is caused by in vivochanges in the microbial parasite. Our own stud-ies on staphylococci or E. coli isolated from theblood stream during the phase of persistent orrising bacteremia would suggest that these micro-organisms do not differ from those originally in-jected.
In certain instances the persistence of resur-gence of bacteremia may relate to active intra-vascular multiplication of microorganisms. Stud-ies by Wright showed that virulent pneumococcidisappeared from the rabbit blood stream onlywhen injected during the lag phase of growth,and clearance of this microorganism could be pre-vented by the injection of actively multiplyingcocci (15). The presence of enormous numbers ofmeningococci in the blood of patients with menin-gococcemia in the absence of obvious foci for
1960] 59
DAVID E. ROGERS
-J
0
L&I0
-J 50
>0
>-* ~ ~~
0~~~*
1- 10 MINUTES I-5 HOURSTIME AFTER INJECTION
Figure 9. Pulmonary trapping of Escherichiacoli. During the first 10 min of bacteremia there islittle evidence of removal within the lung. As bac-teremia progresses, increasing numbers of bacilliare removed in transit through the pulmonaryvasculature (reprinted by permission. See figure6).
seeding the circulation similarly suggests that in-travascular multiplication has occurred, but itseems unlikely that this process is a common
phenomenon.Similarly, while staphylococci may persist in
the circulation because of their survival withinthe cytoplasm of circulating leucocytes, thismechanism cannot explain the reappearance ofmany other bacteria which disappear completelyfrom the blood stream during the early hours ofclearance.The bulk of experimental evidence suggests
that bacteremia persists or resurges only whenthere is active seeding of the blood stream. Stud-ies on streptococcal bacteremia by Hopkins andParker suggested that small numbers of bacteriatrapped in muscles were not subject to immediatedestruction and underwent later multiplicationin these areas to reenter the circulation (19).Our own studies indicate that certain bacteria
may reseed the circulation from the reticulo-en-dothelial tissues in which they are initially se-
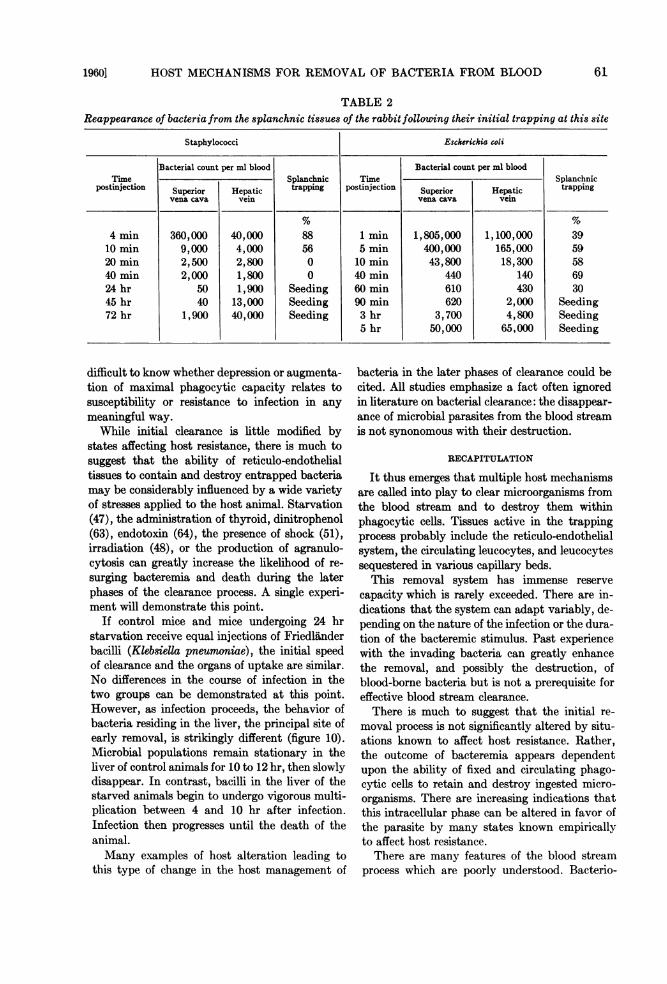
questered. Inflow-outflow cultures obtainedacross the splanchnic bed during the period ofrising staphylococcal or E. coli bacteremia oftenindicate that large numbers of microorganismsreenter the blood from this initial trapping site(table 2).
Whether this represents the usual reason forthe reappearance of bacteremia in virulent in-fections remains to be explored. For the most partthe factors which determine whether bacteria in-gested by the phagocytic cells of the reticulo-en-dothelial system will survive or be killed remaintotally unknown. It has been demonstrated thatprofound depletion of certain host metabolitesmay occur in certain infections (46). One mightpostulate that changes in host substances essen-tial to phagocytic cell function occur during thecourse of certain virulent infections which allowmicroorganisms to emerge from cells that haveinitially phagocytized them.
FACTORS WHICH ACT TO ALTER HOST
CLEARING MECHANISMS
Do changes in the animal host known to affectresistance to infection alter the ability of the hostto clear bacteria from the blood stream? Exten-sive studies have been directed to this question.
Experiments employing bacteria indicate thatthe initial rapid phase of clearance is virtuallyinviolate. Starvation (47), massive irradiation(48, 49), the production of agranulocytosis (14,18, 50), shock (51), experimental diabetes (D. E.Rogers and M. A. Melly, unpublished data),splenectomy (19), renal failure (52), or the pres-ence of overwhelming infection (26, 27) do notdetectably change the early clearance process.The administration of drugs known to affect sus-ceptibility to infections, including corticotropins(53), adrenal steroids (19), or endotoxin (51),similarly does not alter initial clearance. Changesin blood clotting induced by heparin (52) or re-moval of platelets with anti-platelet serum (55,56) are also without effect.Maximal reticulo-endothelial capacity can,
however, be altered by many factors. It has beenshown, for example, that depression of maximalreticulo-endothelial uptake of particulate sub-stances occurs in the immediate period followingadministration of endotoxin (57, 58). Similarchanges in splanchnic capacity have been notedin the presence of serious infection (59). Con-versely, a definite increase in the ability of retic-ulo-endothelial tissues to clear circulating carbonparticles can be obtained at certain intervals fol-lowing injections of endotoxin (57, 58), certainestrogenic steroids (60, 61), or choline (62). Inthe face of findings suggesting that maximal func-tion of the reticulo-endothelial tissues is rarelychallenged by bacterial infections, it is currently
I a
l
60 [VOL. 24
HOST MECHANISMS FOR REMOVAL OF BACTERIA FROM BLOOD
TABLE 2Reappearance of bacteria from the splanchnic tissues of the rabbit following their initial trapping at this site
Staphylococci Esckerichia coli
Bacterial count per ml blood Bacterial count per ml bloodTime Splanchnic Time Splanchnic
postinjection Superior Hepatic trapping postinjection Superior Hepatic trappingvena cava vein vena cava vein
4 min 360,000 40,000 88 1 min 1,805,000 1,100,000 3910min 9,000 4,000 56 5 min 400,000 165,000 5920min 2,500 2,800 0 10 min 43,800 18,300 5840 min 2,000 1,800 0 40min 440 140 6924 hr 50 1,900 Seeding 60min 610 430 3045 hr 40 13,000 Seeding 90 min 620 2,000 Seeding72 hr 1,900 40,000 Seeding 3 hr 3,700 4,800 Seeding
5 hr 50,000 65,000 Seeding
difficult to know whether depression or augmenta-tion of maximal phagocytic capacity relates tosusceptibility or resistance to infection in anymeaningful way.
While initial clearance is little modified bystates affecting host resistance, there is much tosuggest that the ability of reticulo-endothelialtissues to contain and destroy entrapped bacteriamay be considerably influenced by a wide varietyof stresses applied to the host animal. Starvation(47), the administration of thyroid, dinitrophenol(63), endotoxin (64), the presence of shock (51),irradiation (48), or the production of agranulo-cytosis can greatly increase the likelihood of re-
surging bacteremia and death during the laterphases of the clearance process. A single experi-ment will demonstrate this point.
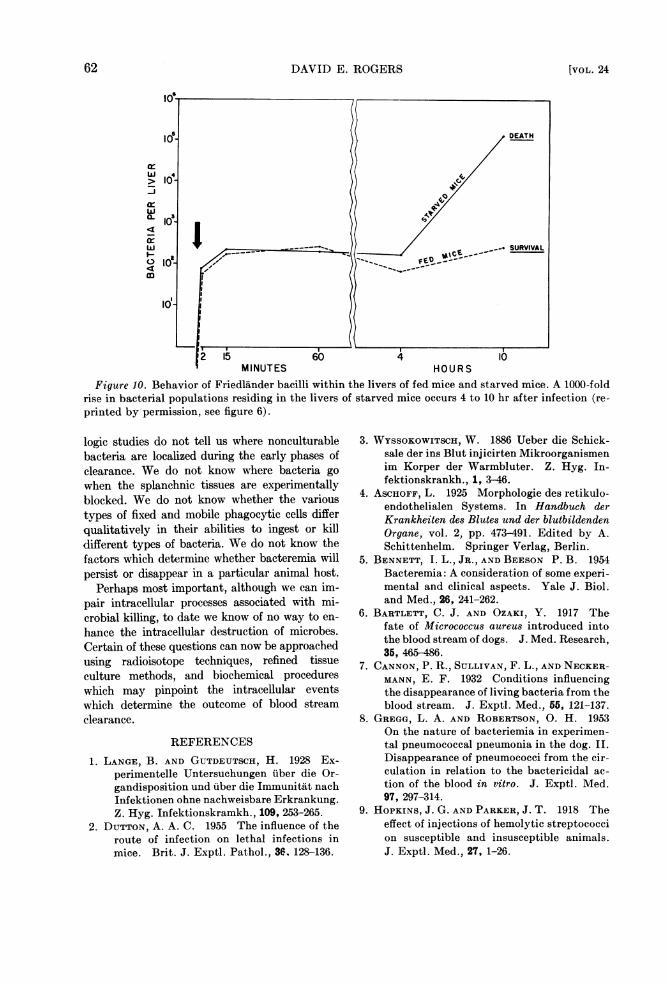
If control mice and mice undergoing 24 hrstarvation receive equal injections of FriedlAnderbacilli (Klebsiella pneumoniae), the initial speedof clearance and the organs of uptake are similar.No differences in the course of infection in thetwo groups can be demonstrated at this point.However, as infection proceeds, the behavior ofbacteria residing in the liver, the principal site ofearly removal, is strikingly different (figure 10).Microbial populations remain stationary in theliver of control animals for 10 to 12 hr, then slowlydisappear. In contrast, bacilli in the liver of thestarved animals begin to undergo vigorous multi-plication between 4 and 10 hr after infection.Infection then progresses until the death of theanimal.Many examples of host alteration leading to
this type of change in the host management of
bacteria in the later phases of clearance could becited. All studies emphasize a fact often ignoredin literature on bacterial clearance: the disappear-ance of microbial parasites from the blood streamis not synonomous with their destruction.
RECAPITULATION
It thus emerges that multiple host mechanismsare called into play to clear microorganisms fromthe blood stream and to destroy them withinphagocytic cells. Tissues active in the trappingprocess probably include the reticulo-endothelialsystem, the circulating leucocytes, and leucocytessequestered in various capillary beds.
This removal system has immense reservecapacity which is rarely exceeded. There are in-dications that the system can adapt variably, de-pending on the nature of the infection or the dura-tion of the bacteremic stimulus. Past experiencewith the invading bacteria can greatly enhancethe removal, and possibly the destruction, ofblood-borne bacteria but is not a prerequisite foreffective blood stream clearance.There is much to suggest that the initial re-
moval process is not significantly altered by situ-ations known to affect host resistance. Rather,the outcome of bacteremia appears dependentupon the ability of fixed and circulating phago-cytic cells to retain and destroy ingested micro-organisms. There are increasing indications thatthis intracellular phase can be altered in favor ofthe parasite by many states known empiricallyto affect host resistance.
There are many features of the blood streamprocess which are poorly understood. Bacterio-
1960] 61L
DAVID E. ROGERS
io 6W 4>101
a-
10
W 'I--SURVIVAL
2 15 ~~~60 4 10MINUTES HOURS
Figure 10. Behavior of Friedlander bacilli within the livers of fed mice and starved mice. A 1000-foldrise in bacterial populations residing in the livers of starved mice occurs 4 to 10 hr after infection (re-printed by permission, see figure 6).
logic studies do not tell us where nonculturablebacteria are localized during the early phases ofclearance. We do not know where bacteria gowhen the splanchnic tissues are experimentallyblocked. We do not know whether the varioustypes of fixed and mobile phagocytic cells differqualitatively in their abilities to ingest or killdifferent types of bacteria. We do not know thefactors which determine whether bacteremia willpersist or disappear in a particular animal host.
Perhaps most important, although we can im-pair intracellular processes associated with mi-crobial killing, to date we know of no way to en-hance the intracellular destruction of microbes.Certain of these questions can now be approachedusing radioisotope techniques, refined tissueculture methods, and biochemical procedureswhich may pinpoint the intracellular eventswhich determine the outcome of blood streamclearance.
REFERENCES
1. LANGE, B. AND GUTDEUTSCH, H. 1928 Ex-perimentelle Untersuchungen uber die Or-gandisposition und uber die ImmunitAt nachInfektionen ohne nachweisbare Erkrankung.Z. Hyg. Infektionskramkh., 109, 253-265.
2. DUTTON, A. A. C. 1955 The influence of theroute of infection on lethal infections inmice. Brit. J. Exptl. Pathol., 36. 128-136.
3. WYSSOKOWITSCH, W. 1886 Ueber die Schick-sale der ins Blut injicirten Mikroorganismenim Korper der Warmbluter. Z. Hyg. In-fektionskrankh., 1, 3-46.
4. ASCHOFF, L. 1925 Morphologie des retikulo-endothelialen Systems. In Handbuch derKrankheiten des Blutes und der blutbildendenOrgane, vol. 2, pp. 473-491. Edited by A.Schittenhelm. Springer Verlag, Berlin.
5. BENNETT, I. L., JR., AND BEESON P. B. 1954Bacteremia: A consideration of some experi-mental and clinical aspects. Yale J. Biol.and Med., 26, 241-262.
6. BARTLETT, C. J. AND OZAKI, Y. 1917 Thefate of Micrococcus aureus introduced intothe blood stream of dogs. J. Med. Research,35, 465-486.
7. CANNON, P. R., SULLIVAN, F. L., AND NECKER-MANN, E. F. 1932 Conditions influencingthe disappearance of living bacteria from theblood stream. J. Exptl. Med., 55, 121-137.
8. GREGG, L. A. AND ROBERTSON, 0. H. 1953On the nature of bacteriemia in experimen-tal pneumococcal pneumonia in the dog. II.Disappearance of pneumococci from the cir-culation in relation to the bactericidal ac-tion of the blood in vitro. J. Exptl. Med.97, 297-314.
9. HOPKINS, J. G. AND PARKER, J. T. 1918 Theeffect of injections of hemolytic streptococcion susceptible and insusceptible animals.J. Exptl. Med., 27, 1-26.
62 [VOL. 24

HOST MECHANISMS FOR REMOVAL OF BACTERIA FROM BLOOD
10. SULLIVAN, F. L., NECKERMANN, E. F., ANDCANNON, P. R. 1934 The localization andfate of bacteria in the tissues. J. Immunol.,26, 49-67.
11. TEALE, F. H. 1935 Some observations onthe relative importance of the reticuloen-dothelial tissues and the circulating anti-body in umminity. J. Immunol., 28, 133-182.
12. OTTENBERG, R. 1931 The rate and locationof removal of bacteria from the blood inhuman disease. Arch. Pathol., 11, 766-774.
13. PARKER, J. T. AND FRANKE, E. 1918 Thefate of thyphoid bacilli injected intrave-nously into normal and typhoid immunerabbits. J. Med. Research, 39, 301-309.
14. WRIGHT, H. D. 1927 Experimental pneumo-coccal septicaemia and anti-pneumococcalimmunity. J. Pathol. Bacteriol., 30, 185-252.
15. THORBECKE, G. J. AND BENACERRAF, B. 1959Some histological and functional aspects oflymphoid tissue in germ free animals. II.Studies on phagocytosis in vivo. Ann. N.Y. Acad. Sci., 78, 247-253.
16. BEESON, P., BRANNON, E. S. AND WARNER, J.V. 1945 Observations on the sites of re-moval of bacteria from the blood in patientswith bacterial endocarditis. J. Exptl.Med., 81, 9-23.
17. MARTIN, S. P., KERBY, G. P., AND HOLLAND,B. C. 1949 A method for measuring re-moval of bacteria from the blood by thevarious organs of the intact animal. Proc.Soc. Exptl. Biol. Med., 72, 63-68.
18. ROGERS, D. E. 1956 Studies on bacterie-mia. I. Mechanisms relating to the persist-ence of bacteriemia in rabbits following theintravenous injection of staphylococci. J.Exptl. Med. 103, 713-742.
19. ROGERS, D. E. AND MELLY, M. A. 1957Studies on bacteriemia. II. Further observa-tions on the granulocytopenia induced bythe intravenous injection of staphylococci.J. Exptl. Med., 106, 99-112.
20. ROGERS, D. E. AND MELLY, M. A. 1957Studies on bacteriemia. III. The bloodstream clearance of Escherichia coli in rab-bits. J. Exptl. Med., 106, 113-124.
21. KERBY, G. P. 1950 A comparison of the re-moval of mucoid and non-mudoid variantsof Klebsiella pneumoniae type B from thesplanchnic circulating blood of the intactanimal. J. Immunol., 64, 131-137.
22. MANWARING, W. H. AND COE, H. C. 1916Endothelial opsonins. J. Immunol., 1, 401-408.
23. MANWARING, W. H. AND FRITSCHEN, W. 1923
Study of microbic-tissue affinity by perfu-sion methods. J. Immunol., 8, 83-89.
24. WARDLAW, A. C. AND HOWARD, J. G. 1959A comparative survey of the phagocytosisof different species of bacteria by Kupffercells. Brit. J. Exptl. Pathol., 40, 113-117.
25. GOVAERTS, P. D. 1925 Un essai d'interpre-tation des septicemies chirurgicales. J.chir. et ann. soc. belge chir., 2, 1-22.
26. REICHEL, H. A. 1939 Removal of bacteriafrom the blood stream: Experiments tendingto determine the rate of removal of injectedbacteria in the blood. Proc. Staff Meet-ings Mayo Clinic, 14, 138-143.
27. MARTIN, S. P. AND KERBY, G. P. 1950 Thespanchnic removal in rabbits during fatalbacteriemias of circulating organisms and ofsuperimposed non-pathogenic bacteria. J.Exptl. Med., 92, 45-49.
28. HALPERN, B. N., BENACERRAF, B., AND Biozzi,G. 1953 Quantitative study of the gran-ulopectic activity of the reticulo-endothelialsystem. I. The effect of the ingredients pres-ent in india ink and of substances affectingblood clotting in vivo on the fate of carbonparticles administered intravenously in rats,mice and rabbits. Brit. J. Exptl. Pathol.,34, 426-440.
29. Biozzi, G., BENACERRAF, B., ,AND HALPERN,B. N. 1953 Quantitative study of thegranulopectic activity of the reticulo-endo-thelial system. II. A study of the kineticsof the granulopectic activity of the R.E.S.in relation to the dose of carbon injected.Relationship between the weight of the or-gans and their activity. Brit. J. Exptl.Pathol., 34, 441-457.
30. BENACERRAF, B. 1958 Quantitative aspectsof phagocytosis. In Symposium on liverfunction, Publ. no. 4, pp. 205-227. Am. Inst.Biol. Sci., Washington, D. C.
31. Biozzi, G., BENACERRAF, B., HALPERN, B. N.,STIFFEL, L., AND HILLEMAND, B. 1959Exploration of the phagocytic system of thereticuloendothelial system with heat dena-tured human serum albumin labelled withI'3s and application to the measurement ofliver blood flow in normal man and somepathologic conditions. J. Lab. Clin. Med.,61, 230-239.
32. MARTIN, S. P., KERBY, G. P., AND HOLLAND,B. C. 1952 The effect of thorotrast on theremoval of bacteria in the splanchnic areaof the intact animal. J. Immunol., 68, 293-296.
33. BULL, C. G. 1915 The agglutination of bac-teria in vivo. J. Exptl. Med., 22, 484-491.
34. BULL, C. G. 1916 Further observations of
19601

DAVID E. ROGERS
the agglutination of bacteria in vivo. J.Exptl. Med., 24, 25-33.
35. GOVAERTS, P. 1921 L'agglutination plas-matique, facteur d'instabilit6 des particulesintroduites dans la circulation. Compt.rend. soc. biol., 85, 244-245.
36. DUDGEON, L. S. AND GOADBY, H. K. 1931The examination of the tissues and someobservations on the blood platelets of rab-bits at intervals of five minutes, and later,after intravenous inoculations of Staphy-lococcus aureus and Indian ink. J. Hyg.,31, 247-256.
37. WOOD, W. B., JR., SMITH, M. R., PERRY, W.D., AND BERRY, J. W. 1951 Studies onthe cellular immunology of acute bacterie-mia. I. Intravascular leukocytic reactionand surface phagocytosis. J. Exptl. Med.,94, 521-534.
38. WOOD, W. B., JR., SMITH, M. R., BERRY, J. W.,AND PERRY W. D. 1951 Studies on thecellular immunology of acute bacteremia.Trans. Assoc. Am. Physicians, 64, 155-159.
39. BENNETT, I. L., JR. 1950 The propertiesand biologic effects of bacterial pyrogens.Medicine, 29, 365-400.
40. WOOD, W. B., JR., AND SMITH, M. R. 1947Surface phagocytosis-its relation to themechanism of recovery in acute pneumoniacaused by encapsulated bacteria. Trans.Assoc. Am. Physicians, 60, 77-81.
41. HOLLINGSWORTH, J. W. AND BEESON, P. B.1955 Experimental bacteremia in normaland irradiated rats. Yale J. Biol. and Med.28, 56-62.
42. HOLLINGSWORTH, J. W., FINCH, S. C., ANDAND BEESON, P. B. 1956 The role oftransfused leukocytes in experimental bac-teremia of irradiated rats. J. Lab. Clin.Med., 48, 227-236.
43. DRINKER, C. K. AND SHAW, L. A. 1921Quantitative distribution of particulate ma-terial (manganese dioxide) administered in-travenously to the cat. J. Exptl. Med., 33,77-98.
44. VEJLENS, G. 1938 The distribution of leu-kocytes in the vascular system. ActaPathol. Microbiol. Scand. Suppl., 33, 5-239.
45. DALAND, G. A., GOTTLIEB, L., WALLERSTEIN,R. O., AND CASTLE, W. B. 1956 Hemato-logic observations in bacterial endocarditis.Especially the prevalence of histiocytes andthe elevation and variation of the white cellcount in blood from the ear lobe. J. Lab.& Clin. Med., 48, 827-845.
46. WOODWARD, J. M., SBARRA, A. J., AND HOLT-MAN, D. F. 1954 The host-parasite rela-tionship in tularemia. I. A study of the in-
fluence of Bacterium tularense on the aminoacid metabolism of white rats. J. Bacteriol.,67, 58-61.
47. SMITH, J. M. AND DuBos, R. J. 1956 Theeffect of nutritional disturbances on thesusceptibility of mice to staphylococcal in-fections. J. Exptl. Med., 103, 109-118.
48. GORDON, L. E., COOPER, D. B., AND MILLER,C. P. 1955 Clearance of bacteria fromthe blood of irradiated rabbits. Proc. Soc.Exptl. Biol. Med., 89, 577-579.
49. CALLAWAY, J. L. AND KERBY, G. 1951Splanchnic removal of bacteria from thecirculating blood of irradiated rabbits.Arch. Dermatol. and Syphilol., 63, 200-206.
50. KERBY, G. P. AND MARTIN, S. P. 1951 Thesplanchnic removal of bacteria from theblood stream of leukopenic rabbits. J.Exptl. Med., 93, 189-195.
51. FINE, J. 1954 Relation of bacteria to thefailure of blood-volume therapy in traumaticshock. New Engl. J. Med., 250, 889-895.
52. BALCH. H. H. AND EVANS, J. R. 1956 Theinfluence of acute renal failure on resistanceto infection: an experimental study. Ann.Surg., 144, 191-197.
53. MARTIN, S. P. AND KERBY, G P. 1952 Ef-fect of adrenal hormone overdosage on bac-terial removal by the splanchnic viscera.Proc. Soc. Exptl. Biol. Med., 81, 73-75.
54. STAVITSKY, A. B. 1951 Factors involved inblood clearance of bacteria. Science, 113,520-522.
55. BULL, C. G. AND MCKEE, C. M. 1922 Therelation of blood platelets to the in vivoagglutination of bacteria and their disap-pearance from the blood stream. Am. J.Hyg., 2, 208-224.
56. GOVAERTS, P. 1921 Action du serum anti-plaquettique sur l'6limination des microbesintroduits dans la circulation. Compt.rend. soc. biol., 85, 667-668.
57. BIozzI, G. BENACERRAF, B., AND HALPERN,B. N. 1955 The effect of Salm. typhi andits endotoxin on the phagocytic activity ofthe reticuloendothelial system in mice.Brit. J. Exptl. Pathol., 36, 226-235.
58. BENACERRAF, B. AND SEBESTYEN, M. M. 1957Effect of bacterial endotoxin on the reticuol-endothelial system. Federation Proc., 16,860-867.
59. BOEHME, D. AND DuBos, R. J. 1958 Theeffect of bacterial constituents on the re-sistance of mice to heterologous infectionand on the activity of their reticuloendo-thelial system. J. Exptl. Med., 107, 523-536.
60. NICHOL, T., BILBEY, D. L., AND WARE, C. C.
64 [VOL. 24

HOST MECHANISMS FOR REMOVAL OF BACTERIA FROM BLOOD
1958 Effect of various stilbene compoundson the phagocytic activity of the reticulo-endothelial system. Nature, 181, 1538-1539.
61. HELLER, J. H., MEISER, R. M., ZUCHER, R.,AND MAST, G. W. 1957 The effect of nat-ural and synthetic estrogens on reticuloen-dothelial system function. Endocrinology,61, 235-241.
62. HELLER, J. H. 1953 Stimulation of thereticuloendothelial system with choline.Science, 118, 353-354.
63. SMITH, J. M. AND DuBos, R. J. 1956 Theeffect of dinitrophenol and thyroxin on thesusceptibility of mice to staphylococcal in-fections. J. Exptl. Med., 103, 119-126.
64. DUBOS, R. J. AND SCHAEDLER, R. W. 1956Reversible changes in the susceptibility ofmice to bacterial infections. I. Changesbrought about by injection of pertussis vac-cine or bacterial endotoxins. J. Exptl. Med.,104, 53-65.
DISCUSSION
Early reactions of the host to bacterial invasionmay or may not be determinants of the eventualoutcome of infection. For example, the rapidclearance by the reticulo-endothellal system ofbacteria injected intravenously may be the samewhether or not later reemergence of bacteremiaand death occur. Early reactions of the host, onthe other hand, have been shown to be associatedwith the late events in infection, as illustrated bythe relationship of the outcome of infection byvirulent and avirulent group A streptococci withthe ability of the phagocyte to capture and de-stroy the bacteria (Rogers, Nashville, and Wood,Baltimore).
Besides the effect of the early antibacterial de-fenses of the host, the result of induced infectionwill vary depending upon the route of inoculation.Indicative of this is the observation that a verysmall number of pneumococci, possibly even one,can produce a lethal infection in mice when in-oculated intraperitoneally, whereas very largenumbers must be given intravenously to producethe same effect (MacLeod, Philadelphia).
In addition to phagocytosis, blood clearance bythe reticulo-endothelial system, and route of in-oculation, other factors may play important rolesin determining the late events in infection. It wasshown, for example, that in the presence of re-emergent bacteremia and lethal infection thereticulo-endothelial system retains its tremendous
reserve capacity to remove heterologous or homol-ogous bacteria injected intravenously. Neverthe-less, the reemergent bacteremia is not necessarilyor usually controlled by the activity of the retic-ulo-endothelial system. This fact suggested thatthe bacteria able to persist in the fixed phagocyticsystem or able to reinvade the circulation may beresistant to the antibacterial action of the reticulo-endothelial cells. It was pointed out, however,that inoculation of blood containing the reinva-sive bacteria into normal animals is followed bythe usual rapid blood clearing phenomenon, pos-sibly illustrating that the persistent, reinvading,and multiplying bacteria are as susceptible toremoval by the reticulo-endothelial system asbacteria grown in vitro (Rogers, Nashville). Thework of Hedley Wright, however, indicated thatpneumococci in the logarithmic phase of growthmay evade the rapid initial blood clearing actionof the reticulo-endothelial system (Wright, H.D., 1927, Experimental pneumococcal septi-caemia and anti-pneumococcal immunity, J.Pathol. Bacteriol., 30, 185-252). Additional stud-ies will be required to elaborate the influence ofthe state of the microorganism upon the lateand early reactions of the host (Scherp, Be-thesda).The effect of opsonins or antibodies upon the
clearance mechanism of the reticulo-endothelialsystem and upon phagocytosis was discussed andexperiments were cited showing the blood clear-ance of Escherichia coli by fixed phagocytes inmice is inefficient, but as little as 0.01 ,ug of spe-cific antibody nitrogen per ml blood will greatlyincrease the clearing action of the reticulo-endo-thelial system (Benacerraf, New York). The fail-ure of the reticulo-endothelial system to phagocy-tize and clear a second phase bacteremia couldtherefore be attributable to depression of serumopsonins. The ability to remove antigenicallyidentical bacteria grown in vitro, however, evenin the presence of lethal infection, is evidenceagainst this possibility.
Specific antibody facilitates phagocytosis ofstaphylococci, and human serum after infancyalmost invariably contains such antibody. Inaddition, the serum of rabbits frequently hasno demonstrable antibody to the staphylococcus,although these animals may acquire such anti-bodies in their serum with increasing age, pos-sibly attributable to natural infection. In thestudy of phagocytosis by leucocytes and fixed
1960] 65

DAVID E. ROGERS
macrophages, therefore, attention must be givento the possible influence of serum opsonins (Row-ley, England).
Studies of clearance by the reticulo-endothelialsystem of bacteria injected intravenously, al-though necessary to elucidate some of the phenom-ena of nonspecific host resistance, may repre-
sent a relatively inefficient method of inoculatingthe microorganism into the peripheral tissues.If this is the case, it is probable that the earlyimmediate clearance of bacteria from the bloodstream does not take place in natural infection.The late phase of reemergent bacteremia, how-ever, may be comparable to bacteremia occur-
ring under natural conditions. Furthermore, dur-
ing the course of infection, the presence ofbacteria in the bloodstream is almost always a
reflection of active infection in the tissues, andit may not be the circulating bacteria thatrender the final outcome poor but the fact thatbacteremia indicates a spreading infection in thetissues which is out of control (Bennett, Balti-more).
Finally, the mechanism of the adherence ofleucocytes to vascular endothelium was discussed,and the experiments of Allison and Wood were
cited as showing that this adherence phenomenonmay be as dependent upon changes in the endo-thelium as in the leucocyte itself (Cluff, Balti-more).
66 [VOL. 24





























