Role of Stem Cells in the Pathogenesis of Chronic Obstructive
Pulmonary Disease and Pulmonary Emphysema
Irene Coppolinoa , Paolo Ruggeria , Francesco Nuceraa , Mario
Francesco Cannavoa , Ian Adcockb, Giuseppe Girbinoa , and Gaetano
Caramoria
aDipartimento di Scienze Biomediche, Unita Operativa Complessa
di Pneumologia, Odontoiatriche e delle Immagini Morfologiche e
Funzionali (BIOMORF), Universita degli Studi di Messina, Messina,
Italy;
bAirways Disease Section, National Heart and Lung Institute,
Royal Brompton Hospital Biomedical Research Unit, Imperial College,
London, UK
Author for correspondence:
Gaetano Caramori
Dipartimento di Scienze Biomediche,
Unita Operativa Complessa di Pneumologia, Odontoiatriche e delle
Immagini Morfologiche e Funzionali (BIOMORF),
Universita degli Studi di Messina,
Messina 98122,
Italy.
Email: [email protected]
KEYWORDS
Stem cell; COPD; pulmonary emphysema; fibrocytes; lung;
endothelial progenitor cells; mesenchymal stem cells; adipose
tissue-derived stem cells
ABSTRACT
There are only few human translational studies performed in the
area of stem cell research in patients with chronic obstructive
pulmonary disease (COPD) and/or pulmonary emphysema. Before
progress to clinical trials with stem cells we strongly believe
that more human translational studies are essential, otherwise, the
clinical rationale would be solely based on limited in vitro and
animal studies. In the future, stem cell therapy could be a
treatment for this incurable disease. As of now, stem cell therapy
is still to be considered as an area of active research, lacking
any strong rationale for performing clinical trials in COPD.
Although stem cells would be likely to represent a heterogeneous
population of cells, the different cell subsets and their
importance in the pathogenesis of the different clinical phenotypes
need to be fully characterised before progressing to clinical
trials. Moreover, the potential side effects of stem cell therapy
are underestimated. We should not ignore that some of the most
deadly neoplasms are arising from stem cells.
Introduction
Chronic obstructive pulmonary disease (COPD) is defined as a
“preventable and treatable disease that is characterised by
persistent respiratory symptoms and airflow limitation that is due
to airway and/or alveolar abnormalities usually caused by
significant exposure to noxious particles or gases” (1). COPD is
currently a leading cause of morbidity and mortality worldwide
affecting an estimated 174.5 million people in the world and
accounting for more than three million deaths annually (2). From
1990 to 2015, the all-age prevalence of COPD increased by 44.2% and
the age-standardised prevalence decreased by 14.7%. The mortality
from COPD worldwide in 2015 increase of 11.6% compared with 1990,
despite a decrease in the age-standardised death rate of 41.9% (2).
The burden of COPD estimated in 63.9 million disability-adjusted
life years (DALYs) represented 2.6% of entire global burden of
disease (GBD) in 2015 and COPD ranked eighth among the 315 GBD
causes in 2015. Age standardised DALY rates due to COPD decreased
of 43.7% in 2015 compared to 1990, mostly in countries with high
middle SDI (2). The burden of COPD is projected to increase in the
coming decades due to continued exposure to COPD risk factors and
the changing age structure of the world population (1).
The aetiology of COPD is due to complex interactions between
environmental factors (particularly cigarette smoking) and genetic
factors. Long-term cigarette smoking is currently the cause of more
than 90% of COPD in Westernised countries, whereas other factors,
such as burning biomass fuels for cooking and heating, may be
important causes of COPD in developing countries (3). Only ~25% of
chronic smokers develop symptomatic COPD by the age of 80 years,
suggesting a genetic component, but the influence of single gene
polymorphisms is weak and the only clearly established, albeit
rare, genetic risk factor for COPD is 1-antitrypsin deficiency
(1-AT). However, there are, so far, very few studies comparing
cigarette-smoking associated COPD and other causes of disease, for
this reason, our review of the literature will be limited to the
cigarette-smoking associated COPD.
We review here the evidence for a role of stem cells in the
pathogenesis of COPD and pulmonary emphysema, both during stable
phase and exacerbations. We will also review the potential role of
stem cells in the pathogenesis of complications of the COPD such as
lung cancer and secondary pulmonary hypertension.
Stem cells have been identified in nearly all adult tissues,
including human lungs (4) and are thought to contribute to tissue
maintenance and repair. These are rare unspecialised cells that are
often localised in specialised niches within tissue. Importantly,
those cells exhibit self-renewal capacity and can evoke daughter
progenitor or transit amplifying cells. A progenitor cell, like a
stem cell, tends to differentiate into a specific type of cell. A
stem cell, however, is already more specific and is pushed to
differentiate into its target cell. Stem cells can replicate
indefinitely, whereas progenitor cells can only divide a limited
number of times. Both stem and progenitor cells may give rise to
differentiated cells of the organ (5).
Stem cells do not only have the ability to self-renew but also
give rise to subsequent generations with variable degrees of
differentiation capacities. This offers significant potential for
the generation of tissue that could potentially replace diseased
and damaged areas in the human body.
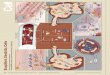
As the lung is a complex organ composed of more than 40
different differentiated cell types, identification of endogenous
progenitor cells has been challenging, and it seems clear that
different progenitor cell populations are localised in different
anatomic regions of the adult human lungs (see Figure 1 for an
example of the lung epithelial stem cells).
Although symptomatic care for COPD has improved over the years,
as of now no knowledge exists of any disease modifying drug
applicable for the treatment of this disease and new options are
desperately needed. As COPD is characterised by loss of lung tissue
and remodelling of the airways, there is growing enthusiasm for
using stem cells to regenerate alveolar tissue and remodel lower
airways and thereby restore lung function in patients with COPD.
Indeed, it could be argued that the destructive, inflammatory
processes induced by smoking that ultimately lead to COPD, could
result from a failure of the regenerative processes in the lungs.
Thus, the homeodynamic process maintaining lung structure and
function could be preserved by enhancing lung regeneration. Recent
studies conducted in animals have revealed that human clonal lung
stem cells may contribute to distal lung tissue regeneration (4),
therefore, administration of stem cells deriving from exogenous
sources may be an innovative way to treat COPD. The main question
is whether we already know enough to start using stem cells to
treat patients with COPD.
Role of stem cells in animal models
In animal models (proteases-treated and transgenic [tight-skin
or Tsk] mice) of pulmonary emphysema, the transplantation of
wild-type mouse bone marrow cells (bone marrow- derived stromal
cells) into emphysematous mice decreases emphysematous lesions in
chimeric mice (6–10). In one study, the complete reversal of
emphysematous lesions was obtained and the transplanted lungs
resembled those of age-controlled normal mice with non-significant
engraftment of donor-derived cells in chimeric mice lungs (8).
Some, but not all, studies performed in animals and humans
suggest that after transplantation bone marrow-derived cells can
migrate to the lungs and create cellular chimerism in the lower
airways’ epithelium. However, the number of these cells is very low
(11–19). Interestingly in animal models of pulmonary emphysema,
there is little or no engraftment of donor-derived cells in
chimeric mice lungs (6–10). These data suggest that adult
bone-marrow-derived stromal mesenchymal stem cells (MSCs) and/or
bone marrow mononuclear cells can migrate to injured areas in
lungs, regenerating the pulmonary parenchyma and repairing
pulmonary emphysema (5, 20). However, the mechanisms con- trolling
this process are unknown and no data is available yet on the
effects of bone-marrow-derived and other stem cells in animal
models of COPD.
Human translational studies in stem cell research in COPD and
pulmonary emphysema
Role of the basal stem cells of the lower airways
The human bronchial basal cells express in vivo on their surface
CD44 (cell-surface glycoprotein) (21), CD49f (integrin subunit
alpha 6 or ITGA6) (22), CD151 (tetraspanin, a member of the
transmembrane 4 superfamily), CD166 (activated leukocyte-cell
adhesion molecule [ALCAM]) a surface marker of all the epithelial
bronchial cells and of the epithelium of the ducts of the bronchial
glands (23), the nerve growth factor receptor (NGFR) (24), the
epidermal growth factor receptor (EGFR, a tyrosine kinase receptor
that is enriched in human airway basal cells and is implicated in
regulation of tissue repair) (25), and in their cytosol the
cytoskeletal cytokeratin (CK) CK5 (22), CK14 (26, 22) and CK17
(27), tissue-factor (TF) and in their cytosol and/or nucleus the
protein Trp-63 (p63) (24). The p63 is a transcription factor of the
p53 family. The human p63 gene encodes at least eight different
isoforms that are generated by transcription from two different
promoters in combination with alternative splicing: there are
transactivating isoforms (TAp63) that contain an NH2-terminal
activation domain (TAD), and truncated isoforms (Np63) lacking the
TAD. TAp63 and DNp63 share a DNA binding domain and an
oligomerisation domain. Only TAp63 and Np63 share a sterile alpha
domain and a transactivation inhibitory domain at carboxy-terminal
region (28, 29). The transactivating isoforms may have functions
like p53 in inducing cell-cycle arrest and apoptosis, whereas Np63
can act inhibiting the activity of p53. The basal cells of human
bronchi and bronchioli constitutively express in their nuclei high
levels of p63, and the Np63 isoforms are the main isoforms
expressed (30, 31). It has been hypothesised that the main
physiological role of p63 is to antagonise p53 functions to ensure
the survival and renewal of basal cells by contrasting apoptosis
and cell cycle arrest (30). Indeed, in the p63-null mice, the
tracheal epithelium is devoid of basal cells (28) and, in vitro,
knockdown of p63 in the human bronchial basal cells reduces
significantly their proliferation and migration with impaired
capacity to generate a pseudostratified-like bronchial epithelium
(31).
Functionally, the basal cells of the human tracheobronchial
epithelium are considered stem cells because both in vivo (in
xenografts) and in vitro are able to reconstitute a fully
differentiated mucociliary epithelium. For example, the human
tracheal basal cells (CD44+/CK5+/p63+) (obtained from healthy
tissue of lung transplants of three donors non- smokers)
xenotransplanted in the denuded rat trachea after five weeks gave
rise to morphologically well-differentiated epithelia and
occasionally submucosal glands (21). Likewise, the transplantation
of human bronchial basal cells in trachea and intrapulmonary
airways of mice post-naphthalene injury gave rise to completely
differentiated epithelium (32).
In vitro, both the basal cells (CD44+/CD166+/NGFR+) of the human
tracheal epithelium and of the ducts cells of tracheal submucosal
glands (CD166+) have the same efficiency in self-renew and
differentiation (23). In vitro, a sub-set of basal cells
CD151-/CK5+/CK14+/TF+ has a high mitotic index (22). Again, in
vitro human bronchial basal cells (CD49f+/NGFR+/p63+) are capable
of self-renewal and to generate ciliated and secretory cells
(24).
In vitro the epidermal growth factor (EGF) stimulating the its
receptors (EGFRs) on the bronchial basal cells (obtained from
bronchial [third and fourth order] brushings of non-smokers and
smokers) changes their differentiation with expression of squamous
cell markers (CK14, CK6, involucrin) and mesenchymal cell markers,
like vimentin, and impairs their ability to generate the junctional
apical complex, necessary for epithelial permeability (25).
Interestingly, tobacco smoking increases EGF expression in ciliated
cells of the bronchial epithelium (25).
The human basal cells (CK5+), isolated by brushings in third and
fourth order of bronchial generation from non-smokers, after
exposure to cigarette smoke extract (CSE) in vitro have an
increased expression of amphiregulin (AREG, an EGF-like growth
factor), an increased proliferation and expression of Ki-67 (marker
of the proliferating cells), and of the major secretory mucins
(MUCs) MUC5AC, and MUC5B, that is prevented with a selective
inhibitor for EGFR. The same basal cells, treated with AREG, have
decreased expression of tight junctions and of the genes implicated
in the ciliogenesis (33).
In vitro, human basal cells (CK5+/p63+), obtained from bronchial
(third and fourth order) brushings of non-smokers and COPD patients
overexpressing the trophoblast cell sur- face antigen 2 (TROP2)
have an increased proliferation rate (34). TROP2 is a transmembrane
glycoprotein highly expressed in many cancers, it has stem
cell-like qualities because provides signals for cellular survival
and proliferation (35).
The treatment in vitro of the human basal cells (CK5+) of
bronchiolar epithelium (10th–12th generations), obtained by
brushing from healthy non-smokers, with EGF or with CSE,
up-regulates genes normally expressed in the large air- ways, as
uroplakin 1B (UPK1B), transcobalamin 1 (TCN1), interleukin 1
receptor type 2 (IL1R2) whereas down-regu- lates the expression of
genes typical of the small airways, like secretoglobin family 3A
member 2 (SCGB3A2), lacto- transferrin (LTF), matrix Gla protein
(MGP), folate receptor 1 (FOLR1), mimicking the phenotype observed
in the bronchiolar epithelium of smokers (36).
In vitro, the basal cells (CK5+/CK14+/p63+) obtained from
bronchial biopsies of stable COPD patients produce significantly
less clone-forming cells and ciliated cells (acetylated tubulin
ACT+), but more basal cells (CK5+) and MUC5B+ cells compared with
control subjects (37).
Likewise, in vitro the basal stem cells (CD151+/CK5+/p63+),
obtained by brushing of the small airway epithelium (from 10th to
12th bronchial order) from COPD patients, have reduced capacity to
successfully differentiate in a normal mucociliary epithelium
compared to smokers with normal lung function and to non-smokers:
at the 28th day of culture, the number of secretory cells
(secretoglobin 1A1, SCGB1A1+) was significantly lower in the
cultures derived from basal cells of COPD patients compared with
nonsmokers samples, whereas were found similar numbers of ciliated
cells (-tubulin IV+) (38).
In vivo, it has compared the gene expression of basal cells
(CD151+/CK5+/p63+) obtained from brushing of large airways from
lifelong non-smoking subjects and smokers with normal lung
function: 676 genes have been identified differentially expressed
in bronchial basal cells with 662 genes significantly up-regulated
in smokers with normal lung function (13 of these 662 genes are
localised in chromosome 19q13.2). Among the most up-regulated were
NF-B inhibitor beta (NFKBIB), egl-9 family hypoxia-inducible factor
2 (EGLN2), latent transforming growth factor beta binding protein 4
(LTBP4), and transforming growth factor beta 1 (TGFB1) (39).
Another study has compared the gene expression of basal cells
obtained by bronchial (up to the sixth bronchial generation) vs.
small airways (from 10th to12th bronchial generation) brushings of
stable COPD and control subjects, both smokers with normal lung
function and non-smokers. The small airway epithelium in smokers
with normal lung function, and more in smokers with COPD, showed
down-regulation of bronchiolar genes, as LTF, SCGB3A2, MGP, FOLR1,
SRY-box 9 (SOX1), GATA binding protein 6 (GATA6), and others, and
up-regulation of uroplakin 1B (UPK1B), carcinoembryonic
antigen-related cell adhesion molecule 5 (CEACAM5), transcobalamin
1 (TCN1), and interleukin 1 receptor type 2 (IL1R2), leading to
shift towards the transcriptomic pattern normally displayed in the
epithelium of large bronchi (33).
In vivo, AREG was up-regulated in the basal cells
CK6+(cytokeratin 6, expressed in squamous differentiation) obtained
from bronchial epithelium (third and fourth order) by both smokers
with normal lung function and COPD subjects (33).
It was shown that the expression of TROP2 in airway basal cells
(CK5+/p63+), obtained in lung samples by lobectomy or pneumonectomy
performed for medical reasons, was significantly increased in COPD
patients and in smokers with normal lung function compared to
lifelong non-smokers. The expression of TROP2 was significantly
correlated with the expression of Ki-67+ in epithelium from COPDs
(34).
The number of bronchial basal cells (CK5+/CK14+/p63+) obtained
by endobronchial biopsies at the secondary bronchus, was
significantly decreased in COPD patients versus control subjects.
It has been identified a subset of non-COPD subjects, former and
current smokers, with lower basal stem cells count and lower
airflows, that may represent an early, pre-diagnostic stage of
COPD, indicating that basal stem cells exhaustion is involved in
COPD pathogenesis (37).
Role of the submucosal bronchial gland stem cells
The duct cells of the human submucosal bronchial glands express
CK5, CK14, p63, and -smooth muscle actin (-SMA) (40). Human
bronchial epithelial cells xenotransplanted in denuded rat trachea
produce many cell clones with different differentiation capacity
including glandular cells (41).
Despite duct glandular stem cells (CD166+/CD44-/NGFR-) have been
isolated from human trachea (23) and similar cells (p63+, CK5+,
CK14+) have been obtained from autopsy lung specimens of patients
with obliterative bronchiolitis due to lung transplantation (42),
there are no studies about these cells in patients with COPD and/or
pulmonary emphysema.
Role of the bronchioalveolar stem cells
The murine bronchioalveolar stem cells (BASCs) express the
alveolar type 2 cell marker prosurfactant protein C (SP-C), the
club cell marker secretoglobin family 1A member 1 (SCGB1A1), the
stem cells antigen-1 (Sca-1), and CD34 (43). Other authors have
identified these murine BASCs as Sca-1+/CD45.2-/CD31- - (platelet
and endothelial cell adhesion molecule 1, also known as PECAM1,
normally found on endothelial cells, platelets, macrophages,
granulocytes, and lymphocytes) (44).
Murine BASCs in vitro have capacity of multilineage
differentiation and self-renewal (43). In mouse lungs these cells
are found exclusively at the bronchioalveolar ductal junction, are
quiescent in the normal lungs and proliferate in response to lung
injury (45).
The human BASCs are also cadherin-1+ (CAM 120/80 or epithelial
cadherin [E-cadherin] or uvomorulin), SPC+ and vimentin+ (46). In
vitro when these cells are co-cultivated with human lung
fibroblasts produce bronchioalveolar structures (46).
The human lungs show stem cells in niches in the distal airways
with capacity in vivo, transplanted in damaged lung of mouse, of
self-renew, and differentiation in structures of endodermal origin
(epithelial cells) and mesodermal origin (vessels), generating
human bronchioles, alveoli, vessels, and partially restoring the
recipient parenchyma. These cells are CD117+ (proto-oncogene c-kit
or tyrosine-protein kinase kit or mast/stem cell growth factor
receptor [SCFR]; a receptor tyrosine kinase protein) but negative
for haematopoietic cell lineage (CD34, CD45, CD133 [prominin 1, a
transmembrane glycoprotein of haematopoietic cells]), mesenchymal
cell antigen (CD44, CD90 [previously termed Thy-1 cell surface
antigen, because expressed in T cells], CD105 [endoglin, expressed
in human vascular endothelium, in bone marrow proerythroblasts and
in activated monocytes]), mast-cell lineage (CD6, CD29 [integrin
subunit beta 1], CD49) and epithelial cells lineage (p63, pan-CK,
SP-C) markers (4). Other authors have found that these CD117+ stem
cells in the early developing human lungs also are CD34+, VEGFR-2+
(vascular endothelial growth factor receptor 2, also termed KDR)
and Tie-2+ (a transmembrane tyrosine-kinase receptor of the
angiopoietins), indicating their endothelial lineage (47).
Recently, in human lungs obtained from subjects ongoing lung
surgery for cancer or nodule have been isolated many different
subsets of CD117+ cells: the mast cells (CD117high/ CD45+/tryptase+
[a serine protease produced purely in the mast cells]) are mainly
located around the peribronchial interstitium and are the majority
(85%) of CD117+ cells; the innate lymphoid cells
(CD117low/CD45+/tryptase-) that represent the 13% of c-kit cells;
the endothelial stem cells (CD117low/CD45-/CD31+/CD34+/tryptase-)
located in the alveolar walls, represent the 1.6% of c-kit
population; the resident stem cells
(CD117low/CD45-/CD31-/CD34-/tryptase-) that represent less than
0.1% of the total cells (48). When stable COPD patients were
compared with smokers with normal lung function and non-smoking
subjects no significant differences of these cells were observed
between these three group of subjects (48).
There are no published studies about the human BASCs in patients
with pulmonary emphysema.
Role of the alveolar stem cells
In mouse models, the subset of alveolar epithelial type 2 cells
(AT2) axin-2+ (a protein regulating the stability of beta-catenin
in the Wnt signalling pathway [regulates selfrenewal and
lineage-specific differentiation in stem cells]) (49) show features
of stem cells (50–52) including self renewal capacity and the
ability to differentiate to alveolar type 1 cells (AT1 cells) (51).
After H1N1 influenza virus injury of mice lungs, there is a large
increase of the number of AT2 and AT1 cells with alveolar
regeneration (52).
The murine alveolar stem cells (AT2) are also epithelial cell
adhesion molecule (EpCAM)+, SP-C+, CD45-, podoplanin- (a small
membrane glycoprotein that belongs to mucin-type proteins, required
for lung development, specifically for maturation of alveolar type
I epithelial cells). After knock-down the enzyme telomerase in
mice, the AT2 cells decreased in association with decreased
alveolar numbers (53).
Human lungs have resident alveolar stem cells that express
markers of human mesenchymal stem cells (hMSCs) (CD73+
[5’-nucleotidase ecto, expressed in lymphocytes B and
T]/CD90/CD105+/vimentin+), surfactant proteins (SPA, SP-C, SP-D) of
the AT2 cells, but are negative for CD117 and for haematopoietic or
endothelial stem cells markers (CD31, CD34, CD45, VEGFR-2) and, in
vitro, have clonogenic ability and can differentiate in AT2 cells.
In normal human lung tissue, these SP-C+/CD90+ cells are located
exclusively in the alveolar walls (54). These human AT2 stem cells
are also transmembrane-4-L-six-family-1 (TM4SF1)+ (a member of the
transmembrane 4 superfamily, also known as tetraspanin family, that
regulates cell movement), anti-AT2+ specific HTII-280 antibody,
EpCAM+ and represent ~29% of the human AT2 total population
(52).
Human alveolar epithelial stem cells (CD90þ, prosurfactant
protein C [pro-SP-C]) exposed in vitro to all-trans retinoic acid,
differentiate in AT1 (aquaporin-5þ[AQP-5]) and AT2 cells (SP-A+)
(55). The same result was obtained stimulating the human alveolar
stem cells (CD90+/pro-SPC+) with Am80 (a synthetic retinoid)
(56).
It was found that phosphoinositide-3-kinase (PI3K)-protein
kinase B (Akt) pathway (pathway that promotes cellular survival in
response to growth factors) is involved in the mechanism of the
human alveolar stem cells differentiation. During in vitro
treatment with all-trans retinoic acid of alveolar stem cells,
which leads to their differentiation, Akt is not phosphorylated
(55). PI3K-inhibitors (e.g. Wortmannin) that block the
phosphorylation of Akt, induces the differentiation of these
alveolar epithelial stem cells in AQP-5+ (AT1) and SP-A+ (AT2)
cells. In addition, in a mouse model of elastase-induced pulmonary
emphysema, lung Akt phosphorylation is enhanced and PI3K-inhibitors
lead to differentiation of the alveolar stem cells (57).
Role of the circulating blood pluripotent haematopoietic stem
cells
A decreased number of circulating CD34þ stem cells has been
found in the blood of patients with moderate to severe stable COPD,
which correlates with hypoxaemia, severity of airflow obstruction
and peak oxygen uptake, and low body mass index (BMI) (58–60).
However, these studies used non-smoking subjects as controls and
not, as in the ideal situation, age-matched smokers with normal
lung function (58–60).
Another study showed that circulating CD34+ stem cells were
significantly decreased both in stable COPD patients and during
COPD exacerbations compared to control smokers with normal lung
function, whereas no significant difference was found between
stable and exacerbated COPD patients. In addition, the number of
CD34+ cells during COPD exacerbations was inversely correlated with
the pulmonary artery systolic pressure (PAPs), the N-terminal
pro-B-type natriuretic peptide serum levels, and with resting heart
rate, and positively correlated with the left ventricular ejection
fraction (61).
Stable COPD patients have a decreased number of circulating
haematopoietic progenitor cells (HPCs) (defined as
CD34+/CD45+/CD133+ or CD34+/CD45+ cells) compared to both smokers
with normal lung function and non-smoking control subjects. Whereas
were not significant differences between the control non-smokers
and smokers with normal lung function, suggesting that the number
of these cells is independent of the smoking habit. In addition,
COPD patients with PAPs >31mmHg have greater number of
CD34+/CD45+/CD133+ cells compared to those with PAPs >31mmHg
(62).
Role of lung mesenchymal stem cells and bone marrow mesenchymal
stem cells
The Mesenchymal and Tissue Stem Cell Committee of the
International Society for Cellular Therapy has provided three
criteria to identify the MSCs in vitro: adherence to plastic when
maintained in standard culture conditions, expression of CD73,
CD90, CD105, and lack of CD11b, CD14, CD19, CD34, CD45, HLA-DR, and
the differentiation capacity into osteoblasts, chondroblasts, and
adipocytes (63).
In vivo, hMSCs are found at low frequency and are defined by
their capacity to differentiate into bone, cartilage, and adipose
tissue, depending on the stimuli and culture conditions under which
they are expanded (64). In addition, under selective conditions of
growth may also differentiate in myogenic cells (65). Lung tissue
(or residents) MSCs (LTMSC) express CD44, CD73, CD90, CD105, CD146
(melanoma cell adhesion molecule) and NG2 (chondroitin sulphate
proteoglycan 4), and lack the expression of haematopoietic,
endothelial and monocyte markers (CD45, CD34, CD14) (66). MSCs can
be isolated from the bronchoalveolar lavage fluid (and are termed
BAL-MSC) of early (within the first 12 post-operative months) lung
transplant recipients, whereas are rarely recoverable in the BAL of
healthy subjects. The immunophenotype of these cells is similar to
those of the LT-MSC (CD73+/CD90+/CD105+/CD45-), but they have lower
differentiation capacity (in particular lack adipogenic
differentiation), increased expression of COL1 and -SMA and express
76 more mRNAs, including those involved in fibroblast activation
(67). In vitro, transforming growth factor beta 1 (TGF-b1)
stimulation of the BAL-MSC increases their expression of -SMA and
COL1 (68). These findings suggest that LT-MSC could acquire a
profibrotic phenotype in pathological conditions (67). These
functional properties explain why lung recipients who develop a
bronchiolitis obliterans syndrome (BOS) have an increased number of
BAL-MSC and that the increase of these cells is a predictor of the
future development of BOS (69).
The bone-marrow MSCs (BM-MSC) are isolated for their expression
of CD29, CD44, CD73, CD90, CD105, and HLAABC, and the absence of
CD31, CD34, CD45, HLA-DR (70). During multiple passages in vitro
these MSC exhibit various surface receptors with different
expression profiles, including C-X-C motif chemokine receptor 3
(CXCR3, also known as CD183), C-X-C motif chemokine receptor 4
(CXCR4, also known as CD184), C-X-C motif chemokine receptor 6
(CXCR6, also known as CD186), C-C motif chemokine receptor 9 (CCR9,
also known as CD199), and C-C motif chemokine receptor 10 (CCR10),
which may be involved in the migration of the MSC to the damaged
tissues (70). The most studied chemokine-chemokine receptor axis in
MSC is C-X-C motif chemokine 12 (CXCL12) (known as stromal
cell-derived factor 1 (SDF1), a potent chemoattractant from bone
marrow of the endothelial progenitor cells [EPCs] and mesenchymal
cells)-CXCR4. But MSC also express adhesion molecules such as C-C
motif chemokine ligand 19 (CCL19) and C-C motif chemokine ligand 21
(CCL21), that are specific ligands of C-C motif chemokine receptor
7 (also known as CCR7 or CD197, expressed in B- and T-lymphocyte
cell), and may also involve in the migration of MSC (70, 71).
In vitro, human lung-MSC have functional properties similar to
BM-MSC: including capacity of self-renewal and to differentiate
into multiple mesodermal lineages: osteocytes, chondrocytes,
adipocytes, and myofibroblasts. Also, these cells have the same
inhibitory effect on T cell proliferation and this may contribute
to a local anti-inflammatory capacity (49, 66, 72). Human lung-MSC
are considered important regulators of tissue repair following
injury (49), through production of cytokines and growth factors
such as fibroblast growth factor-10 and hepatocyte growth factor
(HGF) (46). Human BM-MSC (CD29+/CD44+/CD90+/CD105+), isolated from
bone marrow aspirates from the iliac crest of a 40 years old male
healthy volunteer, in co-culture with human endothelial cells,
produce VEGF, inhibiting the apoptosis of the endothelial cells
(73).
In mouse models of elastase-induced pulmonary emphysema, there
is a significant increase of number of lung-MSC (CD44+/CD73+/CD90+)
(74). In animal models of pulmonary emphysema, lung-MSC or BM-MSC
administrated intravenous or intratracheal, contribute to repair of
the lung damage increasing the levels of epidermal growth factor
(EGF), HGF, VEGF, reducing airway inflammation through the
down-regulation of cyclooxygenase-2 and the inhibition of release
of proteases by inflammatory cells, induce the proliferation of AT1
and AT2 cells and improve lung function (72, 75–79).
The BM-MSC (CD73+/CD90+/CD105+/CD14-/CD34-/CD45-) obtained from
stable COPD patients have reduced levels of CXCR4 mRNA, of its
specific ligand CXCL12, of CCL19 and CCL21 compared to control
smokers with normal lung function. These findings suggest that in
stable COPD there could be an impaired migration capacity of the
stem cells from the bone marrow (80). The human BM-MSC obtained
from patients with stable COPD and pulmonary emphysema compared to
control non-smoking subjects have similar immunophenotype
(CD31+/CD34+/CD45+/CD73+/CD80+/CD90+/CD105+ HLA- DR+),
proliferation, differentiation, migration capacity, but the BM-MSC
from pulmonary emphysema patients have a significantly higher
potential to differentiate in adipocytes than the BM-MSC isolated
from the control subjects (81).
The human lung fibroblasts, isolated from the lung tissue of
stable COPD patients undergoing surgery for carcinoma, express the
LT-MSC-like markers CD44, CD90, CD105, CD166, HLA class I, but not
CD34, CD45, CD133, VEGFR-2 and in vitro, after stimulation with
TGF-1, have decreased differentiation capacity in myofibroblasts
(these cells express the a-smooth muscle actin) compared to smokers
with normal lung function and non-smoking controls (82). The human
lung fibroblasts isolated from non-smoking controls, cultivated in
vitro with osteoblastic, chondrogenic, and adipogenic culture
media, differentiate into osteocytes (expressing alkaline
phosphatase and osteocalcin), chondrocytes (expressing SRY-box 9
[SOX9], a transcription factor essential for cartilage development)
(83), and adipocytes (expressing the nuclear transcription factor
peroxisome proliferator-activated receptor gamma [PPAR-c], that
regulate adipocytes differentiation (84) and the enzyme lipoprotein
lipase). The capacity of these human lung fibroblasts to
differentiate into osteocytes is almost absent in COPD patients and
is significantly decreased in control smokers with normal lung
function compared to non-smoking subjects, whereas no differences
were observed between these 3 groups in their differentiation
capacity into chondrocytes and adipocytes (82).
Role of adipose-derived stromal stem cells
Adipose tissue-derived MSC (ADMSC), as BM-MSC, are
mesoderm-derived cells able to differentiate into multilineage
connective tissue (including osteogenic, chondrogenic, adipogenic,
and myogenic cells) but adipose tissue contains a much higher
number of MSCs than bone marrow (85, 86), in fact, while BM-MSC
constitute 0.0001–0.01% of all BM nucleated cells, the adipose
tissue contains 100.000 MSCs/g of fat (87).
In vitro, human ADMSCs, as BM-MSCs, express CD29, CD44, CD73,
CD90, CD105, and lack CD14, CD34, CD45, CD133, whereas vascular
cell adhesion molecule 1 (VCAM-1 or CD106) is more expressed in
BM-MSC (88). However, other authors have reported that 50% of ADMSC
express CD34 and almost all express CD49e (a5 integrin) (89). In
addition, ADMSCs have higher proliferative capability (88), retain
differentiation potential for a longer period in culture and have
increased immunomodulating capacity compared to BM-MSCs (90, 91),
secreting various immunomodulating molecules including HGF, VEGF,
and interleukin-10 (IL-10) (85).
Human ADMSCs, as murine ADMSCs, administered to mice with
elastase-induced pulmonary emphysema, accumulate in their lungs for
long time, express growth factors such as fibroblast growth
factor-2 (FGF2) and HGF, reduce matrix metalloproteases secretion
and have decreased emphysema (92–94). Human ADMSCs, pre-treated in
vitro with pioglitazone (a PPAR- agonist) and infused intravenously
in mice with elastase-induced and smoke-induced pulmonary emphysema
improve alveolar regeneration and increase lung tissue levels of
FGF2, HGF, and VEGF compared with the same untreated cells
(95).
In an animal model of pulmonary emphysema induced by the
treatment of mice with a VEGFR neutralising antibody intravenous
infusion of the human ADMSCs attenuates cell death (measured by IHC
quantification of active caspase-3) suggesting that in the lung
human ADMSCs have an anti-apoptotic role (96).
Role of circulating endothelial stem cells
Bone-marrow-derived EPCs, capable in vitro cultures of
transforming into mature, functional endothelial cells, can be
isolated from peripheral blood mononuclear cells (97–99). These
cells express on their surface both the endothelial cell markers
CD31 and CD34 (haematopoietic progenitor cell antigen), but lack
the expression of CD14 (monocyte differentiation antigen), CD41a
(integrin alpha chain 2b, mainly expressed on
megakaryocyte-platelet lineage), CD45 (protein tyrosine phosphatase
receptor type C, originally called leukocyte common antigen [LCA]),
CD133 (prominin 1 or AC133; mainly expressed haematopoietic stem
cells and cancer cells), CD235a (glycophorin A;expressed on
erythrocytes and erythroid precursors) (100, 101). However, this
field remains controversial, because other authors, using flow
cytometry, have found that also the
CD31+/CD34+/CD45dim/CD133+/CD14-/CD41a-/CD235a-cells have
angiogenic properties in vitro (102). To further confuse the field
other studies have defined, the circulating EPCs as
CD34+/CD45dim/VEGFR-2+ cells (103). These CD34+/CD133+/VEGFR-2+þ
cells are also expressing other endothelial cell markers such as
the von Willebrand factor (vWF) and the endothelial nitric oxide
synthase (NOS3) (104).
In vitro, the human endothelial colony forming units (CFUs)
cells usually express CD31, CD144 (also termed vascular
endothelial-cadherin [VE-cadherin] or cadherin 5, type 2) VEGFR-2
and vWF, but lack the expression of CD3 (a pan-lymphocytic marker)
and CD45 (105). These cells also express CD184 (CXCR4) (106).
Circulating EPCs seem to serve as a reserve pool of cells to
replace dysfunctional/damaged endothelium in mature blood vessels
(99) and reduced levels of circulating EPCs are associated with
increased risk of death in patients with stable coronary artery
disease (107). Furthermore, the subjects with higher Framingham
risk score have lower endothelial CFUs (108). The patients with
heart failure have lower number of circulating CD34+/VEGFR-2+ cells
and a lower count of these cells is an independent predictor of
increased mortality (109).
Currently, the contribution of the EPCs to the pathogenesis of
COPD has not been fully understood because the studies did not use
the same phenotypic definition of these cells (100). In addition,
many drugs such as statins (110) and systemic glucocorticoids
(111), hypoxaemia (112) and comorbidities (systemic arterial
hypertension, diabetes mellitus, increased age, smoking, and plasma
triglycerides) may influence the number and the function of
circulating EPCs (113) and not all the studies reviewed below have
provided all these details.
Previous studies have suggested a role for an increased
apoptosis of the endothelial cells in the pulmonary capillaries of
the alveolar septa in the pathogenesis of human pulmonary emphysema
(114, 115). In addition, evidence for autoimmune-induced
destruction of endothelial cells has been reported in the lung of
COPD patients (116) providing a further rationale that endothelial
cell loss is a driver of pulmonary emphysema. In animal models,
circulating endothelial stem cells, characterised by the
concomitant expression of CD34, CD133, and VEGFR-2, may contribute
to the repair of lung damage (19). Thus, one can hypothesise that a
decrease in the blood of EPCs may contribute to the pathogenesis of
pulmonary emphysema and/or COPD in humans. Indeed, some studies
(58, 60), but not all (117), have found a significant reduction of
the number of circulating EPCs in patients with stable COPD.
Conversely, this suggests that increased blood EPCs may prevent
pulmonary emphysema.
Again, the number of CD34+/VEGFR2+ cells and their proliferative
ability in vitro, were significantly decreased in stable COPD
patients with PAH (>25 mmHg) compared to both COPD patients
non-PAH and control subjects, whereas there were not significant
differences between the COPD non-PAH patients and control subjects.
Also, the number of these cells was negatively correlated with the
pulmonary artery pressure, suggesting that the number of EPCs could
be related to involvement of pulmonary arteries. All the subjects
had no cardiovascular disease, diabetes mellitus, and did not use
statins, whereas in COPD patients the levels of triglycerides and
arterial hypertension were significantly higher than those in
non-COPD subjects (118).
We have investigated by flow cytometry the number of total
(CD34+) and endothelial stem (triple positive for
CD34/CD133/VEGFR-2) cells in the peripheral venous blood of
age-matched smokers with or without pulmonary emphysema and with or
without COPD, all the subjects were in stable phase (119).
The presence and the severity of pulmonary emphysema were
determined using high resolution computed tomography (HRCT) scans
of the chest with density mask and the National Emphysema Treatment
Trial (NETT) Research Group score (0–4). We found a significant
correlation between the absolute number of circulating CD34+ cells
and the absolute number of circulating endothelial stem cells.
Also, there was a significant correlation between the percentage of
circulating endothelial stem cells and the number of pack-years
smoked, however, no significant correlation was found between total
and endothelial stem cells number and HRCT score of pulmonary
emphysema or lung function data. These data indicate that the
number of circulating endothelial stem cells is not related to the
presence and/or severity of the pulmonary emphysema or the presence
or absence of COPD in stable phase (119).
Using immunohistochemistry another study has demonstrated an
increased number of CD133+ cells in the pulmonary arteries of
patients with mild to moderate COPD compared with a small number of
smokers with normal lung function (120). CD133+ cells are localised
to the endothelial surface and in the vessel wall and their number
in the intima shows a direct relationship with the pulmonary artery
wall thickness (120), indicating their potential for a role in the
pathogenesis of pulmonary artery remodelling and pulmonary
hypertension, however, more research is mandatory in this area
before we can reach firm conclusions.
Another study showed that the number of CD34+/CD133+/VEGFR-2+
cells and of CD34+/VEGFR-2+ cells from stable COPD patients and
from control subjects was similar, and that, in vitro, the
endothelial CFUs was increased in COPD patients compared to
controls. There were no differences between COPD patients and
control subjects in cardiovascular comorbidities (121).
Again, the number of circulating CD34+/CD45dim/VEGFR-2+ cells
and CD34+/CD45dim cells obtained from stable COPD patients was
similar to that of control subjects; instead, the number of
CD34+/CD45+/CD133+/VEGFR-2+ cells was significantly reduced in COPD
patients compared to control subjects and was correlated with FEV1.
The number of these cells was also significantly reduced in
patients with percentage of emphysema >26, whereas there was no
correlation between the levels of CD34+/CD45dim/VEGFR2+ cells and
percentage of emphysema. However, both COPD patients and control
subjects used statins (122).
Circulating CD34+/VEGFR2+ cells and, in particular
CD34+/CD133+/VEGFR2+ cells are reduced in stable severe COPD
patients compared to controls and the number of
CD34+/CD133+/VEGFR2+ cells, but not of CD34+/VEGFR2+ cells, was
significantly and inversely correlated with the severity of the
emphysema. These results were adjusted for confounders such as
smoking status, diabetes mellitus, arterial hypertension, BMI, and
statin use (123).
The dynamic of blood CD34+/CD133+/VEGFR2+ cells had been
measured in COPD patients and control subjects during thoracic
surgery for lung cancer. The number of these cells in the blood is
not affected by thoracic surgery in stable COPD patients. Whereas
in control subjects their number increases 2 hours after starting
surgery and returns to the preoperative levels 24 hours later. In
addition, in vitro, the endothelial CFUs cells from control
subjects increased 2 hours after the operation started, whereas
that from COPD patients unchanged (124).
The blood CD34+/CD133+/VEGFR2+ cells of stable COPD patients
show, in vitro, a reduced proliferation capacity and significantly
impaired migratory capacity in response to CXCR12 compared to those
isolated from non-smoking control subjects (125). Likewise,
circulating CD34+/CD133+/VEGFR2+ cells are decreased, have impaired
proliferative capacity and express lower levels of NOS3 in stable
COPD patients compared to controls non-smoking subjects. However,
there were subjects with systemic arterial hypertension and with
coronary heart disease and other comorbidities and statin use were
not mentioned (104).
Others found a lower number of circulating CD34+/CD45dim/CD133+
cells from stable COPD patients compared to control subjects and
showed an increase of these cells in sputum of COPD patients,
suggesting that the lowering of these cells in the blood could be
due to an increased recruitment in the lungs. However, 25% of COPD
patients had arterial hypertension and hyperlipidaemia, the 17%
used statins and one had cardiovascular disease, whereas the
control subjects had not these comorbidities (126).
Another study shows that blood CD34+/CD31+/CD133+/CD144- cells,
of patients with stable COPD with pulmonary emphysema
differentiate, in vitro, in significantly fewer endothelial CFUs
cells compared to those obtained from the blood of COPD patients
without emphysema, smokers with normal lung function and
non-smoking controls. A significant inverse correlation between the
number of endothelial CFUs cells and the degree of pulmonary
emphysema was observed. Moreover, the EPCs from both group of COPD
patients (emphysema and non-emphysema) showed significantly lower
migration after human recombinant VEGF stimulation compared with
those obtained from non-smoking controls. No patient was receiving
statins, but there are subjects with systemic arterial hypertension
and diabetes mellitus included in the study (127).
The endothelial CFUs from blood samples of stable COPD patients
and of smokers with normal lung function show, in vitro, a slower
growth rate and display significantly increased
senescence-associated -galactosidase activity, increased expression
of p21 and p16 and decreased expression of SIRT1 (sirtuin-1, an
important regulator of genomic stability and inhibitor of
endothelial senescence) compared to controls non-smokers. Only one
COPD patient had diabetes mellitus and one arterial hypertension,
whereas in all groups statins were used (105).
Role of circulating fibrocytes
Fibrocytes are blood stem cells which originate in the bone
marrow and express CD11b (integrin subunit alpha M; expressed on
the surface of monocytes/macrophages, granulocytes, activated
lymphocytes, a subset of NK cells, a subset of dendritic cells),
CD13 (alanyl aminopeptidase membrane, expressed on granulocytes,
endothelial cells, epithelial cells, macrophages), CD14, CD34,
CD45, CXCR4, and major histocompatibility complex class II, but
lack the expression of lymphocyte markers such as CD3, CD4, CD8,
CD19, and CD25; they secrete the type 1 and 3 collagens,
fibronectin, and vimentin (128). A large proportion of the
fibrocytes also express CCR2 (also known as CD192, C-C motif
chemokine receptor 2; produced by endothelial cells, smooth muscle
cells, and macrophages), CCR3 (also known as CD193, C-C motif
chemokine receptor 3, expressed in eosinophils and lymphocytes),
and some express CCR7 (129). In the normal subjects, the fibrocytes
constitute approximately 0.5% of circulating leukocytes (128).
In animal models, as well as in patients with pulmonary
fibrosis, these blood fibrocytes are able to differentiate in vitro
into lung fibroblasts (130). Fibrocytes now are recognised as
mesenchymal cells that arise from circulating monocyte precursors
and participate in the innate response to injury where they have
functions of antigen presentation and tissue remodelling (131). In
the damaged tissue, the fibrocytes can adopt the phenotype of
contractile myofibroblast, losing the CD34 and acquiring the
expression of the -SMA (128).
They can induce the differentiation of the mesenchymal cells
into myofibroblasts, can produce proinflammatory cytokines (such as
IL-6 and IL8), could be involved in the stimulation of angiogenesis
with production of matrix metalloproteinases and proangiogenic
factors such as VEGF and platelet-derived growth factor (PDGF)
(132, 133). The fibrocytes differentiation is reduced by interferon
, tumour necrosis factor , IL-12, serum amyloid P (SAP), and is
augmented by IL-4, IL-13, TGF-1 and endothelin-1 (ET-1) (134).
Two different populations of fibrocytes may be identified: the
classical phenotype CD34+/CD45med/CD14+/collagen type I alpha 1
chain (COL1)+ and the myeloid-derived suppressor cell-like
fibrocytes CD34-/CD45dim/CD14-/COL1+(135).
So far, there are few studies on the number of these fibrocytes
in the peripheral blood of patients with COPD. The level of
myeloid-derived suppressor cell-like fibrocytes
(CD34-/CD45dim/CD14-/COL1+ cells), obtained by blood of stable COPD
patients is significantly increased compared to control subjects,
whereas the number of circulating fibrocytes
(CD34+/CD45med/CD14+/COL1+) is similar between COPD patients and
controls. Also, the number of fibrocytes is not related with lung
function measured by spirometry (135).
There is only one study quantifying the number of fibrocytes in
the lung tissue. The fibrocytes (CD34+/COL1+) are been found
located in subepithelial region of the proximal airways (obtained
from healthy areas of lung resection for cancer) and their number
for mm2 is similar between COPD patients and control subjects
(135).
Role of the satellite cells of the skeletal muscles
Satellite cells are undifferentiated myogenic precursor cells,
localised beneath the basal lamina and the myofiber plasma
membrane. These cells are heterogeneous, but most of them express
the transcription factors involved in myogenesis and skeletal
muscle differentiation termed paired box 3 (Pax3) and 7 (Pax7),
myogenic factor 5 (Myf5), BARX homeobox 2 (Barx2), the molecules
CD34, CXCR4 (136) and the adhesion molecules cadherin 15
(M-cadherin, adhesion molecule highly expressed in developing
skeletal muscle), neural cell adhesion molecule (NCAM) (137). Of
these, Pax7 is the best biomarker as it is expressed in all
quiescent and proliferating human satellite cells (136).
Satellite cells are usually quiescent but may be re-activated
with their proliferation and differentiation into myoblasts after
severe muscle injuries (e.g. extensive physical activity or
exposure to mycotoxins) cause (136).
There are various studies on the satellite cells in skeletal
muscle of stable COPD patients. Many stable COPD patients have a
dysfunction of their skeletal muscles, and it has been hypothesised
that is caused by an impaired muscle regeneration (138). In the
vastus lateralis muscle, the number of satellite cells (Pax7+)
obtained by needle biopsy of stable COPD patients with mid-thigh
muscle cross-sectional area (MTCSA) >70cm2, has been compared
with stable COPD patients with MTCSA <70cm2 and with control
non-smoking subjects without showing any significant differences
between groups. However, the proportion of skeletal muscle fibres
with central nuclei (considered newly formed myofibres) is
increased in COPD patients with MTCSA >70cm2 (138).
Another study has confirmed that the number of Pax7+ in the
biopsies from vastus lateralis muscle is not different at the
baseline between stable COPD patients compared with control
subjects. But after 8 weeks of high-intensity resistance training,
these satellite cells tend (p=0.07) to increase in both groups at
24 hours remaining elevated at week 8 (139).
It has been hypothesised that the satellite cells may be
senescent in stable COPD patients, in fact, the maximal telomere
length of satellite cells (NCAM+/Pax7+) obtained from vastus
lateralis muscle of control subjects is significantly increased
compared to stable COPD patients with MTCSA >70cm2 and COPD
patients with MTCSA <70cm2 (140).
In vitro, the satellite cells (Pax7+) obtained from vastus
lateralis muscle of stable COPD patients show increased
proliferation rates compared to those obtained from control
subjects and a significant increase of Myf5 compared to those from
controls (138).
In another study, the satellite cells (NCAM+) obtained from
biopsy of vastus lateralis muscle of COPD patients in vitro show
similar proliferation and differentiation capacity compared to
those of control subjects, but build myotubes with reduced diameter
with increased expression of proteins, such as myostatin and
atrogin-1, that inhibit myogenesis and promote muscle atrophy
(141).
In vitro, the mRNA expression of markers of autophagy
[microtubule associated protein 1 light chain 3 alpha (LC3),
sequestosome 1 (SQSTM1) and BCL2 interacting protein 3 (BNIP3)] is
significantly increased in myoblasts from stable COPD patients
compared with healthy subjects (142).
Role of the above structural and circulating lung stem cells in
the pathogenesis of COPD exacerbations
Interestingly, the number of blood CD34+/VEGFR2+ cells is
increased in many patients during a severe COPD exacerbation
leading to hospitalisation compared with their phase of clinical
stability (117). However, this may be explained by the blood
leucocytosis seen during COPD exacerbations as the number of EPCs
in stable phase correlated significantly with the number of total
blood leucocytes (119). The functional significance of this
observation is unknown. Cardiovascular morbidity and mortality are
increased in patients with COPD exacerbations but remains to be
demonstrated if COPD patients with no increase in their blood EPCs
level during COPD exacerbations have an increased risk for
cardiovascular morbidity and mortality.
Very small embryonic-like stem cells (VSELs)
(CD34+/CD133+/CXCR4+/CD45-) (cells with very small diameter [~5m]
are found in most human tissues including the bone marrow and are
characterised by the expression of several transcription factors
involved in the self-renewal of undifferentiated embryonic stem
cells, such as the octamer binding transcription factor 4 (Oct-4)
and the homeobox transcription factor Nanog [from “Tir Na Nog,” the
mythologic Celtic land of the ever-young]) are pluripotent stem
cells because can differentiate into osteoblasts, chondrocytes,
adipocytes, and neurons (143, 144). These cells are increased in
the blood of COPD patients during their severe exacerbations (with
arterial oxygen saturation [SaO2] ≤92%), compared to a different
cohort of exacerbating COPD patients (SaO2 >92%) and control
subjects, suggesting that the hypoxaemia may induce the
mobilisation of these cells (145).
The level of circulating fibrocytes (CD45+/COL1+) is increased
in COPD patients during exacerbation compared to stable COPD and to
control subjects (129). Interestingly, the exacerbating patients,
two months after exacerbation, had a number of fibrocytes
significantly reduced compared to the exacerbation phase, and the
percentage of fibrocytes was negatively correlated to FEV1. The
same Authors showed that the fibrocytes in vitro have significantly
increased migration when exposed to enhanced concentrations of
CXCL12, and the fibrocytes from blood of COPD exacerbation patients
have higher migration capacity compared to control subjects. The
migration is abolished with antagonist of CXCR4. These results
suggest that during exacerbation the fibrocytes are recruited by
the CXCR4/CXCL12 axis (129).
Pulmonary epithelial stem cells as a target of the carcinogenic
process of lung carcinoma
Lung cancer and COPD are leading causes of morbidity and
mortality worldwide. They share a common environmental risk factor
in cigarette smoke exposure and a genetic predisposition
represented by the incidence of these diseases in only a fraction
of smokers. COPD is also a major independent risk factor for lung
carcinoma, among long-term smokers. Smokers with COPD also have a
higher risk of developing a specific histological subtype of
non-small cell lung cancer termed squamous cell carcinoma (146).
Despite, current data supporting a direct relationship between
proximal airway basal progenitors and cells associated with
carcinogenesis in murine models, the precise stem cell target of
this process in the development of human squamous cell lung
carcinoma remains unknown (146).
In murine models, a pulmonary stem cell population is found at
the bronchioalveolar duct junction. These cells, termed BASCs, are
resistant to bronchiolar and alveolar damage and proliferate during
epithelial cell renewal in vivo and their transformed counterparts
give rise to adenocarcinoma (45). BASCs can be functionally defined
in vivo because of their resistance to chemical (naphthalene)
injury, their infrequent proliferation relative to other progenitor
cell types, and their co-expression of the bronchiolar and alveolar
secretory cell markers club cell secretory protein and
pro-surfactant protein C, respectively. Their immunophenotype is
CD45-, CD31-, CD34-, Sca-llow, and autofluorescencelow (147).
Interestingly, the protein kinase Ciota (PKCiota) is an oncogene
required for maintenance of the transformed phenotype of NSCLC
cells (148). In mouse, the genetic loss of Prkc dramatically
inhibits Kras initiated hyperplasia and subsequent lung tumour
formation in vivo. This effect correlates with a defect in the
ability of Prkc-deficient BASCs to undergo Kras-mediated expansion
and morphologic transformation in vitro and in vivo (148). This
suggests that both squamous cell lung carcinoma and lung
adenocarcinoma may originate from the same stem cells (146). This
is an area which clearly requires more human translational
research.
Stem cell therapy in patients with COPD and/or pulmonary
emphysema
Recently, good evidence has been provided that new lung growth
can occur in an adult human lung. After pneumonectomy serial
computed tomographic scans of the chest has shown progressive
enlargement of the remaining lung and an increase in tissue density
and magnetic resonance imaging with the use of hyperpolarised
helium-3 gas has shown overall acinar-airway dimensions consistent
with an increase in the alveolar number rather than the enlargement
of existing alveoli, despite that the alveoli in the growing lung
are shallower than in normal lungs (149). Stem cell treatment is a
type of intervention strategy that leads new cells into damaged
tissue, in order to treat a disease or injury. Many potential stem
cell therapies already exist, but most of them, outside of bone
marrow stem cells transplantation, are yet at an experimental
stage. Nevertheless, before stem cell therapy could be applied in a
clinical setting, more research is deemed necessary to understand
stem cell behaviour upon transplantation, as well as the mechanisms
of stem cell interaction with the diseased microenvironment.
Findings that embryonic stem cells and stem cells deriving from
adult human tissues (4) might be used for the repair and
regeneration of diseased lung tissues, has stimulated extensive
investigations with respect to whether these approaches could be
used for lung diseases. Understanding the identity and roles of
endogenous progenitor cells in the lungs as well as the possible
roles as lung cancer stem cells has increased (4, 5). These are
rapidly moving fields that hold promise for improved comprehension
of lung biology and a potential therapeutic approach for many lung
diseases. For example, although MSCs were initially hypothesised to
be the panacea for regenerating tissues, MSCs appear to be more
important in therapeutics to regulate the immune response invoked
in settings such as tissue injury and autoimmunity (64).
Results of controlled clinical trials of stem cells
transplantation
Tables 1 and 2 are summarised the design and the results,
respectively, for the primary and secondary outcomes of the
clinical trials of stem cell therapies performed in stable COPD
patients whereas Table 3 are summarised for comparison the
minimally clinical important differences (MCIDs) for these
outcomes.
Human bone marrow mesenchymal stem cells
During a phase I, controlled trial of PROCHYMAL (ex vivo
cultured adult hMSCs produced by Osiris Therapeutics;
www.osiristx.com) in patients with acute myocardial infarction, an
improvement, in FEV1 and forced vital capacity (FVC) has been
observed through an as yet unknown mechanism (150). These data have
stimulated a multicentre, double-blind, placebo-controlled phase II
trial of PROCHYMAL for patients with moderate-severe COPD
(FEV1/FVC, <0.70, 30% ≤FEV1 post-BD ≤70%), which was initiated
in May 2008. The primary goal of the trial is to determine the
safety of MSCs infusions in patients with COPD. The trial has
recruited 62 patients in 6 participating US sites. No toxicities or
significant adverse events were reported post-infusion. Notably, a
significant decrease in the circulating inflammatory marker
C-reactive protein (CRP), which is commonly increased in COPD
patients, was noted in treated patients as was a trend towards
improvement in quality of life indicators but without any
significant improvement in lung function, in 6-minute walking
distance (6MWD), Borg dyspnoea scale, and number of COPD
exacerbations (www.osiristx.com). The trial ended in the third
quarter of 2010 (151).
A phase I open label and prospective study, performed in the
Netherlands, has enrolled 10 patients with severe pulmonary
emphysema who undergone a lung volume reduction surgery (LVRS) in
two separate occasions. At the time of first surgery, the bone
marrow was collected, the mesenchymal cells were isolate and
expanded in vitro; after 6–10 weeks, the patients received two
autologous BM-MSC infusions at a distance of one week, followed by
the second LVRS. In the first 48 hours and 3 weeks after the second
MSC infusion, no symptoms or toxicity were observed. At 12-month
follow up, the FEV1 had increased from baseline of 390mL (p=0.03;
minimal clinically important difference [MCID] =100–140mL), the
residual volume decreased by 540mL (p=0.053) and the body weight of
all patients significantly increased of mean 4.6Kg (p=0.016). In
addition, after BM-MSC infusion the immunohistochemical analysis of
the resected lung tissue, has shown a threefold increased
expression of the endothelial marker CD31 (p=0.016) in the alveolar
septa (152).
Granulocyte colony-stimulating factor (G-CSF), a haematopoietic
growth factor stimulates the proliferation and mobilisation of bone
marrow haematopoietic cells to the peripheral blood, including an
increase in myeloid progenitor cells and CD34+ stem cells which are
possibly involved in the regeneration of damage tissues in the
lungs (59, 153–156).
In an uncontrolled phase I clinical trial, performed in Brazil,
G-CSF was administered immediately prior to bone marrow harvest and
then bone marrow mononuclear cells were isolated and infused into a
peripheral vein in patients with advanced COPD (stage IV with
dyspnoea) (157). The 12-month follow-up showed an absence of
significant adverse effects but no proliferation of CD34+ and
CD133+ cells (157). The same four patients were followed for 3
years. The authors stated that this treatment with autologous bone
marrow mononuclear cells is safe, however, one patient died 12
months after the administration and one died 27 months after. The
other two patients showed improved FVC, whereas FEV1 remained
stable (158).
Ten patients with severe pulmonary emphysema have been enrolled
in phase I, prospective, patient-blinded, placebo-controlled study,
performed in Brazil from December 2013 and October 2014. All
patients were subjected to bronchoscopic lung volume reduction with
placement of endobronchial valves (EBV), but immediately before,
five patients received allogeneic BM-MSC administration in the
subsegmental bronchi whereas the remaining five patients received
saline solution only. In the MSC-EBV group, no patient experienced
adverse events and the serum CRP levels were significantly reduced
(p<.05) at days 30 and 90 compared to saline-EBV group, without
significant differences in FEV1, FVC, total lung capacity (TLC),
arterial oxygen partial pressure, arterial carbon dioxide partial
pressure, single-breath carbon monoxide diffusing capacity (DLCO),
BMI, and 6MWD. The body mass index, airflow obstruction, dyspnoea
and exercise capacity (BODE) index and St. George’s Respiratory
Questionnaire (SGRQ) score were significantly decreased compared to
baseline (dimensionless unit of BODE index from 8 to 3;
dimensionless unit of SGRQ from 68 to 30, MCID = reduction of 4 or
more units) at days 90 in the patients of the MSC-EBV group
(159).
Human adipose-derived stromal stem cells
A phase I, open label clinical study performed in Mexico, has
enrolled 12 severe stable COPD patients to test the safety of
intravenous administration of autologous stromal vascular fraction.
This contains ADSMC [30–40%], vascular endothelial cells, smooth
muscle cells, pericytes, various circulating cells such as
leucocytes, and not require culture expansion (91) immediately
after taking it from the adipose tissue. During the infusion and at
12th month of follow-up no adverse events were observed and the
SGRQ score was decreased from 73 units at baseline to 45 at 3
months (p=0.005) ant to 44 at 6 months after this treatment
(160).
There are no other published studies treating COPD/pulmonary
emphysema patients with human adipose-derived stem cells, but it
has been published the case report of the intravenously delivery of
human autologous adipose-derived MSCs (CD44+/CD73+/CD90+/CD105+),
in a patient with lung injury after long-term exposure to sulphur
mustard with improvement of 6MWT, as well as SGRQ, and Borg Scale
Dyspnoea Assessment, but not of the FEV1, and without adverse
effects (161).
Ongoing clinical trials of stem cell therapies
Table 4 are summarised the ongoing clinical trials of stem cell
therapies for the treatment of patients with COPD/pulmonary
emphysema.
Because the adipose-derived MSCs are effective in animal models
of pulmonary emphysema and are easy to isolate, most of these
ongoing clinical trials are using these cells (162, 163).
New potential approaches to regenerative medicine
Lasers typically generate electromagnetic radiation which is
relatively uniform in wavelength, phase and polarisation, and
classical medical applications of lasers are considered “high
energy” because of their intensity, which ranges from about 10 to
100 W. Low level lasers (LLLs) elicit effects through non-thermal
means and may activate endogenous pulmonary stem cells and
neoangiogenesis which is optimal for stem cell growth. LLLs are
currently in clinical trials for acute myocardial infarction and
stroke and have been proposed as regenerative photoceutical
treatment for COPD (164).
Side effects of stem cell therapies
The analysis of serious adverse events (including deaths) in
four clinical trials of COPD treatment with stem cells has found
that the total (combination of non-fatal adverse events and deaths)
serious adverse events are more common in cell therapy group
compared to controls, with a 1.5 and 1.3-fold increased odds ratios
for death and non-fatal serious adverse events respectively
(165).
In the clinical trial with the highest number of patients (151),
27 patients treated with stem cells and 28 in the placebo-group
experienced an adverse event in the 2 years of follow-up. Most of
the adverse events are of mild or moderate intensity and was not
reported any fatal adverse event. The most common adverse events
are infections (bronchitis and pneumonia, urinary tract infection),
respiratory disorders (dyspnoea, cough, and respiratory failure),
cardiac disorders, gastrointestinal disorders, dizziness, and
lethargy.
Ethical issues of the stem cell therapies
Stem cell treatment for COPD is associated with certain risks
that have been clearly identified, primarily by using allogenic
bone marrow transplantation. Graft versus host disease, tumour
formation, inappropriate migration of cells to other areas of the
body, and immune system rejection have been observed. Stem cell
therapy for COPD is in its infancy, but even so, its development
already brings great hope to many patients who have exhausted other
forms of COPD treatment. For this reason, it is mandatory that all
the medical institutions worldwide enrol patients with COPD into
controlled clinical trials on experimental stem cell therapy. In
the lay world, there are high expectations of flexible cells that,
in theory, can develop into a lot of cell types, but laymen do not
know that there are many types of stem cells. Moreover, it not
clears to the public that experimental stem cell therapy requires
extensive preparation and monitoring in the laboratory for these
cells to develop the correct features. Importantly, greater
evidence-based studies are essential along with a greater
understanding of the stem cell growth conditions required to obtain
optimal success/growth particularly as some of the most deadly
neoplasms arise from stem cells (166).
That COPD patients decide to try this therapy is not surprising
when patients have exhausted other forms of COPD treatment and only
have, for example, in cases of very severe COPD, two more years to
live and can read on the Internet about the possibilities
available. There is a great need for government and
supra-governmental action in this area to prevent people from
taking such therapies until adequate controlled research is
performed. However, the banning of stem cell therapy in some EU
countries since 2007 has excluded controlled clinical experiments
in some excellent academic centres.
Conclusions
There are only few human translational studies performed in the
area of stem cell research in patients with COPD and/or pulmonary
emphysema. Before progress to clinical trials with stem cells we
strongly believe that more human translational studies are
essential, otherwise, the clinical rationale would be solely based
on limited in vitro and animal studies. In the future, stem cell
therapy could be a treatment for this incurable disease. As of now,
stem cell therapy is still to be considered as an area of active
research, lacking any strong rationale for performing clinical
trials in COPD.
Although stem cells would be likely to represent a heterogeneous
population of cells, the different cell subsets and their
importance in the pathogenesis of the different clinical phenotypes
need to be fully characterised before progressing to clinical
trials. Moreover, the potential side effects of stem cell therapy
are underestimated. We should not ignore that some of the most
deadly neoplasms are arising from stem cells.
Declaration of interest
The authors report no conflict of interest.
References
1. Global Initiative for Chronic Obstructive Lung Disease.
Global Strategy for the Diagnosis, Management and Prevention of
Chronic Obstructive Pulmonary Disease. Latest update 2018;1–123.
goldcopd.org.
2. GBD 2015 Chronic Respiratory Disease Collaborators. Global,
regional, and national deaths, prevalence, disability-adjusted life
years, and years lived with disability for chronic obstructive
pulmonary disease and asthma, 1990–2015: a systematic analysis for
the Global Burden of Disease Study 2015. Lancet Respir Med.
2017;5(9):691–706. doi:10.1016/S2213-2600(17)30293-X.
3. Caramori G, Casolari P, Barczyk A, Durham AL, Di Stefano A,
Adcock I. COPD immunopathology. Semin Immunopathol.
2016;38(4):497–515. doi:10.1007/s00281-016-0561-5.
4. Kajstura J, Rota M, Hall SR, Hosoda T, D’Amario D, Sanada F,
Zheng H, Ogorek B, Rondon-Clavo C, Ferreira-Martins J, et al.
Evidence for human lung stem cells. N Engl J Med. 2011;
364(19):1795–1806. doi:10.1056/NEJMoa1101324.
5. Sueblinvong V, Weiss DJ. Stem cells and cell therapy
approaches in lung biology and diseases. Transl Res. 2010;
156(3):188–205. doi:10.1016/j.trsl.2010.06.007.
6. Adachi Y, Oyaizu H, Taketani S, Minamino K, Yamaguchi K,
Shultz LD, Iwasaki M, Tomita M, Suzuki Y, Nakano K, et al.
Treatment and transfer of emphysema by a new bone marrow
transplantation method from normal mice to Tsk mice and vice versa.
Stem Cells. 2006;24(9):2071–2077. doi:10.1634/stemcells.
2005-0575.
7. Ishizawa K, Kubo H, Yamada M, Kobayashi S, Numasaki M, Ueda
S, Suzuki T, Sasaki H. Bone marrow-derived cells contribute to lung
regeneration after elastase-induced pulmonary emphysema. FEBS Lett.
2004;556(1–3):249–252. doi:10.1016/S0014-5793(03)01399-1.
8. Spees JL, Olson SD, Ylostalo J, Lynch PJ, Smith J, Perry A,
Peister A, Wang MY, Prockop DJ. Differentiation, cell fusion, and
nuclear fusion during ex vivo repair of epithelium by human adult
stem cells from bone marrow stroma. Proc Natl Acad Sci USA.
2003;100(5):2397–402. doi:10.1073/pnas.0437997100.
9. Yuhgetsu H, OhnoY, Funaguchi N, Asai T, Sawada M, Takemura G,
Minatoguchi S, Fujiwara H, Fujiwara T. Beneficial effects of
autologous bone marrow mononuclear cell transplantation against
elastase-induced emphysema in rabbits. Exp Lung Res.
2006;32(9):413–426. doi:10.1080/01902140601047633.
10. Zhen G, Liu H, Gu N, Zhang H, Xu Y, Zhang Z. Mesenchymal
stem cells transplantation protects against rat pulmonary
emphysema. Front Biosci. 2008;13:3415–3422. doi:10.2741/2936.
11. Bittmann I, Dose T, Baretton GB, M€uller C, Schwaiblmair M,
Kur F, Leohrs U. Cellular chimerism of the lung after
transplantation. An interphase cytogenetic study. Am J Clin Pathol.
2001; 115(4):525–533. doi:10.1309/GAFN-5MPA-LY8E-DTPQ.
12. Kleeberger W, Versmold A, Rothamel T, Glockner S, Bredt M,
Haverich A, Lehmann U, Kreipe H. Increased chimerism of bronchial
and alveolar epithelium in human lung allografts undergoing chronic
injury. Am J Pathol. 2003;162(5): 1487–1494.
doi:10.1016/S0002-9440(10)64281-2.
13. Kotton DN, Ma BY, Cardoso WV, Sanderson EA, Summer RS,
Williams MC, Fine A. Bone marrow-derived cells as progenitors of
lung alveolar epithelium. Development. 2001;128(24): 5181–5188.
https://www.ncbi.nlm.nih.gov/pubmed/11748153
14. Kotton DN, Fabian AJ, Mulligan RC. Failure of bone marrow to
reconstitute lung epithelium. Am J Respir Cell Mol Biol.
2005;33(4):328–334. doi:10.1165/rcmb.2005-0175RC.
15. Krause DS, Theise ND, Collector MI, Henegariu O, Hwang S,
Gardner R, Neutzel S, Sharkis SJ. Multi-organ, multi-lineage
engraftment by a single bone marrow-derived stem cell. Cell.
2001;105(3):369–377. doi:10.1016/S0092-8674(01)00328-2.
16. Lama VN, Smith L, Badri L, Flint A, Andrei AC, Murray S,
Wang Z, Liao H, Toews GB, Krebsbach PH, et al. Evidence for
tissue-resident mesenchymal stem cells in human adult lung from
studies of transplanted allografts. J Clin Invest. 2007;
117(4):989–996. doi:10.1172/JCI29713.
17. Schrepfer S, Deuse T, Reichenspurner H, Fischbein MP,
Robbins RC, Pelletier MP. Stem cell transplantation: the lung
barrier. Transplant Proc. 2007;39(2):573–576.
doi:10.1016/j.transproceed.2006.12.019.
18. Suratt BT, Cool CD, Serls AE, Chen L, Varella-Garcia M,
Shpall EJ, Brown KK, Worthen GS. Human pulmonary chimerism after
hematopoietic stem cell transplantation. Am J Respir Crit Care Med.
2003;168(3):318–322. doi:10.1164/rccm.200301-145OC.
19. Yamada M, Kubo H, Kobayashi S, Ishizawa K, Numasaki M, Ueda
S, Suzuki T, Sasaki H. Bone marrow-derived progenitor cells are
important for lung repair after lipopolysaccharide induced lung
injury. J Immunol. 2004;172(2):1266–1272.
https://www.ncbi.nlm.nih.gov/pubmed/14707105
20. Weiss DJ, Kolls JK, Ortiz LA, Panoskaltsis-Mortari A,
Prockop DJ. Stem cells and cell therapies in lung biology and lung
diseases. Proc Am Thorac Soc. 2008;5(5):637–667.
doi:10.1513/pats.200804-037DW.
21. Van de Laar E, Clifford M, Hasenoeder S, Kim BR, Wang D, Lee
S, Paterson J, Vu NM, Waddell TK, Keshavjee S, et al. Cell surface
marker profiling of human tracheal basal cells reveals distinct
subpopulations, identifies MST1/MSP as a mitogenic signal, and
identifies new biomarkers for lung squamous cell carcinomas. Respir
Res. 2014;15:160. doi:10.1186/s12931-014-0160-8.
22. Ghosh M, Ahmad S, Jian A, Li B, Smith RW, Helm KM, Seibold
MA, Groshong SD, White CW, Reynolds SD. Human tracheobronchial
basal cells. Normal versus remodeling/repairing phenotypes in vivo
and in vitro. Am J Respir Cell Mol Biol. 2013;49(6):1127–1134.
doi:10.1165/rcmb.2013-0049OC.
23. Hegab AE, Ha VL, Darmawan DO, Gilbert JL, Ooi AT, Attiga YS,
Bisht B, Nickerson DW, Gomperts BN. Isolation and in vitro
characterization of basal and submucosal gland duct stem/progenitor
cells from human proximal airways. Stem Cells Transl Med.
2012;1(10):719–724. doi:10.5966/sctm.2012-0056.
24. Rock JR, Onaitis MW, Rawlins EL, Lu Y, Clark CP, Xue Y,
Randell SH, Hogan BLM. Basal cells as stem cells of the mouse
trachea and human airway epithelium. Proc Natl Acad Sci USA.
2009;106(31):12771–12775. doi:10.1073/pnas.0906850106.
25. Shaykhiev R, Zuo WL, Chao IW, Fukui T, Witover B, Brekman A,
Crystal RG. EGF shifts human airway basal cell fate toward a
smoking-associated airway epithelial phenotype. Proc Natl Acad Sci
USA. 2013;110(29):12102–12107. doi:10.1073/pnas.1303058110.
26. Evans MJ, Van Winkle LS, Fanucchi MV, Plopper CG. Cellular
and molecular characteristics of basal cells in airway epithelium.
Exp Lung Res. 2001;27(5):401–415. doi:10.1080/01902140120740.
27. Nakajima M, Kawanami O, Jin E, Ghazizadeh M, Honda M, Asano
G, Horiba K, Ferrans VJ. Immunohistochemical and ultrastructural
studies of basal cells, clara cells and bronchiolar cuboidal cells
in normal human airways. Pathol Int. 1998; 48(12):944–953.
doi:10.1111/j.1440-1827.1998.tb03865.x.
28. Daniely Y, Liao G, Dixon D, Linnoila RI, Lori A, Randell SH,
Oren M, Jetten AM. Critical role of p63 in the development of a
normal esophageal and tracheobronchial epithelium. Am J Physiol
Cell Physiol 2004;287(1):C171–181.
doi:10.1152/ajpcell.00226.2003.
29. Gonfloni S, Caputo V, Iannizzotto V. p63 in health and
cancer. Int J Dev Biol. 2015;59(1–3):87–93.
doi:10.1387/ijdb.150045sg.
30. Chilosi M, Doglioni C. Constitutive p63 expression in airway
basal cells. A molecular target in diffuse lung diseases.
Sarcoidosis Vasc Diffuse Lung Dis. 2001;18(1):23–26.
https://www.ncbi.nlm.nih.gov/pubmed/?term=Constitutive+p63+expression+in+airway+basal+cells.+A+molecular+target+in+diffuse+lung+diseases
31. Arason AJ, Jonsdottir HR, Halldorsson S, Benediktsdottir BE,
Bergthorsson JT, Ingthorsson S, Baldursson O, Sinha S, Gudjonsson
T, Magnusson MK. deltaNp63 has a role in maintaining epithelial
integrity in airway epithelium. PLoS One. 2014;9(2):e88683.
doi:10.1371/journal.pone.0088683.
32. Ghosh M, Ahmad S, White CW, Reynolds SD. Transplantation of
airway epithelial stem/progenitor cells: a future for cell-based
therapy. Am J Respir Cell Mol Biol. 2017;56(1):1–10. doi:
10.1165/rcmb.2016-0181MA.
33. Zuo WL, Yang J, Gomi K, Chao I, Crystal RG, Shaykhiev R.
EGF-amphiregulin interplay in airway stem/progenitor cells links
the pathogenesis of smoking-induced lesions in the human airway
epithelium. Stem Cells. 2017;35(3):824–837.
doi:10.1002/stem.2512.
34. Liu Q, Li H, Wang Q, Zhang Y, Wang W, Dou S, Xiao W.
Increased expression of TROP2 in airway basal cells potentially
contributes to airway remodeling in chronic obstructive pulmonary
disease. Respir Res. 2016;17(1):159–173.
doi:10.1186/s12931-016-0463-z.
35. Shvartsur A, Bonavida B. Trop2 and its overexpression in
cancers: regulation and clinical/therapeutic implications. Genes
Cancer. 2015;6(3–4):84–105. doi:10.18632/genesandcancer.40.
36. Yang J, Zuo WL, Fukui T, Chao I, Gomi K, Lee B, Staudt MR,
Kaner RJ, Strulovici-Barel Y, Salit J, et al. Smoking-dependent
distal-to-proximal repatterning of the adult human small airway
epithelium. Am J Respir Crit Care Med. 2017;196(3):340–352.
doi:10.1164/rccm.201608-1672OC.
37. Ghosh M, Miller YE, Nakachi I, Kwon JB, Baron AE, Brantley
AE, Merrick DT, Franklin WA, Keith RL, Vandivier RW. Exhaustion of
airway basal progenitor cells in early and established chronic
obstructive pulmonary disease. Am J Respir Crit Care Med.
2018;197(7):885–896. doi:10.1164/rccm.201704-0667OC.
38. Staudt MR, Buro-Auriemma LJ, Walters MS, Salit J, Vincent T,
Shaykhiev R, Mezey JG, Tilley AE, Kaner RJ, Ho MW, Crystal RG.
Airway basal stem/progenitor cells have diminished capacity to
regenerate airway epithelium in chronic obstructive pulmonary
disease. Am J Respir Crit Care Med. 2014;190(8): 955–958.
doi:10.1164/rccm.201406-1167LE.
39. Ryan DM, Vincent TL, Salit J, Walters MS, Agosto-Perez F,
Shaykhiev R, Strulovici-Barel Y, Downey RJ, Buro-Auriemma LJ,
Staudt MR, et al. Smoking dysregulates the human airway basal cell
transcriptome at COPD risk locus 19q13.2. PLoS One.
2014;9(2):e88051. doi:10.1371/journal.pone.0088051.
40. Wansleeben C, Barkauskas CE, Rock JR, Hogan BL. Stem cells
of the adult lung: their development and role in homeostasis,
regeneration, and disease. Wiley Interdiscip Rev Dev Biol. 2013;
2(1):131–148. doi:10.1002/wdev.58.
41. Engelhardt JF, Schlossberg H, Yankaskas JR, Dudus L.
Progenitor cells of the adult human airway involved in submucosal
gland development. Development. 1995;121(7): 2031–2046.
https://www.ncbi.nlm.nih.gov/pubmed/?term=Progenitor+cells+of+the+adult+human+airway+involved+in+submucosal+gland+development.
42. Swatek AM, Lynch TJ, Crooke AK, Anderson PJ, Tyler SR,
Brooks L, Ivanovic M, Klesney-Tait JA, Eberlein M, Pena T, et al.
Depletion of airway submucosal glands and TP63+ KRT5+ basal cells
in obliterative bronchiolitis. Am J Respir Crit Care Med.
2018;197(8):1045–1057. doi:10.1164/rccm.201707-1368OC.
43. Kim CF, Jackson EL, Woolfenden AE, Lawrence S, Babar I,
Vogel S, Crowley D, Bronson RT, Jacks T. Identification of
bronchioalveolar stem cells in normal lung and lung cancer. Cell.
2005;121(6):823–835. doi:10.1016/j.cell.2005.03.032.
44. Driscoll B, Kikuchi A, Lau AN, Lee J, Reddy R, Jesudason E,
Kim CF, Warburton D. Isolation and characterization of distal lung
progenitor cells. Methods Mol Biol. 2012;879:109–122. doi:
10.1007/978-1-61779-815-3_7.
45. Kim CF. Paving the road for lung stem cell biology:
bronchioalveolar stem cells and other putative distal lung stem
cells. Am J Physiol Lung Cell Mol Physiol. 2007;293(5):L1092–1098.
doi: 10.1152/ajplung.00015.2007.
46. Kato T, Oka K, Nakamura T, Ito A. Bronchioalveolar
morphogenesis of human bronchial epithelial cells depending upon
hepatocyte growth factor. J Cell Mol Med. 2015;19(12): 2818–2826.
doi:10.1111/jcmm.12672.
47. Suzuki T, Suzuki S, Fujino N, Ota C, Yamada M, Suzuki T,
Yamaya M, Kondo T, Kubo H. c-Kit immunoexpression delineates a
putative endothelial progenitor cell population in developing human
lungs. Am J Physiol Lung Cell Mol Physiol. 2014; 306(9):L855–865.
doi:10.1152/ajplung.00211.2013.
48. Lopez-Giraldo A, Cruz T, Molins L, Guirao A, Cuerpo S,
Ramirez J, Agusti A, Faner R. Characterization, localization and
comparison of c-kit+ lung cells in never smokers and smokers with
and without COPD. BMC Pulm Med. 2018;18(1):123. doi:
10.1186/s12890-018-0688-3.
49. Foronjy RF, Majka SM. The potential for resident lung
mesenchymal stem cells to promote functional tissue regeneration:
understanding microenvironmental cues. Cells. 2012;1(4):874.
doi:10.3390/cells1040874.
50. Hogan B. Stemming lung disease? N Engl J Med. 2018;378(25):
2439–2440. doi:10.1056/NEJMcibr1803540.
51. Nabhan AN, Brownfield DG, Harbury PB, Krasnow MA, Desai TJ.
Single-cell Wnt signaling niches maintain stemness of alveolar type
2 cells. Science. 2018;359(6380):1118–1123. doi:10.1126/
science.aam6603.
52. Zacharias WJ, Frank DB, Zepp JA, Morley MP1, Alkhaleel FA,
Kong J, Zhou S, Cantu E, Morrisey EE. Regeneration of the lung
alveolus by an evolutionarily conserved epithelial progenitor.
Nature. 2018;555(7695):251–255. doi:10.1038/nature25786.
53. Chen R, Zhang K, Chen H, Zhao X, Wang J, Li L, Cong Y, Ju Z,
Xu D, Williams BR, et al. Telomerase deficiency causes alveolar
stem cell senescence-associated low-grade inflammation in lungs. J
Biol Chem. 2015;290(52):30813–30829.
doi:10.1074/jbc.M115.681619.
54. Fujino N, Kubo H, Suzuki T, Ota C, Hegab AE, He M, Suzuki S,
Suzuki T, Yamada M, Kondo T, et al. Isolation of alveolar
epithelial type II progenitor cells from adult human lungs. Lab
Invest. 2011;91(3):363–378. doi:10.1038/labinvest.2010.187.
55. Horiguchi M, Kojima H, Sakai H, Kubo H, Yamashita C.
Pulmonary administration of integrin-nanoparticles regenerates
collapsed alveoli. J Control Release. 2014;187:167–174.
doi:10.1016/j.jconrel.2014.05.050.
56. Sakai H, Horiguchi M, Ozawa C, Akita T, Hirota K, Shudo K,
Terada H, Makino K, Kubo H, Yamashita C. Pulmonary administration
of Am80 regenerates collapsed alveoli. J Control Release.
2014;196:154–160. doi:10.1016/j.jconrel.2014.10.004.
57. Horiguchi M, Oiso Y, Sakai H, Motomura T, Yamashita C.
Pulmonary administration of phosphoinositide 3-kinase inhibitor is
a curative treatment for chronic obstructive pulmonary disease by
alveolar regeneration. J Control Release. 2015;213: 112–119.
doi:10.1016/j.jconrel.2015.07.004.
58. Fadini GP, Schiavon M, Cantini M, Baesso I, Facco M, Miorin
M, Tassinato M, de Kreutzenberg S, Avogaro A, Agostini C.
Circulating progenitor cells are reduced in patients with severe
lung disease. Stem Cells. 2006;24(7):1806–1813. doi:10.1634/
stemcells.2005-0440.
59. Huertas A, Testa U, Riccioni R, Petrucci E, Riti V, Savi D,
Serra P, Bonsignore MR, Palange P. Bone marrow-derived progenitors
are greatly reduced in patients with severe COPD and low-BMI.
Respir Physiol Neurobiol. 2010;170(1):23–31. doi:
10.1016/j.resp.2009.10.003.
60. Palange P, Testa U, Huertas A, Calabro L, Antonucci R,
Petrucci E, Pelosi E, Pasquini L, Satta A, Morici G, et al.
Circulating haemopoietic and endothelial progenitor cells are
decreased in COPD. Eur Respir J. 2006;27(3):529–541.
doi:10.1183/09031936.06.00120604.
61. Liu Y, Liu X, Lin G, Sun L, Li H, Xie C. Decreased CD34+
cell number is correlated with cardiac dysfunction in patients with
acute exacerbation of COPD. Heart Lung Circ. 2014;23(9): 875–882.
doi:10.1016/j.hlc.2014.03.008.
62. Pizarro S, Garcıa-Lucio J, Peinado VI, Tura-Ceide O, Dıez M,
Blanco I, Sitges M, Petriz J, Torralba Y, Marın P, et al.
Circulating progenitor cells and vascular dysfunction in chronic
obstructive pulmonary disease. PLoS One. 2014;9(8):e106163.
doi:10.1371/journal.pone.0106163.
63. Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I,
Marini F, Krause D, Deans R, Keating A, Prockop Dj, Horwitz E.
Minimal criteria for defining multipotent mesenchymal stromal
cells. The international society for cellular therapy position
statement. Cytotherapy. 2006;8(4):315–317.
doi:10.1080/14653240600855905.
64. Singer NG, Caplan AI. Mesenchymal stem cells: mechanisms of
inflammation. Annu Rev Pathol. 2011;6:457–478. doi:10.1146/
annurev-pathol-011110-130230.
65. Beier JP, Bitto FF, Lange C, Klumpp D, Arkudas A, Bleiziffer
O, Boos AM, Horch RE, Kneser U. Myogenic differentiation of
mesenchymal stem cells co-cultured with primary myoblasts. Cell
Biol Int. 2011;35(4):397–406. doi:10.1042/CBI20100417.
66. Ricciardi M, Malpeli G, Bifari F, Bassi G, Pacelli L, Nwabo
Kamdje AH, Chilosi M, Krampera M. Comparison of epithelial
differentiation and immune regulatory properties of mesenchymal
stromal cells derived from human lung and bone marrow. PLoS One
2012;7(5):e35639. doi:10.1371/journal.pone.0035639.
67. Sinclair KA, Yerkovich ST, Chen T, McQualter JL, Hopkins PM,
Wells CA, Chambers DC. Mesenchymal stromal cells are readily
recoverable from lung tissue, but not the alveolar space, in
healthy humans. Stem Cells. 2016;34(10):2548–2558.
doi:10.1002/stem.2419.
68. Walker N, Badri L, Wettlaufer S, Flint A, Sajjan U,
Krebsbach PH, Keshamouni VG, Peters-Golden M, Lama VN. Resident
tissue-specific mesenchymal progenitor cells contribute to
fibrogenesis in human lung allografts. Am J Pathol. 2011;178(6):
2461–2469. doi:10.1016/j.ajpath.2011.01.058.
69. Badri L, Murray S, Liu LX, Walker NM, Flint A, Wadhwa A,
Chan KM, Toews GB, Pinsky DJ, Martinez FJ, et al. Mesenchymal
stromal cells in bronchoalveolar lavage as predictors of
bronchiolitis obliterans syndrome. Am J Respir Crit Care Med.
2011;183(8):1062–1070. doi:10.1164/rccm.201005-0742OC.
70. Nejad-Moghaddam A, Panahi Y, Abdollahpour Alitappeh M, Borna
H, Shokrgozar MA, Ghanei M. Therapeutic potential of mesenchymal
stem cells for the treatment of airway remodelling in pulmonary
diseases. Iran J Allergy Asthma Immunol. 2015;14(6):552–568.
https://www.ncbi.nlm.nih.gov/pubmed/26725553
71. Hocking AM. The role of chemokines in mesenchymal stem cell
homing to wounds. Adv Wound Care (New Rochelle).
2015;4(11):623–630. doi:10.1089/wound.2014.0579.
72. Kruk DMLW, Heijink IH, Slebos DJ, Timens W, Ten Hacken NH.
Mesenchymal stromal cells to regenerate emphysema: on the horizon?
Respiration. 2018: 96(2):148–158. doi:10.1159/000488149.
73. Guan XJ, Song L, Han FF, Cui ZL, Chen X, Guo XJ, Xu WG.
Mesenchymal stem cells protect cigarette smoke-damaged lung and
pulmonary function partly via VEGF-VEGF receptors. J Cell Biochem.
2013;114(2):323–335. doi:10.1002/jcb.24377.
74. Skurikhin EG, Pakhomova AV, Krupin VA, Pershina OV, Pan ES,
Ermolaeva LA, Vaizova OE, Rybalkina OY, Dygai AM. Response of
inflammatory mediators, extracellular matrix proteins and stem and
progenitor cel