Embed Size (px)
Citation preview

1521-0103/356/2/410–423$25.00 http://dx.doi.org/10.1124/jpet.115.229880THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS J Pharmacol Exp Ther 356:410–423, February 2016Copyright ª 2016 by The American Society for Pharmacology and Experimental Therapeutics
Identification of Phosphorylation Consensus Sequences andEndogenous Neuronal Substrates of the Psychiatric RiskKinase TNIK
Qi Wang,1 Stephen P. Amato,2 David M. Rubitski, Matthew M. Hayward,Bethany L. Kormos, Patrick R. Verhoest, Lan Xu,3 Nicholas J. Brandon,1,4
and Michael D. Ehlers4
Neuroscience & Pain Research Unit, BioTherapeutics Research and Development, Pfizer Inc. Cambridge, Massachusetts (Q.W.,S.P.A., D.M.R., N.J.B., M.D.E.); Center of Chemistry Innovation and Excellence, Pfizer Inc., Groton, Connecticut (M.M.H.);Neuroscience Medicinal Chemistry, Pfizer Inc., Cambridge, Massachusetts (B.L.K., P.R.V.);and Program in Molecular Medicine,University of Massachusetts Medical School, Worcester, Massachusetts (L.X.)
Received October 7, 2015; accepted December 1, 2015
ABSTRACTTraf2- and Nck-interacting kinase (TNIK) is a serine/threoninekinase highly expressed in the brain and enriched in the postsyn-aptic density of glutamatergic synapses in the mammalian brain.Accumulating genetic evidence and functional data have impli-cated TNIK as a risk factor for psychiatric disorders. However, theendogenous substrates of TNIK in neurons are unknown. Here, wedescribe a novel selective small molecule inhibitor of the TNIKkinase family. Using this inhibitor, we report the identification ofendogenous neuronal TNIK substrates by immunoprecipitationwith a phosphomotif antibody followed by mass spectrometry.Phosphorylation consensus sequences were defined by phos-phopeptide sequence analysis. Among the identified substrateswere members of the delta-catenin family including p120-catenin,
d-catenin, and armadillo repeat gene deleted in velo-cardio-facialsyndrome (ARVCF), each of which is linked to psychiatric orneurologic disorders. Using p120-catenin as a representativesubstrate, we show TNIK-induced p120-catenin phosphorylationin cells requires intact kinase activity and phosphorylation of TNIKat T181 and T187 in the activation loop. Addition of the smallmolecule TNIK inhibitor or knocking down TNIK by two shRNAsreducedendogenousp120-cateninphosphorylation incells. Together,using a TNIK inhibitor and phosphomotif antibody, we identifyendogenous substrates of TNIK in neurons, define consensussequences for TNIK, and suggest signaling pathways by whichTNIK influences synaptic development and function linked topsychiatric and neurologic disorders.
IntroductionTNIK is an abundant kinase highly expressed in the brain
and concentrated at the postsynaptic density of glutamatergicsynapses (Jordan et al., 2004; Peng et al., 2004; Collins et al.,2005; Burette et al., 2015). TNIK is highly enriched in synap-tosome and postsynaptic density fractions where it regulatesthe stability of postsynaptic density proteins (Nonaka et al.,2008; Hussain et al., 2010; Wang et al., 2011). Among itsvarious binding partners, TNIK interacts with disrupted inschizophrenia 1, a multifunctional scaffold protein whosealtered function is linked with psychiatric disorders includ-ing schizophrenia, depression, and bipolar disorder (Blackwood
et al., 2001; Camargo et al., 2007; Brandon and Sawa, 2011).Transcriptional profiling, genome-wide association studies,and functional genomics network analysis further support therole of TNIK as a psychiatric risk gene (Glatt et al., 2005;Matigian et al., 2007; Potkin et al., 2009; Shi et al., 2009;Ayalew et al., 2012).TNIK is a serine/threonine (Ser/Thr) kinase in the Ste20
family of MAP kinase kinase kinase kinases (MAP4K) (Fuet al., 1999). Although the cellular function of TNIK remainsunclear, one of the most reproducible effects of TNIK over-expression in mammalian cells is kinase-dependent inductionof cell rounding (Fu et al., 1999; Taira et al., 2004; Wang et al.,2011). TNIK is required for Wnt target gene induction incolorectal cancer cell lines and leukemia stem cells and thusregulates Wnt-induced proliferation (Mahmoudi et al., 2009;Shitashige et al., 2010; Schurch et al., 2012). In addition, asa downstream effector of the small GTPase Rap2A, TNIKregulates actin dynamics, cell morphology, and neuronal
1Current affiliation: Neuroscience iMED, AstraZeneca, Cambridge, MA2Current affiliation: TissueVision, Cambridge, MA3Current affiliation: Blueprint Medicines, Cambridge, MA4N.S.B. and M.D.E. contributed equally to this article.dx.doi.org/10.1124/jpet.115.229880.
ABBREVIATIONS: ARVCF, armadillo repeat gene deleted in velo-cardio-facial syndrome; DIV, days in vitro; EGFP, enhanced green fluorescentprotein; FMNL2, formin-like protein 2; HB, hydrogen bond; INPP4A, inositol polyphosphate-4-phosphatase type I; IP, immunoprecipitation;MAP4K4, mitogen-activated protein kinase kinase kinase kinase 4; MINK1, misshapen-like kinase 1; Msn, Misshapen; MTSS1L, metastasissuppressor 1-like; OA, okadaic acid; PBS, phosphate-buffered saline; pS/T, phosphoserine/threonine; RFP, red fluorescent protein; SMAD,mammalian homologs of SMA (C. elegans) and MAD (Drosophila); TNIK, Traf2- and Nck-interacting kinase; WT, wild type.
410
http://jpet.aspetjournals.org/content/suppl/2015/12/08/jpet.115.229880.DC1Supplemental material to this article can be found at:
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from
at A
SPET
Journals on Novem
ber 28, 2021jpet.aspetjournals.org
Dow
nloaded from
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from
at A
SPET
Journals on Novem
ber 28, 2021jpet.aspetjournals.org
Dow
nloaded from

structure (Taira et al., 2004; Kawabe et al., 2010; Gloerichet al., 2012). In Drosophila, the single TNIK family orthologMisshapen (Msn) negatively regulates the transforming growthfactor b pathway by phosphorylating an inhibitory site inthe mammalian homologs of the Drosophila protein mothersagainst decapentaplegic, and the C. elegans protein SMA(SMAD) transcription factors (Kaneko et al., 2011). Despitethese initial clues, the functions of TNIK in the brain are notknown, in large part due to the absence of known TNIKsubstrates in neurons.In vitro methods for the identification of kinase substrates
include utilizing peptide libraries to determine sequencespecificity and protein arrays to identify putative substrates(Obata et al., 2000; Hutti et al., 2004; Mok et al., 2009). Aquasi in vivo method is to incubate cell extracts with kinase ofinterest and [g-32P]ATP and identify 32P-labeled proteins asputative substrates (Cohen and Knebel, 2006). A modifiedversion of this method is to use unnatural ATP analogs andthe analog-sensitive kinase bearing kinase domainmutations(Koch and Hauf, 2010). Endogenous substrates may or maynot be identified by these methods because of differences withthe in vivo environments in these assays. Phosphorylationsite-specific antibodies have also been used successfully toidentify physiologic substrates of tyrosine and Ser/Thr ki-nases. However, the phosphorylation consensus sequencemust be known to generate suitable phosphorylation site-specific antibodies against a pool of mixed peptides matchingthe consensus sequence (Gronborg et al., 2002; Kane et al.,2002; Zhang et al., 2002). Therefore, substrate identificationfor novel Ser/Thr kinases without known consensus sequencesremains challenging.In the present study, using a novel specific small molecule
inhibitor of the TNIK family along with a phosphomotifantibody found to broadly recognize TNIK substrates, weidentified multiple physiologic substrates of TNIK in neu-rons and generalized three phosphorylation consensus se-quences, pT/S-L/I/V-D/E-x-x-x-K/R, pT/S-L/I/V-x-K/R, andpT/S-L-P/Q-L/I-x-x-K/R. The identified substrates includemammalian enabled, formin-like protein 2 (FMNL2) andmembers of the delta-catenin family, including p120-catenin,d-catenin, and ARVCF, supporting functions of TNIK in actinregulation, synaptic adhesion, and synaptic plasticity.
Materials and MethodsDNA Constructs and Antibodies. EGFP-TNIK-WT, EGFP-
TNIK-K54R, the empty vector HA, HA-TNIK-wild-type (WT), K54Rwere gifts fromKaibuchi Kariya (University of the Ryukyus). RFP andFlag-p120-catenin were purchased from Origene (Rockville, MD). HA-TNIK-T187A and T181A were generated with the QuickchangeLightning kit (Agilent Technologies, Santa Clara, CA). The scrambledand TNIK shRNAs in the pRFP-C-RS vector were purchased fromOrigene. The shRNA sequences are:
Scrambled: GCACTACCAGAGCTAACTCAGATAGTACTG;TNIK shRNA #1: ATACGAGACCAACCTAATGAGCGACAGGT;TNIK shRNA #2: ATGGCACCAGAAGTTATTGCCTGTGATGA.The HA (sc-805-G) antibody was from Santa Cruz Biotechnology
(Dallas, TX). The Flag antibody (M2) was from Sigma-Aldrich (St.Louis,MO). The pS (612547), phosphoserine/threonine (pS/T; 612548),and p120-catenin (610134) antibodies were from BD Biosciences(Franklin Lakes, NJ). The TNIK antibody was generated in rabbitsby an antigenic peptide located in the intermediate domain of TNIK toensure specificity: LLRQEQAKLNEARK. The pT187 antibody was
raised against the phosphorylated peptide NH2-GRRN(pT)FIGTPC-CONH2 in rabbits and purified by protein G and two steps ofpeptide-affinity columns with nonphosphorylated and phosphory-lated peptides. The pT312 antibody was raised against the peptideCNRNSpTIENTRRHIG in rabbits and affinity purified as describedpreviously (Kaneko et al., 2011).
Cell Line and Primary Neuronal Cultures. HEK293T andCOS7 cells were maintained in Dulbecco’s modified Eagle’s medium(Life Technologies, Carlsbad, CA) supplemented with 10% heat-inactivated fetal bovine serum (Life Technolgies). Rat hippocampaland cortical neuronal cultures were prepared from 18-day-old embry-onic rats as previously described (Wang et al., 2011). Hippocampi orcortices from both male and female rat embryos were pooled forneuronal culture preparations.
Chemical Synthesis and Analysis of PF-794. PF-794 wasdesigned via structure based drug design from previously reportedMAP4K inhibitors (Guimaraes et al., 2011). For the preparation ofPF-794, see Supplemental Data.
Kinase Selectivity Panels. PF-794 (1 mM) was screened in twokinase selectivity panelswith the Z-lyte assay (Life Technologies). TheIC50 of PF-794 for the kinase domain of TNIK and MAP4K4 wasdetermined by the Z-lyte assay performed at the ATP concentrationthat equals the ATP Km of each protein.
Computational Methods for Compound Docking. PF-794was docked into the TNIK X-ray crystal structure, PDB ID: 2X7Fafter structure preparation with the Protein Preparation Wizard inMaestro (version 9.7, Schrödinger, LLC, New York, NY) using defaultoptions. Ligand docking calculations were conducted using Glide SP(version 6.2, Schrödinger, LLC) (Friesner et al., 2004; Halgren et al.,2004). The pose was chosen based on the top-ranked poses andstructure activity relationship knowledge and rendered using thePyMOLMolecular Graphic System, Version 1.7.4.0 Schrödinger, LLC(Schrödinger, LLC).
Cell Rounding Assay. COS7 cells were seeded onto poly-L-lysinecoated 12 mm coverslips (BD Biosciences) and incubated overnight toreach about 50% confluency. Cells were transiently cotransfected withRFP and EGFP or EGFP-TNIK cDNA using lipofectamine2000 (LifeTechnologies) per the manufacturer’s suggested protocol. For exper-iments using TNIK wild-type and K54R kinase dead mutants, DMSOor PF-794 (10mM)was added to the indicated cell populations 15 hoursposttransfection and incubated for 2 hours before fixation. For cellrounding inhibition experiments, cells were transfected with RFP andEGFP-TNIK. Fifteen hours posttransfection, the cells were treatedwith PF-794 (1 mM) and fixed at the indicated time points. For PF-794washout experiments, cells were transfectedwithRFPandEGFP-TNIK.PF-794 (1 mM) was added 3 hours posttransfection and incubated foran additional 15 hours. After this incubation the original cell mediumcontaining PF-794 was removed, and the coverslips were washedthree times with prewarmed 1� phosphate-buffered saline (PBS).Fresh media was added to the well before fixation at the indicatedtime points. All cells were fixed using 4% paraformaldehyde/4%sucrose in 1� PBS. Coverslips were mounted onto microscope slidesusing prolong gold antifade mounting medium (Life Technologies)and kept in the dark at 4°C before imaging.
Images were acquired using a Zeiss (Oberkochen, Germany) wide-field microscope in combination with a 20� objective. Projected cellarea was determined by tracing the RFP signal of cells that werepositive for the expression of EGFP, EGFP-TNIK, or EGFP-TNIK-K54R and measuring the resulting area using ImageJ analysissoftware. The values for projected cell area were normalized to thecontrol COS7 cells expressing RFP and EGFP. Bar graphs representthree independent experiments with 30–40 cells being measured percondition for each trial. All values are reported as mean 6 S.E.M.
Immunoprecipitation. For preparation of cell extracts, rat hip-pocampal cultures or HEK293T cells grown in 6-well plates werewashed once with ice-cold PBS. Cells were then lysed in 120–150 ml ofco-IP buffer (50 mM Tris-Cl pH 7.5, 150 mM NaCl, 1 mM EDTA,1% Triton X-100) with freshly added protease and phosphatase
Substrates and Phosphorylation Consensus Sequences of TNIK 411
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from

inhibitor cocktails (Sigma-Aldrich). After rocking for 30 minutes at4°C, cell lysates were collected by scraping and cleared by centrifuga-tion at 10,000 g for 10 minutes at 4°C. Twenty microliters of proteinG-Sepharose4 beads (GE Healthcare, Little Chalfont, UK) equili-brated with co-IP buffer were mixed with 1–3 mg of antibodies. After1 hour of incubation, beads were washed one time with co-IP bufferand then mixed with cell or tissue lysates and incubated 2 hours toovernight at 4°C with rocking. The beads were washed extensivelywith co-IP buffer to remove unbound proteins, separated by NuPage4–12% Bis-Tris gels (Life Technologies), and subjected to immunoblotanalysis.
In Gel Digestion, TiO2 Phosphopeptide Enrichment, andLiquid Chromatography-Tandem Mass Spectrometry Analy-sis. For in gel protein digestion, proteins were isolated by 1- or2-dimensional polyacrylamide gel electrophoresis before in situenzymatic digestion. For TiO2 enrichment, the phosphopeptideswere enriched by TopTips (Glygen Corporation, Columbia, MD).For liquid chromatography-tandem mass spectrometry , an LTQOrbitrap equipped with a Waters nanoAcquity UPLC system wasused with aWaters Symmetry C18 180 mm� 20mm trap column anda 1.7 mm, 75 mm � 250 mm nanoAcquity UPLC column (35°C) forpeptide separation (Waters, Milford, MA). For database queries, alltandemmass spectrometry spectra were searched in-house using theMascot algorithm (version 2.2.0, Matrix Science, Boston, MA) foruninterpreted tandem mass sprectrometry spectra after using theMascot Distiller program to generate Mascot compatible files. Byusing the Mascot database search algorithm, a protein was consid-ered identified when Mascot lists it as significant and more than twopeptides match the same protein. The database searched is typicallythe NCBInr, which is chosen over genome specific databases becausea match to the correct species has more significance in the largerdatabases and, for some incomplete genomes, a match may be foundbased on homology to another species.
Peptides. Substrate peptides derived fromp120-catenin, d-catenin,and SMAD1 were synthesized at GenScript (Piscataway, NJ). Allpeptides used in the in vitro kinase assay were 20-mer peptides withthe TNIK phosphorylation site as the 10th residue. The peptides usedin this study are
SMAD1-TIEN(TRR): LLSNVNRNSTIENTRRHIGK;SMAD1-AIEN(TRR): LLSNVNRNSAIENTRRHIGK;SMAD1-TAEN(TRR): LLSNVNRNSTAENTRRHIGK;SMAD1-TIAN(TRR), LLSNVNRNSTIANTRRHIGK;SMAD1-TIEA(TRR): LLSNVNRNSTIEATRRHIGK;SMAD1-TIENTRA: LLSNVNRNSTIENTRAHIGK;p120-TLTR: DANPLMANGTLTRRHQNGRF;p120-TATR: DANPLMANGTATRRHQNGRF;p120-TLAR: DANPLMANGTLARRHQNGRF;p120-TLTA: DANPLMANGTLTARHQNGRF;p120-TLPL(IDR): QSSHSYDDSTLPLIDRNQKS;p120-TAPL(IDR): QSSHSYDDSTAPLIDRNQKS;p120-TLAL(IDR): QSSHSYDDSTLALIDRNQKS;p120-TLPA(IDR): QSSHSYDDSTLPAIDRNQKS;p120-TLPLIDA: QSSHSYDDSTLPLIDANQKS;d-TIER: QYHFVASSSTIERDRQRPYS;d-TAER: QYHFVASSSTAERDRQRPYS;d-TIAR: QYHFVASSSTIARDRQRPYS;d-TIEA: QYHFVASSSTIEADRQRPYS.ADP-Glo Kinase Assay. Ten nanomoles of purified TNIK kinase
domain (Carna Biosciences, Natick, MA) was incubated with 20-merpeptide of indicated concentrations in the kinase reaction buffer(50 mM Tris-Cl pH 7.5, 0.01% Triton X-100, 10 mM MgCl2, 50 mMATP) at 25°C for 40 minutes. The ADP-Glo assay (Promega, Madison,WI) was performed as per themanufacturer’s suggested protocol. Fourreplicates were performed for each condition, and the mean value ofADP formed in each condition was plotted as mean 6 S.E.M. Thesubstrate concentration and ADP formed was plotted in a XY graphand fitted with Michaelis-Menten equation to determine kineticconstants, Km, Vmax, and ratio of Vmax/Km.
Dot Blots. In vitro kinase assayswere performed as in theADP-Gloassay. Ten nanomoles of purified TNIK kinase domainwas incubatedwith 30 mM of the 20-mer peptide in the kinase reaction buffer. TheADP formed in the kinase reaction was determined by using theADP-Glo assay. Three microliters of each kinase reaction was dottedto a glutaraldehyde-activated nitrocellulose membrane, followed byimmunoblot with the pT312 antibody. The affinity of pT312 tophosphorylated peptides was determined by normalizing the dotblot signal to ADP formed in the kinase reaction. The affinity ofpT312 to the phosphorylated SMAD1-TIEN, which contained theantigenic sequence of pT312, was plotted as 100, and the affinity ofpT312 to other phosphopeptides was plotted as percentage of itsaffinity to SMAD1-TIEN. Four replicates were performed for eachcondition, and the mean value of ADP formed and pT312 affinity ineach condition was plotted as mean 6 S.E.M.
Statistical Analysis. All column graphs were plotted as mean 6S.E.M. Experiments with more than two conditions were analyzed byone-way analysis of variance test, followed by post hoc Tukey test tocompare the mean of each condition with the mean of every othercondition, or by post hoc Dunnett test to compare the mean of eachcondition with a control condition. Experiments with only twoconditions were analyzed by unpaired t tests.
ResultsA Tool Compound to Study TNIK Biology. Functional
studies of TNIK have been limited to overexpression andknockdown experiments because of the lack of selectivepharmacological tools. In mammalian genomes, two otherkinases, misshapen-like kinase 1 (MINK1) and mitogen-activated protein kinase kinase kinase kinase 4 (MAP4K4;also known as HGK), are closely related to TNIK and arepotential paralogs in mammals (Fu et al., 1999; Yao et al.,1999; Hu et al., 2004) (Fig. 1A). The kinase domains ofTNIK, MINK1, and MAP4K4 are highly similar with approx-imately 90% amino acid identity, and their ATP binding sitesare identical except for a single amino acid difference at aminoacid 113 (Xing et al., 2014). Because of this high ATP bindingsite similarity, discovery of a potent TNIK inhibitor that isselective over MINK1 and MAP4K4 has been a challenge.However, the discovery of compounds that are selective for theTNIK family over much of the kinome has been demonstratedand is hypothesized to be due to an unusual conformationadopted by the P-loop, also known as the glycine-rich loop inthe kinase domains of TNIK, MINK1, andMAP4K4 (Guimaraeset al., 2011).Using structure-based design from previously reported
MAP4K4 inhibitors (Guimaraes et al., 2011), we identifiedPF-794 as a potent, ATP-competitive TNIK inhibitor with acell free IC50 of 39 nM against the kinase domain (Fig. 1B).PF-794 was profiled in vitro in two kinase selectivity panelsthat contain representative kinases from different familiesthat span the human kinome and only displayed greaterthan 60% inhibition of TNIK, MINK1, andMAP4K4 at 1 mM,indicating excellent kinome selectivity (Fig. 1C). DockingPF-794 into the X-ray crystal structure of TNIK (PBD ID:2X7F) suggests that the aminopyridine core forms hydrogenbond (HB) interactions with the hinge residues E106 andC108 (Fig. 1D). The compound could potentially be furtherstabilized in the active site through interactions of thesubstituents with nearby residues, such as HBs with K54and V31, a CH-p HB with G111, and a p-p stacking in-teraction with Y36. The 2X7F TNIK X-ray crystal structure
412 Wang et al.
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from

adopts a folded P-loop in which Y36 forms a HB with Q157,which may help stabilize this conformation. As describedpreviously (Guimaraes et al., 2011), compounds that effi-ciently interact with residues in the hydrophobic tunnelformed by the folded P-loop conformation, especially withthe P5 residue (Y36 in TNIK) have a high selectivity againstthe kinome, suggesting a rational for the selectivity of PF-794for the TNIK family. Although PF-794 is not selective for TNIKover MINK1 and MAP4K4 (Fig. 1, C and E), it is uniquelyselective for the TNIK family and thus was chosen as a toolcompound to study TNIK biology and signaling.To test whether PF-794 can efficiently inhibit TNIK in cells,
we developed a cell rounding assay based on the observationthat TNIK induces cell rounding in a manner requiring intactkinase activity (Fu et al., 1999) (Fig. 2A). COS7 cells werecotransfected with red fluorescent protein (RFP) as a cell fillalong with enhanced green fluorescent protein (EGFP),EGFP-TNIK-WT, or EGFP-TNIK-K54R, a kinase dead mu-tant (Fu et al., 1999). As expected, EGFP-TNIK-WT inducedcell rounding as indicated by amore compact cell morphologycompared with the EGFP or EGFP-TNIK-K54R transfectedcells (Fig. 2, A and B). Addition of the TNIK family inhibitorPF-794 at 15 hours posttransfection (10 mM, 2 hours)inhibited TNIK-induced cell rounding and significantly in-creased the projected area of EGFP-TNIK-WT transfectedcells, indicating that PF-794 efficiently inhibits TNIK in cells(Fig. 2, A and B). Furthermore TNIK-induced cell roundingat 15 hours posttransfection was completely reversed by2 hours of PF-794 treatment (1 mM, 0.5 to 5 hours; Fig. 2, Cand D). When PF-794 (1 mM, 15 hours) was washed out fromcells expressing EGFP-TNIK-WT, the cells rounded up to a
minimal projected area within 2 hours (Fig. 2, E and F).Together, these experiments show that PF-794 inhibitsTNIK in cells. Moreover, TNIK-induced cell rounding andTNIK inhibition by PF-794 occur and can be reversed within2 hours.Detection and Isolation of TNIK Substrates in
Hippocampal Neurons. The protein phosphatase 1 andprotein phosphatase 2A inhibitor okadaic acid (OA) acti-vated TNIK in 293T cells (Kaneko et al., 2011), suggestingthat TNIK may be activated by phosphorylation. Activationof many kinases is triggered by phosphorylation in theactivation loop located between a highly conserved DFG(using the single-letter amino acid codes) motif and an APEmotif (Endicott et al., 2012) (Fig. 3A). In the activation loop ofTNIK, phosphorylated T187 has been identified by massspectrometry (Hornbeck et al., 2012) (Fig. 3A). We hypothe-sized that T187 phosphorylation may be enhanced by OA andrequired for activation of TNIK. To test this hypothesis, wegenerated a pT187 phosphorylation site-specific antibody.It specifically recognized HA-TNIK and HA-TNIK-K54R,but showed no signal with the phospho-blocking mutantHA-TNIK-T187A in transfected COS7 cells (Fig. 3B). As wehypothesized, upon incubation with OA (0.5 mM, 1 hour),T187 phosphorylation in TNIK was greatly increased in rathippocampal cultures (Fig. 3C). The observations that thekinase dead mutant HA-TNIK-K54R can be phosphorylatedat T187 and that PF-794 did not inhibit T187 phosphoryla-tion suggest that T187 is not an autophosphorylation siteand is phosphorylated by a different kinase.In Drosophila melanogaster, the single TNIK family
ortholog Msn was identified by a kinome RNAi screen as an
Fig. 1. PF-794 is a specific small mole-cule inhibitor of the TNIK family ofkinases. (A) Dendogram of the STE20kinase family. The TNIK family kinasesTNIK, MINK1, and MAP4K4 are labeledin the dendogram. Each of the otherkinases in the STE20 family is repre-sented by a short orange line. (B) PF-794is an ATP-competitive kinase inhibitorfrom an aminopyridine series. (C) Kinaseselectivity panel profile of PF-794. At1 mM, PF-794 is highly kinome-selectiveand specifically inhibited TNIK familykinases (MAP4K4, MINK1, and TNIK) inthe Z-lyte assay. (D) Docked structure ofPF-794 with TNIKATP binding site (PBDID: 2X7F). The amino acid residues in-volved in the compound interactions arelabeled. The flexible P-loop (highlightedin green) is a unique selectivity handle inthe TNIK family of kinases. (E) PF-794inhibits TNIK (cell free IC50 = 39 nM) andMAP4K4 (cell free IC50 = 40 nM) equallypotently. IC50 was determined by theZ-lyte assay performed at the ATP Km ofeach kinase.
Substrates and Phosphorylation Consensus Sequences of TNIK 413
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from

upstream kinase of MAD—the fly ortholog of the transforminggrowth factor b pathway effectors SMAD1, SMAD2, andSMAD3. Msn phosphorylates MAD at T312, and this phos-phorylation site is conserved in the mammalian orthologs,SMAD1 and SMAD2 (Kaneko et al., 2011). A phosphoantibodypT312 was generated against a phosphopeptide containing theT312 site of MAD andwas able to recognizeMAD, SMAD1, andSMAD2 phosphorylation by TNIK (Kaneko et al., 2011). As OAinduced TNIK activation loop phosphorylation in rat hippo-campal cultures (Fig. 3, A–C), we expected that immunoblotanalysis using pT312 would detect a band at ∼59 kDacorresponding to increased SMAD phosphorylation in neu-rons treated with OA. Surprisingly, the most prominentbands induced byOA and detected by the pT312 antibody hadhigher molecular masses .100 kDa (Fig. 3D). We reasonedthat the phosphorylation recognition motif present aroundT312 in MAD and recognized by the pT312 antibody may becommon to other yet unidentified TNIK substrates. Consis-tent with this idea, treatment of hippocampal neurons withthe selective TNIK family inhibitor PF-794 decreased theintensity and abundance of pT312 immunoreactive species inOA-treated hippocampal neurons (Fig. 3D). Furthermore,whereas OA increased the abundance of protein speciesimmunoreactive for broad spectrum phosphoserine (pS) and
phosphoserine/threonine (pS/T) antibodies, as well as spe-cies immunoreactive for the pT187 TNIK activation loopphosphoantibody, the abundance of pS, pS/T, and pT187species was not affected by PF-794 treatment (Fig. 3, D andE). Thus, pT312 recognizes multiple phosphoprotein speciesthat are increased by OA and decreased by TNIK inhibitorPF-794, features consistent with putative TNIK substrates.To isolate and identify potential TNIK substrates, immu-
noprecipitation was performed on OA-treated hippocampalneuron lysates using the pT312 antibody (Fig. 3F). Majorproteins immunoprecipitated were between 107 and 195 kDaand control immunoprecipitations were performed from un-treated neurons and neurons treated with both OA andPF-794 (Fig. 3F). Multiple protein species were identified inpT312 immunoprecipitates from OA-treated neuron lysatesthat were absent or reduced in untreated or OA1PF-794treated neurons (Fig. 3F). Gel sections above the antibodyheavy chain to ∼200 kDa in the pT312 IP lanes wereisolated and subjected to in-gel trypsin digestion followedby phosphopeptide enrichment and mass spectrometryanalysis (Fig. 3F). Putative TNIK substrates were definedby their presence in pT312 immunoprecipitates, increaseupon OA treatment, and reduction upon incubation withthe TNIK inhibitor PF-794.
Fig. 2. PF-794 inhibits TNIK in cells.(A) RFP images of COS7 cells cotrans-fected with RFP as a cell fill and EGFP,EGFP-TNIK, or EGFP-TNIK-K54R. Fif-teen hours posttransfection, the cellswere treated with 10 mM of PF-794 for2 hours. (B) Data indicate means 6 S.E.M. of the projected cell area from cellpopulations as in (A). Mean of eachcondition was compared with mean ofEGFP+DMSO. Expression of wild-typeTNIK resulted in a significant reductionin projected cell area that was reversedby treatment with the TNIK inhibitorPF-794. Cells expressing the kinase deadmutant K54R did not show a significantchange in shape whether in the presenceor absence of PF-794. (C) RFP images ofCOS7 cells cotransfected with RFP andEGFP-TNIK. Fifteen hours posttransfec-tion, the cells were treated with 1 mM ofPF-794 for the indicated time. (D) Dataindicate means6 S.E.M. of projected cellarea from cell populations shown in (C).Mean of each condition was comparedwith mean of 0 hour. Within 2 hours ofPF-794 treatment, projected cell areareturned to control levels in COS7 cellsexpressing EGFP. (E) RFP images ofCOS7 cells expressing RFP and EGFP-TNIK. Three hours posttransfection, thecells were treated with 1 mM of PF-794for 15 hours, followed by washout of theindicated time. (F) Data indicate means6S.E.M. of projected cell area from cellpopulations shown in (E). Mean of eachcondition was compared with mean of0 hour. Projected cell area returned tountreated levels after 1–2 hours of PF-794 washout in cell populations express-ing EGFP-TNIK. For (B), (D), and (F),bar graphs represent three independentexperiments with 30–40 cells being mea-sured per condition for each trial. *P #0.05; **P # 0.01; ****P # 0.0001.
414 Wang et al.
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from

Identification of TNIK Substrates and Phosphoryla-tion Consensus Sequences. Mass spectrometry of pT312immunoprecipitates identified multiple phosphopeptides fromhippocampal neuron lysates. We focused on phosphopeptidesthat were induced by OA and inhibited by PF-794 as potentialTNIK substrates (Table 1). Surprisingly, bioinformatic analysisrevealed three distinct consensus sequences shared bymultiplepeptides from different proteins—pT/S-L/I/V-D/E, pT/S-L/I/V-x-R/K, and pT/S-L-P/Q-L/I (Table 1). All phosphopeptides and
phosphomimetic peptides that match these consensus se-quences are listed in Table 1. Three lines of evidence supporta conclusion that these seemingly different sequences arephosphorylation consensus sequences of TNIK. First, pre-viously identified substrates of TNIK family kinases containthese sequences. For example, the Msn phosphorylation sitein MAD (pTIEN) and the corresponding phosphosites inSMAD1 (pTIEN) and SMAD2 (pTVEM), match the con-sensus pT/S-L/I/V-D/E (Kaneko et al., 2011) (Table 1). In
Fig. 3. Detection and isolation of TNIK substrates in hippocampal neurons. (A) Schematic representation of the protein structure of TNIK. The highlyconserved DFG and APE motifs define the activation loop of most kinases. Phosphorylation of T187 was identified by mass spectrometry (Hornbecket al., 2012). Amino acid numbers are labeled above. K, kinase domain. (B) COS7 cells were transfected with HA-TNIK, HA-TNIK-K54R, or HA-TNIK-T187A followed by immunoblot with the indicated antibodies. The pT187 phosphospecific antibody recognized HA-TNIK and HA-TNIK-K54R but notthe phospho-blocking mutant HA-TNIK-T187A. (C) 17 DIV rat hippocampal neurons were treated with OA (0.5 mM, 1 hour) with or without PF-794pretreatment (10 mM, 1 hour) followed by TNIK immunoprecipitation and immunoblot with the indicated antibodies. OA greatly enhanced T187phosphorylation of TNIK, which was not inhibited by PF-794. (D) Protein species detected by indicated phosphoantibodies in 17 DIV hippocampalneurons treated with OA (0.5 mM, 1 hour) with or without PF-794 pretreatment (10 mM, 1 hour). OA increased the abundance of protein speciesimmunoreactive for pT312, pS, pS/T and pT187, but PF-794 specifically decreased the abundance of pT312 immunoreactive protein species induced byOA. Immunoblot for GAPDHwas used as a loading control. (E) Line scan analysis of band intensity in (D). Band intensity was measured along a verticalline between ∼200 and 60 kDa. (F) Protein species isolated by the pT312 antibody (pT312 IP) from 17 DIV hippocampal neurons. Neurons were treatedwith OA (0.5 mM, 1 hour) with or without PF-794 pretreatment (10 mM, 1 hour) followed by immunoprecipitation and immunoblot with pT312. Gelsections were cut above the antibody heavy chain to ∼200 kDa in the pT312 IP lanes and subjected to in-gel trypsin digestion, phosphopeptideenrichment, and mass spectrometry analysis. In (B), (C), (D), and (F), molecular mass markers in kilodaltons are shown at the left.
Substrates and Phosphorylation Consensus Sequences of TNIK 415
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from

addition, Prickle 1 was identified as a substrate of MINK1,and the phosphosite (pTLSR) matches with the consensuspT/S-L/I/V-x-R/K (Daulat et al., 2012) (Table 1). BecauseTNIK, MINK1, and MAP4K4 have almost identical ATPbinding sites and activation loops (Figs. 1 and 3A), they arevery likely to share phosphorylation consensus sequences.Second, the pT312 antibody immunoprecipitated proteinscontaining each of the three consensus sequences, suggest-ing these phosphosites share similar spatial structuresdespite different primary sequences (Fig. 3F and Table 1).Third, the abundance of most of these phosphopeptidesacross the three different consensus sequences was in-creased by OA and reduced by PF-794, suggesting they aresubstrates of TNIK (Table 1). The abundance of thed-catenin peptide DGWSQYHFVASSSpTIER and MAP2peptide SDpTLQITDLLVPGSR was not reduced by PF-794(Table 1), but we included them as potential substratesbecause the PF-794 treatment was only 2 hours and thehalf-life of individual phosphorylation sites may varydepending on kinase and phosphatase activities.Further evidence that pT312 is a phosphomotif antibody
recognizing common structures shared by the three con-sensus sequences came from the observation that proteinswithout any of the three consensus sequences and nodetectable phosphoserine or phosphothreonine were alsoimmunoprecipitated by the pT312 antibody. However, all ofthese proteins contained at least one phosphomimetic pep-tide that matches one of the above consensus sequences
(Table 1; "phosphomimetic peptides"). As expected, theamount of most of the phosphomimetic peptides were neithersignificantly induced by OA nor inhibited by PF-794. To-gether these data provided strong evidence that pT312 is aphosphomotif antibody that recognizes common structures ofTNIK substrates. Moreover, these findings define canonicalconsensus sequences for TNIK family kinases.TNIK Phosphorylates Identified Consensus Se-
quences In Vitro. To test whether the identified pT/S-L/I/V-D/E, pT/S-L/I/V-x-R/K, and pT/S-L-P/Q-L/I consensussequences can be directly phosphorylated by TNIK, weperformed in vitro kinase reactions with purified TNIKkinase domain (amino acids 1–314) followed by a lumines-cent ADP-Glo assay (Promega) to quantify ADP generatedin kinase reactions. Twenty-mer peptides derived fromSMAD1, p120-catenin, and d-catenin were synthesized asrepresentatives of the three consensus sequences (Table 1and Fig. 4A). The d-TIER peptide matches both the pT/S-L/I/V-D/E and pT/S-L/I/V-x-R/K consensus sequences, andphosphorylation of this threonine was induced by OA butnot inhibited by 2 hours of PF-794 treatment (Table 1).Therefore we included it in our experiments to confirm it is asubstrate site for TNIK. Kinase reactions contained 50 mMATP. The relationship between the luminescent signal andADP concentration was linear between 0 and 100 mM (Fig.4B). SMAD1 is a confirmed TNIK substrate (Kaneko et al.,2011), and correspondingly the SMAD1-TIEN peptide wasphosphorylated by TNIK, whereas the alanine substitution
TABLE 1TNIK substrates and phosphorylation consensus sequencesThree consensus sequences were generalized from the phosphopeptides identified by mass spectrometry analysis.Phosphorylated residues are indicated in red.
Protein ID Peptide Identified DMSO OA OA+794
Consensus: pT/S - L/I/V - D/E Normalized AbundanceMENA GSTIETEQKEDRSEDAEPVTSK 0.01 1 0.0063FMNL2 SIEDLHR 0.002 1 0.037d-catenin SPSIDSIQK 0.69 1 0.043P120-catenin GSLASLDSLRK 0.0016 1 0.092MTSS1L GLSLEHQK 0.00022 1 0.61MAP2 VGSLDNAHHVPGGGNVK 0.0024 1 0.68d-catenin DGWSQYHFVASSSTIER 0.087 1 1.17
Consensus: pT/S - L/I/V - x - R/KFMNL2 DANTQVHTLRK 0.0043 1 0P120-catenin VSPQDANSLMANGTLTR 0.028 1 0.0056P120-catenin GSLASLDSLRK 0.0036 1 0.077P120-catenin HYEDGYPGGSDNYGSLSR 0.00074 1 0.1CamKII beta NFSVGR 0.00039 1 0.52d-catenin DGWSQYHFVASSSTIER 0.087 1 1.17
Consensus: pT/S - L - P/Q - L/IINPP4A KYAFNSLQLK 0 1 0P120-catenin SQSSHSYDDSTLPLIDR 0.0059 1 0.064ARVCF GTPNSGGFDDSTLPLVDK 0 1 0.16MAP2 SDTLQITDLLVPGSR 0.045 1 3.01
Phosphomimetic peptidesgrp75 YDDPEVQK 0.52 1 0.35MAPK8IP3 LYHAHTHQHLQDVDIEPYVSK 0.94 1 0.60MAPK8IP3 VDQLSGEQEVLKGELEAAK 0.19 1 0.61grp75 DDIENMVK 0.47 1 0.73MAPK8IP3 IKELEEELKR 0.27 1 0.77histone H2B LLLPGELAK 0.96 1 1.25MAP2 GEEEEIEAEGEYDK 0.02 1 2.63KRT6A YEELQITAGR 2.64 1 19.72
Previously published phosphosites KinaseMAD NRNSTIEN TNIK, MINK1, MAP4K4SMAD1 NRNSTIEN TNIK, MINK1, MAP4K4SMAD2 NRNATVEM TNIK, MINK1, MAP4K4Prickle1 ADDTLSR MINK1
416 Wang et al.
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from
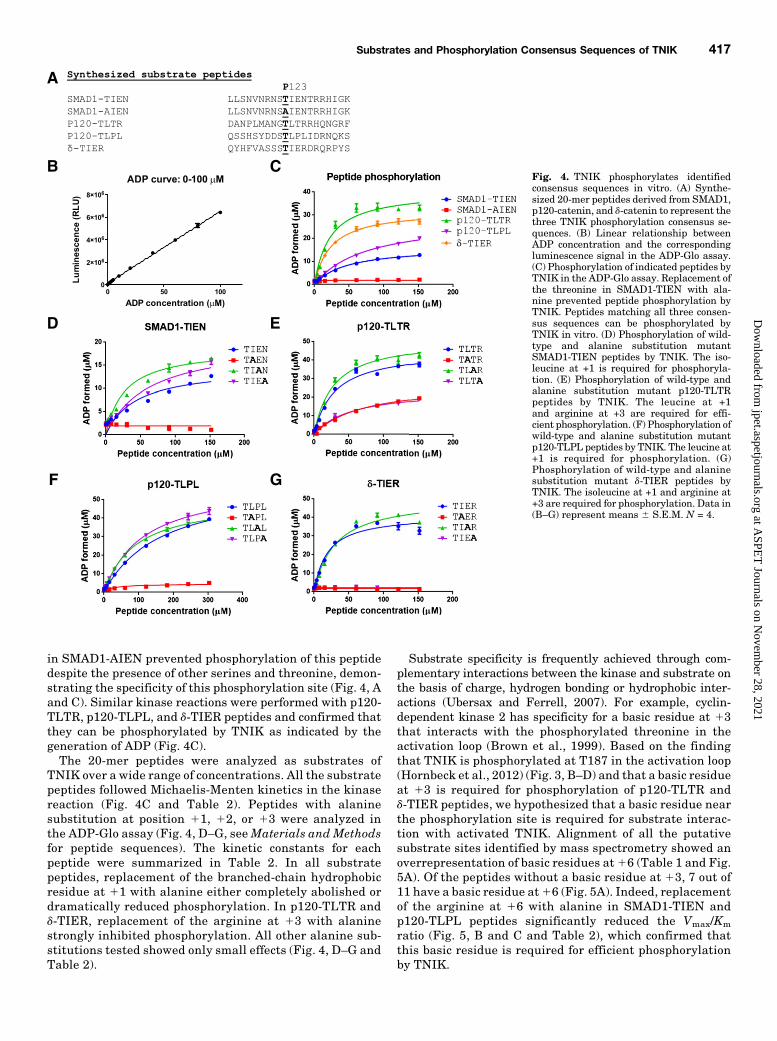
in SMAD1-AIEN prevented phosphorylation of this peptidedespite the presence of other serines and threonine, demon-strating the specificity of this phosphorylation site (Fig. 4, Aand C). Similar kinase reactions were performed with p120-TLTR, p120-TLPL, and d-TIER peptides and confirmed thatthey can be phosphorylated by TNIK as indicated by thegeneration of ADP (Fig. 4C).The 20-mer peptides were analyzed as substrates of
TNIK over a wide range of concentrations. All the substratepeptides followed Michaelis-Menten kinetics in the kinasereaction (Fig. 4C and Table 2). Peptides with alaninesubstitution at position 11, 12, or 13 were analyzed inthe ADP-Glo assay (Fig. 4, D–G, seeMaterials and Methodsfor peptide sequences). The kinetic constants for eachpeptide were summarized in Table 2. In all substratepeptides, replacement of the branched-chain hydrophobicresidue at 11 with alanine either completely abolished ordramatically reduced phosphorylation. In p120-TLTR andd-TIER, replacement of the arginine at 13 with alaninestrongly inhibited phosphorylation. All other alanine sub-stitutions tested showed only small effects (Fig. 4, D–G andTable 2).
Substrate specificity is frequently achieved through com-plementary interactions between the kinase and substrate onthe basis of charge, hydrogen bonding or hydrophobic inter-actions (Ubersax and Ferrell, 2007). For example, cyclin-dependent kinase 2 has specificity for a basic residue at 13that interacts with the phosphorylated threonine in theactivation loop (Brown et al., 1999). Based on the findingthat TNIK is phosphorylated at T187 in the activation loop(Hornbeck et al., 2012) (Fig. 3, B–D) and that a basic residueat 13 is required for phosphorylation of p120-TLTR andd-TIER peptides, we hypothesized that a basic residue nearthe phosphorylation site is required for substrate interac-tion with activated TNIK. Alignment of all the putativesubstrate sites identified by mass spectrometry showed anoverrepresentation of basic residues at16 (Table 1 and Fig.5A). Of the peptides without a basic residue at 13, 7 out of11 have a basic residue at16 (Fig. 5A). Indeed, replacementof the arginine at 16 with alanine in SMAD1-TIEN andp120-TLPL peptides significantly reduced the Vmax/Km
ratio (Fig. 5, B and C and Table 2), which confirmed thatthis basic residue is required for efficient phosphorylationby TNIK.
Fig. 4. TNIK phosphorylates identifiedconsensus sequences in vitro. (A) Synthe-sized 20-mer peptides derived from SMAD1,p120-catenin, and d-catenin to represent thethree TNIK phosphorylation consensus se-quences. (B) Linear relationship betweenADP concentration and the correspondingluminescence signal in the ADP-Glo assay.(C) Phosphorylation of indicated peptides byTNIK in the ADP-Glo assay. Replacement ofthe threonine in SMAD1-TIEN with ala-nine prevented peptide phosphorylation byTNIK. Peptides matching all three consen-sus sequences can be phosphorylated byTNIK in vitro. (D) Phosphorylation of wild-type and alanine substitution mutantSMAD1-TIEN peptides by TNIK. The iso-leucine at +1 is required for phosphoryla-tion. (E) Phosphorylation of wild-type andalanine substitution mutant p120-TLTRpeptides by TNIK. The leucine at +1and arginine at +3 are required for effi-cient phosphorylation. (F) Phosphorylation ofwild-type and alanine substitution mutantp120-TLPL peptides by TNIK. The leucine at+1 is required for phosphorylation. (G)Phosphorylation of wild-type and alaninesubstitution mutant d-TIER peptides byTNIK. The isoleucine at +1 and arginine at+3 are required for phosphorylation. Data in(B–G) represent means 6 S.E.M. N = 4.
Substrates and Phosphorylation Consensus Sequences of TNIK 417
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from

Taken together, these results demonstrate that the consen-sus sequences identified by mass spectrometry are bona fidesubstrates for TNIK in vitro. These data also show that abranched-chain hydrophobic residue at 11 is required for allthree consensus sequences to be phosphorylated by TNIK. Abasic residue at 13 or 16 is also required for efficientphosphorylation by TNIK. The nonessential residues in vitromay be required for optimized phosphorylation in vivo whenboth the kinase and substrate concentrations are low or are
positioned within a multiprotein complex. Based on the anal-ysis of alanine substitutions, we modified the consensussequences to pT/S-L/I/V-D/E-x-x-x-R/K, pT/S-L/I/V-x-R/K,and pT/S-L-P/Q-L/I-x-x-R/K.The pT312 Antibody Recognized Identified Consen-
sus Sequences In Vitro. To confirm that the TNIK sub-strate phosphorylation consensus sequences identified bymass spectrometry can be recognized by the pT312 antibody,30 mM of 20-mer peptides (Fig. 4A and see Materials and
TABLE 2Summary of kinetic constants of TNIK substrate peptidesAll peptides followed Michaelis-Menten kinetics in the ADP-Glo assay. Alanine substitutions are shown in bold in the leftcolumn.
20-mer peptide Km Vmax 10003Vmax/Km
mM mmol/min per mg
…TIENTRR… (SMAD1) 46.9 6 4.6 0.65 6 0.02 13.9…TAENTRR… ADP level too low to calculate kinetic constant…TIANTRR… 27.1 6 4.9 0.75 6 0.04 27.7…TIEATRR… 61.2 6 14.5 0.81 6 0.08 13.3…TIENTRA… 48.6 6 12.3 0.31 6 0.03 6.3
…TLTR… (p120-cat) 25.5 6 1.6 1.72 6 0.03 67.6…TATR… 71.8 6 9.4 1.13 6 0.07 15.8…TLAR… 27.5 6 2.0 2.08 6 0.05 75.5…TLTA… 54.3 6 7.1 0.99 6 0.05 18.2
…TLPLIDR… (p120-cat) 147.1 6 24.7 1.90 6 0.14 12.9…TAPLIDR… ADP level too low to calculate kinetic constant…TLALIDR… 90.7 6 4.5 2.04 6 0.04 22.4…TLPAIDR… 113.3 6 6.9 2.39 6 0.06 21.1…TLPLIDA… 231.5 6 36.6 1.10 6 0.09 4.7
…TIER… (d-cat) 20.4 6 2.3 1.47 6 0.05 72.1…TAER… ADP level too low to calculate kinetic constant…TIAR… 29.8 6 3.3 2.03 6 0.07 68.0…TIEA… ADP level too low to calculate kinetic constant
Fig. 5. A basic residue at +6 is required for efficient TNIKphosphorylation of substrate peptides without a basicresidue at +3. (A) Sequence alignment of all putative TNIKphosphorylation sites identified by mass spectrometry inTable 1. Overrepresentation of a basic residue at +6 wasidentified in phosphorylation sites without a basic residueat +3. (B) Phosphorylation of wild-type and R+6A SMAD1-TIEN peptides by TNIK. The arginine at +6 is required forefficient phosphorylation. (C) Phosphorylation of wild-typeand R+6A p120-TLPL peptides by TNIK. The arginine at+6 is required for efficient phosphorylation. Data in (B) and(C) represent means 6 S.E.M. N = 4.
418 Wang et al.
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from

Methods for peptide sequences) were incubated with purifiedTNIK kinase domain in the presence of ATP in an in vitrokinase assay, followed by dot blot with the pT312 antibody(Fig. 6A). The amount of ADP generated in the kinasereactions, which represented the amount of phosphorylatedpeptides, is shown in Fig. 6, B–E. The relative affinity of thepT312 antibody to each phosphorylated peptide was deter-mined by normalizing the dot blot signal to ADP formed ineach kinase reaction (Fig. 6, F–I).The pT312 antibody was raised against a 14-mer phosphor-
ylated peptide containing T312 of MAD (see Materials andMethods). These 14 amino acid residues are completelyconserved in SMAD1 (Kaneko et al., 2011). As expected, thepT312 antibody showed the highest affinity to phosphorylatedSMAD1-TIEN peptide, which we plotted as a normalizedvalue of 100 (Fig. 6, A, B, and F). The pT312 antibody showedpositive dot blot signals for p120-TLTR, p120-TLPL, andd-TIER peptides, which confirmed all three phosphorylatedconsensus sequences can be detected by pT312, although withlower affinities (Fig. 6, A and F–I). The pT312 antibody did notshow dot blot signals for SMAD1-TAENTRR, p120-TAPLIDR,d-TAER, or d-TIEA, consistent with their inefficient phosphor-ylation by TNIK, and confirming the specificity of pT312 tophosphorylated residues (Fig. 6). It is worth noting that p120-TLALIDR and p120-TLPAIDR showed equivalent phosphor-ylation by TNIK but their dot blot signals were dramatically
different, which ruled out the possibility that pT312 is anonspecific pT or even pT-L/I/V antibody (Fig. 6, A, D, and H).These data provide further evidence that phosphorylatedTNIK substrate sites share a common structure or epitopethat can be recognized by the pT312 phosphomotif antibody.Confirmation of p120-Catenin as an Endogenous
Substrate of TNIK. To confirm the identified substratescan be phosphorylated by TNIK in cells, we tested p120-catenin as a representative substrate. We coexpressed TNIKwith p120-catenin in HEK293T cells. Phosphorylation ofp120-catenin was monitored by immunoblot analysis usingthe pT312 phosphomotif antibody. In the absence of coex-pressed HA-TNIK, phosphorylation of p120-catenin was un-detectable (Fig. 7, A and B). Coexpression of HA-TNIKsignificantly increased pT312-detectable phosphorylationof p120-catenin. Whereas wild-type HA-TNIK increasedphosphorylation of p120-catenin, the kinase dead mutantHA-TNIK-K54R failed to increase their phosphorylation.Moreover, the activation loop phosphorylation site mutantsHA-TNIK-T187A and T181A only modestly increased phos-phorylation of p120-catenin, which confirmed that phos-phorylation of not only T187 but also T181 in the activationloop is required for full TNIK activation and phosphoryla-tion p120-catenin (Fig. 7, A and B). Acute treatment with theselective TNIK family inhibitor PF-794 (3 mM, 2 hours)inhibited TNIK-induced phosphorylation of p120-catenin
Fig. 6. The pT312 antibody recognized identified con-sensus sequences in vitro. (A) pT312 dot blots ofphosphorylated peptides. The indicated 20-mer peptideswere phosphorylated by TNIK in the in vitro kinaseassay followed by dot blot with the pT312 antibody.pT312 recognized phosphorylated peptides matching allthree consensus sequences. (B–E) The amount of TNIKphosphorylated SMAD1-TIEN (B), p120-TLTR (C),p120-TLPL (D), and d-TIER (E) peptides is indicatedby the amount of ADP formed in the ADP-Glo assay.(F–I) The relative affinity of the pT312 antibody to TNIKphosphorylated SMAD-TIEN (F), p120-TLTR (G), p120-TLPL (H), and d-TIER (I) peptides is represented bynormalizing the dot blot signal in (A) to ADP formed in(B–E) for each peptide. The affinity of pT312 tophosphorylated wild-type SMAD1-TIEN containing theantigenic sequence of pT312 was plotted as 100. Theaffinity of pT312 to other peptides was plotted aspercentage of the affinity to wildtype SMAD1-TIEN.Data in (B–I) indicate means 6 S.E.M. Mean of eachmutant peptide was compared with mean of thecorresponding wild-type peptide. N = 4. **P # 0.01;***P # 0.001; ****P # 0.0001.
Substrates and Phosphorylation Consensus Sequences of TNIK 419
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from

(Fig. 7, A and B). Together these data show that TNIKmediates phosphorylation of p120-catenin in cells.To test whether p120-catenin is phosphorylated by TNIK in
neurons, 15–21 days in vitro (DIV) hippocampal cultures weretreated with vehicle or the selective TNIK family inhibitor PF-794 (3 mM, 2 hours). TNIK substrates were isolated fromneuron lysates by pT312 immunoprecipitation before immuno-blot analysis for p120-catenin (Fig. 7C). p120-Catenin waspresent in pT312 immunoprecipitates, and the amount ofpT312-immunoprecipitated p120-catenin was significantly re-duced by PF-794 (Fig. 7, C and D). These results are consistentwith ourmass spectrometric and in vitro data identifying p120-catenin as endogenous TNIK substrates in neurons.To further confirm substrate specificity, two different TNIK
shRNAs were transfected into HEK293T cells and producedabout 50% reduction in the protein level of endogenous TNIKprotein levels (Fig. 7, E and F). p120-Catenin immunoprecip-itation followed by immunoblot with the pT312 antibodyshowed TNIK knockdown reduced endogenous p120-cateninphosphorylation (Fig. 7, E–G). The remaining phosphorylatedp120-catenin was likely due to the incomplete knockdown ofTNIK and could be further reduced by addition of PF-794(Fig. 7, E–G). These data demonstrate p120-catenin is anendogenous substrate of TNIK.
DiscussionPhosphorylation Consensus Sequences for TNIK
Family Kinases. In the present study, although we werelooking at SMAD phosphorylation in neurons, we discoveredthat the pT312 phosphorylation site-specific antibody origi-nally raised against a single phosphorylated MAD peptide wasbroadly reactive to and capable of immunoisolating multipleTNIK substrates (Fig. 3). Mass spectrometric analysis of pT312immunoprecipitates identified multiple phosphopeptides fromwhich we generalized three phosphorylation consensus se-quences for the TNIK kinase family, pT/S-L/I/V-D/E-x-x-x-R/K, pT/S-L/I/V-x-R/K, and pT/S-L-P/Q-L/I-x-x-R/K (Table 1,Figs. 4 and 5).Several lines of evidence support the assignment of pT/S-L/
I/V-D/E-x-x-x-R/K, pT/S-L/I/V-x-R/K, and pT/S-L-P/Q-L/I-x-x-R/K as TNIK phosphorylation consensus sequences. First,all proteins identified here were recognized and immunopre-cipitated by the pT312 antibody, which was originally raisedagainst epitopes corresponding to the Msn phosphorylationsite on MAD (Kaneko et al., 2011) (Fig. 3). Second, all proteinsidentified, without exception, had at least one phosphopep-tide or one phosphomimetic peptide that matched one ofthe consensus sequences (Table 1). Third, the previouslyidentified TNIK family substrates, SMAD1 and SMAD2,
Fig. 7. Confirmation of p120-catenin as an endogenoussubstrate of TNIK. (A) HEK293T cells were cotrans-fected with the indicated HA-TNIK constructs andFlag-p120-catenin followed by immunoblot analysiswith the indicated antibodies. Immunoreactivity forpT312 indicates phosphorylation by TNIK. Wild-typeHA-TNIK induced p120-catenin phosphorylation,which was inhibited by PF-794 (3mM, 2 hours).Expression of the kinase dead mutant HA-TNIK-K54R and the activation loop phosphorylation sitemutants TNIK-T187A and TNIK-T181A resulted inno or weak induction of p120-catenin phosphorylation.(B) Quantification of immunoblots from (A). Datarepresent means 6 S.E.M. of p120-catenin phosphory-lation measured by the ratio of band intensity detectedby the pT312 antibody to total p120-catenin signaldetected by the anti-Flag antibody. N = 3. Mean of eachcondition was compared with mean of every othercondition and all significant pairs were shown. (C) PF-794 inhibited endogenous p120-catenin phosphoryla-tion in hippocampal neurons. 15–21 DIV hippocampalcultures were treated with PF-794 (3 mM, 2 hours)followed by immunoprecipitation with pT312 andimmunoblot with the antibody against p120-catenin.(D) Quantitation of immunoblots from (C). Graph showsmeans6 S.E.M. of phosphorylated p120-catenin.N = 3.(E) HEK293T cells were transfected with a scrambledcontrol or one of two distinct TNIK shRNAs, followed byimmunoprecipitation and immunoblot with the in-dicated antibodies. (F) Quantification of immunoblotsfrom (E). Data represent means 6 S.E.M. Mean of eachcondition was compared with mean of Scrm+DMSO.Expression of either TNIK shRNA sequence resulted ina significant knockdown of endogenous TNIK proteinexpression as compared with the scrambled control. (G)Quantification of immunoblots from (E). Data representmeans 6 S.E.M. Mean of each condition was comparedwith mean of Scrm+DMSO. Knockdown of TNIKresulted in a decrease in p120-catenin phosphorylationthat was further reduced via PF-794 treatment (3 mM,2 hours). *P # 0.05; ***P # 0.001; ****P # 0.0001.
420 Wang et al.
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from

contain the consensus pT/S-L/I/V-D/E-x-x-x-R/K, whereasPrickle 1, a separate TNIK family substrate contains theconsensus pT/S-L/I/V-x-R/K (Kaneko et al., 2011; Daulatet al., 2012) (Table 1). Fourth, the abundance of mostphosphopeptides was increased by two to four orders ofmagnitude upon okadaic acid treatment and reduced signif-icantly by the selective TNIK inhibitor PF-794. The in-creased abundance of substrate phosphopeptides with OAtreatment was consistent with the approximately threeorders of magnitude induction of kinase catalytic activityby the activation loop phosphorylation (Adams, 2001) (Fig. 3and Table 1). Fifth, representative peptides that match eachof the three consensus sequences can be phosphorylated byTNIK in vitro and recognized by the pT312 antibody (Fig.4–6). Sixth, alanine substitutions in ADP-Glo assay demon-strated the branched-chain hydrophobic residue at 11 andbasic residue at 13 or 16 are required for efficient TNIKphosphorylation (Figs. 4 and 5 and Table 2). We thus con-clude that TNIK recognizes and phosphorylates the consen-sus sequences pT/S-L/I/V-D/E-x-x-x-R/K, pT/S-L/I/V-x-R/K,and pT/S-L-P/Q-L/I-x-x-R/K. One attractive hypothesis isthat the basic residue at 13 or 16 in substrates interactswith phosphorylated T181 and/or T187 in the activation loopof TNIK family kinases, a hypothesis that can be tested infuture studies by cocrystallization of TNIK with substratepeptides.Substrate Specificity of TNIK, MINK1, and MAP4K4.
TheN-terminal kinase domains of TNIK, MINK1, andMAP4K4are highly similar (Fig. 1). Key residues in the kinase domaininvolved in active site interactionswithATP are 96% similar andthe activation loops between DFG and APE motifs are identical,suggesting shared phosphorylation consensus sequences (Hanksand Hunter, 1995). Consistent with the sequence similarity,all three kinases phosphorylate SMAD1 when both the kinaseand substrate are expressed in heterologous cells (Kanekoet al., 2011).How might substrate specificity be achieved between these
kinases? First, substrate may interact differentially withindividual kinases. Prickle 1 interacts with MINK1 but notwith TNIK and MAP4K4, allowing it to be a specific MINK1substrate (Daulat et al., 2012). Second, TNIK family membersmaybe be differentially expressed in different cells andtissues. Third, the intermediate domains (amino acid residues
from ∼300 to 1,000) of TNIK, MINK1, and MAP4K4 are lesssimilar than the highly conserved N-terminal kinase and C-terminal CNH domains and may confer specific subcellulartargeting or substrate recognition. Using TNIK shRNAs weshow that endogenous p120-catenin is phosphorylated byendogenous TNIK in cells (Fig. 7). Whether individual sub-strates are phosphorylated byMINK1 orMAP4K4will requirefuture studies with shRNAs or inhibitors that are specificwithin TNIK family kinases.TNIK-Induced Cell Rounding. One of the most repro-
ducible effects of active TNIK is cell rounding (Fu et al., 1999;Taira et al., 2004; Wang et al., 2011) (Fig. 2). A few substratesidentified here could potentially mediate this effect. Byinteracting with the barbed ends of actin filaments andenhances the rate of F-actin polymerization, mammalianenabled regulates cell movement and shape (Gertler andCondeelis, 2011). FMNL2 is a member of the formin family,which is characterized by the ability to polymerize straightactin filaments at the barbed end (Kuhn and Geyer, 2014).FMNL2 is one of the effector proteins of small GTPases, suchas Cdc42, and was shown to regulate actin dynamics and cellmorphology (Kuhn and Geyer, 2014). In addition to function-ing as an important regulator of classic cadherins, p120 is amain regulator of RhoGTPase and thus has a prominent effecton actin dynamics and cell morphology (Schackmann et al.,2013). Whether phosphorylation by TNIK alters the ability ofthese substrates to regulate actin dynamics will requirefurther studies with phosphoblocking and phosphomimeticmutants.Neuronal Substrates of TNIK. Among the 16 phospho-
peptides identified in pT312 immunoprecipitates by massspectrometry, eight phosphopeptides corresponded to se-quences within p120-catenin, d-catenin, and ARVCF, threemembers of the delta-catenin family of armadillo-repeatproteins involved in cell adhesion and intracellular signal-ing (Table 1) (Pieters et al., 2012). Delta-catenin familyproteins are also part of the classic cadherin adhesioncomplexes at synapses where they are required for synapto-genesis, spine growth, and synaptic plasticity (Kosik et al.,2005; Elia et al., 2006; Arikkath et al., 2009; Matter et al.,2009). The fact that multiple phosphorylation sites of threedelta-catenins were identified suggests their phosphoryla-tion by TNIK is of particular functional importance.
Fig. 8. The neuronal substrates of TNIK suggest its role insynaptic development and function linked to psychiatricand neurologic disorders. Many of TNIK substrates aregenetically linked to psychiatric and neurologic disorders.TNIK could regulate synaptic adhesion, synaptic actindynamics, and synaptic signaling by phosphorylating thesesubstrates.
Substrates and Phosphorylation Consensus Sequences of TNIK 421
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from

It is worth noting that almost all of the TNIK substratesidentified in this study (Table 1) are genetically linked topsychiatric or neurologic disorders. Exome sequencing ofsporadic autism patients identified de novo deletion of p120-catenin in an autism patient (O’Roak et al., 2012). The recentreport from the Psychiatric Genomics Consortium on thelargest schizophrenia genome-wide association studies to dateshowed that p120-catenin gene is located in one of the108 genome-wide significant loci (Schizophrenia WorkingGroup of the Psychiatric Genomics, 2014). d-Catenin is one ofthe essential genes deleted in the neurodevelopmental cri duchat syndrome, whose symptoms include severe cognitiveand language impairments, motor delays, and behavioralproblems (Medina et al., 2000). ARVCF is one of the essentialgenes deleted in the 22q11.2 microdeletion syndrome (Sanderset al., 2005), in which the prevalence of schizophrenia and otherpsychotic disorders is as high as 30–40% (Karayiorgou et al.,2010; Schneider et al., 2014). A recessive mutation of metasta-sis suppressor 1-like (MTSS1L) caused neurodegeneration in aconsanguineous family (Alazami et al., 2015). FMNL2 was oneof the essential genes deleted in a severely mentally retardedpatient (Lybaek et al., 2009). Frameshift mutations of inositolpolyphosphate-4-phosphatase type I (INPP4A) caused neuro-nal loss and seizures in both a patient and the weeble mutantmouse line that arose spontaneously (Nystuen et al., 2001;Sheffer et al., 2015).We identified delta-catenin family proteins, actinmodulators,
and signaling molecule as neuronal substrates of TNIK (Fig. 8).Because these substrates have been shown to be important forsynaptic development and function linked to psychiatric andneurologic disorders, TNIK could modulate synaptic adhesion,synaptic actin dynamics, and synaptic signaling by phosphory-lating these substrates (Fig. 8). It will be important for futurestudies to confirmeach of the additional putative substrates andunderstand the consequences of TNIK phosphorylation.A Phosphomotif Antibody to Identify Endogenous
Kinase Substrates. In this study, we found that the pT312phosphoantibody was broadly reactive to multiple TNIKsubstrates in neurons (Fig. 3). Although pT312 was raisedagainst a single phosphopeptide, this antibody was able torecognize phosphopeptides with very different primary se-quences, thus enabling the identification of three substrateconsensus sequences for TNIK family kinases (Table 1). Thestructural basis for pT312 epitope recognition is not known.Alanine substitutions in dot blot assay suggest the branched-chain hydrophobic residue at11, the acidic residue at12, andthe branched-chain hydrophobic residue at13 are involved inthe interaction of the phosphopeptides with pT312 (Fig. 6).Differential affinities of pT312 with similarly phosphorylatedp120-TLALIDR and p120-TLPAIDR demonstrated thatpT312 is not just a broadly reactive pT antibody (Fig. 6, A,D, and H). In particular, it may be that different immunoglob-ulin species within the pT312 polyclonal antisera possessdistinct phosphoepitope recognition. Alternatively, individualimmunoglobulins may recognize multiple phosphoepitopeswith varying affinities. In either case, the implication is thatphosphoepitopes with varying primary sequences can shareepitope recognition features. An understanding of the precisemolecular basis of phosphorylation site recognition awaitscocrystallization of the TNIK kinase domain with substratepeptides from each of the three different consensus sequenceclasses (Table 1).
Prevailing methods using antibodies to identify Ser/Thrkinase substrates have generally required knowledge ofphosphorylation consensus sequences (Kane et al., 2002;Zhang et al., 2002). Antibodies were generated using a poolof mixed peptides that match the consensus sequence. Byusing this method, novel substrates were successfully iden-tified for Akt (Kane et al., 2002). Nonetheless, identificationof initial substrates and generalization of phosphorylationconsensus sequences remain one of the biggest challenges inkinase biology. Here we described a proteomics-based ap-proach whereby endogenous TNIK substrates were identi-fied by a phosphomotif antibody generated against a singlesubstrate (MAD), a specific kinase inhibitor (PF-794), and abroad-acting kinase activator (OA). This relative simplemethod may be applicable to other kinases.
Acknowledgments
The authors thank Dr. Kaibuchi Kariya for his generous gifts ofTNIK-WT and K54R constructs and Veronica Reinhart for technicalassistance.
Authorship Contributions
Participated in research design: Wang, Hayward, Brandon, andEhlers
Conducted experiments: Wang, Amato, and Rubitski,Contributed new reagents or analytic tools: Wang, Hayward,
Verhoest, and XuPerformed data analysis: Wang, Amato, Rubitsk, and KormosWrote or contributed to the writing of the manuscript: Wang,
Amato, Hayward, Kormos, Verhoest, Brandon, and Ehlers.
References
Adams JA (2001) Kinetic and catalytic mechanisms of protein kinases. Chem Rev101:2271–2290.
Alazami AM, Patel N, Shamseldin HE, Anazi S, Al-Dosari MS, Alzahrani F, Hijazi H,Alshammari M, Aldahmesh MA, and Salih MA, et al. (2015) Accelerating novelcandidate gene discovery in neurogenetic disorders via whole-exome sequencing ofprescreened multiplex consanguineous families. Cell Reports 10:148–161.
Arikkath J, Peng IF, Ng YG, Israely I, Liu X, Ullian EM, and Reichardt LF (2009)Delta-catenin regulates spine and synapse morphogenesis and function in hippo-campal neurons during development. J Neurosci 29:5435–5442.
Ayalew M, Le-Niculescu H, Levey DF, Jain N, Changala B, Patel SD, Winiger E,Breier A, Shekhar A, and Amdur R, et al. (2012) Convergent functional genomics ofschizophrenia: from comprehensive understanding to genetic risk prediction. MolPsychiatry 17:887–905.
Blackwood DH, Fordyce A, Walker MT, St Clair DM, Porteous DJ, and Muir WJ(2001) Schizophrenia and affective disorders–cosegregation with a translocation atchromosome 1q42 that directly disrupts brain-expressed genes: clinical and P300findings in a family. Am J Hum Genet 69:428–433.
Brandon NJ and Sawa A (2011) Linking neurodevelopmental and synaptic theories ofmental illness through DISC1. Nat Rev Neurosci 12:707–722.
Brown NR, Noble ME, Endicott JA, and Johnson LN (1999) The structural basis forspecificity of substrate and recruitment peptides for cyclin-dependent kinases. NatCell Biol 1:438–443.
Burette AC, Phend KD, Burette S, Lin Q, Liang M, Foltz G, Taylor N, Wang Q,Brandon NJ, and Bates B, et al. (2015) Organization of TNIK in dendritic spines.J Comp Neurol 523:1913–1924.
Camargo LM, Collura V, Rain JC, Mizuguchi K, Hermjakob H, Kerrien S, BonnertTP, Whiting PJ, and Brandon NJ (2007) Disrupted in Schizophrenia 1 Interactome:evidence for the close connectivity of risk genes and a potential synaptic basis forschizophrenia. Mol Psychiatry 12:74–86.
Cohen P and Knebel A (2006) KESTREL: a powerful method for identifying thephysiological substrates of protein kinases. Biochem J 393:1–6.
Collins MO, Yu L, Coba MP, Husi H, Campuzano I, Blackstock WP, Choudhary JS,and Grant SG (2005) Proteomic analysis of in vivo phosphorylated synaptic pro-teins. J Biol Chem 280:5972–5982.
Daulat AM, Luu O, Sing A, Zhang L, Wrana JL, McNeill H, Winklbauer R,and Angers S (2012) Mink1 regulates b-catenin-independent Wnt signaling viaPrickle phosphorylation. Mol Cell Biol 32:173–185.
Elia LP, Yamamoto M, Zang K, and Reichardt LF (2006) p120 catenin regulatesdendritic spine and synapse development through Rho-family GTPases andcadherins. Neuron 51:43–56.
Endicott JA, Noble ME, and Johnson LN (2012) The structural basis for control ofeukaryotic protein kinases. Annu Rev Biochem 81:587–613.
Friesner RA, Banks JL, Murphy RB, Halgren TA, Klicic JJ, Mainz DT, Repasky MP,Knoll EH, Shelley M, and Perry JK, et al. (2004) Glide: a new approach for rapid,accurate docking and scoring. 1. Method and assessment of docking accuracy.J Med Chem 47:1739–1749.
422 Wang et al.
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from

Fu CA, Shen M, Huang BC, Lasaga J, Payan DG, and Luo Y (1999) TNIK, a novelmember of the germinal center kinase family that activates the c-JunN-terminal kinase pathway and regulates the cytoskeleton. J Biol Chem 274:30729–30737.
Gertler F and Condeelis J (2011) Metastasis: tumor cells becoming MENAcing.Trends Cell Biol 21:81–90.
Glatt SJ, Everall IP, Kremen WS, Corbeil J, Sásik R, Khanlou N, Han M, Liew CC,and Tsuang MT (2005) Comparative gene expression analysis of blood and brainprovides concurrent validation of SELENBP1 up-regulation in schizophrenia. ProcNatl Acad Sci USA 102:15533–15538.
Gloerich M, ten Klooster JP, Vliem MJ, Koorman T, Zwartkruis FJ, Clevers H,and Bos JL (2012) Rap2A links intestinal cell polarity to brush border formation.Nat Cell Biol 14:793–801.
Grønborg M, Kristiansen TZ, Stensballe A, Andersen JS, Ohara O, Mann M, JensenON, and Pandey A (2002) A mass spectrometry-based proteomic approach foridentification of serine/threonine-phosphorylated proteins by enrichment withphospho-specific antibodies: identification of a novel protein, Frigg, as a proteinkinase A substrate. Mol Cell Proteomics 1:517–527.
Guimarães CR, Rai BK, Munchhof MJ, Liu S, Wang J, Bhattacharya SK,and Buckbinder L (2011) Understanding the impact of the P-loop conformation onkinase selectivity. J Chem Inf Model 51:1199–1204.
Halgren TA, Murphy RB, Friesner RA, Beard HS, Frye LL, Pollard WT, and BanksJL (2004) Glide: a new approach for rapid, accurate docking and scoring. 2. En-richment factors in database screening. J Med Chem 47:1750–1759.
Hanks SK and Hunter T (1995) Protein kinases 6. The eukaryotic protein kinasesuperfamily: kinase (catalytic) domain structure and classification. FASEB J 9:576–596.
Hornbeck PV, Kornhauser JM, Tkachev S, Zhang B, Skrzypek E, Murray B, LathamV, and Sullivan M (2012) PhosphoSitePlus: a comprehensive resource for in-vestigating the structure and function of experimentally determined post-translational modifications in man and mouse. Nucleic Acids Res 40:D261–D270.
Hu Y, Leo C, Yu S, Huang BC, Wang H, Shen M, Luo Y, Daniel-Issakani S, PayanDG, and Xu X (2004) Identification and functional characterization of a novel hu-man misshapen/Nck interacting kinase-related kinase, hMINK beta. J Biol Chem279:54387–54397.
Hussain NK, Hsin H, Huganir RL, and Sheng M (2010) MINK and TNIK differen-tially act on Rap2-mediated signal transduction to regulate neuronal structure andAMPA receptor function. J Neurosci 30:14786–14794.
Hutti JE, Jarrell ET, Chang JD, Abbott DW, Storz P, Toker A, Cantley LC, and TurkBE (2004) A rapid method for determining protein kinase phosphorylation speci-ficity. Nat Methods 1:27–29.
Jordan BA, Fernholz BD, Boussac M, Xu C, Grigorean G, Ziff EB, and Neubert TA(2004) Identification and verification of novel rodent postsynaptic density proteins.Mol Cell Proteomics 3:857–871.
Kane S, Sano H, Liu SC, Asara JM, Lane WS, Garner CC, and Lienhard GE (2002) Amethod to identify serine kinase substrates. Akt phosphorylates a novel adipocyteprotein with a Rab GTPase-activating protein (GAP) domain. J Biol Chem 277:22115–22118.
Kaneko S, Chen X, Lu P, Yao X, Wright TG, Rajurkar M, Kariya K, Mao J, Ip YT,and Xu L (2011) Smad inhibition by the Ste20 kinase Misshapen. Proc Natl AcadSci USA 108:11127–11132.
Karayiorgou M, Simon TJ, and Gogos JA (2010) 22q11.2 microdeletions: linking DNAstructural variation to brain dysfunction and schizophrenia. Nat Rev Neurosci 11:402–416.
Kawabe H, Neeb A, Dimova K, Young SM, Jr, Takeda M, Katsurabayashi S,Mitkovski M, Malakhova OA, Zhang DE, and Umikawa M, et al. (2010) Regu-lation of Rap2A by the ubiquitin ligase Nedd4-1 controls neurite development.Neuron 65:358–372.
Koch A and Hauf S (2010) Strategies for the identification of kinase substrates usinganalog-sensitive kinases. Eur J Cell Biol 89:184–193.
Kosik KS, Donahue CP, Israely I, Liu X, and Ochiishi T (2005) Delta-catenin at thesynaptic-adherens junction. Trends Cell Biol 15:172–178.
Kühn S and Geyer M (2014) Formins as effector proteins of Rho GTPases. SmallGTPases 5:e29513.
Lybaek H, Ørstavik KH, Prescott T, Hovland R, Breilid H, Stansberg C, Steen VM,and Houge G (2009) An 8.9 Mb 19p13 duplication associated with precocious pu-berty and a sporadic 3.9 Mb 2q23.3q24.1 deletion containing NR4A2 in mentallyretarded members of a family with an intrachromosomal 19p-into-19q between-arm insertion. Eur J Hum Genet 17:904–910.
Mahmoudi T, Li VS, Ng SS, Taouatas N, Vries RG, Mohammed S, Heck AJ,and Clevers H (2009) The kinase TNIK is an essential activator of Wnt targetgenes. EMBO J 28:3329–3340.
Matigian N, Windus L, Smith H, Filippich C, Pantelis C, McGrath J, Mowry B,and Hayward N (2007) Expression profiling in monozygotic twins discordant forbipolar disorder reveals dysregulation of the WNT signalling pathway. MolPsychiatry 12:815–825.
Matter C, Pribadi M, Liu X, and Trachtenberg JT (2009) Delta-catenin is required forthe maintenance of neural structure and function in mature cortex in vivo. Neuron64:320–327.
Medina M, Marinescu RC, Overhauser J, and Kosik KS (2000) Hemizygosity of delta-catenin (CTNND2) is associated with severe mental retardation in cri-du-chatsyndrome. Genomics 63:157–164.
Mok J, Im H, and Snyder M (2009) Global identification of protein kinase substratesby protein microarray analysis. Nat Protoc 4:1820–1827.
Nonaka H, Takei K, Umikawa M, Oshiro M, Kuninaka K, Bayarjargal M, Asato T,Yamashiro Y, Uechi Y, and Endo S, et al. (2008) MINK is a Rap2 effector forphosphorylation of the postsynaptic scaffold protein TANC1. Biochem Biophys ResCommun 377:573–578.
Nystuen A, Legare ME, Shultz LD, and Frankel WN (2001) A null mutation ininositol polyphosphate 4-phosphatase type I causes selective neuronal loss inweeble mutant mice. Neuron 32:203–212.
Obata T, Yaffe MB, Leparc GG, Piro ET, Maegawa H, Kashiwagi A, Kikkawa R,and Cantley LC (2000) Peptide and protein library screening defines optimalsubstrate motifs for AKT/PKB. J Biol Chem 275:36108–36115.
O’Roak BJ, Vives L, Girirajan S, Karakoc E, Krumm N, Coe BP, Levy R, Ko A, Lee C,and Smith JD, et al. (2012) Sporadic autism exomes reveal a highly interconnectedprotein network of de novo mutations. Nature 485:246–250.
Peng J, Kim MJ, Cheng D, Duong DM, Gygi SP, and Sheng M (2004) Semi-quantitative proteomic analysis of rat forebrain postsynaptic density fractions bymass spectrometry. J Biol Chem 279:21003–21011.
Pieters T, van Roy F, and van Hengel J (2012) Functions of p120ctn isoforms in cell-cell adhesion and intracellular signaling. Front Biosci 17:1669–1694.
Potkin SG, Turner JA, Guffanti G, Lakatos A, Fallon JH, Nguyen DD, Mathalon D,Ford J, Lauriello J, and Macciardi F; FBIRN (2009) A genome-wide associationstudy of schizophrenia using brain activation as a quantitative phenotype.Schizophr Bull 35:96–108.
Sanders AR, Rusu I, Duan J, Vander Molen JE, Hou C, Schwab SG, Wildenauer DB,Martinez M, and Gejman PV (2005) Haplotypic association spanning the 22q11.21genes COMT and ARVCF with schizophrenia. Mol Psychiatry 10:353–365.
Schackmann RC, Tenhagen M, van de Ven RA, and Derksen PW (2013) p120-cateninin cancer - mechanisms, models and opportunities for intervention. J Cell Sci 126:3515–3525.
Schizophrenia Working Group of the Psychiatric Genomics Consortium (2014) Bi-ological insights from 108 schizophrenia-associated genetic loci. Nature 511:421–427.
Schneider M, Debbané M, Bassett AS, Chow EW, Fung WL, van den Bree M,Owen M, Murphy KC, Niarchou M, and Kates WR, et al.; International Con-sortium on Brain and Behavior in 22q11.2 Deletion Syndrome (2014) Psychi-atric disorders from childhood to adulthood in 22q11.2 deletion syndrome:results from the International Consortium on Brain and Behavior in 22q11.2Deletion Syndrome. Am J Psychiatry 171:627–639.
Schürch C, Riether C, Matter MS, Tzankov A, and Ochsenbein AF (2012) CD27signaling on chronic myelogenous leukemia stem cells activates Wnt target genesand promotes disease progression. J Clin Invest 122:624–638.
Sheffer R, Bennett-Back O, Yaacov B, Edvardson S, Gomori M, Werner M, FahhamD, Anteby I, Frumkin A, and Meiner V, et al. (2015) Hindbrain malformation andmyoclonic seizures associated with a deleterious mutation in the INPP4A gene.Neurogenetics 16:23–26.
Shi J, Levinson DF, Duan J, Sanders AR, Zheng Y, Pe’er I, Dudbridge F, HolmansPA, Whittemore AS, and Mowry BJ, et al. (2009) Common variants on chromosome6p22.1 are associated with schizophrenia. Nature 460:753–757.
Shitashige M, Satow R, Jigami T, Aoki K, Honda K, Shibata T, Ono M, Hirohashi S,and Yamada T (2010) Traf2- and Nck-interacting kinase is essential for Wnt sig-naling and colorectal cancer growth. Cancer Res 70:5024–5033.
Taira K, Umikawa M, Takei K, Myagmar BE, Shinzato M, Machida N, Uezato H,Nonaka S, and Kariya K (2004) The Traf2- and Nck-interacting kinase as a pu-tative effector of Rap2 to regulate actin cytoskeleton. J Biol Chem 279:49488–49496.
Ubersax JA and Ferrell JE, Jr (2007) Mechanisms of specificity in protein phos-phorylation. Nat Rev Mol Cell Biol 8:530–541.
Wang Q, Charych EI, Pulito VL, Lee JB, Graziane NM, Crozier RA, Revilla-SanchezR, Kelly MP, Dunlop AJ, and Murdoch H, et al. (2011) The psychiatric disease riskfactors DISC1 and TNIK interact to regulate synapse composition and function.Mol Psychiatry 16:1006–1023.
Xing L, Rai B, and Lunney EA (2014) Scaffold mining of kinase hinge binders incrystal structure database. J Comput Aided Mol Des 28:13–23.
Yao Z, Zhou G, Wang XS, Brown A, Diener K, Gan H, and Tan TH (1999) A novelhuman STE20-related protein kinase, HGK, that specifically activates the c-JunN-terminal kinase signaling pathway. J Biol Chem 274:2118–2125.
Zhang H, Zha X, Tan Y, Hornbeck PV, Mastrangelo AJ, Alessi DR, Polakiewicz RD,and Comb MJ (2002) Phosphoprotein analysis using antibodies broadly reactiveagainst phosphorylated motifs. J Biol Chem 277:39379–39387.
Address correspondence to: Qi Wang, Neuroscience iMED, AstraZeneca,141 Portland Street, Cambridge, MA 02139. E-mail: [email protected] orMichael D. Ehlers, Neuroscience & Pain Research Unit, BioTherapeuticsResearch and Development, Pfizer Inc. 610 Main Street, Cambridge, MA02139. E-mail: [email protected]
Substrates and Phosphorylation Consensus Sequences of TNIK 423
at ASPE
T Journals on N
ovember 28, 2021
jpet.aspetjournals.orgD
ownloaded from

JPET #229880
1
The Journal of Pharmacology and Experimental Therapeutics (JPET)
Identification of Phosphorylation Consensus Sequences and Endogenous Neuronal
Substrates of the Psychiatric Risk Kinase TNIK
Qi Wang, Stephen P. Amato, David M. Rubitski, Matthew M. Hayward, Bethany L. Kormos,
Patrick R. Verhoest, Lan Xu, Nicholas J. Brandon, Michael D. Ehlers
Supplemental Materials and Methods
Chemical synthesis and analysis of PF-794
To a solution of compound 1 (1.5 g, 5.017 mmol) in DME/H2O (5:1) (36 ml) was added
K2CO3 (692 mg, 5.017 mmol), compound 2 (737 mg, 5.017 mmol) and Pd (dppf) Cl2 (184 mg,
0.251 mmol). The mixture was heated to 85oC for 2 hours. Then the mixture was added H2O (10
mL) and extracted with EtOAc (10mL × 3). The organic layers were dried over Na2SO4 and
concentrated to afford the crude product. The crude product was chromatographed on silica gel
(Petroleum ether: EtOAc = 1:1) give compound 3 (1 g, 72.9) as a yellow solid.

JPET #229880
2
To a solution of compound 3 (500 mg, 1.82 mmol) in DME/H2O (5:1) (36 ml) was added
K2CO3 (396 mg, 0.1.82 mmol), compound 4 (453 mg, 2.189 mmol) and Pd (dppf) Cl2 (66.9 mg,
0.091 mmol). The mixture was heated to 85oC for 3 hours. Then the mixture was added H2O (10
mL) and extracted with EtOAc (10mL × 3). The organic layers were dried over Na2SO4 and
concentrated to afford the crude product. The crude product was purified by preparative HPLC to
give PF-794 (233.9 mg, 35.9 %) as a yellow solid. PF-794 (110 mg, 0.309) was dissolved in to
1:1 vol of EtOAc/MeOH (3 ml each) and warmed up to 40oC. 2 M HCl /ether (0.154 mL, 0.309
mmol) was added and the resultant slurry was stirred for 1 h before filtration. The HCl salt (98
mg, 89%) of white solids was recovered after drying. LCMS: m+H 357.2. 1HNMR (400 MHz,
CH3OD): δ 1.25-1.28 (m, 6H), 3.46-3.52 (m, 1H), 7.75-7.79 (m, 2H), 7.92-7.96 (m, 4H), 8.27-
8.31(m, 4H).





























