Embed Size (px)
Citation preview

1
UMEÅ UNIVERSITY MEDICAL DISSERTATIONS
New Series No. 892-ISSN 0346-6612-ISBN 91-7305-652-9
From the Department of Clinical Sciences, Otorhinolaryngology, Umeå University, Sweden
Impaired reparative processes in particular related to hyaluronan in various cutaneous disorders
- A structural analysis
Ulf Bertheim
Umeå 2004

2
Cover illustration Colour light micrograph of histological section from normal skin stained with HABP (brown), demonstrating the distribution of hyaluronan (HA). (Photo; Dr Ulf Bertheim) The PD is intensely stained with an intense, uniform appearance of HA in the BM zone. The epidermis is moderately stained and only weak staining is seen in the RD
Copyright © 2004 by Ulf Bertheim
ISBN 91-7305-652-9
Printed in Sweden by Solfjädern Offset AB
Umeå 2004

3
To my children Jane and Andre’, for your love, patience and inspiration

4
CONTENTS CONTENTS 4 ABSTRACT 5 ABBREVIATIONS 6 ORIGINAL PAPERS 7 INTRODUCTION 8 The Skin 9 Hyaluronan 12 Wound Healing Process 15
Pathologic Scar Tissue Formation 18 Cutaneous Changes In Diabetic Mellitus 19 Basal Cell Carcinomas 20 AIMS OF THE STUDY 22 MATERIAL AND METHODS 23 Scar Tissues 23
Diabetes Mellitus 23 Basal Cell Carcinomas 23 Paraffin Embedded Material 25 Plastic Embedded Material 25 Histochemical Staining for HA 26 Immunohistochemical Analysis 26 Quantitative Analysis of HA in Tissue Samples 27 Statistical Methods 27 Examination 28
RESULTS 29
The Architecture of Normal Skin 29 Scar Tissues 30 Skin in Diabetic Patients 31 Different Types of BCC Tumours 32 DISCUSSION 33 HA in Wounded Skin 34 HA in Skin in Diabetes Mellitus 37
HA in Tumour Progression 39 Clinical Considerations 41 CONCLUSIONS 42 SAMMANFATTNING PÅ SVENSKA 43 ACKNOWLEDGEMENTS 45 REFERENCES 46 PAPERS I-IV

5
ABSTRACT
Impaired reparative processes in particular related to hyaluronan in various cutaneous disorders - A structural analysis
Ulf Bertheim
Cutaneous reparative processes, including wound healing, are highly developed procedures in which a chain of actions occurs to reconstitute the function of the wounded tissue. To prevent a delayed or excessive reparative process it is important to understand how this procedure develops and is maintained. One of the major extracellular matrix components of the skin is the glycosaminoglycan hyaluronan (HA). HA contributes to an extracellular environment, which is permissive for cell motility and proliferation, features that may account for HA’s unique properties observed in scarless foetal wound healing. The molecule is found at high concentration whenever proliferation, regeneration and repair of tissue occur. The aims of the present studies were to analyse the distribution of HA and to investigate its possible role in various cutaneous conditions associated with an impaired reparative process like in scar tissue formation in healing wounds, changed skin characteristics in diabetes mellitus and proliferating activity in basal cell carcinomas. Tissue biopsies were obtained from healthy human skin, type-I diabetic skin and various scar tissues. The samples were analysed in the light microscope with a hyaluronan-binding-probe, antibodies for collagen I, III, PCNA and Ki-67. Ultrastructural analyses were performed on the same tissue samples. In normal skin HA was present mainly in the papillary dermis. In epidermis HA was located in between the keratinocytes in the spinous layer. In the different scar tissues the localization of HA varied, with an HA distribution in mature scar type resembling that in normal skin. In keloids the papillary dermis lacked HA, but the thickened epidermis contained more HA than the other scar types. Ultrastructural studies of keloids revealed an altered collagen structure in the dermal layers, with an abundance of thin collagen fibers in the reticular dermis and thicker collagen fibers in the papillary dermis. Furthermore, the keloids displayed epidermal changes, which involved the basement membrane (BM), exhibiting fewer hemidesmosomes, and an altered shape of desmosomes in the entire enlarged spinous layer. These alterations in epidermis are suggested to influence the hydrodynamic and cell regulatory properties of the wounded skin. In diabetic patients, a reduced HA staining in the basement membrane zone was seen. The staining intensity of HA correlated to the physical properties of the skin reflected by their grades of limited joint mobility (LJM). Furthermore, the HA staining correlated with serum concentration of the HbA1c. In basal cell carcinomas (BCC), HA occurred predominantly in the tumour stroma. The distribution was most intense in the highly developed superficial BCC type, and resembled that of the papillary dermis of normal skin. In contrast, in the infiltrative BCC type, the tumour stroma stained weakly in the infiltrative part of the tumour. Moreover, the surrounding dermal layer was deranged and devoid of HA. The findings suggest that the tumour stroma in superficial BCC causes a slow, well-regulated cell growth in which the tumour cells do not substantially disturb the normal skin function. In the infiltrative BCC type, the tumour cells cause a disintegration of the tumour stroma as well as the normal surrounding dermis, which permits further spreading of the tumour. In fact, the behaviour of the infiltrative BCC tumour, growing beyond its boundaries, resembles that of the keloid. The mapping of the distribution of HA could be a useful tool for prognostic information, for evaluating the degree of progress and for deciding the choice of treatment in various diseases of the skin. In skin malignancies such as BCC it can be used to determine the radicality at the surgical excision of the tumour. Keywords: Hyaluronan, scar tissue, diabetes mellitus, basal cell carcinoma, skin, wound healing

6
ABBREVIATIONS BCC Basal cell carcinoma BM Basement membrane BMZ Basement membrane zone ECM Extracellular matrix GAG Glycosaminoglycan HA Hyaluronan, hyaluronic acid HABP Hyaluronan-binding protein HYA Hyaluronan, hyaluronic acid IDDM Insulin dependent diabetes mellitus LJM Limited joint mobility NIDDM Non insulin dependent diabetes mellitus PD Papillary dermis PBS Phosphate-buffered saline PCNA Proliferating cell nuclear antigen RD Reticular dermis

7
ORIGINAL PAPERS This thesis is based upon the following publications and manuscripts, which will be referred to in the text by their respective roman numerals: I Bertheim U, Hellström S. 1994. The distribution of hyaluronan in human skin
and mature, hypertrophic and keloid scars. British Journal of Plastic Surgery 47: 483-489.
II Bertheim U, Engström-Laurent A, Hofer P-Å, Hallgren P, Asplund J, Hellström
S. 2002. Loss of hyaluronan in the basement membrane zone of the skin correlates to the degree of stiff hands in diabetes patients. Acta Derm Venereol 82: 320-334.
III Bertheim U, Hofer P-Å, Engström-Laurent A, Hellström S. 2003. The stromal
reaction in basal cell carcinomas. A prerequisite for tumour progression and treatment strategy. British Journal of Plastic Surgery (in press).
IV Bertheim U, Engström-Laurent A, Hellström S. 2004. Structural changes
correlated to hyaluronan are associated with an excessive collagen formation in keloids. (in manuscript).

8
INTRODUCTION
The basics of plastic surgery rest on our understanding of the structure and function of normal
skin and the management of its various pathological reactions as in various malignancies and
wound processes. In spite of recent advances in our knowledge of the fundamental processes
of wound healing, awareness of the factors involved in the development of chronic wounds
remains limited (Falanga V et al., 1995). Progress in the treatment of chronic wounds will
require a better understanding of their pathogenesis and failure to heal. Treatment principles
have to be focused on investigations of and guidance concerning the connections between
stimulatory and inhibitory wound healing mechanisms (Diegelmann and Evans., 2004). It is
important to detect a pathologic repair process, in order to reduce the risk of infection, as well
as the pain and discomfort patients may suffer (Moseley et al., 2004). Under normal
circumstances the complex wound healing mechanism acts protectively, but when it fails
healing may be protracted and resulting fibrosis may produce an abnormal disabling scar
(Hunt et al., 1978).
By using various reconstructive techniques, pathological scar tissue can be treated but its
function can rarely be completely restored to normal. The objective for the plastic surgeon is
to guide the repair process toward a scar that resembles normal skin in its structure and
function. An optimal reparative process, includes restoration not only of the cellular
components in the dermis and epidermis but also the epidermal-dermal junction, the basement
membrane (BM), the constituents of the extracellular matrix (ECM), and all the
interconnecting proteins and enzymes. Interest in skin matrix components, including the
glycosaminoglycan (GAG) hyaluronan (HA) and the collagens is increasing, and their various
roles in the developing and restorative processes have been postulated (Gailit and Clark.,
1994; Hellström et al., 1991; Grinnel., 1984). The present studies have been focused on

9
investigating the role and influence of these molecules in the wound healing process and how
they are modified in the skin in the presence of metabolic disorders and malignant processes.
Furthermore, this thesis highlights the importance of early signs of any structural disturbance
that alters the tissue architecture and thereby also the possibility of an optimal reparative
process.
The Skin
Fig 1. Schematic drawing of the skin; Epidermis (Överhuden), Dermis (Läderhuden).
The skin, the largest organ of the human body, constituting the body’s outer covering,
provides protection against physical, chemical or microbial insults from the environment. It
acts as a defensive barrier having a reparative capacity (Wood et al., 1992). In many internal
disorders, different reactions in internal organs and structures as well as various types of
cutaneous manifestations can be observed (Lever and Lever., 1990). The skin consists of three
compartments: epidermis, dermis and subcutis. The subcutaneous tissue is not dealt with in
the present thesis. The superficial epidermal layer is multilayered, consisting mainly of
keratinocytes in different stages of maturation. The epidermis also contains melanocytes,

10
Merkel cells, Langerhans cells and lymphocytes. It is separated from the dermis by a
basement membrane (BM). The deepest epidermal layer, stratum basale, consists of a single
layer of keratinocytes. As the latter develop, they move towards the surface. Above the basal
layer the keratinocytes form part of the spinous layer, characterized by an abundance of
interconnective adhesive proteins called desmosomes. The keratinocytes of the superficial
layer, stratum granulosum, contain keratohyalin granules. At the surface, the keratinocytes
become flattened, and lose their nuclei and intracellular organelles, and so adopt the
characteristics of stratum corneum. The keratinocytes play an important role in the function of
an active skin barrier through their interactions with the immune and inflammatory systems
(Eckert.,1989; Wood et al., 1992). Nevertheless, a normal architecture–and hence normal
functioning of the epidermis– relies on an undisturbed intercellular environment that not only
holds the cells together, but also permits intercellular exchange of gases and nutrients. This
cohesion is effected mainly by interconnecting desmosomes, which are adhesive proteins
consisting of two symmetrical halves, each located in one of two adjoining cells. On the
cytoplasmic side of the plasma membrane is the desmosomal plaque, in association with the
keratin filaments (North et al., 1999). Evidently, desmosomes are not static structures, rather
they are dynamic units, whose composition and structure are critical for normal epidermal
function (Kitajima., 2002). Intra-epidermal and dermal-epidermal cohesion are essential for
the integrity of the skin (Moll and Moll., 1998)
Basement membrane (BM) is a thin layer, intersecting the connective tissue from the basal
keratinocytes of the epidermis. The BM constitutes a contiguous sheet of ECM containing
various proteins, separating the epidermis from the dermis. Such proteins are laminin,
collagen IV and various heparan sulphate proteoglycans. Three main zones in the BM can be
recognized: lamina lucida, lamina densa and sublamina densa. The key structure of the BM is
lamina densa, characterized by a network of cords and intercordal spaces (Leblond and

11
Inoue., 1988). The hemidesmosomes attach the basal cells of the epidermis to the BM by a
complicated intercellular arrangement. Together with specialized structures, primarily
anchoring filaments and fibrils, they form adhesion complexes that are extracellularly
connected to the cytoskeletal network of cytokeratin filaments (Jones et al 1989). As the
constitution of the BM is so complex, it provides for several functions such as attachment of
cells, a support or scaffolding function, regulation of permeability through the membrane, and
plays a part in regulating the growth and differentiation of the keratinocytes (Briggaman.,
1982; Delvoy et al., 1988; Bosman et al., 1989).
The connective tissue of the dermis consists of collagenous and elastic fibres embedded into
the ECM. All three components are synthesized by fibroblasts. Collagen is by far the most
abundant constituent of the dermal connective tissue. The collagen fibres form either a neatly
woven network as in the superficial papillary dermis (PD) or thick bundles buried in the
deeper reticular dermis (RD). A few fibroblasts are present between the collagen bundles.
Another cell type present in normal dermis is the mast cell, often located perivascularly. In
PD, the ECM has a loosely structure, with sparse collagen content, but numerous GAGs such
as HA, heparan sulphate, fibronectin and chondroitin sulphate. The physicochemical
properties of the HA polymer and interaction of the polymer with other macromolecular
components is the prerequisite for rapid adaptive changes of the skin, for instance when
subjected a number of disease conditions, including trauma (Juhlin., 1997; Laurent and Fraser
1992; Ono et al., 1998). As a principal molecule within ECM, the quantity of HA increases
whenever rapid tissue proliferation, regeneration and repair occur (Manuskiatti and Maibachl.,
1996). In RD the ECM is rich in fibroblasts and collagen that give the skin both stability and
rigidity (van Zuijlen et al., 2003).

12
Hyaluronan
Fig 2. Hyaluronan (HA) is a common component of the extra cellular matrix. At higher
concentrations HA exhibit interesting rheological properties due to a random coil formation
of a highly entangled network of flexible polysaccharide molecules with a diameter around
200nm.(from; Smedsröd., 1989).
HA molecules are attached to cells exhibiting specific HA receptors on the cell surface. A
relatively low cell density and an abundance of ECM material characterize loose connective
tissue. The extracellular macromolecular material consists of fibrous proteins such as collagen
and elastin, GAGs, e.g. HA, proteins for cell attachments, and other specific soluble and
insoluble proteins (Laurent, 1987). Hyaluronan (HA), previously called hyaluronic acid
(Balazs et al., 1986), is a linear polysaccharide consisting of alternating disaccharide units of
N-acetyl-D-glucosamine and D-glucuronic acid. The molecular weight of HA is usually in the
order of 106–107, and its mass has a radius of about 200 nm (Laurent, 1987). The
configuration of the molecule is that of a highly expanded random coil, but only about 0.1%
of the content is a polysaccharide. The major constituent is water, mechanically immobilized
within the coil. At a concentration exceeding 1 mg/ml the HA molecules entangle, and at
higher concentrations the chain forms a contiguous flexible network. HA is synthesized in the

13
cell membrane of most cells by the addition of monosaccharide units supplied by UDP-
glucuronic acid and UDP-N-acetylglucosamine to the reducing end of the chain (Laurent,
1987). During elongation, the polysaccharide chain extrudes through the cell membrane into
the extracellular space. The synthesis of HA is influenced by numerous agents such as
hormones, inflammatory mediators, and growth factors (Laurent and Fraser., 1986), most
likely via regulative phosphorylation (Mian N,. 1986a). In general, agents activating adenylate
cyclase appear to stimulate the production of this polysaccharide (Mian N,.1986b).
In some tissues an HA–mediated endocytosis and intracellular deposition occur
(Hua et al .,1993). The turnover rate of HA is surprisingly rapid; its half-life in skin and joints
is about 12 h, in the anterior chamber of the eye about 60-90 min, and in the vitreous body 70
days (Reed et al, 1990). Degradation starts in the ECM of the connective tissue and the
molecules, reduced in size are transported into the general circulation to be metabolised in the
hepatocytes of the liver (Smedsröd et al., 1984). A total amount of 10-100 mg is turned over
in the circulation of an adult human each day (Fraser and Laurent, 1989).
Many physiological and cell biological functions of HA have been associated with the
physico-chemical characteristics of its polymer network and hydrodynamic capacity (Laurent
et al., 1995). Examples are lubrication, viscoelasticity, flow resistance, osmotic pressure,
exclusion properties and filter effects. HA contributes significantly to the water balance in the
body and it affects the transport and distribution of plasma proteins in the tissues (Laurent and
Fraser, 1992). Many studies have demonstrated that HA is involved in the proliferation,
recognition, and locomotion of cells. Increased concentrations of HA and the presence of HA-
receptors, such as CD-44, ICAM-1 and RHAMM (Brown, 2004) have been found in
proliferative tissue.

14
The total content of HA in the human body is calculated to 16g half of which is located in the
skin, contributing to its normal architecture and function (Reed et al., 1988). The distribution
of HA in healthy skin involves both epidermis and dermis. In epidermis HA is present mainly
in the intercellular spaces around the middle and upper parts of the spinous keratinocytes in
the basal layer, whereas in the granular and cornified layers little, or no HA is present (Tammi
et al., 1988; Wells et al., 1990). In dermis, the most intense HA staining is seen in PD (Wells
et al., 1990; Bertheim and Hellström, 1994). Nevertheless, HA is also present in the RD,
showing weak staining around the coarser fibres and stronger staining in the outer lining of
blood vessels, as around the adnexal structures (Wells et al., 1990, Bertheim and Hellström,
1994). The importance of HA in normal skin functioning has been asserted in numerous
studies (Burd et al., 1991; Juhlin, 1997). Due to the many postulated functions of HA (Laurent
et al 1995), its localization directly below the basement membrane zone (BMZ) is of the
greatest interest, since changes here might affect not only the basement membrane but also the
entire epidermis.
Another important role of HA is its influence on cell locomotion, which characterizes tumour
invasion (Brecht et al., 1986). Tumours often have an enriched HA content and in some,
increased concentrations are correlated with invasiveness (Liotta et al., 1977; Barsky et al.,
1983; Zhang et al., 1995). In various tumour types, the stroma too has been shown to contain
increased concentrations of HA, which would seem to correlate with the grade of malignancy
and metastatic capacity of the tumour (Toole, 1981; Turley and Tretiak, 1985; Tammi et al.,
1991; Rooney P et al., 1995).
In wound healing HA has been reported to effect scarless healing in fetal wounds by virtue of
its increased concentration and prolonged presence in that type of tissue, possibly because it
lacks degrading capacity due to absence of the degrading enzyme hyaluronidase (Longaker et

15
al., 1991). In normal wound healing, HA content is increased during the early wound phase,
when the proliferative activity peaks (Weigel et al., 1986, Gailit and clarc., 1994). In this
period it is assumed that HA together with fibrin forms the early granulation tissue (Bartolami
and Donoff., 1978), which attracts inflammatory cells into the wound. CD-44 is suggested to
function as an HA receptor in granulation tissue cells, such as macrophages and fibroblast
(Oksala et al., 1995). This matrix modifies by the cells entering the wound as they secrete
hyaluronidase and plasminogen activator into the ECM to degrade HA and fibrin (Weigel et
al., 1986).
The clinical use of HA is increasing rapidly, mainly because of it has no inflammatory
properties, and has rheological capacity. Today, it is used in joint diseases to promote
cartilage-healing, functions as a lubricant and is now an important agent in ophthalmic
surgery (Laurent et al., 1995). In the skin, it has been used as a dermal expander agent to
eliminate wrinkles or fill out various skin defects. In experimental animal models it has been
shown to increase the healing capacity of chronic wounds in the tympanic membrane
(Hellström and Laurent, 1987; Laurent et al., 1988). Furthermore, it has been used to heal
chronic diabetic ulcers (Abatangelo et al., 1983).
Wound healing process Wound repair is a complex process, where a sequential occurrence with diverse cells and
substances must perform their special task in coordination with each other. Several biological
cell phenomena are involved in this reparative process, such as cell migration, proliferation,
BM regeneration, and the formation of granulation tissue. Thus, wound repair requires
continual interactions among cells, cytokines and matrix. The skin’s response to injury can be
divided into four phases that begins with an orderly process of haemostasis and fibrin

16
deposition, which leads to an inflammatory cell cascade, characterized by neutrophils,
macrophages and lymfocytes within the tissue (Diegelmann et al., 2004). Attraction and
proliferation of fibroblasts and collagen deposition is forming the granulation tissue and
finally deposition of matrix and remodelling of the scar tissue occur (Kirsner and Eaglstein.,
1993). An injury results in extravasations of blood constituents, including mediator release,
which initiates an inflammatory reaction. Activated platelets affect haemostasis by adhering
to de-endothelialized blood vessel walls, by clumping and by opening coagulation pathways.
Platelets also release a cascade of biologically active substances, including ECM molecules
such as hyaluronan, fibronectin, fibrin/fibrinogen, and various cytokines such as growth
factors, platelet-derived growth factor (PDGF), integrins, interleukin-1, epidermal growth
factor, transforming growth factors (TGF-α and TGF-β), connective tissue-activating peptide-
III, and keratinocyte growth factor (KGF) (Berman and Duncan., 1989; Gailit and Clark.,
1994; Diegelmann et al., 2004). These molecules activate the synthesis and growth of
fibroblasts, and promote cell migration into the wounded site.
Within the first few hours after an injury, abundant neutrophils infiltrate the wound site.
Substantial numbers of monocytes accumulate from 24 up to 48 hours later. Both cell types
are attracted to the site of tissue injury by a variety of chemotactic factors. The major function
of the neutrophils is to prevent bacterial invasion. Monocytes, in their activated state as
macrophages, phagocytos and eliminate pathogenic organisms, scavenge tissue debris
including combusted neutrophils, and appear to be critical for the initiation of tissue repair
(Hammar., 1993). Macrophages release a plethora of biologically active substances, many of
which facilitate the recruitment of additional inflammatory cells or remove tissue debris and
decontamination (Diegelmann et al., 2004). Macrophages also secrete growth factors, such as
PDGF and TGF-β, which initiate and sustain new tissue formation. Thus the macrophages are

17
believed to play a pivotal role in the transition between the inflammatory phase of wound
healing and the second phase and the formation of granulation tissue.
Granulation tissue consists of a dense array of macrophages, fibroblasts and neovasculature
embedded in a loosely woven matrix of fibronectin, collagen types I and III and HA.
As fibroblasts migrate to the wounded site, they display several distinct phenotypes, first a
migratory phenotype and ultimately a contractile phenotype, In the contractile state, the so-
called myofibroblasts align themselves along newly deposited ECM in the radial axes of the
wound, form cell-cell and cell-matrix links, and generate tension across the wound to cause
wound contraction (Clark, 1988).
Blood vessel growth, angiogenesis, into the wounded site occurs simultaneously with the
ingrowth of fibroblasts and ECM deposition. Endothelial cells, lining the microvasculature
adjacent to a wound, dissolve the supreme BM, emigrate through the disrupted barrier, and
migrate as a cord of cells on a provisional matrix into the wounded area. As endothelial cell
cords link up to form new arcades of capillaries, lumina appear in the centre of the cords and
blood flow normally begins around the third day, after the trauma. BM rapidly forms between
the endothelium of the capillaries and the neomatrix. When the epithelium is disrupted at the
time of injury, re-epithelialization quickly ensues to re-establish tissue integrity. Within a day
or two, the epithelial cells remaining at the wound margin begin to proliferate. The additional
population of cells then migrate across the wound bed. If the BM has been destroyed,
epidermal cells migrate over a provisional matrix of fibronectin and fibrin. A new BM grows
inward from the wound margin. If the BM has not been destroyed, hemidesmosomes at the
wound margin undergo dissolution, thus facilitating reepithelialization (Clark, 1988).

18
In the absence of firm binding at the epithelial-stromal interface is a prerequisite for epithelial
mobility. Once reepithelialization is complete, the cells revert to their normal phenotype and
reattach firmly to the BM by means of hemidesmosomes.
The final phase of wound repair is matrix remodelling. As with all phases of wound repair,
the third phase overlaps its predecessor. In fact, matrix production and remodelling begin
simultaneously with granulation tissue formation. Nevertheless, in the month that follows
granulation tissue dissolution, the matrix altereds continuously. Most fibronectin is rapidly
eliminated from the matrix and type I collagen fibres slowly grow into large bundles that
provide the residual scar with increasing tensile strength (Clark, 1988). Decorin, a small
dermatan sulfate proteoglycan has demonstrated a high affinity for type I collagen and is
assumed to regulate connective tissue regeneration via collagen fibril formation (Oksala et al
1995).
HA is a major component of early granulation tissue (Clark, 2001), increasing in content until
3 days after injury, before declining. As mentioned earlier, HA content continues to be
increased in the fetal wound, leading to scarless healing in which no inflammatory reaction is
evident (Longaker et al., 1989).
Pathologic scar tissue formation
When wound healing involves the BM, the healing will result in a scar formation. After tissue
injury, resident fibroblasts undergo metabolic activation and exhibit increased growth and
synthesis to repair the injured tissue. Under basal conditions, this activity of resident
fibroblasts is limited and probably controlled by interaction with native connective tissue
matrix components (Diegelmann et al 2004). When something fails in the remodelling phase,

19
this scar tissue formation develops into a hypertrophic scar, or a keloid, instead of a mature
scar type (Ketchum 1974; Rockwell et al., 1989; Phan et al., 2002). Hypertrophic scar is
difficult to distinguish from keloid, as they have similar clinical features and resemble each
other histopathologically (Murray et al., 1981; Muir, 1990; Linares et al., 1972). Keloids are
characterized by an abnormal rate of collagen production, and increased levels of certain
ECM components such as fibronectins and proteoglycans (Babu et al., 1989; Bailey, 1975;
Kischer et al., 1983), but a decrease in others, such as HA (Meyer et al., 2000). The keloidal
growth pattern is characterized by an accelerated fibroblast proliferation with an abnormal,
abundant deposition of collagen and ECM components, growing beyond its boundaries and
invading the neighbouring uninjured tissue (Blackburn and Cosman, 1966). The exact
background to this uncontrolled growth behaviour is still unknown. It has been suggested that
hypoxia as a result of microvessel occlusion might be an initiating factor in the pathogenesis
of excessive collagen synthesis (Hunt et al., 1978, Le et al., 2004). Several investigations have
disclosed some important facts, but many mechanisms remain to be discovered. Recent
studies have suggested the existence of some kind of keratinocyte-fibroblast interaction. The
proliferative rate of keloid fibroblasts seems to be stimulated by certain factors secreted by
keloid derived keratinocytes (Phan et al., 2002).
Cutaneous changes in diabetes mellitus
Cutaneous manifestations and impaired wound healing, most often in conjunction with
various types of disturbance affecting the internal organs, are related to diabetes mellitus
(DM) (Huntley, 1989; Lundbæk., 1957). Furthermore, changes are seen in various tissues due
to their hyperglycaemic state and hormonal aberrations underlying the altered metabolic state
(Lieberman et al., 1980). DM is divided into two groups depending on insulin dependency or
not. In type 1, (IDDM), there is insufficient insulin production due to reduced functioning in

20
Langerhan island cells in pancreas. In type 2, (NIDDM) the glycaemic state is high, because
of a reduced sensibility for insulin. The cutaneous disturbances, seen in DM, are correlated to
the severity of the disease. The contribution to these changes of the skin concerns both the
BM, which becomes more porous due to increased degradation or inadequate synthesis of
heparan sulphate proteoglycan (Rohrbach et al., 1982), and the dermal connective tissue,
demonstrating an increase in size and disorganization of the collagen bundles (Lyons and
Kennedy., 1985). Another well-known feature in diabetes mellitus is stiffening of the
periarticular connective tissue of the hands, cheiroartropathy, also named limited joint
mobility (LJM). This condition is believed to identify a population exceptionally at risk of
early development of microvascular complications. The LJM test can be used to evaluate the
severity of the disease (Rosenbloom et al., 1981).
Basal cell carcinomas
Basal cell carcinomas (BCC) are usually regarded as harmless. They display a slow pattern of
growth and rarely metastasize to other organs (Conley et al., 1985). Nevertheless, BCCs are
malignant tumours and behave accordingly by invading surrounding tissues. The destructive
consequences of invasion derive from the migration of malignant cells into adjacent tissues
and from their subsequent growth in the new location. BCCs occur chiefly in Caucasians,
especially in elderly individuals. Prolonged exposure to sunlight is one suggested cause of
BCCs. The head and neck are the sites of predilection. Most BCCs respond well to local
treatment (Pollack et al., 1982).
Other BCC types grow slowly but aggressively, infiltrating the deeper structures beneath the
skin. Many clinicians and pathologists recognize from experience the aggressive form of
BCC, which is either recurrent or deeply invasive into underlying muscle, cartilage and bone.

21
These tumours are difficult to delineate and tend to recur after treatment. Such tumours often
occur in the nasolabial fold and lacrimal duct, growing downward and being difficult to
circumscribe. They are most often referred to as infiltrating, morphemic, “sclerosing BCC”
by the pathologist (Blackburn and Cosman, 1966).
The characterization of most BCC tumours is otherwise diverse, including the more common
types: superficial, nodular, cystic, and ulcerated. The aggressive BCC types are more
ulcerative and infiltrative in their behaviour, consist of small nests of tumour cells, often
displaying an irregular, spiky appearance with infiltration of cells in cords. The usual
peripheral palisade is absent, and the cells tend not to exhibit any differentiation.
Hyalinisation of the stroma is more common in the aggressive BCC type.
The choice of treatment is often based on a preoperative biopsy, which will provide
information on the histological behaviour and distribution of tumour cells in the tissue. It is
difficult to predict aggressive behaviour in BCCs, but nevertheless important prior to deciding
which treatment to adopt. Even after using modified surgical technique, with a microscope
that provides for precise excision, the recurrence rate is high.

22
AIMS OF THE STUDIES The aim of the present work was to investigate the pathogenesis of impaired skin reparative processes especially with regard to hyaluronan (HA). Studies were performed of various scar tissues, skin biopsies from patients with type-1 diabetes mellitus and basal cell carcinomas (BCC). Against this background, the studies were designed to answer the following specific questions: I Does the distribution of HA in various forms of scar tissue contribute to their
differing clinical characteristics? II Does the distribution and content of HA in the skin of patients with diabetes
mellitus explain the severity of their clinical manifestations? III Does the distribution of HA in various BCC tumours correspond to their
differing proliferative activity and capacity of the tumour to spread? IV Does the ultrastructural architecture of abnormal scars, such as keloid and
hypertrophic scar, provide us with further information as to their different clinical characteristics?

23
MATERIAL
Tissue samples from human skin were obtained for all studies. Each patient was informed of
the study prior to surgery, and tissue samples were obtained with their consent.
Scar tissues All patients, who suffered from an abnormal scar, visited the Department of Plastic Surgery,
at which they were examined and diagnosed by an experienced plastic surgeon prior to
treatment. The scar tissues were excised with narrow margins. In paper I; tissues from 39
patients; mature scar (n=6), hypertrophic scar (n=14) and keloids (n=12), were obtained.
From one person two samples were collected: an earlobe keloid and a hypertrophic scar on the
cheek, both scars a result from a dog bite 3 years earlier. Normal skin (n=8) was excised from
various locations in healthy patients with other diagnoses in which skin biopsies were
included. For the ultrastructural study in paper IV; scar tissue samples from 16 patients;
mature scar (n=3), hypertrophic scar (n=3), keloids (n=5) and normal skin (n=5) were used.
Diabetes mellitus Skin punch biopsies, from 23 patients with insulin dependent diabetes mellitus (IDDM) and
varying grades of limited joint mobility (LJM) were used. The biopsies; diabetic skin with
LJM grade 0 (n=10), LJM grade 1 (n=7), LJM grade 2 (n= 6), were taken from the dorsum of
the hand, 4 cm distal to the ulnar head. Biopsies from the same area were also obtained from
non-diabetic patients (n=6).
Basal cell carcinoma (BCC) Patients, consulting a dermatologist at the department of Dermatology, or a plastic surgeon at
the department of Plastic Surgery at the University hospital in Umeå, were diagnosed

24
preoperatively with some type of BCC tumour. They were treated with surgery and thus
included in this study. Thirty specimens from 28 patients were used: superficial BCC (n=10),
nodular BCC (n=10) and infiltrative BCC (n=10). Two patients had at the same time different
types of BCC tumours, which were diagnosed and both included in the study. The basal cell
carcinomas were surgically removed, prepared and examined by an experienced dermato-
pathologist to the pathologic anatomic diagnosis and ensure the radicality of the surgery.
Pieces of the tumour tissue were then used for this study. As controls, normal skin was
excised from various locations in healthy patients treated for other diagnoses in which skin
excision was included.

25
METHOD
Paraffin embedded material
Papers I-IV: The excised tissue pieces were transferred to saline and within 30 minutes fixed
in a solution containing 2% formaldehyde and 0.5 % glutaraldehyde in 0.1 M phosphate
buffered saline (PBS). For localization of HA, fixation was performed under microwave
irradiation. The specimens were irradiated at 700 W up to 45 C and then transferred to PBS
and kept at cold storage temperature until further processed and embedded. The tissue
samples were dehydrated in upgraded series of ethanol to xylene and embedded in paraffin
wax. Serial paraffin sections (5 m) were cut and prepared for light microscopy.
Plastic embedded material
Paper IV: Pieces of scar tissue were fixed in 3% glutaraldehyde solution in 75 mM sodium
cacodylate buffer with 4 % polyvinylpyrrolidone and 2 mM CaCl2 added. The specimens
were postfixed in 1% OsO4 overnight and dehydrated in increasing concentrations of acetone
and embedded in an epoxy resin. Semithin sections, 0.5-1.0 m were then stained with
toluidine blue for examination in the light microscope, and ultrathin sections 70-85 nm were
contrasted with uranyl acetate and lead citrate for electron microscopy.

26
Histochemical staining for HA
Papers I-IV: Following deparaffinization, the tissue sections were washed in PBS and then
incubated with a fresh solution of 3% H2O2 in methanol 5 min at room temperature to destroy
endogenous peroxidase activity. After 2 washes in PBS the slides were incubated with 1.0%
bovine serum albumin for 30 min at room temperature to block non-specific binding sites.
The slides were washed with PBS and then incubated with approximately 100 l of
biotinylated hyaluronan binding protein probe (HABP) at a dilution of 1:40, overnight, at 4 C
storage temperature. After the washings in PBS, the slides were incubated with the
Vectastain-Elite avidin-biotin complex at a dilution of 1:200, for 40 min, at room temperature.
After 3 washes in PBS for 10 min each, the sections were incubated for 5 min in 0.1%
diaminobenzidine tetrahydrochloride (DAB) and 0.03% H202 in 0.05 M TRIS-HCL buffer,
pH 7.6, at room temperature, which produced a water insoluble brown precipitate. Finally the
slides were washed in tap water for 5 min and cover slipped.
Immunohistochemical analysis
Paraffin sections (4 m) were mounted, deparaffinized and washed in 3% H2O2 in methanol
for 5 min at room temperature to destroy endogenous peroxidase activity. Antigen retrieval of
the sections was performed for all antibody stainings in 10 mM citric acid (pH 6.0) during two
microwave irradiation cycles of 5 min at 700W. In paper III, the proliferative activity was
studied by use of a rabbit anti-Ki-67 polyclonal antibody in 0.05 M TBS (pH 7.4), and a
rabbit anti-PCNA monoclonal antibody. In paper IV, the distribution of collagen I and III was
studied by incubation in normal swine serum 1:20 for 30 min before incubation for 1 hour in a
1:400 solution of rabbit anti- human collagen type I /III, prior to application of a 1:300
dilution of biotinylated rabbit anti-mouse IgG. The immunoreactive sites were visualized with

27
diaminobenzidine, counterstained with Mayer Htx stain and cover slipped. Sections incubated
without the primary antibody but otherwise treated identically served as negative controls.
Quantitative analysis of HA in tissue samples
In paper II, tissue samples were digested with pronase (Protease P-5005 5 units/2 ml buffer
0.05 M TRIS-HCL-0.01M CaCl2, pH 7.2) for 18 h at 55 C. The digests were kept in a bath of
boiling water for 10 min and then stored frozen at 80 C. Prior to analyses, 0.1 ml 0.2 M
phenylmethanyl-sulphonyl fluoride in 99% ethanol was added to inhibit the remaining
enzyme activity. The solution was centrifugated for 10 min at approximately 2,000 rpm and 2
ml of the supernatant was then applied onto a 10 ml Sephadex G-25 column (PD-10,
Pharmacia Fine Chemicals, Uppsala, Sweden) followed by PBS. The first 3 ml of the eluate
was discarded and the subsequent 4 ml was saved for analysis of the HA content, according to
the method described by Laurent & Tengblad (1980). The method utilizes purified HABP
extracted from nasal bovine cartilage. The HABP is used as an antibody in an RIA- like type
of assay.
Statistical methods In paper II, an unpaired t-test was used for calculation of differences between the groups
regarding age, duration of disease, HbA1c and microalbuminuria. A non-parametric test, the
Mann-Whitney U-test, was used to calculate a possible correlation between epidermal
thickness and the histological grading of the HABP staining. In paper IV, acquired data were
tested with the non-parametric Kruskal-Wallis Test.

28
Examination In papers I-IV, the specimens were examined with a Zeiss Axiophot light microscope. All
images were photodocumented by means of a Zeiss Axiophot photomicroscope, using
negative film, developed and processed to photopaper. In paper IV, electron microscopy was
performed with a JEOL 1200EX, (Tokyo, Japan). Electron micrographs were made using
negative film, developed and processed to photopaper.

29
RESULTS
The architecture of normal skin
Papers I-IV; Epidermis is attached to the irregularly shaped basement membrane and the collagen-rich
dermis. The epidermis consists of keratinocytes, which possess intercellular bridges,
desmosomes. The desmosomes are large (0.37µm), elongated, spindle shaped, electron-dense
plaques. The basement membrane forms a continuous sheet of extracellular matrix, with
hemidesmosomes attaching the basal cells of epidermis to it. In epidermis the distribution of
HA was moderate and HA was mainly seen in the interstices of the spinous layer. In the
cornified and granular layers, no HA was observed. The basal layer displayed a weak,
interstitial HA-staining.
Studying the proliferative activity in epidermis by the use of Ki-67 and PCNA,a moderate
nuclear staining localized to the basal cells of epidermis was revealed.
Collagen, embedded with elastic fibers in the ground substance, represents by far the most
abundant constituent of the dermal connective tissue. In PD, HA was present in a dense layer
directly below the epidermis, with the most pronounced staining just beneath the basement
membrane. In contrast, a weak HA-staining was seen in RD, distributed in an irregular, mesh-
like pattern. In the superficial PD the collagen existed in a finely woven network consisting of
thin calibre fibers. In the deeper RD, the collagen fibers, arranged in bundles were of a thick
caliber. Regarding collagen types, the PD contained moderate amounts of collagen I and III.
In the RD, moderate amounts of collagen I were observed, but only a weak distribution of
collagen III. A small number of fibroblasts were seen between the collagen bundles. Another
cell type present in the normal dermis was the mast cell, generally occurring in small numbers
around the vessels.

30
Scar tissue Papers I and IV; In mature scars the localization of HA resembled that of the normal skin, with the most
intense HA staining in PD. An intense HA-staining of PD was also seen in hypertrophic scars.
The positive staining occurred close to the basement membrane. The HA-staining of RD was
scattered.
In keloids the PD was devoid of HA-staining. In contrast, a grossly thickened epidermis
showed intense HA-staining. Also, a massive, bulging RD was strongly stained for HA.
Ultrastructurally, the epidermis of all the scar tissues, except the keloid, closely resembled
that of the normal skin. The keratinocytes of the keloids were swollen in both the basal layer
and the spinous layer, and exhibited a large nucleus that almost filled out the cell cytoplasm.
All scar tissues displayed a comparably thicker epidermis than that of the normal skin. The
epidermal layer of the keloid was the thickest, approximately two times that of the normal
skin, followed by the hypertrophic scar, and the mature scar. Changes were observed
concerning the intercellular adhesion features; the hemidesmosomes and the desmosomes.
Furthermore, the microfilaments were shorter and less well organized in the keloid but
resembled that of the normal skin in the other scar tissues.
Both hypertrophic scars and keloids exhibited rectangular desmosomes of a shorter length
axis than that of the normal skin. The desmosomes of mature scars also presented a
rectangular form, but with a length axis that resembled normal skin. The width of the
desmosomes was 0.2 µm, and did not vary between the normal skin and the various scar
tissues. The density of hemidesmosomes, calculated as the number versus the length of the
basement membrane, seemed to be reduced in all scar tissues compared to that of the normal
skin. The keloid showed the lowest density, whereas the density of the hypertrophic and the
mature scars did not differ, but were slightly reduced compared to normal skin.

31
In keloids, the major finding of the dermis was the accumulation of collagen fibers, which
occupied the entire area below the basement membrane in the PD. This area corresponded to
the area lacking HA in the HA-stained specimens. In the various scar tissues the fiber
diameters displayed a less variation between the dermal layers compared to normal skin.
Thus, the collagen fibers in the RD were thinner and in the PD thicker. The changed fiber
diameter was most striking in the keloid, in which the collagen fiber diameter was almost
identical in the entire dermal region.
Interestingly, the distribution of collagen I was similar in the PD of all scar tissues, whereas in
the RD, the staining was stronger in the mature scar tissue and weaker in the keloids. The
distribution of collagen III was more abundant in the PD in the hypertrophic scar tissues and
weak in the keloids while in the RD, the distribution of collagen III was abundant in all scar
tissues. The number of mast cells was more frequent in the mature scar, than in the normal
skin. The keloid tissue exhibited the fewest number of mast cells.
Skin in diabetic patients Paper II; In the skin from patients with IDDM with no restriction of the mobility of the hands, LJM
grade 0, the distribution of HA resembled that of the normal skin, except that the dense HA
layer of PD, the basement membrane zone, was thinner. In IDDM patients with a moderately
restricted mobility of the hands, LJM grade 1, the HA pattern in PD was even less dense. The
HA pattern showed some variations, such as patches or the entire basement membrane zone
devoid of HA. In IDDM patients with severe restriction in their joint mobility, LJM grade 2,
the HA-staining of PD was weak with the basement membrane zone devoid of HA. The RD
exhibited less HA in the IDDM patients with LJM grade 2.

32
In spite of these findings, a quantitative analysis of HA, did not reveal any significant
difference in HA concentration between the groups.
In the LJM grade 2 group, an increased epidermal thickness as well as a pronounced
epidermal HA staining was evident when compared to other LJM groups as well as the
normal skin.
Significantly higher (p<0.05) HbA1c levels were observed in the IDDM patients with LJM
grade 2 than in the other groups.
Different types of BCC tumours Paper III;
In superficial BCC, the tumour islands showed a moderate HA staining. The proliferative
activity was weak as indicated by a modest mitotic activity and a weak Ki-67 and PCNA
immunoreactivity of the tumour islands. The surrounding tissue resembled normal skin, as no
differentiated tumour stroma was observed. In nodular BCC the HA staining of the tumour
strands was weak to moderate, and the tumour cells expressed an increased proliferative
activity. The surrounding tumour stroma stained strongly for HA. Tumour islands of the
infiltrative BCC stained weakly to moderate for HA and expressed an intense proliferation,
which was present throughout the entire tumour. An intensely HA-stained tumour stroma
ceased abruptly in the deeper, infiltrating part of the tumour where the adjacent connective
tissue was almost devoid of HA. Furthermore, this neighbouring connective tissue also
expressed a deranged appearance.

33
DISCUSSION
The ECM plays a vital role in govering the regulation of normal skin development and
function – mainly by means of its specific arrangement of macromolecules, such as collagens,
proteoglycans, GAGs, and glycoproteins. Cutaneous wound healing may be defined broadly
as the interaction of a complex series of phenomena, resulting in resurfacing, reconstruction
and proportionate restoration of the tensile strength of wounded skin. The process of wound
repair is a sophisticated interplay between ever-changing cell populations, growth factors,
proteases and matrix molecules. These complicated scenarios are closely regulated, changing
in response to the environment. The cellular regulation - or dysfunction of this process - that
results in the formation of abnormal scars is poorly understood (Diegelmann et al., 2004).
Both keloid and hypertrophic scar are characterized as abnormal scars, as they show
aggravated, red and itchy scarring. In contrast to hypertrophic scarring, keloids do not regress
with time, are difficult to treat and recur frequently. They require a different therapeutic
approach even though their characteristics and morphology are alike, which indicates the
importance of distinguishing between them. Several studies have in particular recognized
GAGs - and especially HA – as being important for a normal wound healing process (Clark.,
2001; Burd et al ., 1991; Manuskiatti and Maibach., 1996). A breakthrough in the role of HA
in wound healing research was the discovery of scarless wound healing coupled with the fetal
ECM (Longaker et al., 1989; Longaker et al., 1991; Burd et al., 1990; Siebert et al., 1990),
which is extremely riched in HA.
In postnatal wounds, other studies have shown HA to be an important feature in the healing
process interacting mainly during the early events (Oksala et al., 1995; Weigel et al., 1986;
Brown, 2004).

34
In papers I, III, IV, skin from different parts of the body of individuals of various ages, was
investigated. In paper II, biopsy samples were collected from a standardized location on the
diabetic patients, on the dorsum of the hand. In spite of the various biopsy sites and varying
ages of the patients (in papers I, III, IV) no obvious differences in the distribution of HA
could be observed. Furthermore, in papers IV, and I, the interval between the trauma and the
surgical procedure (including the biopsy) varied with no significant differences in HA
distribution within the scar groups.
HA in wounded skin
Fig 3. Schematic description of the localization of hyaluronan (HA) in; (S) normal skin, (H) hypertrophic scar
and (K) keloid. Ep= epidermis, BM= basement membrane, PD= papillary dermis. RD= reticular dermis.
In the present thesis it was found that the localization of HA in the normal skin was located
chiefly in PD, immediately beneath the BM. In contrast, the distribution of HA in RD was
sparse, a finding that supports the results in other studies on postnatal skin (Tammi et al.,
1988; Wells et al., 1990).
In paper I, a similar pattern of HA was seen in mature scar, whereas the HA in hypertrophic
scar- and particularly in keloid - was lacking in the PD. In these abnormal scars, HA occurred

35
mainly in the bulging RD and the thickened epidermis. In paper IV, it was demonstrated that
in keloid, the region of PD lacking HA, bordering the BM, was filled with densely packed
collagen fibres, thicker than in the normal skin. The altered dimension of the collagen fibres
indicates that they contain another type of collagen than the PD of normal skin. On the
contrary, the RD of the keloid contained collagen fibres, which appeared finer than that in
normal skin. The entire bulging dermis in the keloids was filled with collagen fibres of a
diameter similar to that in PD. This finding contradicts the general opinion that keloids
contain thick collagen fibres (Ehrlich et al., 1994). The immunohistochemical studies showed
an increased occurrence of collagen type III and a decrease in type I in the RD, with a
decrease in type III in the PD in the keloid, compared with the normal skin, which supports
earlier findings (O’Sullivan et al., 1996). The increase in collagen III in RD, was not
pronounced, however, and the altered collagen fibres of the keloid probably belong to another
collagen type than collagens III and I. Considering the keloid as an uncontrolled growing scar
tissue with invading capacity to reach beyond its margins, should be regarded as important.
That the same, fine calibre collagen fibres are present in both PD and RD could be due to an
uncontrolled production of fine calibre collagen. This collagen does not create the appropriate
stability or rigidity in the dermis, and will thus allow the tissue to expand with no other
restriction than the neighbouring normal skin areas. Various alterations in the architecture of
the epidermal region were also noted. The keloid displayed a reduced number of
hemidesmosomes anchoring to the BM, and furthermore, the desmosomes connecting the
intercellular space of the spinous layer appeared different.
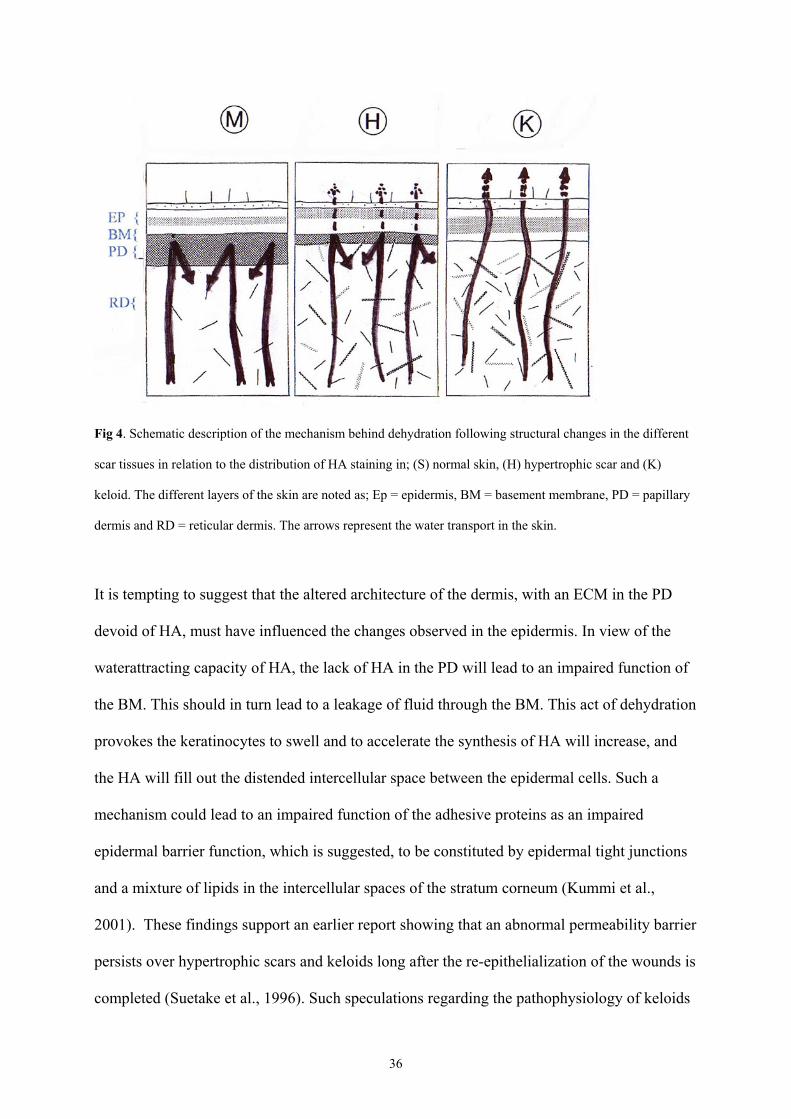
36
Fig 4. Schematic description of the mechanism behind dehydration following structural changes in the different
scar tissues in relation to the distribution of HA staining in; (S) normal skin, (H) hypertrophic scar and (K)
keloid. The different layers of the skin are noted as; Ep = epidermis, BM = basement membrane, PD = papillary
dermis and RD = reticular dermis. The arrows represent the water transport in the skin.
It is tempting to suggest that the altered architecture of the dermis, with an ECM in the PD
devoid of HA, must have influenced the changes observed in the epidermis. In view of the
waterattracting capacity of HA, the lack of HA in the PD will lead to an impaired function of
the BM. This should in turn lead to a leakage of fluid through the BM. This act of dehydration
provokes the keratinocytes to swell and to accelerate the synthesis of HA will increase, and
the HA will fill out the distended intercellular space between the epidermal cells. Such a
mechanism could lead to an impaired function of the adhesive proteins as an impaired
epidermal barrier function, which is suggested, to be constituted by epidermal tight junctions
and a mixture of lipids in the intercellular spaces of the stratum corneum (Kummi et al.,
2001). These findings support an earlier report showing that an abnormal permeability barrier
persists over hypertrophic scars and keloids long after the re-epithelialization of the wounds is
completed (Suetake et al., 1996). Such speculations regarding the pathophysiology of keloids

37
have not been put forward earlier. However, these morphometric changes give support to and
a theoretical explanation of the benefit of a treatment strategy with hydration and occlusion,
which not only improves the clinical symptoms but also permits an improvement of the scars’
morphological properties (Perkins, 1982; Sawada et al., 1998). Regarding a possible dermal –
epidermal relationship (Nowinski et al., 2002; Phan et al., 2002), these epidermal changes
could implicate an impaired regulatory function on the collagen formation, emanating from
the pathophysiological characteristic of the keloid.
Regarding the clinical characteristics of the abnormal scar with reddening and itching, the
mast cells and macrophages have been suggested to be intimately involved in the formation of
keloids and hypertrophic scars. An increase in the number of mast cells has been reported in
these scar tissues (Kischer et al., 1989). Mast cells release a plethora of substances that
intensifies the inflammatory response in the injured area, considered important in regulating
the normal wound healing process (Noli and Miolo., 2001, Diegelmann et al., 2004).
In paper IV, mast cells were quantitated; few were noted in the PD and RD both in keloids
and in hypertrophic scars, compared with those in mature scar and the normal skin. This was a
surprising result, suggesting the possibility of other mechanisms than mast cells to cause the
reddening and itching in the abnormal scar.
HA in skin in diabetes mellitus
Many patients with type-1 diabetes have restricted mobility affecting their palms, called
LJM, combined with a thickened, waxy skin. This combination of symptoms, called
cheiroarthropathy is associated with appreciably increased risk of microangiopathy
(Rosenbloom et al., 1981). This diabetic microangiopathy of the skin includes morphologic

38
and functional changes with thickening of the capillary membranes, and enhanced collagen
deposition around papillary microvessels (McMillan., 1966). Since the biochemical change in
the skin in type-1 diabetes has not yet been identified, but is assumed to involve the ECM, we
wanted to investigate the distribution of HA in the affected skin in an attempt to correlate any
change in the severity of the disease. Our findings demonstrated a modified ECM
composition, with changes in the distribution of HA in the diabetic skin suggesting that
pathophysiological events are not related exclusively to the microvasculature.
We found significantly less HA localized in the PD of the patients most severely affected.
Furthermore, the epidermis of the diabetic skin was thicker and stained more strongly for HA
than the normal skin. This compartmentalization could be one explanation why the analysis of
the total amount of HA showed no significant difference between the diabetic patients and the
healthy controls. The thickening of the epidermis, with intensitied HA staining, and less HA
in the PD, resembles the HA staining pattern in keloids.
HA is recognized not only as an important component of the structural organization of the
skin with its hydrophilic capacity, but also as involved in cell migration, angiogenesis,
immune reactions and phagocytosis. A loss of HA from the superficial dermis could have
serious consequences for the skin’s integrity, as well as in connective tissues of other organs
where similar events occur. This finding is most certainly the consequence of a structural
change in the ECM and collagenous tissue, which should occur parallel with the development
of microangiopathy. Increased glucose concentration in the media in which normal fibroblasts
are grown in culture influences the amount, the type, and possibly the glycosylation of
collagen secreted by them (Ville and Powers., 1977). When evaluating the general status of
diabetic patients, all changes in the ECM should be regarded as important,

39
HA in tumour progression
HA affects the physical environment of cells and has been suggested to promote cell
migration through its cell surface receptors. HA expression is often increased in malignant
tumours, and is assumed to be associated with the invasive potential of various tumour types
(Knudson et al., 1989; Toole., 1991). Several functions of HA in cell migration are
described, including a weakening of the attachment to the adhesive substrata, a facilitation of
the partial detachment, and the creation of a hydrated pathway that may facilitate the invasion
of tissues by separating cellular and fibrous barriers (Toole., 1997). The presence of tumour-
associated HA may therefore play an important role in facilitating tumour cell invasion of
certain host connective tissues (Knudson et al., 1989).
In paper III, we have described the relationship between HA and various types of BCC
tumours. It was found that the proliferative activity of BCC cells is associated with an
increased expression of HA in the tumour stroma. In the superficial and nodular BCC types,
often depicted as slow growing tumours, the HA pattern of the tumour stroma resembled that
of PD in normal skin. Paradoxically, this stromal reaction with intense HA staining may
represent a highly developed protective mechanism generated by the healthy tissue against the
tumour cells. It can be debated whether enrichment with tumour associated HA could serve as
a favourable or unfavourable prognostic factor (Knudson et al., 1989; Pirinen et al., 1998).
In the infiltrative BCC, the staining intensity for Ki-67 and PCNA of the tumour islands
evidenced an intense proliferation, despite a weak staining of HA. The stroma surrounding the
tumour cells showed intense HA staining. However, in the deeper, infiltrating parts of the
tumour the presence of HA decreased and at some distance the adjacent connective tissue was
almost devoid of HA. The modification of the tumour-associated connective tissue indicates a

40
close relationship between the tumour cells and the adjacent matrix. This alteration of the
neighbouring connective tissue could be of utmost importance in the pathophysiology of
tumour infiltration. We regard the modified stromal tissue as a prerequisite for tumour
progression, which also could cause a recurrence of the tumour when treating it surgically, as
these changes can reach much further than the clearly visible extension of the tumour cells.
One can assume that the loss of ECM components, paving way for the tumour invasion,
depends on an increased proteolytic activity (Leake et al., 2003), governed from the tumour
cells.

41
Clinical considerations
The pathophysiological implication of the observations in papers I-IV could provide
us with increased knowledge and understanding of the biological mechanisms behind
the formation of skin tumours, abnormal scars such as keloids and hypertrophic scars
and the impaired function of the diabetic skin.
The distribution of HA in these different tissue samples representing various
pathological conditions, all associated with an impaired wound healing, show various
differences but mainly some similarities that indicate that they might share a
pathophysiologic background.
The role for HA in the papillary dermis is vital for the structural organization and
hydrodynamic constitution of the skin. A decrease can alter the architecture of the
epidermis, provoked by a disturbed basement membrane function with increased
leakage from the skin and an impaired function.
An abundance of thin calibre collagen content characterizes keloids. This could
emanate from a sequential error with uncontrolled and disorganized production of
immature collagen that never undergoes maturation.
Considering an interconnecting relationship with a down regulatory function of the
keratinocytes on the collagen synthesis, the epidermal changes could be of
importantance for the pathogenesis of the keloid formation.
Stromal reaction in basal cell tumours could represent a highly developed protective
mechanism generated from the healthy connective tissue against the adverse tumour
cells and should be considered as a good prognostic sign. However in the infiltrating
basal cell tumour this stroma becomes deranged and serves as a prerequisite for further
tumour growth.

42
CONCLUSIONS
The differing distribution of HA in scar tissues contributes to their different clinical
characteristics
This difference should be used as a diagnostic criteria to distinguish the scar types
from each other
Specific morphologic observations of the scar tissues provide us with novel
information of their pathophysiology and will also explain some of their different
clinical characteristics
A reduction of the HA content in the papillary dermis leading to an impaired
hydrodynamic capacity and a disturbed composition of the basement membrane with
leakage of water from the dermis could be the mechanism behind the thickening of the
skin in severely affected diabetic patients and in the abnormal scar
Parallel to the structural disorganisation observed in the connective tissue of the skin
in diabetics similar changes should be expected concerning the connective tissue of
other internal organs
Stromal reaction in basal cell tumours should be considered together with the
characteristics of the tumour cells when evaluating the malignant behaviour and the
choice of treatment of the tumour

43
SAMMANFATTNING PÅ SVENSKA Hudens normala läkningsförmåga är en välreglerad men invecklad process som innefattar olika molekylära händelser vilka förväntas resultera i ett återskapande av normal funktion hos den skadade vävnaden. För att undvika en försenad eller abnormal läkningsreaktion är det av stor vikt att förstå denna process. Hyaluronan (HA) är en vanligt förekommande glykosaminoglykan (GAG) som har stor betydelse för kroppens bindvävsstrukturer och vävnadsvätskor–det sk extracellulära matrix (ECM). HA har stor vätskebindande förmåga vilket bidrar till att vävnadsmiljön medger bla celltillväxt och transport. Sådana egenskaper har uppfattats ha en särskild betydelse vid sårläkning hos foster, som läker sina sår utan förekomst av synliga ärr. HA molekylen återfinns överallt där celldelning, vävnadstillväxt eller reparation sker. Målsättningen med denna avhandling är att beskriva förekomsten och betydelsen av HA för normal hud, samt hur förekomsten av HA kan förändras vid olika sjukdomstillstånd med fördröjd eller rubbad sårläkning som följd. Vi har studerat keloider och hypertrofiska ärr, basalcells cancer samt hud från patienter med insulin- beroende typ-1 diabetes mellitus. Analyser har utförts av vävnads prover av ärr och tumörvävnad från 50 patienter som opererats samt hudbiopsier från diabetespatienter och friska försökspersoner. Undersökningar av dessa olika vävnadsprover har skett med hjälp av ljusmikroskop och elektronmikroskop. Resultaten visade att HA i den normala huden framförallt förekommer i den övre läderhuden, papillära dermis (PD). En mindre ansamling av HA finns även mellan cellerna i överhuden, epidermis. I de olika ärrvävnaderna fanns HA i olika grad;det mogna ärret (mature scar) uppvisade en likartad förekomst av HA som den normala huden. Mest avvikande var HA i keloiden där övre läderhuden (PD) saknade HA. I motsats innehöll den djupare läderhuden rikligt med HA. Det hypertrofiska ärret intog en mellanställning avseende HA förekomst mellan keloid och mature scar. Detta fynd var betydelsefullt och undersöktes vidare med elektronmikroskopi där framförallt kollagenets struktur analyserades. Även vidhäftningsstrukturer mellan basalmembran och basalceller liksom mellan keratinocyteret var förändrade i fra keloiden. Dessa avvikelser förmodas bidra till uppkomsten av en försämrad barriärfunktion hos basalmembranet (BM) med läckage av vävnadsvätska upp till överhuden som i sin tur leder till en störning av överhudens funktion med förtjockning. I huden hos diabetespatienterna observerades en minskad förekomst av HA i övre läderhuden (PD), liksom ett ökat HA innehåll i en förtjockad överhud. Liksom hos de abnorma ärren kan dessa fynd tyda på en nedsatt barriärfunktion med läckage av vävnadsvätska till epidermis. Denna ändrad struktur i huden hos diabetespatienter skulle delvis kunna förklara en försämrad sårläkning hos dessa patienter. Vid basalcellscancer omges tumörcellerna sk tumörstromat av rikligt med HA. De ytligt växande typerna som karakteriseras av långsamt växtsätt uppvisade ett HA innehåll i basalcells-tumörstromat som inte skiljde sig från frisk bindväv i frisk läderhud. Den basalcellscancer som växer infiltrativt på djupet, uppvisade ett helt annat HA mönster. En

44
minskad mängd HA förekom i tumörstromat och ännu längre ut i bindväven förekom ingen HA alls. Vi tolkar dessa observationer som att bindväven som omger tumörcellerna sk tumörstromat, genomgår en förändring av sin sammansättning under påverkan av tumörcellerna, vilket kan avläsas som en minskad närvaro av HA. Denna rubbning av bindväven har förmodligen styrts av tumören med hjälp av okänd mekanism. Bindväven kan här ha omreglerats, från tidig försvarsmekanism med uppgift att bekämpa de främmande tumörcellerna - till att bli en viktig tillväxtfrämjande struktur som tumörcellerna utnyttjar för vidare invasion i vävnaden. Dessa observationer är av betydelse för såväl prognostisk som terapeutisk värdering vid behandling av de olika basalcellstumörerna. Dessa studier visar att förekomsten av hyaluronan (HA) i huden förändras vid olika sjukdomstillstånd och speglar vävnadens vitalitet. Kartläggningen av HA skulle kunna ha stor betydelse för bedömning av sårläkningsprocessen, om den går i rätt eller fel riktning – en sårläkningsmarkör. Detektionen av HA borde således inkluderas som ett komplement i granskningen av de hudbiopsier vi idag utför när man utvärderar eventuell förekomst av inflammation eller cancerceller som orsak till olika kroniska hudbesvär och sår. Den kliniska nyttan med denna metod skulle kunna vara betydelsefull för att förbättra vår diagnostik, liksom att följa olika sjukdomsförlopp som påverkar huden. Vår behandling av de abnorma, oönskade ärren; keloider och hypertrofiska ärr, skulle bli mer precis och effektiv, med en specifik metod att kunna skilja dem åt. Tillämpningen av HA markören på hudbiopsin vid basalcellscancer borde vara användbar vid planeringen av radikalitet vid kirurgin. Studien av basalcellscancer indikerar nämligen att vi bör utföra vår kirurgi av tumörerna med större marginal för att få med den tumöromvandlade bindväven som kan innehålla omogna ”osynliga” cancerceller som annars kan tillväxa.

45
ACKNOWLEDGEMENTS Many people have been important for me during the process in carrying out this Thesis. I wish to express my sincere gratitude to all who have contributed to it, in particular: Anna Engström-Laurent, my supervisor, co-author and friend, for your guidance, scientific enlightenment’s, optimistic spirit and support. Sten Hellström, my co-supervisor, tutor, co-author and friend, who gave me the opportunity to begin research and guided me through these studies during these years, with intellectual discussion, never-ending enthusiasm, friendship and support. Claude Laurent, my friend, for your enthusiasm, expert advice and support through the years. Per-Åke Hofer, for all your knowledge about cutaneous tumours and all the time we shared behind the microscope. Andrea Lotz, for excellent secretarial assistance and friendly support. The people in the ENT laboratory in Umeå, with special thanks to Cathrine Johansson and Berith Lundström for your professional technical assistance and guidance. My collegues and staff at the Dept of Plastic Surgery, University Hospital of Umeå, for your collaboration and professional support. My collegues and staff at the Dept of Surgery, in Lycksele, for your support and friendship. Per-Olof Eriksson, Lennart Bohlin and Janne Hörnsten, Dept of ENT, for your technical support. Björn Tavelin, for excellent statistical assistance. Max Brandt, for expert linguistic revision of the manuscripts. My family and personal friends, which have made this work possible by supporting me all the time while working with my projects.

46
REFERENCES
Abatangelo G, Martelli M, Vecchia, P. Healing of hyaluronic acid-enriched wounds: Histological observations.
J Surg Res 1983;35:410-416.
Anttila MA, Tammi RH, Tammi MI, Syrjanen KJ, Saarikoski SV, Kosma VM. High levels of stromal
hyaluronan predict poor disease outcome in epithelial ovarian cancer. Cancer Res 2000; 60: 150-5.
Auvinen P, Tammi R, Parkkinen J, et al. Hyaluronan in peritumoural stroma and malignant cells associates with
breast cancer spreading and predicts survival. Am J Path 2000; 156: 529-36.
Babu M, Diegelmann R, Oliver N. Fibronectin is overproduced by keloid fibroblasts during abnormal wound
healing. Mol Cell Biol 1989; 9 (4): 1642-50.
Bailey AJ, Bazin S, Sims TJ, LeLous M, Nicoletis C, Delaunay A. Characterization of the collagen of human
hypertrophic and normal scars. Biochim Biophys Acta 1975;405:412-421.
Balazs EA, Laurent TC. Jeanloz RW. Nomenclature of hyaluronic acid. Biochem J 1986;235:903.
Barsky SH, Siegal GP, Jannotta F, Liotta L. Loss of basement membrane components by invasive tumours but
not by their benign counterparts. Lab Invest 1983; 49: 140-7.
Bartolami CN, Donoff RB. Hyaluronidase activity during open wound healing in rabbits: a preliminary report. J
Surg Res 1978;25:256-259.
Berman B, Duncan MR. Short-term keloid treatment in vivo with human interferon alfa-2b results in a selective
and persistant normalization of keloidal fibroblast collagen, glycosaminoglycan, and collagenase production in
vitro.J Am Acad Dermatol 1989;21:694-702.
Bertheim U, Hellström S. The distribution of hyaluronan in human skin and mature, hypertrophic and keloid
scars. Br J Plastic Surg 1994; 47: 483-9.
Blackburn WR, Cosman B. Histologic basis of keloid and hypertrophic scar differentiation. Arch Path 1966;
82: 65-71.
Bosman FT, Cleutjens J, Beek C, Havenith M. Basement membrane heterogeneity. J Histochem 1989;21:629-
633.
Brecht M, Mayer U, Schlosser M, Prehm P. Increased hyaluronate synthesis is required for fibroblast
detachment and mitosis. Biochem J 1986; 239: 445-50.
Briggaman RA et al-Biochemical composition of the epidermal-dermal junction and other basement membrane.
J Invest Dermatol 1982;78:1-6.
Brown JA. The role of hyaluronic acid in wound healing’s proliferative phase. J Wound Care 2004;13:48-51.

47
Burd DAR, Longaker MT, Adzick NS, Harrison MR, Ehrlich HP. Foetal wound healing in a large animal
model: the deposition of collagen is confirmed. Br J Plast Surg 1990;43:571-577.
Burd DAR, Greco RM, Regauer S, Longaker MT, Siebert JW, Garg HG. Hyaluronan and wound healing: a new
perspective. Br J Plast Surg 1991;44:579-584.
Clark,R A F. Potential roles of fibronectin in cutaneous wound repair. Arch of Dermatology 1988; 124: 201-6.
Clark RAF. Fibrin and wound healing. Ann NY Acad Sci 2001;936:355.
Clore JN, Cohen IK, Diegelmann RF. Quantitative assay of types I and III collagen synthesized by keloid
biopsies and fibroblasts. Biochim Biophys Acta 1979;586:384-390.
Conley J, Sachs ME, Romo T, Labay G, Gillooley J. Metastatic basal cell carcinoma of the head and neck.
Otolaryngol Head Neck Surg 1985; 93: 78-85.
Delvoy P, Pierard D, Noel A, et al. Fibroblasts induce the assembly of the macromolecules of the basement
membrane. J Invest Dermatol 1988;90:276-82.
Diegelmann RF, Cohen IK, McCoy BJ. Growth kinetics and collagen synthesis of normal skin, normal scar and
keloid fibroblasts in vitro. J Cell Physiol 1979;98:341-346.
Diegelmann RF, Evans MC. Wound healing: an overview of acute, fibrotic and delayed healing. Front Biosci
2004;9:283-9.
Eckert RL. Structure, function, and differentiation of the keratinocyte. Physiol Rev 1989;69:1316.
Ehrlich HP, Desmoulier A, Diegelmann RF, Cohen IK, Compton CC, Garner WL, Kapanci Y, Gabbiani G.
Morphologic and immunochemical differences between keloid and hypertrophic scar. Am J Pathol
1994;145:105-13.
Falanga V, Grinnel F, Gilchrest B, Maddox YT, Moshell A. Experimental approaches to chronic wounds.
Wound Rep Reg 1995;3:132-40.
Flint MH, Craig AS, Reilly HC, Gillard GC, Parry DAD. Collagen fibril diameters and glycosaminoglycan
content of skins: indices of tissue maturity and function. Connect Tissue Res 1984;13:69-81.
Fraser JRE, Laurent TC. Turnover and metabolism of hyaluronan. In : Evered D, Whelan J, eds. The biology of
hyaluronan. Chichester: Wiley, 1989;41-59.
Gailit J , Clark RAF. Wound repair in the context of extracellular matrix. Curr Opin Cell Biol 1994;6:717-725.
Grinnell F. Fibronectin and wound healing. J Cell Biochem 1984;26:107-116.
Hammar H. Wound healing. Int J Dermatol 1993;32:6-15.

48
Hellström, S., and C. Laurent. 1987. Hyaluronan and healing of tympanic membrane perforations. An
experimental study. Acta Otolaryngol (Stockh) Suppl 442:54-61.
Hellström S, Bloom GD, Berghem L, Stenfors L-E, Söderberg O. A comparison of hyaluronan and fibronectin
in the healing of tympanic membrane perforations. Eur Arch Otolaryngol 1991;248: 230-235.
Hua G, Knudson CB and Knudson W. Internalization of hyaluronan by chondrocytes occurs via receptor
mediated endocytosis. Journal of Cell Science 1993; 106: 365-375.
Hunt TK, Conolly WB, Aronson SB, Goldstein P. Anaerobic metabolism and wound healing:an hypothesis for
the initiation and cessation of collagen synthesis in wounds. Am J Surg 1978;135:328-32.
Huntley, A.C. Cutaneous manifestations of diabetes mellitus. Dermatologic Clinics1989; 7:531-546.
Jones JCR, Steinman HK, Goldsmith BA. Hemidesmosomes, collagen VII, and intermediate filaments in basal
cell carcinoma. J Invest Dermatol 1989;93:662-671.
Juhlin L. Hyaluronan in the skin (minisymposium: Hyaluronan). J Intern Med 1997; 242: 61-6.
Ketchum LD, Cohen IK, Masters FW. Hypertrophic scars and keloids: a collective review. Plast Reconstr Surg
1974; 53:140-154.
Kischer CW, Hendrix MJ. Fibronectin (FN) in hypertrophic scars and keloids. Cell Tissue Res 1983; 231(1):
29-37.
Kischer CW, Wagner HN Jr, Pindur J, Holubec H, Jones M, Ulreich JB, Scuderi P. Increased fibronectin
production by cell lines from hypertrophic scar and keloid. Con Tissue Res 1989;23:279-288.
Kirsner RS, Eaglstein WH. The wound healing process. Dermatol Clin 1993;11:629-40.
Kitajima Y. Mechanisms of desmosome assembly and dissambly. Clin and Exp Dermatol 2002;27:684-690.
Knudson W, Biswas C, Li X-Q, Nemec R, Toole BP. The role and regulation of tumour-associated hyaluronan.
In: The biology of hyaluronan. Ciba Found Symp 1989; 143: 150-69.
Laurent C, Hellström S, Fellenius E. Hyaluronan improves the healing of experimental tympanic membrane
perforations: a comparison of preparations with different rheologic properties. Arch Otolaryngol Head Neck
Surg 1988;114:1435-1441.
Laurent TC, Fraser JRE. Properties and turnover of hyaluronan. In: Evered D, Whelan J (eds). Functions of the
proteoglycans. Ciba foundation Symposium 124. Chichester: Wiley, 1986:9-29.
Laurent TC. Structure, function and turnover of the extracellular matrix. Adv Microcirc 1987; 13:15-34.
Laurent TC, Fraser JR. Hyaluronan. FASEB 1992;6:2397-2404.
Laurent TC, Laurent UBG, Fraser JRE. Functions of hyaluronan. Ann Rheum Dis 1995;54: 429-32.

49
Laurent UBG, Tengblad A. Determination of hyaluronate in biological samples by a specific radioassay
technique. Anat Biochem 1980; 109: 386-94.
Le AD, Zhang Q, Wu Y, Messadi DV, Akhondzadeh A, Nguyen AL, Aghaloo TL, Kelly AP, Bertolami CN.
Elevated vascular endothelial growth factor in keloids: relevans to tissue fibrosis. Cells Tissues Organs
2004;176:87-94.
Leake D, Doerr TD, Scott G. Expression of urokinase-type plasminogen activator and its receptor in keloids.
Arch Otolaryngol Head Neck Surg 2003;129:1334-38.
Leblond CP, Inoue S. Structure, composition, and assembly of basement membrane. Am J Anat 1988;185:367-
90.
Lever WF, Schaumburg-Lever G. Histopathology of the Skin, 7th ed. Philadelphia: J.B. Lippincott Co 1983. Pp
562-575.
Lieberman LS, Rosenbloom AL, Riley WJ, Silverstein JH. Reduced skin thickness with pump administration of
insulin. N Engl J Med 1980;303:940-941.
Liotta LA, Kleinerman J, Catanzaro P, Rynbrandt D. Degradation of basement membrane by murine tumour
cells. J Natl Cancer Inst 1977; 58: 1427-31.
Linares HA, Kischer CW, Dobrkovsky M, Larson DL. The histiotypic organization of the hypertrophic scar in
humans. J Invest Dermatol 1972;59:323-331.
Longaker MT, Chiu ES, Harrison MR, Stern R. Studies in fetal wound healing. IV. Hyaluronic acid stimulating
activity distinguishes fetal wound fluid from adult wound fluid. Ann Surg 1989;210:667-672.
Longaker MT, Chiu ES, Adzick NS, Stern M, Harrison MR, Stern R. Studies in fetal wound healing: V. A
prolonged presence of Hyaluronic acid levels characterizes fetal wound fluid. Ann Surg 1991;213:292-296.
Lundbaek, K. Stiff hands in long term diabetes. Acta Med Scand 1957;158:447-451.
Lyons, T.J., and L. Kennedy. Non-enzymatic glycosylation of skin collagen in patients with Type 1 (insulin-
dependent) diabetes mellitus and limited joint mobility. Diabetologia 1985; 28:2-5.
Manuskiatti W, Maibach HI. Hyaluronic acid and skin: Wound healing and aging. Int J Derm 1996; 35: 539-
544.
Matrisian LM, Cunha GR, Mohla S. Epithelial–Stromal interactions and tumour progression: Meeting,
summary and future directions. Cancer res 2001; 61: 3844-6.
McMillan, D.E. Deterioration of the microcirculation in diabetes.Diabetes 1975;24:944-957

50
Meyer LJM, Russel SB, Russel JD, Trupin JS,Egbert BM, Shuster S, Stern R. Reduced hyaluronan in keloid
tissue and cultured keloid fibroblasts. J Invest Dermatol 2000;114:953-9.
Mian N.(a) Analysis of cell – growth – phase – related variations in hyaluronate synthase activity of isolated
plasma – membrane fractions of cultured human skin fibroblasts. Biochem J 1986;237:333-42.
Mian N.(b) Characterization of a high-Mr plasma-membrane-bound protein and assessment of its role as a
constituent of hyaluronate synthase complex. Biochem J 1986; 237:343-57.
Moll R, Moll I. Epidermal adhesion molecules and basement membrane components as target structures of
autoimmunity. Virchows Arch 1998;432:487-504.
Moseley R, Stewart JE, Stephens P, Waddington RJ, Thomas DW. Extracellular matrix metabolites as potential
biomarkers of disease activity in wound fluid: lessons learned from other inflammatory diseases? Br J Dermatol
2004;150:401-413.
Muir IFK. On the nature of keloid and hypertrophic scars. Br J Plastic Surg 1990; 43:61-69.
Murray JC, Pollack SV, Pinnell SR. Keloids: A review. Am Acad Dermatol 1981;4:461-470.
Noli C, Miolo A. The mast cell in wound healing. Vet Dermatol 2001;12:303-13.
North AJ, Bardsley WG, Hyam J, Bornslaeger EA et al. Molecular map of the desmosomal plaque. J Cell
Science 1999;112:4325-36.
Nowinski D, Hoijer P, Engstrand T, Rubin K, Gerdin B, Ivarsson M. Keratinocytes inhibit expression of
connective tissue growth factor in fibroblasts in vitro by an interleukin-1alpha-dependent mechanism. J Invest
Dermatol 2002;119:449-55.
Oksala O, Salo T, Tammi R, Häkkinen L, Jalkanen M, Inki P, Larjava H. Expression of proteoglycans and
hyaluronan during wound healing. J Histochem Cytochem 1995; 43: 125-35.
Ono S, Imai T, Aso A, Yamano T, Shimizu N, Nagao K. Alterations of skin glycosaminoglycans in patients with
ALS. NEUROLOGY 1998,51:399-404.
O’Sullivan ST, O’Shaughnessy M, O’Connor TPF. Aetiology and management of hypertrophic scars and
keloids. Ann R Coll Surg Engl 1996;78:168-175.
Perkins K, Davey RB, Wallis KA. Silicone gel: a new treatment for burn scars and contractures. Burns
1982;9:406-10.
Phan TT, Lim IJ, Bay BH, Qi R, Huynh HT, Lee ST, Longaker MT. Differences in collagen production
between normal and keloid-derived fibroblasts in serum-media co-cultured with keloid-derived keratinocytes.
Journal of Dermatol Science 2002; 29:26-34.

51
Pirinen RT, Tammi RH, Tammi MI, Paakko PK, Parkkinen JJ, Ågren UM, Johansson RT, Viren MM,
Tormanen U, Soini YM, Kosma VM. Expression of hyaluronan in normal and dysplastic bronchial epithelial and
in squamous cell carcinoma of the lung. Int J Cancer1998;79:251-5.
Pollack SV, Goslen JB, Sherertz EF, Jegasothy BV. The biology of basal cell carcinoma: a review. J Am Acad
Dermatol 1982; 7: 569-77.
Pummi K, Malminen M, Aho H, Karvonen S-L, Peltonen J, Peltonen S. Epidermal tight junctions: ZO-1 and
occludin are expressed in mature, developing, and affected skin and in vitro differentiating keratinocytes. J
Invest Dermatol 2001;117:1050-58.
Reed RK, Lilja K, Laurent T. Hyaluronan in the rat with special reference to the skin. Acta Physiol Scand
1988;13:405-11.
Reed RK, Laurent UBG, Fraser JRE, Laurent TC. Removal rate of [3H] hyaluronan injected subcutaneously in
rabbits. American J of Physiology 1990;259:H532-35.
Rockwell WB, Cohen IK, Ehrlich HP. Keloids and hypertrophic scars: A comprehensive review. Plast Reconstr
Surg 1989;84:827-837.
Rohrbach DH, Hassel JR, Kleinman HK, Martin GR. Alterations in the basement membrane (heparan sulfate)
proteoglycan in diabetic mice. Diabetes 1982;31:185-188.
Rooney P, Kumar S, Ponting J, Wang M. The role of hyaluronan in tumour neovascularization (review). Int J
Cancer 1995; 60: 632-6.
Rosenbloom, A.L., J.H. Silverstein, D.C. Lezotte, K. Richardson, and M.McCallum. 1981. Limited joint
mobility in childhood diabetes mellitusindicates increased risk for microvascular disease. N Engl J Med 305:191-
194
Sawada Y, Urushidate S, Nihei Y.Hydration and occlusive treatment of a sutured wound. Ann Plast Surg
1998;41:508-12.
Siebert JW, Burd AR, McCarthy JG, Weinzweig J, Ehrlich HP. Fetal wound healing. A biochemical study of
scarless healing. Plast Reconstr Surg 1990;85:495-504.
Smedsröd B, Pertoft H, Eriksson S, Fraser JRE, Laurent TC. Studies in vitro on the uptake and degradation of
sodium hyaluronate in rat liver endothelial cells. Biochem J 1984;223:617-26.
Smedsröd B. Catabolism in liver sinusoids. In: Clinical impact of bone and connective tissue markers (Lindh E
& Thorell JI, eds) pp 5173, Academic Press, London, England.

52
Suetake T, Sasai S, Zhen YX, Ohi T, Tagami H. Functional studies after injury and comparison among keloids,
hypertrophic scars and atrophic scars. Arch Dermatol 1996;132:1453-58.
Tammi R, Ripellino JA, Margolis RU, Tammi M. Localization of epidermal hyaluronic acid using the
hyaluronate binding region of cartilage proteoglycan as a specific probe. J Invest Dermatol 1988;90: 412-414
Tammi R, Tammi M. Correlations between hyaluronan and epidermal proliferations studied by
[3H]Glucosamine and [3H]Thymidine incorporations and staining of hyaluronan on mitotic keratinocytes. Exp
Cell Res 1991; 195: 524-7.
Toole BP. Glycosaminoglycans in morphogenesis. In: Hay ED, ed. Cell Biology of the Extracellular Matrix.
New York: Plenum Press 1981. Pp 259-94.
Toole BP. Hyaluronan in morphogenesis. J Intern Med 1997;242:35-40.
Turley, EA, Tretiak M. Glycosaminoglycan production by murine melanoma variants in vivo and in vitro.
Cancer Res 1985; 45: 5098-105.
Van Zuijlen PPM, Ruurda JJB, van Veen HA, et al. Collagen morphology in human skin and scar tissue: no
adaptions in response to mechanical loading at joints. Burns 2003;29:423-31.
Ville DB, Powers ML. Effect of glycose and insulin on collagen secretion by human skin fibroblasts in vitro.
Nature 1977;268:156-8.
Weigel PH, Fuller GM, LeBoeuf RD. A model for the role of hyaluronic acid and fibrin in the early events
during the inflammatory response and wound healing. J Theor Biol 1986;119:219-234.
Wells AF, Lundin Å, Michaelsson G. Histochemical localization of hyaluronan in psoriasis, allergic contact
dermatitis and normal skin. Acta Derm Venereol (Stockh) 1990;71:232-238.
Wood LC, Jackson SM, Elias –0,Grunfeld C, Feingold KR. Cutaneous barrier perturbation stimulates cytokine
production in the epidermis of mice. J Clin Invest 1992;90:482.
Yang GP, Lim IJ, Phan TT, Lorenz HP, Longaker MT. From scarless fetal wounds to keloids: Molecular studies
in wound healing. Wound Rep Reg 2003;11:411-418.
Zhang L, Underhill CB, Chen L. Hyaluronan on the surface of tumour cells is correlated with metastatic
behaviour. Cancer Res 1995;55:428-33.





























