Embed Size (px)
Citation preview

NeuroImage 83 (2013) 770–781
Contents lists available at ScienceDirect
NeuroImage
j ourna l homepage: www.e lsev ie r .com/ locate /yn img
Image registration of ex-vivo MRI to sparsely sectioned histology ofhippocampal and neocortical temporal lobe specimens
Maged Goubran a,c, Cathie Crukley a, Sandrine de Ribaupierre b,c, Terence M. Peters a,c,d, Ali R. Khan a,d,⁎a Imaging Research Laboratories, Robarts Research Institute, London, Ontario, Canadab Department of Clinical Neurological Sciences, Western University, London, Ontario, Canadac Biomedical Engineering, Western University, London, Ontario, Canadad Department of Medical Biophysics, Western University, London, Ontario, Canada
⁎ Corresponding author at: Imaging Research Laborator100 Perth Drive, London, Ontario N6A 5K8, Canada.
E-mail address: [email protected] (A.R. Khan).
1053-8119/$ – see front matter © 2013 Elsevier Inc. All rihttp://dx.doi.org/10.1016/j.neuroimage.2013.07.053
a b s t r a c t
a r t i c l e i n f oArticle history:Accepted 15 July 2013Available online 26 July 2013
Keywords:Image registrationMRIHistologyEpilepsyAnterior temporal lobectomy
Intractable or drug-resistant epilepsy occurs in up to 30% of epilepsy patients, withmany of these patients under-going surgical excision of the affected brain region to achieve seizure control. Recentmagnetic resonance imaging(MRI) sequences and analysis techniques have the potential to detect abnormalities not identifiedwith diagnosticMRI protocols. Prospective studies involving pre-operative imaging and collection of surgically-resected tissueprovide a unique opportunity for verification and tuning of these image analysis techniques, since direct compar-ison can be made against histopathology, and can lead to better prediction of surgical outcomes and potentiallyless invasive procedures. To carry out MRI and histology comparison, spatial correspondence between the MRimages and the histology imagesmust be found. Towards this goal, a novel pipeline is presented here for bringingex-vivoMRI of surgically-resected temporal lobe specimens and digital histology into spatial correspondence. Thesparsely-sectioned histology images represent a challenge for 3D reconstruction which we address with a com-bined 3D and 2D registration algorithm that alternates between slice-based and volume-based registrationwith the ex-vivo MRI. We evaluated our registration method on specimens resected from patients undergoinganterior temporal lobectomy (N = 7) and found our method to have a mean target registration error of0.76 ± 0.66 and 0.98 ± 0.60 mm for hippocampal and neocortical specimens respectively. This work allowsfor the spatially-local comparison of histology with post-operative MRI and paves the way for eventual correla-tion with pre-operative MRI image analysis techniques.
© 2013 Elsevier Inc. All rights reserved.
Introduction
Intractable or drug-resistant epilepsy occurs in over 30% of epilepsypatients and is commonly characterized by partial refractory seizures(Engel, 1998). When there is a localized focus, the standard of care forthese patients is a surgical excision of the affected brain region in orderto achieve seizure control (Engel et al., 1992). Current clinical imagingprotocols and surface EEG techniques of epileptogenic focus localizationmay not be sufficient for pre-operative planning due to limited sensitiv-ity to deeper brain structures, and low resolution of source localizationtechniques (Smith, 2005). These limitations often necessitate invasiveelectrophysiological monitoring using sub-dural strips or grid elec-trodes. Magnetic resonance imaging (MRI) techniques such as diffusiontensor imaging (DTI), relaxometry mapping, high resolution functionalMRI (fMRI), voxel-based morphometry, and cortical thickness analysiscan detect abnormalities not identified with conventional or diagnosticMRI protocols (Bernasconi et al., 2000, 2004; Bernhardt et al., 2009).
ies, Robarts Research Institute,
ghts reserved.
This is important since it has been shown that post-operative outcomescan be predictedmore accurately in patients where lesions can be iden-tified (de Tisi et al., 2011; Fish et al., 1993). These techniques also havethe potential to improve pre-operative localization of the focus, pavingtheway towards less invasive procedures and better surgical outcomes.With the plethora of MRI sequences and analysis techniques available,the challenge then becomes how to verify and evaluate the sensitivityand specificity of these techniques for detection of abnormal brain tis-sue. Prospective studies involving pre-operative imaging and collectionof resected tissue from surgery (Eriksson et al., 2007; Howe et al., 2010)provide a unique opportunity for verification and tuning of these imageanalysis techniques, since direct comparison can be made against high-resolution ex-vivo imagery, histology and immunohistochemistry. Inorder to carry out this verification, spatial correspondence betweenthe MR images and the tissue-derived histology images must be found,which remains challenging due to the deformations involved duringresection of the tissue and histological processing and the difficulty ofaccurately registering the histology slices to theMR images. Registrationis the task of finding this spatial correspondence by modeling the trans-formations and deformations that occur due to surgical resection andtissue handling. These deformations can be divided into two types:
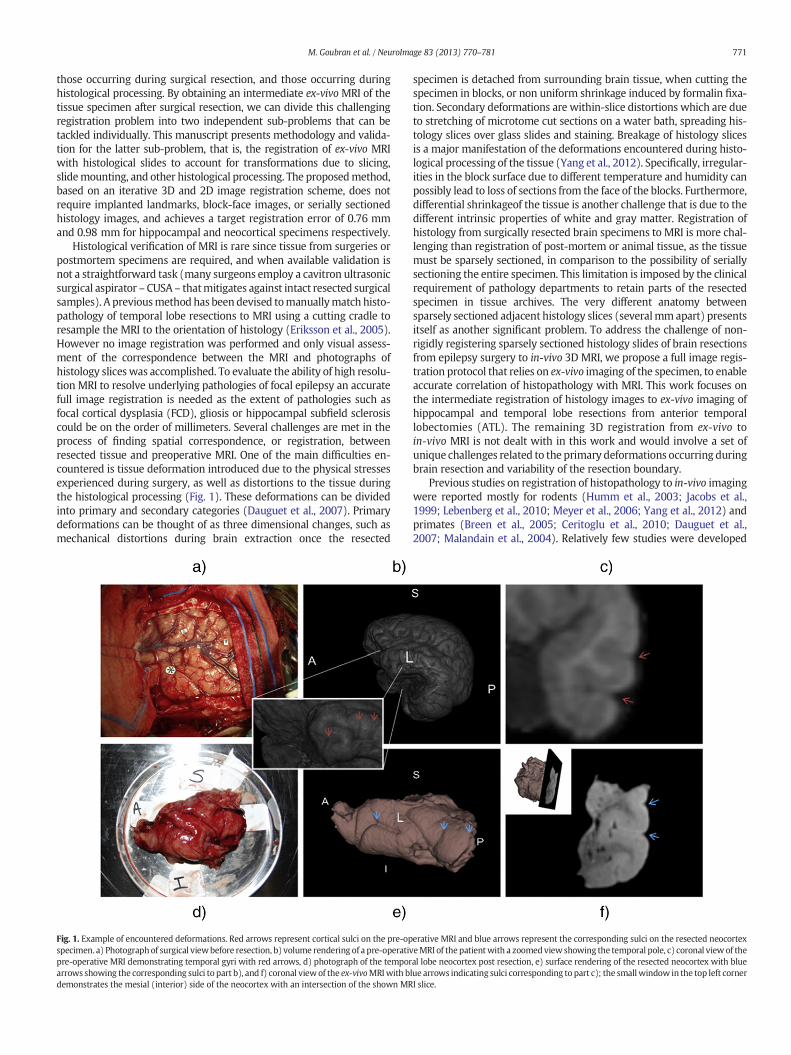
771M. Goubran et al. / NeuroImage 83 (2013) 770–781
those occurring during surgical resection, and those occurring duringhistological processing. By obtaining an intermediate ex-vivo MRI of thetissue specimen after surgical resection, we can divide this challengingregistration problem into two independent sub-problems that can betackled individually. This manuscript presents methodology and valida-tion for the latter sub-problem, that is, the registration of ex-vivo MRIwith histological slides to account for transformations due to slicing,slidemounting, and other histological processing. The proposedmethod,based on an iterative 3D and 2D image registration scheme, does notrequire implanted landmarks, block-face images, or serially sectionedhistology images, and achieves a target registration error of 0.76 mmand 0.98 mm for hippocampal and neocortical specimens respectively.
Histological verification of MRI is rare since tissue from surgeries orpostmortem specimens are required, and when available validation isnot a straightforward task (many surgeons employ a cavitron ultrasonicsurgical aspirator –CUSA – thatmitigates against intact resected surgicalsamples). A previousmethodhas been devised tomanuallymatch histo-pathology of temporal lobe resections to MRI using a cutting cradle toresample the MRI to the orientation of histology (Eriksson et al., 2005).However no image registration was performed and only visual assess-ment of the correspondence between the MRI and photographs ofhistology slices was accomplished. To evaluate the ability of high resolu-tion MRI to resolve underlying pathologies of focal epilepsy an accuratefull image registration is needed as the extent of pathologies such asfocal cortical dysplasia (FCD), gliosis or hippocampal subfield sclerosiscould be on the order of millimeters. Several challenges are met in theprocess of finding spatial correspondence, or registration, betweenresected tissue and preoperative MRI. One of the main difficulties en-countered is tissue deformation introduced due to the physical stressesexperienced during surgery, as well as distortions to the tissue duringthe histological processing (Fig. 1). These deformations can be dividedinto primary and secondary categories (Dauguet et al., 2007). Primarydeformations can be thought of as three dimensional changes, such asmechanical distortions during brain extraction once the resected
Fig. 1. Example of encountered deformations. Red arrows represent cortical sulci on the pre-opspecimen. a) Photographof surgical viewbefore resection, b) volume rendering of a pre-operativpre-operative MRI demonstrating temporal gyri with red arrows, d) photograph of the temporarrows showing the corresponding sulci to part b), and f) coronal viewof the ex-vivoMRIwith bdemonstrates the mesial (interior) side of the neocortex with an intersection of the shown MR
specimen is detached from surrounding brain tissue, when cutting thespecimen in blocks, or non uniform shrinkage induced by formalin fixa-tion. Secondary deformations are within-slice distortions which are dueto stretching of microtome cut sections on a water bath, spreading his-tology slices over glass slides and staining. Breakage of histology slicesis a major manifestation of the deformations encountered during histo-logical processing of the tissue (Yang et al., 2012). Specifically, irregular-ities in the block surface due to different temperature and humidity canpossibly lead to loss of sections from the face of the blocks. Furthermore,differential shrinkageof the tissue is another challenge that is due to thedifferent intrinsic properties of white and gray matter. Registration ofhistology from surgically resected brain specimens to MRI is more chal-lenging than registration of post-mortem or animal tissue, as the tissuemust be sparsely sectioned, in comparison to the possibility of seriallysectioning the entire specimen. This limitation is imposed by the clinicalrequirement of pathology departments to retain parts of the resectedspecimen in tissue archives. The very different anatomy betweensparsely sectioned adjacent histology slices (severalmmapart) presentsitself as another significant problem. To address the challenge of non-rigidly registering sparsely sectioned histology slides of brain resectionsfrom epilepsy surgery to in-vivo 3D MRI, we propose a full image regis-tration protocol that relies on ex-vivo imaging of the specimen, to enableaccurate correlation of histopathology with MRI. This work focuses onthe intermediate registration of histology images to ex-vivo imaging ofhippocampal and temporal lobe resections from anterior temporallobectomies (ATL). The remaining 3D registration from ex-vivo toin-vivo MRI is not dealt with in this work and would involve a set ofunique challenges related to the primary deformations occurring duringbrain resection and variability of the resection boundary.
Previous studies on registration of histopathology to in-vivo imagingwere reported mostly for rodents (Humm et al., 2003; Jacobs et al.,1999; Lebenberg et al., 2010; Meyer et al., 2006; Yang et al., 2012) andprimates (Breen et al., 2005; Ceritoglu et al., 2010; Dauguet et al.,2007; Malandain et al., 2004). Relatively few studies were developed
erative MRI and blue arrows represent the corresponding sulci on the resected neocortexeMRI of thepatientwith a zoomedviewshowing the temporal pole, c) coronal view of theal lobe neocortex post resection, e) surface rendering of the resected neocortex with bluelue arrows indicating sulci corresponding to part c); the smallwindow in the top left cornerI slice.

Table 1Summary of demographics and clinical data, includingMRI and histopathological findings,for the seven recruited patients in the study. Registration was performed on both hippo-campus and neocortex specimens for all patients. In two cases (denoted by *) registrationwasperformed only on theneocortexdue to amissed scan and a fragmentedhippocampusspecimen. MTS: mesial temporal sclerosis. FCD: focal cortical dysplasia.
Patient Gender Age Onset age Seizureorigin
MRI Path. Scanprotocol
1 F 51 10 Right Normal Dysplasia I2 F 22 15 Right Normal Mild MTS I3 F 52 12 Left Non-specific Mild FCD I4* F 26 20 Right Tuberous
sclerosisCorticaltubers
II
5 F 22 15 Right R. MTS MTS II6 M 20 3 Left L. MTS MTS II7* M 19 5 Right Normal Gliosis II
772 M. Goubran et al. / NeuroImage 83 (2013) 770–781
to register human brain MRI to histology, with the majority beingperformed on whole-brain or single hemisphere postmortem data(Kim et al., 2000; Schormann et al., 1995; Singh et al., 2008). Theselandmark-based and image-based registration algorithms, however,are not likely to be applicable to the registration of specimens fromlobectomies to full preoperative MR images, due to the drastic changein shape and coherence when the specimen is separated from neighbor-ing tissue. Othermethods have been proposed that allow co-registrationof histology to other modalities through the use of stereotactic systemsusing target points (Humm et al., 2003; Schmierer et al., 2003), howeverthe design of these systems is tissue-specific and is not broadly applica-ble to other brain resections. To the best of our knowledge, this is the firstinstance where human brain surgery specimens were registered to his-tology using an image-based algorithm in a clinical setting, with a targetregistration error reported and validated.
Presently, there are no automatedhistology toMRI image registrationprotocols that could be widely applicable to focal resections of humanbrain, such as tissue resected during epilepsy surgery. We present herea protocol to register ex-vivo scans of hippocampal and neocortical tem-poral lobe resections to histology as an intermediate step that reducesthe complexity of the preoperative MRI to 2D sparse histology problem.Specifically we describe a novel landmark-free algorithm for simulta-neous reconstruction and alignment of sparsely sectioned histologicaldata to ex-vivo MRI, and a quantitative validation for our registrationmethod. Performing this intermediate step addresses most of the chal-lenges of registration to in-vivo imaging due to the higher resolutionand reduceddeformations of the ex-vivo images. Furthermore, the higherresolution of specimen imaging is advantageous for examining the corre-lation between MRI and histology. The proposed method represents asignificant step towards in-vivo MRI to histology registration in theclinical setting and can be broadly applicable toMRI and histopathologycorrelations of resections other than epilepsy surgery.
Methods & materials
Recruitment, surgery & specimen acquisition
Seven patients suffering from intractable temporal lobe epilepsy(TLE) were recruited as part of an ongoing study. This project has beenapproved by the office of research and ethics of Western University,and informed consent was obtained from all patients prior to theirrecruitment in the study. All such patients were recommended for ATLsurgery by the department of clinical neurological sciences at theUniversity Hospital (UH) of the London Health Sciences Centre, andhad preoperative investigations including neuropsychological testingand 1.5 T clinical MRI scans which included T1w, T2w, FLAIR, anddiffusion-weighted sequences. Patients were monitored with scalp-based electroencephalogram (EEG) video telemetry for seizure charac-terization, with three patients having to undergo monitoring withsubdural placement of strip electrodes. In addition to the 1.5 T clinicalMRI scans performed at the hospital, patients underwent a series ofscans on 3 T and 7 T MRI research scanners, including high-resolutionstructural imaging, diffusion-tensor imaging, relaxation mapping andresting-state functional imaging prior to surgery. Following surgery,the resected tissue specimens were transferred to the Robarts ResearchInstitute for ex-vivo specimen imaging on the same3 T scanner and thento the pathology technologist for histological processing. From the sevenpatients, fourteen resected specimenswere collected (7neocortex and 7hippocampus), but only twelve out of the fourteen were used in thestudy due to a fragmented hippocampus specimen and missed ex-vivohippocampus scan. All the specimens used in the manuscript were enbloc resections, however the size of the neocortex specimens weresmaller if the resection was on the language-dominant side. The meanvolume of the hippocampal specimens was 4.30 ± 0.41 mL; and themean volumes of the neocortical specimens for language-dominantside resections and non-language-dominant resections were 12.9 ±
1.78 mL and 23.2 ± 6.41 mL respectively. It should be noted however,that different surgeons use slightly different techniques, which couldaccount for the variability in the volume of specimens. Table 1 summa-rizes the patients' demographic data, as well as, their clinical MRI andhistopathological findings.
Specimen ex-vivo MR imaging
After resection, each specimenwasplaced in a large petri dishwithina specialized sealed cooler for specimen transport, and orientationlabels were marked on the container by the operating neurosurgeon,with photographs taken for future reference. MR imaging was carriedout on the specimens in two sessions: immediately following surgicalresection, and after overnight fixation in 10% formalin. For the initialsession, referred to as the pre-fixation session, the specimens were im-mediately transferred from the operating room to the scanning suite atthe Robarts Research Institute and prepared for imaging. Each specimenwas wrapped in gauze for stabilization, transferred to suitably-sizedcontainers for imaging, and immersed in a fluorine-based lubricant‘Christo-lubeMCG 1046’ (Lubrication Technology, Inc.) prior to imagingto avoid susceptibility artifacts at the tissue boundaries. Identical prep-aration was performed for the second post-fixation session.
Specimen imagingwas performedon a 3 TDiscoveryMR750 scanner(GEMedical Systems, Milwaukee,WI, USA.). Initially, an in-house devel-oped gradient-insert coil was employed in the scanning setup witheach specimen imaged sequentially using different coils. For improvedtime-efficiency in scanning and setup, the gradient-insert coil was notemployed in later studies, and both specimens were imaged in thesame field of view. Post-fixation T2-weighted scans were used in thesubsequent image processing and registration. Both scanning protocolsare described in detail below:
Scan protocol IThe first protocol utilized a gradient-insert with a 4 channel
TORO coil for the neocortex and a solenoid coil for the hippocampus,with each specimen scanned sequentially. T2-weighted images with amulti-phase balanced SSFP FIESTA sequence with 4 cycled phaseswere acquired for the neocortex (TR = 3.5 ms, TE = 1.75 ms, flipangle = 40°, N = 4, matrix = 200 × 200, slice thickness = 0.3,FOV = 60 mm) and the hippocampus (TR = 3.97 ms, TE = 1.98 ms,flip angle = 40°, N = 4, matrix = 200 × 200, slice thickness = 0.3,FOV = 60 mm).
Scan protocol IIFor the second protocol, a 6 channel coil, designed to image the
carotid artery, was used instead of the gradient-insert coil of the previ-ous configuration. Similar T2-weighted FIESTA images (TR = 8.17 ms,TE = 4.08 ms, flip angle = 40°, N = 2, matrix = 200 × 200, slicethickness = 0.4, FOV = 70 mm) with a resolution of 0.35 × 0.35 ×0.4 mm, as well as Fast gradient echo (fastGRE) scans with sixteen

773M. Goubran et al. / NeuroImage 83 (2013) 770–781
echoes (TR = 65.0 ms, TE = 38.9 ms, flip angle = 40°, matrix =200 × 200, slice thickness = 0.4, FOV = 70 mm) were acquired forthe study. A switch was made to the second protocol due to the signif-icant time savings achieved during the setup and gradient shimmingprocesses (from ~445 min to ~5 min)with minimal loss of image reso-lution. Note that similar SSFP FIESTA sequences were used in both pro-tocols to provide the images used in registration.
Histological processing
Following pre-fixation and post-fixationMRI imaging, the specimensunderwent accessioning and grossing at the Department of Pathology atUH, and were then cut into two halves midway, anterior-posterior,through the specimen. Each half of the specimen was then embeddedin agar for a stabilization effect during slicing. The half-specimens werethen sectioned parallel to the initial cut, into 4.4 mm pieces in the ante-rior to posterior direction using a deli slicer (Globe Food EquipmentCompany, Dayton, OH, USA). Each block was embedded in paraffin andmounted on a microtome where 8 μm thick sections were cut from theface of each block and mounted on slides. The constraint of cuttingonly one section from each block stems from two limitations. The firstbeing the requirement set forth by the committee governing tissue use,which requires that the remaining part of the blocks be reserved intheir tissue archive in case further analysis is needed. The second factoris the significantly increased cost associated with acquiring serial sec-tions of all our blocks for both types of specimens, which would restrictthe number of subjects we could recruit for this ongoing study.
One slide from each block was stained with hematoxylin and eosin(H&E) according to standard clinical neuropathology protocols, andadditional stains or immunohistochemistry (IHC), mainly glial fibrillaryacidic protein (GFAP) and neuronal nuclei (NeuN), as well as neurofila-ment (NF), luxol fast blue (LFB) and cluster of differentiation antibody(CD34) were ordered when deemed necessary by the neuropathologiston duty. The median numbers of Hp and Neo blocks were 8 and 12respectively. For the hippocampus, on average about a quarter of theblocks were additionally stained with GFAP. As for the neocortex,about half of the blocks were additionally stained with a combinationof the previously mentioned stains and immunohistochemistry; whileGFAP and NeuN being the two most common additional stains. Theex-vivo subvolumes, the numbers of histological blocks and sections,as well as NeuN and GFAP sections, are summarized in Table 2.The average physical size of the hippocampal histological sections(slide area (mm2)) was 96.5 ± 60.9, and 281.0 ± 168.9 for neocorticalsections.
The resulting slides were digitized on a ScanScope GL (Aperio Tech-nologies, Vista, CA, USA) brightfield slide scanning systemat amaximumof 20× optical zoom, and stitched to form full-frame multi-resolutionimages stored in BigTIFF file format(maximum pixel resolution0.5 μm). The full resolution BigTIFF histology images were retained forfuture quantitative analysis. Since each specimen was sectioned intoblocks of 4.4 mm thickness, the corresponding H&E stained imageshave a physical spacing of effectively 4.4 mm in the coronal (anterior–
Table 2Summary of ex-vivo subvolumes, the numbers of histological blocks and sections, as well as Ne
Subject Hp volume (mm2) Hp blocks Hp sections
1 4808.4 9 18 (9 GFAP)2 4140.3 9 11 (2 GFAP)3* 3896.1 10 10 (No IHC)4 – – –
5 3992.4 6 6 (No IHC)6* 4677.0 7 8 (1 GFAP)7* – – –
* Denote subjects which had dominant hemisphere resections, as is evident by the reduced ne
posterior) direction. Fig. 2 shows an overview of all these histologicalprocessing steps.
Image registration
To motivate our registration approach we first describe how thenumerous physical processing steps between ex-vivo MR imaging andslide digitization affect the specimen, and how these steps could beaccounted for with registration. As outlined in the previous section,after imaging, the tissue specimen is sectioned coronally, but this slicingplane is not enforced to be along the orthogonal axes corresponding tothe MRI coordinate system. We therefore need to obtain a transforma-tion between the MRI axes and tissue slicing axes. Next, when thespecimen is mounted on the microtome, there may be variability inthe angle at which sections are taken and in the number of partialsections removed before a full section is retained. This effectively leadsto variability in the angle and spacing between sections. For similarprocedures carried out on prostate specimens, Gibson et al. (2012b)quantified the variability to be 1.7 ± 1.1° and 1.0 ± 0.5 mm in angleand spacing respectively. Because of the relatively small magnitude ofvariability, which presumably would be similar for brain sections, aswell as the ease of working with parallel sections, we do not explicitlyaccount for this and instead assume sections are parallel and spacedby 4.4 mm. The tissue being sectioned in themicrotome is highly foldedafter the blade is brought down, thus tomount the section on a slide, it isfirst placed in a water bath to unfold, then eased onto the glass slide.This procedure can introduce folds or tears in the mounted section,and placement on the slide is variable. Histological processing and stain-ing of the section can introduce further distortions, such as differentialshrinkage or expansion of tissue. Since all these deformations are pres-ent in the thin (8 μm) section of tissue mounted on the slide, thesecan be modeled as transformations and warps constrained to the 2Dplane.
In summary, we require a registration approach that can model:1) the transformation between the 3DMRI axes to the specimen slicingaxes (3D rigid transformation), and 2) the transformations and defor-mations of each slide-mounted section constrained to the 2D space ofthe slide (2D rigid transformations and non-rigid deformations).
Iterative registration algorithm
In this sectionwe outline our iterative registration approach to attainthe transformations and deformations and to establish correspondencebetween theMRI and histology images. Note that the registration proce-dures for hippocampus and neocortex images were carried out sepa-rately in each case. Preliminary results for neocortex registration wereshown in Goubran et al. (2012).
If a 3D reconstruction of the histology were given, 3D rigid imageregistration could be used to align the MRI to the histology. However,to generate a 3D reconstruction of the histology, the individual histologyslices would need to be corrected, using the registered MRI as a refer-ence. Thus we see that obtaining the 3D rigid transformation is
uN and GFAP sections.
Neo volume (mm2) Neo blocks Neo sections
21331.0 16 20 (4 GFAP)20363.1 10 14 (2 NeuN, 2 GFAP)14087.3 16 20 (1 NeuN, 1 GFAP)32669.3 14 37 (13 NeuN, 3 GFAP)18472.0 12 21 (3 Neu-N, 3 GFAP)13830.8 12 15 (2 NeuN)10885.8 9 15 (3 NeuN, 2 GFAP)
ocortical resected volume.
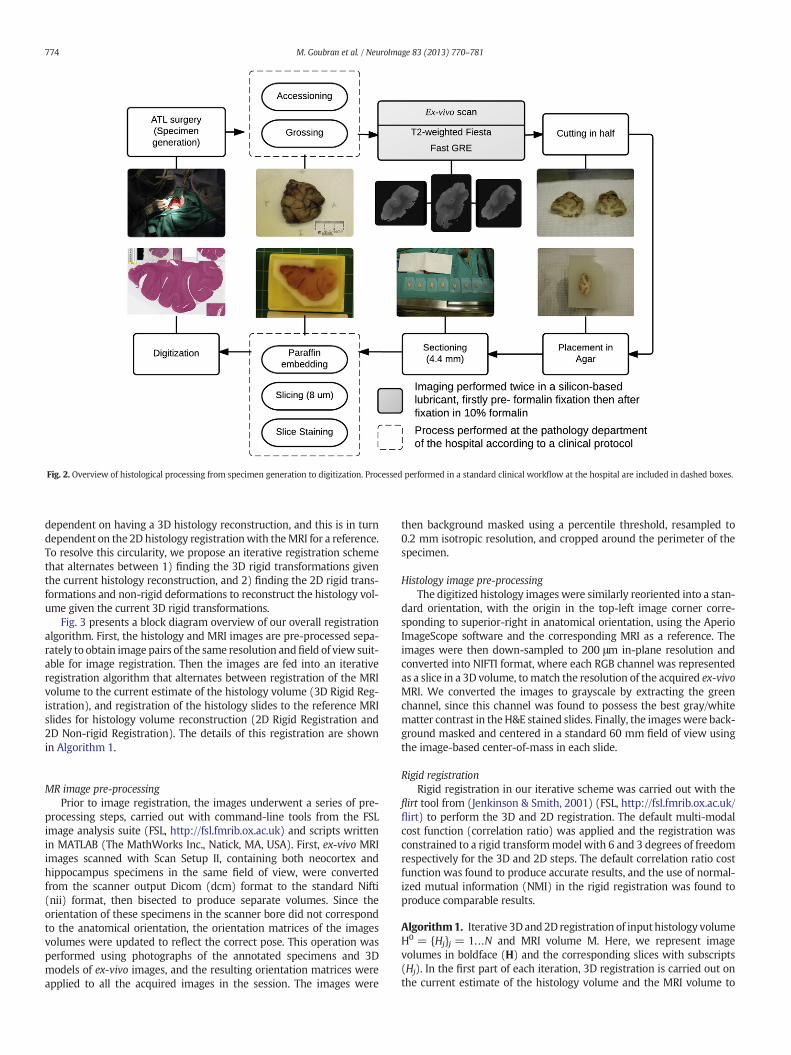
Fig. 2. Overview of histological processing from specimen generation to digitization. Processed performed in a standard clinical workflow at the hospital are included in dashed boxes.
774 M. Goubran et al. / NeuroImage 83 (2013) 770–781
dependent on having a 3D histology reconstruction, and this is in turndependent on the 2D histology registrationwith theMRI for a reference.To resolve this circularity, we propose an iterative registration schemethat alternates between 1) finding the 3D rigid transformations giventhe current histology reconstruction, and 2) finding the 2D rigid trans-formations and non-rigid deformations to reconstruct the histology vol-ume given the current 3D rigid transformations.
Fig. 3 presents a block diagram overview of our overall registrationalgorithm. First, the histology and MRI images are pre-processed sepa-rately to obtain image pairs of the same resolution andfield of view suit-able for image registration. Then the images are fed into an iterativeregistration algorithm that alternates between registration of the MRIvolume to the current estimate of the histology volume (3D Rigid Reg-istration), and registration of the histology slides to the reference MRIslides for histology volume reconstruction (2D Rigid Registration and2D Non-rigid Registration). The details of this registration are shownin Algorithm 1.
MR image pre-processingPrior to image registration, the images underwent a series of pre-
processing steps, carried out with command-line tools from the FSLimage analysis suite (FSL, http://fsl.fmrib.ox.ac.uk) and scripts writtenin MATLAB (The MathWorks Inc., Natick, MA, USA). First, ex-vivo MRIimages scanned with Scan Setup II, containing both neocortex andhippocampus specimens in the same field of view, were convertedfrom the scanner output Dicom (dcm) format to the standard Nifti(nii) format, then bisected to produce separate volumes. Since theorientation of these specimens in the scanner bore did not correspondto the anatomical orientation, the orientation matrices of the imagesvolumes were updated to reflect the correct pose. This operation wasperformed using photographs of the annotated specimens and 3Dmodels of ex-vivo images, and the resulting orientation matrices wereapplied to all the acquired images in the session. The images were
then background masked using a percentile threshold, resampled to0.2 mm isotropic resolution, and cropped around the perimeter of thespecimen.
Histology image pre-processingThe digitized histology images were similarly reoriented into a stan-
dard orientation, with the origin in the top-left image corner corre-sponding to superior-right in anatomical orientation, using the AperioImageScope software and the corresponding MRI as a reference. Theimages were then down-sampled to 200 μm in-plane resolution andconverted into NIFTI format, where each RGB channel was representedas a slice in a 3D volume, tomatch the resolution of the acquired ex-vivoMRI. We converted the images to grayscale by extracting the greenchannel, since this channel was found to possess the best gray/whitematter contrast in theH&E stained slides. Finally, the imageswere back-ground masked and centered in a standard 60 mm field of view usingthe image-based center-of-mass in each slide.
Rigid registrationRigid registration in our iterative scheme was carried out with the
flirt tool from (Jenkinson & Smith, 2001) (FSL, http://fsl.fmrib.ox.ac.uk/flirt) to perform the 3D and 2D registration. The default multi-modalcost function (correlation ratio) was applied and the registration wasconstrained to a rigid transformmodel with 6 and 3 degrees of freedomrespectively for the 3D and 2D steps. The default correlation ratio costfunction was found to produce accurate results, and the use of normal-ized mutual information (NMI) in the rigid registration was found toproduce comparable results.
Algorithm1. Iterative 3Dand 2D registration of input histology volumeH0 = {Hj}j = 1…N and MRI volume M. Here, we represent imagevolumes in boldface (H) and the corresponding slices with subscripts(Hj). In the first part of each iteration, 3D registration is carried out onthe current estimate of the histology volume and the MRI volume to

Fig. 3. Registration pipeline showing pre-processing steps of the data and our iterative 2D–3D approach. The left column demonstrates the pre-processing steps applied to the histologyslides sequentially from top to bottom. Likewise, the right column represents the pre-processing steps applied to the MRI of the specimens. The resulting histology stack acts as a fixedimage to transform the MR image in a 3D rigid registration. The transformed image as well as the stack is split into slices 4.4 mm apart where each histology slice has a correspondingMRI slice. These MR slices act as fixed images to deform the histology slices rigidly then non-rigidly. The resulting deformed histology slices are stacked and fed back into the 3D rigidregistration for the next iteration.
775M. Goubran et al. / NeuroImage 83 (2013) 770–781
obtain the transformation between theMRI axes to the specimen slicingaxes. In the second part, 2D registration is carried out to obtain thetransformations and deformations of each slide-mounted histology sec-tion using the current estimate of the aligned reference MRI.
Non-rigid registrationAdeformable registration between corresponding histology andMRI
slices was employed to account for any anisotropic tissue deformationsthat can occur during histological processing, sectioning, and staining.We used a fast non-rigid registration thatmakes use of a B-spline defor-mationfield and a normalizedmutual information cost-function (Modatet al., 2010; Rueckert et al., 1999) (NiftyReg,http://sourceforge.net/projects/niftyreg/). The B-spline image registration used a three-levelmulti-resolution image pyramid with final control point spacing of2 mm. This control point spacingwas chosen after a number of differentcontrol point spacings (0.1, 0.5,1, 2, 4, 6 mm) were qualitatively evalu-ated on representative sections on multiple specimens and the 2 mmspacing was found to perform best, as it is small enough to account forlocal deformations encountered and large enough to avoid noise andprovide a smooth deformation. Non-rigid registration was carried outstarting at iteration 3 of the algorithm and not having been employedin the first two iterations to ensure the sufficient convergence of therigid registration step. Furthermore, for slices where foreground of theMRI image or histology image was below a specified threshold, non-rigid registration was not performed and a zero deformation wasassumed for the slice. The deformation penalty term (bending energyof the spline at a control point), was successively relaxed after each iter-ation to allow for greater deformations as the alignment is improvedover each iteration. Specifically the sequence of bending energiesemployed at the iterative registration steps were {0.5,0.025,0.01}, foriterations 3–5.
Due to histology tissue breakage and loss, afinal registration stepwasadded where binary ‘ignore’ masks defined on the registered histologysliceswere included in the deformable registration scheme. These ignoremasksweremanually-defined in 3DSlicer (http://www.slicer.org) usinga large 2 mm radius paintbrush on regions of the MRI image where tis-sue loss is readily apparent in corresponding regions of the histologyimage, preventing these regions from contributing to the registration,which would result in incorrect deformations since one-to-one tissue

776 M. Goubran et al. / NeuroImage 83 (2013) 770–781
correspondence is unattainable. The average percentage of sectionsrequiring registration masks was 75% and 82.5% for the neocortex andhippocampus respectively. An example of a registration mask for a hip-pocampal section is shown on Fig. 8-e.
Coregistration of additional stainingAs noted previously additional IHC stainingwas performed on a sub-
set of blocks for clinical diagnostic purposes. To make use of these IHCstained images in future quantitative analysis, a supplementary registra-tion stepwas carried out in order to register the sections with additionalIHC to the sections with H&E stains. This registration step must beperformed for these additional stains or IHC since additional sectionswere cut, placed on slides, and processed for each stain. We performed2D affine registration between the H&E and the each IHC stainedimage with the flirt tool (Jenkinson and Smith, 2001) on histologyimages downsampled to 200 μm resolution and converted to grayscale.We performed this registration for NeuN andGFAP IHC since thesewerethe most common and relevant in diagnosis or quantification of neuro-nal loss, cytoarchitectural abnormalities, and gliosis.
Registration validation
Landmark-based validationTo validate our registration protocol, we computed target registra-
tion error (TRE) based on manually-identified corresponding intrinsiclandmarks on MR images and histology slices. These landmarks wereused as independent targets to assess the accuracy of the registrationat each iteration of the iterative registration scheme, as well as afterdeformable warping of the images.
We found that micro-vasculature or micro-bleeds that were visibleon the H&E histology slides appeared as dark hypo-intense regions inthe ex-vivo T2-weighted MRI, as demonstrated in Fig. 4. First, one rateridentified landmarks on histology slides (downsampled to 10 μm perpixel), restricting selection to vasculature with a transverse diameterof more than 35 pixels wide, assuming an ellipsoid shape. The mostanterior and posterior histology slices in many cases of both specimensdid not contain sufficient intact tissue for reliable placement of anatom-ical landmarks. The landmarkswere 20 per neocortical specimen and 12per hippocampal specimen on average. Fig. 5 demonstrates the distribu-tion of landmarkswithin a neocortical specimen. Three raters then inde-pendently searched through the ex-vivo MRI to locate correspondinglandmarks in the MRI images representing the centroid of thesemicro-vasculature ormicro-bleeds. Since tissue contrast varies through-out the specimen, other MR scans of the specimen were used to facili-tate the localization process. To compute the TRE the coordinates forall three raters were averaged to generate a consensus set of MRI land-marks. A total of 215 pairs of corresponding landmarks were identifiedfor the TRE calculations in the twelve specimens. Note that, to ensureconsistent landmark locations, only a single set of the histology land-marks was used.
Fig. 4. Example of the chosen intrinsic landmarks on histology and their localized correspondinvasculature used as targets for the TLE calculations on both histology and MRI. A zoomed in wcoordinates on MRI and a circle showing the chosen coordinates on histology.
For the separate registration between the original H&E stain and theadditional IHC (NeuN, GFAP), the target registration error of the 2Daffine registration was evaluated for 2 subject neocortical specimens,in a total of 5 blocks, placing a total of 40 pairs of intrinsic landmarks.These histology intrinsic landmarks consisted of micro-vasculature ormicro-bleeds visible on the different types of stains and IHC, resemblingthe histology landmarks used to validate our MRI to histology registra-tion. The 2D euclidean distance was measured between these corre-sponding landmarks to compute the TRE.
Localization error and statistical analysisTarget localization incorporates humanerror in localizing the coordi-
nates which combines with the image registration error to produce theTREmeasurements. The target localization error (TLE)was calculated onex-vivo MRI images as an unbiased estimator of the standard deviationof repeated localizations of the same landmark by the same rater(Fitzpatrick et al., 1998), described by the equation below:
TLE ¼ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi1J
XJ
j¼1
1K−1
XKK¼1
jjP j;k−1K
XK
k¼1
P j;kjj2vuut
where Pj,k is the k-th localization of the j-th landmark. A total of fivelocalization (K = 5) of twenty landmarks (J = 20) was performed.
Inter-rater variability was measured as an estimator of the standardof deviation of repeated localization of the same landmarks by differentraters, where a hundred and twenty eight landmarks (J = 128) wereplaced by three different raters (K = 3). Statistical analyses wereperformed in Prism 5.04 (GraphPad Software, San Diego, CA). To assessfor significant differences between the several iterations of the algo-rithm and across rigid and non-rigid steps, we computed a repeated-measures analysis of variance (ANOVA) of the mean TRE value of eachof these steps followed by Bonferroni multiple-comparison correction.A Bartlett's test for equal variances was conducted between all itera-tions (rigid and non-rigid) of the algorithm to verify the equal varianceassumption of the ANOVA tests.
Results
The proposed methodology required ~85 min (including setup) forex-vivo imaging and b10 min for execution of both the rigid and nonrigid components of the algorithm (excluding the manual re-orientationstep in the pre-processing scheme and the time for definition of ignoremasks). Evaluation of the protocol was performed by localization ofmicro vasculature landmarks seen on both modalities by three raters.The localization protocol yielded 2–4 homologous landmarks on eachof 8–12 sections per neocortex specimen and 1–3 landmarks on eachof 6–8 sections per hippocampus specimen. Our registration protocolproduced amean target registration error of 0.76 ± 0.66 for hippocam-pal specimens, as shown in Table 3, and 0.98 ± 0.60 for neocortical
g landmark on a neocortical ex-vivoMRI specimen. The arrows in blue pinpoint the microindow demonstrates the targets on both modalities with cross hairs indicating the target

Fig. 5. 3D visualization revealing the wide spatial distribution of validation landmarks (blue spheres) within an exemplar neocortical specimen in sagittal and coronal views.
777M. Goubran et al. / NeuroImage 83 (2013) 770–781
specimens, as shown in Table 4. The mean TRE was below 1.2 mm afterthe last step of the registration algorithm in all cases including bothspecimens. The mean landmark localization error for the three raterswas 0.21 mm, relative to an MRI voxel size of 0.35 × 0.35 × 0.4 mm,while the inter-rater reliability between the raters was 0.33 mm.Fig. 6 shows the registration errors across all steps of the algorithm in-cluding both rigid and non-rigid components for both specimens. Theerrors are shown first along iterations 1, 3 and 5 of the rigid componentthen the non-rigid steps beginningwith a step using a high bending en-ergy regularization penalty; then a low penalty weight and finally de-formable registration utilizing ignore masks, that account for tissuebreakage and differential shrinkage. The mean error of the rigid itera-tions reaches a plateau around iteration 5 for both specimens. Themasked imaged based step outperformed the non-masked registrationas expected by avoiding tissue breakage. For the supplemental 2D affineregistration between sections with the original H&E stain and sectionswith the additional IHC (Neu-N, GFAP), the target registration errorwas 0.46 ± 0.31 and 0.41 ± 0.23 for NeuN and GFAP respectively.This validation demonstrates our ability to accurately align, and poten-tially correlate, MRI with additional stains and immunohistochemistry.
Fig. 7 shows three neocortical slices with their corresponding MRIslices, that represent the locations where the histology cuts were madewith respect to the MRI scans of the resections, as well as the transfor-mations of the slices after deformable registration. A picture of theresected specimen along with a volume rendering demonstrating thelocation of these histology slices in respect to the whole specimen arealso shown in the figure. Checkerboard images of both rigid and non-rigid registration for a hippocampal slice are displayed in Fig. 8, whichalso shows a rendered representation of both sides of the hippocampuswhere the histology slice was cut.
The Bartlett's test confirmed the validity of the equal varianceassumption for the ANOVA analyses of both specimens (P N 0.05). Thesignificant results of the ANOVA analysis are shown in the ‘Mean’ rowof Tables 2 and 3. The ANOVA analysis, between the first deformableregistration step (High Bending Energy) and the last rigid step (Iteration5), failed to show a statistically significant difference of the mean TREsfor both the hippocampus (P N 0.05, 95% Confidence Interval [CI] ofdifference −0.2666 to 1.653) and neocortex (P N 0.05, 95% CI −0.0147to 1.54). However, this test did demonstrate a significant decrease inTRE between (High Bending Energy) and the first rigid iteration
Table 3TRE values for hippocampal registration across iterations. H.B.E.: non-rigid with a High Bendingmeans registration step and Rigid Iteration 1. ‡: P ≤ 0.01 between means of registration step a
It. 1 It. 3 It. 5
Subj 1 2.07 ± 0.96 2.05 ± 0.97 2.05 ± 0.98Subj 2 2.81 ± 0.54 2.87 ± 0.63 2.74 ± 0.80Subj 3 2.13 ± 1.78 2.12 ± 1.74 2.09 ± 1.73Subj 4 2.19 ± 1.02 2.09 ± 1.02 2.07 ± 1.21Subj 5 2.05 ± 0.84 1.86 ± 0.85 1.78 ± 0.95Mean 2.25 ± 1.10 2.18 ± 1.11 2.15 ± 1.14
(Iteration 1) for the hippocampus and neocortex respectively(P ≤ 0.01, 95% CI 0.143 to 1.69 and P ≤ 0.05, 95% CI −0.2040 to1.715). In comparison, decreasing the bending energy weight penaltyproduced significantly lower mean TRE than the final rigid iterationstep (Iteration 5) for the hippocampus and neocortex respectively(P ≤ 0.001, 95% CI 0.449 to 2.00 and P ≤ 0.01, 95% CI 0.3516 to 2.271).The proposed masked non-rigid scheme (Masked NR) had as wellsignificantly lower TRE (P ≤ 0.0001) than Iteration 5 for both the hippo-campus (95% CI 0.590 to 2.14 mm) and neocortex (95% CI 0.5143–2.434).
Discussion
In this article, we have described a method to reliably registerex-vivo MRI and sparsely sliced histology slides of neocortex and hippo-campus specimens. Our protocol is a landmark free algorithm that pro-duced sub-millimeter accuracy for hippocampal registration and closeto 1-mm of error for temporal lobe neocortical registration. CorrelatingMRI with histopathology is imperative in the validation of new imagingsequences, since verification of pathological anomalies underlyingsignal changes is needed to enable these sequences to ultimately gainclinical acceptance. The intrinsic higher resolution of ex-vivoMR imagesprovides a superior opportunity to further examine the correlationbetween MRI and histology. By addressing many challenges of thein-vivoMRI to histology registration, our protocol leaves singlemodalityregistration between specimen and preoperative MRI scans as theremaining step. In addition, ex-vivo to MRI registration can be used tovalidate specimen imaging, as has been shown by Madabhushi et al.(2005) in prostate ex-vivo imaging examples.While using an intermedi-ate ex-vivo registration interrupts the clinicalflow for specimen imaging,our algorithm requires ~9 min ± 37 s on average (for the automatediterative 3D/2D rigid approach plus the non-rigid steps) to register100 μm coronally sliced (anterior to posterior) histology slices toex-vivo MRI images.
The vast majority of previous attempts at registeringMRI and histol-ogy using a combination of 2D and 3D transformations, or alternatingbetween 2D and 3D registration, were performed with a reconstructed3D histology volume from serially sectioned brain specimens atb700 μmm (Bardinet et al., 2002; Chakravarty et al., 2006; Dauguetet al., 2007; Humm et al., 2003; Lebenberg et al., 2010). This technique,
Energy penalty, L.B.E.: non-rigid with a Low Bending Energy penalty. †: P ≤ 0.01 betweennd Rigid Iteration 5.
H.B.E. L.B.E. Masked N.R.
1.35 ± 0.85 0.88 ± 0.67 0.73 ± 0.852.52 ± 0.94 1.42 ± 0.94 1.03 ± 0.941.30 ± 0.78 0.73 ± 0.42 0.62 ± 0.401.01 ± 0.54 0.68 ± 0.37 0.68 ± 0.421.15 ± 0.68 0.75 ± 0.43 0.72 ± 0.451.46 ± 0.77† 0.89 ± 0.61‡ 0.76 ± 0.66‡
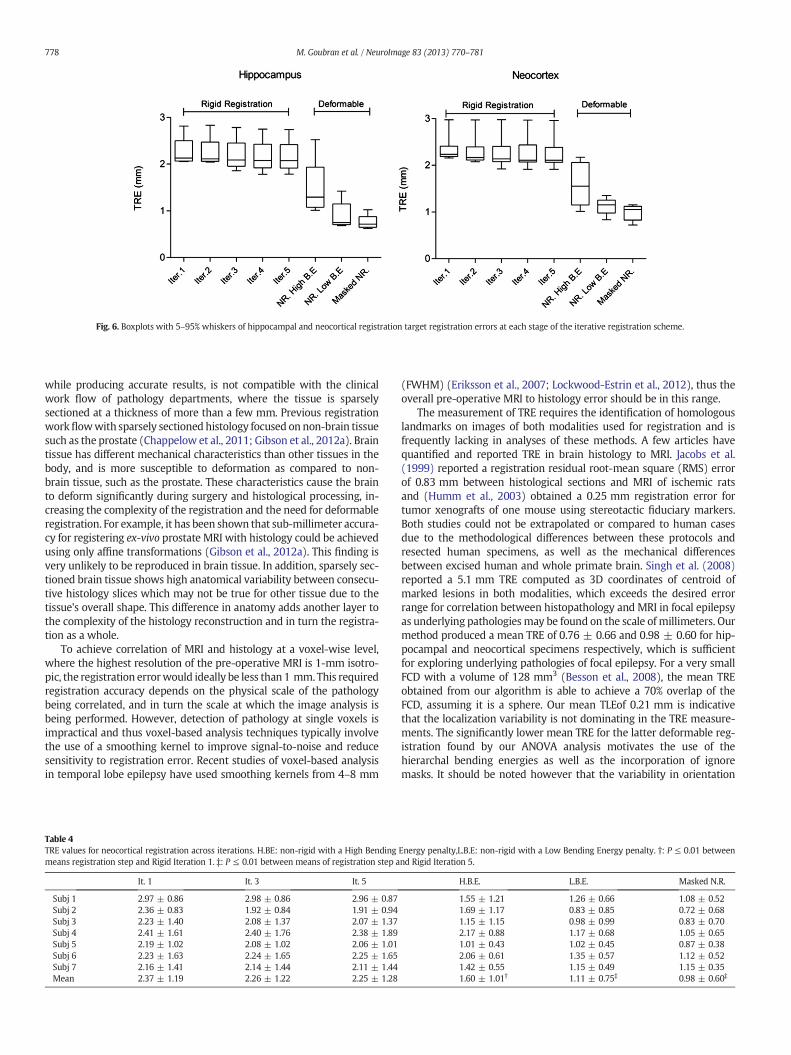
Fig. 6. Boxplots with 5–95% whiskers of hippocampal and neocortical registration target registration errors at each stage of the iterative registration scheme.
778 M. Goubran et al. / NeuroImage 83 (2013) 770–781
while producing accurate results, is not compatible with the clinicalwork flow of pathology departments, where the tissue is sparselysectioned at a thickness of more than a few mm. Previous registrationworkflowwith sparsely sectioned histology focused on non-brain tissuesuch as the prostate (Chappelow et al., 2011; Gibson et al., 2012a). Braintissue has different mechanical characteristics than other tissues in thebody, and is more susceptible to deformation as compared to non-brain tissue, such as the prostate. These characteristics cause the brainto deform significantly during surgery and histological processing, in-creasing the complexity of the registration and the need for deformableregistration. For example, it has been shown that sub-millimeter accura-cy for registering ex-vivo prostate MRI with histology could be achievedusing only affine transformations (Gibson et al., 2012a). This finding isvery unlikely to be reproduced in brain tissue. In addition, sparsely sec-tioned brain tissue shows high anatomical variability between consecu-tive histology slices which may not be true for other tissue due to thetissue's overall shape. This difference in anatomy adds another layer tothe complexity of the histology reconstruction and in turn the registra-tion as a whole.
To achieve correlation of MRI and histology at a voxel-wise level,where the highest resolution of the pre-operative MRI is 1-mm isotro-pic, the registration errorwould ideally be less than 1 mm. This requiredregistration accuracy depends on the physical scale of the pathologybeing correlated, and in turn the scale at which the image analysis isbeing performed. However, detection of pathology at single voxels isimpractical and thus voxel-based analysis techniques typically involvethe use of a smoothing kernel to improve signal-to-noise and reducesensitivity to registration error. Recent studies of voxel-based analysisin temporal lobe epilepsy have used smoothing kernels from 4–8 mm
Table 4TRE values for neocortical registration across iterations. H.BE: non-rigid with a High Bendingmeans registration step and Rigid Iteration 1. ‡: P ≤ 0.01 between means of registration step a
It. 1 It. 3 It. 5
Subj 1 2.97 ± 0.86 2.98 ± 0.86 2.96 ± 0.87Subj 2 2.36 ± 0.83 1.92 ± 0.84 1.91 ± 0.94Subj 3 2.23 ± 1.40 2.08 ± 1.37 2.07 ± 1.37Subj 4 2.41 ± 1.61 2.40 ± 1.76 2.38 ± 1.89Subj 5 2.19 ± 1.02 2.08 ± 1.02 2.06 ± 1.01Subj 6 2.23 ± 1.63 2.24 ± 1.65 2.25 ± 1.65Subj 7 2.16 ± 1.41 2.14 ± 1.44 2.11 ± 1.44Mean 2.37 ± 1.19 2.26 ± 1.22 2.25 ± 1.28
(FWHM) (Eriksson et al., 2007; Lockwood-Estrin et al., 2012), thus theoverall pre-operative MRI to histology error should be in this range.
The measurement of TRE requires the identification of homologouslandmarks on images of both modalities used for registration and isfrequently lacking in analyses of these methods. A few articles havequantified and reported TRE in brain histology to MRI. Jacobs et al.(1999) reported a registration residual root-mean square (RMS) errorof 0.83 mm between histological sections and MRI of ischemic ratsand (Humm et al., 2003) obtained a 0.25 mm registration error fortumor xenografts of one mouse using stereotactic fiduciary markers.Both studies could not be extrapolated or compared to human casesdue to the methodological differences between these protocols andresected human specimens, as well as the mechanical differencesbetween excised human and whole primate brain. Singh et al. (2008)reported a 5.1 mm TRE computed as 3D coordinates of centroid ofmarked lesions in both modalities, which exceeds the desired errorrange for correlation between histopathology and MRI in focal epilepsyas underlying pathologiesmay be found on the scale of millimeters. Ourmethod produced a mean TRE of 0.76 ± 0.66 and 0.98 ± 0.60 for hip-pocampal and neocortical specimens respectively, which is sufficientfor exploring underlying pathologies of focal epilepsy. For a very smallFCD with a volume of 128 mm3 (Besson et al., 2008), the mean TREobtained from our algorithm is able to achieve a 70% overlap of theFCD, assuming it is a sphere. Our mean TLEof 0.21 mm is indicativethat the localization variability is not dominating in the TRE measure-ments. The significantly lower mean TRE for the latter deformable reg-istration found by our ANOVA analysis motivates the use of thehierarchal bending energies as well as the incorporation of ignoremasks. It should be noted however that the variability in orientation
Energy penalty,L.B.E: non-rigid with a Low Bending Energy penalty. †: P ≤ 0.01 betweennd Rigid Iteration 5.
H.B.E. L.B.E. Masked N.R.
1.55 ± 1.21 1.26 ± 0.66 1.08 ± 0.521.69 ± 1.17 0.83 ± 0.85 0.72 ± 0.681.15 ± 1.15 0.98 ± 0.99 0.83 ± 0.702.17 ± 0.88 1.17 ± 0.68 1.05 ± 0.651.01 ± 0.43 1.02 ± 0.45 0.87 ± 0.382.06 ± 0.61 1.35 ± 0.57 1.12 ± 0.521.42 ± 0.55 1.15 ± 0.49 1.15 ± 0.351.60 ± 1.01† 1.11 ± 0.75‡ 0.98 ± 0.60‡
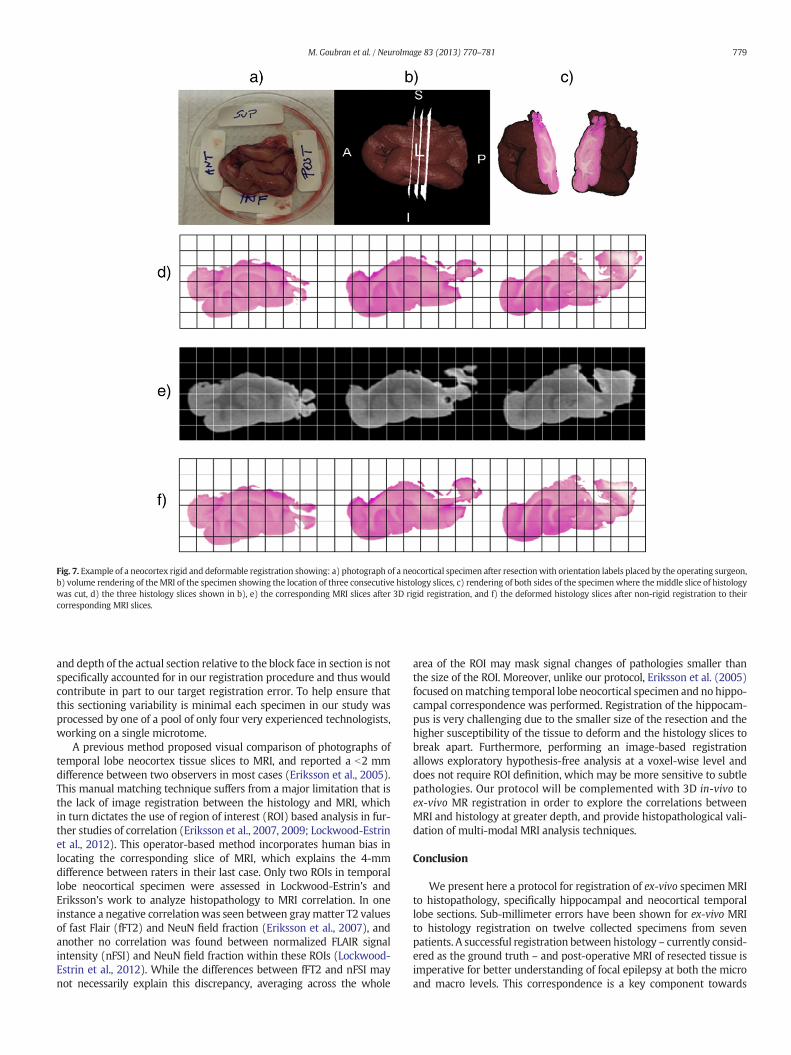
Fig. 7. Example of a neocortex rigid and deformable registration showing: a) photograph of a neocortical specimen after resectionwith orientation labels placed by the operating surgeon,b) volume rendering of theMRI of the specimen showing the location of three consecutive histology slices, c) rendering of both sides of the specimenwhere themiddle slice of histologywas cut, d) the three histology slices shown in b), e) the corresponding MRI slices after 3D rigid registration, and f) the deformed histology slices after non-rigid registration to theircorresponding MRI slices.
779M. Goubran et al. / NeuroImage 83 (2013) 770–781
and depth of the actual section relative to the block face in section is notspecifically accounted for in our registration procedure and thus wouldcontribute in part to our target registration error. To help ensure thatthis sectioning variability is minimal each specimen in our study wasprocessed by one of a pool of only four very experienced technologists,working on a single microtome.
A previous method proposed visual comparison of photographs oftemporal lobe neocortex tissue slices to MRI, and reported a b2 mmdifference between two observers in most cases (Eriksson et al., 2005).This manual matching technique suffers from a major limitation that isthe lack of image registration between the histology and MRI, whichin turn dictates the use of region of interest (ROI) based analysis in fur-ther studies of correlation (Eriksson et al., 2007, 2009; Lockwood-Estrinet al., 2012). This operator-based method incorporates human bias inlocating the corresponding slice of MRI, which explains the 4-mmdifference between raters in their last case. Only two ROIs in temporallobe neocortical specimen were assessed in Lockwood-Estrin's andEriksson's work to analyze histopathology to MRI correlation. In oneinstance a negative correlationwas seen between graymatter T2 valuesof fast Flair (fFT2) and NeuN field fraction (Eriksson et al., 2007), andanother no correlation was found between normalized FLAIR signalintensity (nFSI) and NeuN field fraction within these ROIs (Lockwood-Estrin et al., 2012). While the differences between fFT2 and nFSI maynot necessarily explain this discrepancy, averaging across the whole
area of the ROI may mask signal changes of pathologies smaller thanthe size of the ROI. Moreover, unlike our protocol, Eriksson et al. (2005)focused onmatching temporal lobe neocortical specimen and no hippo-campal correspondence was performed. Registration of the hippocam-pus is very challenging due to the smaller size of the resection and thehigher susceptibility of the tissue to deform and the histology slices tobreak apart. Furthermore, performing an image-based registrationallows exploratory hypothesis-free analysis at a voxel-wise level anddoes not require ROI definition, which may be more sensitive to subtlepathologies. Our protocol will be complemented with 3D in-vivo toex-vivo MR registration in order to explore the correlations betweenMRI and histology at greater depth, and provide histopathological vali-dation of multi-modal MRI analysis techniques.
Conclusion
We present here a protocol for registration of ex-vivo specimen MRIto histopathology, specifically hippocampal and neocortical temporallobe sections. Sub-millimeter errors have been shown for ex-vivo MRIto histology registration on twelve collected specimens from sevenpatients. A successful registration between histology – currently consid-ered as the ground truth – and post-operative MRI of resected tissue isimperative for better understanding of focal epilepsy at both the microand macro levels. This correspondence is a key component towards

Fig. 8. Example of a hippocampal rigid and deformable registration showing: a) photograph of a hippocampus before grossing, b) volume rendering of the MRI of the hippocampusdemonstrating the location of a histology slice through the specimen, c) rendering of both sides of the hippocampus where the histology slice was cut, and d) three orthogonal viewsof the hippocampal MRI (left to right: coronal, sagittal, axial). The bottom row depicts: e) a coronal view of the same histological slice with a registration mask over a breakage regionshown in blue, f) a checkerboard image showing the MRI and histology before non-rigid registration, and g) a checkerboard image showing the MRI and non-rigidly deformed histologyslice.
780 M. Goubran et al. / NeuroImage 83 (2013) 770–781
achieving MRI and histology correlation by bringing together informa-tion from both domains.
Acknowledgments
The authors would like to thank Dr. Seyed Mirsattari, Dr. RobertHammond and Dr. Andrew Parrent for their assistance and supportthroughout the study. This project is funded by the Canadian Instituteof Health Research (CIHR) grant MOP 184807 and the Canada Founda-tion for Innovation (CFI) grant 20994. MG is supported by the NSERCCreate Grant CAMI award at Western University. AK is supported bythe Canadian Institute of Health Research (CIHR) Fellowship.
References
Bardinet, E., Ourselin, S., Dormont, D., Malandain, G., Tandé, D., Parain, K., Ayache, N.,Yelnik, J., 2002. Co-registration of histological, optical and MR data of the humanbrain. Med. Image Comput. Comput. Assist. Interv. 548–555.
Bernasconi, A., Bernasconi, N., Caramanos, Z., Reutens, D.C., Andermann, F., Dubeau, F.,Tampieri, D., Pike, B.G., Arnold, D.L., 2000. T2 relaxometry can lateralize mesialtemporal lobe epilepsy in patients with normal MRI. NeuroImage 12 (6), 739–746.
Bernasconi, N., Duchesne, S., Janke, A., Lerch, J., Collins, D.L., Bernasconi, A., 2004. Whole-brain voxel-based statistical analysis of gray matter and white matter in temporallobe epilepsy. NeuroImage 23 (2), 717–723.
Bernhardt, B.C., Worsley, K.J., Kim, H., Evans, A.C., Bernasconi, A., Bernasconi, N., 2009.Longitudinal and cross-sectional analysis of atrophy in pharmacoresistant temporallobe epilepsy. Neurology 72 (20), 1747–1754.
Besson, P., Andermann, F., Dubeau, F., Bernasconi, A., 2008. Small focal cortical dysplasialesions are located at the bottom of a deep sulcus. Brain 131 (Pt 12), 3246–3255.
Breen, M.S., Lancaster, T.L., Wilson, D.L., 2005. Correcting spatial distortion in histologicalimages. Comput. Med. Imaging Graph. 29 (6), 405–417.
Ceritoglu, C., Wang, L., Selemon, L.D., Csernansky, J.G., Miller, M.I., Ratnanather, J.T., 2010.Large deformation diffeomorphic metric mapping registration of reconstructed3D histological section images and in vivo MR images. Front. Hum. Neurosci. 4,43.
Chakravarty,M.M., Bertrand, G., Hodge, C.P., Sadikot, A.F., Collins, D.L., 2006. The creation ofa brain atlas for image guided neurosurgery using serial histological data. NeuroImage30 (2), 359–376.
Chappelow, J., Bloch, B.N., Rofsky, N., Genega, E., Lenkinski, R., DeWolf,W.,Madabhushi, A.,2011. Elastic registration ofmultimodal prostateMRI and histology via multiattributecombined mutual information. Med. Phys. 38 (4), 2005.

781M. Goubran et al. / NeuroImage 83 (2013) 770–781
Dauguet, J., Delzescaux, T., Condé, F., Mangin, J.-F., Ayache, N., Hantraye, P., Frouin, V.,2007. Three-dimensional reconstruction of stained histological slices and 3D non-linear registration with in-vivo MRI for whole baboon brain. J. Neurosci. Methods164 (1), 191–204.
de Tisi, J., Bell, G.S., Peacock, J.L., McEvoy, A.W., Harkness, W.F.J., Sander, J.W., Duncan, J.S.,2011. The long-term outcome of adult epilepsy surgery, patterns of seizure remission,and relapse: a cohort study. Lancet 378 (9800), 1388–1395.
Engel, J., 1998. Etiology as a risk factor for medically refractory epilepsy: a case for earlysurgical intervention. Neurology 51 (5), 1243–1244.
Engel, J., Levesque, M.F., Shields, W.D., 1992. Surgical treatment of the epilepsies:presurgical evaluation. Clin. Neurosurg. 38, 514–534.
Eriksson, S.H., Free, S.L., Thom, M., Harkness, W., Sisodiya, S.M., Duncan, J.S., 2005. Reliableregistration of preoperative MRI with histopathology after temporal lobe resections.Epilepsia 46 (10), 1646–1653.
Eriksson, S.H., Free, S.L., Thom, M., Martinian, L., Symms, M.R., Salmenpera, T.M., McEvoy,A.W., Harkness, W., Duncan, J.S., Sisodiya, S.M., 2007. Correlation of quantitative MRIand neuropathology in epilepsy surgical resection specimens—T2 correlates withneuronal tissue in gray matter. NeuroImage 37 (1), 48–55.
Eriksson, S.H., Free, S.L., Thom, M., Symms, M.R., Martinian, L., Duncan, J.S., Sisodiya, S.M.,2009. Quantitative grey matter histological measures do not correlate with greymatter probability values from in vivoMRI in the temporal lobe. J. Neurosci. Methods181 (1), 111–118.
Fish, D.R., Smith, S.J., Quesney, L.F., Andermann, F., Rasmussen, T., 1993. Surgical treatmentof children with medically intractable frontal or temporal lobe epilepsy: results andhighlights of 40 years' experience. Epilepsia 34 (2), 244–247.
Fitzpatrick, J.M., West, J.B., Maurer, C.R., 1998. Predicting error in rigid-body point-basedregistration. IEEE Trans. Med. Imaging 17 (5), 694–702.
Gibson, E., Crukley, C., Gaed, M., Gómez, J.A., Moussa, M., Chin, J.L., Bauman, G.S., Fenster,A., Ward, A.D., 2012a. Registration of prostate histology images to ex vivo MR imagesvia strand-shaped fiducials. J. Magn. Reson. Imaging 1402–1412.
Gibson, E., Gómez, J., Moussa, M., Crukley, C., Bauman, G., Fenster, A., Ward, A., 2012b. 3dreconstruction of prostate histologybased onquantified tissue cutting anddeformationparameters. Society of Photo-Optical Instrumentation Engineers (SPIE) ConferenceSeries, vol. 8317, p. 22.
Goubran, M., Khan, A.R., Crukley, C., Buchanan, S., Santyr, B., deRibaupierre, S., Peters, T.M.,2012. Robust registration of sparsely sectioned histology to ex-vivo MRI of temporallobe resections. 8314, 83141V.
Howe, K.L., Dimitri, D., Heyn, C., Kiehl, T.-R., Mikulis, D., Valiante, T., 2010. Histologicallyconfirmed hippocampal structural features revealed by 3 T MR imaging: potential toincrease diagnostic specificity of mesial temporal sclerosis. AJNR Am. J. Neuroradiol.31 (9), 1682–1689.
Humm, J.L., Ballon, D., Hu, Y.C., Ruan, S., Chui, C., Tulipano, P.K., Erdi, A., Koutcher, J.,Zakian, K., Urano, M., Zanzonico, P., Mattis, C., Dyke, J., Chen, Y., Harrington, P.,O'Donoghue, J.A., Ling, C.C., 2003. A stereotactic method for the three-dimensional
registration of multi-modality biologic images in animals: NMR, PET, histology, andautoradiography. Med. Phys. 30 (9), 2303–2314.
Jacobs, M.A., Windham, J.P., Soltanian-Zadeh, H., Peck, D.J., Knight, R.A., 1999. Registrationand warping of magnetic resonance images to histological sections. Med. Phys. 26(8), 1568–1578.
Jenkinson, M., Smith, S., 2001. A global optimisation method for robust affine registrationof brain images. Med. Image Anal. 5 (2), 143–156.
Kim, T., Singh, M., Sungkarat, W., Zarow, C., Chui, H., 2000. Automatic registration of post-mortembrain slices toMRI reference volume. IEEE Trans. Nucl. Sci. 47 (4), 1607–1613.
Lebenberg, J., Hérard, A.S., Dubois, A., Dauguet, J., Frouin, V., Dhenain, M., Hantraye, P.,Delzescaux, T., 2010. Validation of MRI-based 3D digital atlas registration with histo-logical and autoradiographic volumes: an anatomofunctional transgenic mouse brainimaging study. NeuroImage 51 (3), 1037–1046.
Lockwood-Estrin, G., Thom, M., Focke, N.K., Symms, M.R., Martinian, L., Sisodiya, S.M.,Duncan, J.S., Eriksson, S.H., 2012. Correlating 3 T MRI and histopathology in patientsundergoing epilepsy surgery. J. Neurosci. Methods 205 (1), 182–189.
Madabhushi,A., Feldman,M.D.,Metaxas, D.N., Tomaszeweski, J., Chute, D., 2005. Automateddetection of prostatic adenocarcinoma from high-resolution ex vivo MRI. IEEE Trans.Med. Imaging 24 (12), 1611–1625.
Malandain, G., Bardinet, E., Nelissen, K., Vanduffel,W., 2004. Fusion of autoradiographswithanMR volume using 2-D and 3-D linear transformations. NeuroImage 23 (1), 111–127.
Meyer, C.R., Moffat, B.A., Kuszpit, K.K., Bland, P.L., Mckeever, P.E., Johnson, T.D., Chenevert,T.L., Rehemtulla, A., Ross, B.D., 2006. A methodology for registration of a histologicalslide and in vivo MRI volume based on optimizing mutual information. Mol. Imaging5 (1), 16–23.
Modat, M., Ridgway, G.R., Taylor, Z.A., Lehmann, M., Barnes, J., Hawkes, D.J., Fox, N.C.,Ourselin, S., 2010. Fast free-form deformation using graphics processing units.Comput. Methods Programs Biomed. 98 (3), 278–284.
Rueckert, D., Sonoda, L.I., Hayes, C., Hill, D.L., Leach, M.O., Hawkes, D.J., 1999. Nonrigidregistration using free-form deformations: application to breast MR images. IEEETrans. Med. Imaging 18 (8), 712–721.
Schmierer, K., Scaravilli, F., Barker, G.J., Gordon, R., MacManus, D.G., Miller, D.H., 2003.Stereotactic co-registration of magnetic resonance imaging and histopathology inpost-mortemmultiple sclerosis brain. Neuropathol. Appl. Neurobiol. 29 (6), 596–601.
Schormann, T., Dabringhaus, A., Zilles, K., 1995. Statistics of deformations in histology andapplication to improved alignment withMRI. IEEE Trans. Med. Imaging 14 (1), 25–35.
Singh, M., Rajagopalan, A., Kim, T.-S., Hwang, D., Chui, H., Zhang, X.-L., Lee, A.-Y., Zarow, C.,2008. Co-registration of in-vivo human MRI brain images to postmortem histologicalmicroscopic images. Int. J. Imaging Syst. Technol. 18 (5–6), 325–335.
Smith, S.J.M., 2005. EEG in the diagnosis, classification, and management of patients withepilepsy. J. Neurol. Neurosurg. Psychiatry 76 (Suppl. 2), ii2–ii7.
Yang, Z., Richards, K., Kurniawan, N.D., Petrou, S., Reutens, D.C., 2012. MRI-guided volumereconstruction of mouse brain from histological sections. J. Neurosci. Methods 211,210–217.





























