Embed Size (px)
Citation preview

Neuropsychologia 41 (2003) 702–712
Implicit face perception in a patient with visual agnosia?Evidence from behavioural and eye-tracking analyses
Sandra Lea,∗, Eric Raufasteb, Sophie Rousselc, Michèle Puela, Jean-François Démoneta
a INSERM U455, Federation of Neurology, CHU Purpan, 31059 Toulouse Cedex, Franceb Work and Cognition Laboratory, University of Toulouse, Toulouse, France
c Laboratoire Jacques Lordat, University of Toulouse, Toulouse, France
Received 14 May 2002; received in revised form 30 September 2002; accepted 30 September 2002
Abstract
This paper investigates face perception in a visual agnosic and prosopagnosic patient (SB). Despite very extensive lesions of visual areas,SB remains capable of some visual processing [Brain 125 (2002) 58]. However, in everyday situations SB does not exhibit signs of specificface recognition. To investigate how SB may process faces, we tested two hypotheses. According to the ‘spared module hypothesis,’ SBsabilities come from spared modules of implicit face processing. According to the ‘general strategy hypothesis,’ SB may have developedsome deliberate compensatory strategies. A two-session experimental design was constructed. In both sessions, face and non-face pictureswere shown to participants. In Session 1 (implicit condition), participants had to decide whether each picture was a vegetable. In Session2 (explicit condition), participants had to decide whether each picture was a face. Verbal reports showed that SB was not aware of faces inSession 1. However, behavioural results showed that (1) SB could process faces; (2) even when SB was not aware of faces, he processedthem differently than non-faces; (3) when knowing the presence of faces, he did not process faces better. In addition, eye-tracking datasuggested that SB did not change the nature of his processing from Sessions 1 to 2. Pupil diameters showed that fixated facial features wereprocessed similarly as in control participants. Together, these results are not compatible with a general compensatory strategy hypothesisand suggest sparing of an implicit face processing module in SB.© 2002 Elsevier Science Ltd. All rights reserved.
Keywords:Face perception; Prosopagnosia; Visual areas; Eye movements
1. Introduction
This paper addresses, in a patient with a face perceptiondeficit, the notion of covert face processing in the absenceof the patient’s awareness, and the possible involvement ofspecific cortical areas in such processes.
Prosopagnosia is a neurological deficit characterised bythe inability to recognise the faces of previously known per-sons in the absence of severe intellectual, sensory or cogni-tive impairments. Although prosopagnosic patients usuallyexperience a total absence of a feeling of familiarity withfaces of known individuals and a failure of overt recog-nition of these faces, it has been shown that the process-ing of familiar faces can still take place in the absence ofthe patient’s awareness. This was demonstrated by physi-ological (skin conductance[3,46], pupillometry [19], andevent related potentials[37,42]) and behavioural indices
∗ Corresponding author. Tel.:+33-5-61-77-9500;fax: +33-5-61-49-9524.
E-mail address:[email protected] (S. Le).
[8,12,39] of recognition showing significant differences inresponse to familiar and unfamiliar faces (see[9] for areview). Bruyer[9] defines a further area of behaviouralstudy for the covert recognition of faces, namely, the record-ing of eye movements[9]. Neurologically-intact perceiverstend to adopt a facial feature scanning strategy[30], i.e.they explore preferentially the ‘eyes’, ‘nose’, and ‘mouth’[48]. The recording of eye movements in two patients withimpaired facial learning and recognition showed that fixa-tion, pursuit, saccades, and scanning of salient features ofscenes and faces were normal[38]. However, the study ofthe transitional properties of scanning revealed that the scanpaths of personally meaningful familiar faces, whether or notthey were consciously recognised, were less predictable thanthose of other faces[38]. This suggested that as autonomicstudies of covert recognition in prosopagnosia, the proper-ties of scanning can be used as an index of higher neuralprocessing.
Evidence from cognitive psychology[7,44,51], computa-tional vision[47], neuropsychology[5,11], and neurophys-iology [16,36] suggests that face and object recognition
0028-3932/02/$ – see front matter © 2002 Elsevier Science Ltd. All rights reserved.PII: S0028-3932(02)00204-X

S. Le et al. / Neuropsychologia 41 (2003) 702–712 703
involve qualitatively different processes that may occur indistinct brain areas. At a neuroanatomical level, prosopag-nosia is commonly associated with bilateral damage tothe region below the calcarine fissure, in the region ofthe fusiform gyrus[10,20,33,41,50]. However, it has beendemonstrated that unilateral damage to the right fusiformgyrus [13–15,28,34,45,49]is sufficient to induce a similardeficit, whilst object recognition (of same difficulty level)may remain more or less spared. On the other hand, im-paired object recognition along with relatively spared facerecognition can be found after damage to the left inferotem-poral cortex, although visual object agnosia is quite oftenassociated with bilateral damage[20,26,32,35].
Recent neuroimaging studies on normal subjects havedemonstrated that faces and objects activate different lociin the same cortical area, usually the right fusiform gyrus[1,2,24,27,31,40]. This specific cortical area is called the“Fusiform Face Area” or FFA[27,31]. In a study with fMRI,Kanwisher et al.[27] concluded that the FFA is selectivelyinvolved in the perception of faces. However, fMRI studieswhere subjects viewed faces as opposed to non-face objects(tasks usually used to reveal the right FFA) showed activa-tions in several occipito-temporal areas, including areas inthe left middle fusiform gyrus[21,22], bilaterally in the an-terior fusiform gyrus[21,40] and a region in the occipitallobe[17,23,25]. As suggested by Gauthier et al.[22], it ap-pears that the area of the occipital lobe that they called the“Occipital Face Area” or OFA, shows the same kind of se-lectivity to faces (albeit weaker) as FFA. As a part of theearly visual areas mediating the first stages of object per-ception, OFA could be considered as a good candidate forspecific detection of faces.
The present study reports the case of a 31-year-old patient(SB) with a severe pattern of visual agnosia following ameningoencephalitis at the age of 3 years[29]. The ensuingcerebral damage was extensive enough to destroy the rightand left ventral streams as well as the right dorsal stream (seedetailed description inSection 2). As a result, SB becameunable to recognise colours, objects, faces, or letters. Butstrikingly, he is very good at many everyday visually guidedactions (see[29] for more details).
With the noticeable exception of OFA, the cortical areassupposedly involved in face perception are lesioned or dis-connected in SB. It is perhaps unsurprising, therefore, thatSB shows a severe prosopagnosia. On the other hand, de-spite the severity of his visual deficits, SB is able to demon-strate, in experimental contexts, unexpected visual abilities(particularly visuomotor[29]) including in face perception.As the lesions occurred when SB was a 3-year-old child,one should consider the possibility of an unusual maturationof some brain areas as well as of compensatory adaptationsto the primary deficits, in addition to normal properties ofundamaged areas (e.g. OFA).
The present study combines behavioural testing andeye-movement analyses to further investigate implicit andexplicit face processing in SB.
2. The patient SB
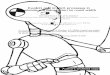
SB is a 31-year-old right-handed man who suffered braindamage as a result of a meningoencephalitis sustained atthe age of 3. MRI conducted in 1998 revealed lesions ofoccipito-parietal and occipito-temporal regions in the righthemisphere, and mainly of the occipito-temporal junctionin the left hemisphere (Fig. 1). The lesions were localisedaccording to atlases from Talairach and Tournoux[43], andDuvernoy[18].
In the right hemisphere, lesions involve most of the ventralvisual areas (i.e. the inferior temporal gyrus, the inferioroccipital gyrus, the fusiform gyrus, and the inferior lingualgyrus). In the dorsal visual areas, lesions involve the middleoccipital gyri and, to a lesser extent, the superior occipitalgyrus. In the temporal lobe, the lesions involve the middletemporal gyrus as well as a limited portion of the superiortemporal gyrus. Partial damage to the right inferior parietallobule is also present in the region of the supra-marginalgyrus. The spared regions in the right occipital pole involvethe posterior inferior part of V2, the primary visual cortex atleast in its rostral and superior aspects and presumably thearea involving the OFA. Among the right parietal regions,the greater part of the angular gyrus and a limited portionof the posterior part of T2 are spared.
In the left hemisphere, lesions are much smaller and con-cern mainly the ventral visual cortex, involving a completedestruction of the ventral part of the occipital-temporal junc-tion (fusiform gyrus). The lesion spreads further dorsallyand partially involves the middle occipital gyrus. On thecontrary to the right hemisphere, occipito-parietal areas arelargely spared by the lesions.
Neuropsychological investigation (see[29] for details) re-vealed a profound visual agnosia, where patient SB showedan inability to recognise faces, colours, words and objectsby sight. The recognition of objects was flawless throughother sensory modalities. SB obtained a verbal IQ of 98 onthe WAIS-R. SBs perception of motion was preserved, aswell as his vision for action, and to a lesser extent his visualimagery and ability to copy certain drawings. In general SBresorts to a slow, feature-by-feature visual strategy when at-tempting to identify objects.
With regard to face recognition, SB showed a severeprosopagnosia. Apart from contextual guessing, SB usedother visual cues to recognise people, such as gait, or silhou-ette. He was not able to readily discriminate between humanfaces and non-face stimuli. It was only after a certain timeand a lot of efforts that SB eventually perceived the char-acteristic features of a face, such as ‘eyes’, ‘nose’, ‘mouth’,etc. Despite this, SB remained totally unable to recognisefacial expressions, nor to discriminate familiar faces fromthose unfamiliar to him. In a face matching task (with im-ages from different viewpoints), like the Benton facial recog-nition test[6], SB obtained a score of 35/54 (65%). Theseperformances are just above chance, which according to theBenton’s test scoring, corresponds to a severe deficit in face

704 S. Le et al. / Neuropsychologia 41 (2003) 702–712
Fig. 1. MRI axial contiguous slices (thickness= 1 mm) parallel to the bi-commissural plane (slice #0) in SB (left hemisphere shown on right) showing themain lesions in this patient. These images confirm the destruction of the right ventral and dorsal stream, and the disconnection of the occipito-temporalpathway in the left hemisphere. Furthermore, it also shows the quasi-isolation of the right occipital pole from the visual output pathways whereas thesuperior portions of the optic radiations are spared.
perception. Therefore, although SB is able to perceive facesto some limited extent, he remains severely impaired in faceidentification. However, a detailed study of SBs behaviourin face processing tasks reveals that his performance maybe well above chance level providing he is given the oppor-tunity to devise strategies to perform the tasks.
We conducted a decision task where SB was presentedwith pictures of normal faces, scrambled faces, and objects.The task of SB was to comment on what he was seeing andto identify the picture when possible. At the beginning, SBdid not recognise faces at all. But after several presentations,and much effort, he seemed to pick out some specific features(like the ‘eyes’) and to adopt a new strategy to explore theitems. Eventually he succeeded in detecting real faces amongother items.
We then conducted a same/different task where SB had tocompare two greyscale photographs of faces. He was asked
to give his responses within 2 s and obtained a score of67/94 (71%), which would correspond to a severe deficit ina normal subject but is rather good when considering SBsgeneral visual deficit and brain damage. In other words, SBwas ‘good’ at comparing details (‘eyes’, ‘ears’, ‘hair’. . . )of two sets of faces, and at deducing whether they wereidentical or not. But this does not mean that SB relied onface processing per se since he did not compare two facestogether but rather two visual details together.
3. Experimental rationale
Although SB seems to use explicit detection of some fa-cial features, the question is to determine whether his abil-ities in face perception derive from an implicit, automaticprocessing that would use some form of spared module,

S. Le et al. / Neuropsychologia 41 (2003) 702–712 705
or whether his abilities result from a more general deliber-ate strategy (based upon detail processing). Therefore, wedecided to design an experiment built on the fact that SBcannot recognise faces as such when he is not cued ex-plicitly. This experiment was divided into two sessions. Ineach, SB was presented with both face and non-face stim-uli. In the first session (Session 1), SB was not told aboutthe presence of faces. In the following session (Session 2),SB was explicitly asked to discriminate faces. Under the‘spared module hypothesis’, SB was expected (1) to processface and non-face stimuli differently during each of the twosessions; (2) to show similar accuracy levels in response tofaces in Sessions 1 and 2, i.e. whether his attention was di-rected to face processing or not. Under the ‘general strat-egy hypothesis’, SB was expected (1) to process face andnon-face stimuli similarly during Session 1, but differentlyduring Session 2; (2) to process faces more accurately inSession 2 than in Session 1. In addition to testing specific hy-potheses with behavioural assessment, the experiment wasdesigned to obtain more specific data on SBs visual explo-ration of faces using eye tracking. Data were also acquiredin age-matched non-brain-damaged participants so as to al-low for comparison between SBs performance and that ofnormal perceivers.
4. Methods
4.1. Participants
In addition to patient SB, nine neurologically normal malestudents (six were right-handed, and three left-handed; meanage,±25.2) from the University Le Mirail (Toulouse) agreedto participate in this experiment.
4.2. Experimental tasks
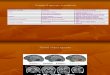
In order to assess SBs behaviour towards faces, a cat-egorisation task was conducted through two different ses-sions. The stimuli were 100 greyscale digitised photographsof faces and 100 greyscale digitised pictures of non-face ob-jects (Fig. 2). The sets of images for the two sessions were
Fig. 2. Examples of stimuli used during Sessions 1 and 2 of the experiment.
different. Each session included a series of 50 faces and 50non-faces. In Session 1, the 50 non-face pictures included25 vegetable and 25 non-vegetable objects. Each stimuluswas presented for an unlimited period of time.
In Session 1, the task was to decide whether the displayeditem was a vegetable or not (“Is it a vegetable?”), whereas inSession 2, the task was to decide whether the stimulus wasa face or not (“Is it a face?”). One should note that SB wasnot told of the presence of faces before the end of Session 1.Between the two sessions, SB was asked (i) whether he hadrecognised any of the stimuli and (ii) whether there wereany faces present in the session. SB answered negatively toboth questions. SBs decision times were recorded with hisresponses, as well as his eye movements.
4.3. Eye-movement recording
Each stimulus was delivered on a dark background, foran unlimited period of time, at the centre of a 21-in. mon-itor with a 640× 480 resolution. Pictures were displayedas large as possible without changing original proportions.Participants sat 90 cm away from the computer screen toallow for eye-movement recording. Thus, they could seepictures having angular heights ranging from 14◦53′ to15◦30′ and angular widths ranging from 10◦31′ to 23◦6′.In order to enable recording of changes in pupil diam-eters, the experimental room was kept dark with only adim neon light. Eye movements were recorded by meansof an eye-tracking system, and were sampled at a rate of50 Hz. The apparatus for eye-movement recording was anASL5000 infra-red eye-tracker, with remote optics, fromApplied Science Laboratory®. By virtue of remote optics,participants were free to move their head (they wore nohelmet) and they could speak as needed for answering ques-tions. Head movements were suppressed in the analysis bymeans of a magnetic head tracker (Flock of Birds®).
4.4. Data analysis
For each picture, both decision times and response accu-racy were recorded. In order to analyse eye movements, fix-ations were computed from eye-position samples using the

706 S. Le et al. / Neuropsychologia 41 (2003) 702–712
eye-tracker manufacturer’s software (‘Eyenal’). Parameterswere set so that elementary samples were aggregated into afixation when at least three consecutive samples were in asame area of 1◦ of visual angle. The (x, y) coordinates ofa fixation represent the median of the aggregated elemen-tary positions. The mean pupil diameter was also recordedby the eye-tracker.
For each face picture, we defined severalAreas of Interest(AOIs), ‘eyes’, ‘nose’, ‘mouth’, ‘ears’, and ‘hair’/foreheadboundary. Each AOI was drawn using either a circle or arectangle, which was placed at the desired location and sizedto the desired proportions (e.g. a circle around an eye). Fix-ations that fell within each of these AOIs were accumulatedso as to produce gaze variables. Two measures were ob-tained for each AOI: thenumber of fixationsthat fell withinthe AOI, and thepupil diametervariation. Under the nullhypothesis, one would expect a positive linear relationshipbetween stimulus size and the number of fixations. Hence,in ANOVAs using number of fixations, the area of the AOIwas introduced as a covariate. Pupil diameter variations werecomputed aszscores using, as mean and standard deviation,the average pupil diameter and the standard deviation ob-tained from the set of all pupil diameters across both sessionsfor each participant. Therefore, pupil diameters coded as be-low zero represent a pupil that was constricted while gazingat the AOI, whereas pupil diameters above zero represent adilated pupil, independently of individual participants’ base-lines of pupil diameter. Under controlled lighting conditions,enlarged pupil diameters can be taken as cues for attentionalor emotional effects (e.g.[4]). A more qualitative approachto visual exploration is to observe when a particular area isfixated, i.e. in which order the areas are fixated. Thus, wecomputed thefirst gaze rankfor each AOI of a given face pic-ture. This variable represents the sequence of participants’fixations on face pictures: low values correspond to areasthat are fixated early, whereas high values correspond to ar-eas that are fixated later. The effects of AOI area on num-ber of fixations and on pupil diameter were partialled outby using residuals of the regression equation with AOI area(relatively to the full screen) as independent variable. Therewas no measurable effect of AOI area on the explorationorder.
5. Results
5.1. Behavioural analyses
5.1.1. Response accuracyDetailed descriptive results about response accuracy ap-
pear inTable 1. During Session 1, SB processed faces betterthan non-face objects, with 88% correct responses to facesversus 60% correct responses to non-faces,χ2 = 10.19;N = 100; P = 0.001. One might argue that correct re-sponses to non-faces in Session 1 include both ‘yes’ and‘no’ answers whereas correct responses to faces include only
Table 1SBs and controls’ response accuracy for Sessions 1 and 2
Stimulus type Number % correct
SB Control group
Session 1 (Is it a vegetable?)Non-faces
Non-vegetable 25 40.0 98.7Vegetable 25 80.0 97.3Total 50 60.0 98.0
Faces 50 88.0 98.0Total
Non-vegetable 75 72.0 98.2Vegetable 25 80.0 97.3Total 100 74.0 98.0
Session 2 (Is it a face?)Non-faces 50 98.0 99.8Faces 50 86.0 99.3Total 100 92.0 99.6
TotalNon-faces 100 79.0 98.9Faces 100 87.0 98.7Total 200 83.0 98.8
‘no’ responses. However, performance within non-face stim-uli for which only ‘no’ responses were correct (pictures thatwere neither faces nor vegetables), was only 40%. Thus,even when restricted to the cases where only the ‘no’ re-sponses were correct, the comparison of face versus non-facestimuli still shows a better performance for face processing(χ2 = 19.05; N = 75; P < 0.001). Normal participantsdid not exhibit these effects with 98% correct responses tofaces and 98% to non-faces. Within Session 2, 86% of SBsresponses to faces were correct versus 98% for non-faces.Since two cells out of four in the contingency table had ex-pected frequencies below 5, phi statistics was used insteadof a chi-square. It showed a significant difference betweenface and non-face performance in Session 2,φ = −2.21;N = 100; P = 0.027. In comparison, the control groupexhibited no significant difference between accuracy of re-sponses to faces and non-faces (P > 0.05).
SB did not produce a different performance on faces inSessions 1 and 2,χ2 = 0.09; N = 100; P > 0.05, as dideight control participants out of nine (allχ2
s < 2.02; Ns =200;Ps > 0.05).
Overall, SBs performance was quite good, significantlybetter than chance within both sessions.
5.1.2. Decision timesOn average, SB produced correct responses approxi-
mately twice as fast as false responses (S.E. = 0.56 s),F(1, 196) = 21.56; P < 0.001. As interpreting RTs forincorrect responses is hazardous, only correct responseswere included in the following analyses. Descriptive resultsabout decision times appear inTable 2.
When comparing face versus non-face stimuli in Ses-sion 1, SBs correct decisions for face stimuli were faster,

S. Le et al. / Neuropsychologia 41 (2003) 702–712 707
Table 2SBs and controls’ decision times for face and non-face stimuli, dependingon the Session (correct responses only)
Stimuli SB Control Group
N Mean RT (s)a S.E. (s) N Mean RT (s) S.E. (s)
Session 1Face 44 3.60 0.37 50 1.24 0.02Object 30 5.81 0.68 50 1.47 0.05
Session 2Face 43 2.89 0.26 50 1.14 0.01Object 49 2.03 0.11 50 1.19 0.01
a RT = reaction times.
t (72) = 3.07; P = 0.003. The same pattern held in normalparticipants, although the difference was only 0.2 s;t (98) =4.229;P < 0.001. All correct responses for faces were ‘no’responses whereas correct responses for non-faces could be‘yes’ or ‘no’. However, when restricted to the subset of stim-uli where only ‘no’ responses were correct, SBs correct re-sponses were still much faster for faces than for non-faces,t (73) = 4.83; P < 0.001.
In Session 2, SBs correct ‘face’ decisions were longerthan his ‘non-face’ decisions,t (90) = 3.00; P = 0.004.The pattern was the same in the control group.
5.1.3. Discussion of behavioural resultsThese results seem in agreement with the spared mod-
ule hypothesis and against the general strategy hypothesis,since performance to face and non-face stimuli was differentwithin each of the two sessions, and performance for facesbetween Sessions 1 and 2 was similar.
It might be objected that the baseline probabilities oftarget items were not the same in both sessions (25% inSession 1 versus 50% in Session 2). However, we do notthink this is a problem, because any effect would haveinduced a difference between the sessions, which can beseen to have not occurred. Therefore, it is unlikely thatdifferences in baseline probabilities can account for ourresults. A second criticism could be the fact that correctresponses to faces were ‘no’ responses in Session 1 and‘yes’ responses in Session 2, thus rendering difficult thecomparison between Sessions 1 and 2. Again, however,the absence of a difference in accuracy argues against thisbeing a problem. Nonetheless, it has been found that sub-jects may have a response bias in favour of saying ‘yes’ or‘no’. Therefore, although the behavioural data have to beinterpreted with caution, we believe that the eye-trackingresults are more informative and can be interpreted withconfidence.
If SB is differentially processing faces by means of a pas-sive ‘implicit’ system, then he would not be expected to scanfacial patterns in a strategic fashion as normal observersdo. He would, however, be expected to process low-levelfeatures normally, and thus to show normal pupillaryresponses.
Table 3Raw eye data for correctly processed face stimuli in SB and controls,depending on the Session (over AOIs)
Sessions SB fixations Control fixations
N Mean S.E. N Mean S.E.
Number of fixations1 220 1.77 0.25 250 0.28 0.022 215 1.97 0.32 250 0.24 0.02
Pupil diameters (fixated AOIs only)1 83 −0.57σ 0.09σ 179 0.05σ 0.05σ2 91 −0.06σ 0.09σ 154 −0.11σ 0.06σ
Pupil diameter is expressed as S.D. units, with zero indicating no change.
5.2. Eye-tracking analyses
Due to a damaged file, eye-movement data are availablefor only eight of the nine control participants. Eye-trackinganalyses are limited to faces that were processed success-fully. Overall descriptive results appear inTable 3.
5.2.1. Number of fixationsIn SB, there was neither an effect of session,F(1, 425) =
0.35;P > 0.05, nor an interaction between session and areasof interest (AOIs),F(4, 425) = 0.48; P > 0.05. However,there was a main effect of the type of AOI, (i.e. ‘hair’,‘eyes’, ‘nose’, ‘mouth’, ‘ears’),F(4, 425) = 59.8; P <
0.001 (Fig. 3). Namely, ‘hair’ was the most fixated area (allpost-hocPs < 0.001). Then ‘eyes’ and ‘nose’ constituted ahomogenous group, with no significant difference betweenthe two. Finally, ‘mouth’ and ‘ears’ were the least fixatedareas.
In the control group there was a significant effect ofthe session for the number of fixations, with significantlymore fixations in Session 1,F(1, 490) = 7.87; P = 0.005.Also, AOIs did not receive the same numbers of fixations,F(4, 490) = 122.5; P < 0.001. Namely, ‘eyes’ were mostfixated (allPs < 0.05) then came ‘nose’ (allPs < 0.001).Then ‘hair’ and ‘mouth’ constituted a homogenous group,with no significant difference between the two. Finally,‘ears’ were the least fixated area (allPs < 0.001).
In the control group there was also a significant interactionbetween sessions and AOI types,F(4, 490) = 7.08; P <
0.001: ‘hair’, ‘nose’ and ‘mouth’ were fixatedlessin Session2 than in Session 1 whereas ‘eyes’ were fixated more inSession 2.
5.2.2. Pupil diameter variationsDescriptive results on pupil diameter variations appear in
Fig. 4.In SB, there was no significant effect of session on pupil
diametersF(1, 164) = 2.57; P > 0.05 (Table 3). On theother hand, there was a significant main effect of the typeof AOIs on pupil diameters,F(4, 164) = 6.53; P < 0.001:‘hair’ was gazed at with significantly smaller pupil diametersthan all other areas (Ps < 0.04) but eyes (P > 0.05). All

708 S. Le et al. / Neuropsychologia 41 (2003) 702–712
Fig. 3. Mean numbers of fixations on the different AOIs for the face stimuli (Sessions 1 and 2) in SB and control participants. Note that the scales ofY-axes are different.
other pair-wise comparisons were not significant (allPs >
0.05).As in SB, there was no significant effect of Session in the
control group,F(1, 323) = 1.62; P > 0.05; and there wasa significant effect of the nature of the AOI on pupil diame-ters,F(4, 323) = 5.69; P < 0.001. By decreasing order ofpupil size, the pattern was ‘ear’, ‘eye’, ‘mouth’, ‘hair’, and‘nose’. However, only three Tukey post-hoc tests reachedsignificance, for the comparison ’eye’–‘nose’ (P < 0.001),
Fig. 4. Pupil sizes (z scores) in the different AOIs for the face stimuli in SB and control participants.
the comparison ‘nose’–‘mouth’ (P = 0.004), and the com-parison ‘nose’–‘hair (P = 0.036). As in SB, there was no in-teraction between AOIs and the Session,F(4, 323) = 1.10;P > 0.05.
5.2.3. First gaze ranksWith regard to gaze ranking, all statistical tests were
non-parametric. AOIs were not gazed by SB in the same or-der (χ2 = 108.3; N = 173; P < 0.001): ‘hair’ areas were

S. Le et al. / Neuropsychologia 41 (2003) 702–712 709
Fig. 5. First gaze ranks in the different AOIs for the face stimuli in SB and control participants. Low values indicate areas that are fixated sooner. Notethat the scales ofY-axes are different.
fixated first, then ‘eyes’, ‘nose’, ‘ear’, and ‘mouth’ (Fig. 5).Applying Mann–Whitney tests to all possible pair-wise com-parisons, ‘hair’ was ranked first against all other AOIs (allPs < 0.001); ‘eye’ came second, allPs ≤ 0.001; finally,‘nose’ came sooner than ‘mouth’ (P = 0.012). In the con-trol group, AOIs were not gazed in the same order (χ2 =13.99; N = 333; P = 0.007): ‘nose’ and ‘hair’ wereranked first, then came ‘eyes’, ‘ears’, and ‘mouth’. UsingMann–Whitney tests, however, the only significant compar-isons were ‘eyes’ fixated before ‘mouth’ (P < 0.004), and‘nose’ before ‘mouth’ (P < 0.001).
Mann–Whitney comparisons of Sessions 1 and 2 weremade, one for each AOI type (e.g. rank of first gaze on ‘hair’,in Session 1 versus in Session 2), but none was significant(all Ps > 0.70). The same pattern was observed in the con-trol group except for ‘eyes’, which were gazed significantlysooner in Session 2 than in Session 1 (P = 0.002). Qualita-tively, it should be noted however, that during the experimentwe could observe that normal participants tended to maketheir decisions while staying centred on one point, generallyin the nose/eyes area.
5.2.4. Discussion of eye-tracking resultsAgain, these results seem in agreement with the spared
module hypothesis and against the general strategy hypoth-esis. Indeed, the eye-tracking analyses reveal no differencebetween SBs behaviour during Sessions 1 and 2, whether weconsider fixation numbers, pupil diameters or explorationorders. In other words, SB looked at the pictures in thesame way whether he was seeking to categorise pictures asvegetable/non-vegetable or as face/non-face. This may ex-plain why his performance on faces is left unchanged by ex-perimental instructions: those instructions were not followed
by a change in the pattern of exploration. On the other hand,systematic differences did appear in control participants.Particularly, they fixated fewer AOIs in Session 2, a fact thatis compatible with the ability to engage in whole-based pro-cessing of faces, an ability presumably not available to SB.
A second point that emerges is a striking difference be-tween SB and neurologically normal participants, regardingthe status of ‘hair’ (e.g.Fig. 3). ‘Hair’ played a big role inSBs visual processing, whereas it played a secondary rolein control participants.Fig. 5shows a pattern of explorationorders in SB that is very different from those of control par-ticipants. This point is far from surprising since SB is knownto rely exclusively on a feature-by-feature visual recogni-tion strategy[29]. He cannot have an exploration guided byproperties involving a whole-based perception and, thus, ex-ploration sequences are organised differently. Our data sug-gest that SB uses a topographical approach to faces, fromhair downwards to mouth. In turn, this might explain theparticular place of ‘hair’ with regard to the number of fixa-tions: ‘hair’ might be looked at more because it would be asystematic entry into the picture. Interestingly,Fig. 4showsthat the pattern of pupil diameters exhibited by SB is notvery different to the pattern exhibited by control participants.This point is striking with regard to the differences found infixation numbers and exploration order. For example, SBspupil diameters on ‘hair’ are quite similar to those of normalparticipants.
6. General discussion
Previous studies showed that despite very extensive le-sions of visual areas, SB remains capable of some visual

710 S. Le et al. / Neuropsychologia 41 (2003) 702–712
processing[29]. This paper investigated the possible originsof his performance in face perception using behavioural andeye-tracking analyses. In order to explain how SB can pro-cess faces we considered two possibilities. According to thespared module hypothesis, SBs abilities come from sparedmodules of normal automatic, implicit, face processing; ac-cording to the general strategy hypothesis, SB makes use ofsome deliberate compensatory strategies. To test these hy-potheses, a two-session experimental design was used wherethe two hypotheses led to two opposite patterns of predictedresults. In both sessions, face and non-face pictures wereshown to the participants. In Session 1, participants had todecide whether each picture was a vegetable or not. In Ses-sion 2, participants had to decide whether each picture wasa face or not. Session 1 required no explicit processing offaces (faces were not the targets), whereas Session 2 did(faces were the targets). Indeed, verbal reports collected be-tween the sessions, showed thatpatient SB was not aware ofthe presence of faces in Session 1. Thus, any potential faceprocessing by SB in Session 1 was necessarily implicit, asrequired for the testing of the hypotheses. Hence, accordingto the spared module hypothesis, faces should be processedin the same way for both sessions but differently than objectsin Sessions 2 and 1. In Session 1, the general strategy hy-pothesis expects the same processing for face and non-facestimuli, whereas the two types of pictures are expected tobe processed differently in Session 2. According to the gen-eral strategy hypothesis, faces should also be processed bet-ter in Session 2 (where SB is aware of their presence) thanin Session 1. In summary, behavioural results showed that(1) SB was quite good at processing faces (given his braindamage); (2) he processed faces differently than non-facestimuli; (3) he did not process faces better in Session 2 thanin Session 1. Eye-tracking data suggested that (1) SB didnot change deeply his processing strategy from Sessions 1to 2, and that (2) once his attentional window is placed on afacial feature (eye, nose,. . . ), this feature is given low-levelprocessing that is not very different to the way normal par-ticipants process the same feature (as shown by the patternsof pupil diameters).
Together, these results are more compatible with the hy-pothesis of a spared module than with the hypothesis of ageneral compensatory strategy hypothesis.
That said, we must remain very cautious because it mightwell be that the stimuli used in this experiment explainsome of the results (e.g. stimuli in Session 1 were notthe same as in Session 2). Also, there might be differ-ences in the spatial-frequency composition of the face ver-sus non-face patterns presented. Further investigations aretherefore needed before asserting with confidence that SB re-tains a spared module. For example, as suggested by anony-mous reviewers, it would be interesting to compare, usingeye-tracking data, SBs processing of normal faces with hisprocessing of scrambled and/or inverted faces.
Most of the studies of prosopagnosia described in the lit-erature relate cases of patients that can perceive faces as
such, but cannot recognise familiar or famous faces. SB isprosopagnosic and, in naıve situations, he cannot even recog-nise that a face is a face. Yet, the results of this experimentsuggested that although SB cannot consciously recognisea face as such, he can infer its non-vegetable nature morereadily than that of other objects (see Session 1). It may besuggested that indeed faces are a particular type of object.But, they are special in the way that they are one of the mostcommon objects we encounter everyday. SB too has exten-sive experience of faces, but his face-selective areas cannotbe located in the ventral visual pathway, as they are for neu-rologically normal perceivers. Instead, such residual visualcompetences of SB in the perception of faces must rely on hisfunctionally spared visual areas. These regions include theprimary visual areas and the left intra-parietal sulcus, sug-gesting the involvement of the left dorsal, occipital-parietalpathway. On the one hand, we have strong reasons to believethat in SB, the dorsal stream is sufficient enough for his ba-sic ‘what’ processing to allow him to perform elementaryperception[29]. Along similar lines, Bauer and Verfaellie[3] suggested that face recognition is mediated by two cere-bral pathways. One would be the ventral visual pathway andwould permit overt recognition, while the other one wouldbe the dorsal visual route (connecting the visual cortex tothe limbic structures via the inferior parietal lobule), andwould support covert face recognition. They argued as wellthat prosopagnosia would result from damage of the ventralpathway, and that those prosopagnosics evidencing physio-logical signs of covert recognition could manifest the pre-served functioning of the dorsal system (see also[9]). Alter-natively, it is also possible that SBs residual vision for faceperception depends primarily on the occipital lobe. Gauthieret al. [22] have demonstrated that part of the occipital lobe,specifically the OFA, seems selectively implicated in faceperception and suggested that it might be involved in thedetection of this specific category of object, i.e. perceivingfaces at the basic level (see also[17,25]).
Like many visual agnosics, SB focuses on parts of ob-jects rather than on objects themselves, whatever the object,including faces. The eye-tracking data reported here cor-roborates this view, with a pattern of face exploration thatseems mainly topographical, from hair/forehead, downwardto mouth. In contrast, normal participants start directly fromrelevant parts like eyes, or stay centred on the nose.
The present observations give insights into how faces canbe processed by a ‘basic’ visual system (such as SBs), whichrelies mainly on its left occipito-parietal pathway and theprimary visual cortex. Hence, we may suggest that, in pa-tient SB, the OFA might be involved in the implicit detec-tion of faces (as in Session 1) and that spared portions ofthe left dorsal stream can then deal in a limited way withits output. However, generalisation to implicit face process-ing in normal perceivers requires further studies because itis possible that SB has developed some variant of face pro-cessing that would take advantage of another spared visualarea, normally unrelated to faces.

S. Le et al. / Neuropsychologia 41 (2003) 702–712 711
SBs case nonetheless suggests that the FFA (lesioned inSB) might not be the only area involved specifically in faceperception. Neuroimaging techniques such as fMRI or ERPswill need to be used to fully elucidate the spatial and tem-poral aspects of face recognition, respectively, particularlyon the role played by the OFA, and should be conducted inparallel so as to complement neuropsychological findings.If nothing else, the results reported here show that neuralmodels of face processing should not be restricted to theFFA but should take into account other areas like the OFA,which might also be implicated in that processing.
Acknowledgements
We wish to thank SB for his always enthusiastic coopera-tion in our research. We gratefully acknowledge commentsfrom Rebecca Gamble, and from two anonymous refereesfor their helpful reviews of the manuscript.
References
[1] Allison T, Ginter H, McCarthy G, Nobre AC, Puce A, Luby M,et al. Face recognition in human extrastriate cortex. Journal ofNeurophysiology 1994;71:821–5.
[2] Allison T, McCarthy G, Nobre AC, Puce A, Belger A. Humanextrastriate visual cortex and the perception of faces, words, numbersand colors. Cerebral Cortex 1994;5:544–54.
[3] Bauer RM, Verfaellie M. Electrodermal discrimination of familiarbut not unfamiliar faces in prosopagnosia. Brain and Cognition1986;8:240–52.
[4] Beatty J. Task-evoked pupillary responses, processing load, and thestructure of processing resources. Psychological Bulletin 1982;91:276–92.
[5] Behrmann M, Winocur G, Moscovitch M. Dissociation betweenmental imagery and object recognition in a brain-damaged patient.Nature 1992;359:636–7.
[6] Benton A, de Hamsher KS, Varney NR, Spreen O. Facialrecognition: stimulus and multiple choice pictures. In: Contributionsto neuropsychological assessment: a clinical manual. Oxford: OxfordUniversity Press, 1983. p. 30–43.
[7] Bruce V, Doyle T, Dench N, Burton M. Remembering facialconfigurations. Cognition 1991;38:109–44.
[8] Bruyer R, Laterre C, Seron X, Feyereisen P, Strypstein E, PierrardE, et al. A case of prosopagnosia with some preserved covertremembrance of familiar faces. Brain Cognition 1983;2:257–84.
[9] Bruyer R. Covert face recognition in prosopagnosia: a review. Brainand Cognition 1991;15:223–35.
[10] Damasio AR, Damasio H, Van Hoesen GW. Prosopagnosia: anatomicbasis and behavioral mechanisms. Neurology 1982;32:331–41.
[11] Damasio AR, Tranel D, Damasio H. Face agnosia and the neuralsubstrates of memory. Annual Review of Neuroscience 1990;13:89–109.
[12] de Haan EH, Young A, Newcombe F. Faces interfere with nameclassification in a prosopagnosic patient. Cortex 1987;23:309–16.
[13] De Renzi E. Prosopagnosia in two patients with CT scan evidenceof damage confined to the right hemisphere. Neuropsychologia1986;24:385–9.
[14] De Renzi E. Current issues are in prosopagnosia. In: Ellis HD, JeevesMA, Newcombe F, Young AW, editors. Aspects of face processing.Dordrecht: Martinus Nijhoff, 1986. p. 243–52.
[15] De Renzi E, Perani D, Cartesimo GA, Silveri MC, Fazio F.Prosopagnosia can be associated with damage confined to the righthemisphere: an MRI and PET study and a review of the literature.Neuropsychologia 1994;32:893–902.
[16] Desimone R. Face-selective cells in the temporal cortex of monkeys:face perception. Journal of Cognitive Neuroscience 1991;3:1–8[special issue].
[17] Dubois S, Rossion B, Schiltz C, Bodart JM, Michel C, Bruyer R, et al.Effect of familiarity on the processing of human faces. NeuroImage1999;9:278–89.
[18] Duvernoy HM. Le cerveau humain. Paris: Springer, 1992.[19] Etcoff N, Freeman R, Cave K. Can we lose memories of faces?
Content specificity and awareness in a prosopagnosic. Journal ofCognitive Neuroscience 1991;3:25–41.
[20] Farah MJ. Visual agnosia: disorders of object recognition and whatthey tell us about normal vision. Cambridge (MA): MIT Press, 1990.
[21] Gauthier I, Tarr MJ, Anderson AW, Skudlarski P, Gore JC. Activationof the middle fusiform ‘face area’ increases with expertise inrecognizing novel objects. Nature Neuroscience 1999;2:568–73.
[22] Gauthier I, Tarr MJ, Moylan J, Skudlarski P, Gore JC, Anderson AW.The fusiform “face area” is part of a network that processes faces atthe individual level. Journal of Cognitive Neuroscience 2000;12:495–504.
[23] Halgren E, Dale AM, Sereno MI, Tootell RB, Marinkovic K,Rosen BR. Location of human face-selective cortex with respect toretinotopic areas. Human Brain Mapping 1999;7:29–37.
[24] Haxby JV, Horwitz B, Ungerleider LG, Maisog JM, Pietrini P, GradyCL. The functional organization of the human extrastriate cortex: aPET rCBF study of selective attention to faces and locations. Journalof Neuroscience 1994;14:6336–53.
[25] Haxby JV, Ungerleider LG, Clark VP, Schouten JL, Hoffman EA,Martin A. The effect of face inversion on activity in human neuralsystems for face and object perception. Neuron 1999;22:189–99.
[26] Hécaen H, Goldblum MC, Masure MC, Ramier AM. Une nouvelleobservation d’agnosie d’objets: déficit de l’association ou de lacatégorisation, spécifique de la modalité visuelle. Neuropsychologia1974;12:447–64.
[27] Kanwisher N, McDermott J, Chun MM. The fusiform face area: amodule in human extrastriate cortex specialised for face perception.Journal of Neuroscience 1997;17:4302–11.
[28] Landis T, Cummings JL, Christen L, Bogen JE, Imhof HG.Are unilateral right posterior cerebral lesions sufficient to causeprosopagnosia? Clinical and radiological findings in six additionalpatients. Cortex 1986;22:243–52.
[29] Le S, Cardebat D, Boulanouar K, Henaff MA, Michel F, Milner D,et al. Seeing, since childhood, without ventral stream: a behaviouralstudy. Brain 2002;125:58–74.
[30] Luria SM, Strauss MS. Comparison of eye movements over faces inphotographic positives and negatives. Perception 1978;7:349–58.
[31] McCarthy G, Puce A, Gore JC, Allison T. Face-specific processingin the human fusiform gyrus. Journal of Cognitive Neuroscience1997;9:605–10.
[32] McCarthy RA, Warrington EK. Cognitive neuropsychology: a clinicalintroduction. San Diego (CA): Academic Press, 1990.
[33] Meadows JC. The anatomical basis of prosopagnosia. Journal ofNeurology, Neurosurgery, and Psychiatry 1974;37:489–501.
[34] Michel F, Poncet M, Signoret JL. Are the lesions responsiblefor prosopagnosia always bilateral? Revue Neurologique (Paris)1989;145:764–70.
[35] Newcombe F, Mehta Z, de Haan EH. Category specificity in visualrecognition. In: Farah MJ, Ratcliff G, editors. The neuropsychologyof high-level vision. Hillsdale (NJ): Erlbaum, 1994. p. 103–32.
[36] Perrett DI, Hietanen JK, Oram MW, Benson PJ. Organisationand functions of cells responsive to faces in the temporal cortex.Philosophical Transactions of the Royal Society of London [SeriesB: Biological Sciences] 1992;335:23–30.

712 S. Le et al. / Neuropsychologia 41 (2003) 702–712
[37] Renault B, Signoret JL, Debruille B, Breton F, Bolgert F.Brain potentials reveal covert facial recognition in prosopagnosia.Neuropsychologia 1989;27:905–12.
[38] Rizzo M, Hurtig R, Damasio AR. The role of scanpaths infacial recognition and learning. Annals of Neurology 1987;22:41–5.
[39] Sergent J, Poncet M. From covert to overt recognition of faces in aprosopagnosic patient. Brain 1990;113:989–1004.
[40] Sergent J, Ohta S, MacDonald B. Functional neuroanatomy of faceand object processing: a positron emission tomography study. Brain1992;115:15–36.
[41] Sergent J, Signoret JL. Varieties of functional deficits in proso-pagnosia. Cereb Cortex 1992;2:375–88.
[42] Small M. Visual evoked potentials in a patient with prosopagnosia.Electroencephalography and Clinical Neurophysiology 1988;71:10–6.
[43] Talairach J, Tournoux P. Co-planar stereotaxic atlas of the humanbrain. Stuttgart: Thieme, 1988.
[44] Tanaka JW, Farah MJ. Parts and wholes in face recognition. TheQuarterly Journal of Experimental Psychology 1993;46A:225–45.
[45] Tovee MJ, Cohen-Tovee EM. The neural substrate of face processingmodels: a review. Cognitive Neuropsychology 1993;10:505–28.
[46] Tranel D, Damasio AR. Knowledge without awareness: anautonomic index of facial recognition by prosopagnosics. Science1985;228:1453–4.
[47] Turk M, Pentland A. Eingenfaces for recognition: face perception.Journal of Cognitive Neuroscience 1991;3:71–86 [special issue].
[48] Walker-Smith GJ, Gale AG, Findlay JM. Eye-movement strategiesinvolved in face perception. Perception 1977;6:313–26.
[49] Warrington EK, James M. An experimental study of facial recognitionin patients with unilateral cerebral lesions. Cortex 1967;3:317–26.
[50] Whiteley AM, Warrington EK. Prosopagnosia: a clinical,psychological, and anatomical study of three patients. Journal ofNeurology, Neurosurgery, and Psychiatry 1977;40:395–403.
[51] Yin RK. Looking at upside-down faces. Journal of ExperimentalPsychology 1969;81:141–5.