Embed Size (px)
Citation preview

Archives Infernationales de Physiologic ei de Biochirnie, 1968, 76, 299-310 299
R e p le 8 mars 1968.
IDENTIFICATION HISTOENZYMOLOGIQUE DES FIBRES LENTES ET RAPIDES
DANS LES MUSCLES SQUELETTIQUES DES VERTeBReS
PAR
J. BROTCHI {Laboratoire d’dnatornie et Clinique Neurochirurgicale de I’ Universiit de Liege)
(15 figures)
Introduction
Apres une longue phiode oh l’etude fonctionnelle des muscles stribs squelettiques portait sur des muscles entiers, on s’est rendu compte de leur complexitk et de la nhcessite d’aborder cette Btude fibre par fibre. En effet, on classe les fibres muscu- laires en extrafusales et intrafusales, e t on repartit les premieres en fibres rouges ou lentes e t en fibres blanches ou rapides. Des recherches prkcises sur un muscle donne exigent, comme stade prdiminaire, la connaissance de la distribution des divers types de fibres dans ce muscle. I1 existe, bien entendu, des muscles dont toutes les fibres extrafusales sont blanches ou, au contraire, rouges. Leur Btude peut apporter des renseignements tres i n t b ressants, mais le materiel est limit6 : la plupart des muscles ont une constitution mixte, et il faut 6tablir une carte montrant la distribution des divers types de fibres dans le muscle, sinon iles recherches sur des fibres individuelles par implantation de microelectrodes ou par isolement deviennent un jcu de hasard. Voyons quelles sont les techniques permettant d’6tablir la topo- graphie interne d’un muscle.
L’examen macroscopique peut donner des indications prC- cieuses. C’est ainsi qu’au premier coup d’oeil, on constate que la partie profonde du tibialis anterior du rat est plus rouge que ses portions superficielles. Mais la pigmentation, intime- ment like A la richesse en myoglobine, n’est pas en relation etroite avec la lenteur de la contraction musculaire (DENNY- BROWN, 1929). De plus, la couleur des muscles n’est pas uni-
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
cgill
Uni
vers
ity o
n 10
/27/
14Fo
r pe
rson
al u
se o
nly.

300 J. BROTCHI
forme dans le rbgnc animal e t il existe des muscles lents e t rapides chez des especes animales oh tous les muscles ont une teinte assez semblable.
L’etude histologique de coupes transversales d’un muscle apporte des elements intkressants. Par exemple, le reperage des fibres musculaires intrafusales ne pose aucun problbme. Mais, pour les fibres extrafusales, leur diamktre, l’abondance e t la rkpar- tition du sarcoplasme ne sont pas des indications toujours valables. WALLS (1960) a repris les caractkres essentiels des fibres rouges e t des fibres blanches e t avec lui, nous sornmes d’accord pour affirmer que la plupart des fibres rouges con- tiennent un sarcoplasme plus abondant que les fibres blanches mais que ce critbre n’est pas genkral car c’est parfois l’inverse qui se produit. KRUGER (1952) distingue parmi les fibres mus- culaires, la Felderstruktur e t la Fibriflenstrukfur, conditionnees par la richesse ou la pauvrete du sarcoplasme, mais l’identifi- cation des deux structures n’est pas toujours facile. De meme, le diamktre des fibres n’est pas un critere absolu m6me si les fibres blanches ont en gkneral un calibre plus important. La majorite des fibres rouges possedent plus de noyaux que les fibres blanches, e t ces noyaux ne sont pas seulement disposes a la periph6rie des fibres mais egalement au sein de celles-ci; cependant, encore une fois, ce caractere ne s’applique pas a toutes les fibres. Quant a la rkpartition des mitochondries, nous ne pouvons pas lui accorder plus de credit qu’aux autres criteres, car elle n’est pas rigoureuse pour chaque type de fibres dans toutes les espbces animales. La forme de la plaque motrice peut renseigner sur la rapidit6 de la fibre, surtout si l’examen histologique des terminaisons nerveuses est com- plete par une appreciation histochimique de l’activite de l’ack- tylcholinesterase au niveau de l’appareil sous-neural ( GEREBT- ZOFF, PHILIPPOT e t DALLEMAGNE, 1954). Toutefois, ces re- cherches ne peuveni, Btre menees a bien que sur des coupes longitudinales de muscle e t elles exigent, en plus de techniques histologiques et histochimiques assez dificiles, une reconstruc- tion graphique du muscle afin de transposer les resultats sur un plan transversal.
L’analyse biochimique a montrk que les fibres blanches ont plus de glycogkne e t moins de lipides que les fibres rouges. Mais le glycoghe est une reserve qui se consume rapidement e t la
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
cgill
Uni
vers
ity o
n 10
/27/
14Fo
r pe
rson
al u
se o
nly.

IDENTIFICATION DES FIBRES LENTES ET RAPIDES 301
diffkrence entre fibres rouges et blanches s’attbnue apres l’effort musculaire. De meme, la teneur en lipides peut varier forte- ment. Comme exemple, citons l’accumulation de graisses dans la periode prbmigratrice dans les muscles des Oiseaux, pouvant aller jusqu’a 43 yo du poids total du corps (FARNER, 1960). D’autres recherches biochimiques pourraient avoir plus de valeur. Malheureusement, dans tous les cas, elles portent sur des masses musculaires relativement considkrables et ne peuvent Btablir la distribution exacte des types de fibres.
L’Btude histochimique restait la seule solution. En ce qui concerne le glycoghe et les lipides, les objections adresskes a l’analyse biochimique s’appliquent ici. Cependant, dans de nom- breux cas, cette recherche reste valable. Comme les techniques, en particulier la detection des lipides, sont faciles et ne demandent pas une instrumentation onkreuse, on peut con- seiller leur emploi dans les services de physiologie ne disposant pas de la collaboration de morphologistes expkrimentks. Mais, dans le cadre des recherches menkes au laboratoire du Pro- fesseur GEREBTZOFF sur l’histoenzymologie des tissus nerveux et musculaire, nous avons Bt6 orient6 vers une mkthode qui permet de classer chaque fibre musculaire en rapide ou lente. Nous avons eu soin de choisir des muscles dont les critkres klectrophysiologiques sont bien connus, soit par leur vitesse de contraction, soit par la qualiti! de la depolarisation locale ou propagee, soit par leur frkquence de fusion par seconde. Notre distinction entre ces differentes fibres repose sur leur activitk .enzymatique respective e t en particulier sur leur activitk deshy- drogknasique (l). Signalons avant tout que cette activite ne se modifie pas de fason perceptible quel que soit l’btat fonctionnel du muscle : il nous est impossible de distinguer entre une coupe de muscle au repos et une coupe de muscle stirnulk, m&me lorsque cette stimulation a Btk realisbe pendant une heure aux limites de la fusion tetanique.
Materiel et metbode
Nous avons utilisk une technique histochimique dkrivke de la m6thode de FRIEDE (1959) pour la dkshydrogknase du succinate
(l) Une note preliminaire a deja 6te publiee : BROTCHI e t MLADENOV (1967).
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
cgill
Uni
vers
ity o
n 10
/27/
14Fo
r pe
rson
al u
se o
nly.

302 J. BROTCHI
(GEREBTZOFF et BROTCHI, 1965, 1966; GEREBTZOFF, 1966). Les muscles pr6lev6s sont immediatement congelCs A -70 O C puis on pratique au cryostat a -20 O C des coupes de 7 a 10 p d’kpais- seur. L’incubation se fait, a la temperature correspondant a la temperature interne de l’animal 6tudi6, dans un milieu tam- ponnb au Tris-HC1 (pH : 7.4) auquel nous ajoutons le substrat adequat (isocitrate, lactate, ... etc), le coenzyme necessaire au transfert d’hydrogkne (NAD, NADP) e t un accepteur d’hydro- gene qui est le tetranitro bleu de tetrazolium (TNBT), lequel est reduit en un precipite colore de formazan. I1 faut noter que la deshydrogenase du succinate agit en l’absence de NAD ou NADP, et c’est pourquoi pour elle, e t pour elle seule, nous n’avons pas ajoute de coenzyme au substrat.
Notre mkthode histochimique met donc en evidence l’acti- vite de deshydrogenases ( DHases) dont l’accepteur d’hydro- gene est le NAD ou le NADP e t en donne la localisation g r h e a une detection de la deshydrogenase de la lipoamide (NAD reduit : lipoamide oxydoreductase, E.C. 1.6.4.3) intimement lihe a la deshydrogknase (GEREBTZOFF e t BROTCHI, 1965).
RBsultats
L’intensitb de la reaction histochimique varie d’une deshydro- gknase A l’autre e t la localisation de grains de formazan n’est pas uniforme pour toutes les fibres musculaires. Certaines montrent une positivite trhs forte, comme les DHases du succinate (succinate : (accepteur) oxydoreductase, E.C. 1.3.99.1) (fig. 1) , du malate (L-malate : NAD oxydoreductase, E.C. 1.1.1.37), de l’isocitrate (thrio-D,-isocitrate : NAD oxydoreductase (dkar - boxylante), E.C. 1.1.1.41), de l’isocitrate (NADP) (thrdo-D,- isocitrate: NADPoxydoreductase (dbcarboxylante), E.C.1.1.1.42), du lactate (L-lactate : NAD oxydoreductase, E.C. 1.1.1.27) e t ses isoenzymes (fig. 2 e t 3). GERFBTZOFF (1966) a montre que la detection histochimique des isoenzymes de la deshydro- genase du lactate met particulihrement en evidence, parmi les cinq isoenzymes isol6s par electrophorkse sur film d’amidon, ceux qui subissent le deplacement le plus rapide (isoenzyme 1) et le plus lent (isoenzyme 5) vers I’anode e t qu’il a denomm6s respectivement isoenzyme F (Fasf-tnouing) e t iFoenzymes S (Slowmoving). L’isoenzyme F de la lactate DHase a une acti-
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
cgill
Uni
vers
ity o
n 10
/27/
14Fo
r pe
rson
al u
se o
nly.
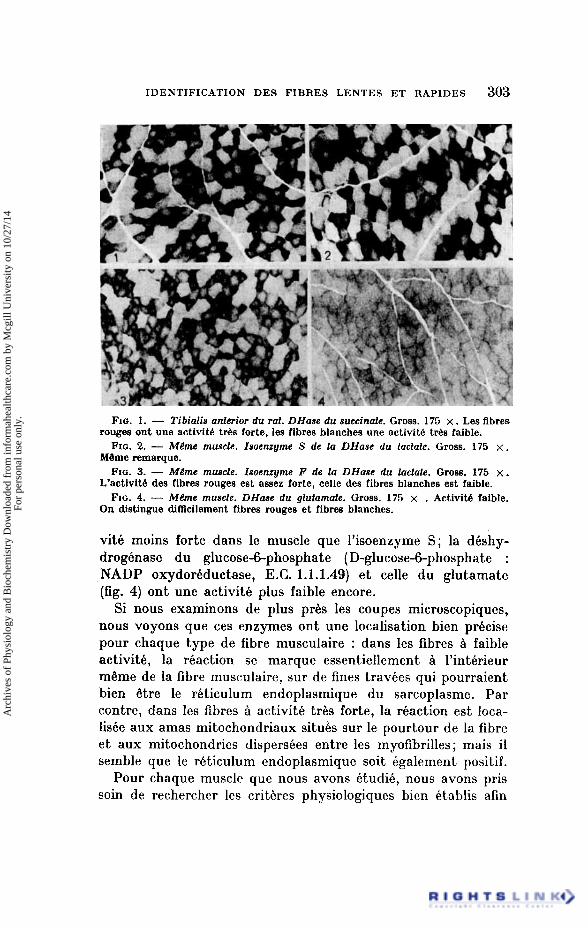
IDENTIFICATION DES FIBRES LENTES ET RAPIDES 303
FIG. 1. - Tibialis anterior du rat. DHase du succinate. Gross. 175 x . Les fibres rouges ont une activitb tds forte, Ies fibres blanches une activitb tr6s faible.
FIG. 2. - Mtme muscle. Isoenzyme S de la DHase du lactate. Gross. 175 x . MBme remarque.
FIG. 3. - MLme muscle. Isoenzyme F de la DHase du lactate. Gross. 175 x. L’activit6 des fibres rouges est assez forte, celle des fibres blanches est faible.
FIG. 4. - Mtme muscle. DHase du glutamate. Gross. 175 x . Activitb faible. On distingue dimcilement fibres rouges et fibres blanches.
vitk moins forte dans le muscle que l’isoenzyme S; la deshy- drogbnase du glucose-6-phosphate (D-glucose-6-phosphate : NADP oxydorbductase, E.C. 1.1.1.49) et celle du glutamate (fig. 4) ont une activite plus faible encore.
Si nous examinons de plus prks les coupes microscopiques, nous voyons que ces enzymes ont une localisation bien precise pour chaque type de fibre musculaire : dans les fibres i faible activitk, la reaction se marque essentiellement Q l’interieur mQme de la fibre musculaire, sur de fines travees qui pourraient bien Qtre le reticulum endoplasmique du sarcoplasme. Par contre, dans les fibres Q activite trhs forte, la reaction est loca- lisee aux amas mitochondriaux situbs sur le pourtour de la fibre e t aux mitochondries disperskes entre les myofibrilles; mais il semble que le reticulum endoplasmique soit Bgalement positif.
Pour chaque muscle que nous avons Btudie, nous avons pris soin de rechercher les criteres physiologiques bien etablis afin
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
cgill
Uni
vers
ity o
n 10
/27/
14Fo
r pe
rson
al u
se o
nly.
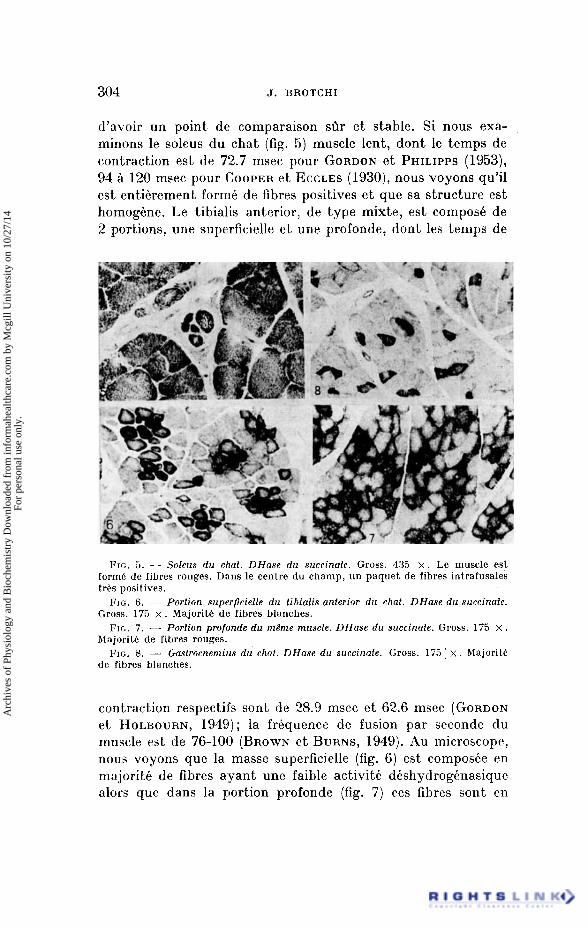
304 J. BROTCHI
d’avoir un point de comparaison sOr e t stable. Si nous exa- minons le soleus du chat (fig. 5) muscle lent, dont le temps de contraction est de 72.7 msec pour GORDON e t PHILIPPS (1953), 94 a 120 msec pour COOPER e t ECCLES (1930), nous voyons qu’il est entikrement forme de fibres positives e t que sa structure est homogene. Le tibia1.i~ anterior, de type mixte, est composi: de 2 portions, une superficielle e t une profonde, dont les temps de
FIG. 5. - Soleus du chat. DHase du succinate. Gross. 435 x . Le muscle est form6 de fibres rouges. Dans le centre du champ, un paquet de fibres intrafusales tres positives.
FIG. 6 . - Portion superpcielle du tibialis anterior du chat. DHase du succinaie. Gross. 175 x . Majorit6 de fibres blanches.
FIG. 7. - Portion profonde du m&me muscle. DHase du succinate. Gross. 175 x . hlajorite de fibres rouges.
FIG. 8. - Gastrocnemiris du chat. DHase du succinate. Gross. 175 x . Majorit6 de fibres blanches.
contraction respectifs sont de 28.9 msec e t 62.6 msec (GORDON e t HOLBOURN, 1949); la frequence de fusion par seconde du muscle est de 76-100 (BROWN e t BURNS, 1949). Au microscope, nous voyons que la masse superficielle (fig. 6) est composee en majorit6 de fibres ayant une faible activiti: dkshydrogenasique alors que dans la portion profonde (fig. 7) ces fibres sont en
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
cgill
Uni
vers
ity o
n 10
/27/
14Fo
r pe
rson
al u
se o
nly.

IDENTIFICATION DES FIBRES LENTES ET RAPIDES 305
minoritk. Le gastrocnemius (fig. 8) a une frkquence de fusion par seconde de 100 (COOPER et ECCLES, 1930), voisine donc ou un peu supkrieure a celle du tibialis anterior, e t nous voyons que sa structure ressemble a la portion superficielle du tibialis anterior, qui est la portion rapide. Le crureus dont la vitesse de contraction est de 58.2 msec (GORDON et PHILIPPS, 1953) a une structure relativement uniforme, ressemblant d’assez prhs a celle du soleus. I1 s’agit bien d’un muscle lent, la difference de vitesse de contraction avec celle du soleus ktant minime. L’extensor digitorum longus a une vitesse de contraction de 19.6 msec (GORDON et PHILIPPS, 1953), e t est donc trCs rapide; au microscope, il comprend un pourcentage trhs klevk de fibres de faible activitk enzymatique et a une structure gknerale assez semblable a celle de la portion rapide superficielle du tibialis anterior.
Pour le diaphragme, les rksultats sont un peu differents. Les physiologistes le considerent comme rapide (GOFFART et RITCHIE, 1952; CREESE, 1949) et sa vitesse de contraction est de 16 a 25 msec chez le rat tandis que sa frkquense de fusion par seconde chez le chat est de 120-140 (ADRIAN et BRONK, 1928). Les pharmacodynamistes le tiennent pour un muscle lent en raison de sa reponse a l’acktylcholine et aux acktylcholi- nomimktiques (ZAIMIS, 1951 ; PHILIPPOT et DALLEMAGNE, 1951 ; DALLEMAGNE et PHILIPPOT, 1951). Les recherches histochi- miques sur les plaques motrices dans ce muscle (GEREBTZOFF, PHILIPPOT et DALLEMAGNE, 1954) tendaient a confirmer le second point de vue. De notre cbtk, si nous examinons l’acti- vitk des dkshydrogenases, nous voyons que le diaphragme est de composition uniformkment mixte, ce qui cadre d’assez pres avec les donnkes de GUNTHER (1950) qui trouve 42 yo de fibres musculaires a Felderstriiktur, c’est-&dire de fibres lentes. Nous reviendrons sur cette question dans la discussion de nos resultats.
Le soleus, le tibialis anterior e t le diaphragme du rat ont une activite deshydrogknasique semblable a celle de ces muscles chez le chat.
Chez le poulet, nous avons 6tudik deux muscles dont les caracteristiques klectrophysiologiques sont bien connues : le latissimus dorsi anterior et le latissimus dorsi posterior avec des temps de contraction respectifs de 100 et de 14 msec (GINSBORG, 1960). La portion anterieure contient uniquement des fibres B
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
cgill
Uni
vers
ity o
n 10
/27/
14Fo
r pe
rson
al u
se o
nly.

306 J . BROTCHI
activitk enzymatique tres intense, alors que la portion postk- rieure ne contient que des fibres a activite trBs faible (fig. 9). Quant au biventer cervicis (fig. lo), electrophysiologiquement mixte (GINSBORG, 1960), il apparait au microscope comme uni- formkment compos6 d’un mklange harmonieiix de fibres po- sitives e t de fibres presque nkgatives.
FIG. 9. - Latissimi tiorsi anlerior fu gauche) et posterior du poulet. DHase du succinate. Gross. 7 x . Remarcper I’opposition d’activitb entre les deux muscles.
F I G . 10. - Biuenter ceroicis du poulet. DHase du succinate. Gross. 175 X .
Chez la grenouille ( R a n a temporaria), nos expkriences re- coupent les donnkes physiologiques apportees par KUFFLER e t VAUGHAN WILLIAMS (1953). Comme muscles rapides ou les potentiels d’action sont propages, nous avons etudii: le sartorius e t l’adductor longus. Le premier (fig. 1 1 ) est formi: essentiel- lement de fibres assez peu positives mais est bordi: sur sa face superficielle par deux a trois couches de fibres activiti: enzy- matique importante, done a caractkre lent. L’adductor longus (fig. 12) a une stucturc relativement homogkne et , dans sa masse de fibres a activitk enzymatique faible, quelques fibres posi- tives seulement sont diss6mini:es. Comme muscles lents oh la depolarisation est locale, non propagke, nous avons ktudii? l’ileofibularis, le seinitendinosus e t le rectus abdominis. L’ileofi- bularis (fig. 13) contient un amas important de fibres positives couvert d’une masse de fibres negatives. Cet amas correspond au Tonus Bundel qui rkpond a l’acktylcholine par une con- tracture prolongee ( SOMMERKAMP, 1928). Le semitendinosus possBde dans son petit chef, un pourcentage eleve de fibres positives tandis que son grand chef (fig. 14) contient un faisceau compact de fibres positives, dans une masse de fibres nkgatives, tout A fait comparable au Tonus Bundel de SOMMERKAMP pour
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
cgill
Uni
vers
ity o
n 10
/27/
14Fo
r pe
rson
al u
se o
nly.

IDENTIFICATION DES FIBRES LENTES ET RAPIDES 307
FIG. 1 1 . - Sartorius de la grenouille. DHase du succinate. Gross. 175 x . Couche
FIG. 12. - Adductor longus de la grenouille. DHase du succinate. Gross. 175 x - FIG. 13. - Ileofibularis de la grenouille. DHase du succinate. Gross. 175 x.
FIG. 14. - Grand chef du semitendinosus de la grenouille. DHase du succinate.
superficielle de fibres rouges.
Les fibres rouges sont dissBmin6es parmi les fibres blanches.
Noter le (( Tonus Bundel )) form6 de fibres rouges.
Gross. 175 x. Dans ce muscle, il y a aussi un (( Tonus Bdndel )).
l’ileofibularis. Le rectus abdominis a une structure uniform& ment mixte mais avec une predominance numkrique de fibres a activitk enzymatique riche.
Chez les Poissons, nous avons btudik les muscles de la carpe dont les critkres biochimiques dkfinis par HAMOIR (1955), corres- pondent bien aux donnkes klectrophysiologiques ktablies par BONE (1966) sur la roussette. On observe une fine couche super- ficielle de fibres rouges au niveau de la ligne latkrale, alors que les muscles rapides, de couleur blanche, forment le reste de la masse musculaire. Nous avons retrouvk chez la carpe un ras- semblement de fibres a activite trbs elevbe des DHases sur toute la longueur de la ligne latkrale (fig. 15). Cet amas affecte la forme d’un triangle a base superficielle e t a sommet interne. Cependant, entre les fibres positives pkriphbriques de la ligne laterale e t les fibres a faible activitk du reste de la musculature, nous devons signaler la presence de fibres A activitk enzyma-
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
cgill
Uni
vers
ity o
n 10
/27/
14Fo
r pe
rson
al u
se o
nly.

308 J. BROTCHI
tique intermkdiaire entre les deux types pr6ckdemment dkcrits. Quelques fibres de ce type intermkdiaire s’observent aussi en plein milieu des muscles de la ligne latkrale.
FIG. 15. - RCgion de la l igne latirale chez la carpe. DHase du succinate. Gross. 175 x . En surface, le muscle de la ligne laterale, tres positif; puis une couche de fibres de type intermediaire; en profonderir, les fibres blanches.
Nous devons maintenant dire un mot des fibres intrafusales que nous avons ktudikes chez le chat e t chez le rat, principa- lement au niveau du tibialis anterior. Ces fibres d’un diametre trks petit ont une activiti: enzymatique tres klevke sur l’en- semble de leur section transversale. Leur sarcosomes, tan t cen- traux que pkriphkriques, sont volumineux e t tres positifs. De plus, on observe une activiti: sarcoplasmique diffuse.
Nous avons enfin ktudik les effets de la dknervation sur la diffkrenciation mktabolique des fibres musculaires du tibialis anterior du rat albinos. En 1962, LEWIS a montrk que la vitesse de contraction du soleus et du flexor digitorum longus ne varie pas dans les quinze premiers jours suivant la dknervation e t diminue durant les troisieme e t quatrieme semaines. La vitesse de contraction du soleus passe de 70-85 msec a 110-150 msec e t celle du flexor digitorum longus de 25-36 msec a 45-65 msec. De notre cBt8, nous avons vu que la diffkrenciation mktabo- lique entre fibres rapides e t fibres lentes s’attknue entre le 20e e t le 3 0 e jour oh le muscle tend B acqukrir une structure ho- mogene surtout par un gain d’activitk enzymatique des fibres rapides. A ce stade, les fibres intrafusales semblent avoir perdu leur intense activitk normale.
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
cgill
Uni
vers
ity o
n 10
/27/
14Fo
r pe
rson
al u
se o
nly.

IDENTIFICATION DES FIBRES LENTES ET RAPIDES 309
Discussion
Ainsi donc, il y a une concordance trks nette entre les para- metres physiologiques et l’activitb des DHases, notamment celle de la DHase du succinate dont la dktection est la plus facile e t la moins onkreuse. Cela s’applique A tous les muscles btudiks, sauf une exception discutke plus loin, e t a toutes les especes animales, du Poisson aux Mammifkres. Signalons que cette btude rbvkle que, s’il existe des muscles constitubs exclu- sivement de fibres lentes, comme le soleus, les muscles rapides des Mammifkres e t de la grenouille possedent un contingent de fibres lentes. Notre technique permet de prbciser le pourcentage de ces fibres dans les divers secteurs du muscle. Chez les Oiseaux, il y a des muscles exclusivement rapides : le latissimus dorsi posterior en est un be1 exemple. Chez les Poissons, les muscles lents de la ligne lathale sont separbs des muscles rapides par une zone de fibres a structure histochimique intermkdiaire. I1 est donc possible d’btablir, griice a cette mbthode, la topo- graphie de la distribution relative des fibres lentes e t rapides pour chaque muscle interessant particulikrement les physio- logis tes.
L’exception signalee plus haut est le diaphragme. Muscle rapide pour les physiologistes, muscle lent pour les pharmaco- dynamistes, muscle constituk presque A kgalitb de fibres rapides et de fibres lentes pour nous. Ce muscle stri6 squelettique a fonction viscbrale reste donc une knigme. I1 en est de m4me pour le muscle dc la ligne latkrale des Poissons dont l’inter- vention dans la motilitb de l’animal est douteuse. Ce muscle ne jouerait-il pas, par rapport a la musculature gknbrale du corps, le rBle des fibres intrafusales?
Conclusions
Nous avons btudib, dans diverses espkces animales, une skrie de muscles stribs squelettiques dont les paramktres physiolo- giques ou biochimiques sont bien btablis. Nous avons vu que les fibres lentes montrent une forte activitk des dbshydrogknases suivantes : deshydrogbnases du succinate, du malate, de l’iso- citrate, de l’isocitrate NADP, du lactate e t isoenzyme lent de cette derniPre. L’activiti: est plus faible pour l’isoenzyme rapide et tres faible pour les dkshydrogbnases du glucose-6-phosphate
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
cgill
Uni
vers
ity o
n 10
/27/
14Fo
r pe
rson
al u
se o
nly.

310 J. BROTCHI
e t du glutamate. Les fibres rapides montrent une activitk beau- coup moins importante que les fibres lentes. Quant aux fibres intrafusales, elles ont une .activiti: enzymatique encore plus elevke que les fibres lentes. Cette activitk histochimique s’est vkrifike Btre en parfait accord avec les critkres physiologiques Zeds ou rapides pour les muscles des diffkrentes especes animales etudiecs, indkpendamment de leur diamhtre, de leur couleur, de l’ahondance et, de la rkpartition de leur sarcoplasme, sauf pour le diaphragme oh on peut soupsonner une adaptation A la fonction respiratoire. Les techniques histochimiques permettent donc non seulement de confirmer la nature rapide, lente ou intrafusale de chaquc fibre musculaire strike squelettique mais de dresser des cartes exactes de la repartition de ces diverses fibres dans chaque muscle.
BIBLIOGRAPHIE
ADRIAN, E. D. e t BRONK, D. W. (1928). - J . of Physiol., 66, 81-101. BONE, Q. (1966). - J . Murine Biol. Ass. U . K., 46, 321-349. RROTCHI, J . et MLADENOV, S . (1967). - Arch. internat. Physiol. et Bioch., 75,
HROWN, G. L. e t BURNS, El. D . (1949). - Proc. Roy. SOC. (London), Ser. B., 136,
COOPER, S. e l ECCLES, J . C . (1930). - J . of Physiol . , 69, 377-385. CREESE, R. (1949). - J . of Physiol . , 110, 450-457. DALLEMAGNE, M . J . e t PHIL.IPPOT, E. (1951). - A r c h . internat. Physiol., 59, 374-376,
Y-BROWN, D. E. (19%!9). - Proc. Roy. Soc. (London), Ser. B . , 104, 371-411.
543-345.
18’1-1 95.
407-422.
FARNER, D. S. (1960). - Proc. X I I t h . Intern. Ornith. Congr. , 1, 197-208. FRIEDE, R . L. (1959). - Exp. Neurol., 1, 441-416. GERERTZOFF, M . A. (1966). - C. R. Soc. Biol., 160, 1323-1325. GEREBTZOFF, M. A. e t BROTCHI, J . (1965). - C. R. SOC. Biol., 159, 2257-2259. GEREBTZOFF, M . A. e t BRCITCHI, J . (1966). - A n n . Histoch., 11, 63-70. GEREBTZOFF, M . A. , PHILIPPOT, E. e t DALLEMAGNE, M. J . (1954). - Acta Ana-
GINSBORG, B. L. (1960). -. J . of Physiol . , 150, 707-717; 154, 581-598. GOFFART, 3%. et RITCHIE, .I . M. (1952). - J . of Physiol., 116, 357-371. GORDOK, G. e t HOLBOURN, A. H. S. (1949). - J . of Physiol . , 110, 26-35. GORDOZ, G. e t PHILI.IPS, C:. G. (1953) . - Quart. J . Exp. Physiol., 38, 35-45. GUNTHER, P . G. (1950). -- Z . Naturforsch., 5, 386-389. I ~ A M O I R , G. (1955). - Adu. Protein Chem., 10, 227-288. KRUGER, P . (1952). - Tetanus und Tonus der quergestreiften Skelettmuskeln der
tomica., 20, 234-257.
Wirbeltiere und des Menschen. Akad. Verlagsges., Leipzig. KCFFLER, S. w‘. e t V A U G H A N \ v I L L I A M S , E. kf. (1953). - J . O f Physiol . , 121,
318-340. LEWIS, D. M . (1962). - J . of Physiol . , 161, 24 P. PHILIPPOT, E. et DALLEMAGNE, M . J . (1953j.- Arch. internat. Physiol., 59, 252-253,
SOUMERKAMP, € I . (1928). -- Arch. exp. Pathol. Pharmakol., 128, 99-115. WALLS, E. \Y. (1960). ~ ~ The Wicroanatomy of Bluscle, in The Structure and Func-
tion of Muscle (edit. by G. H. BOURNE), Academic Press, New York, Vol. 1 ,
357-373.
pp. 21-61. Z A I M I S , E. J. (1951). - J . of Physiol . , 112, 176-190.
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
cgill
Uni
vers
ity o
n 10
/27/
14Fo
r pe
rson
al u
se o
nly.





























