Embed Size (px)
Citation preview

ICRAF SOILS WORKING GROUP. NAIROBI, 26-31 MARCH,1979
THB EFFECT OF :JaIl, f/fICROORGANIGMG
ON PLANT PRODUCTIVITY
Y.R.DOMMERGUBG (1 )H.G.DIEM (1 )and F.GANRY
(1) OW3TOM, D.1'. 1 juG, DAKAH, :JBNRGA.L
(2) Ingénieur de Recherche à l'IRAT détaché à
l'ISRA, CNRA, BAMBEY, SRNEGAL

SUMMARY==:::=====
Soil microorganisms favorably or unfavorably affect plant
productivity either indirectly, by actinG upon soil physical
or chemical properties, or directly by interaction with plant
roots.
(1) Benefical or detrimental effects on soil properties con-
cern structure, coating of particles with water-repellent
compounds, redox potential, soil nitrogen status (gains by
N2 fixation, losses through denitrification), availability
of nutrients (especially N,r) and accumulation or elimination
of phyto·toxic inorganic and orgunic c ornpourid e ,
(~) floi l rn i.c r-o o r gu.n.i umo di io c t Ly ll.ffect plan L growth b.y .L1n-
proving or reducing the absorption of nutrients or water up
take(~~m~'~are w~~l .known such as ecto-or endomycorrhizae;
others are not yet even characterized, such as microorganisms
inrlll~inr: prnt:r·(\·jrl "()(îLr;). 'l'llf',v III/l,V ~1.11\\) l'l'(l,l1lt:I' 1~I'(Jwl.ll-I'I'I·~U-
lating substances or protect the plant against certain patho-
gens.
Manipulation of the soil microflora appears to be highly
desirable, but it is difficult to accomplish. Sorne succesS
has already been achieved with direct inoculation, especially
in the case of N2-fixers
and mycorrhizae. Indirect control
of soil microflora by methods involving classical means, ste
rilisation, or the application of specifie compounds is possi-
l i lo lU'UV.ld()J tltuL :jU!tll! rcl1ull'èrnentu are I'u.l.Li.Ll.eu , Altering
the soil microflora by acting through the plant is another
promising possibility.
These processes are discussed with special reference to
th!"i r i rnpo r-trm c o n.nrl UCC1lt'(,'UCI.' 111 t.ro p.i cu.]. uo I Lo .

1. INTRODUCTION------------------------------
By now many agronomists would readily agree that soil micro
organisms affect plant productivity, especially in the tro
pics. Yet this idea took a long time to catch on, except in
the case of Rhizobium, because microbiologists were mainly
concerned with the physiology of microorganisms which had
been isolated and studied in test-tubes or Petri dishes and
were, therefore, out of their natural environment. Another
reason is that the study of complex systems such as the soil
-plant-microorganism system is more difficult than that of
pure cultures.
~n this paper, we shall consider sorne of the mechanisms by
which soil microorganisms favorably or unfavorably affect
plant growth by altering the soil physical or chemical pro
perties, or by directly acting upon the plant itself. Since
other contributors have covered the interactions between
plante and mycorrhizae, or N2-fixin
g microorganisme, (Keya,
1979; Redhead, 1979) we shall only briefly mention the role
of those microorganisms, focusing our attention upon other
groups whose influence is still not always recognized. Two
preliminary remarks should be made here which are related
to the unique conditions that prevail in the tropics. The
first is that when soil water content is not limiting, the
temperature is generally high enough to allow much more vi
gorous microbial activity than in temperate areas. The second
~.~~~ refe~$ to th~ ~hi~~$~h~~~ effect. Since the Qrganic
materials which originate from the plant debris are only to
a slight extent stored as humic compounds and are readily .d·~
composed, most of the microbial life is located on or around
the root system of plants.
2

3
2. INFLUENCE OF MICROORGANISrJIS ON SOIT.J PROPERTIES
==============================================
2.1. Effect on soil physical properties
The role of microorganisms in the genesis and maintenance of
BQ!l ntnlcture hR8 recently bepn revipwP~ (Hppper, 1975).
Our aim here is merely to emphasièe the importance of this
process in the rhizosphere. It has been demonstrated that
there are more water-stable aggregates in the bhizosphere
than in the non-rhizosphere soil (Harris et al., 1964). Sin
ce the number of polysaccharide-producing microorganisms is
characteristically higher in the rhizosphere, it can be assu
mcd that Goil stabilization arounJ the root can, at least to
sorne extent, be due ta the rhizosphere microflora. In tropi
cal ooile, where most of -the microbial population io conccn
trateù in the root zone, it would really be definitively
worthwhile to elucidate the relative importance of the root
itself and that of associated microorganisms in soil struc
ture otabillzBtion. Duch invectigationc chould not bo reo
tricted to free-living microorganisms, (such as Azotobacter
sp, Beij erinckia indica or Li ')omyces starkeyi, whi.ch are well
known polysaccharide producers), but extended to mycorrhizae
which were reported to be involved in sand aggregation and
dune stabilization in colder climates (Koske et al., 1975).
In contrast to this beneficial activity, microorganisms can
be harmful in two ways : by decomposing the aggregating com
pOlmùs originating from plants or microorganisms; and by
coating 0011 vartlcules with water-repellent films (Bonu anû
Harris, 1964). By altering the advancing contact angle of wa
ter with the particles, such films disturb the infiltratmon
of water into the soil, inducing a patchy distribution of
plants and 0. rnur-lce d Lo ao 01' productivi t y , Water r-e peLLeo c y ,
which was mostly attributeù ta basidiomycete hyphae is ~
thought by Griffin (1969) to be of potentially wide impor
tance, especially in semi-arid conditions.

4
Microorganisms may also alter the soil redox potential. Thus the
growth of aerobic microorganisms, most of which grow at the ex-
pens~ of decaying plant debris, may lead to a reduction in the
redox potential, inducinc pr0~csscS which nr0 deleterious to the
plant (sect. 2.2.3.). Alternatively, photosynthetic aleae might
produce oxygen and raise the red8x potential, thus acting direct-
ly or indirectly u pori the p.Lanc .
2.2. Influence on soil ch~~ical-Eroperties
2.2.1. Nitrogen gains and losses through biological
processes
2.2.1 .1. Symbiotic N2
fixation
Since this process has already been reviewed (Keya, 1979), the
discussion of this topic here will be restricted to the effect
of limiting factors, an aspect which is often overlooked. Be
sides the possible inadequacy of native N2-fixin
g micropopula
tions and the attacks of pathogens, especially nernatodes (Ger
mani, 1979), four major factors appear to limit symbiotic N2
fixation in the tropics: moisture stress (especially in semi
arid conditions), soil acidity and associated toxicity, mineral
deficiencies and, in sorne Si+'l~tions, an excess of combined
nitrogen in the soil (Table 1). As long as one factor is oper
ntine, N2
fixation is low or nil and the input of nitrogen to
the ecosystem negligiblc or inexistant. Two examples will illus
trate the unfavorable effect o~ limiting factors. These examples
are related to peanut anù result from field-experiments carried
out at the Bambey Experimental Station, in Central Senegal, over
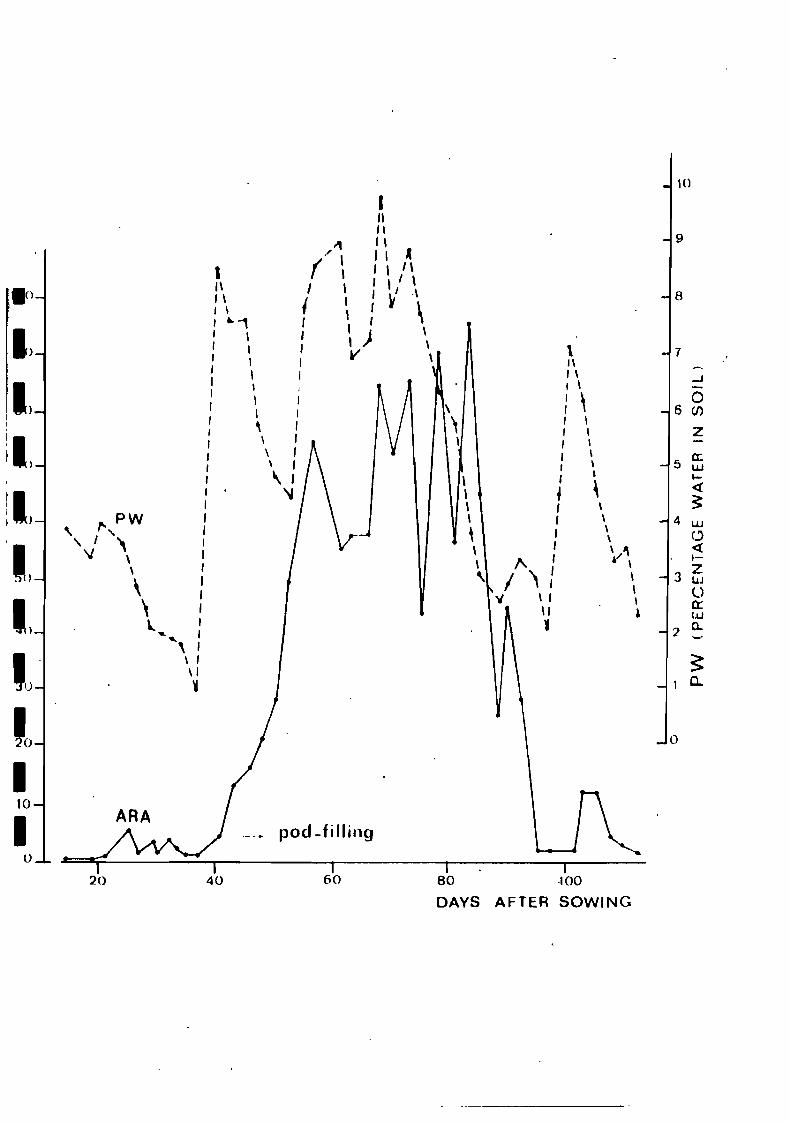
the last 3 years. The first is illustrated by Fig. 1, which
shows that in the arid conditions prevailing in Central Senegal
N2 fixation (measured by the acetylene assay) is closely related
to soil water content. The second example concerns the limiting
effect of inorganic nitrogen. Using the A value method, Ganry
(1976) found that by incrcélsinG' the rate of application of nitro
gen fertilizer from 15 to GO kG per ha, N2
fixation by peanut
decreased from 52 to 25 kg per ha. In spite of those limitations,
sorne N2-fixinc systeme

5
can remain active, which explains that Casuarina equisetifolia,
a non-Ieguminous nodule-bearing tree, largely used for refores-
of West Africa, was reported to-1year on the Cap-Vert peninsula
tins sandy soils on the coast-1fix as much as 60 kg N2ha
(Dommergues, 1963).
2.2.1.2. Losses through nitrification and denitrification.
The activity of nitrifying bacteria varies considerably accor
ding to the soil characteristics and to the nature of the vege
tation. It is weIl known that these bacteria are typically neu
trophilic. However, that does not Mean that nitrification is
restricted to neutral soils; it means that it is restricted to
neutral micro-habitats. Since such habitats may occur (e.g. in
the vicinity of organic debris) in soils whose overall pH is
acid, nitrification can be very active in such soils. Thus
aoid tropical soils grown with banana, maize, or rain-fed
rice exhibit a high nitrifying activity when ammonium fertili
zer is applied. (Dommergues et al), 1978; Chabalier, 1978). In
forest soils, nitrification May be hindered by antibacterial
eubat6noea releaaed by the litterj when the foreet ie oloared,
a flush of nitrification usually occurs (Dommergues, 1954).
There is increasing agreement that nitrification is a detri
mental process Binee it is responsible for two types of nitro
gen losses : losses through leaching, since nitrate is of an
anionic nature and losses through denitrification (Focht and
Verstraete, 1977). Such losses are highly variable, but they
are seldom lower than 20-30% of nitrogen applied as fertili
zero The increased cost and shortage of fertilizer nitrogen,
especially in the tropics, must prompt soil microbiologists
to gather more information on factors that could limit nitri-
fication in soils, since this process is presumably more easi
ly controlled than denitrification. Recents advances in the
field of methodology (especially direct detection of bacteria
in the soil by the fluorescent-antibody technique) promise to
be Most helpful (Schmidt, 1978).

6
2.2.2. Availability of nutrients
2.2.2.1. Nitrogen
#1D tropical soils, amonification is usually very active so
.tbat the potential for the relGase oï ammonium from soil or
sanic nitrogen is high. Unfortunately, the organic nitrogen
,inpu~s (through N2
fixation, root and litter deposition) into
~he soil are often limited, so that ammonium release is not
high enough to meet the plant requirements. It is not clear
whether nitrate, which is the end product of nitrificatio~ is
more available to plants than ammonium.
2.2.2.2. Phosphorus
Microorganisms,·especially those thriving in the rhizosphere,.
are often thou'ght'":' to be able to increase the phosphate avai-
lable to plante by dieeolving water-ineoluble mineraI phoepha
te or by mineralizing phosphate from soil organic matter.
Ae far as mycorrhizae are concerned , their role as solubili
zing·agents has not yet been demonstrated. Other soil micro-
organisms might be involved. In vitro experiments have olear
ly shown that many common microorganisms, including Pseudomonas,
Aohromobacter, Flavobacteriurl, Steptomyces, and especially
Aspergillus and Arthrobacter can solubilize soil phoephorus
(Lowand Walker, 1959; Hayman, 1975; Barber, 1978). However
sorne authors argue that the increased uptake of phosphate may
not only result from ~ increase of the availibility of phos
phate, but could also be explained by the effect on plant
growth of stimulating substances synthesized by the micro-
organisme (sect. 3.5.). With regard to organic phosphate, it
is readi~mineralised by plant phosphatases of the root sur
face. The soil microflora does not seem to increase this pro
cess significantly,
2.2.2.3. Trace elements
Microbially-induced increases and decreases of available trace1;
elements have been recently discussed (Baber, 1978). Since a

7
('WaZ"iety of microorganisms (as well as plants) synthesize" somef .
~xamic acids known to be powerful chelating agents, it is
DOt eurprising that soil microorganisms play a prominent role
. 1D the .i.aon metabolism of plE.~'lts (Waid, 1 975:. A class ical
exemple of the decreased availability of trace elements is
~hat of manganese. Manganese deficiency of oats was shown to
ooour when the activity of manganese-oxidizing microorganisms
Was too high. Soil fumigation reduced the population of these
microorganisms and eliminated the manganese deficiency symp
toms (Timonin, 1946).
2.2.3. Soil toxicity
Phytotoxic compounds that may accumulate in soils are of
microbial or plant origine A classical example of phytotoxicity
induced by microorganisms is that of hydrogen sulfide produced
by sulfate-reducing bacteria. The growth and activity of these
bacteria is triggered in the rhizosphere when the following
environmental conditions exist concurrently : active root exu
dation, soil sulfate content of the rhizospheric soil above a
minimum threshold, and strict anaerobiosis. Accumulation of
hydrogen sulfide can be high enough to lead to the death of
plants (Dommergues et al., 1 s'69. ;Jacq and ROber, 1978). IJlanga
nese toxicity which occurs in acid soils which are relatively
rich in m6.l1ganese may be z-eLrif'oz-c ed by rhizosphere microorganisms
capable of reducing manganic sources. Partial sterilisation of
Buch soils may prevent toxicity (Barber, 1978).
Phytotoxic compounds of plant origin are responsible for dimin
.ishing plant growth when they are not decomposed. Many examples
of such toxic effects have been described by Rice (1974) in his
book on allelopathy. Recently, investigations carried out at the
Agronomie Research Center of Bambey in Central Senegal showed
that sorghum roots contained phytotoxic compounds which, in
Borne circumtances, could significantly reduce the yield of
subsequent crops, especielly sorghum. When sorghum is grown
once in a two-course rotation (peanut-sorghum) instead of once
in a four-course rotation (green manure-peanut-sorghum-peanut)

8
yields are severely depressed. Such a deleterious effect
(known as soil sickness) is induced by the accumulation in
the soil of a phytotoxic compound after the first crop. The
phytotoxic compound, hich is specifically inhibitory to sor
ghum, remains in the soil aS long as environmental conditions
prevent its biodegradation by soil microorganisms. Since such
unfavorable oonditions may prevail in sandy soils for soven)
to eieht monthe ~, the phytotoxic compounds are still
present when sorghum is resown too soon after its last crop
ping. It should be pointed out that while soil sickness does
occur in sandy soils containing kaolinite-type clays and
showing a poor microbial activity, no symptoms are noted in
vertisols, which contain montmorilloni~e-type clays and where
microorganisms are significantly more active. In vertisols,
the sorghum rhizosphere is spontaneously colonized by a rich
soil microflora comprising s~rains that oan actively dooomposB
the phytotoxic compound (Dommergues, 1978b).
Another example of phytotoxicity, which is of importance in
forestry is relnted:.to:· the failure of Grevillea
robusta regeneration in Australia. Seedlings of this species
were reported to be killed by sorne water-transferable factor
associated with the roots of parent trees. The resulting
regulation of population in Q. robusta, is thought to explain
the maintenance of floristic diversity in complex tropical
rain forests (Webb et ~., 1967).

3. DIRECT EFFECT ON THE PLANT=================~========
·As a root grows throue~ soil,it encounters diverse components
of soil microflora and it is directly affected by the activity
of soil microorganisms. Rhizoplane and rhizosphere populations
affect the host plant in mariy we.ys 'out there is now increasing
eyidence that the most important effects of microorganisms on
plant growth concern the mod Lf'd ca.u Lori of plant nutrition and
water uptake, the production of growth-regulating substances
and the protection of roots agaimd; pathogens.
3.1. Modification of plant nutrition and water uptake
by mycorrhizae
The best example of the role of microorganisms aS regulating
agents of plant nutrition is illustrated by mycorrhizal aBSo
ciations, the plant's'mai~ response to nycorrhizal infection
being the increased nptalce of r..utrients, especially of phospho
rus. Mineral nutrition of plants aD stimulated by ectomycorrhi
zae has been weIl treatod by Dowen (19739 and the effects of
vesicular-arbuscular mycorrhizee (VAM)bave been reviewed by
Tinker (1 975) Redhead (1 979) :-,--::d. O-~'!::9rs.
Many theories have already been proposed to explain theinc~CaBed
uptake of phosphorus by ectomycorrhizal reots (Bowen,
1 973). Sorne of them conld a.ppLy 't o VAM since Gerdemann (1 968)
considers that the function cf VAM may also be very similar to
that of the ectomycorrhizae.
It includes the formation of more efficient nutrient-absorbing
structures than non-mycorrhizal roo~s. The extensive etrande of
extramatrical hyphae in VM1 may also explore a much greater vo~-
. ume of soil than non-infected roots, as pyphae' of ectomycorrhi
zal fungi. The postulate of a longer duration of mycorrhizas than
non-mycorrhizal roots as absorbillg organe ae etatad by Bowen and
Theodorou (Bowen, 1973) for eCTootrophic mycorrhizao should also
apply to VAM (Gerdemann, 1 960: ?l.J_cheugh evidence is still lacking.
Another interesting facet of the blology of mycorrhizae is rela
ted to the behaviour of in:fec'~ed z-oo-t e under low water regimes
in the soil. Apart from a high temperature which is the

1 a
common characteristic, tropical soils are quite different from
one another in water content because there is a wide range of
.011 textures and climatesin the tropics.
In sandy soils and is the semi-arid regions plants are often
eubjected to a relatively long period of water stress. A most
~teresting question is whether soil water supplies could be
.1mproved by mycorrhizae. The physiology of water absorption by
~oorrhizae has been little studied but sorne investigations
have indicated a greater drought resistance in a number of
~oorrhizal seedlings (Bo.wen, 1 973).
ID 1971, Safir et al. indicated that VAM could probably decrease
; t the resistance to water transport in soybean. But later
,,(a.fir .21 al., 1 972) they concluded that increased plant growth~
~ater_streSBed conditions wes due to the improvement of phos-
. rus nutrition. Recently Menge et al.(1 978) reported that my-~.,. --
. bizal infection enabled avocado plants to resist transplant
ok, suggesting that mycorrhizae could improve water uptake;:'r ~he host-plant. Drought resistance of rnycorrhizal plants may
be related to the grater exploration of soil by extensive hyphal
growth, but also to large differences between infected and non-
infeoted roota in their own biology. As stateà by Cromer (nowon,
1973), mycorrhizal roots of Pinus radiata seemed to renew growth
more quk:kly than non infected roots when they are subjected to
severe water stress. Another interesting hypothesis on the rela
~ionship between soil water regime and mycorrhizal infection is
given by Sieverding (Moawad, 1978) who found that the amount of
water used to produce 19 of dry matter was much lower in mycor
r.bizal than in non-mycorrhizal plants growing in dry soil ferti-
In.Od with C"5(P04)30H :. (Tab~B 2), Acoording to Moawad, Siever
~te findings may simply be due to the better utilization of
f~ter by p1ants growing in phosphorus deficient eoils. If we wiehr~~ explain the greater drought resistance of plants, the theory
~ water oonsumption economy as stated aboya eeeme to be more
~.'usible and more attractive than the principle of increased
iPtake or transport of water in plants (Safir et al., 1 971 ) •

1 1
'.2. Mycorrhizae under tropical conditions
The impact of mycorrhizal symbiosis in the growth of tropical
plants has been recently discussed by Bowen (1 978) and Black
(1978). Black noticed that the number of tropical plants asSo
ciated with ectomycorrhiz€e appears to be very limited as com
red to the wide range of ectomycorrhizal plants in the temperate
region. The only crop recorded with ectomycorrhizae is Pinus
(Redhead, 1978). Inventories and other informations concerning
ectomycorrhizal forest trees are given in Alewis and Abeynayake
(1978).
As for endomycor~~izae, although sorne " families
. . such as Casuarinaceae, Chenopodiaceae,
yrticaceae are devoid of VAM (Khan, 1974), most tropical plant
species of economic importance are infected : cocoa, tobacco,
ootton, corn, sweet potato, peanut, sugar cane, sorghum, rubber,
~ea, citrus and many species of timber trees (Redhead, 1978).
Spores of VAM are widely distributed in Nigerian soils from the
moist lowland forest to the dry Sahel and Savanna regions (Redhead,
1977). A few olive specimens (Olea cuspidata) may also be infected
by both ectomycorrhizal and endomycorrhizal fungi. Significance
of mycorrhizal symbiosis in the cultivation of olives in Pakistan
bas been discussed by Khan and Saif (1973).
In different soils of the arid and semi-arid regions, it is pro
bable that mycorrhizal associations play an important part in the
crowth and drought-resistance of a number of plants because of
~eir ability to regulate uptake of nutrients and soil water.
Unfortunately, little is known about the mycorrhizal response of
plants usually growing unde~r~onditions. Studies of mycorrhizal
effects in these regions of the world would be of great practical
interest particularly in the case of afforestation with plant
epecies that usually are transplanted. In our laboratory, obser
vations of AZadirachta indica rooto, a trce WhOB0 8ro~th iB wid&-
'spread in dry sandy soils in Senegal, indicate that most roots,
if not all, are infected with VAM (Fig. 2). It is significant to
note that Azadirachta indica is able to grow vigorously in non
fertilized Boils and in arid conditions.

12
,.,. Effect of VAM infection on legume-Rhizobium symbiosie
Aocording to a number of papers, VAM also occurs iB many tro
pical legumes of economic importance e.g. peanuts, cow-pea,
~cropti].ium. atropurpureum, Stylosanthes spp. (Sanni, 1976;
Possingh~o ~t al., 19711 Graw and Rehm, 1977). As legumes have
been shown to require high levels of phosphate for nodulation,
it is likely that mycorrhizal infection may affect the nodula
~ion process and also even N2
fixation. Many authors have alre
ady explored this facet of mycorrhizal response in legumes
(Crush, 1974; Islam et al., 1976; Mosse et al., 1976; Smith
and Daft, 1977). Rooently, in an oxoellent Qseay on the role
of mycorrhizae in legume nutrition on marginal soils, Mosse
(1977) summarized our knowledge on this subject and reported
evidence that endomycorrhizal inoculations associated with the
eupply of rock phosphate stimulated growth and nodulation of
many ~egume&.
On the mechanism of legume stimulation by mycorrhizae, Mosse
wrote that the principal cause is undoubtedly increased assi
milation of phosphate, but mycorrhizae may have other secondary
affects possibly of a hormonal nature.
3.4. Microbially induced proteoid roots
Mycorrhizal associations are not the sole process which stimula
tés the phosphorus nutrition of host plants. Despite resistance
of aeveral lupin species to mycorrhizal infections, they are,
nevertheless, able to grow in sandy soils which are highly defi
oient in phosphorus. The ability of lupin to absorb soil phos
phate has been attributed to the formation of clusters of root
1ets in lovalized parts of the lupin root system. These clusters
of rootlets resemble the dense clusters known as proteoid roots
which have been described in the family of Proteaceae by Purnell
(Trinick, 1977). Other proteoid roots ha~~' also been recorded by
Lamont on Viminaria juncea and by Malajczuk on Kennedia (Trinick,
1977). It h&s now been shown that proteoid roots play an impor
tant role in phosphorus nutrition of plants due to their increa
sed absorbing ability as compared with normal roots (Jeffrey,
1967; Malajczuk and Bowen, 1974).

13
Acoording to the literature published, very few plant species
form proteoid roots. In Senegal, one of the authors (R.G.D.)
observed that rootlet clusters similar to proteoid roots can be
found in Casuarina ~uisetifolia usually growing in sandy and
deficient soils. In the cluster , lateral rootlets are so nume
rous that they resemble fingers (Fig. 3). It is interesting to
note that, c. equisetifolia does not seem sensitive to mycorrhi
zal infection, so that proteoid roots may be an alternative for
this plant to take up phosphate from deficient soils. Investi
gations are now in progress in our laboratory to demonstrate the
effects of these root formations on the physiology of C. esuise
tif01ia. Although the mechanisms of the initiation of proteoid
roots are not clear, sorne inoculation experiments indicate that
proteoid roots may be induced by microorganisms colonizing the
root surface (Malajczuk and Bowen, 1974).
'.5. Effect on plant growth of phytohormone. -producing
microorganisms
As microbial numbers and activity are more intense in the rhizos
phere than in soil, it hardly seems conceivable that the develop
ment of such rhizosphere microflora would not directly affeot the
development of the roots. Microbial effects may be detrimental to
root growth for example roots of tomato, subterranean clover and
radiate pine were stunted by the presence of soil microo~ganisms
(Rovira and ~ic Dougall, 1967). However, particular attention has
been paid to the benefical effect exerted by rhizosphere inhabi
tants. It has been a long tœme since typical rhizosphere bacteria
such as Arthrobacter, Pseudomonas and Agrobacterium were found to
he able to produce substances promoting plant growth (Krasilnikov,
1958) •
Ectomycorrhizal fungi also provide the host plant with phyto
hormones and growth-regulating B vitamins (Slankis, 1973).
Detailed discussion about the direct effects of bacteria on
root growth by the production of plant growth reg~lating fac
k~rs can be found in many reviews (Krasilnikov, 1958; Katne1~!M""8OD, 1965; Brown, 1975). The influence of ectomycorrhizal hor-
~~~8 on the development of roots of the host plant has BlsoIl ~.t. ..

14
been amply demonqtrated in Slankis (1973). However, careful ~~'.
scrutiny of the literature dealing with increased plant growth
resulting from interactions between soil microorganisms and
plants lead us to remark that when plants are artificia1ly inoc-
:ulated with a particular microorganism known for a determined
biological activity (e.g. N2
fixation; phosphorus solubiliza
tion), stimulation of plant growth often appeared to be putative-
:ly attributed to the effect of this specifie activity, although
it may simply be due to the production of phytohormones by the
sarne microorganism. Three examples found in different fields
reinforce this point of view :
(1) Thirty years ago, Gerretsen (1948) thought that the increased
.: growth of plants in sterilized sand containing insoluble
phosphate compounds Was due to inoculation with solubilizing
phosphate bacteria. This conclusion is now criticized by Brown
(1975) who attributes the improved growth reported by Gerretsen
to the production of gibberellin-like substances ,by the bacteria
used for inoculation.
(2) Increases in plant growth and crop yields have often been
recorded after inoculation with Azotobacter, but it is now well
known that these effects are caused, not by fixation of signi
ficant amounts of N2,
but by the production of small amounts of
highly active growth-promoting substances by the bacteria (Brown,
1975). Similarly, inoculation with Azospirillum brasiliense, a
free living N2-fixing bacterium in vogue nt present, can also
induce increased plant growth. Table 3 shows that inoculating
rice with a non N2-fixin
g bacterium stimulated plant growth even
more actively than A. brasiliense. Moreover, since inoculation
with Azoapir;i,llum E{enerf\lly dOflA not nignifioantly improve N2fixation, Gasking and Hubbell (1978) and Tien et~. (1979)
suggested that the effect of Azosp~i+~ inoculation on plant
growth could be due to growth-stimulating substances produced
by this bacterium, as in the case of Azotobacter.
(3) In some experiments mf biological control, root disease of
wheat associated with Rhizoctonia solan~·.was reduced and grain
yield increased by seed inoculation with bacteria and actinomy
oetes. Merriman ~ al. (1974) sugggested that the yield increases

1 5
•
-.. t prir,arily due to plant growth-stimulating factors
~o ~he biological control of root disease.f,t,
j~1';'.6. Improvement of plant resistance to infection
rather
Discussion will be restricted to the control of pathogens through
the improvement of plant resistance by symbiotic microorganisms
or microorganisms more or less loosely associated with the roots,
which is only one aspect of the vast problem of biological con
trol. Two types of mechanisms may be involved in the type of
con~rol studied here.
3.6.1. Protection by mechanical barriers
In his review, Marx (1975) indicatcd that if pine .roots were
:"a8sociated with Leucopaxillus cerealis var. piceina to form
-;eotomycorrhizas, they became rcsistant to infections caused by
euch pathogenic fungi as Phytophthora cinnamomi. Many mechanisms
oould be involved to explain the protective role of ectomycorrhi
.&1 pine roots. Apart from the explanation that antibiotic pro
~duction inhibits fungal pathogens (Marx, 1975), the fungal man-fI'
~~l. of ectomycorrhiza~ also creates effective mechanical barriers
:~acainst penetration by R. cinnamomi. There was further evidence
~~ fungal mantles formed by non-antibiotic-producing ectomy
r·oorrhizal fungi also protected roots from pathogenic root infec1;.
is also suggested that endophytic mycorrhizae may pro
(Wj lhelm, 1 9~·3). In this case, there is no
~"1ons. ItRI~;vide plant protectionl''' r
mPhYsical barrier, but early territorial occupation of living root
H'tissues by the endophyte is anothur principle applicable for bio-1
~10lical oontrol.
3.6.2. Protection by chemicnl response
X~ is well known that ,in. ·thè Pl'e'sence of saprophytic microflora,
~ plants produce a multitude of compounds, especially the so
oa11ed phytoalexins, which can play a role in root diseas~ resis
tance. Most have' been identified in aerial portions, but i t is

16
1ikely that the same compounds cen also be formed in the root
system (Paxton, 1975). For instance, pisatin, the well known
phytoalexin of the pea plant, occurs in the roots as well as
in most other parts of the plant and has a wide spectrum of
antibiotic activity. Strawberry roots also produce phytoalexins
in response to Phytophthora fra~ariae infections (Mussell and
Staplee, 1971).

17
4. MANIPULATING THE SOIL MICROFLORA===================================
lince the major part of the soil population in tropical condi
tions is made up of the rhizosphere microflora, and since the
Z'himosphere microflora must be viewed as a component of the
,~o1e soil-plant-atmosphere system (Dommergues, 1978a), the
[8011 microflora could predictably be manipulated, not only by
;tibectly acting u.pori the microorganisms, but also- indirectly
,~ acting upon the soil and the plant. Actually, direct manip~
r~ation of the soil microflora has been achieved by inoculation
lpractioes, sterilization and application of specifio inhibitors
or specifie substrates. Indirect manipulation of the soil-plant
.~mosphere system has been achieved by classical or non-conven
~ional soil management practices, or by acting upon the plant
oomponent itself.
4.1. Inoculation
In spite of the fact that root colonization by non-pathogenio
microorganisms is still poorly undGrstood, soil miorobiologiste
and agronomists have been trying for' many years to alter the
rhizosphere microflora by introduci~g selected microbial strains,
either by coating seeds with an inoculum, or by placing the ino
oulum into the sail close ta Dhe seed or the seedling.
4.1.1. Inoculation of plants with symbiotic microorganisms
The value of legume inoculation is well recognized, provided :
that the strain used is highly effective and efficient in its
symbioeis with the selected legume cultivar; that it is good
colonizer of the roots and is able to compete with any native
root mioroorganism , and that the proper environmental prereq-
.uisites are fulfilled. However,legume inoculation by classical
Methode ie not alwaye fully eatiefactory (see sect. 2.2.2.).

18
!he value of ec~omycorrhizal inocula~ion if ~he proper environ
.en~al condi~ions are me~ (e.g. Hacskaylo, 1972; Marx and Krupa,
1978) is also generally acknowledged. Inocula~ion by endomyoor-
rhizae is curren~ly a~ ~he experimen~al s~age excep~ in special
~.itua~ions (sec~. 4.2.1.). Preliminary repor~s sugges~ ~ha~ larger
'responses are more likely in ~ropical regions ~han in ~empera~e
regions, because Qf higher ~emperatures and ~he na~urally low
phosphorus level of soils (Hayman, 1978).
aecen~ experiments carried out in the nor~hern coastal area of
Senegal showed ~ha~ inoculating Casuarina equisetifolia with
orushed nodules improved the plant grow~h markedly (Dubreuil
and Andeque, personal communica~ion). Fur~her inves~iga~ion on
the endophy~e of Casuarina is needed in order ~o improve ~he
CUrren~ method of inoculation, which is obviously hazardous
sinoe crushed nodules used as inoculurn May carry pathogens.
4.1 .2. Inoculation of plan~s wi~h microorganisms
which are not stric~ly symbiotic
Whereas ~echniques of inoculation with typicallY symbio~ic micro
organisme (e.g. Rhizobium) are already in use in ~he field, or
could predic~ably be in use in ~he near fu~ure (e.g. endomycor
rhizae), ~echniques of inocula~ion with loosely symbio~ic or non
symbio~ic microorganisms (e.g. rhizosphere N2
fixers or phosphate
80lubilizing bac~eria) cannot ye~ be safely recommended.
;~e first at~empts at using N2-fixin
g rhizosphere bacteria to~fiDoou1a~e grasses or cereale were made as early as 1902 (Ruben-
~, 1963). Since tha~ date many experiments have been performed,
:,t firs~ wi~h ~z~~2bac~~L or BeijerinolEl~ and l&ter with ~
Apiri11um (e.g. Smith et al., 1976; Dobereiner, 1978). Yield
have generally been inconsis~en~.
Field-experiments with phosphate-solubilizing bacteria (espe-
oially Baoillue mega~herium) did no~ show any consis~en~ effec~
on plant yield. According to Barber (1978), "this lack of
response is not really surprising for two ressons.

19
Pirstly, since a considerable proportion of soil phosphorus is
present in organic compounds and up to 90% of the rhizosphere
microflora are capableof~prodUoingphosphatases,the introduction
of other organisms, which would have to compete for available
oarbon sources, is unlikely to cause any increase in the supply
of phosphate to plants. Secondly, the inoculum used, Bacillus
megatherium ~. phosphatic~ is a spore-forming bacterium and
such organisms grow far less ream.ly in the rhizosphere than do
other types of bacteria".
When stimulation of plant growth consecutive to inoculation by
N,..···fixers or phosphate-solubilizing bacteria was observed, it
"'oould not be explained by N2 fixation, ~or by an increase of
phosphate solubilization. The stimulation of plant growth prcb-
ably resulted, at least in part, from the effect of growth
substances produced by the microorganisms added with the inocu-
lum, a.e already mentioned (sect. 3. -5") . In spi te of sorne
recent improvements in the preparation of the inoculum itself
(Dommergues et al., 1979) or in the introduction of mixed cul
tures (Dommergues et al., 1978), there would seem to bé no easy
solution to the difficulties which arise when attempting to
inoculate non-sterile soils.
4.1 .3. Inoculation of soil containing phytotoxic residues
Soil sickness can result from the presence of plant residues in
the soil,especially root litter co,ntaining phytotoxic substances
(sect. 2.2.3.), Inoculating such soils with microorganisms which
actively decompose the root litter appears to be a promising
approach to curing these soils. Thus inoculating a ferrallitic
sandy soil which contained phytotoxic root debris with Enterobao
ter cloacae restored soil fertility (Table 4). Phytotoxic suh
stsno~~,pre-existing in plant residues·or formed during decompo
sition can possees a broad spectrL~m of e~fects which are injuri
".:ous to the roots and stems of pl':l.nts (Tousson and Patriok,
1963). Such a deleterious effect Bould probably be reduced by
Boil inoculation with proper microbial strains.

20
4.2. Soil sterilization and application of specifie compounds
4.2.1. Soil sterilization
If sterilization by heating, irradiation and drying is used in
certain circumstances, sterilization is often achieved by fumi-
sation with such chemicals as chloroform, carbon-sulfide methyl
bromide or chloro-picrin. Such treatments often improve plant
growth even in the absence of pathogens (Wilhelm, 1966; Rovira,
1976). This beneficial effect can be attributed to differer.t ..
Causes: chemical modifications, especially increase of NH4~
oontent, flush of organic matter decomposition, including dead
microorganisms (Anderson and Domch, 1978), elimination of nitri
fying bacteria, which are particularly vulnerable to fumigation
'(Jenkinson and Powlson, 1976), and re-colonization of soil by
non-pathogenic microorganisms, especially pseudomonads, which
are thought to stimulate plant growth (Ridge, 1976).
Soil sterilization prior to inoculation with mycorrhizae appears
~o be most helpful in special situations (Lamb and Richards,1978).
Among these are fumigated nursery soils where severe stunting of
oitrus was reported; inoculation with vesicular-arbuscular mycor
'rbizae appcared to be the best method to overcome this stunting
(Lamb and Richards, 197f.; Timmer a~d Leyden, 1978; Hayman, 1978).
4.2.2. Specifie inhibitors
Among the different specifie inhibitors that have been studied
(e.s. Anderson and Domch, 1975), nitrification inhibitors have
received much attention becausB of their possible use in the
field 1. Besides agronomie practices reported in sect. 4.2.1.,
inhibitors such as 2-chloro-6- (trichloromethyl)-pyridine have
been successfully used to inhibit nitrification, thus increasing
the efficiency of nitrogen fertilizers by reducing deni
~rification and leaching of the nitrate ion. Unfortunatoly,
especially in tropical conditions, the inhibitor is readily
decomposed by the soil microflora so that nitrification occurs
before the plant requirements for nitrogen are at their peak.

21
er reason for the restricted use of nitrification inhibitors
priee. However sorne inexpensive substitutes have been
such aS neem cake (cake mo.de out of the seeds of
rachta indica), but this material is not as effective as
oro-6 (trichloromethyl)-pyridine (Prasad and de Datta1978).
4.2.3. Specifie substr.ates
stimulation of a given component of the microflora can be
adding a specifie substrate to the soil. A classi
example is that of the selective multiplication of actino
etes in a soil amended by chitin, Streptomycetes, and to a
',••er extent Nocardia, constituting the bulk of the chitin
.~ oomposing microflora (Alexander, 1961).
ther example is that of the solubilization of rock-phosphate
F Thiobacilli· These chemoautotrophic bacteria are introduced.'IDto the soil together with sulphu.r which is oxidized to Bulphu-
tric acid, thus dissolving the phosphate (Swaby, 1975).
4.3. Fertilization and soil management
lnooulation ev en with opacifie rni~roorganisrns, especially
Rhizobium, is unsuccessful when one of the environmental limiting
~ .... factors listed in Table 1 is still operating. Therefore,
improvement of the environmental conditions is a prerequisite
which can be achieved by different soil management practices,
such as irrigation, liming, application of organic amendments
or slow-release fertilizers. The benefical effect, of liming is
illustrated by Table 5 (Expt. 1) which reports on a study of
soybean nodulation in a ferrallitic acid soil from Casamance,
Senegal. The increased nodulation was attributed to the elimi
nation of Mn and Al toxicity by liming. Table 5 (Expt. 2),
shows that the application of organic matter even at low rates
(400 kg of peat per ha) favorably affected the growth and nodu
lation of soybean. This last r-e au.Lb confirms those obtnined .. '
by Dart et al., 1973 with Vig~ mungo and y. radiata. Nei
ther species grew well in a nitrogen free sand-grit mixture.

22
:But adding 10;6 of Kettering loam by volume improved growth and~~
rnodulation. When added loam had been previously ignited at~(
_450°C for 4 h to remove soil organic matter, plant growth wae
'poor and the plants eventually died.
The combination of liming, ploughing and farm-yard manure
application was reported to significantly increase peanut
yields in Central Senegal, probably through increasing N2
fixa
tior ('Wey and Obrrtori , 1 978) .
Bince N2
fixation is not always active enough to meet the
legume's requirements, it is necessary to use nitrogen fertilizers.
But it is known that such applications inhibit N2
fixation
_(sect. 2.2.1.1.). To prevent this inhibition in legumes, Hardy
.t al. (1973) suggested the use of other forms of nitrogen
fertilizers which do not inhibit N2
fixation, while providing
the plants with the complementary nitrocrcn roquired for thoir
growth. Such new forms of chemical fertilizers,which they desig
nated as compatible fertilizers, could also be recommended for
use. The possibility, though promising,has not yet been serious
ly explored.
Nitrification can be controlled by such classical methods as
split application of ammonium fertilizers, localization in mud
balls (International Rice Research Institute, 1978), or banding,
which inhibits nitrification due to the high concentration effect
on nitrifying bacteria (Wetselaar et al., 1972; Myers, 1978). The
use of slow-release fertilizers is also recommended to avoid the
harmful effects of nitrification (Fochts and Verstr~cte, 1977).
4.4. Manipulation of the plant component of the eoil-plant-
microorganisms system
4.4.1. Rotations
Introducing a specific'crop in the rotation system has been
used successfully as a basis- of the biological control of sorne
pests. Thus in Florida, soils infested by nematodes pathogenic
to tomato, are cured by growing a grass, Digitaria decumbens,

2'. after the tomato crop (Salette, personal communication). Crop
rotation is often the best method of control of soil-borne phy
topathogenic fungi in cereals (see Baker and Cook, 1974). The
possibility of increasing populations of microorganisms benefi
oial to plants through proper crop rotation was suggested by
Krasilnikov (1958) but has not yet oeen exploited. Though crop
sequence to manipulate the microhilogical balance is a promising
approach, investigations in that field will probably be diffic~t
to initiate and develop because of the large variability of cli
mate and soil conditions.
4.4.2. Plant breeding
Genetic variability in plants responding to Rhizobium infection
is well known. This variability could be used as a basis for the
breeding programs of legumes. The future of this approach was
envisioned a.e follows by Holl and La Rue (1974).. "Plant genes
controlling fixation do occur, and experience shows that we oan
obtain informative and useful variants. There is no obvious rea-
son why symbiotic fixation cannot be increased by genetic means.
We oan envisage cultivars which nodulnte early in ha:t·sh soil
oonditions, fix dinitrogen, even in the presence,"of high Boil
nitrate levels, and continue ~ixing throughout t~eir life. It
appears that ~ixation may be limited by the supply o~ photosyn
thate to the roots. Increased fixation may then require greater
photosynthesis,decreased photorespiration, delayed lodging, or
~ess pod-nodule competition for carbon". Two examples may serve
as an illustration of such a promising approach which has not
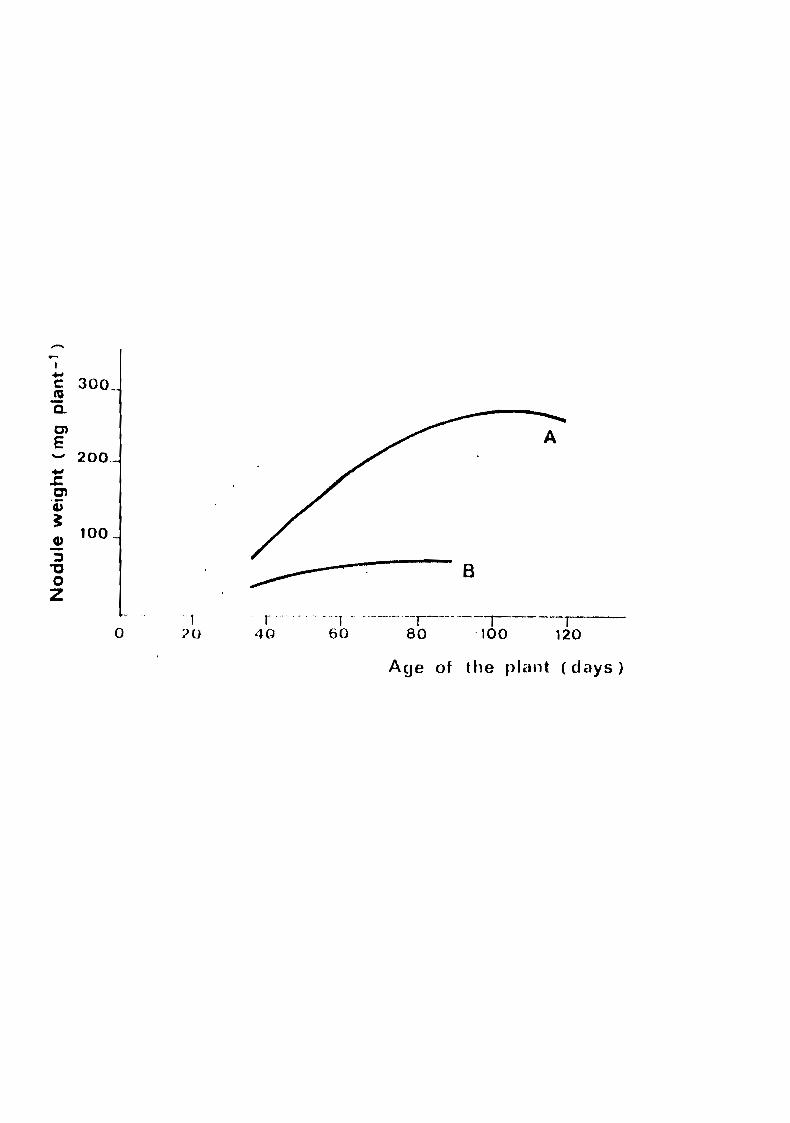
yet been seriously exploited. The first concerns the nodulation
of peanut. Comparing the time course of nodule dry weight of
three peanut cultivars grown at the same time in identical con
ditions (Lior soil, Central Senegal, 1977), Germani (1979) found
that the maximum nodule weight of two of them was much higher
than that of the third (Fig. 4). However, such results should be
interpreted with caution since diiferences in nodule woight are
also ••••••

24
observed from one year to another. Thus the maximum nodule weight
of cv. 55-437 which was only 70 mg in 1977 could reach 100 mg
in ~he sarne soil during more humid years (1973 and 1975) and
even more than 200mg during an even more humid year (1974)
(Wey and Oba~on, 1978). The other example is rela~ed to soybean.
In West African soils, certain soybeen cultivars, such as Mala
yan, are readily nodulated by na~ive Rhizobium of the cow-pea
group, whereas other cultivars, such as Bossier, a high yielding
cv. from the USA, require inoculation with the specifie Rhizobium
japonicum strains. Selection
of high-yielding soybeans which could nodulate with native
Rhizobium of the cow-pea group would allow the developmen~ of
this crop in Africa without requiring any inoculation, since
na~ive Rhizobium of the cow-pea group are common in most African
soils (International Institute of Tropical Agriculture, 1979).

~. 1. Uctho~s ta control Lho cffects of euvLronmen t a I
fat=lors limiling sym1)iotic Il2
fixation
factors
~11 acidity anù toxicity
~neral deficiencies,
~pecially phosphorus dcfi.ciency
.So11 inorganic nitrogcn
[Iathugcns
Mctho~s of control
- Irrigation
- Search for drought-rcsisting
cv. of legumes anù drought
rcsisting ahizobium
- Stimulating VA mycorrhizal
infection
- Liming
A~dition of organic matter
- ALidition of phosphorus
- Stimulating VA mycorrhizal
infection
- Split application of
nitrogen fertili~ers
- Slow-release nitrogen
fertilizers
- Use of compatible fertili~ers
- Search for legwnes \'li th c.J,
lower capacity for nitrate
assimilation
- Chcmical, biological or
integratcd cOntrol
- Crop rotations

'I'ub l o 2. W':lter con sump t Lou (expr o s scd in ml/g Jry wcight) by
EupaLuriullI oJorûLuln I,. a nd 'l'aycLes ercctù L. at; two
Lov e Ls o I' soi1 W.:lt~r cou t.un t, (ÙO urul 20%) (1) und with
two l' [oruis (a tt.c r :lodwdLl, 1978).
NM 1208 1207 2860 4112
ri 1237 1177 1574 1436
NM 1073 1005 2563 3397
r-1 923 1060 1180 1424
inoculateJ wi~h VA mycorrhiza
80% (I) 20% (I)sos (1) 20% (I)'1'realmcnL
ritp,;cies
~r.:ltum
"
l)rcctûrr.:IJ: Non
.•: Inoculatcù with VA mycorrhiza
~oil W.:ltcr I::ontont cxprcsscu as ~ aVûilablc water.

~e ~n~nc..a ~ i~la~ ~ A 'ir~ ..' __
bacterial s~ain (?e=~.) and a ~ixt~e 0= both s~ains
parts or roots of 17 day-old rice seedlings grown in a
or a mixture of the same soil and coarse sand (Exp~.2)
anpublishe~ data, 1979).
__s_e~ 7~ ~in"'i:::1
upon the dry weight 0= aeria:
sterile alluvial soil (EX?~.l)
(Gauthier and Rina~jo,
Dr~l weight 0= aerial parts (::tg) ~ry weiqht 0= roots (mg)
Control Sp 7 Pect. Sp7 + Pec~. ~ontrol Sp 7 ?e~~. Spi' + ?ect.
EXDt. 1 62 12S xxXn.c:.. ,.. ~ 48 84 xXX
n.o.. n •.:l •• ...... ~ .Expt. 2 173 196 xXX
203xXX 233x XX 13ô 149 161x 187x X
n.d. non decermineë.
x,XX,xxx value significantly different =ro~ control, P=.J5, P=.~l, P=.OOl respectively.
Tne pectinolytic strain (Pect.) .did not fix N2.

'l'au1e 4. Ln ï' ~uence or so i I inoculation with EntcrobActor ~1;oto9
on, the growth .:1I1U ni trogen content of sorghua growR iD
a phytotoxic or non-phytotoxic soil. (Burgos, Tho.1B,
University of Nancy, 1979)
Phytotoxic soil
Non-phytotoxic
No inoculation Inoculation soil
(control)
li parts
~ht (cm) 30 65 60
ili(Jhl, plant) 0.5 . 3.9 2.7
al 1-1 content
Jr plant) 13.9 413.7 4.7
tt i CJ h t
plant) \J.u 8.l 1.3

Table 5. Effcct·of aùdition of caco3
.o r sterile peat on nodul~tlon
and dry weight of acrial parts of soyuean cv. Ch1ppcW4(Iloureau, unpulJlisheù data, 1979)
--
fxPt. 1
'Itrol
aCO
lo6k9 per ha)
1
It. 2
:entrol
rit1400 kg per ha)
1
pH
4 '.0 a
7.0 b
4.0 a
4.0 a
Nodul.e
numlJer
(pcr plant)
14 a
39 b
213 cl
41 b
Noùule
dry weight
(mg per plant)
42 a
112 b
44 a
88 b
Dry wei(jht
of ùerial
(mg par plan t)
3.38 a
3.78 ù
2.16 a
3.03 b
One plant per pot containing 5 k~ of sail froln Sefa Rcsearch Station,
s eneqa l , All plants were inoculated with. IrnL of a 3-ùay o Ld culture
of Rhizobiu~1\ japonicum G2Sp (108 lJactcria per m.l) ; Observations were
made when plants were 6 wecks old. In each cxperiment, numbers in
columns nat having the same lctter diffet, P =.05.

FIGUR~ CAPTIONS
Fj~. 1. Variations of acptylene reduaing activity (ARA per plant)
of fif>ld-grown peanut and of soil water content throughout
the t->p.anut growth cycle as observed in 1977 ut the Bamb.ey
Experimental Station,Centra.l Senegal (Ducerf. 1978).
FIg. 2. Azadirachta indica roote infected ·wi.th VA mycorrhizae.
Fi g •.}. Clueter of root lets (proteoid roote) of Caeuarina equieeti-
foLi n grow i nI{ in 8 sandy BO i l (SanAgal).
Pige 4. Timu couree .of nodule dry weight of peanut expreeeed as mg
par plant. A : cv. 28-206 and GH 119-20; B : cv. 55-437.
Al L d u Ln w('r{~ c oLl.nc t ed 'In lq77 ut Patu e , Central Senegal
(ci p rman i, 1 97 q ) •
~la..
..J
o6 (J)
Z
o
5 ~1e:(
~4 UJ
C)e:(1-
3 ~oCt:tu
'} CL
\l\,\1 \
l ,1 \l ,, ,1 1l 'l '4 \1 \, \1 \1 \,,, ~/\
l " : \v \ l ,\1 ll
,,,\
~\\,
\
~\
80 ·100
DAYS AFTER SOWING
60
1"1\
" 11,/ 1 1 ~
(\ " 1\l ,1 \( \ i~/\
: \ ) "1 V \1
pod ifillinq
4020
11\1 \, \1 .. -,, \1 1l ,
1 \l ,1 \, 1
: \, \1 \1 \, \. 1, ~
1
'''''pw :, l , 1
"\ 1\ ,, 1, :
l 1........ 1
, 1\ ,\ ,~
u
!l,l,1)lu
'0110
1
B
..-'---T ... ------·--r---.----r------ 160 80 '100 120
......-
1+'c 300_CO-C.
C)
E- 200......cC)
·cü~
100CI)-:::J"C0Z
0. 1~o
A
Age of the plant ( day s j

A 1.~:j(ANj)I~H, M. (lO(Jl). l n t r-od u c tl o n to o o i I mi c r-o b i o l o gy , John WïJ(',y,'
Nnw Yo r-k , 4'12 l'P.
AI,WI:;, Il.1'. d(~ llnd AB~~YNAYAK(~, K. (1 ~'(B) • ln: Proe. InternatJonal
'NorkHhop on 'froId ca 1 Mycorrhiza Heoearch. Kumars d ,
::ip.ptembnr lq'TH. IP~). Stockholm. 135-1?5.
ANDI~IW()N, .r . l' .1';. ami ]lOM~)CH, K .11, (1 07B). :30il Bit)l. Hiochem .. 10'.
HAKIm. K."'. IInd COOK. H.,I. ( 1 q74 ) • Aiological control of' plant
fll1thr>f~l'nn. V.H. 1·'r·N.... man. ~lu.n ~'rnncloc('). 433 pp.
( 1 Cl'TH) • ln: Inte['action6 b .. twcp.n non-pathosenic soil
microorgl1ni~mo and planta. (Y.H. Dommergues and
"V L' !~i~)• r , • 1\ ru pu ,l, S • ~nnpvjer, Amnter·dum. 131-162 •
HI,ACK. H. ( 1 I)'{H ) • ln: l'roc. lnl~~rnatjonal Workohop on Tropical
BONII. fi.ll.
Myeordl i zu. Htw('Hrclh. KUlnua.i. Beptt'~mbl'r 1978, IFS,
~it;ockho1rn. '15-B6.
BONIJ, H.II. und IJAHHI:i ••J.H. (1 1164 ) . ~tr. J. ~50il HfHl. , 2,111-
BOWr~N, G.U. ( 1 Q'n) . ln: l~etom.Yeorrh.izH.p.. (<i.e. Mf.t.rko and T.T.
KOZI.OW:;K l '·::10.) Academie l'r"pAB, Np.w York H.nd
l.ond o n , 1~1-1l)·T.

BOWBN, G.D. (1978). Ina Proc. International Workshop on Trppi 0a1
MYcorrhiza Research. Kumaei, September 1978, IFS,
Stookholm, 31-71.
CALDWELL, B.B. and VEBT. H.G. (1977). In:A'Treatise on Dinitrogen
Fixation. (R.W.F. Hardy and W.S. Silver Eda.) Wiley
and Bons, New York, 557-576.
,.~-.
CHABALIBR, P.P. (1978). Ina Proc. Workshop on Nitrogen Cyoling in
West Atrican Agrosystems. Ibadan, December 1978 (in preaa~
CRUSH, J.R. (1974). New Phytol., 13, 743-749.
DART, P., DAY, J., IBLAM, R. ar-d DOBERElNER, J. (1973). Inl Sypbio
tie Nitrosen Fixation in Plants. (P.S. Nutman Ed.)
Ca~bridge University Press. 361-384.
DODBRElNER, J. (1977). Ina Proc. Advisory Group Meeting op the
Potential Use n1_J~~~~~n thu Study or Biolo,ieel
Dinitrosen Fixation.. Vienna, November 1977 (in press).
DOMMBRGUBS, Y.R. (1954). Mem. Inst. S~t. ~~dagasoar. D, 6, 115-148.
DOMMBRGUBS, Y.R. (1977). In: Proc. ~Ao/sIDA Regional Workshop on
Atrica on Crianto Recycling in Agriculture. Buea
Cam~roan, Deoember 1977 (in press).
DOMMBRGUB8, Y.R. (1978a). Int Interactions between non-patholenie
Boil Microorganisme and Plants. (Y~R. Dommergues and
B.V. Krupa Ede.) Elsevier, Amsterdam, 1-36.
DOMMBRGUBS, Y.R. (1978b). In: Interactions between non-pathogenic
Soil Microorganisme and Plants. (Y.R. Dommergues and
S.V. Krupa Ede.) Elsevier, Amsterdam, 443-458.

DOMMEIWUltJU, y .R •• CO~1H1mMONT, H•• HBèK, G. and Or.r.AT, C. (1 ~·6o).
Hev. l~co.1. IHol. :Jol., 6, 115-129.
1JOMM~~HGU~lU, Y.R •• GANRY, F. and GARCIA, J.L. (1978). In: Prec.
Regional Workshop on Nitrogcn Cycling in West African
Ecosyt:ltema. Ibadan, pecember 1978. (in press).
UOMM~RGUlm, Y.R., DJEM. Il.G. and UlVTES, Ch.' (197 Q ) . Appl. Environ
M.i crobiol. Cin press) .
IJlJCERF, l'. (1 97H) • Synthèse d e a travaux effectués sur la modé1 iso.
tion ùe la fixation d'azote d'une culture d'arachide
QU ~énéga1 (année 1 976-1 977). Inati tut Bénégillais de
Recherches Agricoles, CNRA Bambey. Doc. mimeo. 42 pp.
FOCHT. V.U. nnd V]ŒSTRA~T~. W. (1977). In: AdvanceB in Miorobio.l
Ecology.Vol. 1. (M. Alexander Ed.) Plenum Press, 135
154.
GA:JKTN~~. ·.M.JI. and llUUUEI,T., D.H. (1978). In: l'roc. The :::loil-Root
fnterfaCA 8ympoGium. OXfOld i~78. (ahstrnct).
GE(~D~MANN, J.W. (1960). Ann. Hev. l'hyto., 6,397-418.
GERMANl, G. ('1979)-. (SHme Workshop).
GERR~TS~JN, Jo'.C. (1 948). Plant Soil, 1, 51'-81.
GRAW, D. and REHM, G. (1977). J. Agron. Crop geL, 144,75-78.
GRIFFIN, D.M. (1969). Ann. Rev. l'hytopo.tbol., 7, 2B9-'}05.
JlACSKAYLO, "Jo (1 072). Bio. Sc:ience, 22, 577-583.
HARDY, R.W.fo'., BURNS, H.C. and HOLSTEN, R.S. (1973). Soil Biol,
Biochem., 5, 47-b1.

AHHH;, R.}<'., GHf<:;)T\<:H:i, G., AI,LEN, D.N •. n nd ATTOR, 0 ••1.
~"AN, . 1> • S • Jn: ~joj 1 MicrobioloK,y. (N. Walhf>r Bd.)
Buttf.'rwnr·thA, London, G7- Ql.
\YMAN, 1). :.i • ( 1 <j'W) • Ill: Inb~ract.lona bf~tween non-pllthogonjc
,'>0]1 Microorgan]amH und PLa.n t e , (Y.R. Dommerguea
and ~LV. Kr-upn \<;ds.) I.;Iopvier, Amsterda.m, 401-442.
~ /' PBH, C. M• ln: ~.ioll Microbiology. (N. Wlllkpr Ed.)
f.\uttf!rworthH, London, 03-110.
>." P.H. and LA mm, 'r.A. 1n: )'roc. 1 st l nternationa1.
:J,yrnpouium Ni Lr'OW't) 1"ixtLt:ion. (w. rL NI,!wton und C. J •
N,ymun r~d8.) I'uLl.man , Wa.shington ~)tu.tp llni.voraity
( 1 /l'lB) • ~)U,"I",U·.Y HI! r.lOrt on
th~> Firf:lt und ~;pcond TnternutionflJ' Trif.l.1u on Nitro
gl!l1 1·'ertj1.i~'>r l~'fricienc,y in Hicn (1 W(?-l ~)'7'l). ],08
Ihu1()(;, 1,H./{unn.. l'h i l i ppi nr-e , . 56 pp.
'.M. H., AYANAHA, A. rrrrd ~;ANlJgR~j, fo'.!':. (1 0'76) • In: l'roc. 5th
u.nnua 1 ConferencCJ' of Ni gurj an :~oc ie t.,y of Mi croblo
&.' Lbud nrr , Ni gHrla. De c e-mbe r- 1 9'(6. Doc. m.Lmeo , 2:5 pp.
C'l!!P. V. A. und fWG rJH , l'. A •
( i n pr0BD).
(1 Q'(8). Cuh. ORSTOM ~k~l·. Biol. 1 13,
~'jlLnr)Y, u. W. (1 <I()l). AUlJtr ••1. 1:30l.! 1~, 40)-411.
t'JKINnON, IJ.~~. a.nd l'OWj,~jON, u.u ,
16'(-17'(.
(1 Q.,6). ~oi l Bjol. H:ioch'~m.! 8,

KATZNfo:l.~ON, fi. ln: r':coJog,v of :;oil-horn(~ flunt l'nthogerUJ.
(K.Jo'. B,LifPl' und W.C. Bnydp.l· Jo~do.) UnJvoroity o I'
CA,lifornia l'rAHs, Be.rk~l(~.y,] A l,OB nge es, 186-20C).
3.0. Worknho fi) •
KHAN, A.G. (,1 lI'l4 ) • J • Ci on. Mie ra b j 01. ., li1, '7 -1 4 •
KHAN, A.G. and SAIP, 8.R. (1 (l73). Po,k. J. Bot. 1 5, 65-70.
KOSKE, ICIL, ~-H1TTON, .I.C. ruid ~)II!,jJ'I'AHD, n.n , (1 Q75) C J B t, . an... O.,
(tranHIHtion 1 11(1). O-rîicn o f Technical Services.
U.:J., /)'~pt. Commor , , Waohington, 474 pp.
LAMB, R.J. Hnd HICHAHD8, B.N. (1978). Uoil Biol. Biochem .• la.
2Tj-276.
MAI,AJCZUI<. N. und BOW!':N, G.1>. (19'74). Nature, London. 251,316-317.
MARX. u, H • ( 1 075 ). In: BioJotdcul control of Soi l-borrH~ Plant
] 'u. thogeno. (Ci. W. Br-u e h 1. gd.) The Ame rican Phyto po.-
thological :Joei e t y , St . Pau L; 112-115.
MARX, O.B. and KRUl'A, .~~.V. (1978). In: Interactions betwcen non-
VB·thoglmic ~)oil Microorganisme and Plante. (Y.R.
Dommerguee and ~J. V. Krupa Ede.) Eloevier, Amsterdam,
';'7':5-400.
MENG~, J.A .. DAVID. R.M •• ~Ollm~Of'l, ~.J,V. and ZgNTMYER. G.A. (lQ78).
Californin Agriculture. 6-7.
MERRIMAN, l'.H •• l'ltICE, H.!). nnd BAKER, ICF. (1~74). Auetr •.J. Agric.

MOAWAD, M. (1978). Tn: "roc. International Workehop on Tropical
MYCOtTh i zn HeBl'Ht'ch. Kumue i , 3eptember 1978, IF3 ,
~tockholm. 101-110,
MaSSE, B. (19'/7). ln: Exploiting th,~ Legume-Rhizobium Symbiosia
in Tropical Agriculture. (.I.M. Vincent. A,S, Whitney
and J. Bose Ede.) University of Hawaii, NIFTAL Pro-
jüct. 275-292.
MOSSE, B., POwr~',L, C.I.l and HAYMAN, D.S. (P176). New Phy"tol •• 76,
331-:542.
MUSSELL, n.w. and STAI'J,r:a, H.C. (1()71). 'l'hytopr.l.tholoBY, 61, 515-
517.
MYERS, R.J.K. (1978). ,Auatr .•r. r:xp. Agrie. Anim Huab., 18 (in
préss) •
PAXTON, a .n, (1~}'75). [II: Riol Bical and cont],l of ~oil-borne
Plant l'El thogena. (G. W. Bruehl Rd.) The American
t'hytoputhological ~)OCif~ty. 8t. Paul. 185-193.
POSSINGHAM.•J.V •• OBBINK, .J.G. nrid JONr~S, R.K. (1971). J. Austr.
'Agrie. ~ci •• Tr, 160-161.
PRASAD, R. and DE DATTA, S.K. (lQ7H). In: Nitrogen and Riee
SympOB 1um , r,ou Banou , :Jo p t ombe r 1 978 (in press).
Rl'JIJHEAD, J.F. (1977). Trans. Br. M,ycol. Soc •• 69, 275-280.
REDHEAD, J.F. (1978). In: l'rop. In j... ernational Workshop on TropicaJ
Hesearch. KUlOlJH-Ï. :.:leptember 1978, IFS, Stockholm,
121-133.
REDHEAD, J.F. (1979). (Same Workahop).

RICR. E.L. (1974). AllelopathY. Academie Press, New York. 353 pp.
HTDGE, JLH. (1976). Sail Biol. Biochem., 8. 249-253.
ROVIRA, A.D. (1976). Soil Biol. Biochem., 8, 241-247.
IWVIRA, A.D. and McDOUGAI,L, B.M. (1967). In: Sail Biochemis-try.
(A. U. Mcl~a.ren and G. H. Peterson Ede.) Marcel. Dekker,
Tnc, New York, 417-463.
RUBENCHICK, [,.T. (1963). Azotobacter and i'ts use in Agriculture.
Oldbourne J'rase, Lorid ori , 2'{8 pp.
IIlJCHMIDT, .a.j., (1978). Microbiol.ogy, 288-291.
~LANKI:3, V'. (1973). In: 1'Jctornycorrhizae. (G.C. Marks and T.T.
KozlowBki Eda.) Academie Press, New York and
London. 232-291.
eMITH, O.E. and DAFT, M.J. (1977). Austr. J. Plant Physiol., 4.
403-413.
SMITH, R.L., BOUTON, J.H •• SCHANK. S.C., QUESENBERRY. K.H •• TYLER.
l'ot.E•• MILAN, J.R •• GASKIN, M.JI. and LITTELL. J.C. (1976). Science.
ill. 1003-1005.
SWABY, R.J. (1975). In: Sulphur in Auatral.ian Agrieul.ture. (K.D.
McLaehlan Ed.) CSIRO. 'Glen Osmond. 213-222.

TIRN. T.M •• GA:JKIN~. M.H. nnd H1JRHi'~LIJ. D.H. (1 ~79). Apple Environ.
Microhiol. (jn press).
'rlMMEIt. i..«. atlJ l,blYlJgN. H.!".. (1 Q7B). ,r. Amer. :Joc. Hort. ~)cj.,
TIMONIN. M.L. (1°46). ;;oi1 :jci. ~;oc. Amer. l'roc •• 11, 284-292.
'l'INl\lm. \'.h.Jl. (197?). ln: ~;;yrnponia. of the ~'3ocj(?ty for Experimental
Biolo'~Y. ~lymbiotljfJ. Number 29. Cambridge. University
Pr-onn , Cnmbrjdgc. '325-349'.
TOU330N, T.A. and PATRICK. Z.A. (1963). PhytopntholoKY. 53. 265-
~70.
TRINICK. M.J. (1017). New Phytol •• 18. 2 Q7- 304 .
WArD, .J.::J. (1"75). :Joil Hioch"mjatry, 4. (r~.A. l'nul and A.D.
WEBB. L.J •• THACJo:Y. J.G. nnd HAYDOCK, K.P. '(1967) ..J. Appl.
Kcology. 1. 13-2~~
WgT~ELAAH. H•. })A~l~JOURA, .1.130 flnd SINGH. B.H. (1972). Plant Soil,
L!i. 15 Q-175.
WEY, J. and OHATON. M. (1 97H) •.
Agronomie Tropicale, 3~. 129-135.
WIUŒM. s. (1966). Ann. Rcw. l'hytopath., 4. 53-78.
WJJ.H1~M.:J. (19'73). :"Joil Biol. Riochem., 5.729-737.





























