Embed Size (px)
Citation preview

Identi® cation of Diaphragmatic
Crural Component of Gastroesophageal
Barrier in the Rat
CARMEN SOTO, MD, BAOQUAN QI, MD, JUAN A. DIEZ-PARDO, MD, PhD, and
JUAN A. TOVAR, MD, PhD
Manometric assessment of the diaphragm atic contribution to the human gastroe sophage albarrie r is dif® cult because it overlaps with that of the lower esophage al sphincte r. Our aim wasto investigate the barrie r compone nts in the rat in which the gastroesophage al junction iswidely separated from the hiatus. Rats unde r anesthesia (N 5 119) and after musclerelaxation (N 5 14) underwent stationary and pull-through perfusion manometry. Inspiratorytransdiaphragm atic pressure gradient was 5.79 6 1.69 mm Hg and lower esophage al sphincte rpressure was 14.76 6 8.63 mm Hg. A 13.78 6 3.13-mm intraabdominal segment of theesophagus was interposed cranially between the sphincte r and a group of phasic oscillationswith frequency identical to the respiratory rate and pressure of 13.81 6 6.54 mm Hg, whichdisappe ared after muscle relaxation. Both compone nts of the gastroe sophageal barrie r in therat are wide ly separate d by a long intraabdominal esophagus. This arrange ment allowsinve stigation of the behavior of both components unde r challe nging conditions.
KEY WORDS: gastroe sophageal re¯ ux; sphincter; diaphragm; crural sling; manome try; rat; esophagus.
The positive pressure gradie nt between both sides of
the diaphragm accounts for the major gastroesopha-
geal re¯ ux-driving force and is permanently oppose d
in man and many animals by the antire ¯ ux barrier.
The main component of this barrie r is the tone of the
lower esophageal sphincte r (LES), but the participa-
tion of the contractions of the diaphragmatic crural
sling during inspiration has also been acknowledged
and studied under several conditions (1± 5). However,
this second compone nt is dif® cult to identify mano-
metrically because the crura surround the esophagus
in close contact with the LES and cause overlap of
the ir respective manometric pro® les. The participa-
tion of the crural sling in the defense against re¯ ux
and the contribution of its dysfunctions to the patho-
genesis of this phenomenon are not fully known be-
cause of the dif® culties for its inve stigation.
In the past we studied the behavior of the LES in
the rat submitted to various experimental conditions
and con® rmed that intrathoracic and intraabdom inal
pressures generate a gradient similar to that found in
the human which is oppose d by a high-pre ssure zone
corresponding to the LES (6, 7). In addition, some
tracings suggested that the anatomy of its gastro-
esophage al junction and, in particular, the length of
the intraabdominal segment of the esophagus could
allow separate study of the crural sling component of
the barrie r. We undertook the present study aiming at
investigating in more detail the manometric feature s
of the gastroesophageal barrier in this animal with
Manuscript received May 6, 1997; revised manuscript receive dAugust 18, 1997; acce pted August 20, 1997.
From the Department of Pediatric Surgery and Expe rimentalLaboratory, Hospital Infantil Universitario ª La Paz,º Madrid,Spain.
Drs. Soto and Qi were Research Fellows under FIS grants93/0031-1, 95/5451, and 96/5577.
Work supported in part by FIS grants 93/0031-1 and 95/498.Address for reprint requests: Prof. Dr. Juan A. Tovar, Depto. de
Cirugõ Â a, Hospital Infantil Unive rstario ª La Paz,º P. de la Caste l-lana, 261, 28046 Madrid, Spain.
Digestive Diseases and Sciences, Vol. 42, No. 12 (Decem ber 1997), pp. 2420 ± 2425
2420 Digestive Diseases and Sciences, Vol. 42, No. 12 (Decem ber 1997)
0163-2116/97/1200-2420$12.50/0 Ñ 1997 Plenum Publishing Corporation

particular attention to the diaphragmatic crural com-
ponent.
MATERIALS AND METHODS
Adult male Wistar rats (Institute of Biomedical Re-search, Madrid, Spain) weighing 270 ± 350 g were fastedovernight, but were allowed free access to tap water. Ket-amine hydrochloride (62.5 mg/kg) and diazepam (5 mg/kg)were injected intraperitoneally to achieve anesthesia beforethe manometric studies were performed with the animals inthe supine position. A ® rst group of animals (N 5 119) wasstudied after anesthesia while in stable spontaneous respi-ration. Aiming at assessing the role of the diaphragmaticmuscle contractions on the manometric pro® les, a secondgroup of tracheostomized and intubated rats (N 5 14) wasinvestigated afte r muscle relaxation with atracurium besy-late (Tracrium) (5.5 mg/kg intraperitoneally). During relax-ation, which lasted 15± 20 minutes, intermittent positivepressure ventilation was administered with a rodent volu-metric respirator (Nemi scienti ® c, New England MedicalInstruments, Boston, Massachusetts) set to 85± 90 respira-tions per minute and tidal volume of 1 ml/100 g, whichallowed for adequate maintenance of both hemodynamicstability and gas exchanges. Animals in this group wereinvestigated manometrically with identical techniques in thebaseline status (afte r tracheostomy) while breathing spon-taneously, after relaxation (under IPPV), and again onspontaneous respiration afte r the effects of the relaxantdrug were over.
The techniques of catheter placement and manometricrecording were described in detail previously (6, 7). Brie¯ y,an open-tip catheter (1.2 mm OD, 0.5 mm ID) ® lled withheparinized saline was introduced through the femoral veininto the inferior vena cava for continuous recording of theintraabdominal pressure. A tip-occluded catheter (1.0 mmOD, 0.5 mm ID) with a distal side hole (1.0 3 0.5 mm)continuously perfused by a pneumohydraulic pump (MuiScienti® c, Mississauga, Ontario) at a rate of 0.4 ml/min waslocated in the mid-esophagus for measuring the intratho-racic pressure. After the resting pressures in both the thoraxand the abdomen were recorded, the esophageal catheterwas advanced into the stomach and subsequently it waspulled back through the esophagogastric junction into theesophagus with a purposefully-made mechanical device atconstant speed (1 mm/sec) to measure the pressures in thehigh-pressure zone. The values recorded are the average ofthree successive pull-through recordings. The atmosphericpressure at the leve l of the atrium served as zero reference.
We considered the following variables: (1) thoracic end-inspiratory and end-expiratory pressures; (2) abdominalend-inspiratory and end-expiratory pressures; (3) transdia-phragmatic end-inspiratory pressure gradient, ie, the differ-ence between abdominal and thoracic end-inspiratory pres-sures; (4) transdiaphragmatic end-expiratory pressuregradient, ie, the difference between abdominal and thoracicend-expiratory pressure; (5) lower esophageal sphincterpressure (LESP), (6) length of the intraabdominal segmentof the esophagus, and (7) crural sling pressure. Values aredescribed as means 6 standard deviations and the units aremillimeters and millimeters mercury. The Kolmogorov-
Smirnov test between the actual values and the ideal onesfor identical means and variances was used to assesswhether the distribution of values ® tted to a normal patte rn.In the second group of rats, comparisons among the varioussituations were done by repeated-measures analysis of vari-ance with Bonferroni-Dunn post hoc tests. Differences wereconsidered signi® cant at P , 0.05 or those required by thespeci ® c post hoc tests when appropriate.
Some animals were killed and dissected in order to de ® nethe anatomy of the crural sling and the lower esophagus.The experimental protocol was approved by the local insti-tutional research committee and met the regulations foranimal research in Europe (EC86/L609) .
RESULTS
The gross anatomy of the esophagogastric junction
in the rat is basically similar to that of the human
except for a cardial junction, which is located lower in
the lesser curvature leaving an ample fundus. The
hiatus opens almost centrally in the diaphragm and
the crura encircle the esophagus quite high as a
cle arly identi® able poste riorly opened, U-shape d
muscular structure (Figure 1) fused anteriorly and
late rally with the tendinous portion of the diaphragm,
which is in continuity with the muscular part inse rted
in the thoracic rim. There is a 10- to 20-mm infradia-
phragmatic segment of the esophagus, which appears
proportionally longer than its human counte rpart,
and no anatomical structure identi® able as the lower
esophage al sphincte r was appare nt on close inspe c-
tion and dissection.
Upon stationary manometry unde r spontaneous
respiration, the intrathoracic pressure measured at
the end of inspiration was clearly negative , whereas
the intraabdominal pressure recorded simultaneously
was slightly positive , revealing a positive transdia-
phragmatic pressure gradie nt. End-expiratory ab-
dominal and thoracic pressures remained positive
throughout the respiratory cycle (Table 1). Mechan-
ical ventilation under relaxation reversed thoracic
inspiratory pressures to the positive side and signi® -
cantly decreased the corresponding gradient. Tho-
racic expiratory pressure was also signi® cantly de-
creased. After recovery from relaxation and again
unde r spontaneous respiration, the value s returned to
the previously normal levels (Table 2).
When the cathe ter was withdrawn through the gas-
troesophageal junction, the ® rst (distal) high-pre ssure
zone corresponding to the LES was detected as a
well-de ® ned plateau or, more appropriate ly, as a wide
tonic peak. After this, the tracing returned to the
abdominal pressure leve l for some millimeters corre-
sponding to the infradiaphragm atic segment of the
CRURAL SPHINCTER IN THE RAT
2421Digestive Diseases and Sciences, Vol. 42, No. 12 (Decem ber 1997)

esophagus. Then, a second high-pre ssure zone was
constantly observed just prior to the exposure of the
measuring device to the in¯ uence of the thoracic
pressure. It consisted of a group of rapid phasic
oscillations of frequency identical to the respiratory
rate separated by short periods during which the
intraluminal pressure dropped until it reached ap-
proximate ly the intraabdominal expiratory pressure
leve l. However, this double plateau patte rn was not
constant in all rats, and three different types of pres-
sure pro® les could be de ® ned: in 100/119 rats (84.1% )
both the LESP and the phasic component were re-
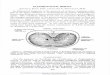
Fig 1. Abdominal side of the rat diaphragm and gastroesophage al region seen afterthe liver and small bowel have been removed. The U-shaped crural sling (CS) is
clearly visible. CR 5 costal rim, E 5 esophagus, IVC 5 inferior vena cava ori® ce andP 5 pylorus (ligated). The remarkable length of the intraabdominal segment of the
esophagus should be noted above the gastroesophage al junction (GEJ).
TABLE 1. PRESSURES, GRADIENTS, AND BARRIER
COMPONENTS IN 119 RATS*
Pressures and gradients (mm Hg)Thoracic inspiratory pressure 2 3.98 6 1.41
Abdominal inspiratory pressure 1.73 6 0.65Transdiaphragmatic gradient (insp.) 5.79 6 1.69
Thoracic expiratory pressure 1.61 6 1.06Abdominal expiratory pressure 1.09 6 0.72
Transdiaphragmatic gradient (exp.) 2 0.51 6 1.35Barrier components
Lower esophageal sphincter pressure (mm Hg) 14.76 6 8.63Crural sling pressure (mm Hg) 13.81 6 6.54
Intraabdominal esophage al length (mm) 13.78 6 3.13
*Values are means 6 SD.
SOTO ET AL
2422 Digestive Diseases and Sciences, Vol. 42, No. 12 (Decem ber 1997)

corded (Figure 2A), whereas in 10/119 rats (8.4% )
only the LESP could be measured (Figure 2B), and in
the remaining 9/119 rats (7.5% ) only the phasic com-
ponent appeared (Figure 2C). The mean pressure of
both components was twofold highe r than the trans-
diaphragmatic gradient throughout the respiratory
cycle (Table 1). Muscle relaxation and mechanical
ventilation temporarily abolishe d the crural sling
compone nt and signi® cantly shortened the intraab-
dominal esophagus. These effects disappe ared after
recovery from relaxation (Table 2 and Figure 3).
DISCUSSION
This study con® rms that rats, like humans, have a
positive transdiaphragm atic pressure gradie nt that is
more promine nt at the end of inspiration and that this
re¯ ux-driving force is permanently oppose d by a con-
siderably stronger tonic pressure barrie r located at
the gastroe sophage al junction and corresponding to
the LES. In addition, in the vast majority of animals,
there is another phasic high-pre ssure segment that we
attribute to the diaphragm atic crural contractions
during inspiration for the following reasons: (1) The
manometric distance between the extreme limits of
both pressure tracing components and the anatomic
separation of the cardia and the point where the
diaphragmat ic crura encircle the esophagus were
closely coincide nt. (2) These phasic de¯ ections ap-
pear about 10 ± 20 mm orad to the LESP, immediate ly
before the respiratory reversal point where the posi-
tive inspiratory waves corresponding to the abdome n
become negative . (3) The oscillating frequency of the
phasic waves is identical to the respiratory rate and
alte rnates with pressure drops to the basal intraab-
dominal leve l. (4) The disappe arance of this compo-
nent occurs with preservation of the LES plateau
unde r striated muscle relaxation. The fact that we
could not demonstrate the phasic component in 8.4%
of the rats studied can be explaine d on the basis of the
wide range of individual variations present in rats as
well as in man.
The crural sling compone nt of the barrier in the rat
is equivale nt to that previously described in human
subjects and in some animals. Boyle et al demon-
strated in the cat, by anchoring the pressure recording
device to the LES in order to avoid transhiatal axial
displacements caused by respiratory movements, that
the resting expiratory pressure accounted for by the
LES was maintaine d in apnea, whereas the increased
pressure oscillations accompanying inspiratory efforts
represented the effect of active diaphragmatic con-
tractions on the esophagus since they were abolishe d
by neuromuscular paralysis and were proportional to
the depth of respiratory excursions (8).
Mittal, in a manometric study in which the artifacts
produce d by axial displacement of the probe were
minimize d by using a sleeve pressure-measuring de-
vice , described two distinct pressure compone nts in
the human lower esophageal sphincte r: the ª smooth
muscle LESº and the crural diaphragm that he
termed the ª external lower esophage al sphincte rº (9).
This author acknowledged the dif® culties of measur-
ing effective ly the crural diaphragm pressures in hu-
mans by side-hole manome try and con® rmed that the
inspiratory oscillations are correlated with the depth of
inspiration and the force of diaphragmatic contraction.
Peck et al assessed manometrically the crural com-
ponent of the high-pre ssure zone in re¯ uxing individ-
uals without and with hiatal hernia before and after
antire ¯ ux operations including crural repair (10) .
They found that crural pressure was abolishe d in
patients with hiatal hernia and that crural repair
TABLE 2. PRESSURES, GRADIENTS, AND BARRIER COMPONENTS IN BASELINE CONDITIONS, AFTER
RELAXATION WITH MECHANICAL VENTILATION, AND AFTER RECOVERY IN 14 RATS*
Baseline
(spontaneou s)
Relaxation
(mech. ven t.)
Recovery
(spon taneou s)
Pressures and gradients (mm Hg)
Thoracic inspiratory pressure 2 3.55 6 1.83 3.57 6 1.82² 2 3.05 6 1.38Abdominal inspiratory pressure 2.19 6 1.2 2.81 6 1.3 2.23 6 0.89
Transdiaphragmatic gradient (insp.) 5.83 6 2.74 2 0.75 6 1.92² 5.3 6 1.24Thoracic expiratory pressure 2.3 6 1.38 0.84 6 0.72 2.87 6 1.69
Abdominal expiratory pressure 1.43 6 1.03 1.9 6 1.13 1.54 6 1.03Transdiaphragmatic gradient (exp.) 2 0.86 6 1.39 1.15 6 0.89² 2 1.33 6 1.51
Barrie r componentsLower esophageal sphincter pressure (mm Hg) 20.11 6 9.38 22.23 6 9.23 18.87 6 9.85
Crural sling pressure (mm Hg) 14.7 6 6.8 0² 10.82 6 4.14Intraabdominal esophagus length (mm) 17.57 6 1.78 6.83 6 1.08² 17.25 6 2.23
*Values are means 6 SD.
² Equivalent to P , 0.05 against baseline and recove ry.
CRURAL SPHINCTER IN THE RAT
2423Digestive Diseases and Sciences, Vol. 42, No. 12 (Decem ber 1997)

reestablishe d normal leve ls in both groups. Animal
studie s demonstrated that the crural diaphragm con-
tracts shortly before the costal component, leading to
an increase of the intrae sophage al pressure (11) . The
problems encounte red for identifying this crural com-
ponent of the anti-re ¯ ux barrier in human subje cts
are probably due to the fact that they have a relative ly
short intraabdominal esophagus and a relative ly long
LES (12) . The overlapping of both compone nts in the
tracings usually masks the participation of crural con-
tractions, which are often inte rpreted as respiratory
artifacts (this is the main reason for the routine use of
rapid pull-throughs when measuring the LESP).
Our observations in the rodent reveal a very pow-
erful gastroe sophage al barrier mechanism. Unfortu-
nately, we could not study other functional aspects
such as esophage al body peristalsis and the timing
and circumstance s of deglutory or spontaneous
sphincte r relaxations in this model because of the size
of the instrumentation necessary. Nevertheless, the
data provide d by the present investigation suggest
that the study of the behavior of the antire ¯ ux barrie r
unde r several challenging conditions can be under-
taken in this animal mode l in which both components
of the antire ¯ ux barrier are so neatly separated.
REFERENCES
1. Mittal RK, Rochester DF, McCallum RW: Sphincteric action
of the diaphragm during a relaxed lower esophage al sphincter
in humans. Am J Physiol 256:G139 ± G144, 1989
Fig 2. Pull-through (1 mm/sec) perfusion manometry tracings ofthe high-pressure zone in three adult normal rats showing the three
different patterns encountered: (A) In this animal there was aLESP peak or plateau followed by a long intraabdominal segme nt
of the esophagus before the phasic positive oscillations correspond-ing to the crural sling contractions appear. These oscillations
precede the negative inspiratory intrathoracic esophageal wavesthat reveal the respiratory inversion point (this pattern was present
in 100/119 rats or 84.1% ). (B) In this animal only the LESP pro® lecan be seen (pattern observed in 10/119 rats or 8.4% ), and (C) in
this animal only the crural oscillations are visible (pattern observedin 9/119 animals or 7.5% ). st 5 stomach; e 5 esophagus. Paper
speed 5 2.5 mm/se c.
Fig 3. The upper tracing corresponds to a baseline pull-throughrecording in a tracheostomized rat during spontaneous respiration.
Both tonic and phasic components are clearly seen. The lowertracing corre sponds to a pull-through recording under re laxation
and mechanical ventilation. The phasic or crural pressure compo-nent has been abolished, and no negative intrathoracic pressures
can be demonstrated.
SOTO ET AL
2424 Digestive Diseases and Sciences, Vol. 42, No. 12 (Decem ber 1997)

2. Mittal RK, Fisher MJ: Electrical and mechanical inhibition of
the crural diaphragm during transient re laxation of the lower
esophageal sphincter. Gastroenterology 99:1265± 1268, 1990
3. Mittal RK, Fisher M, McCallum RW, Rochester DF, Dent J,
Sluss J: Human lower esophageal sphincter pressure response
to incre ase d intra-abdominal pre ssure . Am J Physiol
258:G624 ± G630, 1990
4. Klein WA, Parkman HP, Dempsey DT, Fisher RS: Sphincter-
like thoracoabdominal high pressure zone after esophagogas-
trectomy. Gastroenterology 105:1362± 1369, 1993
5. Mittal RK: The crural diaphragm, an exte rnal lower esopha-
geal sphincter: A de® nitive study. Gastroenterology 105:1565±
1567, 1993
6. Wang WL, Tovar JA, Eizaguirre I, Aldazabal P: Airway obstruc-
tion and gastroesophageal re¯ uxÐ an experimental study on the
pathogenesis of this association. J Pediatr Surg 28:995± 998, 1993
7. Wang WL, Tovar JA, Eizaguirre I, Aldazabal P: Continuous
positive airway pressure and gastroesophage al re ¯ ux: An ex-
perimental study. J Pediatr Surg 29:730 ± 733, 1994
8. Boyle JT, Altschuler SM, Nixon TE, Tuchman DN, Pack AI,
Cohen S: Role of the diaphragm in the genesis of lower
esophageal sphincter pressure in the cat. Gastroente rology
88:724 ± 730, 1985
9. Mittal RK: Infusion manome try and detection of sphincteric
function of crural diaphragm. Dig Dis Sci 36( suppl):6 ± 13, 1991
10. Peck N, Callander N, Watson A: Manometric asse ssment of
the e ffect of the diaphragmat ic crural sling in gastro-
oesophageal re ¯ ux: Implications for surgical manageme nt. Br J
Surg 82:798 ± 801, 1995
11. DeTroye r A, Samson M, Sigrist S, Macklem P: Action of the
costal and crural parts of the diaphragm on the rib cage in dog.
J Appl Physiol 53:30 ± 39, 1982
12. HoÈ llwarth ME, Uray E, Pesendorfer P: Esophageal manome -
try. Pediatr Surg Int 1:177± 183, 1986
CRURAL SPHINCTER IN THE RAT
2425Digestive Diseases and Sciences, Vol. 42, No. 12 (Decem ber 1997)