Embed Size (px)
Citation preview

PRIMER
How to make a hippocampal dentate gyrus granule neuronDiana X. Yu, Maria C. Marchetto and Fred H. Gage*
ABSTRACTGranule neurons in the hippocampal dentate gyrus (DG) receive theirprimary inputs from the cortex and are known to be continuouslygenerated throughout adult life. Ongoing integration of newbornneurons into the existing hippocampal neural circuitry providesenhanced neuroplasticity, which plays a crucial role in learning andmemory; deficits in this process have been associated with cognitivedecline under neuropathological conditions. In this Primer, wesummarize the developmental principles that regulate the processof DG neurogenesis and discuss recent advances in harnessingthese developmental cues to generate DG granule neurons fromhuman pluripotent stem cells.
KEY WORDS: Dentate gyrus, Granule neurons, Hippocampalneurogenesis, Stem cells
IntroductionThe hippocampal dentate gyrus (DG) and the olfactory bulb are twoareas of the mammalian brain where neurogenesis continues tooccur throughout life (Eriksson et al., 1998). A recent study usingradioactive carbon dating reported a striking annual turnover rate of1.75% in the human DG where approximately 700 new neurons areadded every day (Spalding et al., 2013). These new neurons aregenerated at the subgranular zone (SGZ) of the DG and integrateinto the existing hippocampal circuitry, where they play afundamental role in learning and spatial memory formation byperforming pattern separation on inputs from the entorhinal cortex,an area of the brain located in the medial temporal lobe (McHughet al., 2007; Zhao et al., 2008; Aimone et al., 2011). Granule neuroncells are the most prevalent type of excitatory glutamatergic neuronin the DG, and their ongoing production in adults is regulated byphysical exercise, dietary restriction and enriched environmentthrough upregulation of brain-derived neurotrophic factor (BDNF)(van Praag et al., 1999; Rossi et al., 2006; Zhao et al., 2008).Aberrant hippocampal neurogenesis is implicated in a number of
neurological pathologies, and interventions aimed at regulatingneurogenesis have been proposed as potential therapeutic strategies(Jun et al., 2012). Epileptic seizures have been shown to affectneurogenesis and the emergence of abnormal hilar basal dendritesin newborn granule neurons. Normal granule neurons extenddendrites towards the molecular layer and project axons through thehilus. By contrast, dendrites from seizure-induced neurons growectopically towards the hippocampal hilus instead of the molecularlayer (Jessberger et al., 2007; Hattiangady and Shetty, 2010). Inaddition, a significant decline in neurogenesis is observed in theaging brain (Jessberger and Gage, 2008; Villeda et al., 2011;Spalding et al., 2013). These changes have been linked to depletionin the neural progenitor pool, owing to reduced Notch signaling, and
an increase in astrogliogenesis at the expense of producing neurons(Lugert et al., 2010; Encinas et al., 2011). Finally, hippocampalneurogenesis deficits have been linked to cognitive defectscharacteristic of depression (Patricio et al., 2013), impairment inearly Alzheimer’s disease pathology (Demars et al., 2010; Faureet al., 2011; Mu and Gage, 2011; Rodriguez et al., 2011), bipolardisorder (Valvezan and Klein, 2012; Walton et al., 2012) andschizophrenia (SCZD) (Tamminga et al., 2010; Walton et al., 2012;Hagihara et al., 2013). More specifically, alterations in DG granuleneuron maturation have been implicated in the etiology andpathogenesis of schizophrenia and bipolar disorder (see Box 1;Reif et al., 2006; Yamasaki et al., 2008; Walton et al., 2012;Hagihara et al., 2013; Shin et al., 2013).
To date, it has been difficult to investigate the role of hippocampalneurogenesis in the early stages of these CNS disorders in the humansystem, and it is not clear whether findings from studies usingrodent models will translate across species. Recent advances inreprogramming technology now offer a renewable source of humanneurons to investigate the basic biology of mechanisms ofpathogenesis using cells that harbor the same genetic backgroundand propensity for the disease as the patient. This technologyprovides the opportunity for drug screening using disease-relevantneurons in the hope that new therapeutics to target neurological
Box 1. Immature dentate gyrus in neuropsychiatricdiseasesSeveral studies have recently identified the ‘immature dentate gyrus’(iDG) as a conserved pathophysiological alteration that is found inneuropsychiatric disorders, including schizophrenia and bipolar disorder(Reif et al., 2006; Yamasaki et al., 2008; Walton et al., 2012; Hagiharaet al., 2013; Shin et al., 2013). The observation of iDG was first reportedin mice with a heterozygous mutation in the α-calcium/calmodulin-dependent protein kinase II (Camk2a) gene. These mice demonstratedcognitive and behavioral deficits similar to symptoms seen inschizophrenia, bipolar mood disorder and other psychiatric disorders(Yamasaki et al., 2008). The DG in these animals showed increased cellproliferation, hyper-excitability in DG granule cells and reduced numbersof calbindin+ mature neurons, suggesting a deficit in DG neurogenesis.Transcriptome analysis of the mutant hippocampi revealed significantchanges in the expression levels of more than 2000 genes, several ofwhich were also found to be expressed in a similar manner in 16 out of 18examined human post-mortem schizophrenia brains. Additional analysisof human cohorts of 15 schizophrenia and 15 bipolar disorder patientsamples revealed elevated numbers of calretinin+ immature NPCs at theexpense of calbindin+ mature neurons when compared with controlsubjects and with individuals suffering from major depression (Waltonet al., 2012). Despite these molecular insights, the contribution of iDG tothe cognitive impairments observed in these neuropsychiatric disordersremains unclear. One hypothesis is that the increased proportion ofimmature neurons in the DG causes deficits in pattern separation andhyper-activity in CA3-CA1, thereby conferring the psychosis observed inthe patients (Tamminga et al., 2012). However, much additional work isneeded to better understand the pathogenesis and functionalconsequences of iDG.
The Salk Institute for Biological Studies, 10010 North Torrey Pines Road, La Jolla,CA 92037, USA.
*Author for correspondence ([email protected])
2366
© 2014. Published by The Company of Biologists Ltd | Development (2014) 141, 2366-2375 doi:10.1242/dev.096776
DEVELO
PM
ENT

pathologies such as SCZD may be developed. In addition,reprogramming technology can be used to develop in vivo celltherapies (for a review, seeYuet al., 2013).However,muchadditionalwork is needed tomake cell transplantation a useful therapy.Althougha number of preclinical studies using transplantation of stem cell-derived neurons in animals models have demonstrated functionalimprovements (Lu et al., 2003; Roy et al., 2006; Wernig et al., 2008;Hargus et al., 2010; Jiang et al., 2011;Ma et al., 2012), elucidating theprecisemechanisms of functional recovery is still crucial for designingefficient and safe human trials.In this Primer, we will discuss the advances that have been made
in the past two decades to elucidate the process of DG developmentand identify the key regulators that drive DG granule neuronspecification. In addition, we will discuss how these developmentalprinciples have guided efforts to generate neural progenitor cells(NPCs) from pluripotent and somatic cells in vitro and, from these,mature DG granule neurons. Finally, we will discuss the currentlimitations and challenges of these approaches, as well as theirsuitability for disease modeling and therapeutic applications.
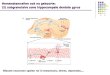
Early cortical patterning events during DG developmentThe formation of the central nervous system (CNS) requires preciseorchestrationof proliferation, cell fate determination anddifferentiation.Following gastrulation, cells derived from the mesoderm form thenotochord and define the primitive axis of the embryo. The notochordthen induces the formation of the neural plate, which later develops intothe neural tube during primary neurulation (Colas and Schoenwolf,2001). Following formation of the neural tube, neuroectodermal cellsalong the neural tube are patterned by gradients of signaling moleculesalong the dorsoventral and anterior-posterior axes to define regions ofthe forebrain, midbrain, hindbrain and spinal cord. The dorsal regionof the forebrain, called the telencephalon, gives rise to the cerebralcortex, which comprises the neocortex, plaeocortex and archicortex,also known as the hippocampus, whereas the ventral telencephalonforms the basal ganglia. Dorsoventral and rostrocaudal differences areestablished by diffusible morphogens released from distinct locations,the temporal and spatial dynamics of which are crucial in patterningthe distinct regions of the brain. Well-studied examples of thesemorphogens include fibroblast growth factor 8 (FGF8), which isproduced rostrally at the anterior neural ridge, wingless (WNT3A,WNT2B, WNT5A) and bone morphogenetic proteins (BMPs), whichare released dorsomedially by the cortical hem, and sonic hedgehog(SHH), which is released ventrally by the floor plate (Shimogori et al.,2004; Hebert and Fishell, 2008) (Fig. 1A). For the purpose ofdeveloping a differentiation protocol to generate the DG granuleneurons, we focus our discussion on the role of these factors in thegeneration and patterning of the hippocampal DG and cornu ammonis(CA) subfields.
Hippocampal and DG formationThe hippocampus arises from the caudomedial edge of the dorsaltelencephalic neuroepithelium adjacent to the cortical hem, a transientstructure found during development after the invagination ofthe telencephalic roof at the dorsal midline (Lee et al., 2000). Thehippocampal primordium is initially populated by Cajal-Retzius cellsand radial glial cells, which proliferate in the dentate neuroepithelium(DNe). As the radial glial cells proliferate to expand the tissue, theyproduce embryonic neural stem cells (NSCs) that will later migrate tothe SGZof theDG to give rise to the prospero homeobox 1 (PROX1)+
DG granule neurons (Altman and Bayer, 1990a,b) (Fig. 2). The DGhas a protracted developmental period that extends from embryonicday 13 (E13) to postnatal day 15 (P15) in mice, and begins in the
dorsomedial region of the telencephalic neuroepithelium that directlyabuts the cortical hem (Machon et al., 2007). The close proximity tothe cortical hem is the first indicator that WNT activity featuresprominently inDGdevelopment and that themedial-lateral gradient ofWNT signaling is essential for determining the cellular specificity inthe hippocampal DG and CA subfields. Indeed, a number of studieshave already demonstrated that WNT/β-catenin function is crucial forthe correct development of the hippocampus.Micedeficient inWnt3a,which encodes canonical WNT signaling protein expressed by thecortical hem, displayed severe defects in the size and organization ofhippocampal DG (Lee et al., 2000). A mutation in Lef1 (lymphoidenhancer binding factor 1), the nuclearmediator of theWNTpathway,resulted in impaired granule cell development (Galceran et al., 2000),whereas a deficiency in the WNT co-receptor LRP6 (low densitylipoprotein receptor-related protein 6) produced amarked reduction inthe number of Prox1+ granule neurons in both the migratory route tothe dentate and within the DG itself (Zhou et al., 2004). In additionto these loss-of-function phenotypes, gain-of-function approachesshowed that sustained canonical WNT activity in the lateral andmedial cortical primordium in D6-CLEF mutant mice resulted inectopic generation of Prox1+ cells in the dorsomedial cortex at theexpense of cortical neurons, providing further evidence that inductionof the DG granule cell fate requires high levels of canonical WNTsignaling (Machon et al., 2007).
In addition toWNT activity, SHH signaling has been shown to beinvolved in DG development, where it is important in expanding thegranule neural progenitor population during perinatal development(Machold et al., 2003). At early postnatal periods, expression of Shhand its receptor patched (Ptch1) can be found in CA3, the hilarregion of the DG and along the fimbria fibers that project to thehippocampus (Lai et al., 2003; Machold et al., 2003). Embryonicablation of smoothened (Smo), an essential component of theSHH signaling, resulted in abnormalities in regions of postnatalneurogenesis, including a reduction in the number of granule cells inDG relative to other regions of the hippocampus (Machold et al.,2003; Han et al., 2008). Pharmacological inhibition of SHHsignaling with cyclopamine also reduced the cell proliferation in theDG (Lai et al., 2003). Conversely, elevated hedgehog signalingproduced increased cell proliferation in the DG, further confirmingthe mitogenic effects of SHH in vivo (Lai et al., 2003; Macholdet al., 2003; Han et al., 2008). Interestingly, it appears that the SHHsignaling required for DG proliferation is mediated via primary cilia.Mice lacking cilia genes, for exampleKif3a (kinesin family member3A) and Ift88 (intraflagellar transport 88), showed defects inadult neural progenitors, but rescue of DG proliferation with aconstitutively activated Smo failed to restore neurogenesis in thesemutants (Han et al., 2008).
Although it is clear that certain morphogens such as SHH andWNT are important for DG development, it is essential to considerthe function of these pathways at different developmental timepoints. For example, for both WNT and SHH signaling, mutationsin these pathways during an earlier developmental window producemuch more substantial alterations in the regionalization of the brainthan if the perturbations were introduced at a later time. Disruptionof WNT or SHH signaling at E8-E10 in mice has been shown toresult in strong ventralization or dorsalization of the telencephalon,respectively, whereas changes after E12 brought about alterationsmostly attributed to a failure of neural progenitors to proliferateadequately (Chiang et al., 1996; Rallu et al., 2002; Backman et al.,2005). In addition, the interactions between the various signalingpathways active within the neurogenic niche must be closelyexamined. It has been shown that both SHH and WNT3A regulate
2367
PRIMER Development (2014) 141, 2366-2375 doi:10.1242/dev.096776
DEVELO
PM
ENT

the NSC population in the perinatal and adult DG throughSox2-dependent autocrine mechanisms (Favaro et al., 2009) andthat each may mediate the proliferation and differentiation indifferent populations of neural progenitors in the DG during earlypostnatal and adult neurogenesis (Choe and Pleasure, 2012). Thesenuances in the function of developmental cues need to be carefullyconsidered when taking on the challenge of recapitulating them inthe in vitro setting.
Transcriptional regulation of embryonic DG developmentWork is ongoing to elucidate the cellular and molecular mechanismsthat regulate the different stages of neurogenesis during embryonichippocampal development (Li and Pleasure, 2007). A number ofproneural genes encoding transcription factors of the basic helix loophelix (bHLH) class that were previously identified for their role inneocortical development have recently been shown to also mediate
cell fate specification and differentiation during DG neurogenesis(Fig. 3). Pro-neurogneic bHLH genes are activated downstream ofsignaling cues and have traditionally been divided into two groups:neuronal determination genes that are involved in the initialcommitment to the neuronal fate and neuronal differentiation genesthat regulate later events during differentiation to confer the identity ofthe terminally differentiated neuron (Pleasure et al., 2000).
Neuronal determination genesThe proneural transcription factor Ascl1 (achaete-scute complexhomolog 1;Mash1) was one of the first neuronal determination genesto be identified and is expressed in hippocampal progenitors duringdevelopment at different locations along the hippocampal primordium(Pleasure et al., 2000). However, although Ascl1 has been reported topromote acquisition of neuronal fate by inhibiting astrogliogenesis,Ascl1-null mice do not show significant defects in the proliferation of
Fig. 1. Endogenous mechanisms of embryonic hippocampal and DG development. Location of some key morphogen (A) and gene (B) expression inthe developing mouse telencephalon at E13.5. Additional Wnt activity also occurs along the rostrocaudal axis (not shown). Lhx2 gene expression (not shown)follows a highly similar pattern to Emx1/2. The hippocampus arises from the caudomedial edge of the dorsal telencephalic neuroepithelium adjacent to theCH. BMP, bone morphogenetic protein; CH, cortical hem; CTX, cortex; Dlx, distal-less homeobox; Emx, empty spiracles homeobox; FGF8, fibroblast growthfactor 8; FP, floor plate; LGE, lateral ganglionic eminence; MGE, medial ganglionic eminence; Nkx2.1, NK2 homeobox 1; Olig2, oligodendrocyte transcriptionfactor 2; Pax2, paired box 2; SHH, sonic hedgehog; WNT, wingless.
Fig. 2. Proliferation and migration of NSCs from the DNe region within the primordial hippocampus to form the laminar organization andcytoarchitecture of the postnatal DG.NSCs give rise to INPs, whichmigrate and eventually give rise toProx1+ granule neurons in the DG. In the postnatal brain,NSCs reside in the SGZ and continue to generate Prox1+ granule neurons. CA pyramidal neurons derive from the AN within the primordial hippocampus.TheCH, a transient structure formed during development, is the source of a number of patterningmorphogens, such asWNTandBMP proteins, which play crucialroles in cell fate specification during hippocampal neurogenesis. AN, ammonic neuroepithelium; CA, cornu ammonis; CH, cortical hem; CTX, cortex; D, dorsal;DNe, dentate neuroepithelium; DG, dentate gyrus; E, embryonic day; F, fimbria; INP, intermediate neural progenitor; L, lateral; LGE, lateral ganglionic eminence;M, medial; MGE, medial ganglionic eminence; NSC, neural stem cell; P, postnatal day; SGZ, sub-granular zone; V, ventral.
2368
PRIMER Development (2014) 141, 2366-2375 doi:10.1242/dev.096776
DEVELO
PM
ENT

neural progenitors or generation of granule neurons in the DG(Nieto et al., 2001). Interestingly, upregulation of Ascl1 expression inadult DG progenitors promotes oligodendrocyte fate acquisition(Jessberger et al., 2008). Similar to Ascl1, neurogenin 2 (Ngn2) hasbeen identified as a pro-neurogenic gene in cortical development, andits expression in the developing hippocampus overlaps with that ofAscl1 to a great extent (Galichet et al., 2008). However,Ngn2mutantmice have reduced numbers of DG neural progenitors in the DNe, thedentate migratory stream and the tertiary matrix, resulting inthe complete loss of the lower DG blade at birth. In addition,analysis of the radial glial cells in theDGofNgn2mutantmice showedan absence of GFAP+ radial glial fibers across the developing DG,likely contributing to the abnormal distribution of cells and DGmorphology at birth (Galichet et al., 2008).
Neuronal differentiation genesIn addition to regulating the proliferation of DG neural progenitors,bHLH transcription factors can modulate the dentate migratorystream. Emx2 (empty spiracles homeobox 2) is a neuroepithelialtranscription factor that was previously reported to play animportant role in the development of the telencephalon bydelineating the regionalization of the choroid plexus (von Froweinet al., 2006). However, it has been observed that Emx2mutant micealso lacked the entire DG granule cell layer at birth, although thecorresponding neuroepithelium appeared to be correctly specified
(Savaskan et al., 2002). Follow-up studies showed that, in additionto significant decreases in cell proliferation in the DNe, the dentatemigratory stream was almost entirely absent in the Emx2 mutants,resulting in the accumulation of Notch1+ dentate migratoryprecursor cells in the germinal layers around E15, and the ectopicpresence of Neurod1+ and Prox1+ cells in the ventricular andsubventricular zones at E18.5 (Oldekamp et al., 2004).
The T-box transcription factor Tbr2 (Eomes) has been found toregulate glutamatergic neuron fate commitment in several brainregions, including the DG (Hodge et al., 2012b). Loss of Tbr2 in theintermediate neuronal progenitor (INP) population during DGneurogenesis resulted in decreased granule cell neurogenesis due toINP depletion (Hodge et al., 2012b). This phenotype appears to besimilar to that observed in the embryonic neocortex, where the lossof Tbr2 in INPs was shown to affect INP proliferation, leading toreduced production of neocortical pyramidal neurons (Arnold et al.,2008; Sessa et al., 2008). In addition, Tbr2 has been shown topromote neuronal lineage progression from NSC to INP, in part viathe repression of Sox2 [SRY (sex determining region Y)-box 2],a key determinant of NSC identity (Hodge et al., 2012b).Interestingly, Tbr2 was recently found to also be expressed inCajal-Retzius cells in the developing DG, and it is important forestablishing and maintaining the radial processes of radial glial cellsto facilitate the migration of newborn neurons (Hodge et al., 2013).
The specification of DG granule neuronal fate is distinct fromother classes of DG or glutamatergic neurons and depends onactivity of Prox1, a homeobox transcription factor homolog of theDrosophila melanogaster gene prospero. While Prox1 expressioncan be transiently found in several regions of the embryonic brainduring development, its expression in the forebrain is highlyrestricted to the granule cell layer of the DG, starting at late gestationand persisting through adulthood (Torii et al., 1999; Lavado andOliver, 2007). Furthermore, even though there are overlaps in themechanisms that regulate cell fate and neuronal differentiationbetween the neocortex and hippocampal DG during development,the co-expression of Prox1 and Neurod1, a bHLH transcriptionfactor that is required for neuroblast differentiation, is uniquelylocalized to the DG in late gestation at E18.5 (Liu et al., 2000;Lavado and Oliver, 2007; Gao et al., 2009). More than just a marker,Prox1 expression has been shown to be crucial for DG granule cellfate and can even instruct DG granule neuron identity at the expenseof CA3 pyramidal neurons. In a recent study, overexpression ofProx1 in immature CA3 neurons resulted in the acquisition of DGgranule neuron properties, demonstrating the important role ofProx1 as a postmitotic cell fate determinant for DG granule cellsover CA3 pyramidal cells (Iwano et al., 2012). Indeed, Prox1 isessential for the maintenance of granule cell identity throughout life,as elimination of Prox1 in postmitotic immature DG neurons resultsin manifestation of CA3 pyramidal neuron properties, and thecontinual presence of Prox1 is required in mature DG cells forcorrect gene expression (Lavado et al., 2010; Iwano et al., 2012).
Temporal regulation of transcriptional networksSimilar to the differential roles observed for morphogens at differentdevelopmental stages, transcription factors in the hippocampus mayexert temporally distinct roles in the regulation of DG neurogenesisdepending on the developmental time point (Figs 1 and 3). Forexample, prior to E10.5, the LIM-homeodomain transcription factorLHX2 is important for defining the boundary of the cortical hem.Conditional knockout of Lhx2 results in an expanded cortical hemregion at the expense of the neocortical epithelium, as well as mis-expression of WNT3A that leads to the formation of ectopic
Fig. 3. The neurogenic niche at the SGZ comprises radial and horizontaltype 1 NSCs, early (type 2) and late (type 3) stage INPs, immature granuleneurons, andmature granule neurons.The progression from type 1NSCs tomature granule neurons is regulated by the sequential expression of keyneurogenic transcription factors during development. WNT3A and SHH aretwo extracellular signaling factors that have been previously shown to playimportant roles in the neuronal determination and differentiation of DG granuleneurons. INP, intermediate neural progenitor; NSC, neural stem cell; SGZ,sub-granular zone. ASCL1, achaete-scute complex homolog 1 (MASH1);DCX, doublecortin; EMX2, empty spiracles homeobox 2; FOXG1, forkheadbox G1; NEUN, RNA binding protein, fox-1 homolog 3; NGN2, neurogenin 2;PROX1, prospero homeobox 1; SHH, sonic hedgehog; SOX2, SRY (sexdetermining region Y) box 2; TBR2, T-box transcription factor; WNT, wingless.
2369
PRIMER Development (2014) 141, 2366-2375 doi:10.1242/dev.096776
DEVELO
PM
ENT

hippocampi (Mangale et al., 2008). However, at later stages, LHX2regulates the cell fate decisions of hippocampal progenitors.Downregulation of Lhx2 in the embryonic hippocampus wasfound to promote astrogliogenesis at the expense of neurogenesis,and overexpression of Lhx suppressed GFAP promoter activity,thereby favoring neuronal differentiation (Mangale et al., 2008).The temporal regulation of neurogenesis and DG granule neurondevelopment in particular must be considered to properly assess thecorrect specification of this cell type in vitro. Many studies haverevealed different transcription factors that are involved in thegeneration and maturation of the granule DG neuron [i.e. FOXG1(forkhead box G1), NEUROD1, TBR2, SOX11; for a review, seeHsieh, 2012). In addition, further characterization of temporaltranscription factor expression dynamics during the directeddifferentiation process may provide greater insights into theunderlying mechanisms involved in lineage specification andprogression during in vivo DG granule neuron development.
Specification of DG granule neurons in vitroThe most intuitive approach to specifying the DG granule cell identityin vitro is to follow a stepwise lineage progression through an NPCstage, as occurs during in vivo development. Not surprisingly, thepatterning cues that instruct NPC formation in vitro significantlyoverlap with those that have been identified in vivo. There have beenmultiple attempts to generate NPCs and neurons in vitro, mostly viadirected differentiation of pluripotent stem cells (PSCs) but also morerecently via direct reprogramming of somatic cells to NPCs andneurons via the forced activation of specific transcriptional networks(Kim et al., 2011; Lujan et al., 2012; Zhou and Tripathi, 2012;Wapinski et al., 2013; Ruggieri et al., 2014). These induced NPC andneuron (iNPC and iN) strategies can theoretically produce a morehomogenous cell population, and may provide an advantage fortranslational studies by reducing the tumorigenic risk associated withtransplanting pluripotent stem cell-derived cells in vivo. However, acurrent challenge for iNPC and iN approaches is the specification ofneuronal subtypes. With the exception of one study that showed directconversion of mouse and human fibroblasts into spinal motor neurons(Son et al., 2011), most direct reprogramming strategies producefunctional iNs that appear to belong tomultiple neuronal subtypes. In asingle culture, these may include dopaminergic neurons marked bytyrosine hydroxylase (TH), glutamatergic neuronsmarkedbyvesicularglutamate transporter, interneurons marked by γ-aminobutyric acid,and motor neurons marked by choline acetyltransferase (Ambasudhanet al., 2011; Pang et al., 2011; Qiang et al., 2011;Yoo et al., 2011). In asimilar way, neurons differentiated from iNPCs thus far also seem tocontainmultiple neuronal subtypes (Han et al., 2012;Ring et al., 2012;Wanget al., 2013;Zouet al., 2014). It is clear fromthese studies that themethod of NPC production and the precise patterning cues that areprovided have a direct bearing on the potential of the cells to reliablydifferentiate into specialized cell types. For this reason it is essential toevaluate the suitability of each of thesemethods not only in generatingNPCs but also for correctly specifying the NPC population.
Generating hippocampal NPCs from PSCs via directed differentiationThe unlimited self-renewal and largely unrestricted fate of PSCsmakes them ideal candidates for generating specialized cell typesin vitro. With the advent of iPSCs (Yamanaka and Takahashi,2006; Yamanaka, 2007), there is also the possibility of creatingdisease-specific PSCs, opening up unparalleled opportunities fordisease modeling and drug screening. Currently, there are only afew basic strategies for the directed differentiation of NPCs fromPSCs, all of which rely on subtle variations in early patterning cues
that occur during neuralization in vivo. Depending on exactlywhich patterning cues are provided in vitro, the resultingself-renewing NPC population can generate neurons enriched fora particular subtype, perhaps reflecting to some extent theendogenous NPC heterogeneity found in vivo (Perrier et al.,2004; Watanabe et al., 2005; Roy et al., 2006; Di Giorgio et al.,2008; Dimos et al., 2008; Eiraku et al., 2008; Gaspard et al., 2008;Marchetto et al., 2008; Kriks et al., 2011; Ma et al., 2011; Boyeret al., 2012; Shi et al., 2012; Vanderhaeghen, 2012; Maroof et al.,2013; Nicholas et al., 2013).
To specify a DG granule neuron subtype from NPCs, it istherefore essential to pattern the cells as per their normal in vivodevelopment. Recently, Yu and colleagues (2014) did just this,using an embryoid body approach to recapitulate key morphologicalcues that specify hippocampal DG identity during development.Previously, the formation of free-floating EBs under serum-freeconditions had been demonstrated to be particularly useful for thegeneration of naive telencephalic neural progenitors for the purposeof obtaining cortical neurons (Watanabe et al., 2005; Eiraku et al.,2008; Gaspard et al., 2008; Shi et al., 2012). These studiesdemonstrated that three-dimensional aggregates of the ESCs couldself-organize to form pallial tissue under serum-free conditions(Eiraku et al., 2008; Kadoshima et al., 2013). Furthermore, thisculture system could be patterned to enrich for telencephalon
Fig. 4. Generation of DG granule neurons from human pluripotent stemcells using embryoid bodies and monolayer neural progenitors cells.This strategy is based on the protocol reported by Yu and colleagues (2014)and represents the first complete differentiation protocol from pluripotent stemcells to mature DG granule neurons. BDNF, brain-derived neurotrophic factor;DG, dentate gyrus; DKK1, dickkopf homolog 1; FGF, fibroblast growth factor;TC, tissue culture.
2370
PRIMER Development (2014) 141, 2366-2375 doi:10.1242/dev.096776
DEVELO
PM
ENT

precursors by treatment with antagonists of WNT and NODALpathways (Watanabe et al., 2005), and directed to impose dorsalattributes via antagonism of SHH signaling (Gaspard et al., 2008)during the early stages of EB formation. Building on these studies,Yu and colleagues developed a protocol for DG granule neurondifferentiation that generated hippocampal NPCs, either entirely viaEB formation or via EB formation followed by NPC monolayerformation. To generate the NPCs, the authors first mimicked theanterior-posterior patterning of the forebrain with factors such asDKK1 (dickkopf homolog 1), NOGGIN and the TGFβ inhibitorSB431542, to instruct the formation of telencephalic neuralprecursors. Antagonism of SHH signaling further enriched fordorsal forebrain progenitors. Overall, the treatment regimepromoted upregulation of the pro-neurogenic genes found duringearly stages of DG neurogenesis, such as Sox2, Pax6 (paired box 6),Emx2 (empty spiracles homeobox 2) and Foxg1, which wereexpressed in a pattern that mimicked in vivo DG development(Fig. 4) (Yu et al., 2014). As will be discussed in the section belowon DG granule neuron specification, these cells gave rise to Prox1+
DG granule neural cells, demonstrating the importance ofrecapitulating key developmental pathways to correctly patternPSC-derived NPCs.
Generating mature DG granule neuronsTo date, there has been only one study demonstrating the in vitrogeneration of DG granule neurons (Yu et al., 2014). Following theinitial period of patterning designed to obtain the hippocampal NPCs,the cells were further treated with two key hippocampal neurogenicmolecules:WNT3A andBDNF.WNT3Ahad previously been shownto be important for the maintenance of hippocampal NPCs and theirdifferentiation into DG granule neurons, whereas BDNF is aneurotrophic factor that had been previously shown to promotehippocampal neurogenesis throughout life (Fig. 4) (Scharfman et al.,2005; Erickson et al., 2010). Quantitative real-time PCR (qPCR)analysis of whole populations of immature neurons patterned in thisway showed concomitant expression ofNeurod1 togetherwithProx1,as is uniquely found in the developing DG in vivo (Yu et al., 2014),although whetherNeurod1 and Prox1were co-expressed in immatureneurons at the single-cell level was not reported. Importantly,however, the mature neurons differentiated from these hippocampalNPCs triple-labeled for PROX1, a marker specific to DG granuleneurons in the hippocampus (Hodge et al., 2012a; Iwano et al., 2012),as well as MAP2AB (microtubule-associated protein 2) and NEUN(RBFOX3; RNA binding protein, fox-1 homolog 3), markers ofpostmitotic neurons (Fig. 5A) (Hodge and Hevner, 2011). Thesehuman DG granule neurons could be dissociated from the EBs andmaintained on co-cultures with hippocampal astrocytes for up to3 months in vitro. Further electrophysiology and calcium imaginganalyses demonstrated the maturation, connectivity and networkproperties of these cells in the astrocyte co-culture system (Fig. 5B).The isolated NPC population was also successfully transplanted intothe developing DG of P10 animals to produce electrophysiologicallyactive Prox1+ neurons. These neurons were able to functionallyintegrate into the endogenous neural circuitry and produce glutamine-mediated postsynaptic currents following stimulation of the perforantpath, the main afferent input into the DG from the entorhinal cortex(Fig. 5C) (Witter, 2007). The fact that the PSC-derived NPCs wereable to functionally integrate and produce DG granule neurons in vivosupports the hypothesis that the NPCs were correctly patternedin vitro and highlights the importance of recapitulating regionallyspecific patterning of NPCs to produce specific neuronal subtypesin vitro.
Conclusions and future perspectivesStem cell biology has opened up new avenues in the field ofneuroscience research as well as for therapeutic approaches forneurological diseases. Human ESCs are fundamental tools for thestudy of human neurodevelopment in an in vitro setting, providing
Fig. 5. Characterization of DG granule neurons generated in vitro. (A) Theoverlapping expression patterns of PROX1, NEUN and MAP2 (left) arecharacteristic of mature DG granule neurons. Additional markers (right) can beused to further assess the differentiation stage of the cells. (B) Co-culture ofimmature DG granule neurons on hippocampal astrocytes promotes theformation of functional neural networks after 3 weeks of differentiation, whichcan be assessed using fluorescent Ca2+ imaging (left). Somatic intracellularCa2+ dynamics calculated from Ca2+ traces (middle) panel obtained fromregions of interest (left panel) reflect neuronal activity patterns of hippocampalgranule and can be used to assess functional connectivity between neurons(right). (C) Transplantation of pre-patterned hippocampal NPCs into the DGof perinatal mice gives rise to functional neurons in the GCL that are properlyintegrated into the hippocampal neural circuitry. Electrophysiologicalproperties such as Na+/K+ currents, action potentials, and spontaneouspostsynaptic currents (depicted below) can be assessed to demonstrate thematurity of the grafted cells and allow comparison with the endogenousneuronal population. CA1, cornu ammonis 1; CA3, cornu ammonis 3; DCX,doublecortin; DG, dentate gyrus; EMX2, empty spiracles homeobox 2;FOXG1, forkhead box G1; GCL, granule cell layer; MAP2, microtubule-associated protein kinase 2; NPC, neural progenitor cell; NEUN, RNA bindingprotein, fox-1 homolog 3; PAX6, paired box 6; PROX1, prospero homeobox 1;TBR1, T-box brain gene 1.
2371
PRIMER Development (2014) 141, 2366-2375 doi:10.1242/dev.096776
DEVELO
PM
ENT

glimpses into crucial developmental windows via their differentiationprocess. In addition, cellular reprogramming has made it possible tomodify somatic cells of adults and neonates into human iPSCs, NPCsand even neurons, thus providing a renewable source of humanneurons that can be used to investigate the mechanism ofpathogenesis and assess the safety and efficacy of candidate drugs.Despite the promise of recent advances such as direct
reprogramming and directed differentiation, considerable hurdlesremain before cells produced via these strategies can be used fortherapeutic purposes. Chief among these is a deeper understandingof the precise molecular and functional attributes of the in vitro-derived cells. In the example of generating the DG granule neurons,an ideal approach would require single-cell expression profilecharacterization from in vitro-derived cells followed by comparisonwith endogenous newborn and adult granule neuron cell profilesfrom living animals. Further immunocytochemistry at selecttimepoints may provide additional evidence for the developmentalprogression of PSC-derived DG granule neurons. Such comparisonscould help to identify specific populations that correspond todistinct DG granule neuron developmental timepoints in vivo, andenable selection of these cell types in vitro. Another tool forcharacterizing cellular subtypes, translating ribosome affinitypurification (TRAP) has been reported to be useful for obtainingthe translated mRNA profiles in genetically defined cell populations(Doyle et al., 2008; Heiman et al., 2008), and has recently facilitatedthe identification cortical neuron subtypes that mediate responses tochronic antidepressant treatments (Schmidt et al., 2012). Indeed,characterization of neuronal subtypes, both of endogenous originand their counterparts produced in vitro, is an ongoing effort thatwill continue to shape the stem cell field by redefining the conceptof cellular plasticity and the properties, both cell-autonomous andnon-cell-autonomous, that confer cellular subtype specificity.As cognitive dysfunction associated with aberrant hippocampal
neurogenesis has been implicated in CNS disorders, generation of DGgranule neurons from patient cells may offer a unique opportunity fordisease modeling and drug discovery (Jun et al., 2012). Althoughtranslation of these findings to a clinical setting is still far away, it is stillpossible to gain insight into the physiological processes thatmanifest incertain neuropathological diseases, perhaps hinting at possible targetsfor pharmaceutical intervention. The recent studybyYuand colleaguesreported reduced neuronal network activity by calcium imaging andelectrophysiological recording in DG granule neurons generated fromreprogrammed schizophrenia patient-derived fibroblasts, comparedwith the same neuronal sub-type generated from normal humanfibroblasts. Importantly, the disease phenotypewas not observed undera pan-neuronal differentiation protocol, but only manifested whenthe correct neuronal sub-type was specified (Brennand et al., 2011;Yu et al., 2014) (see Box 1 on role of DG granule neurons inschizophrenia). This finding supports the concept that selectiveenrichment of clinically relevant neuronal subtypes is an importantstrategy to improve the accuracy of disease modelling. As ourunderstanding of neuronal sub-type specification continues to evolvewith new insights gained from developmental studies, so too must thedifferentiation strategy used to generate the desired cell type. Forexample, recent work from Choe and colleagues showed that BMP7, amember of the bone morphogenetic proteins family, is crucial forinitiating neurogenesis in the DG progenitors by signaling through theactivin receptor type I (ACVR1) and inducing Lef1 expression (Choeet al., 2013). As both BMP7 and WNT3A are present in highconcentrations and mediate the Lef1 expression during DGdevelopment, it would be very interesting to examine the potentiallysynergistic effects of these two molecules on the production of Tbr2+
intermediate neural progenitors and the subsequent Prox1+ neuronsduring DG granule neuron differentiation in vitro.
In this Primer, we have discussed the development and design ofapproaches to generate hippocampal DG granule neurons in vitrofor the purposes of disease modeling and drug screening. Thedifferentiation process recapitulates important aspects of DGneurogenesis and provides the opportunity to examine the etiologyand pathogenesis of diseases in developmental time. However, this isonly the first step in the process of developing an even more completeplatform to model the complexity of the dynamic connections thatexist between different classes of neurons in the hippocampus nicheand to investigate the cellular and molecular mechanisms underlyinghippocampus-related disorders. The fundamental principle ofrecapitulating DG developmental cues to generate DG granuleneurons could also be used to develop additional differentiationapproaches to produce the various subtypes of pyramidal neuronsfound in the CA subfields. If multiple different neuronal subtypes canbe successfully generated in vitro, then this may provide theopportunity to recapitulate neural circuits via engineered platforms,e.g. by using microfluidics devices with microfabricated substrates(see Box 2). This approach would facilitate the dissection of thefunction and interactions between subpopulations of neurons under
Box 2. Recapitulating endogenous neural circuitryin vitro
Although two-dimensional cell culture techniques have been fundamentalto cell biology, drug discovery and tissue engineering, they have beenunable to fully recapitulate the complex and dynamic three-dimensional(3D) environment of the living tissue in vivo. Microfluidics technology canprovide an engineered platform for 3D cell culture that offers precisecontrol over the individual cell microenvironment with high reproducibility.Microfluidic devices fabricated from oxygen-permeable materials suchas polydimethylsiloxane (PDMS) can support long-term neural culturesand preserve compartmentalization of different cell types, whileoccupying less space and using significantly fewer reagents thantraditional tissue culture techniques. These factors make microfluidiccell culture technology ideal for generating specific cell types frompluripotent stem cells for the purposes of disease modeling and drugscreening. The microscale dimensions of each individual microchannelare comparable to in vivo cytoarchitectural dimensions and can be usedto create multiple chemical gradients to simulate endogenous autocrineand paracrine signaling cues. In addition, micropatterning usingbiomaterials, e.g. collagen or laminin, allows for isolation of dendritesand axons, as well as compartmentalization of cellular subtypes. Suchsophisticated micropatterning and microfluidics can create highlyorganized structures that mimic the organization of the correspondingendogenous tissue or organ. For example, chemical gradients of nervegrowth factor delivered viamicrofluidics canbeused toorient thedirectionof neurite outgrowth in 3D micropatterned neuronal cultures (Kunzeet al., 2011). Micropatterned cultures have also been used to supportneuron-astrocyte co-cultures for disease modeling (Kunze et al., 2013).
2372
PRIMER Development (2014) 141, 2366-2375 doi:10.1242/dev.096776
DEVELO
PM
ENT

physiological and pathophysiological conditions (for additionalreading, see Yu et al., 2013). Such high-resolution in vitromodeling of neural circuitry will allow unprecedented control andinsight into how themammalian brain functions and how this functionis derailed during the progression of neuropathological disease.
AcknowledgementsThe authors thank M. L. Gage for editorial comments.
Competing interestsThe authors declare no competing financial interests.
FundingThis work was supported by the CIRM, by the Annette C. Merle-Smith and MathersFoundation, by The Leona M. and Harry B. Helmsley Charitable Trust, by Sanofi-Aventis, by JPB Foundation, and by generous donations fromMary Jane and RobertEngman.
ReferencesAimone, J. B., Deng, W. and Gage, F. (2011). Resolving new memories: a criticallook at the dentate gyrus, adult neurogenesis, and pattern separation. Neuron 70,589-596.
Altman, J. andBayer, S. A. (1990a). Migration and distribution of two populations ofhippocampal granule cell precursors during the perinatal and postnatal periods.J. Comp. Neurol. 301, 365-381.
Altman, J. and Bayer, S. A. (1990b). Mosaic organization of the hippocampalneuroepithelium and the multiple germinal sources of dentate granule cells.J. Comp. Neurol. 301, 325-342.
Ambasudhan, R., Talantova, M., Coleman, R., Yuan, X., Zhu, S., Lipton, S. A.and Ding, S. (2011). Direct reprogramming of adult human fibroblasts tofunctional neurons under defined conditions. Cell Stem Cell 9, 113-118.
Arnold, S. J., Huang, G.-J., Cheung, A. F. P., Era, T., Nishikawa, S.-I., Bikoff, E.K., Molnar, Z., Robertson, E. J. and Groszer, M. (2008). The T-box transcriptionfactor Eomes/Tbr2 regulates neurogenesis in the cortical subventricular zone.Genes Dev. 22, 2479-2484.
Backman, M., Machon, O., Mygland, L., van den Bout, C. J., Zhong, W., Taketo,M. M. and Krauss, S. (2005). Effects of canonical Wnt signaling on dorso-ventralspecification of the mouse telencephalon. Dev. Biol. 279, 155-168.
Boyer, L. F., Campbell, B., Larkin, S., Mu, Y. and Gage, F. H. (2012).Dopaminergic differentiation of human pluripotent cells. Curr. Protoc. Stem CellBiol. Chapter 1, Unit1H 6.
Brennand, K. J., Simone, A., Jou, J., Gelboin-Burkhart, C., Tran, N., Sangar, S.,Li, Y., Mu, Y., Chen, G., Yu, D. et al. (2011). Modelling schizophrenia usinghuman induced pluripotent stem cells. Nature 473, 221-225.
Chiang, C., Litingtung, Y., Lee, E., Young, K. E., Corden, J. L., Westphal, H. andBeachy, P. A. (1996). Cyclopia and defective axial patterning in mice lackingSonic hedgehog gene function. Nature 383, 407-413.
Choe, Y. andPleasure, S. J. (2012).Wnt signaling regulates intermediate precursorproduction in the postnatal dentate gyrus by regulating CXCR4 expression. Dev.Neurosci. 34, 502-514.
Choe, Y., Kozlova, A., Graf, D. and Pleasure, S. J. (2013). Bone morphogenicprotein signaling is a major determinant of dentate development. J. Neurosci. 33,6766-6775.
Colas, J.-F. and Schoenwolf, G. C. (2001). Towards a cellular and molecularunderstanding of neurulation. Dev. Dyn. 221, 117-145.
Demars, M., Hu, Y.-S., Gadadhar, A. and Lazarov, O. (2010). Impairedneurogenesis is an early event in the etiology of familial Alzheimer’s disease intransgenic mice. J. Neurosci. Res. 88, 2103-2117.
Di Giorgio, F. P., Boulting, G. L., Bobrowicz, S. and Eggan, K. C. (2008). Humanembryonic stem cell-derived motor neurons are sensitive to the toxic effect of glialcells carrying an ALS-causing mutation. Cell Stem Cell 3, 637-648.
Dimos, J. T., Rodolfa, K. T., Niakan, K. K., Weisenthal, L. M., Mitsumoto, H.,Chung, W., Croft, G. F., Saphier, G., Leibel, R., Goland, R. et al. (2008).Induced pluripotent stem cells generated from patients with ALS can bedifferentiated into motor neurons. Science 321, 1218-1221.
Doyle, J. P., Dougherty, J. D., Heiman, M., Schmidt, E. F., Stevens, T. R., Ma, G.,Bupp, S., Shrestha, P., Shah, R. D., Doughty, M. L. et al. (2008). Application ofa translational profiling approach for the comparative analysis of CNS cell types.Cell 135, 749-762.
Eiraku, M., Watanabe, K., Matsuo-Takasaki, M., Kawada, M., Yonemura, S.,Matsumura, M., Wataya, T., Nishiyama, A., Muguruma, K. and Sasai, Y.(2008). Self-organized formation of polarized cortical tissues from ESCs and itsactive manipulation by extrinsic signals. Cell Stem Cell 3, 519-532.
Encinas, J. M., Michurina, T. V., Peunova, N., Park, J.-H., Tordo, J., Peterson,D. A., Fishell, G., Koulakov, A. and Enikolopov, G. (2011). Division-coupledastrocytic differentiation and age-related depletion of neural stem cells in the adulthippocampus. Cell Stem Cell 8, 566-579.
Erickson, K. I., Prakash, R. S., Voss, M. W., Chaddock, L., Heo, S., McLaren, M.,Pence, B. D., Martin, S. A., Vieira, V. J.,Woods, J. A. et al. (2010). Brain-derivedneurotrophic factor is associated with age-related decline in hippocampal volume.J. Neurosci. 30, 5368-5375.
Eriksson, P. S., Perfilieva, E., Bjork-Eriksson, T., Alborn, A.-M., Nordborg, C.,Peterson, D. A. and Gage, F. H. (1998). Neurogenesis in the adult humanhippocampus. Nat. Med. 4, 1313-1317.
Faure, A., Verret, L., Bozon, B., El Tannir El Tayara, N., Ly, M., Kober, F.,Dhenain, M., Rampon, C. and Delatour, B. (2011). Impaired neurogenesis,neuronal loss, and brain functional deficits in the APPxPS1-Ki mouse model ofAlzheimer’s disease. Neurobiol. Aging 32, 407-418.
Favaro, R., Valotta, M., Ferri, A. L. M., Latorre, E., Mariani, J., Giachino, C.,Lancini, C., Tosetti, V., Ottolenghi, S., Taylor, V. et al. (2009). Hippocampaldevelopment and neural stem cell maintenance require Sox2-dependentregulation of Shh. Nat. Neurosci. 12, 1248-1256.
Galceran, J., Miyashita-Lin, E. M., Devaney, E., Rubenstein, J. L. andGrosschedl, R. (2000). Hippocampus development and generation of dentategyrus granule cells is regulated by LEF1. Development 127, 469-482.
Galichet, C., Guillemot, F. and Parras, C. M. (2008). Neurogenin 2 has anessential role in development of the dentate gyrus.Development 135, 2031-2041.
Gao, Z., Ure, K., Ables, J. L., Lagace, D. C., Nave, K.-A., Goebbels, S., Eisch, A. J.and Hsieh, J. (2009). Neurod1 is essential for the survival and maturation of adult-born neurons. Nat. Neurosci. 12, 1090-1092.
Gaspard, N., Bouschet, T., Hourez, R., Dimidschstein, J., Naeije, G., van denAmeele, J., Espuny-Camacho, I., Herpoel, A., Passante, L., Schiffmann, S. N.et al. (2008). An intrinsic mechanism of corticogenesis from embryonic stem cells.Nature 455, 351-357.
Hagihara, H., Takao, K., Walton, N. M., Matsumoto, M. and Miyakawa, T. (2013).Immature dentate gyrus: an endophenotype of neuropsychiatric disorders.NeuralPlast. 2013, 318596.
Han, Y.-G., Spassky, N., Romaguera-Ros, M., Garcia-Verdugo, J.-M., Aguilar, A.,Schneider-Maunoury, S. and Alvarez-Buylla, A. (2008). Hedgehog signalingand primary cilia are required for the formation of adult neural stem cells. Nat.Neurosci. 11, 277-284.
Han, D. W., Tapia, N., Hermann, A., Hemmer, K., Hoing, S., Arauzo-Bravo, M. J.,Zaehres, H., Wu, G., Frank, S., Moritz, S. et al. (2012). Direct reprogramming offibroblasts into neural stem cells by defined factors. Cell Stem Cell 10, 465-472.
Hargus, G., Cooper, O., Deleidi, M., Levy, A., Lee, K., Marlow, E., Yow, A.,Soldner, F., Hockemeyer, D., Hallett, P. J. et al. (2010). Differentiated Parkinsonpatient-derived induced pluripotent stem cells grow in the adult rodent brain andreduce motor asymmetry in Parkinsonian rats. Proc. Natl. Acad. Sci. U.S.A. 107,15921-15926.
Hattiangady, B. and Shetty, A. K. (2010). Decreased neuronal differentiation ofnewly generated cells underlies reduced hippocampal neurogenesis in chronictemporal lobe epilepsy. Hippocampus 20, 97-112.
Hebert, J. M. and Fishell, G. (2008). The genetics of early telencephalonpatterning: some assembly required. Nat. Rev. Neurosci. 9, 678-685.
Heiman, M., Schaefer, A., Gong, S., Peterson, J. D., Day, M., Ramsey, K. E.,Suarez-Farinas, M., Schwarz, C., Stephan, D. A., Surmeier, D. J. et al. (2008).A translational profiling approach for the molecular characterization of CNS celltypes. Cell 135, 738-748.
Hodge, R. D. and Hevner, R. F. (2011). Expression and actions of transcriptionfactors in adult hippocampal neurogenesis. Dev. Neurobiol. 71, 680-689.
Hodge, R. D., Kahoud, R. J. and Hevner, R. F. (2012a). Transcriptional control ofglutamatergic differentiation during adult neurogenesis. Cell. Mol. Life Sci. 69,2125-2135.
Hodge, R. D., Nelson, B. R., Kahoud, R. J., Yang, R., Mussar, K. E., Reiner, S. L.and Hevner, R. F. (2012b). Tbr2 is essential for hippocampal lineage progressionfrom neural stem cells to intermediate progenitors and neurons. J. Neurosci. 32,6275-6287.
Hodge, R. D., Garcia, A. J., III, Elsen, G. E., Nelson, B. R., Mussar, K. E., Reiner,S. L., Ramirez, J.-M. and Hevner, R. F. (2013). Tbr2 expression in Cajal-Retziuscells and intermediate neuronal progenitors is required for morphogenesis of thedentate gyrus. J. Neurosci. 33, 4165-4180.
Hsieh, J. (2012). Orchestrating transcriptional control of adult neurogenesis.GenesDev. 26, 1010-1021.
Iwano, T., Masuda, A., Kiyonari, H., Enomoto, H. andMatsuzaki, F. (2012). Prox1postmitotically defines dentate gyrus cells by specifying granule cell identity overCA3 pyramidal cell fate in the hippocampus. Development 139, 3051-3062.
Jessberger, S. and Gage, F. H. (2008). Stem-cell-associated structural andfunctional plasticity in the aging hippocampus. Psychol. Aging 23, 684-691.
Jessberger, S., Zhao, C., Toni, N., Clemenson, G. D., Li, Y. and Gage, F. H.(2007). Seizure-associated, aberrant neurogenesis in adult rats characterizedwith retrovirus-mediated cell labeling. J. Neurosci. 27, 9400-9407.
Jessberger, S., Toni, N., Clemenson, G. D., Jr, Ray, J. and Gage, F. H. (2008).Directed differentiation of hippocampal stem/progenitor cells in the adult brain.Nat. Neurosci. 11, 888-893.
Jiang, M., Lv, L., Ji, H., Yang, X., Zhu, W., Cai, L., Gu, X., Chai, C., Huang, S.,Sun, J. et al. (2011). Induction of pluripotent stem cells transplantation therapy forischemic stroke. Mol. Cell. Biochem. 354, 67-75.
2373
PRIMER Development (2014) 141, 2366-2375 doi:10.1242/dev.096776
DEVELO
PM
ENT

Jun, H., Mohammed Qasim Hussaini, S., Rigby, M. J. and Jang, M.-H. (2012).Functional role of adult hippocampal neurogenesis as a therapeutic strategy formental disorders. Neural Plast. 2012, 854285.
Kadoshima, T., Sakaguchi, H., Nakano, T., Soen, M., Ando, S., Eiraku, M. andSasai, Y. (2013). Self-organization of axial polarity, inside-out layer pattern, andspecies-specific progenitor dynamics in human ES cell-derived neocortex. Proc.Natl. Acad. Sci. U.S.A. 110, 20284-20289.
Kim, J., Efe, J. A., Zhu, S., Talantova, M., Yuan, X., Wang, S., Lipton, S. A.,Zhang, K. and Ding, S. (2011). Direct reprogramming of mouse fibroblasts toneural progenitors. Proc. Natl. Acad. Sci. U.S.A. 108, 7838-7843.
Kriks, S., Shim, J. W., Piao, J., Ganat, Y. M., Wakeman, D. R., Xie, Z., Carrillo-Reid, L., Auyeung, G., Antonacci, C., Buch, A. et al. (2011). Dopamine neuronsderived from human ES cells efficiently engraft in animal models of Parkinson’sdisease. Nature 480, 547-551.
Kunze, A., Giugliano, M., Valero, A. and Renaud, P. (2011). Micropatterningneural cell cultures in 3D with a multi-layered scaffold. Biomaterials 32,2088-2098.
Kunze, A., Lengacher, S., Dirren, E., Aebischer, P., Magistretti, P. J. andRenaud, P. (2013). Astrocyte-neuron co-culture on microchips based on themodel of SOD mutation to mimic ALS. Integr. Biol. 5, 964-975.
Lai, K., Kaspar, B. K., Gage, F. H. and Schaffer, D. V. (2003). Sonic hedgehogregulates adult neural progenitor proliferation in vitro and in vivo.Nat. Neurosci. 6,21-27.
Lavado, A. and Oliver, G. (2007). Prox1 expression patterns in the developing andadult murine brain. Dev. Dyn. 236, 518-524.
Lavado, A., Lagutin, O. V., Chow, L. M. L., Baker, S. J. and Oliver, G. (2010).Prox1 is required for granule cell maturation and intermediate progenitormaintenance during brain neurogenesis. PLoS Biol. 8, e1000460.
Lee, S. M., Tole, S.,, Grove, E. and and McMahon, A. P. (2000). A local Wnt-3asignal is required for development of the mammalian hippocampus.Development127, 457-467.
Li, G. and Pleasure, S. J. (2007). Genetic regulation of dentate gyrusmorphogenesis. Prog. Brain Res. 163, 143-152.
Liu, M., Pleasure, S. J., Collins, A. E., Noebels, J. L., Naya, F. J., Tsai, M.-J. andLowenstein, D. H. (2000). Loss of BETA2/NeuroD leads to malformation of thedentate gyrus and epilepsy. Proc. Natl. Acad. Sci. U.S.A. 97, 865-870.
Lu, P., Jones, L. L., Snyder, E. Y. and Tuszynski, M. H. (2003). Neural stem cellsconstitutively secrete neurotrophic factors and promote extensive host axonalgrowth after spinal cord injury. Exp. Neurol. 181, 115-129.
Lugert, S., Basak, O., Knuckles, P., Haussler, U., Fabel, K., Gotz, M., Haas, C. A.,Kempermann, G., Taylor, V. and Giachino, C. (2010). Quiescent and activehippocampal neural stem cells with distinct morphologies respond selectively tophysiological and pathological stimuli and aging. Cell Stem Cell 6, 445-456.
Lujan, E., Chanda, S., Ahlenius, H., Sudhof, T. C. and Wernig, M. (2012). Directconversion of mouse fibroblasts to self-renewing, tripotent neural precursor cells.Proc. Natl. Acad. Sci. U.S.A. 109, 2527-2532.
Ma, L., Liu, Y. and Zhang, S.-C. (2011). Directed differentiation of dopamineneurons from human pluripotent stem cells. Methods Mol. Biol. 767, 411-418.
Ma, L., Hu, B., Liu, Y., Vermilyea, S. C., Liu, H., Gao, L., Sun, Y., Zhang, X. andZhang, S.-C. (2012). Human embryonic stem cell-derived GABA neurons correctlocomotion deficits in quinolinic acid-lesioned mice. Cell Stem Cell 10, 455-464.
Machold, R., Hayashi, S., Rutlin, M., Muzumdar, M. D., Nery, S., Corbin, J. G.,Gritli-Linde, A., Dellovade, T., Porter, J. A., Rubin, L. L. et al. (2003). Sonichedgehog is required for progenitor cell maintenance in telencephalic stem cellniches. Neuron 39, 937-950.
Machon, O., Backman, M., Machonova, O., Kozmik, Z., Vacik, T., Andersen, L.and Krauss, S. (2007). A dynamic gradient of Wnt signaling controls initiation ofneurogenesis in the mammalian cortex and cellular specification in thehippocampus. Dev. Biol. 311, 223-237.
Mangale, V. S., Hirokawa, K. E., Satyaki, P. R. V., Gokulchandran, N., Chikbire, S.,Subramanian, L., Shetty, A. S., Martynoga, B., Paul, J., Mai, M. V. et al. (2008).Lhx2 selector activity specifies cortical identity and suppresses hippocampalorganizer fate. Science 319, 304-309.
Marchetto, M. C. N., Muotri, A. R., Mu, Y., Smith, A. M., Cezar, G. G. and Gage, F.H. (2008). Non-cell-autonomous effect of human SOD1 G37R astrocytes onmotor neurons derived from human embryonic stem cells. Cell Stem Cell 3,649-657.
Maroof, A. M., Keros, S., Tyson, J. A., Ying, S.-W., Ganat, Y. M., Merkle, F. T.,Liu, B., Goulburn, A., Stanley, E. G., Elefanty, A. G. et al. (2013). Directeddifferentiation and functional maturation of cortical interneurons from humanembryonic stem cells. Cell Stem Cell 12, 559-572.
McHugh,T.J., Jones,M.W.,Quinn,J.J.,Balthasar,N.,Coppari,R.,Elmquist, J.K.,Lowell, B. B., Fanselow, M. S., Wilson, M. A. and Tonegawa, S. (2007). DentategyrusNMDAreceptorsmediate rapidpattern separation in thehippocampalnetwork.Science 317, 94-99.
Mu, Y. and Gage, F. H. (2011). Adult hippocampal neurogenesis and its role inAlzheimer’s disease. Mol. Neurodegener. 6, 85.
Nicholas, C. R., Chen, J., Tang, Y., Southwell, D. G., Chalmers, N., Vogt, D.,Arnold, C. M., Chen, Y.-J., Stanley, E. G., Elefanty, A. G. et al. (2013).Functional maturation of hPSC-derived forebrain interneurons requires an
extended timeline and mimics human neural development. Cell Stem Cell 12,573-586.
Nieto, M., Schuurmans, C., Britz, O. andGuillemot, F.(2001). Neural bHLH genescontrol the neuronal versus glial fate decision in cortical progenitors. Neuron 29,401-413.
Oldekamp, J., Kraemer, N., Alvarez-Bolado, G. and Skutella, T. (2004). bHLHgene expression in the Emx2-deficient dentate gyrus reveals defective granulecells and absence of migrating precursors. Cereb. Cortex 14, 1045-1058.
Pang, Z. P., Yang, N.,, Vierbuchen, T., Ostermeier, , A., Fuentes, D. R., Yang, T.Q., Citri, A., Sebastiano, V., Marro, S., Sudhof, T. C. et al. (2011). Induction ofhuman neuronal cells by defined transcription factors. Nature 476, 220-223.
Patricio, P., Mateus-Pinheiro, A., Sousa, N. and Pinto, L. (2013). Re-cyclingparadigms: cell cycle regulation in adult hippocampal neurogenesis andimplications for depression. Mol. Neurobiol. 48, 84-96.
Perrier, A. L., Tabar, V., Barberi, T., Rubio, M. E., Bruses, J., Topf, N., Harrison,N. L. and Studer, L. (2004). Derivation of midbrain dopamine neurons fromhuman embryonic stem cells. Proc. Natl. Acad. Sci. U.S.A. 101, 12543-12548.
Pleasure, S. J., Collins, A. E. and Lowenstein, D. H. (2000). Unique expressionpatterns of cell fate molecules delineate sequential stages of dentate gyrusdevelopment. J. Neurosci. 20, 6095-6105.
Qiang, L., Fujita, R., Yamashita, T., Angulo, S., Rhinn, H., Rhee, D., Doege, C.,Chau, L., Aubry, L., Vanti, W. B. et al. (2011). Directed conversion ofAlzheimer’s disease patient skin fibroblasts into functional neurons. Cell 146,359-371.
Rallu, M., Machold, R., Gaiano, N., Corbin, J. G., McMahon, A. P. and Fishell, G.(2002). Dorsoventral patterning is established in the telencephalon of mutantslacking both Gli3 and Hedgehog signaling. Development 129, 4963-4974.
Reif, A., Fritzen, S., Finger, M., Strobel, A., Lauer, M., Schmitt, A. and Lesch, K.-P. (2006). Neural stem cell proliferation is decreased in schizophrenia, but not indepression. Mol. Psychiatry 11, 514-522.
Ring, K. L., Tong, L. M., Balestra, M. E., Javier, R., Andrews-Zwilling, Y., Li, G.,Walker, D., Zhang, W. R., Kreitzer, A. C. and Huang, Y. (2012). Directreprogramming of mouse and human fibroblasts into multipotent neural stem cellswith a single factor. Cell Stem Cell 11, 100-109.
Rodriguez, J. J., Noristani, H. N., Olabarria, M., Fletcher, J., Somerville, T. D. D.,, Yeh, C. Y. and Verkhratsky, A. (2011). Voluntary running and environmentalenrichment restores impaired hippocampal neurogenesis in a triple transgenicmouse model of Alzheimer’s disease. Curr. Alzheimer Res. 8, 707-717.
Rossi, C., Angelucci, A., Costantin, L., Braschi, C., Mazzantini, M., Babbini, F.,Fabbri, M. E., Tessarollo, L., Maffei, L., Berardi, N. et al. (2006). Brain-derivedneurotrophic factor (BDNF) is required for the enhancement of hippocampalneurogenesis following environmental enrichment. Eur. J. Neurosci. 24,1850-1856.
Roy, N. S., Cleren, C., Singh, S. K., Yang, L., Beal, M. F. and Goldman, S. A.(2006). Functional engraftment of human ES cell-derived dopaminergic neuronsenriched by coculture with telomerase-immortalized midbrain astrocytes. Nat.Med. 12, 1259-1268.
Ruggieri, M., Riboldi, G., Brajkovic, S., Bucchia, M., Bresolin, N., Comi, G. P.and Corti, S. (2014). Induced neural stem cells: methods of reprogramming andpotential therapeutic applications. Prog. Neurobiol. 114, 15-24.
Savaskan, N. E., Alvarez-Bolado, G., Glumm, R., Nitsch, R., Skutella, T. andHeimrich, B. (2002). Impaired postnatal development of hippocampal neuronsand axon projections in the Emx2-/- mutants. J. Neurochem. 83, 1196-1207.
Scharfman, H., Goodman, J., Macleod, A., Phani, S., Antonelli, C. and Croll, S.(2005). Increased neurogenesis and the ectopic granule cells afterintrahippocampal BDNF infusion in adult rats. Exp. Neurol. 192, 348-356.
Schmidt, E. F., Warner-Schmidt, J. L., Otopalik, B. G., Pickett, S. B., Greengard,P. and Heintz, N. (2012). Identification of the cortical neurons that mediateantidepressant responses. Cell 149, 1152-1163.
Sessa, A., Mao, C.-a., Hadjantonakis, A.-K., Klein, W. H. and Broccoli, V. (2008).Tbr2 directs conversion of radial glia into basal precursors and guides neuronalamplification by indirect neurogenesis in the developing neocortex. Neuron 60,56-69.
Shi, Y., Kirwan, P. and Livesey, F. J. (2012). Directed differentiation of humanpluripotent stem cells to cerebral cortex neurons and neural networks.Nat. Protoc.7, 1836-1846.
Shimogori, T., Banuchi, V., Ng, H. Y., Strauss, J. B. and Grove, E. A. (2004).Embryonic signaling centers expressing BMP, WNT and FGF proteins interact topattern the cerebral cortex. Development 131, 5639-5647.
Shin, R., Kobayashi, K., Hagihara, H., Kogan, J. H., Miyake, S., Tajinda, K.,Walton, N. M., Gross, A. K., Heusner, C. L., Chen, Q. et al. (2013). Theimmature dentate gyrus represents a shared phenotype of mouse models ofepilepsy and psychiatric disease. Bipolar Disord. 15, 405-421.
Son, E. Y., Ichida, J. K., Wainger, B. J., Toma, J. S., Rafuse, V. F.,Woolf, C. J. andEggan, K. (2011). Conversion of mouse and human fibroblasts into functionalspinal motor neurons. Cell Stem Cell 9, 205-218.
Spalding, K. L., Bergmann, O., Alkass, K., Bernard, S., Salehpour, M., Huttner,H. B., Bostrom, E., Westerlund, I., Vial, C., Buchholz, B. A. et al. (2013).Dynamics of hippocampal neurogenesis in adult humans. Cell 153, 1219-1227.
2374
PRIMER Development (2014) 141, 2366-2375 doi:10.1242/dev.096776
DEVELO
PM
ENT

Tamminga, C. A., Stan, A. D. and Wagner, A. D. (2010). The hippocampalformation in schizophrenia. Am. J. Psychiatry. 167, 1178-1193.
Tamminga,C.A., Thomas,B.P.,Chin,R.,Mihalakos,P.,Youens,K.,Wagner,A.D.and Preston, A. R. (2012). Hippocampal novelty activations in schizophrenia:disease and medication effects. Schizophr. Res. 138, 157-163.
Torii, M., Matsuzaki, F., Osumi, N., Kaibuchi, K., Nakamura, S., Casarosa, S.,Guillemot, F. and Nakafuku, M. (1999). Transcription factors Mash-1 and Prox-1delineate early steps in differentiation of neural stem cells in the developing centralnervous system. Development 126, 443-456.
Valvezan, A. J. and Klein, P. S. (2012). GSK-3 and Wnt signaling in neurogenesisand bipolar disorder. Front. Mol. Neurosci. 5, 1.
van Praag, H., Kempermann, G. and Gage, F. H. (1999). Running increases cellproliferation and neurogenesis in the adult mouse dentate gyrus.Nat. Neurosci. 2,266-270.
Vanderhaeghen, P. (2012). Generation of cortical neurons from pluripotent stemcells. Prog. Brain Res. 201, 183-195.
Villeda, S. A., Luo, J., Mosher, K. I., Zou, B., Britschgi, M., Bieri, G., Stan, T. M.,Fainberg, N., Ding, Z., Eggel, A. et al. (2011). The ageing systemic milieunegatively regulates neurogenesis and cognitive function. Nature 477, 90-94.
von Frowein, J., Wizenmann, A. and Gotz, M. (2006). The transcription factorsEmx1 and Emx2 suppress choroid plexus development and promoteneuroepithelial cell fate. Dev. Biol. 296, 239-252.
Walton, N. M., Zhou, Y., Kogan, J. H., Shin, R., Webster, M., Gross, A. K.,Heusner, C. L., Chen, Q., Miyake, S., Tajinda, K. et al. (2012). Detection of animmature dentate gyrus feature in human schizophrenia/bipolar patients. Transl.Psychiatry 2, e135.
Wang, L., Wang, L., Huang,W., Su, H., Xue, Y., Su, Z., Liao, B.,Wang, H., Bao, X.,Qin, D. et al. (2013). Generation of integration-free neural progenitor cells fromcells in human urine. Nat. Methods 10, 84-89.
Wapinski, O. L., Vierbuchen, T., Qu, K., Lee, Q. Y., Chanda, S., Fuentes, D. R.,Giresi, P. G., Ng, Y. H., Marro, S., Neff, N. F. et al. (2013). Hierarchicalmechanisms for direct reprogramming of fibroblasts to neurons. Cell 155,621-635.
Watanabe, K., Kamiya, D., Nishiyama, A., Katayama, T., Nozaki, S., Kawasaki, H.,Watanabe, Y., Mizuseki, K. and Sasai, Y. (2005). Directed differentiation oftelencephalic precursors from embryonic stem cells. Nat. Neurosci. 8, 288-296.
Wernig, M., Zhao, J.-P., Pruszak, J., Hedlund, E., Fu, D., Soldner, F., Broccoli, V.,Constantine-Paton, M., Isacson, O. and Jaenisch, R. (2008). Neurons derivedfrom reprogrammed fibroblasts functionally integrate into the fetal brain and improvesymptoms of rats with Parkinson’s disease. Proc. Natl. Acad. Sci. U.S.A. 105,5856-5861.
Witter, M. P. (2007). The perforant path: projections from the entorhinal cortex to thedentate gyrus. Prog. Brain Res. 163, 43-61.
Yamanaka, S. (2007). Strategies and new developments in the generation ofpatient-specific pluripotent stem cells. Cell Stem Cell 1, 39-49.
Yamanaka, S. and Takahashi, K. (2006). Induction of pluripotent stem cells frommouse fibroblast cultures. Tanpakushitsu Kakusan Koso 51, 2346-2351.
Yamasaki, N., Maekawa, M., Kobayashi, K., Kajii, Y., Maeda, J., Soma, M.,Takao, K., Tanda, K., Ohira, K., Toyama, K. et al. (2008). Alpha-CaMKIIdeficiency causes immature dentate gyrus, a novel candidate endophenotype ofpsychiatric disorders. Mol. Brain 1, 6.
Yoo, A. S., Sun, A. X., Li, L., Shcheglovitov, A., Portmann, T., Li, Y., Lee-Messer,C., Dolmetsch, R. E., Tsien, R. W. and Crabtree, G. R. (2011). MicroRNA-mediated conversion of human fibroblasts to neurons. Nature 476, 228-231.
Yu, D. X., Marchetto, M. C. and Gage, F. H. (2013). Therapeutic translation ofiPSCs for treating neurological disease. Cell Stem Cell 12, 678-688.
Yu, D. X., Di Giorgio, F. P., Yao, J., Marchetto, M. C., Brennand, K., Wright, R.,Mei, A., McHenry, L., Lisuk, D., Grasmick, J. M. et al. (2014). Modelinghippocampal neurogenesis using human pluripotent stem cells. StemCell Rep. 2,295-310.
Zhao, C., Deng, W. and Gage, F. H. (2008). Mechanisms and functionalimplications of adult neurogenesis. Cell 132, 645-660.
Zhou, Q. and Tripathi, P. (2012). How to remake a fibroblast into a neural stem cell.Cell Stem Cell 10, 347-348.
Zhou, C.-J., Zhao, C. and Pleasure, S. J. (2004). Wnt signaling mutants havedecreased dentate granule cell production and radial glial scaffoldingabnormalities. J. Neurosci. 24, 121-126.
Zou, Q., Yan, Q., Zhong, J., Wang, K., Sun, H., Yi, X. and Lai, L. (2014). Directconversion of human fibroblasts into neuronal restricted progenitors. J. Biol.Chem. 289, 5250-5260.
2375
PRIMER Development (2014) 141, 2366-2375 doi:10.1242/dev.096776
DEVELO
PM
ENT