Embed Size (px)
Citation preview

of May 12, 2018.This information is current as
LymphopoiesisRegulate an Early Checkpoint in Hoxa9 and Flt3 Signaling Synergistically
Zhixin L. Huang and Kay L. MedinaKimberly A. Gwin, Mariya B. Shapiro, Joseph J. Dolence,
http://www.jimmunol.org/content/191/2/745doi: 10.4049/jimmunol.12032942013;
2013; 191:745-754; Prepublished online 14 JuneJ Immunol
Referenceshttp://www.jimmunol.org/content/191/2/745.full#ref-list-1
, 28 of which you can access for free at: cites 50 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2013 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Hoxa9 and Flt3 Signaling Synergistically Regulate an EarlyCheckpoint in Lymphopoiesis
Kimberly A. Gwin, Mariya B. Shapiro, Joseph J. Dolence, Zhixin L. Huang, and
Kay L. Medina
Hoxa9 and Flt3 signaling are individually important for the generation of lymphoid lineage precursors from multipotent hema-
topoietic progenitors (MPP) in bone marrow. Mice deficient for Hoxa9, Flt3, or Flt3 ligand (FL) have reduced numbers of lymphoid-
primed multipotential progenitors (LMPP), common lymphoid progenitors (CLP), and B/T cell precursors. Hoxa9 regulates lym-
phoid development, in part, through transcriptional regulation of Flt3. However, it was unclear whether Hoxa9 has functions in
lymphopoiesis independent of, or alternatively, synergistically with Flt3 signaling. In this study, we show that Hoxa92/2Flt3l2/2
mice have more severe deficiencies in all B lineage cells, CLP, LMPP, and total Flt3+ MPP in bone marrow than the single
knockouts. Although LMPP and Flt3+ CLP contain precursors for NK and dendritic cell lineage cells, no deficiencies in these
lineages beyond that in Flt3l2/2 mice was found. Thymocyte cellularity was significantly reduced in the compound knockout,
although peripheral T cell numbers mirrored Flt3l2/2 mice. Analysis of the hematopoietic progenitor compartment revealed
elevated numbers of CD150+hiCD342CD41+ myeloid–biased stem cells in Hoxa92/2Flt3l2/2 mice. In contrast, CD1502 MPP
enriched for lymphoid potential were synergistically reduced, suggesting Hoxa9 and Flt3 signaling function coordinately to
regulate lymphopoiesis at a very early stage. Real-time PCR analysis of CD1502Flt3+ cells from wild-type control, Hoxa92/2,
and Flt3l2/2 single knockouts revealed decreased lymphoid transcripts, corroborating the importance of these regulators in
lymphoid development. Taken together, these studies reveal a very early checkpoint in lymphopoiesis dependent on the combi-
natorial activities of Hoxa9 function and Flt3 signaling. The Journal of Immunology, 2013, 191: 745–754.
Steady-state production of lymphoid–lineage precursorsfrom pluripotent hematopoietic stem cells (HSC) is a step-wise process driven by the concerted activities of tran-
scription factors and signaling molecules (1). For example, at leastsix developmental intermediates have been characterized betweenHSC and committed pro-B cells: short-term repopulating cells,Flt3+ MPP, LMPP that are Flt3hi and evidence lymphoid priming,all lymphoid progenitors (ALP), B lineage–restricted progenitors(BLP), and pre-pro-B cells. The ability to resolve these develop-mental intermediates makes it possible to more accurately defineprecursor–progeny relationships and assemble unique and inter-connected gene regulatory modules that instruct the B cell fatedecision. Importantly, it also allows more precise identificationof developmental stage-specific blocks imposed by gene-targetingstrategies. The end result is a better understanding of the roles ofindividual molecular and cellular determinants in regulation oflymphoid development and B cell differentiation.In bone marrow (BM), most HSCs are quiescent, and few are
actively participating in blood cell genesis at any one time (2).
Pluripotent HSC are enriched in the Lineage-negative (Lin2),
Sca-1+, c-kit+hi (collectively referred to as LSK+) fraction of BM
cells expressing the signaling lymphocyte activation molecule
family marker CD150 (3). Within LSK+CD150+ cells, HSC can
be phenotypically distinguished from multipotential progenitors
(MPP) by differential expression of a variety of cell surface markers
including CD34, CD48, CD27, and Flt3 (2, 4–6). HSC are LSK+
CD150+CD342lo, whereas MPPs are CD1502lo and express CD34,
CD48, CD27, and Flt3. Although most HSC are not actively cy-
cling, the majority of MPP are proliferating to maintain blood cell
production.Surface expression of Flt3 is an early event in HSC differentiation
and largely coincides with downregulation of CD150 (7, 8). Ac-
quisition of Flt3 denotes lymphoid–myeloid-biased differentiation
potential and reduced capacity to generate erythroid/megakaryocyte
progeny (9). A subset of Flt3+ MPPs expressing high levels of Flt3
express the early program of lymphoid–lineage gene expression and
have been denoted early lymphoid progenitors or lymphoid-primed
multipotential progenitors (LMPP) (10). Flt3 signaling plays a crit-
ical role in regulation of lymphoid priming in primitive hemato-
poietic progenitors (10, 11). However, a direct regulatory connection
between Flt3 signaling and the expression and/or activity of any
lymphoid–lineage specification factor remains to be established.
LMPP can be prospectively identified within the LSK+ fraction
using differential expression of VCAM-1 and Flt3 (VCAM-12
Flt3+hi) or by expression of GFP using RAG-1GFP knockin reporter
mice (12–14). LMPP are enriched for precursors biased for T, B, NK,
and/or dendritic cell (DC) differentiation potential (15, 16). At pres-
ent, it is unclear whether individual LMPP retain multilineage lym-
phoid differentiation potential. Regardless, data obtained from numer-
ous gene-targeting strategies have established that failure to generate
and/or maintain the LMPP pool has significant consequences on the
production of B lineage lymphocytes in BM (11, 17–21).
Department of Immunology, Mayo Clinic, Rochester, MN 55905
Received for publication November 29, 2012. Accepted for publication May 15,2013.
This work was supported by Grant R01HL096108 from the National Heart, Lung,and Blood Institute (to K.L.M.).
Address correspondence and reprint requests to Dr. Kay L. Medina, Department ofImmunology, Mayo Clinic, 200 First Street Southwest, Rochester, MN 55905. E-mailaddress: [email protected]
Abbreviations used in this article: ALP, all lymphoid progenitor; B6, C57BL/6; BCP,B cell precursor; BLP, B lineage–restricted progenitor; BM, bone marrow; CLP, commonlymphoid progenitor; DC, dendritic cell; FL, Flt3 ligand; GMLP, granulocyte-macrophage lymphoid progenitor; HSC, hematopoietic stem cell; LMPP, lymphoid-primedmultipotential progenitor; LSK+, Lineage-negative, Sca-1+, c-kit+; MPP, multipotentialprogenitor; SP, single-positive; WT, wild-type.
Copyright� 2013 by The American Association of Immunologists, Inc. 0022-1767/13/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1203294
by guest on May 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
Downregulation of c-kit and surface expression of IL-7R denotesthe transition from LMPP to CLP (16). BLP are enriched in a subsetof CLPs that express the cell surface marker Ly6D, whereas Ly6D2
CLPs exhibit B, T, and NK potentials and are referred to as ALP(22). Resolution of ALP and BLP is an important advance becauseit enables more accurate determination of the roles of regulatoryfactors in lymphoid lineage restriction as well as B lineage com-mitment from lymphoid-restricted progenitors. In that vein, thisstudy seeks to determine individual versus combinatorial roles oftwo key factors, the homeodomain transcription factor Hoxa9, andthe cytokine, Flt3 ligand (FL), in regulation of ALP and BLP.Hoxa9 is a transcription factor important for the generation of
normal numbers of B lineage precursors in BM (17, 19, 23). Hoxa9is expressed at low levels in HSC, upregulated in MPP and CLP,and then downregulated in B cell precursor (BCP) (17). Hoxa9regulates B cell development, in part, through transcriptional reg-ulation of Flt3 (17). It is presently unclear whether the B cell de-ficiency in Hoxa92/2 mice is due solely to impaired Flt3 expres-sion or whether Hoxa9 and Flt3 signaling provide nonoverlappingfunctions that guide B cell development. We theorized that if theB lineage deficiency in Hoxa92/2 mice was due to impaired Flt3alone, then combined loss of Hoxa9 and FL would not exacer-bate the B lineage deficiency beyond that observed in the singleknockouts. However, if Hoxa9 has additional nonredundant rolesor functions cooperatively with Flt3 signaling to promote lymphoiddevelopment and B cell differentiation, then combined loss couldexacerbate the B lineage deficiency.In this study, through generation and analysis of Hoxa92/2
Flt3l2/2 mice, we show that Hoxa9 and Flt3 signaling functioncooperatively to establish the lymphoid progenitor pool in BM.Intriguingly, LSK+CD150hiCD342loCD41+ cells that are myeloid-biased HSC are overrepresented in the LSK+ compartment andincreased in number in the compound knockout mice. In contrast,LSK+CD1502Flt3+ cells that are enriched for lymphoid progenitorsare significantly reduced (5, 24). ALP, BLP, and B lineage lym-phocytes are ablated in BM of Hoxa92/2Flt3l2/2 mice. NK andDC lineage cells are not impacted beyond what has been docu-mented as a consequence of FL deficiency, suggesting alternativepathways of generation (25). Thymic cellularity is significantly
reduced in Hoxa92/2Flt3l2/2 mice compared with the singleknockouts consistent with the ablation of LMPP in BM that seed thethymus. However, T cell development on the whole appears essen-tially normal, and numbers of peripheral T cells mirror Flt3l2/2
mice. These findings establish that there is an extremely earlycheckpoint in lymphoid lineage development dependent on thecombinatorial activities of Hoxa9 and Flt3 signaling.
Materials and MethodsMice
C57BL/6 (referred to in this paper as wild-type [WT] or B6) mice werepurchased from The Jackson Laboratory (Bar Harbor, ME). Hoxa92/2 andFlt3l2/2 mice have been described previously (11, 17). Hoxa92/2 andFlt3l2/2 mice were bred, and genotyping was performed to identifycompound knockouts (11, 17). All mice in this study were maintained inthe Mayo Clinic animal facility and used between 8 and 16 wk of age.Age-matched mice of all four genotypes were used in each experiment,and no differences in phenotype were observed in that range or in maleversus female mice. All experiments were carried out in accordance withMayo Clinic Institutional Animal Care and Use Committee guidelines.
Abs and flow cytometry
Methods for flow cytometry and progenitor isolation have been describedpreviously (11, 14, 17). Flow cytometric analysis was performed on theLSRII cytometer (BD Biosciences, San Jose, CA) and data analysis withFlowJo software (Tree Star, Ashland, OR). All Abs were purchased fromeBioscience, BioLegend, or BD Pharmingen. Ab conjugations and combi-nations used to delineate progenitor subsets in BM were the following:LSK+, Lineage mixture (FITC-conjugated CD45R/B220, CD11b/Mac1,Ly6G/Gr1, TER119, CD3ε, CD8a, and NK1.1), c-kit–allophycocyanin–efluor 780, Sca-1-PerCP-Cy5.5, CD150 Pe-Cy7, Flt3-PE, CD34-eFluor450, and CD41-allophycocyanin; CLP, Lineage mixture (FITC-conju-gated CD45R/B220, CD11b/Mac1, Ly6G/Gr1, TER119, CD3ε, CD8a,CD11c, NK1.1, and Ly6C), c-kit–allophycocyanin–efluor780, Sca1-PerCPCy5.5, biotin IL-7R (visualized with streptavidin PeCy7), Flt3-PE,and Ly6D-allophycocyanin; BCP (CD19-allophycocyanin, B220-PE-Cy7,IgM-FITC, and CD43-PE); myeloid cells, CD11b/Mac-1-allophycocyanin,and Gr-1-FITC; erythroid cells, Ter119-PeCy7; NK subsets, CD3ε-PeCy7,CD122-FITC, and NK1.1-PerCP-Cy5.5; total DCs, CD11c-PE; and thy-mocyte subsets, CD3ε-allophycocyanin, CD4-FITC, and CD8a-PE. Gatesfor discriminating negative and positive subsets were determined based onstaining of BM mononuclear cells or previously characterized positive/negative BM subsets. Spleens and thymi were harvested, and single-cellsuspensions were made by crushing the organs through a 70-mm nylon
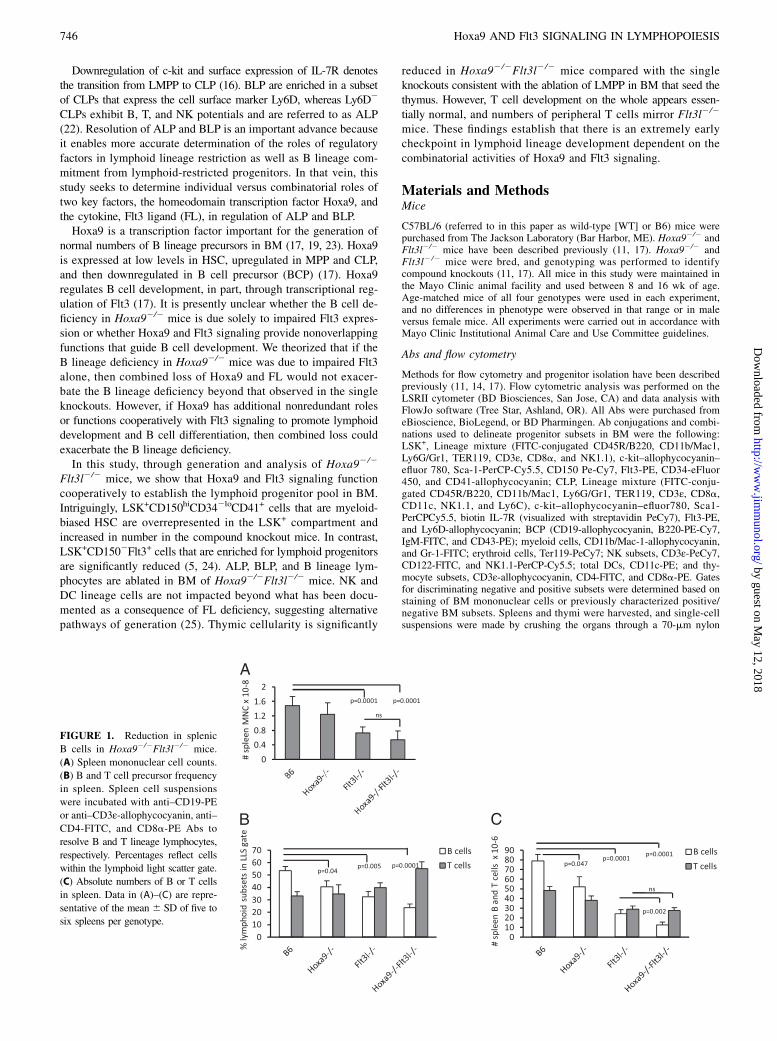
FIGURE 1. Reduction in splenic
B cells in Hoxa92/2Flt3l2/2 mice.
(A) Spleen mononuclear cell counts.
(B) B and T cell precursor frequency
in spleen. Spleen cell suspensions
were incubated with anti–CD19-PE
or anti–CD3ε-allophycocyanin, anti–CD4-FITC, and CD8a-PE Abs to
resolve B and T lineage lymphocytes,
respectively. Percentages reflect cells
within the lymphoid light scatter gate.
(C) Absolute numbers of B or T cells
in spleen. Data in (A)–(C) are repre-
sentative of the mean 6 SD of five to
six spleens per genotype.
746 Hoxa9 AND Flt3 SIGNALING IN LYMPHOPOIESIS
by guest on May 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
mesh. Mononuclear cell counts were determined by counting nuclei afterlysis in 3% acetic acid/water solution.
Isolation of progenitor subsets for real-time PCR analysis
BM cells were harvested and pooled from six B6 controls, sevenHoxa92/2,and eight Flt3l2/2 mice in each of two independent experiments. Lin2
cells were enriched from the three mouse groups by incubating BM cellsuspensions with a biotin-labeled Lin+ Ab mixture (B220, Gr-1, Ter119,CD3e, CD8a, CD11c, NK1.1, and CD19), followed by incubation withstreptavidin-conjugated microbeads and magnetic separation. The Lin2
enriched BM fractions were incubated with c-kit-eFluor 780, Sca-1-PerCP-Cy5.5, CD150-PE, Flt3-PE, and streptavidin-FITC (to identify residualLin+lo cells). Two LSK+ subsets were sorted from B6, Hoxa92/2, orFlt3l2/2Lin2 cells: LSK+CD1502Flt3lo and LSK+ CD1502Flt3+. Aftercentrifugation, the cell pellets were resuspended in 100 ml extractionbuffer and then stored at 280˚C or processed immediately for RNA iso-lation using the Arcturus PicoPure RNA isolation kit (Applied Biosystems,Foster City, CA). To remove contaminating genomic DNA, the RNA wastreated with DNase I kit (Qiagen, Germantown, MD). cDNA amplificationwas performed using the Ovation PicoSL WTA System (NuGEN, SanCarlos, CA). Amplified cDNAwas purified using the MinElute ReactionCleanup kit (Qiagen). Real-time PCR was performed using TaqMan probes(Applied Biosystems) or gene-specific SYBR Green assays as we de-scribed previously. Gene expression was normalized to 18S RNA forTaqMan assays or gapdh for SYBR Green assays. All cDNA samples were
assayed in triplicate. Relative transcript abundance was determined usingthe 22DDCT method with B6 transcript levels for each sorted subset as-signed the comparator.
Statistical analysis
Statistical significance was determined using the Student t test. A pvalue # 0.05 was deemed significant and is indicated by asterisks.
ResultsSynergistic reduction in B lymphopoiesis in Hoxa92/2Flt3l2/2
mice
To determine whether Hoxa9 provides nonredundant roles orfunctionally coordinates with Flt3 signaling in the generation oflymphoid progenitors and BCPs, Hoxa92/2Flt3l2/2 mice weregenerated. The compound knockouts were born at normal Men-delian ratios and exhibited no gross abnormalities. Peripheral bloodanalysis of the double knockouts mirrored Flt3l2/2 mice (i.e., re-duced numbers and percentages of lymphocytes; data not shown).Consistent with the reduction in blood lymphocytes, spleen cellu-larity was reduced ∼60% in Hoxa92/2Flt3l2/2 mice, comparedwith 39% in Flt3l2/2 mice (Fig. 1A). Flow cytometric analysis of
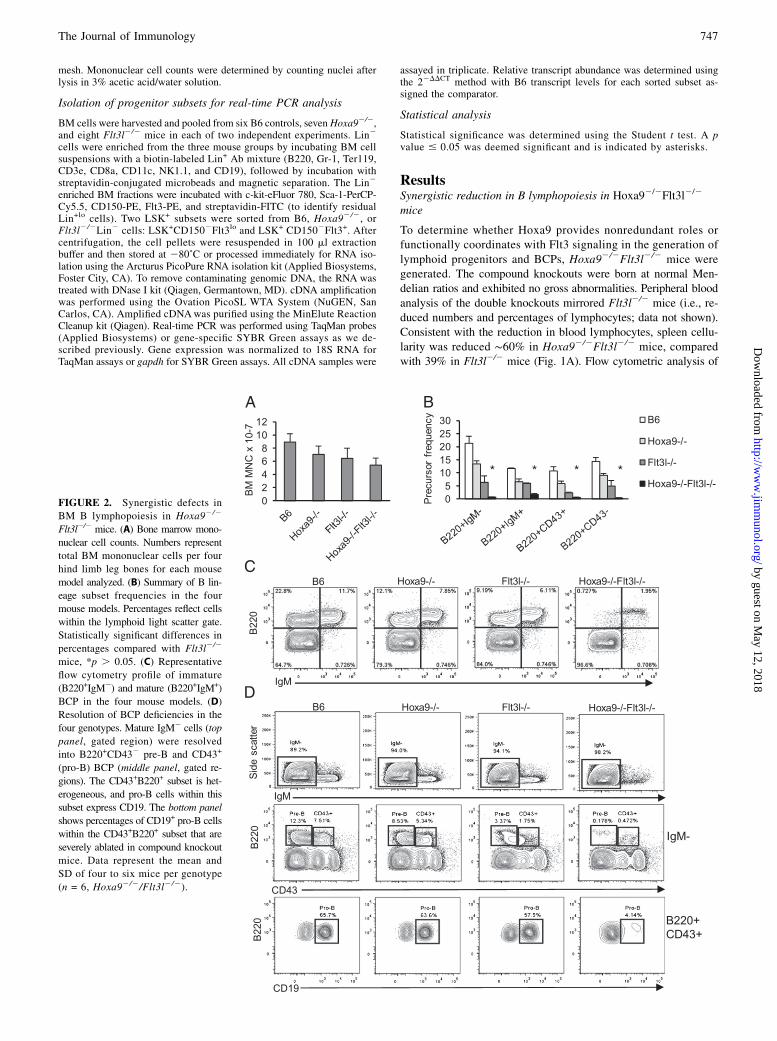
FIGURE 2. Synergistic defects in
BM B lymphopoiesis in Hoxa92/2
Flt3l2/2 mice. (A) Bone marrow mono-
nuclear cell counts. Numbers represent
total BM mononuclear cells per four
hind limb leg bones for each mouse
model analyzed. (B) Summary of B lin-
eage subset frequencies in the four
mouse models. Percentages reflect cells
within the lymphoid light scatter gate.
Statistically significant differences in
percentages compared with Flt3l2/2
mice, *p . 0.05. (C) Representative
flow cytometry profile of immature
(B220+IgM2) and mature (B220+IgM+)
BCP in the four mouse models. (D)
Resolution of BCP deficiencies in the
four genotypes. Mature IgM2 cells (top
panel, gated region) were resolved
into B220+CD432 pre-B and CD43+
(pro-B) BCP (middle panel, gated re-
gions). The CD43+B220+ subset is het-
erogeneous, and pro-B cells within this
subset express CD19. The bottom panel
shows percentages of CD19+ pro-B cells
within the CD43+B220+ subset that are
severely ablated in compound knockout
mice. Data represent the mean and
SD of four to six mice per genotype
(n = 6, Hoxa92/2/Flt3l2/2).
The Journal of Immunology 747
by guest on May 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
T cells in the spleen showed no significant differences in percent-ages or absolute numbers in the compound knockouts beyond thatobserved in the single knockouts (Fig. 1B, 1C). No alterations inratios of CD4+ or CD8+ splenic T cells were observed (data notshown). In contrast, percentages of CD19+ splenic B cells weresynergistically reduced in the compound knockout mice (splenicCD19+ cells, B6 = 53.4 6 8.4% [n = 6], Hoxa92/2 = 40.6 6 9.4%[n = 5], Flt3l2/2 = 32.5 6 11.8% [n = 6], and Hoxa92/2Flt3l2/2 =23.8 6 6.6% [n = 6]) (Fig. 1B). The reduced percentages togetherwith reduction in cellularity in the compound knockout culminatedin a statistically significant reduction in numbers of splenic CD19+
B lymphocytes (12.5 6 7.4 versus 24.1 6 10.2 3 106, Hoxa92/2
Flt3l2/2 versus Flt3l2/2, respectively; *p = 0.047). These datasuggest that combined loss of Hoxa9 and Flt3 signaling has moresevere consequences on B lymphopoiesis than deficiency in Hoxa9or FL alone. Splenic T cells are reduced but comparable to FLdeficiency alone.
Severe reduction in BM BCP in Hoxa92/2Flt3l2/2 mice
The spleen results suggested that the compound knockout micemight have selective abnormalities associated with B lympho-poiesis in BM. Hoxa92/2 and Flt3l2/2 single knockout mice areknown to have reduced BM cellularity, and in this study, we de-termined that the same was true for Hoxa92/2Flt3l2/2 mice (Fig.2A) (23, 25). However, the reduction in BM cellularity in the
compound knockouts was not statistically significant when com-pared with the single knockouts.To determine the consequence of combined loss of Hoxa9 and
Flt3 signaling on B lymphopoiesis, BM cells were harvested fromstrain-matched B6 controls, Hoxa92/2, Flt3l2/2, and Hoxa92/2
Flt3l2/2 mice and stained with Abs to CD45R/B220, CD19, CD43,and IgM to resolve B lineage subsets. As summarized in Fig. 2Band shown in Fig. 2C and 2D, combined loss of Hoxa9 and FLseverely ablated B lymphopoiesis. To document the stages of Blymphopoiesis affected in the compound knockout mice, mature(B220+IgM+) and immature (B220+IgM2) B lineage subsets wereexamined (Fig. 2C, top panel). Indeed, the prominent subset ofBCPs in marrow in the compound knockout mice were IgM+ andlikely represent recirculating B lymphocytes (26). It is known thatnaive B cells undergo homeostatic proliferation under conditionsof B cell deficiency, and the IgM+ cells in the marrow uniformlyexpressed high levels of B220, characteristic of recirculatingB cells (26). In contrast, all IgM2 BCP subsets, including B220+
CD432 pre-B cells and B220+CD43+CD19+ pro-B cells, were ablatedin the compound knockout mice (Fig. 2C). We conclude fromthese experimental findings that Hoxa9 and Flt3 function in ashared pathway to regulate B lymphopoiesis in BM.The consequence of combined loss of Hoxa9 and Flt3 signaling
was selective for B lineage lymphocytes. We found no significantalterations in frequencies of Mac-1/Gr1+ myeloid or Ter119+ ery-
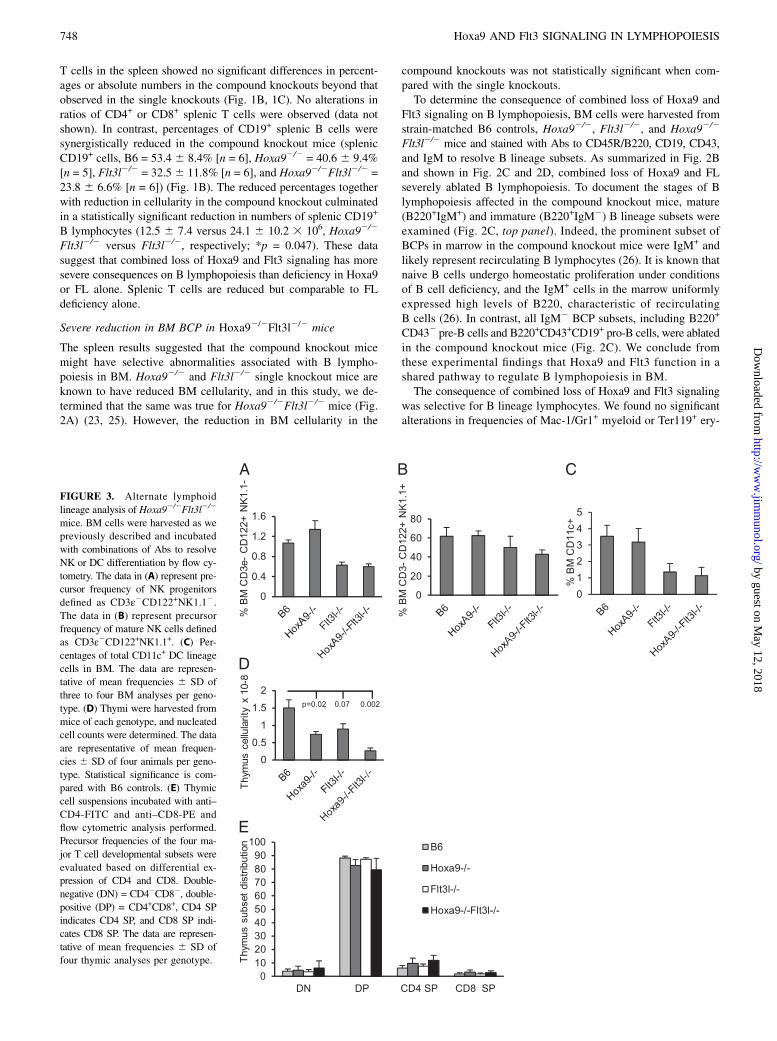
FIGURE 3. Alternate lymphoid
lineage analysis of Hoxa92/2Flt3l2/2
mice. BM cells were harvested as we
previously described and incubated
with combinations of Abs to resolve
NK or DC differentiation by flow cy-
tometry. The data in (A) represent pre-
cursor frequency of NK progenitors
defined as CD3ε2CD122+NK1.12.The data in (B) represent precursor
frequency of mature NK cells defined
as CD3ε2CD122+NK1.1+. (C) Per-
centages of total CD11c+ DC lineage
cells in BM. The data are represen-
tative of mean frequencies 6 SD of
three to four BM analyses per geno-
type. (D) Thymi were harvested from
mice of each genotype, and nucleated
cell counts were determined. The data
are representative of mean frequen-
cies 6 SD of four animals per geno-
type. Statistical significance is com-
pared with B6 controls. (E) Thymic
cell suspensions incubated with anti–
CD4-FITC and anti–CD8-PE and
flow cytometric analysis performed.
Precursor frequencies of the four ma-
jor T cell developmental subsets were
evaluated based on differential ex-
pression of CD4 and CD8. Double-
negative (DN) = CD42CD82, double-
positive (DP) = CD4+CD8+, CD4 SP
indicates CD4 SP, and CD8 SP indi-
cates CD8 SP. The data are represen-
tative of mean frequencies 6 SD of
four thymic analyses per genotype.
748 Hoxa9 AND Flt3 SIGNALING IN LYMPHOPOIESIS
by guest on May 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

throid cells (data not shown). Thus, the profound deficiency in BCPin the compound knockout mice is not due to a requirement forHoxa9 and Flt3 signaling in regulating the lymphoid versus myeloid–erythroid fate decision in primitive hematopoietic progenitors.NK and DC lineage cells in BM share a common progenitor
pathway with BCP (15). Flow cytometric analysis of NK precursorsdefined as CD3e2CD122+NK1.12 or mature NK cells defined asCD3e2CD122+NK1.1+ (27) revealed no significant alteration inNK development or maintenance in the compound knockouts be-yond that because of Flt3l deficiency (Fig. 3A, 3B). The same wastrue for dendritic lineage cells defined broadly as CD11c+ (Fig. 3C).T cells and B cells arise from Flt3+ MPPs in BM (15). Numbers
of total thymocytes were similarly reduced in Hoxa92/2 andFlt3l2/2 mice in accord with previous reports (25, 28, 29). Com-pound loss of Hoxa9 and FL resulted in a more severe reduction intotal thymocyte cellularity than the single knockout mice (∼5.8-foldreduced compared with B6 controls and 3-fold from Hoxa92/2
[p = 0.0058] or Flt3l2/2 mice [p = 0.0078]) (Fig. 3D). The syn-ergistic reduction in thymocyte cellularity is likely due to theablation of Flt3+ MPPs in BM that seed the thymus, thus reducingthe efficiency of BM progenitor seeding of the thymus (see below)(30). Although total thymocyte numbers are reduced, we found nosignificant alteration in major hallmarks of T cell development(Fig. 3E). Percentages of CD42CD82 (double-negative), CD4+
CD8+ (double-positive), or CD4+ or CD8+ single-positive (SP)thymocyte subsets were comparable in all four mouse models.Regardless, the production of T lymphocytes overall is reducedin the compound knockout compared with WT or single mutationcontrols.We conclude from these analyses that compound loss of Hoxa9
and Flt3 signaling severely and synergistically reduces the gen-eration of B and T lineage lymphocytes.
Severe deficiency in ALP and BLP in Hoxa92/2Flt3l2/2 mice
Functional BCP are enriched in Lin2ckitloIL-7R+Flt3+ CLPs.Hoxa92/2 and Flt3l2/2 mice each have significantly reducednumbers of Flt3+ CLPs (11, 17, 19). Recently, it was determinedthat Flt3+ CLPs could be fractionated into Ly6D2 ALP and Ly6D+
BLP (22). Both categories of Flt3+ CLPs were depressed infrequencies and numbers in the single knockouts (Fig. 4A andsummarized in Fig. 4B, 4C). Combined loss of Hoxa9 and FLexacerbated the reduction of Flt3+Ly6D2 ALP (∼70% reductionin Ly6D2 compared with Flt3l2/2 mice) and Flt3+Ly6D+ BLP(∼37-fold reduction compared with Flt3l2/2 mice). We note anincrease in abundance of Flt3+lo CLPs in Flt3l2/2 and Hoxa92/2
Flt3l2/2 mice (Fig. 4A). This observation suggests that Flt3 sig-naling might play a role in autoregulation of Flt3 in CLPs or atan earlier stage (see below). The dramatic loss of Flt3+ CLPs inHoxa92/2Flt3l2/2 mice suggests that Hoxa9 and Flt3 signalingare critical components of a genetic network that regulates thegeneration or maintenance of ALP and BLP.
Reductions in LMPP and granulocyte-macrophage lymphoidprogenitor in Hoxa92/2Flt3l2/2 mice
We previously reported decreased Flt3+LSK+ cells in Hoxa92/2
and Flt3l2/2 mice (11, 17). The dramatic reduction in ALP inthe compound knockout mice suggested critical roles for Hoxa9and Flt3 signaling upstream of IL-7R expression. LMPP arethe presumed immediate precursors of ALP as il7ra transcriptsare detectable by real-time PCR at this stage (10). LMPP canbe distinguished within LSK+ cells based on differential ex-pression of VCAM-1 and Flt3 (31). As shown in Fig. 5A andsummarized in Fig. 5B, LSK+VCAM-12Flt3+hi LMPP are re-duced in Flt3l2/2 and Hoxa92/2 mice and ablated in Hoxa92/2
Flt3l2/2 mice.
FIGURE 4. Ablation of CLP subsets in Hoxa92/2Flt3l2/2 mice. (A) Representative flow cytometry profiles of Lineage (Lin2) IL-7R+ CLP subsets in
the four mouse models. Lin2 cells (top panels) were discriminated based on differential expression of c-kit and IL-7R to identify CLPs. Lin2c-kitloIL-7R+
cells (middle panels) were further fractionated into ALP and BLP based on differential expression of Flt3 and Ly6D (bottom panels). The plots shown
are representative of three to four mice per genotype. (B) Precursor frequency of Flt3+ CLP subsets in the four mouse models. (C) Absolute numbers of ALP
(white bar) and BLP (gray bar). Data represent the mean and SD of four mice per genotype. Statistical significance, indicated by the asterisks, is
determined relative to B6 WT controls. **p . 0.005, ***p . 0.0001.
The Journal of Immunology 749
by guest on May 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
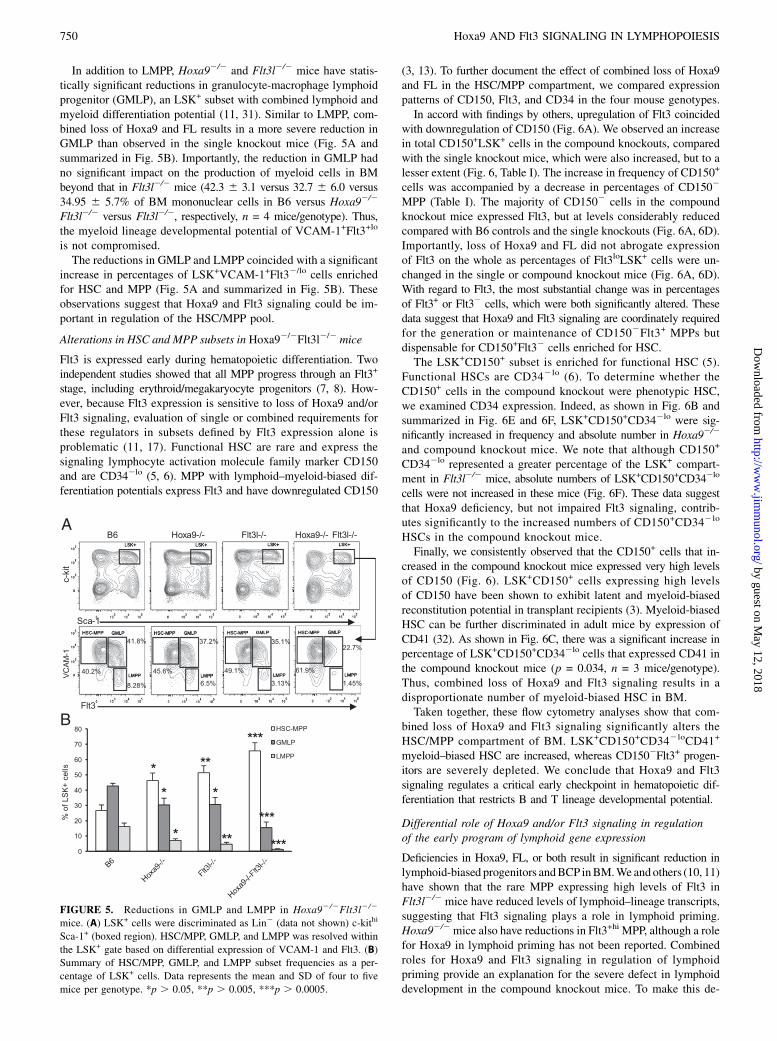
In addition to LMPP, Hoxa92/2 and Flt3l2/2 mice have statis-tically significant reductions in granulocyte-macrophage lymphoidprogenitor (GMLP), an LSK+ subset with combined lymphoid andmyeloid differentiation potential (11, 31). Similar to LMPP, com-bined loss of Hoxa9 and FL results in a more severe reduction inGMLP than observed in the single knockout mice (Fig. 5A andsummarized in Fig. 5B). Importantly, the reduction in GMLP hadno significant impact on the production of myeloid cells in BMbeyond that in Flt3l2/2 mice (42.3 6 3.1 versus 32.7 6 6.0 versus34.95 6 5.7% of BM mononuclear cells in B6 versus Hoxa92/2
Flt3l2/2 versus Flt3l2/2, respectively, n = 4 mice/genotype). Thus,the myeloid lineage developmental potential of VCAM-1+Flt3+lo
is not compromised.The reductions in GMLP and LMPP coincided with a significant
increase in percentages of LSK+VCAM-1+Flt32/lo cells enrichedfor HSC and MPP (Fig. 5A and summarized in Fig. 5B). Theseobservations suggest that Hoxa9 and Flt3 signaling could be im-portant in regulation of the HSC/MPP pool.
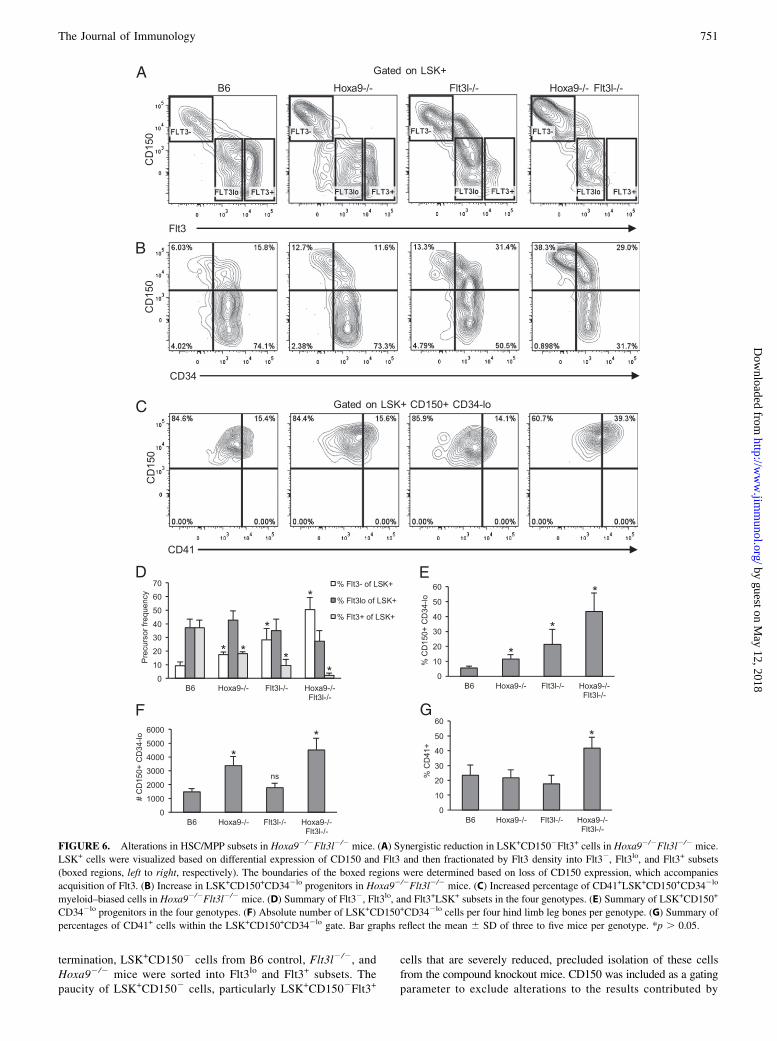
Alterations in HSC and MPP subsets in Hoxa92/2Flt3l2/2 mice
Flt3 is expressed early during hematopoietic differentiation. Twoindependent studies showed that all MPP progress through an Flt3+
stage, including erythroid/megakaryocyte progenitors (7, 8). How-ever, because Flt3 expression is sensitive to loss of Hoxa9 and/orFlt3 signaling, evaluation of single or combined requirements forthese regulators in subsets defined by Flt3 expression alone isproblematic (11, 17). Functional HSC are rare and express thesignaling lymphocyte activation molecule family marker CD150and are CD342lo (5, 6). MPP with lymphoid–myeloid-biased dif-ferentiation potentials express Flt3 and have downregulated CD150
(3, 13). To further document the effect of combined loss of Hoxa9and FL in the HSC/MPP compartment, we compared expressionpatterns of CD150, Flt3, and CD34 in the four mouse genotypes.In accord with findings by others, upregulation of Flt3 coincided
with downregulation of CD150 (Fig. 6A). We observed an increasein total CD150+LSK+ cells in the compound knockouts, comparedwith the single knockout mice, which were also increased, but to alesser extent (Fig. 6, Table I). The increase in frequency of CD150+
cells was accompanied by a decrease in percentages of CD1502
MPP (Table I). The majority of CD1502 cells in the compoundknockout mice expressed Flt3, but at levels considerably reducedcompared with B6 controls and the single knockouts (Fig. 6A, 6D).Importantly, loss of Hoxa9 and FL did not abrogate expressionof Flt3 on the whole as percentages of Flt3loLSK+ cells were un-changed in the single or compound knockout mice (Fig. 6A, 6D).With regard to Flt3, the most substantial change was in percentagesof Flt3+ or Flt32 cells, which were both significantly altered. Thesedata suggest that Hoxa9 and Flt3 signaling are coordinately requiredfor the generation or maintenance of CD1502Flt3+ MPPs butdispensable for CD150+Flt32 cells enriched for HSC.The LSK+CD150+ subset is enriched for functional HSC (5).
Functional HSCs are CD342lo (6). To determine whether theCD150+ cells in the compound knockout were phenotypic HSC,we examined CD34 expression. Indeed, as shown in Fig. 6B andsummarized in Fig. 6E and 6F, LSK+CD150+CD342lo were sig-nificantly increased in frequency and absolute number in Hoxa92/2
and compound knockout mice. We note that although CD150+
CD342lo represented a greater percentage of the LSK+ compart-ment in Flt3l2/2 mice, absolute numbers of LSK+CD150+CD342lo
cells were not increased in these mice (Fig. 6F). These data suggestthat Hoxa9 deficiency, but not impaired Flt3 signaling, contrib-utes significantly to the increased numbers of CD150+CD342lo
HSCs in the compound knockout mice.Finally, we consistently observed that the CD150+ cells that in-
creased in the compound knockout mice expressed very high levelsof CD150 (Fig. 6). LSK+CD150+ cells expressing high levelsof CD150 have been shown to exhibit latent and myeloid-biasedreconstitution potential in transplant recipients (3). Myeloid-biasedHSC can be further discriminated in adult mice by expression ofCD41 (32). As shown in Fig. 6C, there was a significant increase inpercentage of LSK+CD150+CD342lo cells that expressed CD41 inthe compound knockout mice (p = 0.034, n = 3 mice/genotype).Thus, combined loss of Hoxa9 and Flt3 signaling results in adisproportionate number of myeloid-biased HSC in BM.Taken together, these flow cytometry analyses show that com-
bined loss of Hoxa9 and Flt3 signaling significantly alters theHSC/MPP compartment of BM. LSK+CD150+CD342loCD41+
myeloid–biased HSC are increased, whereas CD1502Flt3+ progen-itors are severely depleted. We conclude that Hoxa9 and Flt3signaling regulates a critical early checkpoint in hematopoietic dif-ferentiation that restricts B and T lineage developmental potential.
Differential role of Hoxa9 and/or Flt3 signaling in regulationof the early program of lymphoid gene expression
Deficiencies in Hoxa9, FL, or both result in significant reduction inlymphoid-biased progenitors andBCP inBM.We andothers (10, 11)have shown that the rare MPP expressing high levels of Flt3 inFlt3l2/2 mice have reduced levels of lymphoid–lineage transcripts,suggesting that Flt3 signaling plays a role in lymphoid priming.Hoxa92/2mice also have reductions in Flt3+hi MPP, although a rolefor Hoxa9 in lymphoid priming has not been reported. Combinedroles for Hoxa9 and Flt3 signaling in regulation of lymphoidpriming provide an explanation for the severe defect in lymphoiddevelopment in the compound knockout mice. To make this de-
FIGURE 5. Reductions in GMLP and LMPP in Hoxa92/2Flt3l2/2
mice. (A) LSK+ cells were discriminated as Lin2 (data not shown) c-kithi
Sca-1+ (boxed region). HSC/MPP, GMLP, and LMPP was resolved within
the LSK+ gate based on differential expression of VCAM-1 and Flt3. (B)
Summary of HSC/MPP, GMLP, and LMPP subset frequencies as a per-
centage of LSK+ cells. Data represents the mean and SD of four to five
mice per genotype. *p . 0.05, **p . 0.005, ***p . 0.0005.
750 Hoxa9 AND Flt3 SIGNALING IN LYMPHOPOIESIS
by guest on May 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
termination, LSK+CD1502 cells from B6 control, Flt3l2/2, andHoxa92/2 mice were sorted into Flt3lo and Flt3+ subsets. Thepaucity of LSK+CD1502 cells, particularly LSK+CD1502Flt3+
cells that are severely reduced, precluded isolation of these cellsfrom the compound knockout mice. CD150 was included as a gatingparameter to exclude alterations to the results contributed by
FIGURE 6. Alterations in HSC/MPP subsets in Hoxa92/2Flt3l2/2 mice. (A) Synergistic reduction in LSK+CD1502Flt3+ cells in Hoxa92/2Flt3l2/2 mice.
LSK+ cells were visualized based on differential expression of CD150 and Flt3 and then fractionated by Flt3 density into Flt32, Flt3lo, and Flt3+ subsets
(boxed regions, left to right, respectively). The boundaries of the boxed regions were determined based on loss of CD150 expression, which accompanies
acquisition of Flt3. (B) Increase in LSK+CD150+CD342lo progenitors in Hoxa92/2Flt3l2/2 mice. (C) Increased percentage of CD41+LSK+CD150+CD342lo
myeloid–biased cells in Hoxa92/2Flt3l2/2 mice. (D) Summary of Flt32, Flt3lo, and Flt3+LSK+ subsets in the four genotypes. (E) Summary of LSK+CD150+
CD342lo progenitors in the four genotypes. (F) Absolute number of LSK+CD150+CD342lo cells per four hind limb leg bones per genotype. (G) Summary of
percentages of CD41+ cells within the LSK+CD150+CD342lo gate. Bar graphs reflect the mean 6 SD of three to five mice per genotype. *p . 0.05.
The Journal of Immunology 751
by guest on May 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
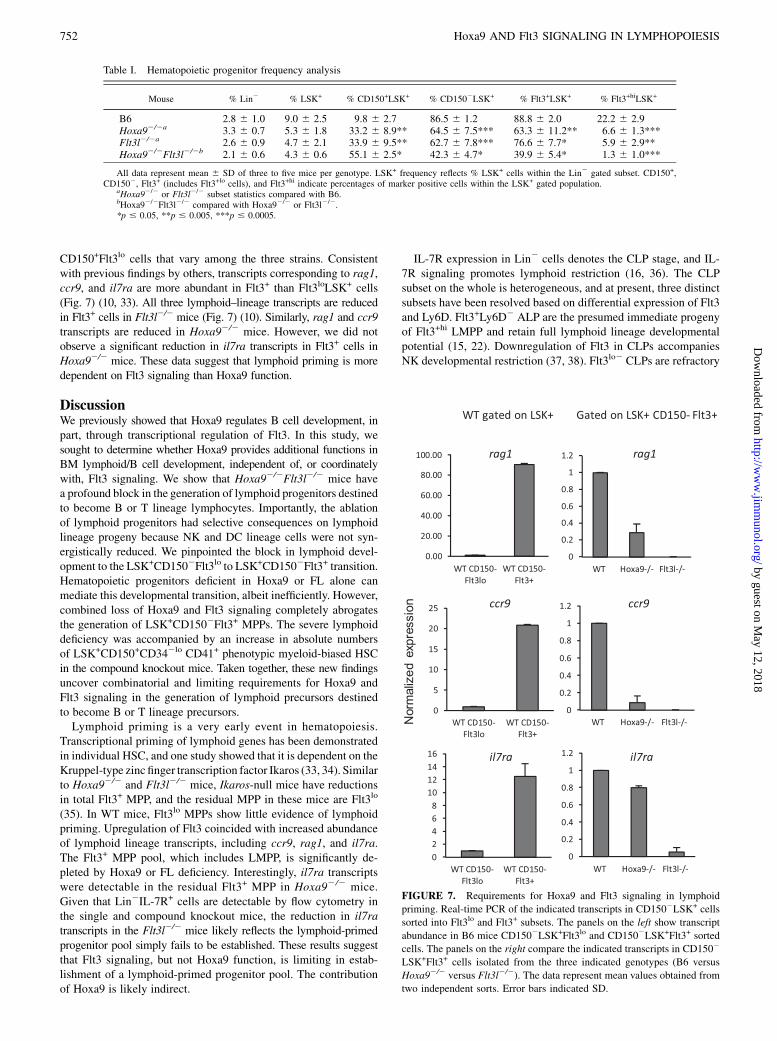
CD150+Flt3lo cells that vary among the three strains. Consistentwith previous findings by others, transcripts corresponding to rag1,ccr9, and il7ra are more abundant in Flt3+ than Flt3loLSK+ cells(Fig. 7) (10, 33). All three lymphoid–lineage transcripts are reducedin Flt3+ cells in Flt3l2/2 mice (Fig. 7) (10). Similarly, rag1 and ccr9transcripts are reduced in Hoxa92/2 mice. However, we did notobserve a significant reduction in il7ra transcripts in Flt3+ cells inHoxa92/2 mice. These data suggest that lymphoid priming is moredependent on Flt3 signaling than Hoxa9 function.
DiscussionWe previously showed that Hoxa9 regulates B cell development, inpart, through transcriptional regulation of Flt3. In this study, wesought to determine whether Hoxa9 provides additional functions inBM lymphoid/B cell development, independent of, or coordinatelywith, Flt3 signaling. We show that Hoxa92/2Flt3l2/2 mice havea profound block in the generation of lymphoid progenitors destinedto become B or T lineage lymphocytes. Importantly, the ablationof lymphoid progenitors had selective consequences on lymphoidlineage progeny because NK and DC lineage cells were not syn-ergistically reduced. We pinpointed the block in lymphoid devel-opment to the LSK+CD1502Flt3lo to LSK+CD1502Flt3+ transition.Hematopoietic progenitors deficient in Hoxa9 or FL alone canmediate this developmental transition, albeit inefficiently. However,combined loss of Hoxa9 and Flt3 signaling completely abrogatesthe generation of LSK+CD1502Flt3+ MPPs. The severe lymphoiddeficiency was accompanied by an increase in absolute numbersof LSK+CD150+CD342lo CD41+ phenotypic myeloid-biased HSCin the compound knockout mice. Taken together, these new findingsuncover combinatorial and limiting requirements for Hoxa9 andFlt3 signaling in the generation of lymphoid precursors destinedto become B or T lineage precursors.Lymphoid priming is a very early event in hematopoiesis.
Transcriptional priming of lymphoid genes has been demonstratedin individual HSC, and one study showed that it is dependent on theKruppel-type zinc finger transcription factor Ikaros (33, 34). Similarto Hoxa92/2 and Flt3l2/2 mice, Ikaros-null mice have reductionsin total Flt3+ MPP, and the residual MPP in these mice are Flt3lo
(35). In WT mice, Flt3lo MPPs show little evidence of lymphoidpriming. Upregulation of Flt3 coincided with increased abundanceof lymphoid lineage transcripts, including ccr9, rag1, and il7ra.The Flt3+ MPP pool, which includes LMPP, is significantly de-pleted by Hoxa9 or FL deficiency. Interestingly, il7ra transcriptswere detectable in the residual Flt3+ MPP in Hoxa92/2 mice.Given that Lin2IL-7R+ cells are detectable by flow cytometry inthe single and compound knockout mice, the reduction in il7ratranscripts in the Flt3l2/2 mice likely reflects the lymphoid-primedprogenitor pool simply fails to be established. These results suggestthat Flt3 signaling, but not Hoxa9 function, is limiting in estab-lishment of a lymphoid-primed progenitor pool. The contributionof Hoxa9 is likely indirect.
IL-7R expression in Lin2 cells denotes the CLP stage, and IL-7R signaling promotes lymphoid restriction (16, 36). The CLPsubset on the whole is heterogeneous, and at present, three distinctsubsets have been resolved based on differential expression of Flt3and Ly6D. Flt3+Ly6D2 ALP are the presumed immediate progenyof Flt3+hi LMPP and retain full lymphoid lineage developmentalpotential (15, 22). Downregulation of Flt3 in CLPs accompaniesNK developmental restriction (37, 38). Flt3lo2 CLPs are refractory
Table I. Hematopoietic progenitor frequency analysis
Mouse % Lin2 % LSK+ % CD150+LSK+ % CD1502LSK+ % Flt3+LSK+ % Flt3+hiLSK+
B6 2.8 6 1.0 9.0 6 2.5 9.8 6 2.7 86.5 6 1.2 88.8 6 2.0 22.2 6 2.9Hoxa92/2a 3.3 6 0.7 5.3 6 1.8 33.2 6 8.9** 64.5 6 7.5*** 63.3 6 11.2** 6.6 6 1.3***Flt3l2/2a 2.6 6 0.9 4.7 6 2.1 33.9 6 9.5** 62.7 6 7.8*** 76.6 6 7.7* 5.9 6 2.9**Hoxa92/2Flt3l2/2b 2.1 6 0.6 4.3 6 0.6 55.1 6 2.5* 42.3 6 4.7* 39.9 6 5.4* 1.3 6 1.0***
All data represent mean 6 SD of three to five mice per genotype. LSK+ frequency reflects % LSK+ cells within the Lin2 gated subset. CD150+,CD1502, Flt3+ (includes Flt3+lo cells), and Flt3+hi indicate percentages of marker positive cells within the LSK+ gated population.
aHoxa92/2 or Flt3l2/2 subset statistics compared with B6.bHoxa92/2Flt3l2/2 compared with Hoxa92/2 or Flt3l2/2.*p # 0.05, **p # 0.005, ***p # 0.0005.
FIGURE 7. Requirements for Hoxa9 and Flt3 signaling in lymphoid
priming. Real-time PCR of the indicated transcripts in CD1502LSK+ cells
sorted into Flt3lo and Flt3+ subsets. The panels on the left show transcript
abundance in B6 mice CD1502LSK+Flt3lo and CD1502LSK+Flt3+ sorted
cells. The panels on the right compare the indicated transcripts in CD1502
LSK+Flt3+ cells isolated from the three indicated genotypes (B6 versus
Hoxa92/2 versus Flt3l2/2). The data represent mean values obtained from
two independent sorts. Error bars indicated SD.
752 Hoxa9 AND Flt3 SIGNALING IN LYMPHOPOIESIS
by guest on May 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

to loss of Hoxa9 function and Flt3 signaling, and we recentlyshowed that Hoxa9 is not essential for the generation or differ-entiation of NK lineage cells (39). At present, there is no evidencefor an LMPP/CLP-independent pathway in NK development. IL-7R signaling is not required for NK cell genesis (40). It is possiblethat NKP can develop directly from LSK+CD1502Flt3lo MPP in-dependent of LMPP/CLP. Future studies using adoptive transferand in vitro assays will directly test this possibility.Flt3+ CLP also have DC differentiation potential (15). Like NK
cells, Hoxa9 is dispensable for the generation and differentiationof DC (39). The generation of committed DC progenitors is notdependent on Flt3 signaling although the cytokine is importantfor DC differentiation and homeostasis (25, 41). An IL-7R2Flt3+
common DC progenitor was described recently (42). Because DCprogeny have multiple development origins and the transcriptionfactor PU.1 is a critical regulator of Flt3 in this lineage, it is notunexpected that DC are not perturbed in the compound mice (15,43–45).We previously showed that Flt3loLSK+ cells from Flt3l2/2 mice
have reduced expression of prosurvival factors, suggesting that Flt3signaling plays a critical role in regulation of progenitor survival(11). Hoxa9, in contrast, has been shown to regulate the prolif-eration of multipotent hematopoietic progenitors in vitro (46).Hoxa92/2 mice are deficient in the serine–threonine kinase Pim-1(47). Flt3 signaling activates Pim-1, and reductions in Pim-1 im-pact B lymphopoiesis (48, 49). Importantly, Pim-1 has been linkedto c-Myb activity, and c-Myb plays a critical role in lymphoidpriming and B cell development (21, 50). Hoxa9 has been impli-cated in regulation of c-myb in leukemic blasts (51). These ex-perimental findings together with the profound lymphoid/B lineagedeficiency manifested by the compound knockout mice suggestthat Hoxa9 and Flt3 signaling regulate lymphoid/B cell develop-ment through regulatory circuits that impact the survival and pro-liferation of Flt3lo MPP. Preliminary analyses of Flt3lo MPPisolated from the single knockout mice did not reveal significantalterations in expression of c-Myb or Pim-1. However, these find-ings do not preclude that similar analysis of Flt3lo MPP from thecompound knockout mice might reveal different results. Futurestudies will be aimed at comparing molecular signatures ofHoxa92/2, Flt3l2/2, and Hoxa92/2Flt3l2/2 total CD150+ andCD1502Flt3lo hematopoietic progenitors to identify genetic circuitssensitive to combinatorial inputs by Hoxa9 and Flt3 signaling.Myeloid-biased HSC exhibit latent reconstitution capabilities in
the early phase in transplant recipients, and the myeloid lineagewas more reconstituted than the lymphoid (3). Lymphoid recon-stitution after hematopoietic stress is contingent on Hoxa9 functionand Flt3 signaling (46, 52). The reduction in LSK+Flt3+ MPP inHoxa92/2Flt3l2/2 mice is accompanied by a marked increase inLSK+CD150+hiCD342loCD41+ phenotypic myeloid-biased HSC.We note that the increase in myeloid-biased HSC did not in-crease myeloid–lineage output in marrow under conditions ofhomeostasis. However, it remains possible that the poor lymphoidreconstitution capability of this subset may contribute to the lym-phoid deficiency in the compound knockout mice.We conclude that there is an extremely early checkpoint in
hematopoietic differentiation critical for B and T lymphopoiesisthat can only be transversed through the cooperative action ofHoxa9 and Flt3 signaling. Our current analysis suggests that thecheckpoint is localized to the LSK+CD1502Flt3lo stage. The se-lective deficiency in lymphoid precursors destined to be B or T cellprecursors makes the Hoxa92/2Flt3l2/2 an exemplary model toidentify and characterize gene regulatory modules and their com-ponents essential for generation of cells of the adaptive immunesystem.
AcknowledgmentsWe thank Virginia Smith Shapiro, Richard Bram, and Paul W. Kincade for
helpful discussions and critical reading of the manuscript.
DisclosuresThe authors declare no competing financial interests.
References1. Singh, H., K. L. Medina, and J. M. Pongubala. 2005. Contingent gene regulatory
networks and B cell fate specification. Proc. Natl. Acad. Sci. USA 102: 4949–4953.2. Wilson, A., E. Laurenti, G. Oser, R. C. van der Wath, W. Blanco-Bose, M. Jaworski,
S. Offner, C. F. Dunant, L. Eshkind, E. Bockamp, et al. 2008. Hematopoietic stemcells reversibly switch from dormancy to self-renewal during homeostasis andrepair. Cell 135: 1118–1129.
3. Morita, Y., H. Ema, and H. Nakauchi. 2010. Heterogeneity and hierarchy withinthe most primitive hematopoietic stem cell compartment. J. Exp. Med. 207: 1173–1182.
4. Wiesmann, A., R. L. Phillips, M. Mojica, L. J. Pierce, A. E. Searles,G. J. Spangrude, and I. Lemischka. 2000. Expression of CD27 on murine he-matopoietic stem and progenitor cells. Immunity 12: 193–199.
5. Kiel, M. J., O. H. Yilmaz, T. Iwashita, O. H. Yilmaz, C. Terhorst, and S. J. Morrison.2005. SLAM family receptors distinguish hematopoietic stem and progenitor cellsand reveal endothelial niches for stem cells. Cell 121: 1109–1121.
6. Osawa, M., K. Hanada, H. Hamada, and H. Nakauchi. 1996. Long-term lym-phohematopoietic reconstitution by a single CD34-low/negative hematopoieticstem cell. Science 273: 242‑245.
7. Boyer, S. W., A. V. Schroeder, S. Smith-Berdan, and E. C. Forsberg. 2011. Allhematopoietic cells develop from hematopoietic stem cells through Flk2/Flt3-positive progenitor cells. Cell Stem Cell 9: 64–73.
8. Buza-Vidas, N., P. Woll, A. Hultquist, S. Duarte, M. Lutteropp, T. Bouriez-Jones,H. Ferry, S. Luc, and S. E. Jacobsen. 2011. FLT3 expression initiates in fullymultipotent mouse hematopoietic progenitor cells. Blood 118: 1544–1548.
9. Adolfsson, J., R. Mansson, N. Buza-Vidas, A. Hultquist, K. Liuba, C. T. Jensen,D. Bryder, L. Yang, O. J. Borge, L. A. Thoren, et al. 2005. Identification of Flt3+
lympho-myeloid stem cells lacking erythro-megakaryocytic potential a revisedroad map for adult blood lineage commitment. Cell 121: 295–306.
10. Sitnicka, E., N. Buza-Vidas, H. Ahlenius, C. M. Cilio, C. Gekas, J. M. Nygren,R. Mansson, M. Cheng, C. T. Jensen, M. Svensson, et al. 2007. Critical role ofFLT3 ligand in IL-7 receptor independent T lymphopoiesis and regulation oflymphoid-primed multipotent progenitors. Blood 110: 2955–2964.
11. Dolence, J. J., K. Gwin, E. Frank, and K. L. Medina. 2011. Threshold levels ofFlt3-ligand are required for the generation and survival of lymphoid progenitorsand B-cell precursors. Eur. J. Immunol. 41: 324–334.
12. Igarashi, H., S. C. Gregory, T. Yokota, N. Sakaguchi, and P. W. Kincade. 2002.Transcription from the RAG1 locus marks the earliest lymphocyte progenitors inbone marrow. Immunity 17: 117–130.
13. Lai, A. Y., and M. Kondo. 2006. Asymmetrical lymphoid and myeloid lineagecommitment in multipotent hematopoietic progenitors. J. Exp. Med. 203: 1867–1873.
14. Medina, K. L., K. P. Garrett, L. F. Thompson, M. I. Rossi, K. J. Payne, andP. W. Kincade. 2001. Identification of very early lymphoid precursors in bonemarrow and their regulation by estrogen. Nat. Immunol. 2: 718–724.
15. Karsunky, H., M. A. Inlay, T. Serwold, D. Bhattacharya, and I. L. Weissman.2008. Flk2+ common lymphoid progenitors possess equivalent differentiationpotential for the B and T lineages. Blood 111: 5562–5570.
16. Kondo, M., I. L. Weissman, and K. Akashi. 1997. Identification of clonogeniccommon lymphoid progenitors in mouse bone marrow. Cell 91: 661–672.
17. Gwin, K., E. Frank, A. Bossou, and K. L. Medina. 2010. Hoxa9 regulates Flt3 inlymphohematopoietic progenitors. J. Immunol. 185: 6572–6583.
18. Mackarehtschian, K., J. D. Hardin, K. A. Moore, S. Boast, S. P. Goff, andI. R. Lemischka. 1995. Targeted disruption of the flk2/flt3 gene leads to defi-ciencies in primitive hematopoietic progenitors. Immunity 3: 147–161.
19. So, C. W., H. Karsunky, P. Wong, I. L. Weissman, and M. L. Cleary. 2004.Leukemic transformation of hematopoietic progenitors by MLL-GAS7 in theabsence of Hoxa7 or Hoxa9. Blood 103: 3192–3199.
20. Dias, S., R. Mansson, S. Gurbuxani, M. Sigvardsson, and B. L. Kee. 2008. E2Aproteins promote development of lymphoid-primed multipotent progenitors.Immunity 29: 217–227.
21. Greig, K. T., C. A. de Graaf, J. M. Murphy, M. R. Carpinelli, S. H. Pang,J. Frampton, B. T. Kile, D. J. Hilton, and S. L. Nutt. 2010. Critical roles forc-Myb in lymphoid priming and early B-cell development. Blood 115: 2796–2805.
22. Inlay, M. A., D. Bhattacharya, D. Sahoo, T. Serwold, J. Seita, H. Karsunky,S. K. Plevritis, D. L. Dill, and I. L. Weissman. 2009. Ly6d marks the earlieststage of B-cell specification and identifies the branchpoint between B-cell andT-cell development. Genes Dev. 23: 2376–2381.
23. Lawrence, H. J., C. D. Helgason, G. Sauvageau, S. Fong, D. J. Izon,R. K. Humphries, and C. Largman. 1997. Mice bearing a targeted interruption ofthe homeobox gene HOXA9 have defects in myeloid, erythroid, and lymphoidhematopoiesis. Blood 89: 1922–1930.
24. Sambandam, A., I. Maillard, V. P. Zediak, L. Xu, R. M. Gerstein, J. C. Aster,W. S. Pear, and A. Bhandoola. 2005. Notch signaling controls the generation anddifferentiation of early T lineage progenitors. Nat. Immunol. 6: 663–670.
The Journal of Immunology 753
by guest on May 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

25. McKenna, H. J., K. L. Stocking, R. E. Miller, K. Brasel, T. De Smedt,E. Maraskovsky, C. R. Maliszewski, D. H. Lynch, J. Smith, B. Pulendran, et al.2000. Mice lacking flt3 ligand have deficient hematopoiesis affecting hemato-poietic progenitor cells, dendritic cells, and natural killer cells. Blood 95: 3489–3497.
26. Hardy, R. R., C. E. Carmack, S. A. Shinton, J. D. Kemp, and K. Hayakawa.1991. Resolution and characterization of pro-B and pre-pro-B cell stages innormal mouse bone marrow. J. Exp. Med. 173: 1213–1225.
27. Rosmaraki, E. E., I. Douagi, C. Roth, F. Colucci, A. Cumano, and J. P. Di Santo.2001. Identification of committed NK cell progenitors in adult murine bonemarrow. Eur. J. Immunol. 31: 1900–1909.
28. Izon, D. J., S. Rozenfeld, S. T. Fong, L. Komuves, C. Largman, andH. J. Lawrence.1998. Loss of function of the homeobox gene Hoxa-9 perturbs early T-cell de-velopment and induces apoptosis in primitive thymocytes. Blood 92: 383–393.
29. So, C. W., H. Karsunky, E. Passegue, A. Cozzio, I. L. Weissman, andM. L. Cleary. 2003. MLL-GAS7 transforms multipotent hematopoietic pro-genitors and induces mixed lineage leukemias in mice. Cancer Cell 3: 161–171.
30. Schwarz, B. A., A. Sambandam, I. Maillard, B. C. Harman, P. E. Love, andA. Bhandoola. 2007. Selective thymus settling regulated by cytokine and che-mokine receptors. J. Immunol. 178: 2008–2017.
31. Lai, A. Y., S. M. Lin, and M. Kondo. 2005. Heterogeneity of Flt3-expressingmultipotent progenitors in mouse bone marrow. J. Immunol. 175: 5016–5023.
32. Gekas, C., and T. Graf. CD41 expression marks myeloid biased adult hemato-poietic stem cells and increases with age. Blood.
33. Mansson, R., A. Hultquist, S. Luc, L. Yang, K. Anderson, S. Kharazi, S. Al-Hashmi, K. Liuba, L. Thoren, J. Adolfsson, et al. 2007. Molecular evidence forhierarchical transcriptional lineage priming in fetal and adult stem cells andmultipotent progenitors. Immunity 26: 407–419.
34. Ng, S. Y., T. Yoshida, J. Zhang, and K. Georgopoulos. 2009. Genome-widelineage-specific transcriptional networks underscore Ikaros-dependent lym-phoid priming in hematopoietic stem cells. Immunity 30: 493–507.
35. Yoshida, T., S. Y. Ng, J. C. Zuniga-Pflucker, and K. Georgopoulos. 2006. Earlyhematopoietic lineage restrictions directed by Ikaros. Nat. Immunol. 7: 382–391.
36. Purohit, S. J., R. P. Stephan, H. G. Kim, B. R. Herrin, L. Gartland, andC. A. Klug. 2003. Determination of lymphoid cell fate is dependent on the ex-pression status of the IL-7 receptor. EMBO J. 22: 5511–5521.
37. Carotta, S., S. H. Pang, S. L. Nutt, and G. T. Belz. 2011. Identification of theearliest NK-cell precursor in the mouse BM. Blood 117: 5449–5452.
38. Fathman, J. W., D. Bhattacharya, M. A. Inlay, J. Seita, H. Karsunky, andI. L. Weissman. 2011. Identification of the earliest natural killer cell-committedprogenitor in murine bone marrow. Blood 118: 5439–5447.
39. Gwin, K., J. J. Dolence, M. B. Shapiro, and K. L. Medina. 2013. Differentialrequirement for Hoxa9 in the development and differentiation of B, NK, and DC-lineage cells from Flt3+ multipotential progenitors. BMC Immunol. 14: 5.
40. Peschon, J. J., P. J. Morrissey, K. H. Grabstein, F. J. Ramsdell, E. Maraskovsky,B. C. Gliniak, L. S. Park, S. F. Ziegler, D. E. Williams, C. B. Ware, et al. 1994.Early lymphocyte expansion is severely impaired in interleukin 7 receptor-deficient mice. J. Exp. Med. 180: 1955–1960.
41. Waskow, C., K. Liu, G. Darrasse-Jeze, P. Guermonprez, F. Ginhoux, M. Merad,T. Shengelia, K. Yao, and M. Nussenzweig. 2008. The receptor tyrosine kinaseFlt3 is required for dendritic cell development in peripheral lymphoid tissues.Nat. Immunol. 9: 676–683.
42. Onai, N., A. Obata-Onai, M. A. Schmid, T. Ohteki, D. Jarrossay, andM. G. Manz. 2007. Identification of clonogenic common Flt3+M-CSFR+ plas-macytoid and conventional dendritic cell progenitors in mouse bone marrow.Nat. Immunol. 8: 1207–1216.
43. Karsunky, H., M. Merad, A. Cozzio, I. L. Weissman, and M. G. Manz. 2003. Flt3ligand regulates dendritic cell development from Flt3+ lymphoid and myeloid-committed progenitors to Flt3+ dendritic cells in vivo. J. Exp. Med. 198: 305–313.
44. Manz, M. G., D. Traver, T. Miyamoto, I. L. Weissman, and K. Akashi. 2001.Dendritic cell potentials of early lymphoid and myeloid progenitors. Blood 97:3333–3341.
45. Carotta, S., A. Dakic, A. D’Amico, S. H. Pang, K. T. Greig, S. L. Nutt, andL. Wu. 2010. The transcription factor PU.1 controls dendritic cell developmentand Flt3 cytokine receptor expression in a dose-dependent manner. Immunity 32:628–641.
46. Lawrence, H. J., J. Christensen, S. Fong, Y. L. Hu, I. Weissman, G. Sauvageau,R. K. Humphries, and C. Largman. 2005. Loss of expression of the Hoxa-9homeobox gene impairs the proliferation and repopulating ability of hemato-poietic stem cells. Blood 106: 3988–3994.
47. Hu, Y. L., E. Passegue, S. Fong, C. Largman, and H. J. Lawrence. 2007. Evidencethat the Pim1 kinase gene is a direct target of HOXA9. Blood 109: 4732–4738.
48. Kim, K. T., K. Baird, J. Y. Ahn, P. Meltzer, M. Lilly, M. Levis, and D. Small.2005. Pim-1 is up-regulated by constitutively activated FLT3 and plays a role inFLT3-mediated cell survival. Blood 105: 1759–1767.
49. Domen, J., N. M. van der Lugt, D. Acton, P. W. Laird, K. Linders, and A. Berns.1993. Pim-1 levels determine the size of early B lymphoid compartments in bonemarrow. J. Exp. Med. 178: 1665–1673.
50. Leverson, J. D., P. J. Koskinen, F. C. Orrico, E. M. Rainio, K. J. Jalkanen,A. B. Dash, R. N. Eisenman, and S. A. Ness. 1998. Pim-1 kinase and p100cooperate to enhance c-Myb activity. Mol. Cell 2: 417–425.
51. Huang, Y., K. Sitwala, J. Bronstein, D. Sanders, M. Dandekar, C. Collins,G. Robertson, J. MacDonald, T. Cezard, M. Bilenky, et al. 2012. Identificationand characterization of Hoxa9 binding sites in hematopoietic cells. Blood 119:388–398.
52. Buza-Vidas, N., M. Cheng, S. Duarte, H. Nozad, S. E. Jacobsen, and E. Sitnicka.2007. Crucial role of FLT3 ligand in immune reconstitution after bone marrowtransplantation and high-dose chemotherapy. Blood 110: 424–432.
754 Hoxa9 AND Flt3 SIGNALING IN LYMPHOPOIESIS
by guest on May 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from





























