Embed Size (px)
Citation preview

Section IIIE Other Crops
Improving Crop Resistance to Abiotic Stress, First Edition.Edited by Narendra Tuteja, Sarvajeet Singh Gill, Antonio F. Tiburcio, and Renu Tuteja� 2012 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2012 by Wiley-VCH Verlag GmbH & Co. KGaA.
j 1369

53Cotton: Genetic Improvement for Drought StressTolerance – Current Status and Research NeedsE. AbouKheir, M.S. Sheshshayee, T.G. Prasad, and Makarala Udayakumar
A significant part of cotton production comes from water-limited rain-fed areas.Being a commercially important crop, improvement of drought tolerance hasreceived increasing attention in crop improvement programs. The marked geneticvariability in desirable traits among cotton germplasm has great potential forexploitation through conventional and/or molecular breeding approaches. Whilemost research activities targeted the cotton speciesGossypium hirsutum, very limitedphysiological information is available in other cotton species. At present, theemphasis is to identify and assess variability in specific traits that have relevancein enhancing drought stress tolerance. Ensuing global research has enumerated afew traits such as roots, water use efficiency, and cellular-level tolerance as the mostimportant besides several other physiological traits. Being quantitatively inherited,numerous research attempts have been initiated to identify the relevant QTL and topyramid them on to superior genetic backgrounds. Likewise, phenomenal progresshas also beenmade in understanding the molecular regulation of drought tolerance,and to identify the functional/regulatory genes for development of drought-toleranttransgenics in various crops. Such approaches are minimal in cotton and it isimperative that such approaches have great promise to develop genotypes withagronomically desirable traits through molecular breeding and/or transgenics toimprove cotton yield and fiber quality. Hence, research priority should be laid onutilization of the available genomic resources through effective integration ofconventional breeding with molecular breeding tools. Furthermore, cotton species,namely,G. arboreum andG. herbaceum need to be explored to assess the variability instress-adaptive traits. This chapter examines the cotton drought response andprovides an insight into the relevant traits that have a role in enhancing droughttolerance. In addition, the prospects of conventional crop improvement approachesvis-a-vis molecular breeding and transgenic approaches that have relevance toachieving crop drought adaptation without sacrificing the yield and quality. It alsoprovides insight into the research needs on the quantitative and association geneticswith an ecophysiological understanding of the cotton genomic resources and tobetter inform crop improvement program.
j 1371
Improving Crop Resistance to Abiotic Stress, First Edition.Edited by Narendra Tuteja, Sarvajeet Singh Gill, Antonio F. Tiburcio, and Renu Tuteja� 2012 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2012 by Wiley-VCH Verlag GmbH & Co. KGaA.

53.1Introduction
Cotton belongs toMalvaceae family and tribeGossypieae that includes eight genera [1,2] with four of these genera (Lebronnecia, Cephalohibiscus, Gossypioides, and Kokia)having restricted geographic distributions [1, 2]. The genera Hampea, Cienfuegosia,Thespesia, and Gossypium are moderately larger genera with broader geographicrange. Fiber cotton belongs to Gossypium, which is the largest and most widelydistributed genus inGossypieae tribe with approximately 50 species [3] grown in aridto semiarid regions of the tropic and subtropic regions [3–5]. Of them, only twospecies, Gossypium arboreum and G. herbaceum, are commercially cultivated andknown collectively as �Desi� cottons, and are the Asiatic or Old World short-staplecotton. Owing to their ability to withstand drought stress, these diploid species aregenerally cultivated inmarginal, drought-prone environments of Asia. These diploidDesi cottons are also known for their ability to resist sucking pests such as hoppers,white flies, thrips, and aphids, and leaf curl virus. However, owing to their short,coarse, and weak fiber and undesirable boll and plant features, the Desi cottons weregradually displaced by G. hirsutum in Asia.
Evolutionarily, G. hirsutum has seven landraces, namely, palmeri, morilli, rich-mondii, yucatenanse, punctatum,marie galante, and latifolium. The race latifolium iswidely cultivated in the world and yucatannse appears to be the wild progenitor ofupland cotton [6]. G. barbadense is the other domesticated allotetraploid that yieldsextralong staples or extrafine quality cotton fiber. This species shares about 3% ofthe total world cotton market [7]. The exploitation of the genetic variability of thesespecies is yet to be realized using modern biological tools in conjunction withconventional approaches.
Most cotton fiber production comes from species G. hirsutum and G. barbadense,with a minor contribution from G. arboreum and G. herbaceum. Although cotton iscultivated in more than 70 countries [8] ranging from tropical to subtropical regionswithmore than 350millionha [9], almost 65%of global area is in four countries India,China, the United States of America, and Pakistan (Table 53.1). India ranks first interms of cultivation area, occupying 26% of the world cotton area, followed by China,the United States, and Pakistan. Globally, 115 million bales were produced during2006–2007 season. China contributes 24% of total production followed by India, theUnited States, and Pakistan. Although Australia ranks first in productivity (1887 kgha�1), it contributes to only 2% of total cotton production with less than 1% of globalcotton area (Table 53.1). However, among themajor cotton growing countries, Chinaholds the highest productivity (1130 kg ha�1) followed by the United States andPakistan with productivity of 858 and 703 kg ha�1, respectively. India with a yield of494 kg ha�1 is the lowest.
Major area of cotton cultivation in India is under rain-fed conditions, which occupyabout 50%. The cultivated area is concentrated in the central part of India occupyingabout 66.66%, with only 23% under irrigated conditions followed by SouthernIndia (40%) [10]. Several abiotic stresses such as temperature, soil ion concentrations,
1372j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs
light, and so on adversely affect cotton growth and development. However, themost devastating abiotic stress is soil moisture deficit and about 45% of the world�sarable land suffers from chronically inadequate supplies of water for agricul-ture [12] including cotton [13]. It is projected that burgeoning human populationand urbanization will further aggravate the increasing demand for fresh water bothfor human consumption and for agriculture. Furthermore, erratic and ofteninsufficient rainfall witnessed in the recent past due to anthropogenic changesin the climate might further reduce the availability of fresh water. Although waterand soil management practices have a significant role in sustaining productivityunder water-limited conditions, a genetic enhancement of cotton genotypes withincreased water use efficiency (WUE) as a seed-based technology has greateracceptability.
Despite increased interest to exploit the genomic resources for the development ofdrought-tolerant crops, it has been hindered by lack of knowledge of precisephysiological characteristics that form the index diagnostic parameter of geneticvariability and potential for improved productivity and quality under water-limitedconditions. Using genetic mapping to dissect the inheritance of different complextraits is a powerful tool to distinguish heritability from casual associations betweensuch traits [14]. In principle, it permits a direct analysis of the role of specificphysiological traits in determining genetic variability and potential for productivityand quality parameters under abiotic stress conditions.
Cotton (Gossypium spp.) is a potential taxon to study the variability in genomicresources with potential for adaptation to water-limited conditions. Cotton originatesfrom wild perennial plants adapted to semiarid, subtropical environments thatexperience periodic drought stress conditions. However, themodern cotton cultivarsare the result of intensive selection to improve the yield of lint and its quality. Such atraditional selection with emphasis on maximal productivity under high-input andirrigated conditions has inadvertently eroded the genetic variability in droughttolerance [15]. However, considerable variation persists within and between the twocultivated tetraploid cotton species, G. hirsutum and G. barbadense, in physiologicaltraits, which can be still exploitable.
Table 53.1 Major cotton-growing countries of the world.
Country Area (�000 ha) Production (�000 bales) Yield (kg ha�1)
USA 5186 20 431 858China 5300 27 500 1130India 9250 21 000 494Pakistan 3250 10 500 703Australia 300 2600 1887World 35175 115 590 715
Source: FAO [11].
53.1 Introduction j 1373
53.2Response of Cotton to Drought Stress
Water deficit stress has profound effects on plant growth, development, and quality.The first effect of themoisture stressmay well be loss of turgor that affects the rate ofcell expansion and ultimate cell size. The result is a decrease in growth rate, stemelongation, leaf expansion, and stomatal aperture with an overall decrease in theperformance of the plant.
Such reduction in leaf area index (LAI) results in a reduced crop growth rate (CGR)under conditions of lower water supply [16–18]. Similar inferences where moisturestress during vegetative stage causes reduced leaf growth and leaf area have also beenreported [19]. Smaller leaves, increased leaf senescence and abscission [20–22], lesserleaf numbers [23], reduced leaf area index atmaturity and interception of less light bythe crop, lesser plant height [19], and lower shoot-to-root ratio [19, 24] eventuallydecrease total dry matter produced by plant.
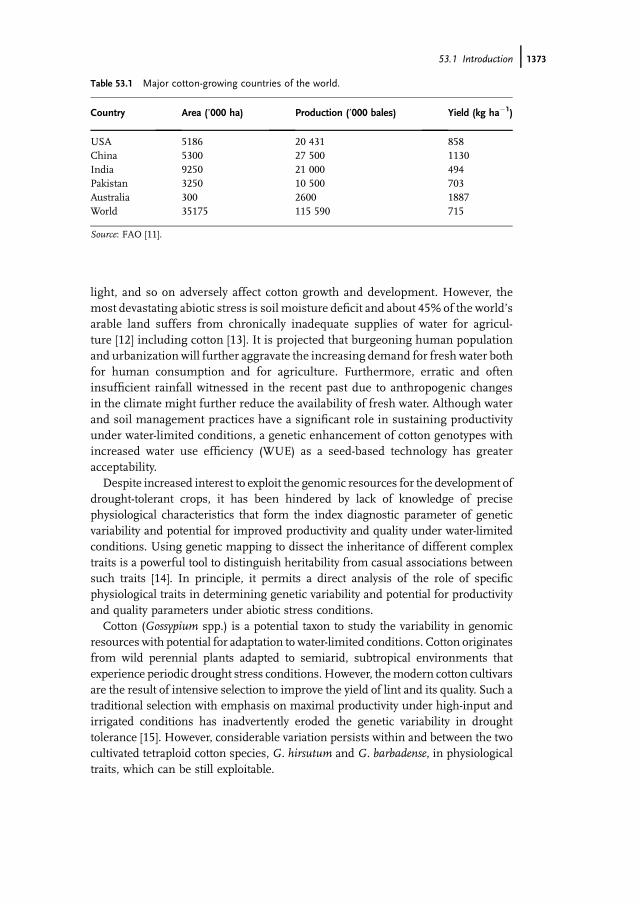
In one of our experiments [25], three upland cotton genotypes grown in twodifferent water regimes (100 and 55% field capacity) showed significant reduction inleaf area (Figure 53.1a) and total dry matter (Figure 53.1b). This reduction in growthrates often corresponds linearly with water used by the crop [26]. A linear decrease instem elongation has been reported as leaf water potential (LWP) declined and nofurther growth was observed when LWP was as low as �2.5MPa [27–29].
These results clearly indicate thatmaintenance of leaf tissuewater relations ismostcrucial to sustain growth rates under water-limited conditions. Therefore, greateremphasis needs to be laid on assessing the variability in root traits and the root growthresponse to drought stress. Significant changes in root growth patterns have beennoticed during a drying cycle [30]. Stimulation of root proliferation is significantlygreater as soil drying continues, a trend not observed in well-watered plants.Furthermore, during drying cycles, root growth appears closely associated withshoot growth, with elongation ceasing concurrently with cessation of growth in theshoot [31] and root elongation ceased when the soil water potential declined to less
Figure 53.1 Effect of moisture stress on (a) leaf area and (b) total dry matter among three uplandcotton genotypes grown into two different water regimes [25].
1374j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs

than �0.1MPa. The taproot length was greater but not the root dry weight undermoisture stress compared to control condition [19]. The leaf expansion was moresensitive to water stress than root elongation [32, 33].
Water deficit stress also affects boll production and thus cotton yield [34–36]. Theabrupt cessation inboll formationdependson imbalancebetweenplantsphotosyntheticcapacity and assimilate demand [37, 38]. Moisture stress occurrence during the first 14days after anthesis leads to boll abscission [39], but occurrence of stress after that periodgenerally does not cause abscission [40]. However, stress later during boll formationadversely affects fiber quality. Fiber elongation was decreased when drought stressoccurred during the fiber elongation stage [41–43] and if stress severity increasedbeyond the fiber elongation period, led to fiber immaturity and lowmicronaire [44]. Onthe contrary,milddrought stress canbebeneficial due to increase in lint percentage overirrigated cotton [45] and occasionally increasedmicronaire [46]. Krieg [23] summarizedthe drought stress effects on cotton growth and productivity and demonstrated that thetimingandseverity ofwaterdeficit stresshasa largeeffect onyield. Levi etal. [47] showed31% reduction in seed cotton yield under moisture stress.
Soil water deficit causes a progressive and significant reduction in evapotranspi-ration (ET) with the extent of reduction dependant on the severity and duration ofstress [27]. This reduced transpiration would lead to an increase in the canopytemperature predominantly due to a stress-induced stomatal closure [22]. Thereduction in transpiration can also be attributed to a stress-induced reduction inleaf expansion [48–50], stomatal closure [51–53], and reduction in leaf metabolicactivities leading to increased senescence. In cotton, photosynthesis ismore sensitiveto low water potential than the stomatal conductance [54, 55] mainly due to decreasein the synthesis and activity of photosynthetic enzymes under moisture stressconditions [56].
53.3Trait-Based Breeding to Improve Tolerance to Abiotic Stress
Considering the economic importance of cotton and that a significant area of cottonproduction comes fromwater-limited rain-fed conditions, attempts to sustain or evenincrease yield potentials under stress is a major challenge. Through crop improve-ment efforts for higher absolute yields, further success in such conventionalbreedingmaynot provide the requisite yield advantage because of a narrow variabilityin yield among the improved cultivars [57, 58]. Therefore, understanding the plantphysiological characteristics underlying yield formation can direct breeding both forspecific integral traits and for crop response to stress leading to increased yield andcotton fiber quality.
Drought stress is a very complex stress, and both stress response and cropadaptation to stress are equally complicated. For achieving a comprehensive improve-ment in drought tolerance coupled with improved productivity under water-limitedconditions, it appears that several diverse traits need to be introgressed into a singleelite genetic background. Among a large number of adaptive and constitutive traits
53.3 Trait-Based Breeding to Improve Tolerance to Abiotic Stress j 1375

that have been identified, traits associated with tissue–water relations need to beimproved [59–61]. Traits associatedwithminingwater fromdeeper soil layers (roots),water use efficiency [62, 63], and high-degree of cellular level tolerance [64] deservegreatest emphasis. Toward a trait-based breeding strategy, the first step is tosystematically characterize the available cotton germplasm accessions for the vari-ability in these traits and identify superior trait donor lines. In addition, it is alsoextremely important to assess variability in several morphophysiological traits suchas the canopy architecture and leaf expansion traits.
53.3.1Canopy Characters
Manipulating the canopy architecture through leaf characters can be a useful strategyin breeding for drought tolerance.Other traits such as earlymaturity [65, 66], reducedleaf area index, and higher canopy CO2 uptake [67], high light-saturated single-leafphotosynthetic rates [68], a shorter sympodial plastochron [69], and increasednumbers of flowers per season [70] also deserve exploitation. Meredith [71] foundthat a copy with subokra-type leaves yielded greater than okra, superokra, and normalleaf types. Stiller et al. [72] confirmed that cotton with the okra leaf morphology wasmore suitable for improved yield under dry land conditions.
53.3.2Root Traits
Cotton root consists of a primary or �tap� root from which branch the secondary andtertiary roots [73]. The cotton root, whether primary or branch root, has a single layerof epidermal cells surrounding the root cortex. The endodermis, single-cell layer,surrounds the stele that contains the xylem and phloem vascular elements and a celllayer called the pericycle. The xylem elements are arranged either in a tetrarch (fourdistinct xylem bundles, the most common) or in a greater bundle arrangement [74].The difference in arrangement is apparently genetically controlled andmay result insignificant genetic diversity in root characteristics.
The depth of root penetration depends on a number of factors such as soilmoisture, structure, pH, temperature, and so on, but in general the taproot canreach depths of over 3m and can elongate at a rate from less than 1 to over 6 cm perday. The elongation rate of the lateral or secondary roots would generally fall withinthe same range [75]. In general, the root system continues to grow and increase inlength until young bolls begin to form [31] at which time root length declines as olderroots die. New roots continue to be formed past this point, but the net result is adecline in total length [76].
A number of different root traits and morphophysiological traits have beenimplicated as important mechanisms that impart drought tolerance in cotton. Theseinclude distance from transition zone to the first main lateral root, taproot weight,number of lateral roots, seedling vigor, rapidity of root systemdevelopment, and root-to-shoot ratio [24] and longer taproot length [19].
1376j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs

Several of the root-related traits show significant response to stress. An increase inroot density in the soil layer between 70 and 180 cm was reported in cotton in dry soilprofile compared towell-watered condition [30]. Pace et al. [19] found that under droughtstress, there was increase in taproot length without any change in cotton taproot dryweight. Detrimental effects of drought on root growthwere observed only after recoverywhere both taproot� and secondary roots� dry weight decreased [19, 33, 77]. Thus,increase in taproot length, at the expense of thickening, in response to droughtmay be acommon response in cotton. Such a responsemay have evolved to permit cotton plantsto survive drought by accessing water from deeper soil profiles [78]. Contrastingevidence exists that drought stress reduces cotton root growth, root elongation, rootlength density, and root distribution [32, 79, 80]. However, such root growth reductionsare normally noticed when the stress is severe or when it is prolonged.
Although superior drought tolerance is associated with enhanced root growth,rapid root water uptake, maintenance of root viability at the soil surface, and rapidroot regeneration after rewatering [81], no serious breeding efforts have beeninitiated till date to improve root traits [82]. Lack of appropriate phenotyping strategyfor root traits is perhaps themost important constraint. Several techniques have beendeveloped to study root parameters such as hydroponics [83, 84],minirhizotrons [85],minilysimeters [86], and tubes [87]. More recently, sophisticated imaging techniqueshave been developed for assessing root growth and development. Although thesetechniques have been used widely to measure root parameters, they suffer eitherfrom cumbersomeness of procedure or inability to screen large number of acces-sions. In addition, some of the techniques such as tubes, minilysimeters, and so ondo not represent the correct phenotypic expression of roots, as they do not experienceinterplant competitions [88]. Similarly, hydroponic techniques also cannot beadopted to assess root growth variations over extended crop growth periods.
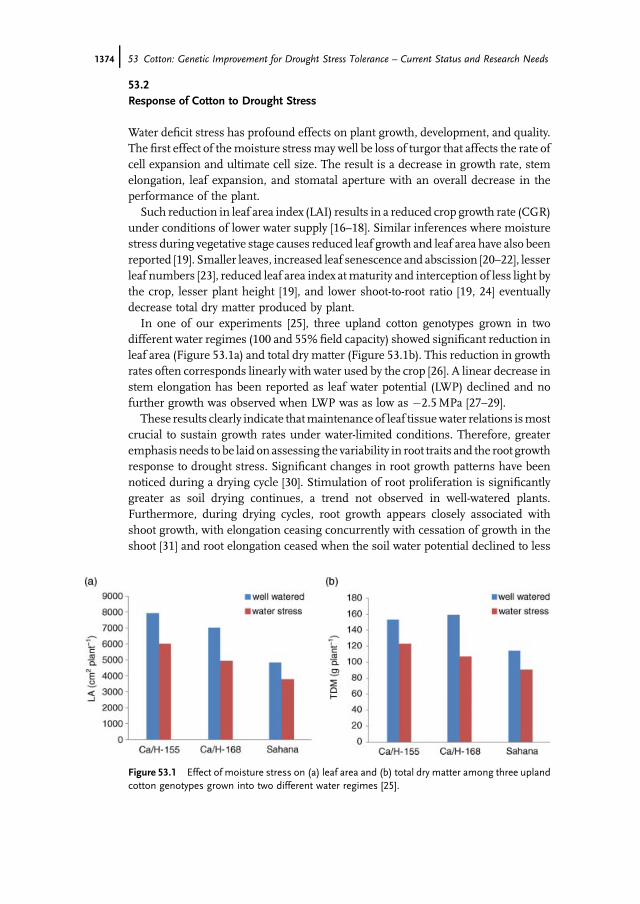
Most of these disadvantages can be overcome by growing plants in speciallyconstructed �field root phenotyping structures� [88]. Briefly, these root structures areconstructed using bricks and are raised beds 5 feet above ground and are 10 feet wide.The length of the structures can be varied based on the crop species and the numberof accessions to be studied (Figure 53.2). Soil of the required type can befilled in thesestructures and plants are raised with the recommended spacing between plants.Thus, in such field root phenotyping structures plants are raised under near-naturalconditions that experience interplant competition and hence would represent thebest phenotypic assessment while determining the genetic variability [64, 88].
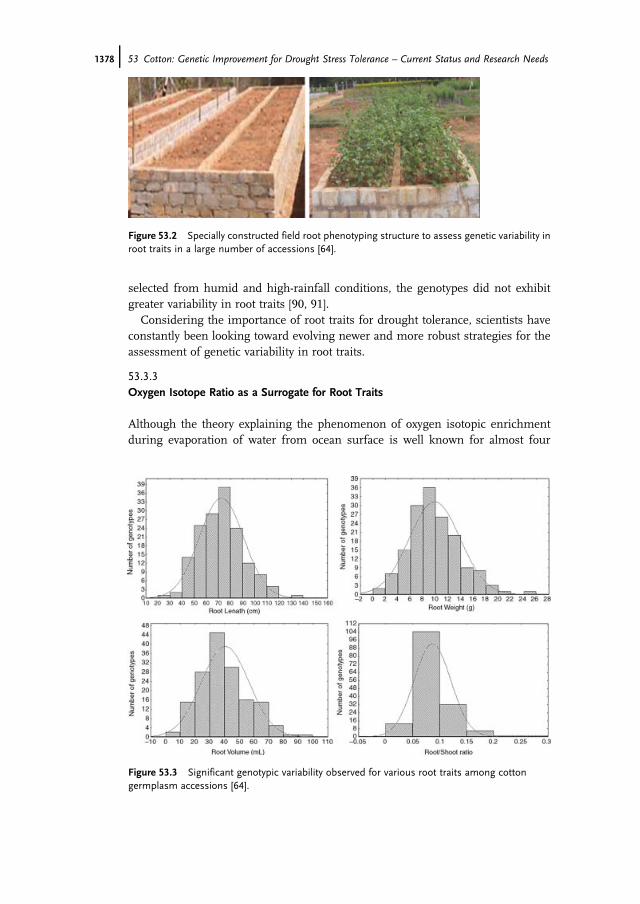
In a field study done at our center, 158 upland cotton germplasm accessions werescreened for variability in root traits by raising plants in field root phenotypingstructures (Figure 53.2). A significant genotypic variability in root length, root dryweight, root volume, and root-to-shoot ratio was observed [64]. Each of theseparameters displayed a normal distribution illustrating the polygenic regulation ofroot traits (Figure 53.3).
Similarly, Basal et al. [89] reported a considerable genetic variation in root length,lateral root number, root freshweight, lateral root dryweight, and total root dryweightof 68 cotton landraces. On the contrary, Roark and Quisenberry [90] observed lowgenetic variability in cotton cultivars. Asmost of the cultivars examined by themwere
53.3 Trait-Based Breeding to Improve Tolerance to Abiotic Stress j 1377
selected from humid and high-rainfall conditions, the genotypes did not exhibitgreater variability in root traits [90, 91].
Considering the importance of root traits for drought tolerance, scientists haveconstantly been looking toward evolving newer and more robust strategies for theassessment of genetic variability in root traits.
53.3.3Oxygen Isotope Ratio as a Surrogate for Root Traits
Although the theory explaining the phenomenon of oxygen isotopic enrichmentduring evaporation of water from ocean surface is well known for almost four
Figure 53.2 Specially constructed field root phenotyping structure to assess genetic variability inroot traits in a large number of accessions [64].
Figure 53.3 Significant genotypic variability observed for various root traits among cottongermplasm accessions [64].
1378j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs
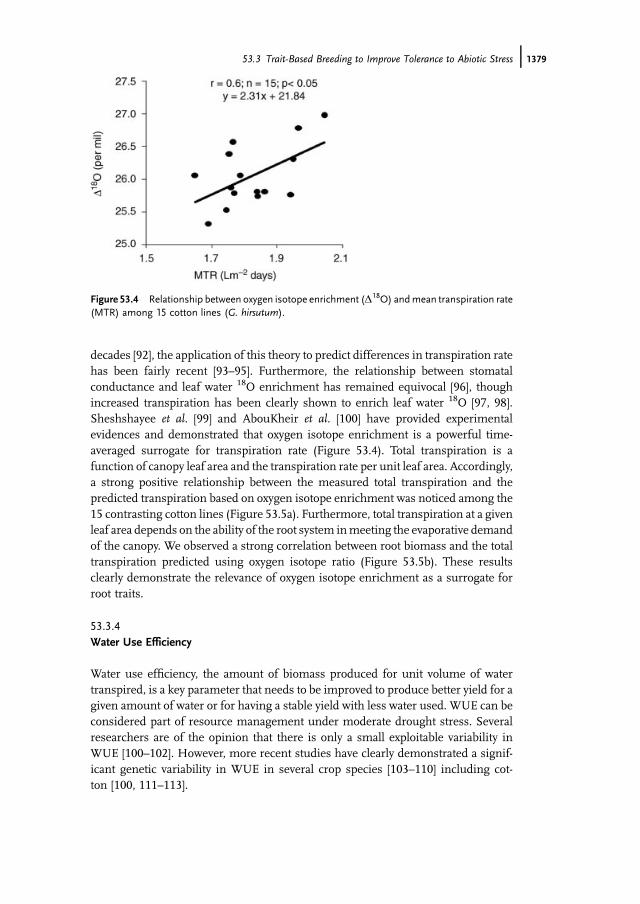
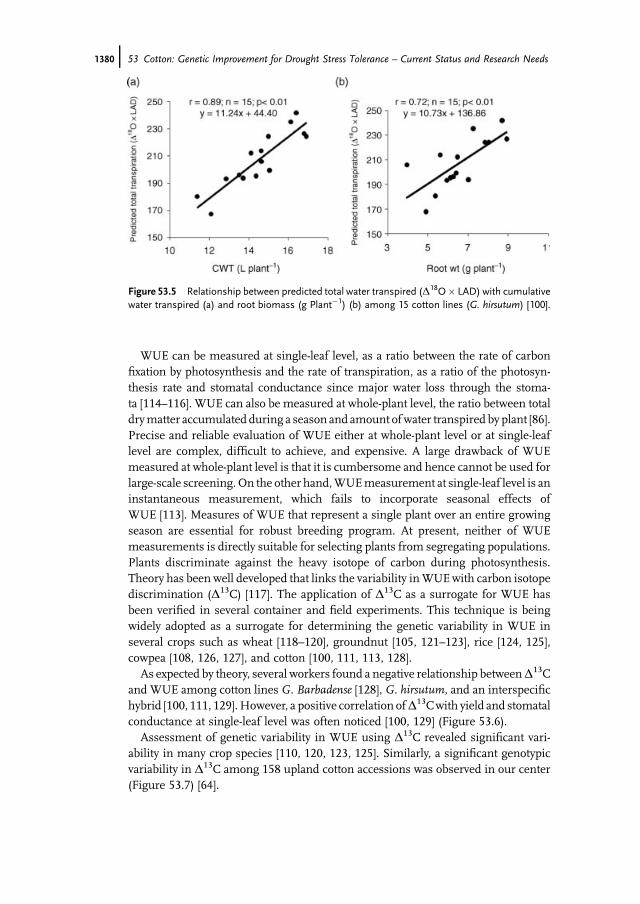
decades [92], the application of this theory to predict differences in transpiration ratehas been fairly recent [93–95]. Furthermore, the relationship between stomatalconductance and leaf water 18O enrichment has remained equivocal [96], thoughincreased transpiration has been clearly shown to enrich leaf water 18O [97, 98].Sheshshayee et al. [99] and AbouKheir et al. [100] have provided experimentalevidences and demonstrated that oxygen isotope enrichment is a powerful time-averaged surrogate for transpiration rate (Figure 53.4). Total transpiration is afunction of canopy leaf area and the transpiration rate per unit leaf area. Accordingly,a strong positive relationship between the measured total transpiration and thepredicted transpiration based on oxygen isotope enrichment was noticed among the15 contrasting cotton lines (Figure 53.5a). Furthermore, total transpiration at a givenleaf area depends on the ability of the root system inmeeting the evaporative demandof the canopy. We observed a strong correlation between root biomass and the totaltranspiration predicted using oxygen isotope ratio (Figure 53.5b). These resultsclearly demonstrate the relevance of oxygen isotope enrichment as a surrogate forroot traits.
53.3.4Water Use Efficiency
Water use efficiency, the amount of biomass produced for unit volume of watertranspired, is a key parameter that needs to be improved to produce better yield for agiven amount of water or for having a stable yield with less water used. WUE can beconsidered part of resource management under moderate drought stress. Severalresearchers are of the opinion that there is only a small exploitable variability inWUE [100–102]. However, more recent studies have clearly demonstrated a signif-icant genetic variability in WUE in several crop species [103–110] including cot-ton [100, 111–113].
Figure 53.4 Relationship between oxygen isotope enrichment (D18O) andmean transpiration rate(MTR) among 15 cotton lines (G. hirsutum).
53.3 Trait-Based Breeding to Improve Tolerance to Abiotic Stress j 1379
WUE can be measured at single-leaf level, as a ratio between the rate of carbonfixation by photosynthesis and the rate of transpiration, as a ratio of the photosyn-thesis rate and stomatal conductance since major water loss through the stoma-ta [114–116]. WUE can also be measured at whole-plant level, the ratio between totaldrymatter accumulated during a season and amount ofwater transpired by plant [86].Precise and reliable evaluation of WUE either at whole-plant level or at single-leaflevel are complex, difficult to achieve, and expensive. A large drawback of WUEmeasured at whole-plant level is that it is cumbersome and hence cannot be used forlarge-scale screening. On the other hand,WUEmeasurement at single-leaf level is aninstantaneous measurement, which fails to incorporate seasonal effects ofWUE [113]. Measures of WUE that represent a single plant over an entire growingseason are essential for robust breeding program. At present, neither of WUEmeasurements is directly suitable for selecting plants from segregating populations.Plants discriminate against the heavy isotope of carbon during photosynthesis.Theory has been well developed that links the variability inWUEwith carbon isotopediscrimination (D13C) [117]. The application of D13C as a surrogate for WUE hasbeen verified in several container and field experiments. This technique is beingwidely adopted as a surrogate for determining the genetic variability in WUE inseveral crops such as wheat [118–120], groundnut [105, 121–123], rice [124, 125],cowpea [108, 126, 127], and cotton [100, 111, 113, 128].
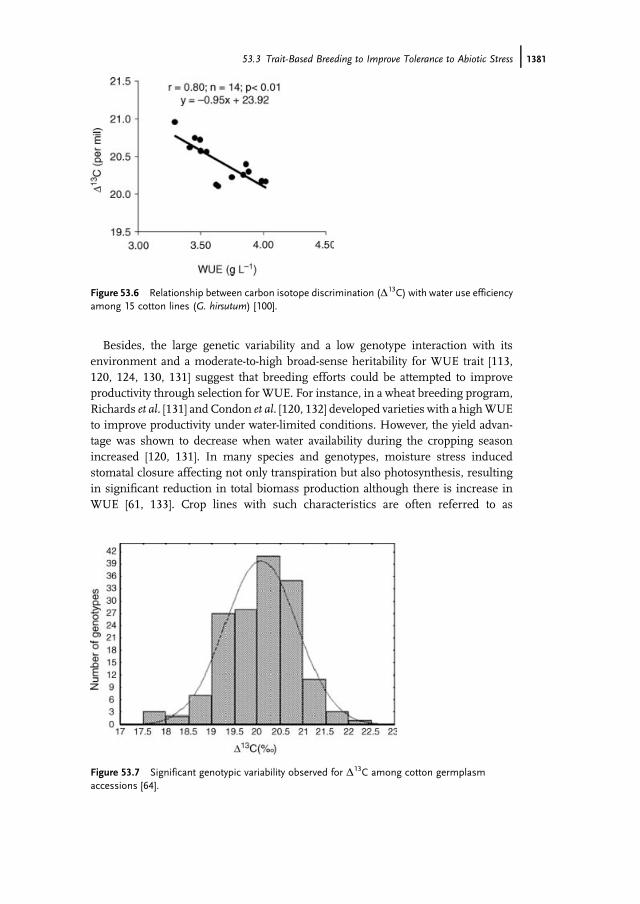
As expected by theory, several workers found a negative relationship betweenD13Cand WUE among cotton lines G. Barbadense [128], G. hirsutum, and an interspecifichybrid [100, 111, 129].However, a positive correlation ofD13Cwith yield and stomatalconductance at single-leaf level was often noticed [100, 129] (Figure 53.6).
Assessment of genetic variability in WUE using D13C revealed significant vari-ability in many crop species [110, 120, 123, 125]. Similarly, a significant genotypicvariability in D13C among 158 upland cotton accessions was observed in our center(Figure 53.7) [64].
Figure 53.5 Relationship between predicted total water transpired (D18O� LAD) with cumulativewater transpired (a) and root biomass (g Plant�1) (b) among 15 cotton lines (G. hirsutum) [100].
1380j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs
Besides, the large genetic variability and a low genotype interaction with itsenvironment and a moderate-to-high broad-sense heritability for WUE trait [113,120, 124, 130, 131] suggest that breeding efforts could be attempted to improveproductivity through selection for WUE. For instance, in a wheat breeding program,Richards et al. [131] and Condon et al. [120, 132] developed varieties with a highWUEto improve productivity under water-limited conditions. However, the yield advan-tage was shown to decrease when water availability during the cropping seasonincreased [120, 131]. In many species and genotypes, moisture stress inducedstomatal closure affecting not only transpiration but also photosynthesis, resultingin significant reduction in total biomass production although there is increase inWUE [61, 133]. Crop lines with such characteristics are often referred to as
Figure 53.6 Relationship between carbon isotope discrimination (D13C) with water use efficiencyamong 15 cotton lines (G. hirsutum) [100].
Figure 53.7 Significant genotypic variability observed for D13C among cotton germplasmaccessions [64].
53.3 Trait-Based Breeding to Improve Tolerance to Abiotic Stress j 1381
�conductance types� [95, 108, 110]. On the other hand, increase in WUE arisespredominantly by maintaining high photosynthetic capacity in a few species andgenotypes; selection in such types for higher WUE will not affect the plant biomassand yield that are called �capacity types� [106, 110, 134]. The classification of somecrops into capacity and conductance types is provided in Table 53.2.
Cotton is classified as conductance types (Table 53.2), where WUE increases byreducing transpiration through stomatal closure, thus associated with reduction inthe biomass produced [132, 147]. Recently, AbouKheir et al. [100] demonstrated stableisotopes (13C and 18O) as high-throughput technique for accurate identification ofcapacity type among cotton genotypes. The carbon and oxygen isotope ratios of plantorganic matter integrate the diurnal and seasonal variations in Ci and gs, respec-tively [110, 135]. Therefore, demonstration of carbon- and oxygen-stable isotope ratio(D13C/D18O) is a good time-averaged estimate of in vivo photosynthetic capacity andthus to identify capacity type genotypes.
53.4Water Conservation Mechanisms
53.4.1Epicuticular Wax
Avariety of functions have been attributed to thesewaxes related to their involvementin physical and physiological processes, including the reduction of transpiration andgas exchange. Commonly, drought-stressed cotton leaves exhibit an increase in thewaxy cuticular layer of the leaf [148, 149] for water conservation. Bondada et al. [150]showed that water stress increased total wax concentration in cotton and induced theproduction of long-chain alkanes whose number and levels were higher in the totalwax of the leaf, bract, and boll compared to the corresponding well-watered cotton
Table 53.2 Classification of some crop species as capacity and conductance types.
Capacity (gm) types Conductance (gs) type
Crop Reference Crop Reference
Groundnut [104] Wheat [141]Wheat [119] Wheat [142]Groundnut [135] Barley [143]Groundnut [103] Wheat [144]Groundnut [121] Wheat [145]Groundnut [136] Coffee [146]Beans [137] Cotton [128]Grasses [138] Cotton [64]Canola [139] Rice [124]Spruce [140] Cowpea [108]
1382j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs

plants. It has been reported that cuticular thickness and waxiness of leaf surface aregenetically controlled by a large number of genes affecting the transpiration [151].
53.4.2Cuticular/Nonstomatal Water Loss
Most of the nonstomatal water loss from the leaf would be from the epidermis due todifferences in cuticular thickness. It is predominantly influenced by cuticular waxes,which is an intrinsic drought-tolerant character affecting water retention in leaves.The trait has great relevance in water conservation under drought conditions. Thegenetic variation in cuticular water loss has been reported in crop species and used asselection criterion for drought resistance [152–155]. Genotypic variation in cuticularwater loss was assessed among the 158 cotton germplasm accessions in our center bymeasuring the rate of water loss from excised leaves. Results revealed a considerablegenotypic variation among cotton accessions tested in moisture retention capacity at5 h that ranged from 40.04 to 83.27% (Figure 53.9) [25].
53.5Cellular-Level Tolerance
53.5.1Intrinsic Tolerance
Under natural conditions, development of abiotic stress levels is a progressivephenomenon. Accordingly, plants initially experience a milder stress level beforebeing exposed to severe stress. In response to milder stress, several genes have beenshown to be either upregulated or downregulated, which leads to the development ofa considerable degree of tolerance in plants when stress becomes severe. Such anacclimatization response of plants is termed �acquired tolerance� and it has beendemonstrated in several species [156–160] including cotton [161].
The induction of stress triggers several signaling pathways, resulting in theexpression of an array of stress-responsive genes and the gene products may alterthe physiological and biochemical processes relevant to stress tolerance. Whileseveral genes are suppressed, a large number of genes are upregulated in responseto stress [162–165]. Drought stress-responsive genes have their own cognate recep-tors, which operate independently or cooperatively to initiate downstream signalingevents. Regulation of both upstream and downstream components such as heatshock protein, protein kinases, secondary messengers (reactive oxygen species(ROS), calcium, etc.), and transcription factors is directly or indirectly involved inestablishing the required responses that are well studied [166–168].
On the basis of the concept that seedlings acquire a certain degree of stresstolerance when exposed to milder stress levels, the novel and high-throughput�temperature induction response� (TIR) technique has been developed and validatedin many crop species [162, 164, 165] including cotton [161]. The technique involves
53.5 Cellular-Level Tolerance j 1383
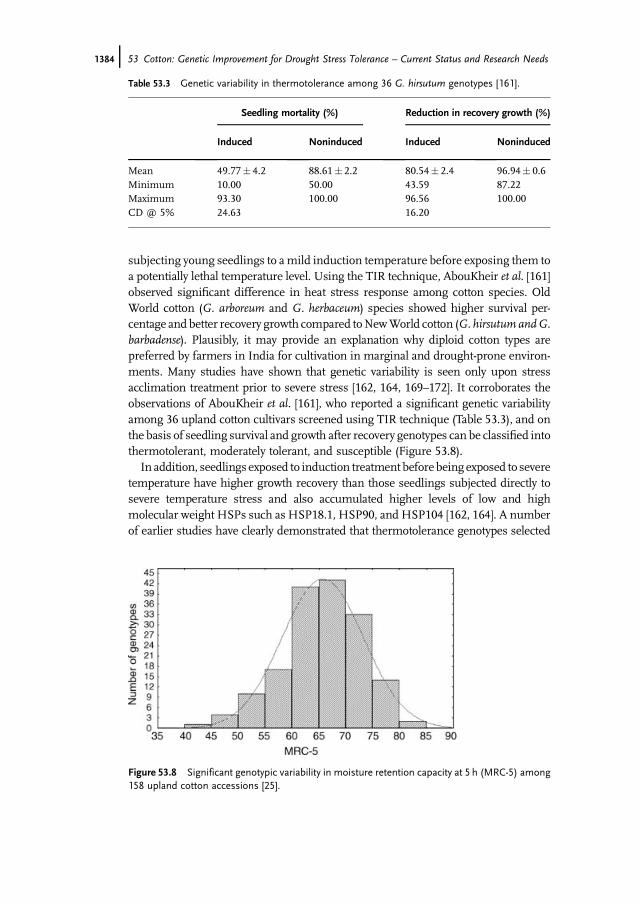
subjecting young seedlings to amild induction temperature before exposing them toa potentially lethal temperature level. Using the TIR technique, AbouKheir et al. [161]observed significant difference in heat stress response among cotton species. OldWorld cotton (G. arboreum and G. herbaceum) species showed higher survival per-centage and better recovery growth compared toNewWorld cotton (G. hirsutum andG.barbadense). Plausibly, it may provide an explanation why diploid cotton types arepreferred by farmers in India for cultivation in marginal and drought-prone environ-ments. Many studies have shown that genetic variability is seen only upon stressacclimation treatment prior to severe stress [162, 164, 169–172]. It corroborates theobservations of AbouKheir et al. [161], who reported a significant genetic variabilityamong 36 upland cotton cultivars screened using TIR technique (Table 53.3), and onthe basis of seedling survival and growth after recovery genotypes can be classified intothermotolerant, moderately tolerant, and susceptible (Figure 53.8).
In addition, seedlings exposed to induction treatment beforebeing exposed to severetemperature have higher growth recovery than those seedlings subjected directly tosevere temperature stress and also accumulated higher levels of low and highmolecular weight HSPs such as HSP18.1, HSP90, and HSP104 [162, 164]. A numberof earlier studies have clearly demonstrated that thermotolerance genotypes selected
Table 53.3 Genetic variability in thermotolerance among 36 G. hirsutum genotypes [161].
Seedling mortality (%) Reduction in recovery growth (%)
Induced Noninduced Induced Noninduced
Mean 49.77� 4.2 88.61� 2.2 80.54� 2.4 96.94� 0.6Minimum 10.00 50.00 43.59 87.22Maximum 93.30 100.00 96.56 100.00CD @ 5% 24.63 16.20
Figure 53.8 Significant genotypic variability in moisture retention capacity at 5 h (MRC-5) among158 upland cotton accessions [25].
1384j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs
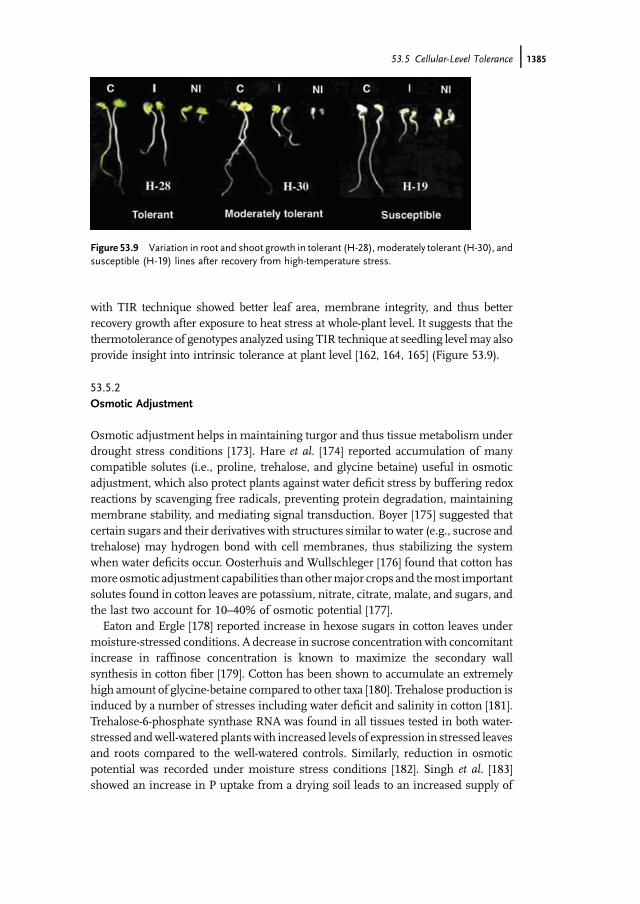
with TIR technique showed better leaf area, membrane integrity, and thus betterrecovery growth after exposure to heat stress at whole-plant level. It suggests that thethermotolerance of genotypes analyzed using TIR technique at seedling levelmay alsoprovide insight into intrinsic tolerance at plant level [162, 164, 165] (Figure 53.9).
53.5.2Osmotic Adjustment
Osmotic adjustment helps in maintaining turgor and thus tissue metabolism underdrought stress conditions [173]. Hare et al. [174] reported accumulation of manycompatible solutes (i.e., proline, trehalose, and glycine betaine) useful in osmoticadjustment, which also protect plants against water deficit stress by buffering redoxreactions by scavenging free radicals, preventing protein degradation, maintainingmembrane stability, and mediating signal transduction. Boyer [175] suggested thatcertain sugars and their derivatives with structures similar to water (e.g., sucrose andtrehalose) may hydrogen bond with cell membranes, thus stabilizing the systemwhen water deficits occur. Oosterhuis and Wullschleger [176] found that cotton hasmore osmotic adjustment capabilities than othermajor crops and themost importantsolutes found in cotton leaves are potassium, nitrate, citrate, malate, and sugars, andthe last two account for 10–40% of osmotic potential [177].
Eaton and Ergle [178] reported increase in hexose sugars in cotton leaves undermoisture-stressed conditions. A decrease in sucrose concentration with concomitantincrease in raffinose concentration is known to maximize the secondary wallsynthesis in cotton fiber [179]. Cotton has been shown to accumulate an extremelyhigh amount of glycine-betaine compared to other taxa [180]. Trehalose production isinduced by a number of stresses including water deficit and salinity in cotton [181].Trehalose-6-phosphate synthase RNA was found in all tissues tested in both water-stressed andwell-watered plantswith increased levels of expression in stressed leavesand roots compared to the well-watered controls. Similarly, reduction in osmoticpotential was recorded under moisture stress conditions [182]. Singh et al. [183]showed an increase in P uptake from a drying soil leads to an increased supply of
Figure 53.9 Variation in root and shoot growth in tolerant (H-28), moderately tolerant (H-30), andsusceptible (H-19) lines after recovery from high-temperature stress.
53.5 Cellular-Level Tolerance j 1385

osmotically active inorganic solutes for the cells in the growing cotton leaves. As aconsequence, plants accumulate both free and bound water, ultimately leading toincreased leaf expansion rate. Thus, root traits that enhance the P uptake could be abeneficial mechanism to alleviate drought stress.
Quisenberry et al. [184] reported significant differences among cotton cultivars forosmotic potential at zero turgor, further suggesting genotypic variation in osmo-regulation in cotton. However, a significant negative correlation between shoot dryweight and increased osmoregulation was reported from the same group, implyingthat if cotton genotypes are selected on the basis of the degree of osmotic adjustmentunder water deficit stress alone, a decrease in potential growth could be a possibilityin selected species. Evidently, documented reports suggest that relationship betweenosmotic adjustment and yield were inconsistent, which can be positive [82], nega-tive [184, 185], or no association [186, 187].
53.5.3Chlorophyll Fluorescence
Fv/Fm is often used as an early indicator of photosynthetic damage caused by abioticstress in many crops [188–191]. Burke [192] developed a novel bioassay for theidentification of drought stress in cotton that utilizes chlorophyll fluorescence tomonitor cell viability under high-temperature dark incubation. Differences betweenwell-watered and drought-stressed plants can be established since, under stress,plants will not mobilize carbohydrate reserves overnight and will therefore maintainhigher fluorescence values during high-temperature dark incubation, with theopposite being true for well-watered plants. Normal metabolic processes have beenshown to be disrupted by drought leading to a reduction in the translocation ofphotosynthate from leaves to other plant tissues [193]. The okra leaf genotypes didhave 14% greater F0
v=F0m across treatments compared to the normal leaf cottons.
Higher photosynthetic rates per unit leaf area have been observed in okra leafgenotypes [194]. Higher chlorophyll fluorescence values in drought-stressed plantswere observed in 20 genotype of diverse germplasm evaluated in a 2 year fieldevaluation under two water regimes [195], indicating the importance of using thisbioassay to evaluate and identify the drought tolerance cotton genotypes.
53.6Conceptual Approach to Cotton Improvement for Abiotic Stress
A variety of approaches have been used to alleviate the problem of drought stressthrough the enhancement of adaptive mechanisms of plants. It appears to be aformidable task as drought is a complex abiotic stress controlled by many physio-logical and biochemical characteristics, besides phenotypic characteristics of plants.Plant breeding, either conventional or molecular breeding, is being extensively usedto tailor and develop genotypeswith genetically superior traits to enable them to adaptto drought-stress environments. There has been commendable progress made
1386j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs

during the last century by plant breeders in developing drought-tolerant lines/varieties/cultivars of some crops through conventional breeding. However, theapproach is highly time consuming and labor and cost intensive. Alternatively,marker-assisted selection (MAS) [196] has been realized to be a more efficientapproach, which identifies the usefulness of thousands of genomic regions of acrop under stress conditions. Quantitative trait loci (QTL) for drought tolerance havebeen identified for a variety of traits in different crops. The development ofcomprehensive molecular linkage maps and marker-assisted selection has openedup options to pyramid desirable traits to achieve crop drought tolerance. To date, suchcrop improvement programs have been hampered by significant genetic� environ-ment interaction, large number of genes encoding yield and use an inappropriatemapping population involved in mapping of QTL for better growth and yield underwater deficit conditions. Advancement of scientific knowledge on genomics andproteomics has been increasingly beneficial for precise molecular breeding andtransformation approaches in crop improvement. With the available tools andtechniques, the realization of trait pyramidingposes a daunting challenge. Therefore,the emphasis must be placed to devising appropriate strategies and tools for precisetrait pyramiding to improve drought tolerance.
The first step in this direction would be to identify genes and QTL that control thespecific traits of relevance to drought tolerance and such desirable traits� variabilityamong the genotypes. Hence, precise identification of trait donor parents/lines/genotypes is critical. It forms the basis for the development of genetic and genomicresources for focusedmolecular breeding and transgenic program.Despite the realizedpotential of molecular breeding strategy, the success has not been encouraging. It ismainly attributed to the inability to identify robustQTLdue to thenonavailability of largenumber of codominantmarker systems such as SSR. Therefore, emphasis has to be ongeneration of large genomic resources of marker systems (SNP and SSR) followed bydevising robust and high-throughout phenotyping techniques.
Transgenics is yet another potential option to introduce novel alleles for improvingdrought tolerance of crop. In recent years, several transgenic programs have providedleads in improving crop adaptations, even at field level, suggesting the potentials andprospects of trait pyramiding by introducing regulatory genes with abilities toupregulate the several downstream functional genes. Suggestively, choice of regulatorygenes and development ofmultigene cassettes with pathway-engineered genes appearto be an effective drought-tolerant trait pyramiding strategy by transgenic approach.
53.7Potential of Genomic Resources for the Enhancement of Drought Tolerance
In the last decade, extensive research in genetics and genomics has improved ourunderstanding of structure and function of plant genomes and has integrated basicknowledge to enhance drought stress tolerance through molecular and transgenicapproaches. Cotton geneticmapshave been developed for variousGossypium crosses,mostly between G. barbadense species and G. hirsutum. These maps collectively
53.7 Potential of Genomic Resources for the Enhancement of Drought Tolerance j 1387

include more than 5000 public DNAmarkers (�3300 RFLPs, 700 AFLPs, more than2000 SSRs, and 100 SNPs). Many thousands of additional SSRs have beendescribed [197], but only subsets of these have been mapped [198–200]. The otheraspect of genomic resources is transcriptome, a cDNA sequence, which is beginningto receive wide attention. However, available knowledge of the Gossypium spp.transcriptome is heavily biased toward genes expressed in fibers. The total numberofGossypium ESTs available is 378 184 in different cotton species. On the basis of thecurrent knowledge on cotton genomic resources, Gene Chip (Affymetrix) has beenconstructed with 23 977 probe sets representing 21 854 cotton transcripts.
Over the past two decades, whole-genome sequencing has been achieved in severalorganisms including plant species, and it is clear that most of the major crop specieswill also be sequenced in near future including cotton genome. In cotton, however,there is need to sequence not only cultivated (tetraploid) genotypes but also theirdiploid progenitors, to understand how tetraploid cottons have come to �transgress�the productivity and quality of their progenitors. As a long-termgoal of characterizingthe spectrum of diversity among eight genome types, the first genetically anchoredwhole-genome physical map of a cotton species (G. raimondii) was built throughintegration of agarose-based fingerprinting and high information content finger-printing [201] since G. raimondii (D genome) has the smallest genome (60% of the�A� genome)with 880Mb, and a detailed genetic linkagemap is available. Studies arein progress on the species G. arboreum and G. hirsutum, the next priorities after G.raimondii in the international strategy for characterizing the spectrum of Gossypiumdiversity [202]. The economic importance of cotton fibers and scientific interest inpolyploidy suggest an ultimate goal of sequencing a G. hirsutum tetraploid. Thepossibility of intergenomic concerted evolution, much like the presence of recentlyamplified repetitive DNA families, may tend to support the need for a BAC-basedrather than awhole-genome shotgun approach.Using a finished diploid genome as atemplate and guide, a BAC-based sequence of a tetraploid will elucidate the types andfrequencies of changes that have distinguished polyploid from diploid cottons. Aneffective utilization of the existing genomic resources for precise identification ofQTL/genes of relevance for enhancing the adaptive capability to moisture-limitedconditions is need of the day.
53.7.1Molecular Breeding
The marker technology has enabled breeders to utilize DNA markers for selectingdesirable plantswithout exposing them to a critical and growth-limiting environmentalconditions [203]. The range of DNAmarkers that can be easily used is quite extensive.The techniques utilize the polymorphisms in the actual DNA sequence of plantgermplasm and identify the variations and use different DNAmarkers such as RFLP,AFLP, SSR, and so on [204–206]. SuchDNAmarkers have been developed and used incotton for fingerprinting [207, 208], linkage map construction [198, 199, 209–212],gene mapping [213, 214], and genetic diversity studies [209, 210]. Several studies andreviews have reported that QTL may play a critical role in mitigating abiotic stress in
1388j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs
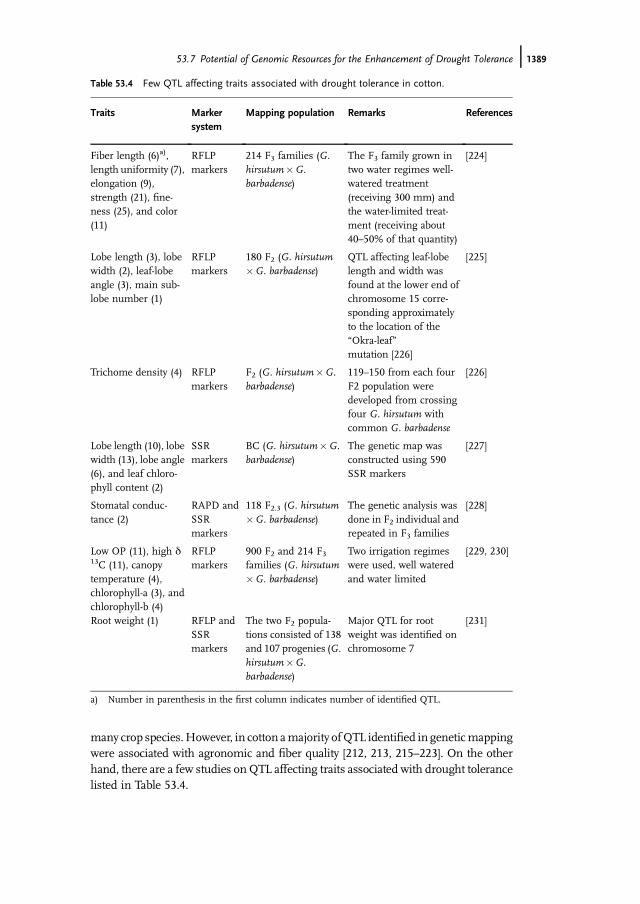
many crop species.However, in cotton amajority ofQTL identified in geneticmappingwere associated with agronomic and fiber quality [212, 213, 215–223]. On the otherhand, there are a few studies on QTL affecting traits associated with drought tolerancelisted in Table 53.4.
Table 53.4 Few QTL affecting traits associated with drought tolerance in cotton.
Traits Markersystem
Mapping population Remarks References
Fiber length (6)a),length uniformity (7),elongation (9),strength (21), fine-ness (25), and color(11)
RFLPmarkers
214 F3 families (G.hirsutum�G.barbadense)
The F3 family grown intwo water regimes well-watered treatment(receiving 300 mm) andthe water-limited treat-ment (receiving about40–50% of that quantity)
[224]
Lobe length (3), lobewidth (2), leaf-lobeangle (3), main sub-lobe number (1)
RFLPmarkers
180 F2 (G. hirsutum�G. barbadense)
QTL affecting leaf-lobelength and width wasfound at the lower end ofchromosome 15 corre-sponding approximatelyto the location of the�Okra-leaf�mutation [226]
[225]
Trichome density (4) RFLPmarkers
F2 (G. hirsutum�G.barbadense)
119–150 from each fourF2 population weredeveloped from crossingfour G. hirsutum withcommon G. barbadense
[226]
Lobe length (10), lobewidth (13), lobe angle(6), and leaf chloro-phyll content (2)
SSRmarkers
BC (G. hirsutum�G.barbadense)
The genetic map wasconstructed using 590SSR markers
[227]
Stomatal conduc-tance (2)
RAPD andSSRmarkers
118 F2.3 (G. hirsutum�G. barbadense)
The genetic analysis wasdone in F2 individual andrepeated in F3 families
[228]
Low OP (11), high d13C (11), canopytemperature (4),chlorophyll-a (3), andchlorophyll-b (4)
RFLPmarkers
900 F2 and 214 F3families (G. hirsutum�G. barbadense)
Two irrigation regimeswere used, well wateredand water limited
[229, 230]
Root weight (1) RFLP andSSRmarkers
The two F2 popula-tions consisted of 138and 107 progenies (G.hirsutum�G.barbadense)
Major QTL for rootweight was identified onchromosome 7
[231]
a) Number in parenthesis in the first column indicates number of identified QTL.
53.7 Potential of Genomic Resources for the Enhancement of Drought Tolerance j 1389

Genomic approaches offer unique opportunities to dissect quantitative traits intotheir single genetic determinants (QTL), thus enabling transfer of specific genomicregions between different genetic backgrounds through MAS [196]. In cotton, MAShas been used in backcrossing transgenes from transformed cultivars to the elitecultivars and further genetic advancement of cultivars. QTL for yield and drought-related physiological traits such as osmotic potential (OP), carbon isotope ratio(d13C), and leaf chlorophyll content have been introgressed via MAS to 12 NILobtained from elite cultivars of the two cotton species G. barbadense cv. F-177 and G.hirsutum cv. Siv�on. Among nine NILs introgressed with OPQTL, five NILs exhibitedsignificant reduction in OP under water-limited conditions and/or improved OAduring two field trials. Similarly, among six NILs introgressed with high d13C (asindirectly measurement of WUE) only four showed significantly higher d13C undermoisture stress conditions. Likewise, only two out of three NILs showed higherchlorophyll content [182].
Over the past decades, conventional linkage analysis (biparenteral QTL mapping)is being successfully used for dissection of trait inheritance mechanism. Theapproach has facilitated the identification of major genes and QTL in plant andanimal species, particularly in model organisms. However, efficient gene discoverywith this approach will probably continue to be largely limited to loci that have a largeeffect on quantitative variation. Although biparenteralmapping is used extensively toidentify QTL associated with different traits, it is time consuming and takes severalyears to develop populations for fine scale mapping. Apart from inherently poorresolution (long-distance associations betweenmarker andQTL, for example, 10 cM)resulting from limitedmeiotic crossover events in pedigreed populations, developinglarge full sib families for each major gene/trait may not be practical for plant geneticimprovement, particularly in tree crops. A more efficient approach that does notrequire generation of large pedigreedmapping populations with higher resolution istherefore needed to complement conventional QTL mapping strategy. Recently, apopulation genomics tool termed �association mapping� [232, 233] has been devel-oped. Association mapping seeks to identify specific functional variants (i.e., loci,alleles, etc.) linked to phenotypic differences in a trait, to facilitate detection of traitcausing DNA sequence polymorphisms, and/or selection of genotypes that closelyresemble the phenotype. In cotton (Gossypium spp.), association studies are verylimited [25, 234–236].
Abdurakhmonov et al. [234] reported the extent of genome-wide LD in uplandcotton (G. hirsutum) and association mapping of fiber quality traits using 95microsatellite markers in a total of 285 exotic G. hirsutum accessions. A genome-wide average of LD declined (r2� 0.1) within the genetic distance at <10 cM in thelandrace stocks of germplasm and >30 cM in varietal germplasms. Furthermore,genome-wide LD (r2� 0.2) reduced on an average �1–2 cM in the landrace stockgermplasms and 6–8 cM in varietal germplasms. Recently, association mapping forfiber-quality traitswas conductedusing 202microsatellitemarkers in 335G. hirsutumgermplasm grown in two diverse environments. The result indicates that genome-wide average of LD (r2� 0.1) extended up to a genetic distance of 25 cM in assayedcotton variety accessions. Genome-wide LD (r2� 0.2) was reduced to �5–6 cM
1390j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs
providing evidence for the potentiality of association mapping of agronomicallyimportant traits in cotton [235]. Similarly, Kantartzi and Stewart [236] evaluated 56G.arboreum (diploid cotton) germplasm accessions for eight fiber characters thatgenotyped with 98 SSR markers. A total of 30 marker-trait associations wereidentified with 19 SSR markers located on 11 chromosomes and identified mark-er-trait associations (P¼ 0.05) for all traits evaluated.
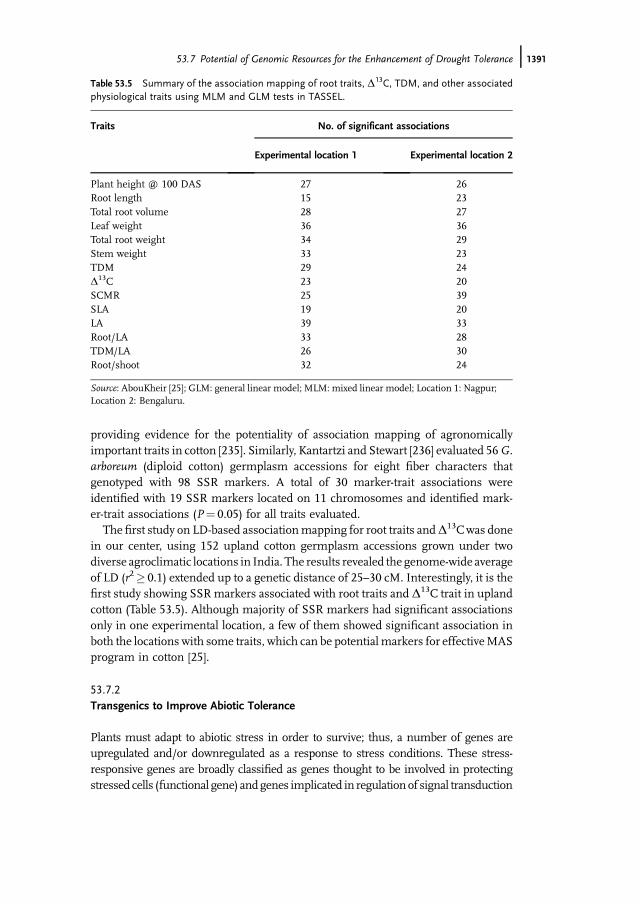
The first study on LD-based associationmapping for root traits andD13Cwas donein our center, using 152 upland cotton germplasm accessions grown under twodiverse agroclimatic locations in India. The results revealed the genome-wide averageof LD (r2� 0.1) extended up to a genetic distance of 25–30 cM. Interestingly, it is thefirst study showing SSRmarkers associated with root traits and D13C trait in uplandcotton (Table 53.5). Although majority of SSR markers had significant associationsonly in one experimental location, a few of them showed significant association inboth the locations with some traits, which can be potential markers for effectiveMASprogram in cotton [25].
53.7.2Transgenics to Improve Abiotic Tolerance
Plants must adapt to abiotic stress in order to survive; thus, a number of genes areupregulated and/or downregulated as a response to stress conditions. These stress-responsive genes are broadly classified as genes thought to be involved in protectingstressed cells (functional gene) andgenes implicated in regulationof signal transduction
Table 53.5 Summary of the association mapping of root traits, D13C, TDM, and other associatedphysiological traits using MLM and GLM tests in TASSEL.
Traits No. of significant associations
Experimental location 1 Experimental location 2
Plant height @ 100 DAS 27 26Root length 15 23Total root volume 28 27Leaf weight 36 36Total root weight 34 29Stem weight 33 23TDM 29 24D13C 23 20SCMR 25 39SLA 19 20LA 39 33Root/LA 33 28TDM/LA 26 30Root/shoot 32 24
Source: AbouKheir [25]; GLM: general linear model; MLM: mixed linear model; Location 1: Nagpur;Location 2: Bengaluru.
53.7 Potential of Genomic Resources for the Enhancement of Drought Tolerance j 1391

and expression (transcription factor genes) [237, 238]. These genes have been usedextensively to enhance the stress tolerance in several of plant species. Some of therelevant genes, which enhance the stress tolerance in cotton, are provided in Table 53.6.
Although the first transgenic cotton plant (Bt cotton) was released and commer-cially cultivated since 1996, to date there is no transgenic cotton for abiotic stresstolerance. This is perhaps due to the complexity of abiotic stress and lack of efficienttransformation technique in cotton. Therefore, there is a need to develop efficienttransformation technique that can be used effectively in cotton. Recently, efficient
Table 53.6 Genes used in transformation to enhance stress tolerance in cotton.
Functional genes Remarks References
1. AntioxidantsSuperoxide dismutase(SOD), ascorbate peroxi-dase (APX), and glutathi-one reductase (GR)
Enhanced recovery of photosynthesis in plantsexposed to chilling temperatures and high lightintensity
[239, 240]
GlutathioneS-transferase(GST)
Seedlings did not show improved tolerance tosalinity, chilling conditions, or herbicides
[241]
2. Membrane transportersTonoplast Naþ/Hþ anti-porters (NHX1)
Transgenic plants had more biomass produced andfibers under salinity stress compared to the wild type
[242]
Hþ -PPase gene (TsVP) Transgenic plants showed high shoot and rootgrowth, better photosynthetic performance, andaccumulated more Naþ , Kþ , Ca2þ , Cl�, and solublesugars in their root and leaf tissues under salinityconditions compared to the wild type
[243, 244]
3. Osmotic potentialCholine monooxygenase(CMO)
The osmotic potential, electrolyte leakage, andmalondialdehyde (MDA) accumulation weresignificantly lower in leaves of the transgenic lines thanin wild type after salt stress. Net photosynthesis and Fv/Fm was less affected by salinity stress in transgenic
[245]
4. Protein kinasesSOS2 Transgenic plant had better salt tolerance than the
wild type[246]
5. Transcription factorsGF14l, encodes 14-3-3group of regulatoryproteins
Overexpression of GF14l in cotton conferred a �stay-green� phenotype under well-watered conditions anddisplayed increase water stress tolerance and main-tained higher photosynthetic rates under conditions oflow water availability
[247]
ABF3 Transgenic cotton plants enhanced expression of ABA-responsive genes under noninducting conditions,better root development, and enhanced survival undersevere water deficit conditions
[246]
1392j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs

embryogenic calli-based method of obtaining transgenic cotton has been standard-ized in two chinese cultivars [248]. In cotton, due to its recalcitrance and poorregeneration, there has been limited progress in achieving effective transformation.A recent alternative method, tissue culture-independent in planta transformationstrategy, has been successfully used for transformation in cotton [249].
53.8Summary
Drought is perhaps the most complex abiotic stress and the varied adaptive mechan-isms evolved by plants have made the task of improving drought tolerance aformidable challenge. Imperatively, projected prevalence and spread of droughtstress due to water deficit situations worldwide has prioritized the efforts in cropimprovement program to dissect the molecular and physiological basis of adapta-tions to drought stress and/or water-limited conditions by various crops. Pastresearch efforts have emphasized on improving constituent drought tolerance traitsthrough a �trait-based breeding� strategy. Thus, the major emphasis has to identifydrought-tolerant traits such as root traits, WUE, and so on, and develop precise andhigh-throughput phenotyping technique for screening a large number of germplasmto select trait donor lines that can be utilized in breeding program.Modern genomictechniques have aided tremendously in identifying stress QTL and diagnostic DNAmarkers for specific stress-related traits in a number of crops including cotton.Improvement-specific traits by introducingnewalleles by transgenic approachwouldbe an essential option to pyramid traits for improving the adaptation. Furthermore,turning the gene-tagging efforts frombiparental crosses to germplasmcollection andfrom traditional linkage mapping to linkage disequilibrium (LD)-based associationstudy holds promise formost effective utilization of ex situ conserved natural geneticdiversity of cotton germplasm resources irrespective of their origin. The applicationof LD-based association mapping for cotton will not only accelerate the MASprograms in cotton but also add to our knowledge and understanding of the cottongenome and its evolution. Furthermore, genomic resources with drought-toleranttraits can be compatibly used for pyramiding to superior genetic background, whichfacilitates genetic improvement for higher yield of cotton germplasm and enhancedadaptability to drought stress conditions.
References
1 Fryxell, P.A. (1968) Bot. Gazette, 129,296–308.
2 Fryxell, P.A. (1979) The Natural History ofthe Cotton Tribe, Texas A&M UniversityPress, College Station, TX.
3 Fryxell, P.A. (1992) Rheedea, 2,108–165.
4 Wendel, J.F., Brubaker, C., Alvarez, I.,Cronn, R., and Stewart, J.M.C.D., (2009)Evolution and Natural History of theCotton Genus In: Genetics andGenomics of Cotton, A.H. Paterson (ed.),Plant Genetics andGenomics: Crops and
References j 1393

Models 3, Springer ScienceþBusinessMedia, LLC (2009), p3–22.
5 Ulloa, M., Stewart, J.M.C.D., Garcia,C.E.A. et al. (2006) Genet. Resour. CropEvol., 53, 653–668.
6 Brubaker, C.L. and Wendel, J.F. (1994)Am. J. Bot., 81, 1309–1326.
7 ICAC (2005) Cotton Production Practices.Technical Information Section,International Cotton AdvisoryCommittee, Washington DC, USA.
8 ICAC (2004) Cotton: Review of the WorldSituation, International Cotton AdvisoryCommittee, Washington D.C., p. 20.
9 FAO (Food and AgricultureOrganization) (2006) FAO ProductionYear Book, FAO.
10 Khadi, B.M., Santhy, V., and Yadav, M.S.(2010) Cotton: an introduction, inCotton,Biotechnology in Agriculture and Forestry65 (ed. U.B. Zehr), Springer, Berlin,pp. 1–14.
11 FAO (Food and AgricultureOrganization) (2007) FAO ProductionYear Book, FAO.
12 Bot, A.J., Nachtergaele, F.O., and Young.,A. (2000) World Soil Resources Reports 90.Land and Water Development Division,FAO, Rome.
13 Sharp,R.E., Poroyko,V.,Hejlek, L.G. et al.(2004) J. Exp. Bot., 55, 2343–2351.
14 Paterson, A.H., Lander, E.S., Hewitt, J.D.et al. (1988) Nature, 335, 721–726.
15 Rosenow, D.T., Quisenberry, J.E., Wendt,C.W. et al. (1983)Agric. Water Manage., 7,207–222.
16 Hearn, A.B. (1969a) J. Agric. Sci. Camb.,73, 75–86.
17 Marani, A. and Levi., D. (1973) Agron. J.,65, 637–641.
18 Constable, G.A. and Hearn, A.B. (1981)Irrigation Sci., 3, 17–28.
19 Pace, P.F., Cralle, H.T., El-Halawany, S.H.M.et al. (1999) J. Cotton Sci., 3, 183–187.
20 Constable, G.A. and Rawson, H.M.(1980) Aust. J. Plant Physiol., 7, 89–100.
21 Krieg, D.R. (1981) Leaf development andfunction as related to water stress, inProceedings of the Beltwide CottonProduction Reserach Conference (ed. J.M.Brown), New Orleans, LA, 4–8 January,1981, National Cotton Council,Memphis, TN, pp. 41–42.
22 Marani, A., Baker, D.N., Reddy, V.R. et al.(1985) Crop Sci., 25, 798–802.
23 Krieg, D.R. (1997) Proceedings of theBeltwide Cotton Production ReserachConference, National Cotton Council,Memphis, TN, pp. 1337–1347.
24 Cook, C.G. (1985) MSc thesis, TexasA&M University, College Station, TX.
25 AbouKheir, E. (2010) PhD thesis,University of Agricultural Sciences,Bengalaru, India.
26 Hearn, A.B. (1969) J. Agric. Sci. Camb.,73, 87–97.
27 Grimes, D.W. and Yamada, H. (1982)Crop Sci., 22, 134–139.
28 Parida, A.K., Dagaonkar, V.S., Phalak,M.S. et al. (2007) Plant Biotech. Rep., 1,37–48.
29 Parida, A.K., Dagaonkar, V.S., Phalak,M.S. et al. (2008) Acta Physiol. Plant., 30,619–627.
30 Klepper, B., Taylor, H.M., Huck, M.G.et al. (1973) Agron. J., 65, 307–310.
31 Taylor, H.M. and Klepper, B. (1974)Agron. J., 66, 584–588.
32 Ball, R.A., Oosterhuis, D.M., andMauromoustakos, A. (1994) Agron. J., 86,788–795.
33 Fern�andez, C.J., Cothren, J.T., andMcInnes., K.J. (1996) J. Plant Nutr., 19,595–617.
34 Mauney, J.R. (1986) Vegetative growthand development of fruiting sites, inCotton Physiology (eds J.R. Mauney andJ.McD. Stewart), The cotton Foundation,Memphis, TN, pp. 11–28.
35 Jackson, B.S. andGerik, T.J. (1990)Agron.J., 82, 483–488.
36 Morrow, M.R. and Krieg, D.R. (1990)Agron. J., 82, 52–56.
37 Baker, D.N., Bruce, R.R., and McKinion.,J.M. (1973) Proceedings of the 1972Cotton Production Research Conference,pp. 110–114.
38 Wullschleger, S.D. and Oosterhuis, D.M.(1990) Crop. Sci., 30, 1259–1264.
39 Gerick, T.J., Faver, K.L., Thaxton, P.M.et al. (1996) Crop. Sci., 36, 914–921.
40 McMichael, B.L., Jordan, W.R., andPowell, R.D. (1973) Agron. J., 65,202–204.
41 Hearn, A.B. (1976) J. Agric. Sci. Camb.,86, 257–269.
1394j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs

42 Guinn, G. and Mauney, J.R. (1984a)Agron. J., 76, 90–94.
43 Guinn, G. and Mauney, J.R. (1984b)Agron. J., 76, 94–98.
44 Antony, A.K. and Kutty, K.E. (1975) Ind. J.Agr. Sci., 45, 199–203.
45 Longenecker, D.E. and Erie, L.J. (1968)Irrigation water management, inAdvances in Production and Utilization ofQuality Cotton: Principle and Practices (edsF.C. Elliot, M. Hoover, and W.K. Poter),Iowa State University Press, San Diego,CA, pp. 321–345.
46 Bilbro, J.D. (1962)Texas Agri. Exp. StationM.P., 611, 1–8.
47 Levi, A., Ovnat, L., Paterson, A.H. et al.(2009a) Plant Sci., 177, 88–96.
48 Mahmood, S., Irfan, M., Raheel, F. et al.(2006) Int. J. Agri. Biol., 8 (6), 796–800.
49 Cutler, J.M. and Rains, D.W. (1977) CropSci., 17, 329–335.
50 Radin, J.W. and Mauney, J.R. (1986) Thenitrogen stress syndrome, in CottonPhysiology, The Cotton Foundation,Memphis, pp. 91–105.
51 McMichael, B.L. andHesketh, J.D. (1977)Proceedings of the 31st CottonPhysiology Conference, pp. 62–63.
52 Hearn, A.B. (1979) Outlook Agric., 10,159–166.
53 Cothren, J.T. (1999) Physiology of thecotton plant, in Cotton (eds C.W. Smithand J.T. Colthern), John Wiley & Sons,Inc., New York, pp. 207–268.
54 Krieg, D.R. (1983) Agric. Water Manage.,7, 249–263.
55 Krieg, D.R. (1986) Feedback control andstress effects on photosynthesis, inCotton Physiology (eds J.R. Mauney andJ.McD. Stewart), The Cotton Foundation,Memphis, TN, pp. 227–242.
56 Jones, H.G. (1973) New Phytol., 72,1095–1105.
57 Iqbal, J., Reddy, O.U.K., EL-Zik, K.M.et al. (2001) Theor. Appl. Genet., 103,547–554.
58 Rungis, D., Llewellyn, D., Dennis, E.S.et al. (2005) J. Agric. Res., 56, 301–307.
59 Miflin, B. (2000) J. Exp. Bot., 51 (342), 1–8.60 Collins, N.C., Tardieu, F., and Tuberosa,
R. (2008) Plant Physiol., 147, 469–486.61 Blum, A. (2009) Field Crops Res., 112,
119–123.
62 Passioura, J.B. (1976) Aust. J. PlantPhysiol., 3, 559–565.
63 Passioura, J.B. (1986) Aust. J. PlantPhysiol., 13, 191–201.
64 AbouKheir, E., Sheshshayee, M.S.,Udayakumar, M. et al. (2008) AABInternational Conference on ResourceCapture by Crops: Integrated Approach,14–16 September 2008, University ofNottingham at Sutton BoningtonCampus, UK.
65 Andries, J.A., Jones, J.E., Sloane, L.W.et al. (1969) Crop Sci., 9, 705–710.
66 Heitholt, J.J. (1993) Crop. Sci., 33,486–490.
67 Kerby, T.A., Buxton, D.R., and Matsuda,K. (1980) Crop Sci., 20, 208–213.
68 Pettigrew, W.T., Heitholt, J.J., andMeredith, W.R. (1993) Agron. J., 85,821–825.
69 Kerby, T.A. and Buxton, D.R. (1978)Agron. J., 70, 535–538.
70 Wells, R. and Meredith, W.R. (1986) CropSci., 26, 223–228.
71 Meredith, W.R. (1984) Crop Sci., 24,855–857.
72 Stiller, W.N., Reid, P.E., and Constable,G.A. (2004) Agron. J., 96, 656–664.
73 McMichael, B.L. (1986) Growth of roots,inCotton Physiology (eds J.R. Mauney andJ.McD. Stewart), The Cotton Foundation,Memphis, TN, pp. 29–38.
74 McMichael, B.L., Burke, J.J., Berlin, J.D.et al. (1985) Environ. Exp. Bot., 25, 23–30.
75 McMichael, B.L., Oosterhuis, D.M., Zak,J.C. et al. (2010)Growth and developmentof root systems, in Cotton Physiology (edsJ.McD. Stewart, D.M. Oosterhuis, J.J.Heitholt, and J.R. Mouney), SpringerScience, pp. 57–71.
76 Hons, F.M. and McMichael, B.L. (1986)Field Crops Res., 13, 147–158.
77 McMichael, B.L. and Quisenberry, J.E.(1991) Environ. Exp. Bot., 31, 461–470.
78 Ludlow, M.M. and Muchow, R.C. (1990)Adv. Agron., 43, 107–153.
79 Prior, S.A., Rogers, H.H., Runion, G.B.et al. (1995) J. Environ.Qual., 24, 678–683.
80 Plaut, Z., Carmi, A., and Grava, A. (1996)Irrigation Sci., 16, 107–113.
81 Huang, B., Duncan, R.R., andCarrow., R.N. (1997) Crop Sci., 37,1863–1869.
References j 1395

82 Blum, A. (2005) Aust. J. Agri. Res., 56,1159–1168.
83 Martinez, F., Merino, O., Garcia, M.D.et al. (1998) Plant Soil, 201, 209–216.
84 Tuberosa, R., Sanguineti, M.C., Landi, P.et al. (2002) Plant Mol. Biol., 48 (5/6),697–712.
85 Drouet, J.L., Pag�es, L., and Serra, V.(2005) Euro. J. Agron., 22, 185–193.
86 Udayakumar, M., Rao, R.C.N., Wright,G.C. et al. (1998) J. Plant Biol., 1,69–75.
87 Taylor, H.M., Upchurch, D.R., Brown,J.M. et al. (1991) Some methods of rootinvestigation, in Plant Roots and TheirEnvironment (eds B.L. McMichael and H.Persson), Elsevier Science Publishers,Inc., New York.
88 Sheshshayee, M.S., AbouKheir, E.,Sreevathsa, R. et al. (2011) Sustainingcrop productivity through improvementin root traits: phenotyping and breedingfor relevant traits, in Root Genomics (ed.A.D. de Oliveira), Springer, TheNetherlands, pp. 205–232.
89 Basal, H., Bebeli, P., Smith, C.W. et al.(2003) Crop Sci., 43, 1983–1988.
90 Roark, B. and Quisenberry, J.E. (1977)Proceedings of the Beltwide CottonProduction Research Conference,Atlanta, GA. 10–12 January, 1977,National Cotton Council of America,Memphis, TN, pp. 49–50.
91 Quisenberry, J.E., Jordan, W.R., Roark,B.A. et al. (1981) Crop Sci., 21, 889–895.
92 Craig, L. and Gordon, L.I. (1965)Deuterium and oxygen-18 variations inthe ocean and the marine atmosphere, inProceedings of a Conference on StableIsotopes in Oceanographic Studies andPaleotemperatures (ed. E. Tongiorgi),Spoleto, Italy, pp. 9–130.
93 Flanagan, L.B., Bain, J.F., and Ehleringer,J.R. (1991) Oecologia, 88, 394–400.
94 Flanagan, L.B., Phillips, S.L., Ehleringer,J.R. et al. (1994) Aust. J. Plant Physiol., 21,221–234.
95 Farquhar, G.D. and Lloyd, J. (1993)Carbon and oxygen isotope effects in theexchange of carbon dioxide betweenterrestrial plants and the atmosphere, inStable Isotopes and Plant Carbon–WaterRelations (eds J.R. Ehleringer, A.E. Hall,
andG.D. Farquhar), Academic Press, SanDiego, pp. 47–70.
96 Farquhar,G.D.,Cernusak,L.A.,andBarnes,B. (2007) Plant Physiol., 143, 11–18.
97 Gonfiantini, R., Gratziu, S., andTongiorgi, E.,Technical Report Series No.206. Isotopic Atomic EnergyCommission, Vienna, 405–410. (1965)
98 DeNiro, M.J. and Epstein, S. (1979)Science, 204, 51–53.
99 Sheshshayee, M.S., Bindumadhava, H.,Ramesh, R. et al. (2005) J. Exp. Bot., 56,3033–3039.
100 AbouKheir, E., Prasad, T.G.,Sheshshayee, M.S. et al. (2010) AsianAust. J. Plant Sci. Biotech., 4 (1), 12–18.
101 Sinclair, T.R., Tanner, C.B., and Bennett,J.M. (1984) Water-use efficiency in cropproduction. BioSci., 34, 36–40.
102 Tanner, C.B. and Sinclair, T.R. (1983)Efficient water use in crop production:research or re-search? in Limitations toEfficient Water Use in Crop Production (edsH.M. Taylor, W.R. Jordan, and T.R.Sinclair), The American Society ofAgronomy (ASA),Crop Science Society ofAmerica (CSSA), and Soil Science Societyof America (SSSA) Madison, Wisconsin,USA, pp. 1–25.
103 Hubick, K.T., Farquhar, G.D., andShorter, R. (1986) Aust. J. Plant Physiol.,13, 803–816.
104 Wright, G.C.,Hubick, K.T., and Farquhar,G.D. (1988) Aust. J. Plant Physiol., 15,815–825.
105 Wright, G.C., Rao, R.C.N., and Farquhar,G.D. (1994) Crop. Sci., 34, 92–97.
106 Udayakumar, M., Sheshshayee, M.S.,Nataraj, K.N. et al. (1998) Current Sci., 74,994–1000.
107 Rao, R.C.N., Williams, J.H., Wadia,K.D.R. et al. (1993) Ann. Appl. Biol., 122,357–367.
108 Ashok, R.S., Aftab Hussein, I.S., Wright,G.C. et al. (1999) Funct. Plant Bio., 26,503–510.
109 Ebdon, J.S., Petrovic, A.M., and Dawson,T.E. (1998) Crop Sci., 38 (1), 157–162.
110 Sheshshayee, M.S., Bindumadhava, H.,Shankar, A.G. et al. (2003) J. Plant Biol.,30 (2), 253–268.
111 Saranga, Y., Flash, I., and Yakir, D. (1998)Crop Sci., 38, 782–787.
1396j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs

112 Pettigrew, W.T. (2004b) Crop Sci., 44,1265–1272.
113 Stiller,W.N.,Read, J.J.,Constable,G.A., andReid, P.E. (2005) Crop. Sci., 45, 1107–1113.
114 Baker, D.A. (1984) Water relations, inAdvanced Plant Physiology (ed. M.B.Wilkins), Pitman, London.
115 Voloudakis, A.E., Kosmas, S.A., Tsakas, S.etal.(2002)Funct.PlantBiol.,29,1237–1245.
116 Basal,H., Smith, C.W., Thaxton, P.S. et al.(2005) Crop Sci., 45, 766–771.
117 Farquhar, G.D., Ehleringer, J.R., andHubick, K.T. (1989) Ann. Rev. PlantPhysiol. Plant Mol. Bio., 40, 503–537.
118 Farquhar, G.D. and Richards, R.A. (1984)Aust. J. Plant Physiol., 11, 539–552.
119 Condon, A.G., Farquhar, G.D., andRichards, R.A. (1990) Aust. J. PlantPhysiol., 17, 9–22.
120 Condon, A.G., Richards, R.A.,Rebetzke, G.J. et al. (2004) J. Exp. Bot., 55,2447–2460.
121 Wright, G.C., Hubick, K.T., Farquhar,G.D. et al. (1993) Genetic andenvironmental variation in transpirationefficiency and its correlation with carbonisotope discrimination and specific leafarea in peanut, in Stable Isotope and PlantCarbon–Water Relations (eds J.R.Ehleringer, A.E. Hall, and G.D.Farquhar), Academic Press, San Diego,USA, pp. 247–267.
122 Nageswara Rao, R.C., Udayakumar, M.,Farquhar, G.D. et al. (1995) Aust. J. PlantPhysiol., 22, 545–551.
123 Bindu Madhava, H., Sheshshayee, M.S.,Shashidhar, G. et al. (2005) Curr. Sci.,189 (7), 1256–1258.
124 Impa, S.M., Nadaradjan, S.,Boominathan, P. et al. (2005) Crop. Sci.,45, 2517–2522.
125 Nadaradjan, S., Impa, S.M., Sheshshayee,M.S. et al. (2005) Second InternationalCongress of Plant Physiology, 8–12January 2003, New Delhi, India.
126 Ismail, A.M. and Hall, A.E. (1992) CropSci., 32, 7–12.
127 Ismail, A.M. and Hall, A.E. (1993) CropSci., 33, 498–503.
128 Lu, C., Richards, G.P., Sharti, M.R. et al.(1996)Aust. J. Plant. Physiol., 23, 127–132.
129 Leidi, E.O., Lopez, M., Gorham, J. et al.(1999) Field Crops Res., 61, 109–123.
130 Rebetzke, G.J., Condon, A.G.,Richards, R.A. et al. (2002) Crop Sci., 42,739–745.
131 Richards, R.A., Rebetzke, G.J., Condon,A.G. et al. (2002) Crop Sci., 42, 111–121.
132 Condon, A.G., Richards, R.A.,Rebetzke, G.J. et al. (2002) Crop Sci., 42,122–131.
133 White, J.W., Castillo, J.A., and Ehleringer,J. (1990) Aust. J. Plant Physiol., 17, 189–198.
134 Scheindegger, Y., Saurer, M., Bahn, M.et al. (2000) Oecologia, 125, 350–357.
135 Hubick, K.T., Shorter, R., and Farquhar,G.D. (1988) Aust. J. Plant Physiol., 15,799–813.
136 Rao, R.C.N., Udayakumar, M., Farquhar,G.D. et al. (1995)Aust. J. Plant Physiol., 22,545–551.
137 White, J.W. (1993) Implications of carbonisotope discrimination studies forbreeding common bean under waterdeficits, in Stable Isotopes and PlantCarbon–Water Relations (eds J.R.Ehleringer, A.E. Hall, and G.D.Farquhar), Academic Press, pp. 389–396.
138 Johnson, D.A., Asay, K.H., and Read, J.J.(1993) Stable Isotopes and Plant Carbon–Water Relations (eds J.R. Ehleringer, A.E.Hall, and G.D. Farquhar), AcademicPress, New York, pp. 269–280.
139 Matus, A., Slinkard, A., and Kessel, C.V.(1995) Crop Sci., 35, 1267–1271.
140 Sun, Z.J., Livingston, N.J., Guy, R.D. et al.(1996) Plant Cell Environ., 19, 887–894.
141 Condon, A.G., Richards, R.A., andFarquhar, G.D. (1993) Aust. J. Agri. Res.,44, 1693–1711.
142 Farquhar, G.D., Hubick, K.T., Condon,A.G. et al. (1988) Carbon isotopefractionation and plant water useefficiency, in Stable Isotopes in EcologicalResearch (eds P.W. Rundel, J.R.Ehleringer, and K.A. Nagy), Springer,New York, pp. 21–40.
143 Acevedo, E. (1993) Stable isotopes andplant carbon–water relations, in StableIsotopes and Plant Carbon–Water Relations(eds J.R. Ehleringer, A.E. Hall, and G.D.Farquhar), Academic Press, San Diego,USA, pp. 398–417.
144 Ehdaie, B.,Hall, A.E., Farquhar,G.D. et al.(1991) Crop Sci., 31, 1282–1288.
References j 1397

145 Richards, R.A. and Condon, A.G. (1993)Stable Isotopes and Plant Carbon–WaterRelations (eds J.R. Ehleringer, A.E. Hall,and G.D. Farquhar), Academic Press,New York, pp. 451–462.
146 Meinzer, F.C., Rundel, P.W., and Schultz,L.M. (1990) Cov. Funct. Ecol., 4, 579–584.
147 Richards,R.A. (1996)PlantGrowthRegul.,20, 157–166.
148 Weete, J.D., Leek, G.L., Peterson, C.M.et al. (1978) Plant Physiol., 62, 675–677.
149 Oosterhuis, D.M., Hampton, R.E., andWullschleger, S.D. (1991) J. Prod. Agric.,4, 260–265.
150 Bondada, B.R., Oosterhuis, D.M.,Murphy, J.B. et al. (1996) Envi. Exp. Bot.,36 (1), 61–69.
151 Haque, M.M., Mackill, D.J., and Ingram,K.T. (1992) Crop Sci., 32, 865–868.
152 Clarke, J.M, and Townley-Smith, T.F.(1986) Crop Sci., 26, 289–292.
153 Clarke, J.M.(1987)Useofphysiologicalandmorphological traits in breedingprogrammes to improve droughtresistanceof cereals, inDroughtTolerance inWinter Cereals (eds J.P. Srivastiva, E.Proceddu,E.Acevedo,andS.Verma),WileyInterscience, New York, pp. 171–189.
154 Clarke, J.M.,DePauw, R.M., andTownley-Smith, T.F. (1992) Crop Sci., 32, 723–728.
155 Ahmad, R.T., Malik, T.A., Khan, I.A. et al.(2009) Int. J. Agric. Biol., 11, 235–240.
156 Burke, J.J., O�Mahony, P.J., and Oleiver.,M.J. (2000) Plant Physiol., 123, 575–587.
157 Sun, W., Bernard, C., van de Cotte, B.M.et al. (2001) Plant J., 27, 407–415.
158 Sung, D.Y., Kaplan, F., Lee, K.J. et al.(2003) Trends Plant Sci., 8, 179–187.
159 Larkindale, J., Hall, J.D., Knight, M.R.et al. (2005) Plant Physiol., 138, 882–897.
160 SenthilKumar, M., Kumar, G.,Srikanthbabu, V. et al. (2006) J. PlantPhysiol., 164, 111–125.
161 AbouKheir, E., Sheshshayee,M.S., Prasad,T.G. et al. (2010) J. Cotton Sci. (in press).
162 Kumar, G., Krishnaprasad, B.T., Savitha,M. et al. (1999) Theor. Appl. Genet., 99,359–367.
163 Gurley, W.B. (2000) Plant Cell, 12,457–460.
164 Srikanthbabu, V., Kumar, G.,Krishnaprasad, B.T. et al. (2002) J. PlantPhysiol., 159, 535–545.
165 SenthilKumar, M., Srikanthbabu, V.,Mohan Raju, B. et al. (2003) J. Exp. Bot.,54 (392), 2569–2578.
166 Kuznetsov, V.V., Rakitin, V.Y., andZholkevich, V.N. (1999) Physiol. Plant.,107, 399–406.
167 Seki, M., Narusaka, H., Abe, H. et al.(2001) Plant Cell, 13, 61–72.
168 Zhu, J.-K. (2002) Ann. Rev. Plant Bio., 53,247–273.
169 Krishnan, M., Nguyen, H.T., and Burke,J.J. (1989) Plant Physiol., 90, 140–145.
170 Uma, S., Prasad, T.G., and Udayakumar,M. (1995) Ann. Bot., 76, 43–49.
171 Jayaprakash, T.L., Ramamohan, G.,Krishna Prasad, B.T. et al. (1998) Ann.Bot., 82, 513–522.
172 Burke, J.J. (2001)Physiol. Plant., 112, 167–170.
173 Zimmerman, U. (1978) Ann. Rev. PlantPhysiol., 29, 122–148.
174 Hare, P.D., Cress, W.A., and van Staden,J. (1998) Plant Cell Environ., 21, 535–553.
175 Boyer, J.S. (1996) Adv. Agron., 56, 187–218.
176 Oosterhuis, D.M. and Wullschleger, S.D.(1988) Arkansas Farm Res., 37, 12.
177 Cutler, J.M. and Rains., S.W. (1978)Physiol. Plant., 42, 261–268.
178 Eaton, F.M. and Ergle., D.R. (1948) Plant.Physiol., 23, 169–187.
179 Murray, A.K. (1999) Myo-inositol,sucrosyl oligosaccharide metabolism anddrought stress in developing cottonfibers, in vivo, in vitro and in plant, inProceedings of the Beltwide CottonProduction Research Conference, January1999 (ed. J.M. Brown), National CottonCouncil, Memphis, TM, pp. 518–520.
180 Gorham, J. (1996) Phytochemistry, 43,367–369.
181 Kosmas, C., Marathianou, M.,Gerontidis, St. et al. (2001) Agric. WaterManage., 48, 61–78.
182 Levi, A., Paterson, A.H., Barak, V. et al.(2009b) Mol. Breeding, 23, 179–195.
183 Singh, V., Pallaghy, C.K., and Singh, D.(2006) Field Crops Res., 96, 199–206.
184 Quisenberry, J.E., Roark, B., andMcMichael, B.L. (1982) Crop Sci., 22,918–922.
185 Singh, D.P., Chaudhary, B.D., Singh, P.,Sharma, H.C., and Karwasra, S.P.S.
1398j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs

(1990) Drought Tolerance in OilseedBrassicas and Chickpea, HaryanaAgriculture University, Hisar, India,pp. 1–60.
186 Leport, L., Turner, N.C., French, R.J. et al.(1999) Eur. J. Agron., 11, 279–291.
187 Turner, N.C., Abbo, S., Berger, J.D. et al.(2007) J. Exp. Bot., 58, 187–194.
188 Kuk, Y.I., Shin, J.S., Burgos, N.R. et al.(2003) Crop Sci., 43, 2109–2117.
189 Netondo, G.W., Onyango, J.C., and Beck.,E. (2004) Crop Sci., 44, 806–811.
190 Belkhodja, R.,Morales, F., Abad�ıa,A. et al.(1999) Photosynthetica, 36, 375–387.
191 Jimenez, M.S., Gonzalez-Rodriguez,A.M., Morales, D. et al. (1997)Photosynthetica, 33, 291–301.
192 Burke, J.J. (2007) Plant Physiol., 143,108–121.
193 Wilson, R.F., Burke, J.J., andQuisenberry, J.E. (1987)Plant Physiol., 84,251–254.
194 Pettigrew, W.T. (2004a) Photosynthetica,42, 567–571.
195 Longenberger, P.S.,PhD thesis,submitted to the Texas A&M University,USA. (2008)
196 Tuberosa, R. and Salvi, S. (2006) TrendsPlant Sci., 11, 405–412.
197 Xiao, J., Wu, K., Fang, D.D. et al. (2009) J.Cotton Sci., 13, 75–157.
198 Rong, J.K., Abbey, C., Bowers, J.E. et al.(2004) Genetics, 166, 389–417.
199 Lacape, J.M., Nguyen, T.B., Thibivilliers,S. et al. (2003) Genome, 46, 612–626.
200 Yu, J.W., Yu, S.X., Lu, C.R. et al. (2007) J.Integr. Plant Biol., 49, 716–724.
201 Lin, L., Pierce, G.J., Bowers, J.E. et al.(2010) BMC Genomics, 11, 395–411.
202 Chen, Z.J., Scheffler, B.E., andDennis, E.(2007) Plant Physiol., 145, 1303–1310.
203 Helentjaris, T., Slocum,M.,Wright, S. et al.(1986) Theor. Appl. Genet., 72, 761–769.
204 Vos, P., Hogers, R., Blecker, M. et al.(1995) Nucl. Acids Res., 23, 4407–4414.
205 Ulloa, M., Saha, S., Jenkins, J.N. et al.(2005) J. Hered., 96, 132–144.
206 Paterson, A., Estill, J., Rong, J. et al. (2002)Cotton Sci., 14, 31.
207 Rahman, M., Hussain, D., and Zafar, Y.(2002) Crop Sci., 42, 2137–2144.
208 Rahman,M., Yasmin, T., Tabassum,N. et al.(2008) Genet. Res. Crop Evo., 55, 331–339.
209 Reinisch, A.J., Dong, J.M., Brubaker, C.L.et al. (1994) Genetics, 138, 829–847.
210 Ulloa,M.,Meredith,W.R., Shappley, Z.W.et al. (2002) Theor. Appl. Genet., 104, 200–208.
211 Zhang, J., Guo, W., and Zhang, T. (2002)Theor. Appl. Genet., 105 (8), 1166–1174.
212 Mei, M., Syed, N.H., Gao, W. et al. (2004)Theor. Appl. Genet., 108, 280–291.
213 Shappley, Z.W., Jenkins, J.N., Zhu, J. et al.(1998) J. Cotton Sci., 4, 153–163.
214 Ulloa, M. and Meredith, W.R. (2000) J.Cotton Sci., 4, 161–170.
215 Jiang,C.X.,Wright, R.J., El-Zik, K.M. et al.(1998) Proc. Natl. Acad. Sci. USA, 95,4419–4424.
216 Kohel, R.J., Yu, J., Park, Y.H. et al. (2001)Euphytica, 121, 163–172.
217 Zhang, T.Z., Yuan, Y.L., Yu, J. et al. (2003)Theor. Appl. Genet., 106, 262–268.
218 Lin, Z., He, D., Zhang, X. et al. (2005)Plant Breed., 124, 180–187.
219 Zhang, L., Cao, W., Zhang, S. et al. (2005)Acta Phytoecol. Sci., 29, 266–273 (inChinese with an English abstract).
220 Shen, X., Guo, W., Zhu, X. et al. (2005)Mol. Breed., 15, 169–181.
221 Shen, X., Guo, W., Lu, Q. et al. (2007)Euphytica, 155, 371–380.
222 Park, Y.H., Alabady, M.S., Ulloa, M. et al.(2005) Mol. Genet. Genomics, 274,428–441.
223 Lacape, J.M., Nguyen, T.B., Courtois, B.et al. (2005) Crop. Sci., 45, 123–140.
224 Paterson, A.H., Saranga, Y., Menz, M.et al. (2003) Theor. Appl. Genet., 106,384–396.
225 Jiang, C., DelMonte, T.A., Paterson, A.H.et al. (2000)Theor. Appl.Genet., 100, 409–418.
226 Wright, R.J., Thaxton, P.M., El-Zik, K.H.,and Paterson, A.H. (1999) J. Hered., 90,215–219.
227 Song, X.L., Wang, K., Guo, W.Z. et al.(2005) Acta Bot. Sin., 47 (11), 1382–1390.
228 Ulloa, M., Zeiger, E., Lu, Z. et al. (2000) J.Cotton Sci., 4, 10–18.
229 Saranga, Y., Menz, M., Jiang, C.X. et al.(2001) Genome Res., 11, 1988–1995.
230 Saranga, Y., Jiang, C.X., Wright, R.J. et al.(2004) Plant Cell Environ., 27, 263–277.
231 Shen, X., Van Becelacre, G., Kumar, P.et al. (2006) Theor. Appl. Genet., 13 (8),1539–1549.
References j 1399

232 Chakraborty, R. and Weiss, K.M. (1988)Proc. Natl. Acad. Sci. USA, 85, 9119–9123.
233 Kruglyak, L. (1999) Nat. Gene., 22,139–144.
234 Abdurakhmonov, I.Y., Kohel, R.J., Yu, J.Z.et al. (2008) Genomics, 92, 478–487.
235 Abdurakhmonov, I.Y., Saha, S., Jenkins,J.N. et al. (2009) Genetica, 136, 401–417.
236 Kantartzi, S.K. and Stewart, J.M. (2008)Plant Breed., 127, 173–179.
237 Shinozaki, K. and Yamazuchi-Shinozaki,K. (1996)Curr. Opin. Biotech., 7, 161–167.
238 Bartels, D. and Sunkar, R. (2005)Crit. Rev.Plant Sci., 24, 23–58.
239 Payton, P., Allen, R.D., Trolinder, N. et al.(1997) Photosyn. Res., 52, 233–244.
240 Payton, P., Webb, R.P., Kornyeyev, D.et al. (2001) J. Exp. Bot., 52, 2345–2354.
241 Light, G.G., Mahan, J.R., Roxas, V.P. et al.(2005) Planta, 222, 346–354.
242 He, C., Yan, J., Shen, G. et al. (2005) PlantCell Physiol., 46, 1848–1854.
243 Lv, S., Zhang, K., Gao, Q. et al. (2008)Plant Cell Physiol., 49 (8), 1150–1164.
244 Lv, S.L., Lian, L.J., Tao, P.L. et al. (2009)Planta, 229, 899–910.
245 Zhang, H., Dong, H., Li, W. et al. (2009)Mol. Breed., 23, 289–298.
246 Allen, R.D. (2010) Opportunities forengineering abiotic stress tolerance incotton plants, in Cotton, Biotechnology inAgriculture andForestry 65 (ed.U.B.Zehr),Springer, Berlin, pp. 127–160.
247 Yan, J.Q., He, C.X., Wang, J. et al. (2004)Plant Cell Physiol., 45, 1007–1014.
248 Zhang, W.X., Nie, Y., and Luo, X. (2005)Plant Breed., 124, 142–146.
249 Keshamma, E., Rohini, S., Rao, K.S.et al. (2008) J. Cotton Sci., 12,264–272.
1400j 53 Cotton: Genetic Improvement for Drought Stress Tolerance – Current Status and Research Needs





























