Embed Size (px)
Citation preview
Impact of Enhanced Metabolic Stability on In Vivo Performance of GalNAc-siRNA ConjugatesAuthors: Martin A. Maier, Donald J. Foster, Jayaprakash K. Nair, Husain Attarwala, Christopher R. Brown, Xuemei Zhang, Minggeng Gao, Ju Liu, Ramesh Indrakanti, Sally Schofield, Philip Kretschmer, Qianfan Wang, Sarfraz Shaikh, Casey Trapp, Mark K. Schlegel, Satya Kuchimanchi, Kun Qian, Alfica Sehgal, Krishna Aluri, Swati Gupta, Jennifer L.S. Willoughby, Julie A. Boshar, Klaus Charisse, Tracy Zimmermann, Kevin Fitzgerald, Kallanthottathil G. Rajeev, Muthiah Manoharan, Akin Akinc, Renta Hutabarat, Vasant Jadhav, Stuart Milstein
Affiliations: Alnylam Pharmaceuticals, Cambridge, MA 02142, USA
AbstractThe combination of a synthetic triantennary N-acetylagalactosamine (GalNAc) ligand to chemically modified siRNA has enabled asialoglycoprotein (ASGPR)-mediated targeted delivery of therapeutically active siRNAs to hepatocytes in vivo.1 This approach has become transformative for the delivery of RNAi therapeutics as well as other classes of investigational oligonucleotide therapeutics to the liver.2-4 However, even in the context of fully modified siRNAs, enhanced stabilization against nucleolytic degradation is essential for optimal in vivo performance of GalNAc-siRNA conjugates.5,6 Here we summarize a multi-pronged approach to further elucidate the relationship between siRNA chemistry and metabolic stability and to guide the development of advanced designs with improved potency and duration.7 This includes a screening paradigm for siRNA design optimization, which is based on a positional model for chemical modifications and relies on parallel screening across multiple sequences.8 These efforts led to the discovery of enhanced stability designs, which have fundamentally transformed the siRNA conjugate platform and helped to advance multiple RNAi therapeutics into pre-clinical and clinical development.9-11
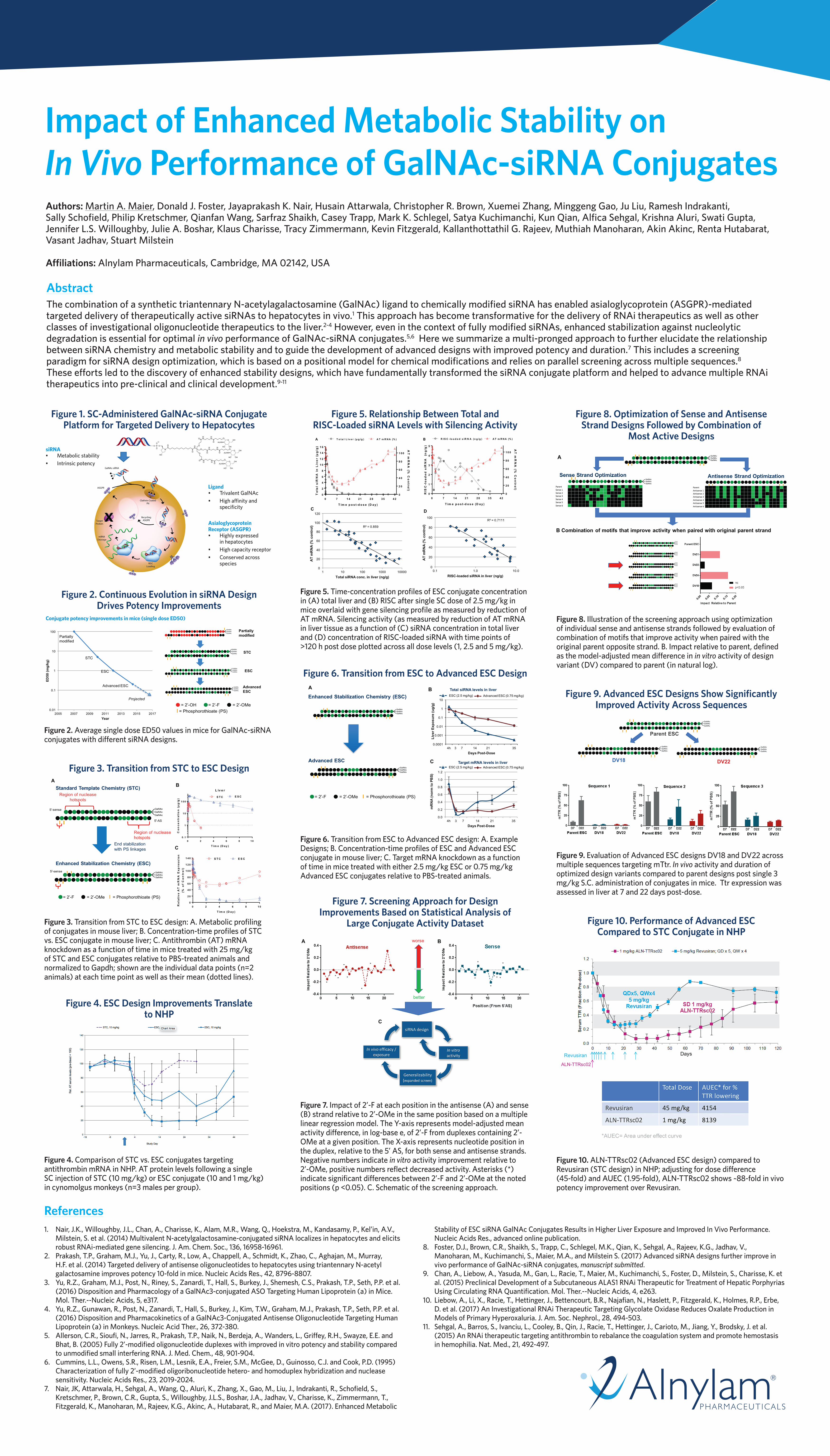
Figure 1. SC-Administered GalNAc-siRNA Conjugate Platform for Targeted Delivery to Hepatocytes
Figure 2. Continuous Evolution in siRNA Design Drives Potency Improvements
Figure 2. Average single dose ED50 values in mice for GalNAc-siRNA conjugates with different siRNA designs.
Figure 3. Transition from STC to ESC Design
Figure 3. Transition from STC to ESC design: A. Metabolic profiling of conjugates in mouse liver; B. Concentration-time profiles of STC vs. ESC conjugate in mouse liver; C. Antithrombin (AT) mRNA knockdown as a function of time in mice treated with 25 mg/kg of STC and ESC conjugates relative to PBS-treated animals and normalized to Gapdh; shown are the individual data points (n=2 animals) at each time point as well as their mean (dotted lines).
Figure 4. ESC Design Improvements Translate to NHP
Figure 4. Comparison of STC vs. ESC conjugates targeting antithrombin mRNA in NHP. AT protein levels following a single SC injection of STC (10 mg/kg) or ESC conjugate (10 and 1 mg/kg) in cynomolgus monkeys (n=3 males per group).
Figure 5. Relationship Between Total and RISC-Loaded siRNA Levels with Silencing Activity
Figure 5. Time-concentration profiles of ESC conjugate concentration in (A) total liver and (B) RISC after single SC dose of 2.5 mg/kg in mice overlaid with gene silencing profile as measured by reduction of AT mRNA. Silencing activity (as measured by reduction of AT mRNA in liver tissue as a function of (C) siRNA concentration in total liver and (D) concentration of RISC-loaded siRNA with time points of >120 h post dose plotted across all dose levels (1, 2.5 and 5 mg/kg).
Figure 6. Transition from ESC to Advanced ESC Design
Figure 6. Transition from ESC to Advanced ESC design: A. Example Designs; B. Concentration-time profiles of ESC and Advanced ESC conjugate in mouse liver; C. Target mRNA knockdown as a function of time in mice treated with either 2.5 mg/kg ESC or 0.75 mg/kg Advanced ESC conjugates relative to PBS-treated animals.
Figure 7. Screening Approach for Design Improvements Based on Statistical Analysis of
Large Conjugate Activity Dataset
Figure 7. Impact of 2’-F at each position in the antisense (A) and sense (B) strand relative to 2’-OMe in the same position based on a multiple linear regression model. The Y-axis represents model-adjusted mean activity difference, in log-base e, of 2’-F from duplexes containing 2’-OMe at a given position. The X-axis represents nucleotide position in the duplex, relative to the 5’ AS, for both sense and antisense strands. Negative numbers indicate in vitro activity improvement relative to 2’-OMe, positive numbers reflect decreased activity. Asterisks (*) indicate significant differences between 2’-F and 2’-OMe at the noted positions (p <0.05). C. Schematic of the screening approach.
Figure 8. Optimization of Sense and Antisense Strand Designs Followed by Combination of
Most Active Designs
Figure 8. Illustration of the screening approach using optimization of individual sense and antisense strands followed by evaluation of combination of motifs that improve activity when paired with the original parent opposite strand. B. Impact relative to parent, defined as the model-adjusted mean difference in in vitro activity of design variant (DV) compared to parent (in natural log).
Figure 9. Advanced ESC Designs Show Significantly Improved Activity Across Sequences
Figure 9. Evaluation of Advanced ESC designs DV18 and DV22 across multiple sequences targeting mTtr. In vivo activity and duration of optimized design variants compared to parent designs post single 3 mg/kg S.C. administration of conjugates in mice. Ttr expression was assessed in liver at 7 and 22 days post-dose.
Figure 10. Performance of Advanced ESC Compared to STC Conjugate in NHP
Figure 10. ALN-TTRsc02 (Advanced ESC design) compared to Revusiran (STC design) in NHP; adjusting for dose difference (45-fold) and AUEC (1.95-fold), ALN-TTRsc02 shows ~88-fold in vivo potency improvement over Revusiran.
1. Nair, J.K., Willoughby, J.L., Chan, A., Charisse, K., Alam, M.R., Wang, Q., Hoekstra, M., Kandasamy, P., Kel’in, A.V., Milstein, S. et al. (2014) Multivalent N-acetylgalactosamine-conjugated siRNA localizes in hepatocytes and elicits robust RNAi-mediated gene silencing. J. Am. Chem. Soc., 136, 16958-16961.
2. Prakash, T.P., Graham, M.J., Yu, J., Carty, R., Low, A., Chappell, A., Schmidt, K., Zhao, C., Aghajan, M., Murray, H.F. et al. (2014) Targeted delivery of antisense oligonucleotides to hepatocytes using triantennary N-acetyl galactosamine improves potency 10-fold in mice. Nucleic Acids Res., 42, 8796-8807.
3. Yu, R.Z., Graham, M.J., Post, N., Riney, S., Zanardi, T., Hall, S., Burkey, J., Shemesh, C.S., Prakash, T.P., Seth, P.P. et al. (2016) Disposition and Pharmacology of a GalNAc3-conjugated ASO Targeting Human Lipoprotein (a) in Mice. Mol. Ther.--Nucleic Acids, 5, e317.
4. Yu, R.Z., Gunawan, R., Post, N., Zanardi, T., Hall, S., Burkey, J., Kim, T.W., Graham, M.J., Prakash, T.P., Seth, P.P. et al. (2016) Disposition and Pharmacokinetics of a GalNAc3-Conjugated Antisense Oligonucleotide Targeting Human Lipoprotein (a) in Monkeys. Nucleic Acid Ther., 26, 372-380.
5. Allerson, C.R., Sioufi, N., Jarres, R., Prakash, T.P., Naik, N., Berdeja, A., Wanders, L., Griffey, R.H., Swayze, E.E. and Bhat, B. (2005) Fully 2’-modified oligonucleotide duplexes with improved in vitro potency and stability compared to unmodified small interfering RNA. J. Med. Chem., 48, 901-904.
6. Cummins, L.L., Owens, S.R., Risen, L.M., Lesnik, E.A., Freier, S.M., McGee, D., Guinosso, C.J. and Cook, P.D. (1995) Characterization of fully 2’-modified oligoribonucleotide hetero- and homoduplex hybridization and nuclease sensitivity. Nucleic Acids Res., 23, 2019-2024.
7. Nair, JK, Attarwala, H., Sehgal, A., Wang, Q., Aluri, K., Zhang, X., Gao, M., Liu, J., Indrakanti, R., Schofield, S., Kretschmer, P., Brown, C.R., Gupta, S., Willoughby, J.L.S., Boshar, J.A., Jadhav, V., Charisse, K., Zimmermann, T., Fitzgerald, K., Manoharan, M., Rajeev, K.G., Akinc, A., Hutabarat, R., and Maier, M.A. (2017). Enhanced Metabolic
Stability of ESC siRNA GalNAc Conjugates Results in Higher Liver Exposure and Improved In Vivo Performance. Nucleic Acids Res., advanced online publication.
8. Foster, D.J., Brown, C.R., Shaikh, S., Trapp, C., Schlegel, M.K., Qian, K., Sehgal, A., Rajeev, K.G., Jadhav, V., Manoharan, M., Kuchimanchi, S., Maier, M.A., and Milstein S. (2017) Advanced siRNA designs further improve in vivo performance of GalNAc-siRNA conjugates, manuscript submitted.
9. Chan, A., Liebow, A., Yasuda, M., Gan, L., Racie, T., Maier, M., Kuchimanchi, S., Foster, D., Milstein, S., Charisse, K. et al. (2015) Preclinical Development of a Subcutaneous ALAS1 RNAi Therapeutic for Treatment of Hepatic Porphyrias Using Circulating RNA Quantification. Mol. Ther.--Nucleic Acids, 4, e263.
10. Liebow, A., Li, X., Racie, T., Hettinger, J., Bettencourt, B.R., Najafian, N., Haslett, P., Fitzgerald, K., Holmes, R.P., Erbe, D. et al. (2017) An Investigational RNAi Therapeutic Targeting Glycolate Oxidase Reduces Oxalate Production in Models of Primary Hyperoxaluria. J. Am. Soc. Nephrol., 28, 494-503.
11. Sehgal, A., Barros, S., Ivanciu, L., Cooley, B., Qin, J., Racie, T., Hettinger, J., Carioto, M., Jiang, Y., Brodsky, J. et al. (2015) An RNAi therapeutic targeting antithrombin to rebalance the coagulation system and promote hemostasis in hemophilia. Nat. Med., 21, 492-497.
References
siRNA• Metabolic stability• Intrinsic potency
Ligand• Trivalent GalNAc • High affinity and
specificity
Asialoglycoprotein Receptor (ASGPR)• Highly expressed
in hepatocytes• High capacity receptor• Conserved across
species
Conjugate potency improvements in mice (single dose ED50)





























