Embed Size (px)
Citation preview

i
Improving the nutritional value of soybean meal through fermentation using
newly isolated bacteria
By
Samantha Medeiros
A Thesis presented to
The University of Guelph
In partial fulfillment of requirements for the degree of
Master of Science In
Animal & Poultry Science
Guelph, Ontario, Canada ©Samantha Medeiros, March, 2015

ii
ABSTRACT
IMPROVING THE NUTRITIONAL VALUE OF SOYBEAN MEAL THROUGH FERMENTATION USING NEWLY ISOLATED BACTERIA
Samantha Medeiros Advisor: University of Guelph, 2015 Dr. Julang Li
Soybean meal has limited use in piglet feed due to anti-nutritional factors. Past studies show that
using fermented soybean meal can reduce these factors and improve pig growth performance. In
this thesis research is reported aimed at using newly isolated strains of bacteria to improve the
feeding value of soybean meal through fermentation. Bacteria were isolated from fermented
foods and the intestines of grass carp (Ctenpharyngodon idella), screened for enzyme activity,
characterized, and then used to ferment soybean meal. The fermented product was analyzed for
protein profile, as well as nutrient composition and allergenicity. Results indicate that the
bacteria identified as Bacillus amyloliquefaciens decreased large proteins, eliminated allergenic
proteins, reduced the oligosaccharide concentration and increased the concentrations of crude
protein and amino acids in partly fermented soybean meal. Further studies are needed to
investigate the non-starch polysaccharide concentration and the growth performance of piglets
fed the fermented soybean meal product.

iii
ACKNOWLEDGEMENTS
First and foremost I would like to thank my advisor Dr. Julang Li. Throughout this
journey she has been more than an advisor to me and provided me with guidance and
encouragement through the roughest of times. She is a brilliant scientist and it was a privilege to
learn under her supervision.
I would also like to thank my advisory committee members, Dr. Kees de Lange and Dr.
Hugh Cai, for all of their advice and insightful comments to this project. I especially would like
to thank Dr. Cai for all of his assistance in the identification of the isolates using MALDI-TOF
MS. I would also like to thank our collaborator Dr. Zhang and Jingjing Xie for their contribution
of the crude protein, amino acid, and oligosaccharide analyses.
I would also like to acknowledge all of my lab members, especially Dr. Paul Dyce who
helped complete the gyrB gene sequencing and the western blot analysis. All of my friends in
the department have provided moral support and scientific advice. I was very fortunate to work
with such an intelligent and amiable group of people which made my laboratory experience a
very pleasant one.
I would also like to thank Ontario Ministry of Agriculture Food and Rural Areas for
awarding me their Highly Qualified Personnel Scholarship.
Lastly, I would like to thank my family and my friends for all of their support through
these last two years. Being surrounded by such a loving group of people has always made me
feel blessed and I could not have come as far as I have without them.

iv
TABLE OF CONTENTS ABSTRACT .................................................................................................................................... ii
ACKNOWLEDGEMENTS ........................................................................................................... iii
LIST OF FIGURES ...................................................................................................................... vii
LIST OF TABLES ....................................................................................................................... viii
LIST OF ABBREVIATIONS.........................................................................................................ix
1.0 INTRODUCTION: LITERATURE REVIEW ......................................................................... 1
1.1 Challenges faced by piglets .................................................................................................. 1
1.1.1 Physiological factors ...................................................................................................... 1
1.1.2 Social factors .................................................................................................................. 3
1.1.3 Diet composition ............................................................................................................ 3
1.2 Soybeans ............................................................................................................................... 4
1.2.1 Positive factors of soybeans ........................................................................................... 4
1.2.2 Negative factors of soybeans ......................................................................................... 7
1.3 Current applications used to improve soybean ................................................................... 19
1.3.1 Processing .................................................................................................................... 19
1.3.2 Biotechnology and selective breeding strategies ......................................................... 20
1.3.3 Addition of enzymes .................................................................................................... 21
1.3.4 Microbial fermentation ................................................................................................ 22
1.4 Summary and rationale ....................................................................................................... 26

v
2.0 HYPOTHESES ....................................................................................................................... 27
3.0 OBJECTIVES ......................................................................................................................... 27
4.0 MATERIALS AND METHODS ............................................................................................ 28
4.1 Isolating and screening bacteria with high enzyme activity ............................................... 28
4.1.1 Primary screening for bacteria with protease activity from fermented food ............... 28
4.1.2 Primary screening for cellulase activity from grass carp source ................................. 29
4.1.3 Second round of screening ........................................................................................... 31
4.2. Identification of bacteria selected ...................................................................................... 33
4.2.1: MALDI-TOF MS:....................................................................................................... 33
4.2.2: PCR Amplification of partial 16S rRNA gene and gyrB gene: .................................. 33
4.3. Soybean meal fermentation................................................................................................ 36
4.3.1. Preparation of soybean meal and inoculums .............................................................. 36
4.3.2. Liquid-state fermentation ............................................................................................ 37
4.3.3. Solid-state fermentation .............................................................................................. 38
4.4 Analysis of soybean meal fermentation products ............................................................... 39
4.4.1 Proximate analysis and amino acid profiling ............................................................... 39
4.4.2 Determination of soluble protein fractions and distribution ........................................ 39
4.4.3 Determination of oligosaccharide concentrations ........................................................ 40
4.4.4 Detection of Allergenic Proteins. ................................................................................. 41
4.5 Statistical analysis: .............................................................................................................. 43

vi
5.0 RESULTS ............................................................................................................................... 43
5.1 Qualitative measurement of enzymes activity. ................................................................... 43
5.1.1 Primary Screening for Protease Producing Bacteria .................................................... 43
5.1.2 Primary Screening of Cellulase Producing Bacteria .................................................... 44
5.1.3 Second round of screening ........................................................................................... 44
5.2 Identification of selected bacteria ....................................................................................... 51
5.2.1 Characterization of isolates using MALDI-TOF MS .................................................. 51
5.2.2 Characterization of isolates using 16S rRNA and gyrB gene sequencing ................... 51
5.3 Liquid-state fermentation .................................................................................................... 55
5.3.1 Influence of selected bacterium on SBM protein profile ............................................. 55
5.4 Solid-state fermentation ...................................................................................................... 57
5.4.1 Influence of selected bacterium on SBM protein profile ............................................. 57
5.4.2 Improved crude protein of fermented soybean meal ................................................... 58
5.4.3 Amino acid profile of the fermented soybean meal ..................................................... 62
5.4.4 Detection of Allergenic Proteins .................................................................................. 65
5.4.5 Oligosaccharide concentration of fermented soybean meal ........................................ 65
6.0 DISCUSSION ......................................................................................................................... 68
REFERENCES: ............................................................................................................................ 82
Appendix A ................................................................................................................................... 96

vii
LIST OF FIGURES
Figure 1: Simplified figure illustrating the degradation of soy oligosaccharides ........................ 17
Figure 2: A simplified diagram illustrating the degradation of cellulose .................................... 18
Figure 3: Measuring protease activity using soymilk agar plates ............................................... 47
Figure 4: Measuring cellulase activity using CMC agar .............................................................. 48
Figure 5: Measuring amylase activity using corn starch agar ...................................................... 49
Figure 6: Gel electrophoresis of PCR products used to identify the bacteria isolates. ................ 53
Figure 7 : The fermentation effects of using different isolates on the protein profile of SBM in a
liquid-state..................................................................................................................................... 56
Figure 8: The relative changes of total soluble protein of soybean meal when fermented for
different lengths and with different bacterial isolates.. ................................................................ 59
Figure 9: Visualizing the changes of total soluble protein profile of soybean meal when
fermented for different lengths and with different bacterial isolates with Coomassie Blue gel ... 60
Figure 10: The effects of solid-state fermentation length on crude protein of soybean meal ...... 61
Figure 11: The effect of fermentation length using Isolate-2 on amino acid concentrations ...... 63
Figure 12: Western blot detecting soy allergens in the unfermented and fermented soybean meal
products ......................................................................................................................................... 66

viii
LIST OF TABLES
Table 1: Summary of the undesirable nutrient components of soybean meal……..………......10
Table 2: Summary of the first round of screening……………………………………………....45
Table 3: Summary of the enzyme activity of best performing bacteria isolated from food and
grass carp intestine…………………...…………………………………………………………..50
Table 4: Summary of the gene sequencing nucleotide BLAST strain identification of the
selected isolates..………………….………………………………………………..…………….54
Table 5: Quantitative representation of the effect of fermentation with Isolate-2 over a 24 and a
48 hour period……………………………………………………………………………………64
Table 6: The oligosaccharide concentration in soybean before and after fermentation with
Isolate-2 for 24 and 48 hours………………………………………………………………….....67

ix
LIST OF ABBREVIATIONS
AOAC Association of Analytical Communities
ATCC American Type Culture Collections
BHI Brain Heart Infusion bacteria culture media
BLAST Basic local alignment search tool
CFU Colony forming units
CMC Carboxymethyl cellulose
DM Dry matter
EAA Essential amino acids
GIT Gastrointestinal tract
gyrB DNA gyrase β subunit
HPLC High-performance liquid chromatography
LAB Lactic acid bacteria
LB Luria Bertani bacteria culture media
MALDI-TOF MS Matrix assisted laser desorption ionization- time of flight mass spectrometry
MRS deMan, Rogosa and Sharpe bacteria culture media
NCBI National
NEAA Non-essential amino acids
NRC National Research Council
NSP Non-starch polysaccharide
P.con Positive control
PCR Polymerase chain reaction
PGA Poly-ɣ-glutamate

x
PVDF polyvinyl difluoride
RFO Raffinose family of oligosaccharides
SB Sodium borate
SBM Soybean meal
SDS-PAGE sodium –dodecyl sulfate-polyacrylamide gel electrophoresis
SPC Soy protein concentrate
SPI Soy protein isolate
16S rRNA 16S ribosomal ribonucleic acid

1
1.0 INTRODUCTION: LITERATURE REVIEW
1.1 Challenges faced by piglets
Weaning is the process of separating piglets from the sow’s milk. In nature, piglets are
weaned gradually at the age of 10-12 weeks (Lalles et al., 2007); however, in industrial settings
piglets are weaned abruptly and much earlier between 21-26 days of age. At this time, the piglet
is still undergoing rapid intestinal development. Early weaning is typically associated with a
decrease in piglet performance due to decreased feed intake brought on by three main factors:
physiological factors, social factors, and change in diet composition. These challenges caused by
early weaning decrease as the pig ages, however the losses due to piglet performance during the
weaning period remain a major loss to the swine industry.
1.1.1 Physiological factors
When a piglet is first born it consumes colostrum from the sow, a liquid feed. It takes
approximately 10-12 weeks post-partum for the piglet’s digestive tract to be able to switch from
a liquid feed to a solid feed (Lalles et al., 2007). This abrupt transition when weaning piglets
early negatively impacts the piglet because its gastrointestinal tract is still immature and cannot
adequately handle solid food yet. There are two intestinal morphological characteristics that
have been associated with negative aspects of early weaning: villus atrophy and crypt
hyperplasia (Pluske et al., 1997). Villus atrophy observed during early weaning is likely due to
an increased rate of cell loss and consequently, an increased crypt-cell production leading to an
increased crypt death (Pluske et al., 1997). Consequently, the decreased villi height also has a
negative impact on the brush-border enzymes lactase and maltase (Pluske et al., 1997). These

2
changes in gut morphology led to malabsorption of nutrients which increases incidences of
diarrhea (Pluske et al., 1997). Early weaning has also been found to effect pancreatic enzyme
secretions. The activity of important enzymes responsible for protein digestion, such as: trypsin,
chymotrypsin, carboxypeptidase A, and carboxypeptidase B, were found to decrease after early
weaning (Hedemann & Jensen, 2004). Without these enzymes, piglets cannot efficiently digest
found in soybean meal or successfully hydrolyse the allergenic proteins. The gastric pH of the
piglet during weaning also contributes to this decrease in performance. Piglets have a decreased
capacity to secrete hydrochloric acid which leads to the gastric pH of 3-4, which is high when
compared to that of a mature pig (pH of 1.6-1.7) (Cranwell et al., 1976; Snoeck et al., 2004).
Gastric pH levels that are high negatively affects piglet performance by resulting in an
incomplete digestion of protein as well as allowing pathogenic bacteria to survive and colonize
the gut (Snoeck et al.,2004). When pathogenic bacteria, such as E.coli, colonize the gut, it leads
to the piglet developing the clinical sign of scours.
Apart from of the immature gastrointestinal tract, the piglet also has an immature immune
system which contributes to the decrease in piglet performance observed with early weaning.
Piglets acquire immunity when they are first born through the sow’s colostrum but this passive
immunity only lasts for a short time (Lalles et al., 2007). At the age of weaning, the piglet has
built up a mucosal immune system which is developed enough to activate the immune response
(Lalles et al., 2007). The challenges faced when piglets are fed soybean meal result in damage to
the intestine resulting in further malabsorption (Li et al., 1991). During weaning the immune
system is still developing which causes the piglet to be more susceptible to bacterial infections

3
(Kelly and King, 2001). Combined, all of these developmental factors affect piglet performance
associated with early weaning.
1.1.2 Social factors
A piglet during weaning also faces a number of non-physiological factors that may be
contributing to the negative piglet performance observed during early weaning. During weaning
the piglet faces a lot of social challenges such as separation from the sow, mixing litters, and
social dominance. When a piglet is separated from its sow, it exhibits separations calls which
have been thought to express a need for the sow (Held & Mendl, 2001). It was also observed
that the earlier the separation, the more separation calls were exhibited which indicated
separation stress (Weary et al., 1999). When piglets from different litters are mixed into one
pen, an elevation of cortisol levels is observed (Merlot et al., 2004). These elevated levels of
cortisol did not exhibit long-term effects. When piglets are introduced into a new pen a social
hierarchy is established. It was observed that dominant piglets showed lower cortisol levels than
submissive piglets (Merlot et al., 2004). It was also observed that dominant piglets had a higher
weight gain than submissive piglets (Held & Mendl, 2001). All of these factors combined can
contribute to the lower feed intake observed during weaning which contributes to a poor growth
performance.
1.1.3 Diet composition
Transitioning from sow’s milk which is highly digestible and highly palatable (Klobasa et
al., 1987), to a solid-feed which is less palatable presents another challenge. Due to all the anti-
nutritional factors associated with plant-based proteins, highly digestible and palatable animal-

4
based proteins are commonly used in piglet diets. These include ingredients like: dried whey,
spray dried plasma, poultry by-product meal, and fish meal. In recent times, there has been a
move away from using animal-based protein mostly due to the ban in Europe prohibiting any
animal-based proteins in animal feed. The rising cost of fish meal is also another reason why
researchers are trying to find an alternative protein source for agricultural animals.
1.2 Soybeans
Soybean (Glycine max (L.) Merril) is a legume that is widely used for both extraction of
oil and animal feed. Canada produces 4.25 million tons of soybean/year and approximately 90%
of that is processed into soybean meal in solvent extraction plants in Ontario (Canadian Institute
Soy 20/20, 2008). More than half of the soybean meal produced in Canada goes to poultry and
about a quarter goes to swine operations (Canadian Institute Soy 20/20, 2008). Soybeans carry
many advantages such as: high protein level, excellent digestible amino acid profile, as well as
their antioxidant properties. Unfortunately, there are also many negative properties of soybeans
such as allergenic proteins and other anti-nutritional factors which can limit their use, especially
in young animal diets. Soybeans can be processed into several different products such as
soybean meal and soybean protein concentrates which eliminates some, but not all, of the anti-
nutritional factors, allowing them to be used in animal feeds.
1.2.1 Positive factors of soybeans
There are several factors that contribute to soybean’s positive nutritional properties.
Soybeans are considered to be a high protein source with approximately 40% protein as dry
matter (DM) (Saz & Marina, 2007). Soybeans processed into soybean meal can have crude

5
protein values ranging from 44- 48% depending on if hulls are reintroduced into the meal.
Soybeans contain an excellent profile of amino acids, providing most of the essential amino
acids (Saz & Marina, 2007). Soybeans are particularly high in lysine and tryptophan, which are
the amino acids low in cereal grains (Stein et al., 2008). The digestibility of amino acids can
differ depending on heat processing of the soybean flakes. Soybean meal which has been
autoclaved at 125 °C contained lower concentrations and reduced digestibility of the amino
acids: arginine, lysine, and cysteine (González-Vega et al., 2011). This reduction in digestibility
of amino acids was attributed to the Maillard reactions and melanoidins which occur when
heating products (González-Vega et al., 2011). Maillard browning reactions decreases the
digestibility of amino acids by irreversibly converting available lysine to unavailable lysine
complexes (Friedman & Brandon, 2001; González-Vega et al., 2011). Soybeans are also high in
arginine which is a great benefit to poultry as they cannot synthesize arginine due to the inactive
urea cycle (Stein et al., 2008). However, soybeans are not the perfect protein source as they are
deficient in methionine, the first limiting amino acid for poultry. The amino acids found in
soybeans are more digestible than amino acids found in other oilseed products (Stein et al.,
2008). The amino acid digestibility and metabolizable energy values can differ depending on if
the soybean meal was expelled or solvent extracted. Soybean meal which has been extruded-
expelled, rather than extracted, was found to have a greater amino acid digestibility in pigs
(Baker & Stein, 2009). This is likely due to the higher percentage of soybean oil in the extruded-
expelled soybean meal. It was also observed that soybean meal had 11-25% higher
metabolizable energy content when compared to canola meal, dehulled sunflower meal,
cottonseed meal, and peanut meal (Stein et al., 2008). Despite some of the limitations stated
above, soybean meal in general is still a great protein source for livestock and poultry feeds.

6
Soybeans have been termed as a functional food. Asian populations with high soy
consumption, have been known to have decreased risk of certain cancers (Friedman & Brandon,
2001). Some of the functional properties of the soybean has been attributed to its high levels of
specific phenols, which are secondary metabolites produced by plants under normal development
or under stressful conditions (Stalikas, 2010). Flavanoids belong to polyphenols and can be
classified as anthocyanins, flavanols, flavones, flavonoes, and flavanols (Stalikas, 2010).
Flavonols are more commonly found in vegetables and fruits, whereas isoflavones are found in
legumes (Stalikas, 2010). Isoflavones, which are isomeric to flavonoids, have been of interest to
the research community. There are twelve types of isoflavones: three free aglycones genistein,
daidzein, and glycitein, and their three glucosidic, three acteyl glucosidic, and 3 malonyl
glucosidic conjugates (Friedman & Brandon, 2001). Isoflavones, genistein, daidzein, glycitein,
biochanin A, and formononectin, are the phytoestrogen molecules found in soybean which have
had a particular interest in human health research. These phytoestrogen molecules are able to
bind to the estrogen receptor and can act as natural selective estrogen receptor modulators (Fritz
et al., 2013). Depending on local estrogen levels, these isoflavones have been found to play
either agonistic or antagonistic on the estrogen receptor (Fritz et al., 2013). Isoflavones and their
role in certain cancers, especially breast and colon cancers, have been controversial. Some in
vitro models suggest that genistein can directly or indirectly interact with the endoplasmic
reticulum and promote tumor cell growth in breast tissue (Van Duursen et al., 2011).
Conversely, other studies have reported genestein to have antitumor and anti-angiogenic activity
(Farina et al., 2006). Other effects of soy on human health include inhibition of vascular
endothelial growth factor and inhibiting tyrosine kinase, and inducing tumor suppression proteins
(Fritz et al., 2013).

7
The role of isoflavones in manipulating animal performance has also been investigated.
The concentration of isoflavones in soy can be altered through processing however; soybean
meal still contains isoflavones (Kuhn et al., 2004). Because of its estrogen-like properties, soy
isoflavones and their effect on carcass characteristic have been studied. The effects of soy
isoflavones have been contradictory. Some studies have observed no effect on growth
performance, carcass composition, or meat quality of pigs (Kuhn et al., 2004). Other studies
indicate isoflavones produce leaner meat in barrows if fed at the level found in conventional
soybean meal (Payne et al., 2001). The effects of isoflavones on gilts was also investigated
which resulted in no effect on carcass trait (Payne et al., 2001).
1.2.2 Negative factors of soybeans
Despite the apparent benefits of eating soybeans, there are also many negative factors that
can impact animal production. Unfortunately, these anti-nutritional factors are part of the
legume and are often hard to eliminate. The first anti-nutritional factor to be discussed is the
large proteins which may be allergenic to humans and animals. The seed of the soybean is
important for storage of nitrogen, sulfur, and carbon (Wilson et al., 2005). Soy seed protein
consists of two main fractions, 11S globulin fraction and 7S globulin fraction, which together
account for 70 % to 80% of the total protein composition of soybean (Wilson et al., 2005). Soy
protein has been listed as one of the “big eight” foods with the most allergens (Wilson et al.,
2005). Two important allergens in soy are glycinin (an 11S globulin fraction) and β-conglycinin
(a 7S globulin fraction). Glycinin is a polypeptide which is composed of multiple acidic (35-40
kDa) (Beardslee et al., 2000) and basic (22 kDa) (Helm et al., 2000) subunits held together by
disulfide bonds. There is approximately 40-70 mg of glycinin /g in soybean meal, as indicated in
Table 1 (American Soybean Association, 2004). Glycinin has been well studied in farm animals

8
and has been known to decrease growth performance in piglets (Sun et al., 2008a; Zhao et al.,
2008; Zhao et al., 2010; Li et al., 1990). This decrease in performance can be attributed to the
damage to the intestinal morphology, disorder in the immune system, and increased diarrhea
(Sun et al., 2008a; Li et al., 1990; Li et al., 1991; Zhao et al., 2008). Glycinin is a small peptide
resistant to heat and most enzymatic digestion. Glycinin is partially insoluble in a piglet’s
stomach (Zhao et al., 2008), which presents a further issue to the piglet as it escapes this initial
digestion step and can stimulate an immune response when it reaches the small intestine. This
was confirmed by the increased levels of immunoglobulins of the mucosal layer of the small
intestine (IgA) and immunoglobulins in the serum (IgG) when piglets were fed glycinin (Sun et
al., 2008b; Hao et al., 2009). Piglets fed glycinin had increased intestinal mast cells numbers,
leading to histamine released (Sun et al., 2008a). Histamine has been known to play a role in
increasing intestinal motility and secretion of water which may explain the decrease performance
results and increased diarrhea in piglets fed glycinin (Liu et al., 2000; Sun et al., 2008a). It has
also been reported that piglets fed increased levels of glycinin experienced villus atrophy and
crypt hypertrophy in the jejenum which also indicates intestinal damage (Sun et al., 2008a). The
polypeptide glycinin is composed of two subunits; an acidic and a basic subunit. The properties
of the different subunits allow glycinin to escape digestion in the duodenum of the small
intestine; however evidence suggest that the number of immunoreactive glycinin proteins
decreases in the middle of the jejunum (Zhao et al., 2008). However, the digestion of glycinin is
inefficient due to its disulfide bonds and hydrophobicity of its subunits (Zhao et al., 2008). The
disulfide bonds provide strength and support to the molecular structure. Glycinin’s basic
subunits are hydrophobic, and thus can aggregate, hiding cleavage sites that enzymes need to
bind in order for the peptide to be digested (Kuipers et al., 2007).

9
β-conglycinin has also been of great interest. Like glycinin, β-conglycinin is a storage
protein consisting of three different subunits: α (72 kDa), α’ (76 kDa) and β (53 kDa) (Doyle et
al., 1986). Soybean meal contains approximately 10-40 mg/g of β-conglycinin (American
Soybean Association, 2004; Table 1). Similar to glycinin, β-conglycinin has been known to
negatively affect piglet growth performance (Zhao et al., 2010). This decrease in growth
performance is likely due to the allergenic effects, direct effect on intestinal cells, and its
molecular structure. β-conglycinin can illicit an immune response, increasing IgE levels in the
serum and stimulating various cytokines to be released, destroying the integrity of the intestine
(Hao et al., 2009). Apart from its allergenic effects, β-conglycinin has also been found to
directly affect the small intestine by altering the cytokine expression, function, and replication in
piglet intestinal cells (Chen et al., 2011). Subunits of β-conglycinin can aggregate in the small
intestine, hiding enzymatic cleavage sites, leading to its ability to escape digestion (Kuipers et
al., 2007). Although both allergenic proteins are mostly resistant to enzymatic digestion, β-
conglycinin is found in higher concentrations than glycinin in the lower parts of the small
intestine, signifying that β-conglycinin is more resistant to digestion than glycinin (Zhao et al.,
2008). Fortunately, piglets can become desensitized to both glycinin and β-conglycinin as they
get older (Wang et al., 2010).
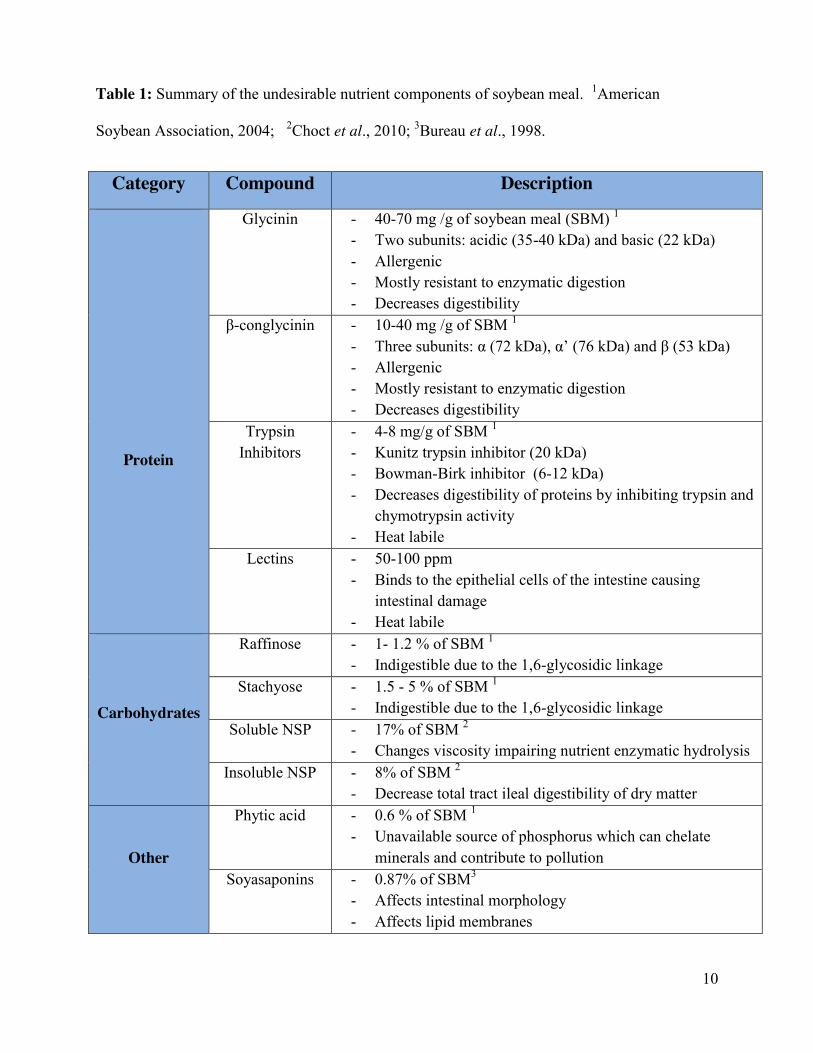
10
Table 1: Summary of the undesirable nutrient components of soybean meal. 1American
Soybean Association, 2004; 2Choct et al., 2010; 3Bureau et al., 1998.
Category Compound Description
Protein
Glycinin - 40-70 mg /g of soybean meal (SBM) 1 - Two subunits: acidic (35-40 kDa) and basic (22 kDa) - Allergenic - Mostly resistant to enzymatic digestion - Decreases digestibility
β-conglycinin - 10-40 mg /g of SBM 1 - Three subunits: α (72 kDa), α’ (76 kDa) and β (53 kDa) - Allergenic - Mostly resistant to enzymatic digestion - Decreases digestibility
Trypsin Inhibitors
- 4-8 mg/g of SBM 1 - Kunitz trypsin inhibitor (20 kDa) - Bowman-Birk inhibitor (6-12 kDa) - Decreases digestibility of proteins by inhibiting trypsin and
chymotrypsin activity - Heat labile
Lectins - 50-100 ppm - Binds to the epithelial cells of the intestine causing
intestinal damage - Heat labile
Carbohydrates
Raffinose - 1- 1.2 % of SBM 1 - Indigestible due to the 1,6-glycosidic linkage
Stachyose - 1.5 - 5 % of SBM 1 - Indigestible due to the 1,6-glycosidic linkage
Soluble NSP - 17% of SBM 2 - Changes viscosity impairing nutrient enzymatic hydrolysis
Insoluble NSP - 8% of SBM 2 - Decrease total tract ileal digestibility of dry matter
Other
Phytic acid - 0.6 % of SBM 1 - Unavailable source of phosphorus which can chelate
minerals and contribute to pollution Soyasaponins - 0.87% of SBM3
- Affects intestinal morphology - Affects lipid membranes

11
Apart from its allergenic aspect, soybeans also contain a high amount of other anti-
nutritional factors. These anti-nutritional factors are trypsin inhibitors, lectins, and phytic acid.
Trypsin and chymotrypsin are two of the key enzymes responsible for protein degradation in
mammals and birds. They belong to the family of serine proteases and is produced by the
pancreas and secreted in the duodenum. Soybeans, similar to other plants, use trypsin inhibitors
as a defence mechanism against pests (Jamal et al., 2013). There are two well-known trypsin
inhibitors in soybeans: the Kunitz inhibitor and the Bowman-Birk inhibitor. The Kunitz trypsin
inhibitor (20 kDa) inhibits only trypsin, whereas the Bowman-Birk inhibitor (6-12 kDa) can also
inhibit chymotrypsin (Birk, 1985). Soybeans have approximately 32-123 mg of trypsin inhibitor/
gram of protein while soybean meal only has 4-8 mg/gram (Anderson & Wolf, 1995; American
Soybean Association, 2004). Protease inhibitors bind with proteases which inactivates the
enzyme, reducing the animal’s ability to digest proteins. This is evident in rats fed raw soybean
which experienced pancreatic hypertrophy and a 60% growth depression (Rackis et al., 1979;
Struthers et al., 1983). Although pigs fed 40% raw soybean flour did not experience a
hypertrophic pancreas there was 29% reduction in trypsin activity and an 80% reduction in
chymotrypsin activity with an overall 84% growth depression (Struthers et al., 1983).
Fortunately, these trypsin inhibitors are heat labile proteins and can be mostly inactivated with
heat treatment (Anderson & Wolf, 1995).
Soybeans also contain glyco-proteins called hemagglutinins, commonly referred to as
lectins. Lectins are globulins that contain a carbohydrate-binding domain which allows it to bind
to red blood cells of rabbit but not cattle or sheep (Csaky & Fekete, 2004). Lectins are not
susceptible to proteolytic digestion in the duodenum which presents the opportunity for lectins to

12
bind to the epithelial cells of the small intestine (Csaky & Fekete, 2004). Soybeans contain 5.7
mg of lectin/g which is greatly reduced to 0.88 mg/g in soybean meal (Canadian National Grain
Institute, 2010). Feeding animals soybeans in which lectins have not been denatured has resulted
in decrease growth performance in pigs, poultry, and mice (Palacios et al., 2004; Zhang et al.,
1993; Friedman et al., 1991). In a study which compared the growth performance of chicks and
piglets fed modified soybeans, conventional soybeans, and conventional soybean meal, it was
observed that chicks and piglets fed lectin-free soybean meal had improved average daily gain
and feed conversion ratios compared to that of animals fed raw soybeans (Palacios et al., 2004).
There was also an additive benefit effect when both trypsin inhibitor and lectins were removed
from the feed (Palacios et al., 2004). This result may be attributed to lectin’s ability to bind to
intestinal mucosa layer which results in an increased secretion of proteins, increasing
endogenous nitrogen losses (Palacios et al., 2004). It was also observed that binding to the brush
boarder of the intestinal epithelial cells can affect cell viability, the crypt, and the weight of the
tissue (Liener, 1994). Lectins do not present a problem if the soybeans are heat treated as in the
case of soybean meal. It was reported, however, that lectins can be recovered in soybean meal,
which are able to still demonstrate epithelial binding ability, suggesting that a portion of lectins
are not destroyed with heat treatment (Maenz, Irish, & Classen, 1999). It is interesting to note
that not all types of lectins affect piglet performance in a negative manner. Lectins from kidney
beans can actually improve the small intestine development of a piglet and could be potentially
used in improving piglet performance during early weaning (Thomosson et al., 2007).
Plants store phosphorus in a form called phytic acid. When phytic acid is bound to a
mineral in the seed it is referred to as phytate. Approximately 0.6% of soybean meal consists of

13
phosphorus bound as phytic acid as demonstrated in Table 1 (American Soybean Association,
2010). Phytase can convert phytic acid into an available source of phosphorus but non-ruminant
animals do not have phytase and thus require the addition of inorganic phosphorus to be
supplemented in diets. This inefficient use of phosphorus has led to issues regarding pollution to
the environment. In addition, phytic acid also has the ability to chelate divalent metal ions such
as zinc and calcium, decreasing its bioavailability in the gut, which in extreme cases could result
in parakeratosis and a depressed growth rate in pigs (Oberleas, Muhrer, & O'dell, 1962).
Carbohydrate concentration is variable by region but is generally around 35% of the
soybean (Karr-Lilienthal et al., 2005). These carbohydrates can be categorized as non-structural
and structural carbohydrates. Non-structural carbohydrates consists mainly of low molecular
weight sugars, polysaccharides (mainly starch), and oligosaccharides. Soy oligosaccharides
raffinose and stachyose have also been thought to have a negative impact when fed to animals.
Raffinose and stachyose are structurally very similar and belong to a family called raffinose
family of oligosaccharides (RFO) (Figure 1). Raffinose is a tri-saccharide made up of one
galactose, glucose, and fructose unit. Stachyose is a tetra-saccharide consisting of two galactose
units, one glucose unit, and one fructose unit. Together, raffinose and stachyose make up 5.5-6.2
% of soybean meal (American Soybean Association, 2004; Table 1). RFOs are considered to be
anti-nutritional because they are indigestible to non-ruminant animals due to the lack of α-
galactosidase enzyme required to digest the raffinose and stachyose. Feeding a non-ruminant
animal feed containing RFOs can result in gastrointestinal discomfort and may result growth
performance reductions. Piglets fed soy protein isolates, which is soybean meal processed to
remove oligosaccharides and other factors, were found to have improved growth rates when

14
compared to piglets fed conventional soybean meal (Sohn et al., 1994). Other studies have
reported that stachyose can negatively affect ileal digestibility of dry matter and energy in pigs
(Van Kempen et al., 2006). A decrease in piglet performance and an increase in diarrhea has
been observed when stachyose and raffinose levels were increased (Liying et al., 2003). The rate
of passage also increased when stachyose levels increased, reducing the amount of time the
lower gut flora has to metabolize the oligosaccharides and thus results in higher excretion in the
feces (Liying et al., 2003). On the contrary, other studies have found no effect of
oligosaccharides on the incidences of diarrhea with only an effect on apparent or true ileal dry
matter, nitrogen or amino acid digestibility to a very small degree (Smirick et al., 2002).
Soybeans, as well as other plants, contain steroid glycosides called saponins. There is a
class of saponins, called soyasaponin, are found in soybeans and may have an anti-nutritional
effect on fish (Bureau et al., 1998). Their anti-nutritional affect may be due to their ability to
alter intestine function or their effect on calcium-dependent potassium channels (Bureau et al.,
1998). Unlike other anti-nutritional factors, processing does not eliminate soyasapoinins, unless
it has been extracted with alcohol, such as in soy protein concentrate (Bureau et al., 1998). Fish
fed soybean meals have been found to suffer from enteritis (Sorensen et al., 2011). It was
observed that there are components in purified alcohol extracts of soybeans that can negatively
affect the growth performance of both chinook salmon and rainbow trout (Bureau et al., 1998).
One study showed that Atlantic salmon fed soybean molasses with different subfractions of
saponins were found to develop enteritis (Knudsen et al., 2007). The authors attributed the
results due to soyasponins alone or in combination with other soybean anti-nutritional factors
(Knudsen et al., 2007). Another study found no negative effects on nutrient digestibility or
growth performance of Atlantic salmon when fish meal was supplemented with soyasaponins but

15
did observe a decreased cholesteric effect (Sorensen et al., 2011). It was also found that
soysaponins can be hydrolyzed into free aglycones by the intestinal microflora in humans,
chicks, rats, and mice (Hu et al., 2004; Gu et al., 2002).
Structural carbohydrates consist of the insoluble and soluble non-starch polysaccharides
(NSP) which together can account for 25% of soybean meal (Choct et al., 2010). Of this 25%,
17% consists of the soluble NSP pectin (Choct et al., 2010). There are three types of pectin
polysaccharides commonly present in the soybean meal: rhamnogalacturonans, arabinogalacton,
and xylogalacturonon (Choct et al., 2010). Soluble NSP are easily fermentable which increases
digesta transit time by forming a gel-like matrix which delays gastric empting (Fabek et al.,
2014; Montagne et al., 2003). In non-ruminants there has been evidence suggesting that soluble
NSP can have an anti-nutritional effect and impact overall animal performance. In poultry, diets
containing high amounts of soluble NSP increased the viscosity of the digesta which decreased
the nutrient and enzyme interaction in the intestine, negatively affecting the digestibility of other
nutrients (Smits & Annison, 1996). The increase in viscosity, as well as the decrease in gastric
emptying, allow for other bacteria to repopulate the intestine, negatively disrupting the balance
of the intestine (Angkanaporn et al., 1994; Choct et al., 2010). Similar changes were observed in
the piglet but were decreased as the piglet aged (Choct et al., 2010). Increasing soluble NSP in
the diet of weaning piglets demonstrated an increase in E.coli proliferation and an increased risk
of developing swine dysentery (Choct et al., 2010). An explanation for these observations is that
the increase in digesta transit time allows the bacteria to proliferate (Kim et al., 2012).
Approximately 8% of soybean meal consists of the insoluble NSP cellulose (Choct et al.,
2010). Cellulose is a high-energy molecule consisting of multiple glucose subunits linked

16
together through a β-1,4 glycosidic linkages (Figure 2). The breakdown of cellulose is complex
and requires three different groups of enzymes: cellobiohydrolases, endo-β-1,4-glucanases, and a
β-glucosidase, collectively referred to as the cellulase enzyme complex (Perez et al., 2002).
Ruminant animals have microbes in their rumen that can digest cellulose; however to non-
ruminant animals cellulose is virtually indigestible until fermentation in the large intestine.
Insoluble NSP are indigestible and do not increase the viscosity and thus decrease digesta transit
time and have been found to increase faecal bulking emptying (Montagne et al., 2003). These
properties cause insoluble NSP to have both beneficial and anti-nutritional qualities, depending
on if above a minimal level. Microbes in the intestine can utilize insoluble NSP and produce
beneficial short chain fatty acids, such as butyrate (Molist et al., 2009). Increases in butyrate
concentration are likely due to the longer fermentation time and the more suitable substrate
(Molist et al., 2009). Butyrate has a trophic effect on the colonocytes which have been found to
promote both small and large intestine growth (Molist et al., 2009; Montagne et al., 2003).
Young animals utilize insoluble NSP less efficiently than older animals (Montagne et al., 2003)
and thus have an anti-nutritional effect on piglets. Feeding piglets diets with increasing
percentages of NSP found in soybean hulls was found to negatively affect the daily growth rate
and feed conversion compared to that of the control group (Freire et al., 2000).
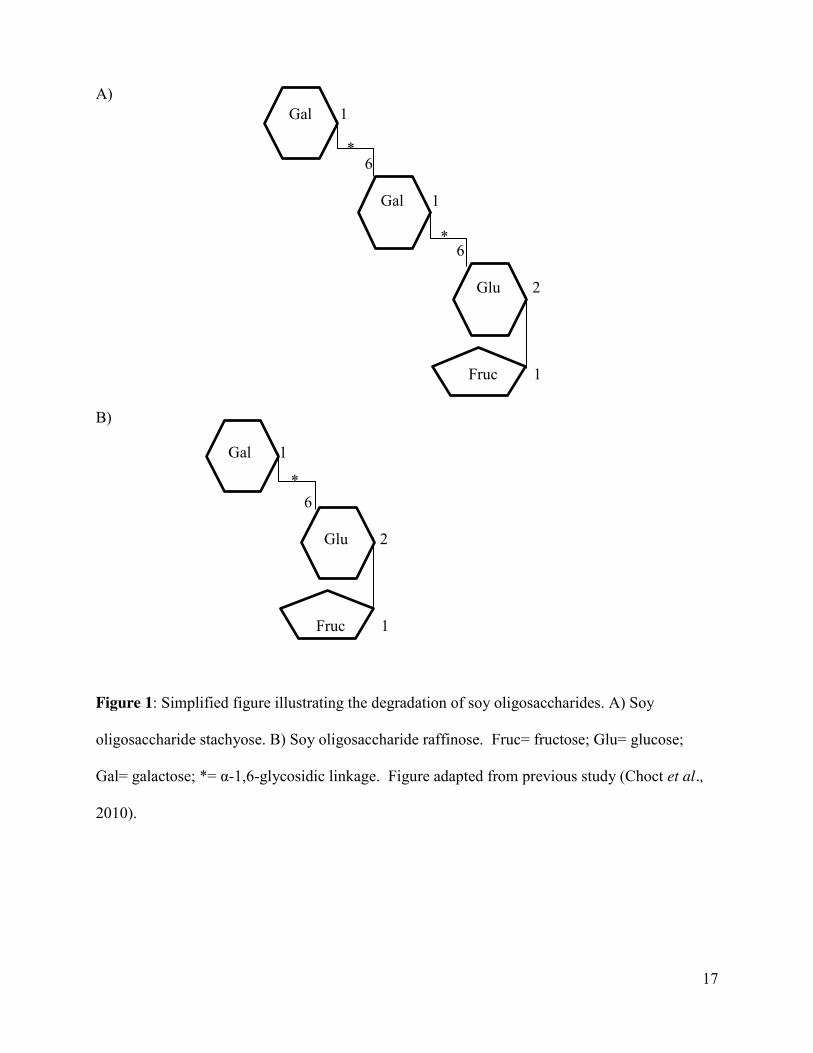
17
A)
B)
Figure 2
Figure 1: Simplified figure illustrating the degradation of soy oligosaccharides. A) Soy
oligosaccharide stachyose. B) Soy oligosaccharide raffinose. Fruc= fructose; Glu= glucose;
Gal= galactose; *= α-1,6-glycosidic linkage. Figure adapted from previous study (Choct et al.,
2010).
Gal 1
6
Gal 1
6
Glu 2
Fruc 1
Gal 1
6
Glu 2
Fruc 1
*
*
*

18
Figure 3
Figure 2: A simplified diagram illustrating the degradation of cellulose. Glu= glucose subunit;
n= number of glucose subunit repeats; 1= cleavage site for cellobiohydrolase; 2= cleavage site
for endo-β-1,4-glucanase; 3= cleavage site for β-glucosidase. Figure was adapted from Deacon,
2005.
Glu Glu 1 Glu 2 Glu Glu 2 Glu 2 Glu Glu 2
Cellulose
Glu 3 Glu Glu 3 Glu Glu 3 Glu
Cellobiose Cellobiose Cellobiose
Glu Glu Glu Glu Glu Glu
Multiple glucose molecules
n

19
1.3 Current applications used to improve soybean
As previously mentioned, there are various negative aspects of soybeans that put
restrictions on the quantity of soybean meal that can be used in piglet diets. Fortunately, some of
the anti-nutritional factors, such as trypsin inhibitors and lectins are heat-labile and can be
reduced during the processing of soybeans into soybean meal. However, some of the anti-
nutritional factors of soybeans are heat stable and are still present when young animals are fed
soybean meal. There are some current applications being used in the agricultural industry which
try to overcome these anti-nutritional factors such as: further processing, biotechnology,
breeding strategies, addition of enzymes, and microbial fermentation.
1.3.1 Processing
Processing soybean into soybean meal is one step which reduces the anti-nutritional
factors that are sensitive to heat damage. However, there are some negative factors that are not
completely eliminated even after the processing into soybean meal. Soybean white flakes can
still be further processed into soy protein concentrates (SPC) and soy protein isolates (SPI)
which has been found to improve the use of soy as a protein source for young animals. The
additional processing involved in creating SPC removes most of the oligosaccharides raffinose,
stachyose, and sucrose, effectively increasing the percentage of crude protein to as high as 67%
(Lenehan et al., 2007). There are three main processes that effectively remove the
oligosaccharides: extraction with aqueous ethanol, extraction with water at its isoelectric pH
(4.5) and denaturing the protein with moist heat before extraction with water (Lusas & Riaz,
1995). SPC has higher amino acid digestibility than soybean meal which resulted in an
improved pig growth performance compared to pigs fed regular soybean meal (Sohn et al.,

20
1994). It was also noted that different processing techniques, moist-extraction or dry-extraction,
can affect pig performance with moist-extracted SPC having a more positive effect than dry-
extraction (Friesen et al., 1993). Further processing of soybeans appears to be beneficial in
removing the anti-nutritional factors of soybean but additional heating must be properly
controlled as excess heat can result in amino acid damage which greatly reduces piglet
performance (Friesen et al., 1993). Additional processing may resolve the carbohydrate issue
with soybean meal; however, some commercially available SPI demonstrated proteins larger
than 37 kDa with less intense bands of the two subunits of β-conglycinin still being present
(Song et al., 2010). Although the additional processing of soybeans into SPI and SPC is
beneficial, the high cost of the product limits the use in animal agricultural industries (Opazo et
al., 2012).
1.3.2 Biotechnology and selective breeding strategies
Biotechnology can also be used to improve the anti-nutritional aspect of soybeans.
Biotechnology may involve the manipulation of the genome of an animal or plant for the purpose
of improving the organism for a particular industry. In Canada, soybeans have already been
modified to be resistant to herbicides to try to reduce costs to farmers, this is considered to be
first-generation traits (Canadian International Grain Institute, 2010). Manipulation of the
soybean genome to reduce anti-nutritional factors or to improve the nutrients for animal and
human consumption is considered to be second-generation traits (Canadian International Grain
Institute, 2010). It has been observed that raffinose and stachyose metabolism is not needed for
seed germination (Dierkin & Bilyeu, 2009) and Kerr and Sebastian (2000) have developed a
strain of soybean that contains high sucrose but a low oligosaccharide content. Similarly, there

21
have been mutations introduced into the phytic acid gene which results in a low phytic acid
soybean (Yuan et al., 2007). Biotechnology is a very appealing solution to some of the anti-
nutritional factors of soybean meal; however, it is still a controversial solution due to food safety
and fears of the public.
Instead of introducing mutations into the genome, plant breeders have been able to
identify plants with spontaneous mutations which have been beneficial for industrial uses. Plant
breeders use this selective breeding technique to successfully produce soybeans with reduced
anti-nutritional factors. There have been mutations identified in the genes responsible for β-
conglycinin and glycinin proteins which results in these proteins to be absent in the soybean
(Clarke and Wiseman, 2000). The literature also reveals genetic selection for soybeans with low
phytic acid, low trypsin inhibitor, and low lectins which have already been developed and may
be a solution to a lot of the issues caused by eating soybean products (Clarke and Wiseman,
2000). The major drawback to selective breeding for positive traits in soybean meal is that it is
time consuming. Identifying plants which possess genetic mutations involves screening of
multiple plants and using different selection pressures, which are time consuming. Selective
breeding may be a strategy to remedy one of the anti-nutritional factors of soybean at a time but
it is not a complete solution for all of the negative factors.
1.3.3 Addition of enzymes
Most of the negative factors of soybeans are anti-nutritional because young animals do
not possess a source of endogenous enzymes that could digest the factors. Research has shown
that adding exogenous enzymes to the soybean can actually reduce the anti-nutritional factors. It

22
has been found that treating soybean meal with proteases produced by Bacillus and Aspergillus
species was successful in reducing the allergenic proteins of soybean meal, effectively improving
piglet performance (Rooke et al., 1998). Several microbes have the ability to produce phytase
which can digest phytic acid, releasing phosphorus which can be used by animals. Several
studies have shown that supplementing soybean meal with purified phytase of microbial origin
can effectively increase the bioavailability of phosphorus and decrease any chelating effects
observed due to phytic acid (Jongbloed et al., 1992). Treating soybean meal with α-
galactosidases isolated from Lactobacillus or Aspergillus species also decreased the
oligosaccharide stachyose concentrations which improved the nutrient digestion leading to an
improved piglet performance (Yoon & Hwang, 2008; Pan et al., 2002). Supplementing β-
glucanases and xylases originally from Trichoderma longibrachiatum resulted in a decrease of
NSP, leading to an increase in apparent ileal digestibility of amino acids, decrease in the
viscosity of the digesta, which all led to an improvement in overall piglet performance (Yin et
al., 2001). The literature suggests that most of the anti-nutritional factors in soybean meal could
be reduced using enzyme supplementation. One of the downfalls to using this approach is it is
costly to produce and purify all necessary enzymes.
1.3.4 Microbial fermentation
Lastly, microbial fermentation is another way of improving soybeans in order to be fed to
humans and animals. Fermentation is one of the oldest techniques widely used in the many
cultures. There are many soy products that, when fermented, have improved nutritional value.
Several different microbes can be used to ferment soybeans. Some of the most commonly used
microorganism used are: Aspergillus species, Bacillus species, and Lactobacillus species. One

23
of the first studies to experiment with utilizing the fermentation method in soybean meal was
Hong et al., (2004). The authors found that the soybean meal fermented with Aspergillus oryzae
for 48 hours was successful in degrading the large allergenic proteins, increasing crude protein
content, and improving the non-essential amino acid profile (Hong et al., 2004). One of the short
comings of this study was the essential amino acid profile was unchanged which suggested that
the mould preferentially uses some amino acids (Hong et al., 2004). Similar to these results,
soybeans fermented with A.egypticus has been found to increase the content of free amino acids
and antioxidant capacity (Zhang et al., 2007). The effect of feeding soybean meal previously
fermented with A.oryzae to animals was also investigated. It was observed that chicks fed the
fermented soybean meal had increased villus lengths and deeper crypts (Feng et al., 2007a). It
was also observed that the intestinal enzymatic activities were improved in birds fed fermented
soybean meal rather than conventional soybean meal, while growth performance was improved
but more so starter birds than grower birds (Feng et al., 2007a). Similar to poultry, piglets were
also seen to have improved performance when fed fermented soybean meal. Fermented soybean
meal at inclusion levels of 10-15% were found to improve feed efficiency, amino acid
digestibility, and increase blood urea nitrogen in piglets (Cho et al., 2007). The increase in blood
urea nitrogen levels was attributed to the high utility value of the fermented soy protein (Cho et
al., 2007). Protein from fermented soybean meal was found to contain more digestible amino
acids than conventional soybean meal but had similar standardized ileal digestibility of amino
acids when compared to that of enzymatically hydrolyzed soybean meal as well as fish meal
(Cervantes-Pahm & Stein, 2010). The improvement in piglet performance and overall increase
in digestibility of fermented soybean meal is due to the elimination multiple types of anti-
nutritional factors found in soybeans. Trypsin inhibitor is heat labile and thus is mostly
eliminated with heat treating of soybean meal. However residual trypsin inhibitor activity in

24
heat treated soybean meal which was successfully decreased with fermentation (Hong et al.,
2004). Trypsin inhibitor negatively affects protein digestibility in piglets and thus, reducing this
increases the digestibility of soybean meal (Kim et al., 2010). Moulds like Aspergillus species
are known for secreting a large amount of extracellular enzymes, including proteases, amylases,
α-galactosidases (Cho et al., 2007).
Bacteria have also been used to ferment soybeans with similar beneficial aspects seen
with fermented soybean meal using mould. Several Bacillus strains have been used and isolated
from different traditional fermented foods. Fermenting soybeans with Bacillus natto has been
found to improve antioxidant capacity compared to that of soybeans (Hu et al., 2010) and
soybean meal fermented with Bacillus subtilis (Wongputtisin et al., 2007). Soybean meal
fermented with Bacillus subtilis has also been found to have a positive effect on piglets. It was
observed that piglets fed fermented soybean meal had improved feed intakes, increased villus
heights, and improved intestinal enzymatic activity (Feng et al., 2007b). Soymilk fermented
with Bacillus natto was found to have reduced IgG reactivity to allergens found in soymilk
indicating the bacteria was successful in degrading the allergenic proteins (Yamanishi et al.,
1995). B.subtilis fermented soybean was seen to be superior to that of A.oryzae or R.oryzae
fermentation, in terms of reducing allergenic proteins and improving amino acid profiles (Frias et
al., 2007). There have also been other beneficial effects of fermenting soybeans using Bacillus
as the inoculum. Spores of some species of Bacillus are commonly used as a probiotic in animal
feeds with observed improvements in both sow and piglet performance (Kyriakis et al., 1999;
Alexopolilos et al., 2004). Interestingly, growing pigs that were fed a balanced diet with the
inclusion of 5% Bacillus subtilis fermented soybean meal were found to have similar average

25
daily intake and average daily gain values as the piglets fed balanced diet with the addition of
100 mg of chlorotetracycline/ kg of feed when challenged with Salmonella Typhimurium; both
treatments improved growth performance compared to the piglets in the antibiotic-free control
group (Gebru et al., 2010). Some strains of Bacillus licheniformis previously isolated from
fermented foods were found to have antimicrobial activity (Kim et al., 2004) which may be a
beneficial way of reducing the use of antibiotics in farm animals. Similar to the mould
fermentation, the elimination of anti-nutritional factors found in soybean meal can be one of the
factors contributing to the beneficial effects on animal performance. The probiotic and increase
antioxidant effects of fermented soybean meal with Bacillus species may be another factor
contributing to the beneficial effects.
Lactic acid bacteria, such as Lactobacillus and Lactococcus species, are commonly used
to ferment milk products such as cheese. Lactobacillus species can secrete α-galactosidase
(Refstie et al., 2005) which is suggests that these bacteria can be used to break down soy
oligosaccharides. Fermenting soy white flakes with Lactobacillus brevis for 36 hours resulted in
a moderate decrease of raffinose concentrations to 36.7 g/kg from the 41.0 g/kg found in the
unfermented white flakes (Refstie et al., 2005). Contents of some essential amino acids, notably
arginine, methionine, threonine and valine were increased with fermentation compared to
unfermented white flakes (Refstie et al., 2005). The fermentation was also able to reduce trypsin
inhibitor levels compared to the unfermented control (Refstie et al., 2005). Lactobacillus
plantarum also has the proteolytic activity necessary to degrade the allergenic proteins found in
soy flour when fermented in a liquid-state (Frias et al., 2007). It was also found that this method
of fermentation resulted in reduced extractable protein compared to the solid-state method of

26
fermentation (Frias et al., 2007). Lactobacillus plantarum with additional supplemented
proteases, were able ferment soybean meal and increase the amount of free amino acids and
decrease allergenic proteins (Amadou et al., 2011). Similar to fermented soybean meal using
Bacillus species, soybeans fermented with Lactobacillus species were found to have improved
antioxidant profiles (Pyo et al., 2005). This increased antioxidant activity was attributed to that
bacteria’s ability to convert the soy isoflavones daidzin and genistein into their bioactive
components daidzein and genistein (Pyo et al., 2005). Some Lactobacillus species had also been
found to be probiotic and which could lead to an additional benefit to piglet performance (De
Angelis et al., 2006). Atlantic salmon fed 40% protein from fermented soy white flakes were
found to have improved feed efficiency and less intestinal pathologies when compared to the fish
fed unfermented white flakes (Refstie et al., 2005).
1.4 Summary and rationale
Soybean meal remains to be one of the most widely used protein sources in animal feeds.
Its high content of crude protein, excellent amino acid profile, and isoflavones are just some of
the benefits of using soybeans. Unfortunately soybean’s unprocessed form contains several
allergenic proteins and other anti-nutritional factors. Industry pressures have adopted the early
weaning practice for piglets. Feeding the desired amount of soybean protein to the early weaned
piglet becomes a challenge due to its allergens, other anti-nutritional factors and the
underdeveloped gut of the piglet. Microbial fermentation of soybean meal is a means for
decreasing these undesirable factors, improving the amino acid profile, increasing antioxidant
capacity and potentially providing a protective benefit (probiotic) for the piglet depending on the
bacterial strain used. Identifying new food-derived bacteria that are capable of effective

27
fermentation would offer opportunity for more cost effective fermentation of soybean meal, and
thus be beneficial to the animal production industry.
2.0 HYPOTHESES
Isolating bacteria derived from fermented food and intestine of the Ctenopharyngodon idella
(grass carp) should yield bacteria that demonstrate high protease, cellulase, and amylase activity.
Using bacteria with high enzymatic activity should improve the nutritional value of soybean
meal largely through the reduction of allergenic proteins and other anti-nutritional factors
typically found in soybean meal.
3.0 OBJECTIVES
1. Screen bacteria from various fermented food sources and grass carp to isolate the bacteria
with the highest protease, amylase, and cellulase activity.
2. Characterize the potentially useful bacteria
3. Determine the efficiency of fermenting soybean meal using isolated bacteria, by
characterizing the soluble protein content, protein profile, crude protein and amino acid
concentrations, detect soy allergens, and raffinose and stachyose concentrations in partly
fermented soybean meal.

28
4.0 MATERIALS AND METHODS
4.1 Isolating and screening bacteria with high enzyme activity
4.1.1 Primary screening for bacteria with protease activity from fermented food
One of the main ways of improving the nutrient value of soybean meal is decreasing the
high-molecular weight and allergenic proteins. To successfully accomplish this, various bacteria
from different fermented foods were screened. Five types of fermented bean curds and
fermented bean products were obtained from local Chinese supermarkets for bacteria isolation.
Approximately 1 cm3 of each source was cut using a clean scalpel and put into a 1.5 mL
Eppendorf tube previously filled with 500 µL of sterile water. The mixture inside the tube was
mixed using a vortex. Serial dilutions were performed by taking 10 µL of the solution and re-
suspending it in 990 µL of sterile water to obtain a dilution of 10-2. Subsequently, 10 µL of the
10-2 diluted sample was re-suspended in 990 µL of sterile water, obtaining a dilution of 10-4.
This was repeated to obtain 10-6 and 10-8 dilutions. Serial dilutions were performed to obtain
approximately 100 colonies of bacteria per culture plate to allow isolation of clearly defined
individual colonies. Diluted samples were screened for ability to grow on soymilk agar using
methods previously described with modifications (Amoa-Awua et al., 2006). In brief, soy milk
agar was made by mixing together 1 gram of soy milk powder (Bulk Barn, Canada) and 2.5
grams of BactoAgar (Becton, Dickinson & Company, U.S.A) with 100 mL of water and
autoclaved for 25 minutes at 121°C. 10 mL of this soymilk agar was poured into petri dishes and
left to cool. 100 µL of the 10-4 and 10-6 diluted of each of the five sources was pipetted onto the
soymilk agar plates and spread using sterile glass beads, aseptically using a microbiology
cabinet. Plates were inverted and incubated for 24 hours in 37 °C. Cultures that were able to
grow on this minimal media and were able to demonstrate a clear zone around its colony was

29
selected for the second round of screening. A glycerol stock of each selected bacteria was
created and stored at – 80 °C in a cryopreservation vial. To do this, a liquid culture for each of
the selected bacteria was generated by inoculating the colony into 1 mL of previously prepared
sterile Bacto Brain Heart Infusion (BHI) (Becton, Dickinson & Company, U.S.A) culture media
according to manufacturer’s protocol. Each of the liquid cultures were incubated for 24 hours at
37 °C. Glycerol stocks were prepared by adding 250 µL of 50% glycerol and 250 µL of bacteria
liquid culture into a cryopreservation vial. Bacteria from this liquid culture was also streaked
onto a previously prepared BHI agar plate and incubated for 24 hours at 37 °C. BHI agar plates
were prepared following manufacturing recommended procedures. These plates were used for
phenotypic characterization of the bacteria using MALDI-TOF MS.
4.1.2 Primary screening for cellulase activity from grass carp source
In order to reduce the amount of non-starch polysaccharides in soybean meal, bacteria
which secreted cellulase, in particular β-1,4-glucanase, were screened. In order to achieve this
we chose the herbivorous fish, the grass carp (Ctenopharyngodon idella) as the source for
selecting cellulolytic bacteria. Two sets of intestines from freshly killed grass carp were
obtained from a local Chinese supermarket and kept on ice until processed. Bacteria were
isolated according to the methods previously described (Li et al., 2009). In brief, the intestines
of each of the two fish were stretched out and the proximal and distal ends of the intestine were
identified. The intestine was sectioned by introducing two cuts using a scalpel at approximately
1/3 and 2/3 length from the proximal end of the intestine. The most proximal third of the
intestine was considered to be the “beginning” of the intestine, the second third the “middle” and
the most distal third the “end”. Contents from each of the three sections were emptied into a 15

30
mL sterile conical tube and mixed using a vortex. One mL of the content was diluted in 9 mL of
sterile deionized water and vortexed to create a more homogenous solution. Serial dilutions of
each section’s contents of both fish intestines were performed as described above. 100 µL of
liquid from each of the serial diluted solutions (10-2 , 10-4, 10-6, and 10-8) were plated on
previously prepared Luria Bertani (LB), BHI, and DeMan, Rogosa and Sharpe (MRS) (Oxoid,
England) agar plates and spread using sterile glass beads. BHI and MRS agar plates were
prepared according to manufacturer’s procedures with the addition of 1% (w/v) BactoAgar
(Becton, Dickinson, & Company, U.S.A) and autoclaved. LB agar plates were prepared by
adding 1.0 gram of Tryptone Powder (Bio Basics Canada INC, Canada), 0.5 grams of Yeast
Extract (Bio Basics Canada INC, Canada), 1.0 grams of NaCl (Fisher Scientific, U.S.A) to 80
mL of deionized water stirred on a magnetic plate with a magnetic stir bar. The pH of the
solution was adjusted to 7.5 using NaOH (Fisher Scientific, U.S.A), 1.5 grams of BactoAgar was
added, and more deionized water was added to reach 100 mL before being autoclaved for 25
minutes at 121 °C. These plates were inverted and incubated at 37 °C overnight. Any colonies
present on the plates were blotted onto previously prepared soymilk agar and carboxymethyl
cellulose (CMC) (Acros Organics, U.S.A) plates using sterile filter paper, to test if the colony
can grow using soybean nutrients and has cellulose degrading activity. CMC agar plates were
prepared according to previously described procedures with slight modifications (Sazci et al.,
1986). CMC agar plates were prepared by adding 1.0 grams of CMC and 1.5 grams of
BactoAgar to 100 mL of deionized water and autoclaved for 25 minutes at 121 °C. Soymilk and
CMC agar plates were inverted and incubated at 37 °C for 72 hours. CMC agar plates were
stained by flooding the agar plates with 1.0 % (w/v) Congo Red Staining Solution for 15
minutes; Congo Red solution was prepared by adding 0.1 gram of Congo Red Dye (Fisher
Science Education, U.S.A) to 10 mL of deionized water and mixed thoroughly until dye was

31
dissolved. Stained agar plates were de-stained with 1M NaCl (Fisher Scientific, U.S.A) by first
by discarding any excess stain into a waste container and flooding the plates with 1M NaCl with
gentle agitation for 15 minutes (Ruijssenaars & Hartmans, 2001 ). Colonies that exhibited an
orange halo after being de-stained were matched up to the colonies that grew on the soymilk
agar. Colonies that exhibited an orange halo and were able to grow on the soymilk agar were
selected for the second round of screening.
4.1.3 Second round of screening
The first round of screening was intended to identify isolates, from each of the five
fermented food sources and the grass carp, which demonstrated high protease and cellulase
activity, respectively. One of the factors required to create an optimal fermentation is a shorter
length of fermentation. In order to achieve this, the bacteria selected must grow and secret large
concentration of extracellular enzymes in a short period of time. The second round of screening
sought out to rank the protease, cellulase, and amylase activity of the isolates. To identify the
bacteria which demonstrate the highest protease, amylase, and cellulase activity, we needed to
first extract the extracellular enzymes of each bacterium. Crude enzymes of the bacteria were
extracted according to methods previously described (Chantawannakul et al., 2002). First, a
single colony from each of the selected bacteria was obtained from the previously streaked BHI
agar. The colony was inoculated into 1 mL of BHI culture media and incubated for 18 hours at
37 °C. A commercially available source of B. subtilis ATCC 6633 was used as a positive control
due to its previous observation for production of protease activity (Dias et al., 2008), amylase
activity (Mitrica & Granum, 1979) and cellulase activity (Mawadza et al., 2000). After the 18
hours of incubation, each of the liquid cultures were emptied into a clean 1.5 mL Eppendorf tube

32
and centrifuged at 10,000 rpm for 15 minutes at 4 oC. Centrifuging the liquid culture allows the
enzymes to be separated from the bacteria pellet so that only secreted enzyme containing
supernatant can be harvested. Agars for screening for protease activity, using 1.0% (w/v)
soymilk agar, and cellulase activity, using 1.0% (w/v) CMC agar, were prepared in similar
methods as described in the first round of screening. Identification of amylase activity was
conducted by using 1.0% corn starch agar plates as previously described (Amoa-Awua et al.,
2006). To prepare starch agar plates, 1 gram of corn starch (Bulk Barn, Canada) and 1.5 grams
of BactoAgar was added to 100 mL of deionized water and autoclaved for 25 minutes at 121 °C.
To screen for each of the three enzymes, 15 mL of their respectful nutrient agars was poured
onto a petri dish (90 mm x 15 mm) and allowed to settle on a flat surface. Once the agars were
solidified, a 7 mm hole was punctured into the middle of the plate using the end of a sterile P-
100 pipette tip. Subsequently, 20 µL of the crude enzymes extract was pipetted into the 7 mm
hole for each of the three nutrient agar plates and, after five minutes, were inverted and incubated
at 37 °C for 72 hours. An additional plate, for each of the three select agars, was filled with 20
µL of just blank culture media to be used as a negative control. For protease activity no
destaining was required and clearance zone measurements were obtained from measuring the
length from the edge of the clearance zone to the edge of the bacterial growth. For cellulase
activity, plates were stained with 1.0% (w/v) Congo Red Staining solution and destained using
1M NaCl, as detailed in first round screening. Clearance zone measurements were taken from
the edge of the punctured hole to the outer edge of the orange halo. To detect amylase activity,
corn starch agar plates were stained with Gram’s Iodine Stain as previously described (Amoa-
Awua et al., 2006). Amylase clearance zone measurements were obtained by measuring the
length from the edge of the punctured hole to the edge of the clearance zone. Each observation
was replicated three times using fresh inoculums each time.

33
4.2. Identification of bacteria selected
4.2.1: MALDI-TOF MS:
All the bacteria that were selected for the second round of screening were also identified.
Each isolate was streaked from the glycerol stock onto a previously prepared BHI agar plate,
inverted, and incubated for 18 hours at 37 °C. Colonies on agar plates were submitted to the
Animal Health Laboratory to be identified using a matrix-assisted laser desorption ionization-
time of flight mass spectrometry (MALDI-TOF MS, Bruker, Canada). Briefly, a fresh cultured
bacterial colony was spotted onto the target plate (Bruker) and left to air dry. The sample was
overlayed with 1 µL matrix solution mixed with an organic solvent solution made of 50%
acetonitrile (Sigma Aldrich, Canada) and 2.5% trifluoroacetic acid (AnalaR Normapur through
VWR, Canada). The target plate was air dried then placed onto the MALDI-TOF instrument
(Bruker) for analysis. Peptide mass fingerprint spectra was read from the target plate and scored
using MALDI-TOF MS software and database (MALDI Biotyper 3.0, Bruker). The software
automatically identified an isolate to species level if the score was 2.0 to 3.0, or to genus level if
the score was 1.7 to 1.999, and no reliable identification is generated if the score was less than
1.7.
4.2.2: PCR Amplification of partial 16S rRNA gene and gyrB gene:
The next step required for identification of the bacteria used in the second round of
screening is sequence analysis of two genes: partial 16S ribosomal RNA gene and gyrB gene.
Genomic DNA of each isolate was extracted from the bacterial cells using the PureLink
Genomic DNA Mini Kit (Invitrogen, U.S.A) according to manufacturers’ instructions. The 16S
rRNA gene in the DNA was amplified using PCR previously described (Cai et al., 2003). In

34
brief, primers BSF8/20 (5’-AGAGTTTGATCCTGGCTCAG-3’) and BSR534/18 (5’-
ATTACCGCGGCTGCTGGC-3’) were used to amplify the 16S rRNA gene with the expected
product size of around 500 bp. For the three genes the PCR reaction had 2 µl of extracted DNA
as a template, 3 U/100 µL of Taq Polymerase (Invitrogen, U.S.A), 0.1 mM dNTPs, 0.2 µM of
each primer, and 1X PCR Buffer for a total of 25 µL per reaction. For the 16S rRNA gene, PCR
was performed using the PCR thermo cycler under the following conditions: 95 °C for 3 min; 35
cycles of amplification with template denaturing at 95 °C for 20 sec, primer annealing at 55 °C
for 30 sec, and primer extension at 72 °C for 90 sec; with a final extension at 72 °C for 5 min.
After the reaction was complete, the concentration of the PCR products was measured using a
BioPhotometer Plus spectrometer (Eppendorf, U.S.A) to ensure that the minimum concentration
of DNA needed for sequencing was obtained.
In order to further identify the strain of the isolates the DNA gyrase subunit B (gyrB)
gene was sequenced. The gyrB gene is very fast evolving gene, necessary for DNA replication,
and is distributed universally among bacteria species (Yamamoto & Harayama, 1995), thus
making it a perfect candidate gene for bacteria identification. To sequence the gyrB gene, two
sets of primers were ordered from Integrated DNA Technology (Iowa, U.S.A). The first set of
primers was:
UP-1: 5’-GAAGTCATCATGACCGTTCTGCA(TC)GC(TCAG)GG(TCAG)GG(TCAG)
AA(AG)TT(TC) GAGAAGTCATCATGACCGTTCTGCAYGCNGGNGGNAARTTYGA- 3’
UP-2r: 5′-AGCAGGGTACGGATGTGCGAGCC(AG)TC(TCAG)AC(AG)TC(TCAG)GC(AG)
TC(TCAG)GTCATAGCAGGGTACGGATGTGCGAGCCRTCNACRTCNGCRTCNGTCAT-
3’ and the second set of primers were UP1S : 5’-GAAGTCATCATGACCGTTCTGCA-3’ and

35
UP2SR – 5’-AGCAGGGTACGGATGTGCGAGCC-3’ according to previous study (Yamamoto
& Harayama, 1995). The UP-1 and UP-2r primers were designed by reverse transcribing the
gyrB protein of three different types of bacteria (Yamamoto & Harayama, 1995). The first
twenty-three nucleotides of primers UP-1 and UP-2r are not degenerate and therefore may not
bind to the complementary strand on the target gene (Yamamoto & Harayama, 1995). This
allows the 5’ regions of both primers to be used as a tag sequence for sequencing (Yamamoto &
Harayama, 1995) which is what UP1S and UP2SR primers are for. The PCR mix contained 1.0
µM of each UP-1 and UP-2r primers, 1.25 mM MgCl2, 0.2 mM dNTP, 1.5 U Taq Polymerase,
1X PCR Buffer and 1.0 µL of template DNA. The PCR was performed using the PCR thermo
cycler under the following conditions: 94 °C for 3 min; 35 cycles of amplification with template
denaturing at 94 °C for 1 min, primer annealing at 48 °C for 1 min, and primer extension at 72°C
for 3 mins. After the reaction was complete, the concentration of the PCR products was
measured using a BioPhotometer Plus spectrometer (Eppendorf, U.S.A) to ensure that the
minimum concentration of DNA needed for sequencing was obtained.
To verify the correct PCR product sizes, the PCR products of both gene reactions were
subjected to agarose gel electrophoresis in separate gels. A 1% agarose gel for electrophoresis
was prepared by mixing 0.7 grams of agarose powder (Invitrogen, U.S.A) with 70 mL of 1X
sodium borate (SB) buffer and boiling the mixture for approximately 2 minutes in the
microwave. Once the agarose was cooled, 10 µL of RedSafe was added in order to visualize the
DNA under UV light. The solution was then slowly added to the plate of the gel-casting tray and
a sample comb with 10 wells was added to create wells in the finished gel. Once the gel was
completely solidified, the sample comb was removed and the gel was placed in the

36
electrophoresis equipment filled with 1X sodium borate (SB) buffer. Each of the wells were
loaded with 9 µL of PCR product, mixed with 1 µL of 10X loading buffer, and the gel
electrophoresis was set to run for 35 minutes at 100 mV. The DNA on the gel was visualized
using UV illumination for verification of the expected product size. Once the correct product
size was observed, the remaining PCR products were sent for sequencing at a DNA Sequencing
Facility (CBS, University of Guelph, Canada) by using an ABI Prism 3100 Automated
Sequencer. For the 16S rRNA gene sequencing the same primers used for the PCR reaction
were used for sequencing. For the gyrB gene sequencing primers UP1S and UP2SR were used
for sequencing. Bacterial strains were identified by using the sequencing results from both genes
and comparing the nucleotide sequences in the GeneBank database
(http://blast.ncbi.nlm.nih.gov/Blast.cgi) using the Basic Local Alignment Search Tool (BLAST)
search algorithm (Altschul et al.,1990).
4.3. Soybean meal fermentation
4.3.1. Preparation of soybean meal and inoculums
After the isolates with high enzyme activities were identified, their ability to ferment
soybean meal was investigated. The first step in preparing for the fermentation was carefully
heat treating the soybean meal to reduce the number of bacteria present in the stock soybean
meal. Sterilizing soybean meal in the autoclave has been found to be detrimental to lysine
(González-Vega et al., 2011) and would be costly and impractical to do in the feed industry. To
prevent any damage to lysine and other amino acids, the soybean meal used was gently heated
according to previous methods which showed no negative effects to lysine (González-Vega et
al., 2011). This was performed by spreading approximately 50 grams of 48 % crude protein

37
soybean meal (Grand Valley Fortifiers, Canada) on an aluminum tray and heated in the oven-
dryer for 30 minutes at 125 °C (González-Vega et al., 2011). Treated soybean meal was
removed from the oven, covered in aluminum foil, and stored in a previously autoclaved glass jar
until needed.
Bacteria inoculums of Isolate-2, Isolate-4, Isolate-7, Isolate-8, F-8, F-9, and F-11
(selected based on enzyme activity measure described above) were prepared fresh for each
fermentation. Each isolate was streaked onto a previously prepared BHI agar plate from each of
their glycerol stocks. Bacteria on the agar plates were grown for 18 hours in 37 °C. Each
inoculum was prepared by inoculating a single colony from each isolate’s agar plate, obtained
using a sterile inoculating loop, into a 50 mL sterile flask containing 10 mL of BHI culture
media. Liquid cultures were incubated for 18 hours at 37 °C and a colony forming unit (CFU)
measurement was taken by diluting the inoculum by a serial dilution and plating 20 µL of the
dilutions on BHI agar plates. To obtain the CFU/mL the number of colonies on the plate was
multiplied by the dilution number and divided by 0.02 mL. The liquid cultures incubated under
these conditions obtained a culture of 108 CFU/mL.
4.3.2. Liquid-state fermentation
The first fermentation investigated was a high-moisture fermentation further referred to
as liquid-state fermentation. The entire preparation of the fermentation was performed under
sterile conditions. In the liquid-state fermentation only the bacteria isolated from food sources
(Isolate-2, Isolate-4, Isolate-7, and Isolate-8) were used. The fermentation process was initiated
by weighing 2 grams of the previously treated soybean meal into a sterile 50 mL flask.

38
Subsequently, 10 mL of sterilized deionized water was added and the mixture was set aside for
an hour to allow the soybean meal to absorb the water. After the hour, 1 mL of each bacteria
liquid culture was inoculated into the soybean meal mixture. There were three replicates per
treatment. This mixture was secured into a shaker incubator (New Brunswick, U.S.A) set at 37
°C, 350 rpm, for 48 hours. There was an additional soybean meal fermentation set-up that used
the same soybean meal as the treatments with the exception of only being inoculated with blank
culture media. This was considered to be the negative control and was directly stored in the -80
°C freezer once the water was added to the soybean meal, representing the soybean meal pattern
completely unfermented. Samples of the fermented soybean meal were taken at 24 and 48 hours
and stored at -80 °C. All of the samples of each treatment group were were lyophilized with a
freeze dryer (Labconco, U.S.A) for 24 hours and ground into a fine powder, using a mortar and
pestle, and used for further analysis.
4.3.3. Solid-state fermentation
The second fermentation condition was low-moisture fermentation, further referred to as
solid-state. The solid state fermentation was initiated by weighing 2 grams of previously heated
soybean meal into a sterile 50 mL flask. There were seven treatment groups and two controls for
each fermentation. Each treatment and control groups were replicated three times. The
treatment groups consisted of inoculum prepared from isolated with high enzymatic activity
(Isolate-2, Isolate-4, Isolate-7, Isolate-8, F-8, F-9, and F-11). The negative control contained
blank culture media instead of inoculums and was directly stored in -80 °C. The treatment
groups were set up by mixing 2 mL of each inoculum (108 CFU/mL) with 2 mL of sterile
deionized water. This was done in order to evenly distribute the inoculum. This diluted liquid

39
culture was inoculated into the 2 grams of soybean meal for each of the treatment groups. This
mixture was set to ferment in an incubator at 42 °C for 48 hours without any agitation. Samples
were collected at 24 and 48 hours and stored at -80 °C until lyophilized. Samples were
lyophilized using a freeze dryer (Labconco, U.S.A) for 24 hours and then ground into a fine
powder using a mortar and pestle. Only lyophilized, finely ground unfermented and fermented
products were used for further analysis.
4.4 Analysis of soybean meal fermentation products
4.4.1 Proximate analysis and amino acid profiling
Finely ground lyophilized solid-state unfermented and fermented samples were sent to
Dr. Zhang’s group at the Institute of Animal Husbandry, China Academy of Agricultural
Science (Beijing, China) to determine dry matter (AOAC official method 934.01), crude protein
(AOAC official method 954.01), and amino acid concentrations (AOAC official method 994.12).
4.4.2 Determination of soluble protein fractions and distribution
Total soluble protein of the unfermented and fermented soybean meal was isolated as
previous described with slight modifications (Hong et al., 2004). First, 0.05 grams of
lyophilized finely ground fermented product was homogenized using an ultrasonic cell disruptor
(Misonix, U.S.A) with 1 mL of deionized water for 30 seconds on ice. The mixture was then
centrifuged at 10,000 rpm for 15 minutes at 4 °C. The supernatants of each product were
transferred into a clean 1.5 mL Eppendorf tube and used for total soluble protein quantification
using the DC Assay Kit (Bio-Rad Laboratories, U.S.A).

40
Samples of soluble protein(supernatants) were analyzed by sodium –dodecyl sulfate-
polyacrylamide gel electrophoresis (SDS-PAGE) using an 11% tris gel which used a 1X Tris
running buffer (0.12 M Tris, 0.96 M Glycine, and 0.5% SDS). The total volume loaded per well
was 20 µL which consisted of 15 µL of the crude protein sample diluted with 5 µL of 5X loading
buffer (5% SDS, 20% glycerol, 0.05% β-Mercaptoehtanol, and 0.001% Bromophenol) and
boiled for 7 minutes at 90 °C prior to being loaded and subsequently subjected to
electrophoresis for 100 minutes using 100 V. Ten µL of Unstained Protein Ladder (Bio-Rad,
U.S.A) was used as a marker for molecular mass from 10-250 kDa. Following electrophoresis,
the gel was stained using a Coomassie blue staining solution which consists of 0.5% Coomassie
Brilliant Blue G-250, 50% methanol, and 10 % acetic acid for 5 minutes. The gel was de-stained
using solution containing 40% methanol and 10% acetic acid, for two hours replacing the de-
staining solution every 20 minutes and left to continue de-staining overnight in Milli Q water.
An image of the gel separating the proteins of the liquid-state fermentations was captured using
white light conversion screen on the GelDoc (Syngene, U.S.A). An image of the gel separating
the proteins from the solid-state fermentation was taken using white light conversion screen on
the ChemiDoc XRS+ (Bio-Rad Laboratories, U.S.A) and analyzed using the Image Lab
software.
4.4.3 Determination of oligosaccharide concentrations
Finely ground lyophilized unfermented and fermented samples were sent to Dr. Zhang’s
group at the Institute of Animal Husbandry, China Academy of Agricultural Science (Beijing,
China) to determine concentrations of raffinose and stachyose. Contents of stachyose and

41
raffinose were determined using high-performance liquid chromatography (HPLC) (Shimadzu
LC-15C, Japan) equipped with refractive index detector (Shimadzu RID-10A, Japan). Briefly,
stachyose and raffinose were first extracted using ethanol. A total amount of 2.00 g of soybean
meal samples were mixed with 50 ml of 70% (v/v) ethanol, microwaved to boiling and then
cooled down to room temperature. The liquid was removed and transferred to EP tube. The
extraction procedure was repeated and the remaining was rinsed with 70% (v/v) ethanol. All
liquid was collected in the same EP tube for each sample. The collected liquid was centrifuged
at 5000 r/min for 10 min and the supernatant was concentrated to a volume less than 10 ml by
rotary evaporation and finally filled with deionized water to the final volume of 25 ml. The
solution was then centrifuged at 4000 rpm for 10 min to obtain the supernatant, which was
filtrated with 0.22 μm filter before injected into the HPLC system. A volume of 20 μl of each
sample was injected into the HPLC system with 70% acetonitrile (v/v = 7:3) as the mobile phase.
The temperature was set at 35 °C, and the flow rate was 1 ml/min. Standards of stachyose (Dr.
Ehrenstorfer, China) and raffinose (Dr. Ehrenstorfer, China) were used.
4.4.4 Detection of Allergenic Proteins.
A Western blot was performed to detect the allergenic proteins of the unfermented and
fermented soybean meal. The total soluble protein of the unfermented and fermented soybean
meal samples of 50 µg were loaded into an 11% polyacrylamide gel similar to the procedures
described above. Once the electrophoresis step was completed, the proteins were transferred to
an Immobolin-P (Millipore, Billerica, USA) polyvinyl difluoride (PVDF) transfer membrane
using an electro-blotting transfer tank (Biorad, U.S.A). To do this, the PVDF membrane was
immersed in methanol and then washed in deionized water for two minutes. Subsequently, the

42
SDS-PAGE and the membrane were equilibrated with chilled 1X transfer buffer (25 mM Tris,
192 mM glycine, and 20% methanol) in order to remove buffer salts, detergents, and to allow
methanol-dependent shrinking. A transfer cassette was assembled in the following order: fiber
pad, filter paper, SDS-PAG, PVDF membrane, filter paper, fiber pad. The assembled cassette
was placed in the transfer tank and the transfer was run at 100 V for 100 minutes. Once the
proteins have been successfully transferred onto the membrane, the cassette was disassembled
and the membrane washed for 8 minutes in 1X TBS. The membrane was then blocked using
blocking buffer comprised of 5% skim milk, 0.1% Tween 20, and 1X TBS for an hour. The
blocked membrane was subsequently washed for 30 minutes using washing buffer (0.1% Tween
and TBS) and replacing the buffer every ten minutes on an orbital shaker. After the wash, the
membrane was incubated in primary antibody in 4 °C overnight on an orbital shaker. The
primary antibody was prepared following a published protocol (Song et al., 2010). Briefly,
plasma was collected from eight piglets (8-week old) that were previously exposed to feed
containing soybean meal. Piglets develop an immune response to soybean allergens by 21 days
of exposure, and thus should have pre-formed antibodies that should react to the two allergenic
proteins glycinin and β-conglycinin. The primary antibody used was comprised of 20 µL of
pooled pig serum and 10 mL of blocking buffer for a dilution ratio of 1:500. The membrane was
washed again for 30 minutes using washing buffer replacing the buffer every ten minutes on an
orbital shaker. Following the wash, the membrane was incubated with secondary antibody (1:
100,000, rabbit anti-pig horseradish peroxidase-linked IgG; Abcam) for 1 hour at room
temperature. The membrane was washed again for 30 minutes using washing buffer and
replacing the solution every ten minutes. Proteins were detected by using Clarity Western
Enhanced Chemiluminescence (ECL) Substrate (Bio-Rad Laboratories, U.S.A) according to
manufacturer’s instructions. In brief, equal parts of solution A and solution B were mixed and

43
poured onto the membrane and incubated for 1 minute. The membrane was developed by using
the ChemiDoc XRS+ (Bio-Rad Laboratories, U.S.A) and analyzed using the Image Lab
software.
4.5 Statistical analysis:
All statistical analysis was performed using PRISM GraphPad Prism Version 5.03 (San
Diego, CA). The screening data was analyzed using one-way ANOVA with a Dunnet post-test
to determine if any treatment groups were significantly different from the positive control. For
the screening data, observations were made in triplicate and simultaneously. The statistical
analysis of the unfermented and fermented samples was performed using one-way ANOVA with
a Tukey post-test; observations were made in triplicate, with one observation obtained daily.
The assumption of no day effect was believed to be true. Data was expressed as mean ± standard
error of the mean and considered to differ significant by p< 0.05.
5.0 RESULTS
5.1 Qualitative measurement of enzymes activity.
5.1.1 Primary Screening for Protease Producing Bacteria
The first round of screening identified one colony which demonstrated the largest
clearance zone from each of the five sources (Table 1). One source had two colonies which
demonstrated similar clearance zones and were both selected for further screening. Of the six
colonies selected, two of the bacteria demonstrated an additional, different bacterial colony
growth on the agar plate (Isolate-3 and Isolate-6). These two colonies as well as the original six,
were selected for the second round of screening.

44
5.1.2 Primary Screening of Cellulase Producing Bacteria
Of all the bacteria screened, only thirty colonies were able to grow on both soymilk agar
and on CMC agar. Of these 30 bacteria, only 20 colonies demonstrated an orange halo when
stained with congo red dye. Of these 20 colonies only three isolates demonstrated high cellulase
activity, i.e. formed distinguishably larger halos than the other colonies. These three colonies (F-
8, F-9, and F-11) were selected for the second round of screening to be ranked with the other
food-derived bacteria isolates.
5.1.3 Second round of screening
Figure 3A shows the representative images of proteinase positive (right image), and
negative (left image), respectively. The results of the secondary screening for protease activity is
shown in Figure 3. The radius of each clearance zones, reflecting protease activity, were
measured. Of the eight isolates from food sources, only six demonstrated protease activity on
the soymilk agar plates (Figure 3B). From the fermented food source, the isolate which had the
largest clearance zone, and thus the most protease activity, was Isolate-4 (Figure 3B). The three
bacteria isolated from the grass carp demonstrated protease activity. Among them, F-11 had the
largest clearance zone (Figure 3B). Protease activity of Isolate-4 and F-11 were significantly
higher than that of B.subtilis ATCC6633, the positive control (Figure 3).
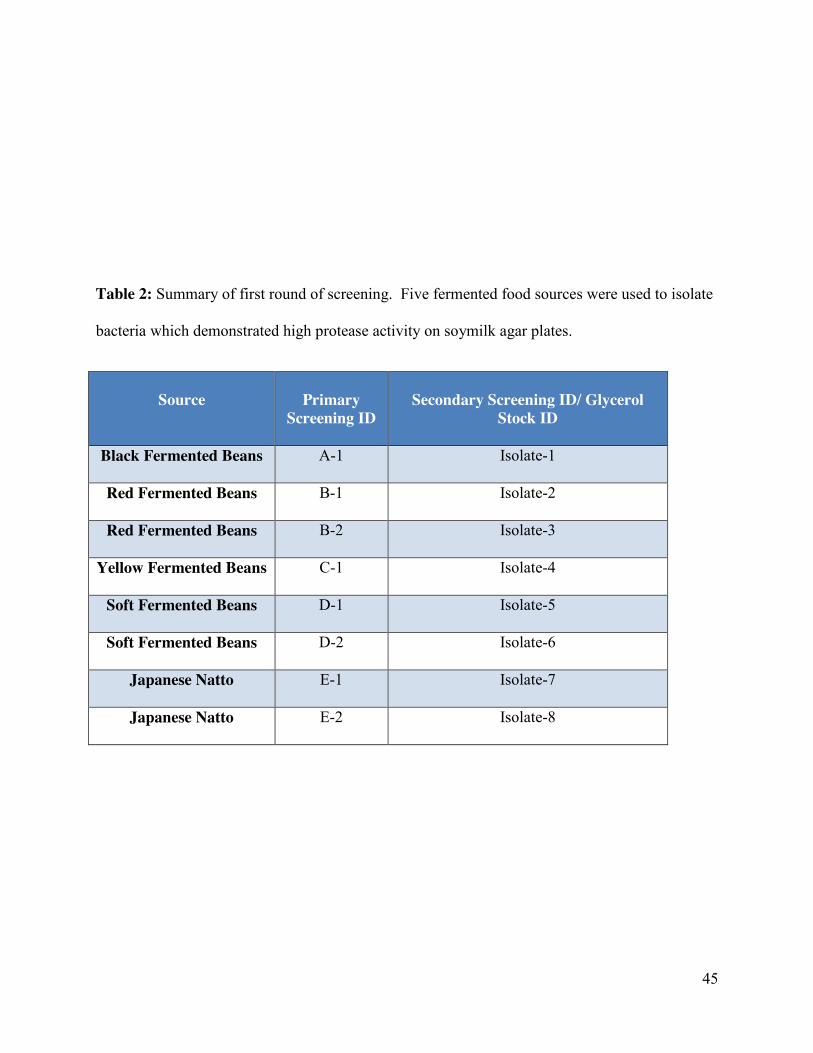
45
Table 2: Summary of first round of screening. Five fermented food sources were used to isolate
bacteria which demonstrated high protease activity on soymilk agar plates.
Source
Primary
Screening ID
Secondary Screening ID/ Glycerol
Stock ID
Black Fermented Beans A-1 Isolate-1
Red Fermented Beans B-1 Isolate-2
Red Fermented Beans B-2 Isolate-3
Yellow Fermented Beans C-1 Isolate-4
Soft Fermented Beans D-1 Isolate-5
Soft Fermented Beans D-2 Isolate-6
Japanese Natto E-1 Isolate-7
Japanese Natto E-2 Isolate-8

46
For the cellulase screening, the positive control plates demonstrated an orange halo,
indicating cellulase activity, as expected (Figure 4 A, right image). All of the negative control
plates did not have orange halos (Figure 4 A, left image). Of the eight bacteria isolated from
food sources, only three isolates (Isolate-2, Isolate-7 and Isolate-8) were able to demonstrate
cellulase activity (Figure 4 B). All three of the bacteria isolated from the fish tested positive for
cellulase activity. Of all the isolates tested, F-8 had a significantly different level of cellulase
activity compared to the positive control (p< 0.05; Figure 4B).
Figure 5A shows representative amylase positive (right), and negative (left) images. Of
all the bacteria isolated from food sources, only three demonstrated amylase activity (Figure 5
B). All of the three bacteria isolated from the fish also tested positive for amylase activity but
only Isolate-2 and Isolate-8 demonstrated significantly (p<0.05) higher amylase activity than the
positive control. A summary of the activities of the three enzymes from bacteria isolated from
food and fish are shown in Table 3.
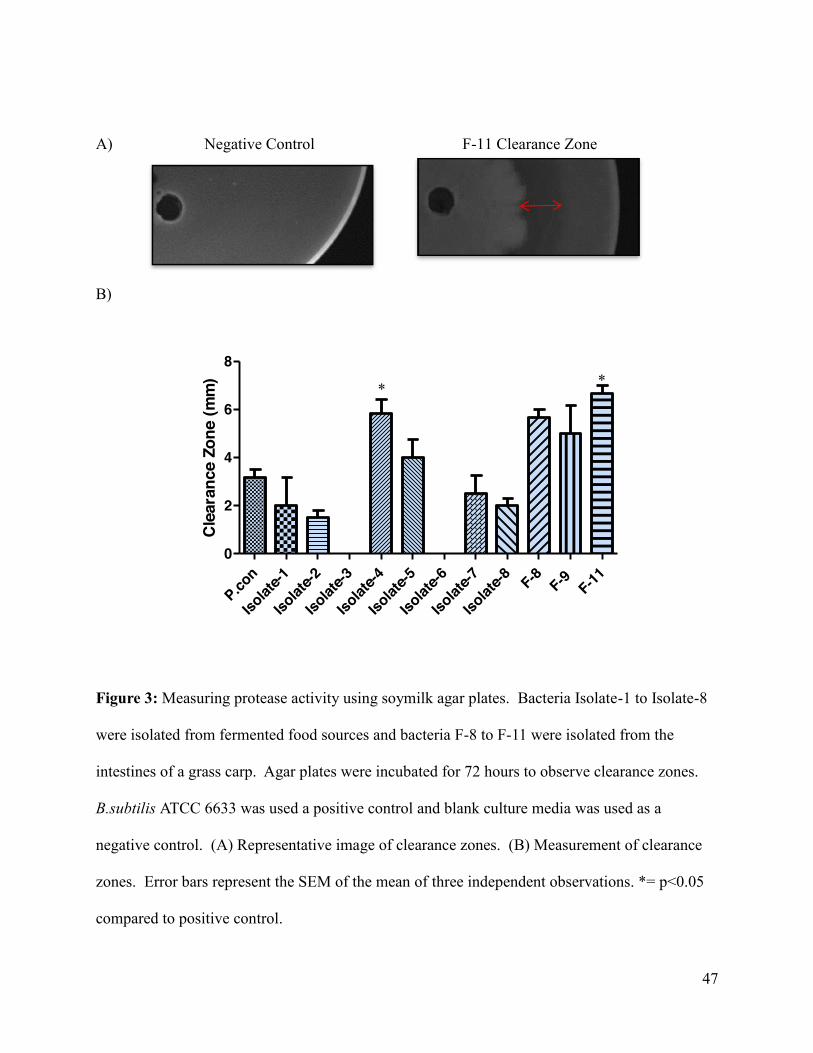
47
A) Negative Control F-11 Clearance Zone
B)
Cle
aran
ce Z
one
(mm
)
P.con
Isolat
e-1
Isolat
e-2
Isolat
e-3
Isolat
e-4
Isolat
e-5
Isolat
e-6
Isolat
e-7
Isolat
e-8 F-8 F-9 F-110
2
4
6
8
Figure 4
Figure 3: Measuring protease activity using soymilk agar plates. Bacteria Isolate-1 to Isolate-8
were isolated from fermented food sources and bacteria F-8 to F-11 were isolated from the
intestines of a grass carp. Agar plates were incubated for 72 hours to observe clearance zones.
B.subtilis ATCC 6633 was used a positive control and blank culture media was used as a
negative control. (A) Representative image of clearance zones. (B) Measurement of clearance
zones. Error bars represent the SEM of the mean of three independent observations. *= p<0.05
compared to positive control.
* *
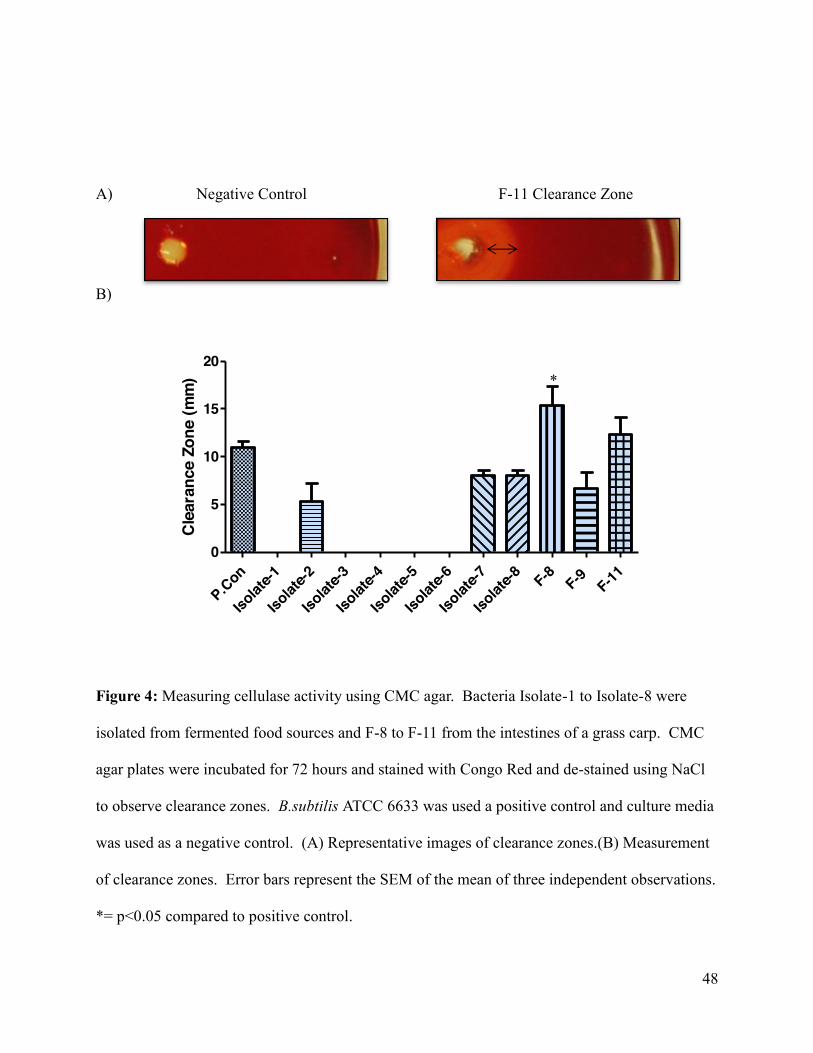
48
A) Negative Control F-11 Clearance Zone
B)
Clea
ranc
e Zo
ne (m
m)
P.Con
Isolat
e-1
Isolat
e-2
Isolat
e-3
Isolat
e-4
Isolat
e-5
Isolat
e-6
Isolat
e-7
Isolat
e-8 F-8 F-9 F-110
5
10
15
20
Figure 5
Figure 4: Measuring cellulase activity using CMC agar. Bacteria Isolate-1 to Isolate-8 were
isolated from fermented food sources and F-8 to F-11 from the intestines of a grass carp. CMC
agar plates were incubated for 72 hours and stained with Congo Red and de-stained using NaCl
to observe clearance zones. B.subtilis ATCC 6633 was used a positive control and culture media
was used as a negative control. (A) Representative images of clearance zones.(B) Measurement
of clearance zones. Error bars represent the SEM of the mean of three independent observations.
*= p<0.05 compared to positive control.
*
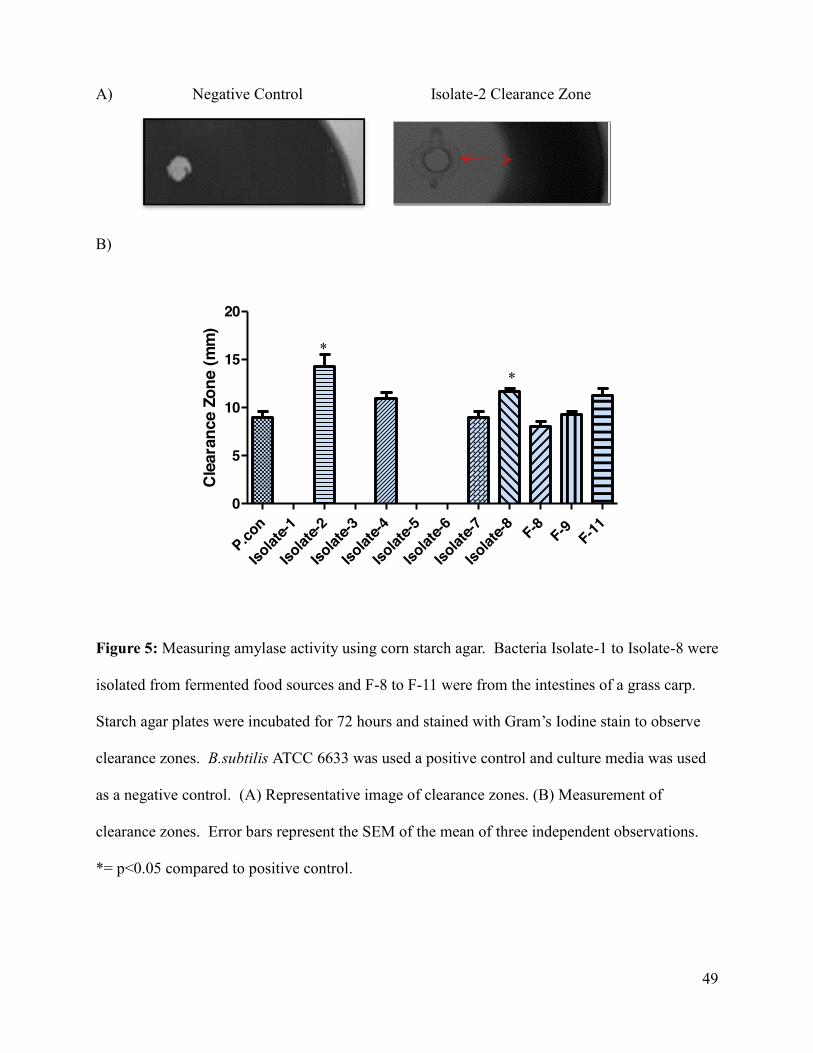
49
A) Negative Control Isolate-2 Clearance Zone
B)
Cle
aran
ce Z
one
(mm
)
P.con
Isolat
e-1
Isolat
e-2
Isolat
e-3
Isolat
e-4
Isolat
e-5
Isolat
e-6
Isolat
e-7
Isolat
e-8 F-8 F-9 F-110
5
10
15
20
Figure 6
Figure 5: Measuring amylase activity using corn starch agar. Bacteria Isolate-1 to Isolate-8 were
isolated from fermented food sources and F-8 to F-11 were from the intestines of a grass carp.
Starch agar plates were incubated for 72 hours and stained with Gram’s Iodine stain to observe
clearance zones. B.subtilis ATCC 6633 was used a positive control and culture media was used
as a negative control. (A) Representative image of clearance zones. (B) Measurement of
clearance zones. Error bars represent the SEM of the mean of three independent observations.
*= p<0.05 compared to positive control.
*
*
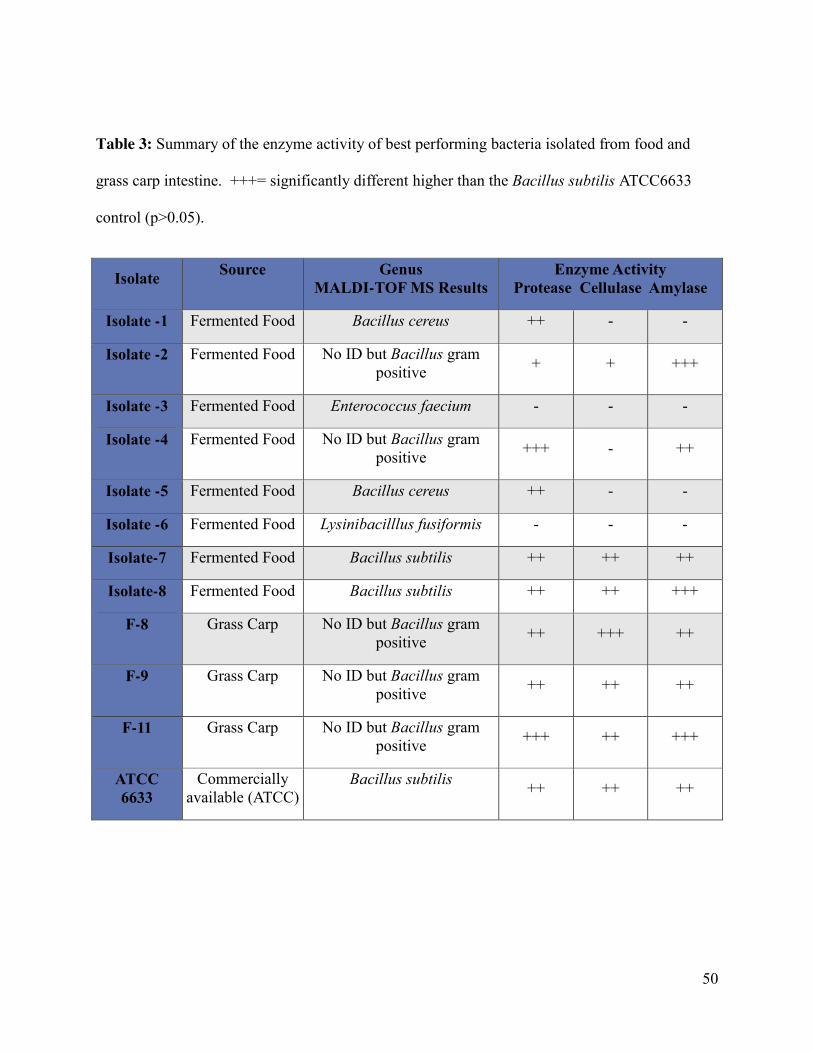
50
Table 3: Summary of the enzyme activity of best performing bacteria isolated from food and
grass carp intestine. +++= significantly different higher than the Bacillus subtilis ATCC6633
control (p>0.05).
Isolate Source Genus MALDI-TOF MS Results
Enzyme Activity Protease Cellulase Amylase
Isolate -1 Fermented Food Bacillus cereus ++ - -
Isolate -2 Fermented Food No ID but Bacillus gram positive + + +++
Isolate -3 Fermented Food Enterococcus faecium - - -
Isolate -4 Fermented Food No ID but Bacillus gram positive +++ - ++
Isolate -5 Fermented Food Bacillus cereus ++ - -
Isolate -6 Fermented Food Lysinibacilllus fusiformis - - -
Isolate-7 Fermented Food Bacillus subtilis ++ ++ ++
Isolate-8 Fermented Food Bacillus subtilis ++ ++ +++
F-8 Grass Carp No ID but Bacillus gram positive ++ +++ ++
F-9 Grass Carp No ID but Bacillus gram positive ++ ++ ++
F-11 Grass Carp No ID but Bacillus gram positive +++ ++ +++
ATCC 6633
Commercially available (ATCC)
Bacillus subtilis ++ ++ ++

51
5.2 Identification of selected bacteria
5.2.1 Characterization of isolates using MALDI-TOF MS
The group of bacteria that were identified for second round screening, were characterized
using matrix assisted laser desorption ionization- time of flight mass spectrometry (MALDI-TOF
MS). Table 3 summarizes all of the MALDI-TOF MS results. Of the eight bacteria isolated
from food, only five were identified to species level. Isolate-1 and Isolate-5 were identified as
Bacillus cereus. Isolate-3 was identified as Enterococcus faecium, Isolate-6 was Lysinibacilllus
fusiformis and Isolate-7 and Isolate-8 were Bacillus subtilis. Isolate-2 and Isolate-4 and the
bacteria isolated from fish intestine were unable to be identified using MALDI-TOF MS but
gram staining identified the bacteria as Bacillus gram positive.
5.2.2 Characterization of isolates using 16S rRNA and gyrB gene sequencing
Figure 6 is a gel image showing the PCR product amplified with the expected size. The
BLAST results revealed that Isolate-2, Isolate-4, and F-9 were of a Bacillus amyloliquefaciens
strain, summarize in Table 4. The partial 16S rNA gene sequence of Isolate-2 had a 99%
identity with B.amyloliquefaciens strain IHB B 3373 (GeneBank accession # KF475869.1) and a
98% identity with B.amyloliquefaciens subsp. plantarum str. FZB42 (GeneBank accession #
NC_009725.1) for its gyrB gene sequence. The partial 16S rNA gene sequence of Isolate-4 had a
99% identity with B.amyloliquefaciens strain BAB-68 16S (GeneBank accession # KC250107.1)
and a 95% identity with B.amyloliquefaciens subsp. plantarum str. FZB42 (GeneBank accession
# NC_009725.1) for its gyrB gene sequence. Isolate-9 had a 99% identity with
B.amyloliquefaciens strain IHB B 3373 (GeneBank accession # KF475869.1) and a 98% identity
with B.amyloliquefaciens subsp. plantarum str. FZB42 (GeneBank accession # NC_009725.1)

52
for its partial 16S rRNA and gyrB gene sequences, respectively. F-11’s gene sequences had a
99% identity to B.amyloliquefaciens supsp. plantarum strain UCMB5033 (GeneBank Accession
# HG328253.1) and a 97% identity to B.amyloliquefaciens subsp. plantarum str. FZB42
(GeneBank Accession # NC_009725.1) to partial 16S rRNA and gyrB sequences, respectively.
Isolate-7, Isolate-8, and F-8 were all of a Bacillus subtilis strain, summarized in Table 4. Isolate-
7’s gene sequences had a 99% identity to B.subtilis strain GLMP6 (GeneBank Accession #
KF364495.1) and a 98% identity to B.subtilis subsp. subtilis str. 168 (GeneBank Accession #
NC_000964.3) to partial 16S rRNA and gyrB sequences, respectively. Isolate-8’s gene
sequences had a 99% identity to B.subtilis strain YLB-P1 (GeneBank Accession # KF220577.1)
and a 97% identity to B.subtilis subsp. subtilis str. 168 (GeneBank Accession # NC_000964.3) to
partial 16S rRNA and gyrB sequences, respectively. F-8’s gene sequences had a 99% identity to
B.subtilis strain IHB B 4270(GeneBank Accession # KF475879.1) and a 97% identity to
B.subtilis subsp. subtilis str. 168 (GeneBank Accession # NC_000964.3) to partial 16S rRNA
and gyrB sequences, respectively.

53
A) 1 2 3 4 5 6 7
B)
Figure 7
Figure 6: Gel electrophoresis of PCR products used to identify the bacteria isolates. (A) The
partial sequence of 16S rRNA gene product. PCR products were electrophoresed on a 1.0%
agarose gel and visualized under UV light. Low molecular weight DNA ladder (New England
Labs, U.K) was used to confirm expected size of product (500 kp). (B) The gyrB gene product.
1 kb DNA Plus Ladder (Life Technologies, U.S.A) was used to confirm expected size of product
(1,200 bp).1=Isolate-2, 2= Isolate-4, 3= Isolate-7; 4= Isolate-8; 5= F-8, 6= F-9, and 7= F-11
500
1 2 3 4 5 6 7
1650 1000
500
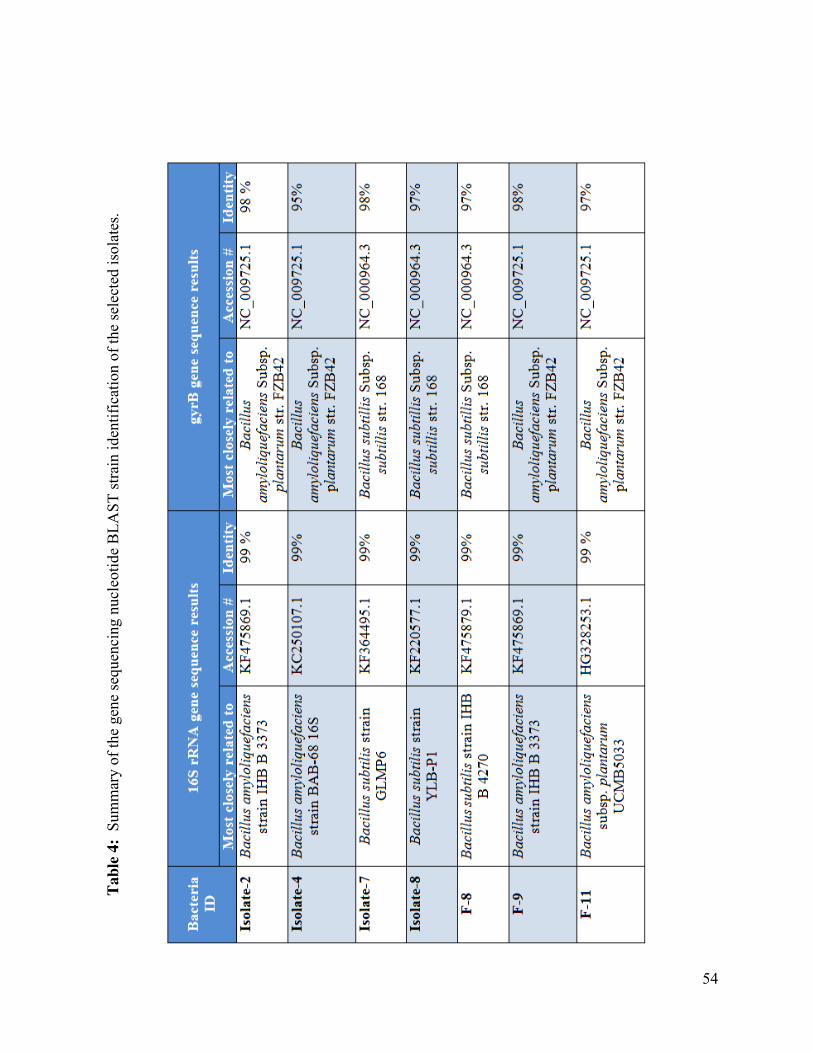
54
Tabl
e 4:
Sum
mar
y of
the
gene
sequ
enci
ng n
ucle
otid
e B
LAST
stra
in id
entif
icat
ion
of th
e se
lect
ed is
olat
es.

55
5.3 Liquid-state fermentation
5.3.1 Influence of selected bacterium on SBM protein profile
Figure 7B is a representative image of the results. It was observed that the abundance of
high molecular weight proteins had decreased, and the abundance of low molecular weight
proteins increased, with fermentation. The large allergenic subunits of β-conglycinin (72 kDa,
76 kDa and 53 kDa) were no longer visible in the gel, suggesting the allergenic proteins have
been reduced. Of all the isolates, Isolate-2 was determined to be the best isolate in terms of
reducing high molecular weight proteins, increasing low molecular weight proteins, and reducing
the allergenic proteins.
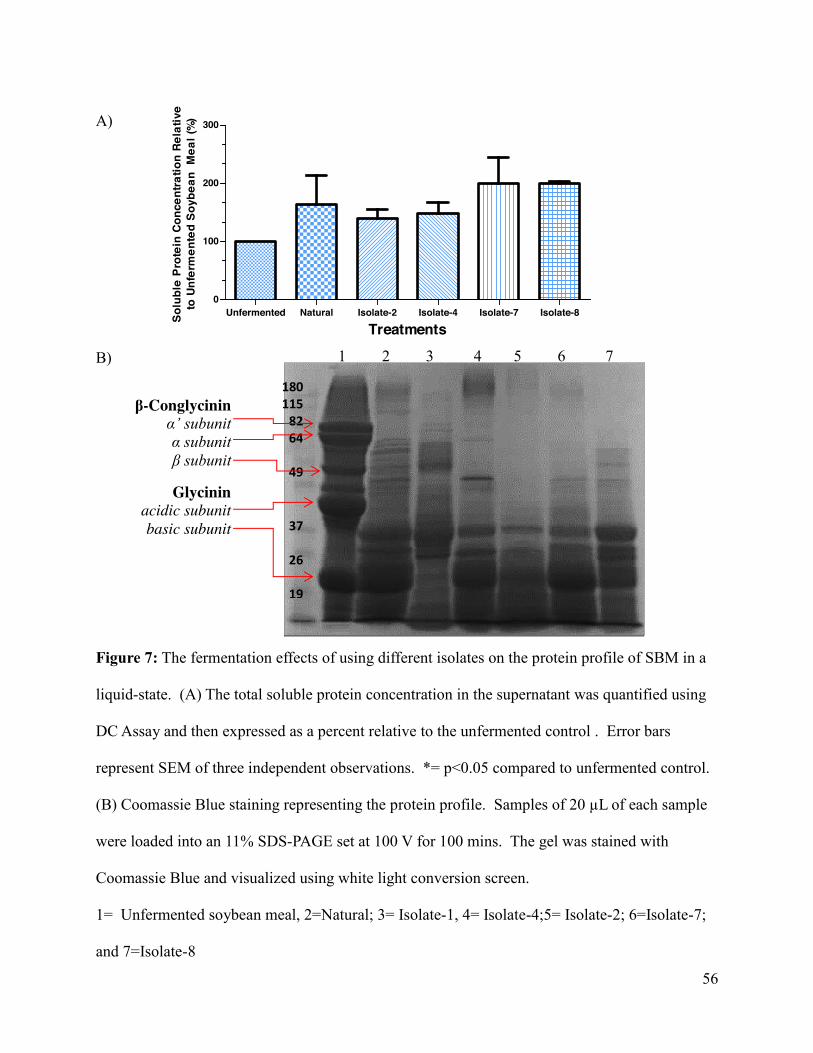
56
A)
B)
Figure 7: The fermentation effects of using different isolates on the protein profile of SBM in a
liquid-state. (A) The total soluble protein concentration in the supernatant was quantified using
DC Assay and then expressed as a percent relative to the unfermented control . Error bars
represent SEM of three independent observations. *= p<0.05 compared to unfermented control.
(B) Coomassie Blue staining representing the protein profile. Samples of 20 µL of each sample
were loaded into an 11% SDS-PAGE set at 100 V for 100 mins. The gel was stained with
Coomassie Blue and visualized using white light conversion screen.
1= Unfermented soybean meal, 2=Natural; 3= Isolate-1, 4= Isolate-4;5= Isolate-2; 6=Isolate-7;
and 7=Isolate-8
1 2 3 4 5 6 7
180 115
82 64
49
37
26
19
β-Conglycinin α’ subunit α subunit β subunit
Glycinin acidic subunit basic subunit
Treatments
Solu
ble
Prot
ein
Conc
entra
tion
Rela
tive
to U
nfer
men
ted
Soyb
ean
Mea
l (%
)
Unfermented Natural Isolate-2 Isolate-4 Isolate-7 Isolate-80
100
200
300

57
5.4 Solid-state fermentation
5.4.1 Influence of selected bacterium on SBM protein profile
After obtaining the encouraging result from liquid-state fermentation, we next examined
the feasibility of the more industry-applicable solid-state fermentation of SBM. Soybean meal
was fermented at a liquid: substrate ratio of 2:1 at 42 °C for 48 hours, with sampling at 24 and 48
hours. Total soluble protein concentrations were increased in five fermentations, compared to
the unfermented SBM control, as shown in Figure 8. No effect of fermentation time on total
soluble protein levels (Figure 9) of the same isolate were observed except for Isolate-2 and
Isolate-4. The total soluble protein of soybean meal fermented with Isolate-2 appears to lower at
48 hours as compared to 24 hours. The opposite effect is observed with soybean meal fermented
with Isolate-4 which was increased at 48 hours when compared to 24 hours. In addition,
fermentation with the selected isolates was able to decrease the high molecular weight proteins
while increasing the low molecular weight proteins (Figure 9). This was different among
bacteria isolated from food rather than from the intestine of the grass carp. In general, bacteria
isolated from food effectively reduced most proteins to approximately less than 25 kDa (Figure
9A). Most of the bacteria isolated from food demonstrated maximal SBM peptide degradation at
24 hours of fermentation. Bacteria isolated from the intestine of the grass carp were able to
degrade the proteins to less than 35 kDa (Figure 9B). This is 10 kDa higher than that fermented
by bacteria isolated from the food sources. Similar to the bacteria isolated from fermented food,
no real improvement in peptide profile was observed after the 24 hour fermentation. The
fermentations from both sources were also successful in reducing the allergenic proteins of β-
conglycinin (72 kDa, 76 kDa and 53 kDa). Of all the bacteria used for fermentation, Isolate-2
was the isolate that degraded most of the high molecular proteins and increase most of the low

58
weight proteins at the 24 hour time point, and thus was selected and further tested to investigate
changes in crude protein and amino acid profiles.
5.4.2 Improved crude protein of fermented soybean meal
Once the most efficient isolate was identified, its ability to improve soybean meal
nutrient composition was examined. The best isolate to improve the peptide profile visualized
using Coomassie Blue gel was Isolate-2, a bacterium isolated from food source. The lyophilized
ground fermented product of Isolate-2 and the unfermented product were tested for crude protein
and amino acid profile changes. It was determined that fermentation with Isolate-2 significantly
increased crude protein concentrations from 48.1 ± 0.5% to 53.3 ± 0.67% (24 hours) and 55.9±
0.9% (48 hours), respectively (Figure 10; Table 5), although the difference is not significant
between 24 hours and 48 hours.
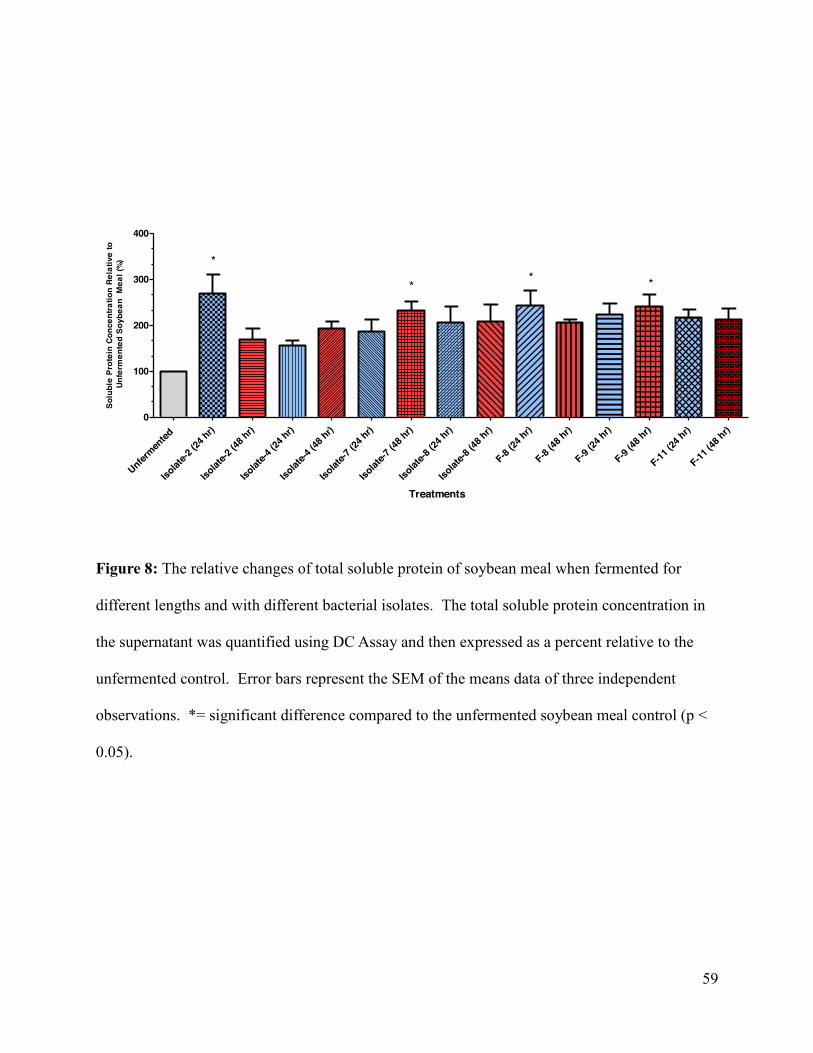
59
Figure 8
Treatments
Solu
ble
Prot
ein
Conc
entra
tion
Rela
tive
to U
nfer
men
ted
Soyb
ean
Mea
l (%
)
Unfermen
ted
Isolat
e-2 (2
4 hr)
Isolat
e-2 (4
8 hr)
Isolat
e-4 (2
4 hr)
Isolat
e-4 (4
8 hr)
Isolat
e-7 (2
4 hr)
Isolat
e-7 (4
8 hr)
Isolat
e-8 (2
4 hr)
Isolat
e-8 (4
8 hr)
F-8 (24
hr)
F-8 (48
hr)
F-9 (24
hr)
F-9 (48
hr)
F-11 (2
4 hr)
F-11 (4
8 hr)
0
100
200
300
400
*
** *
Figure 8: The relative changes of total soluble protein of soybean meal when fermented for
different lengths and with different bacterial isolates. The total soluble protein concentration in
the supernatant was quantified using DC Assay and then expressed as a percent relative to the
unfermented control. Error bars represent the SEM of the means data of three independent
observations. *= significant difference compared to the unfermented soybean meal control (p <
0.05).
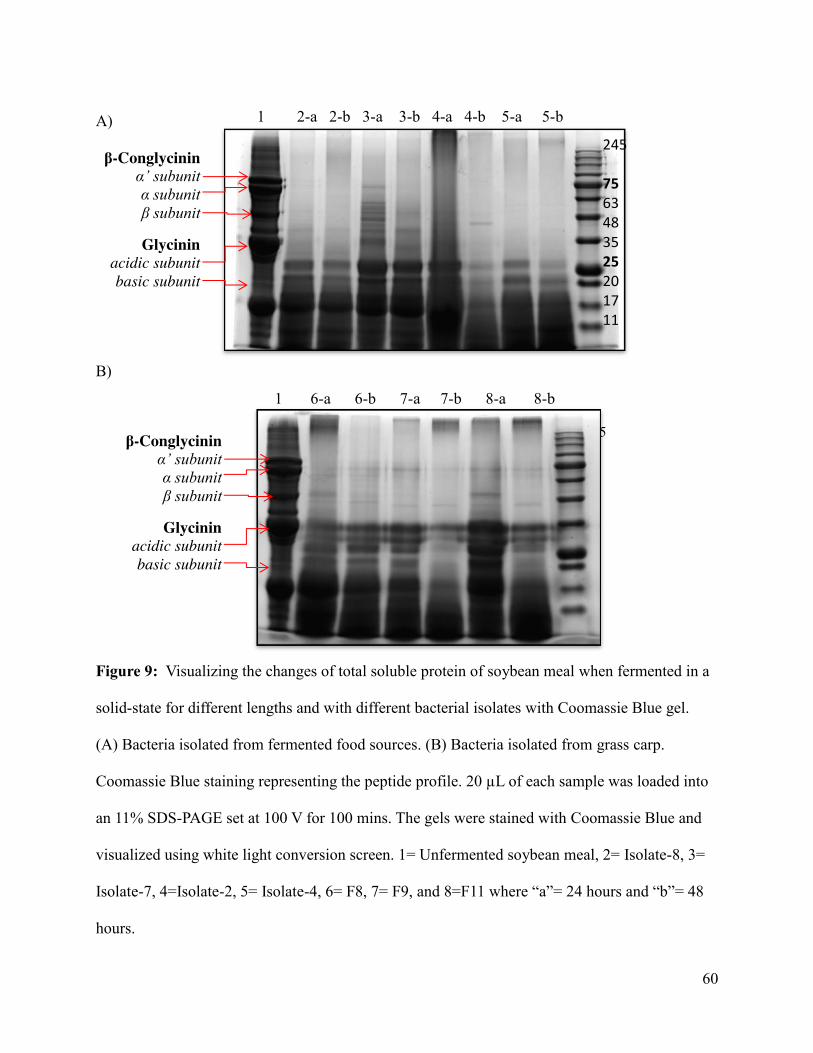
60
Figure 9
A)
B)
Figure 9: Visualizing the changes of total soluble protein of soybean meal when fermented in a
solid-state for different lengths and with different bacterial isolates with Coomassie Blue gel.
(A) Bacteria isolated from fermented food sources. (B) Bacteria isolated from grass carp.
Coomassie Blue staining representing the peptide profile. 20 µL of each sample was loaded into
an 11% SDS-PAGE set at 100 V for 100 mins. The gels were stained with Coomassie Blue and
visualized using white light conversion screen. 1= Unfermented soybean meal, 2= Isolate-8, 3=
Isolate-7, 4=Isolate-2, 5= Isolate-4, 6= F8, 7= F9, and 8=F11 where “a”= 24 hours and “b”= 48
hours.
245 75 63 48 35 25 20 17 11
1 2-a 2-b 3-a 3-b 4-a 4-b 5-a 5-b
1 6-a 6-b 7-a 7-b 8-a 8-b
245 75 63 48 35 25 20 17 11
β-Conglycinin α’ subunit α subunit β subunit
Glycinin acidic subunit basic subunit
β-Conglycinin α’ subunit α subunit β subunit
Glycinin acidic subunit basic subunit
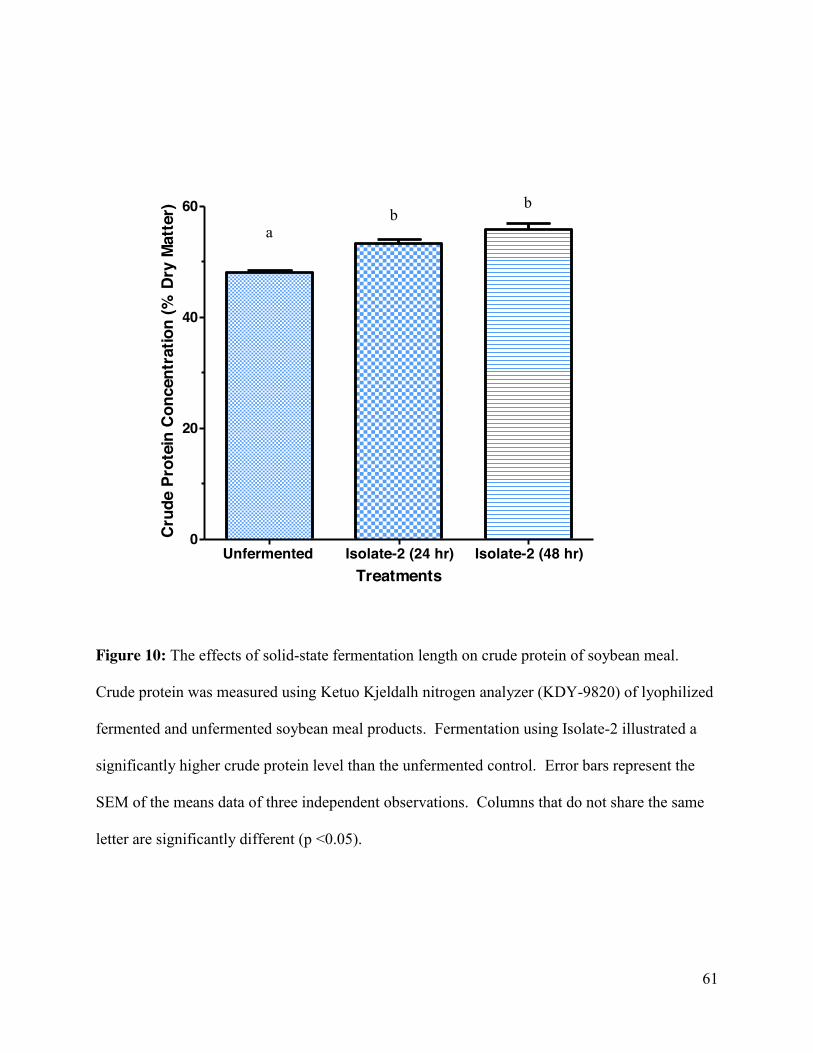
61
Figure 10Treatments
Cru
de P
rote
in C
once
ntra
tion
(% D
ry M
atte
r)
Unfermented Isolate-2 (24 hr) Isolate-2 (48 hr)0
20
40
60
Figure 10: The effects of solid-state fermentation length on crude protein of soybean meal.
Crude protein was measured using Ketuo Kjeldalh nitrogen analyzer (KDY-9820) of lyophilized
fermented and unfermented soybean meal products. Fermentation using Isolate-2 illustrated a
significantly higher crude protein level than the unfermented control. Error bars represent the
SEM of the means data of three independent observations. Columns that do not share the same
letter are significantly different (p <0.05).
a b
b

62
5.4.3 Amino acid profile of the fermented soybean meal
As shown in Figure 11, at 24 hours, both the total essential (Figure 11 A) and total non-
essential (Figure 11 B) amino acids concentrations were significantly increased by fermentation
using Isolate-2, when compared to unfermented control. By 48 hours of fermentation, the
essential amino acid (EAA) profile decreased to the same level of the control whereas the non-
essential amino acids remained unchanged. Table 5 depicts the change detected in all the amino
acids when fermented with Isolate-2 for 24 and 48 hours, respectively. With the exception of
arginine, all the essential amino acids analysed show a numerical increase at 24 hour of
fermentation when compared to the unfermented control, with the changes in valine and
phenylalanine being the only amino acids that increased statically significantly (p<0.05). At 48
hours of fermentation, the concentration of phenylalanine and histidine were significantly higher,
but concentrations of arginine were significantly lower than the unfermented control. Of the
eight non-essential amino acids analysed, levels of aspartic acid, glutamic acid, cysteine,
tyrosine, and proline were found to be significantly from than the unfermented control. Levels
of aspartic acid, glycine, and proline significantly increased at 24 hours of fermentation, but
declined back to the same level of unfermented control at 48 hours. Levels of glutamic acid
were significantly higher than the unfermented control at 24 and 48 hours of fermentation.
Levels of cysteine and tyrosine increased with fermentation to a significantly different level than
the control at 48 hours. The total amino acid concentration in soybean meal was significantly
increased by fermentation at 24 hour, but not at 48 hours (Table 5).
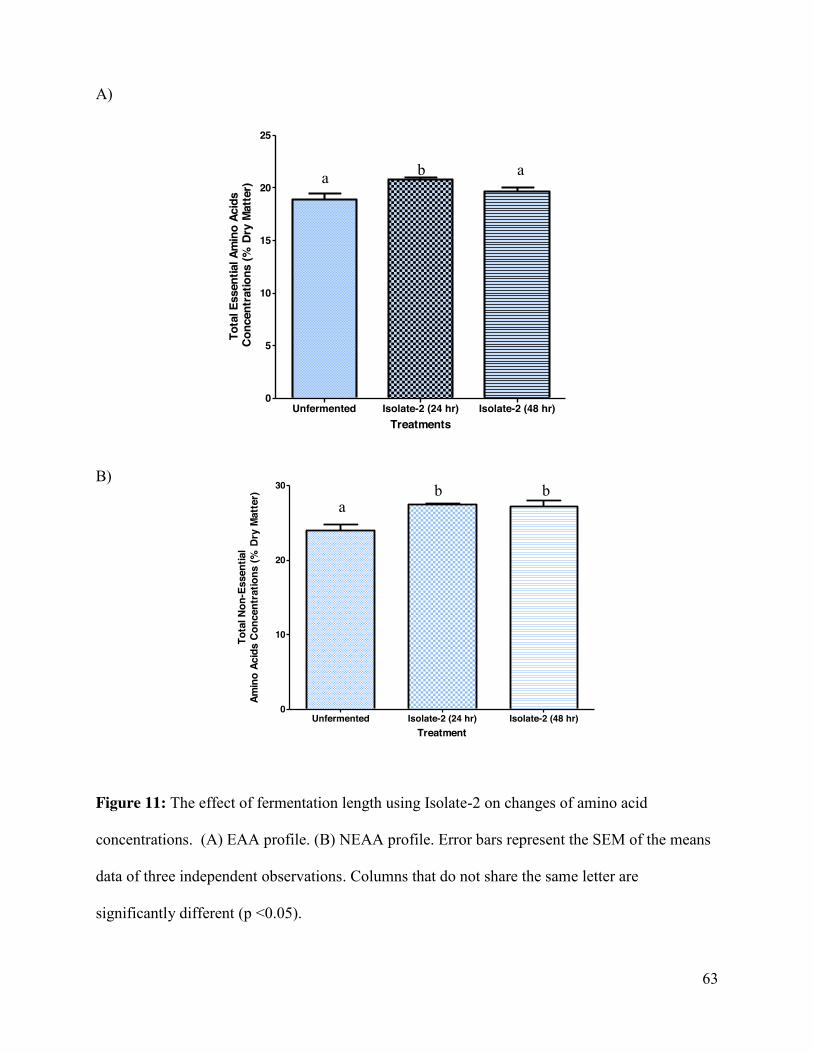
63
A)igure 11
Treatments
Tota
l Ess
entia
l Am
ino
Acid
s C
once
ntra
tions
(% D
ry M
atte
r)
Unfermented Isolate-2 (24 hr) Isolate-2 (48 hr)0
5
10
15
20
25
B)
Figure 11: The effect of fermentation length using Isolate-2 on changes of amino acid
concentrations. (A) EAA profile. (B) NEAA profile. Error bars represent the SEM of the means
data of three independent observations. Columns that do not share the same letter are
significantly different (p <0.05).
Treatment
Tota
l Non
-Ess
entia
lAm
ino
Acid
s C
once
ntra
tions
(% D
ry M
atte
r)
Unfermented Isolate-2 (24 hr) Isolate-2 (48 hr)0
10
20
30
b a
a
b b a
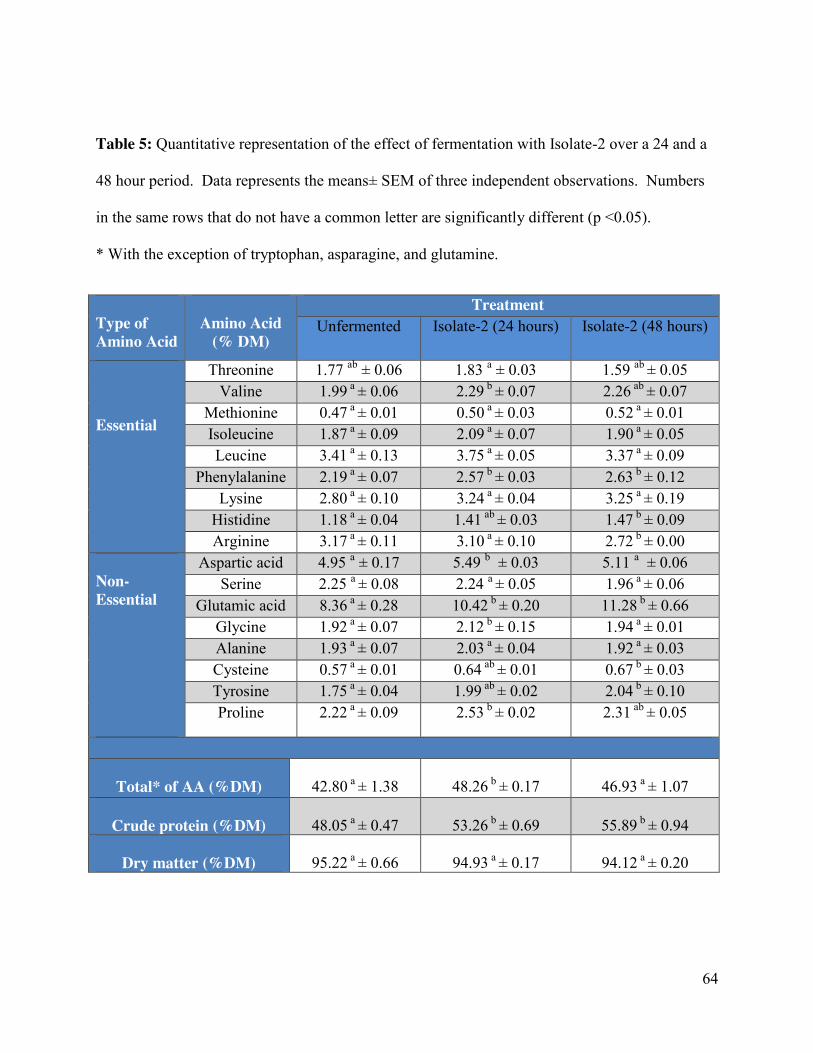
64
Table 5: Quantitative representation of the effect of fermentation with Isolate-2 over a 24 and a
48 hour period. Data represents the means± SEM of three independent observations. Numbers
in the same rows that do not have a common letter are significantly different (p <0.05).
* With the exception of tryptophan, asparagine, and glutamine.
Type of Amino Acid
Amino Acid
(% DM)
Treatment Unfermented Isolate-2 (24 hours) Isolate-2 (48 hours)
Essential
Threonine 1.77 ab ± 0.06 1.83 a ± 0.03 1.59 ab ± 0.05 Valine 1.99 a ± 0.06 2.29 b ± 0.07 2.26 ab ± 0.07
Methionine 0.47 a ± 0.01 0.50 a ± 0.03 0.52 a ± 0.01 Isoleucine 1.87 a ± 0.09 2.09 a ± 0.07 1.90 a ± 0.05 Leucine 3.41 a ± 0.13 3.75 a ± 0.05 3.37 a ± 0.09
Phenylalanine 2.19 a ± 0.07 2.57 b ± 0.03 2.63 b ± 0.12 Lysine 2.80 a ± 0.10 3.24 a ± 0.04 3.25 a ± 0.19
Histidine 1.18 a ± 0.04 1.41 ab ± 0.03 1.47 b ± 0.09 Arginine 3.17 a ± 0.11 3.10 a ± 0.10 2.72 b ± 0.00
Non-Essential
Aspartic acid 4.95 a ± 0.17 5.49 b ± 0.03 5.11 a ± 0.06 Serine 2.25 a ± 0.08 2.24 a ± 0.05 1.96 a ± 0.06
Glutamic acid 8.36 a ± 0.28 10.42 b ± 0.20 11.28 b ± 0.66 Glycine 1.92 a ± 0.07 2.12 b ± 0.15 1.94 a ± 0.01 Alanine 1.93 a ± 0.07 2.03 a ± 0.04 1.92 a ± 0.03 Cysteine 0.57 a ± 0.01 0.64 ab ± 0.01 0.67 b ± 0.03 Tyrosine 1.75 a ± 0.04 1.99 ab ± 0.02 2.04 b ± 0.10 Proline 2.22 a ± 0.09 2.53 b ± 0.02 2.31 ab ± 0.05
Total* of AA (%DM)
42.80 a ± 1.38
48.26 b ± 0.17
46.93 a ± 1.07
Crude protein (%DM)
48.05 a ± 0.47
53.26 b ± 0.69
55.89 b ± 0.94
Dry matter (%DM)
95.22 a ± 0.66
94.93 a ± 0.17
94.12 a ± 0.20

65
5.4.4 Detection of Allergenic Proteins
The results of the SDS-PAGE suggest that the subunits of the soy allergens β-conglycinin
(76, 72, and 53 kDa) and glycinin (35 and 22 kDa) were decreased or eliminated by fermentation
with the different isolates. To validate that the allergens in fermented soybean meal did
decrease, a western blot was performed. The results of the western blot indicated that antibodies
found within the piglet plasma were able to bind to three distinct proteins found in the
unfermented soybean meal product (Figure 12). These three distinct soy proteins were found
approximately at: 75 kDa, 40-50 kDa, and 30-40 kDa. At 24 hours soybean meal fermented with
Isolate-2 or F-9 showed no reactivity to the piglet antibodies. However, piglet antibodies reacted
to a peptide of 37 kDa at 24 hours with Isolate-4, F-8, or F-11 (Figure 12A). By 48 hour of
fermentation, no allergen reactive band was detected (Figure 12B).
5.4.5 Oligosaccharide concentration of fermented soybean meal
Two major problematic soy oligosaccharides for piglets are raffinose and stachyose. As
shown in Table 6, the concentration of stachyose and raffinose in the lyophilized unfermented
soybean meal were 41.73 mg/g and 9.88 mg/g, respectively. Starting from 24 hours,
fermentation with Isolate-2 reduced the amount of both oligosaccharides in SBM to non-
detectable level (Table 6).
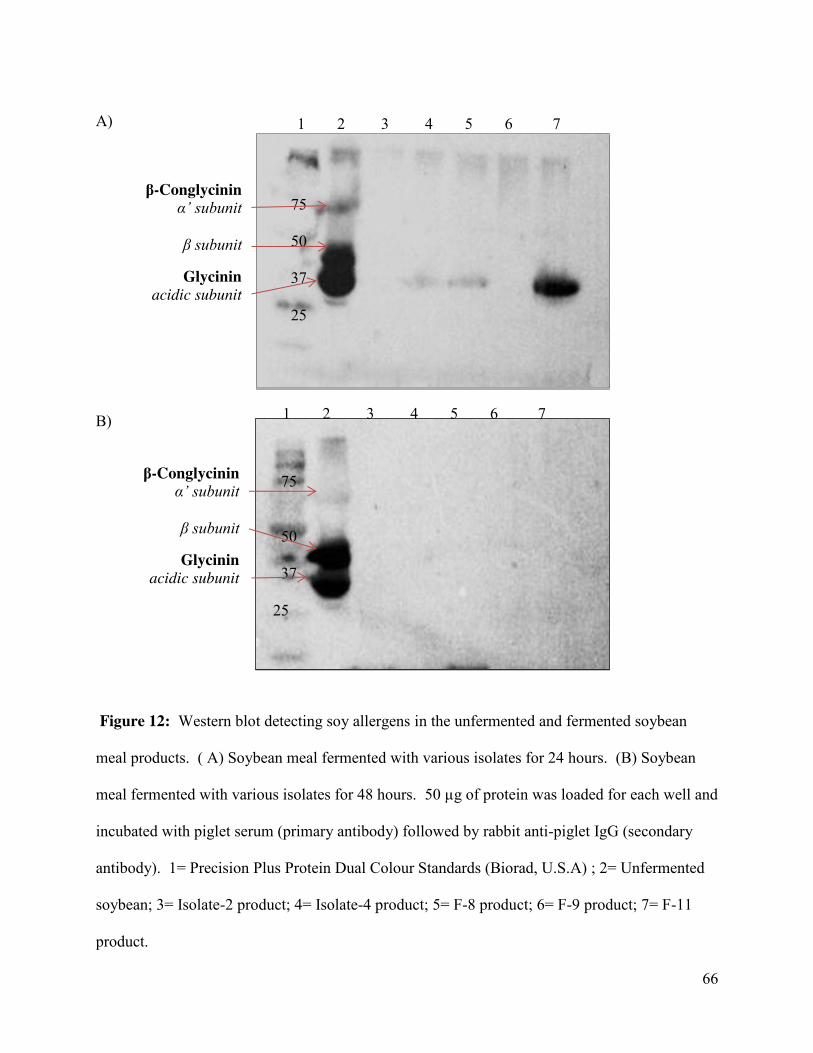
66
Figure 12
A)
B)
Figure 12: Western blot detecting soy allergens in the unfermented and fermented soybean
meal products. ( A) Soybean meal fermented with various isolates for 24 hours. (B) Soybean
meal fermented with various isolates for 48 hours. 50 µg of protein was loaded for each well and
incubated with piglet serum (primary antibody) followed by rabbit anti-piglet IgG (secondary
antibody). 1= Precision Plus Protein Dual Colour Standards (Biorad, U.S.A) ; 2= Unfermented
soybean; 3= Isolate-2 product; 4= Isolate-4 product; 5= F-8 product; 6= F-9 product; 7= F-11
product.
75
50
37
25
75
50
37
25
β-Conglycinin α’ subunit
β subunit
Glycinin acidic subunit
β-Conglycinin α’ subunit
β subunit
Glycinin acidic subunit
1 2 3 4 5 6 7
1 2 3 4 5 6 7
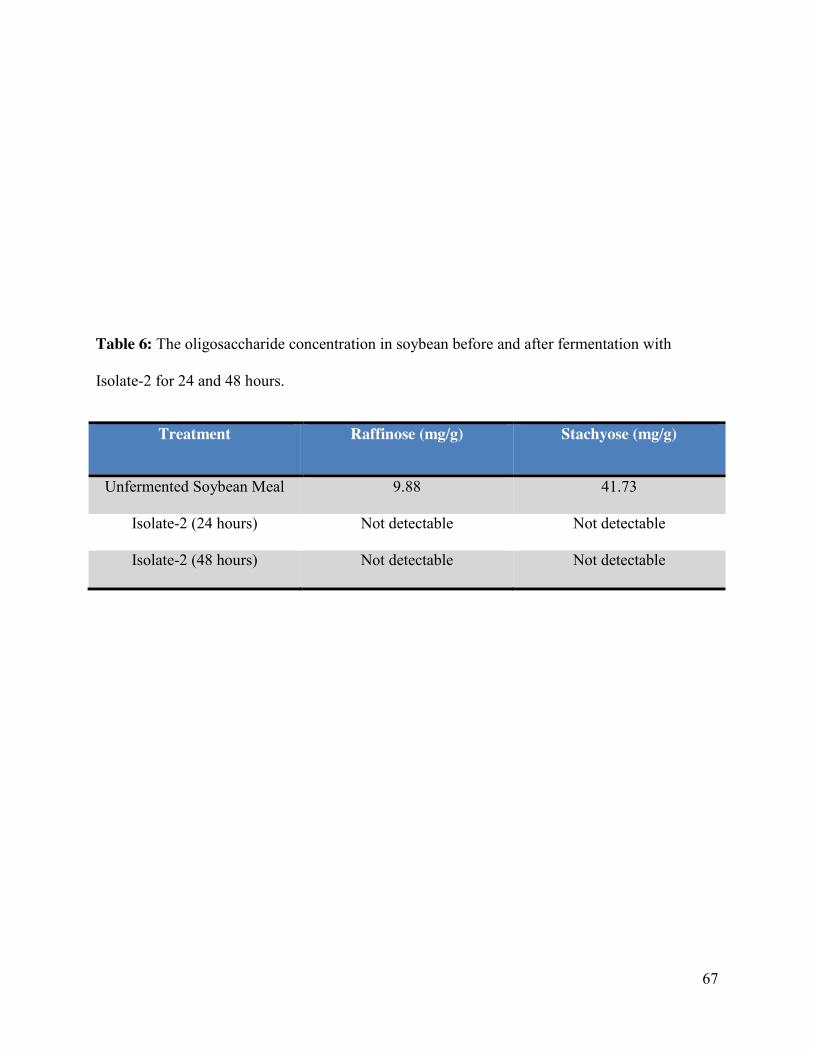
67
Table 6: The oligosaccharide concentration in soybean before and after fermentation with
Isolate-2 for 24 and 48 hours.
Treatment Raffinose (mg/g) Stachyose (mg/g)
Unfermented Soybean Meal 9.88 41.73
Isolate-2 (24 hours) Not detectable Not detectable
Isolate-2 (48 hours) Not detectable Not detectable

68
6.0 DISCUSSION
Fermenting soybean meal with bacteria isolated from fermented food sources, or from the
intestine of grass carp, was found to be successful in improving the nutritional value of soybean
meal. The eight candidate bacteria selected for fermentation of soybean meal were able to
improve the nutrient value of soybean meal through the reduction of allergenic and other high
molecular weight proteins, and an increase in low-weight proteins. Of these eight candidate
fermentations, Isolate-2 was considered to be the best fermented product and further analysis
indicated that it also improved the amino acid profile, and decreased the concentration of soy
oligosaccharides stachyose and raffinose. Six of the eight bacteria isolated from traditional
fermented food sources, successfully demonstrated protease activity. The two colonies (Isolate-3
and Isolate-5) that did not demonstrate protease activity were considered to be contaminating
Isolate-2 and Isolate-4 and thus, were also selected for second round screening. Traditional
Chinese fermented soy foods undergo natural spontaneous fermentation (Han et al., 2001),
which suggests that the bacteria isolated from these sources break down soybean components to
utilizable nutrients. Another benefit to using bacteria derived from fermented foods and intestine
is the safety of the bacteria. Bacteria isolated from fermented foods are considered to be safe as
they are consumed by humans and may contain other beneficial properties such as being
probiotic or having antimicrobial activity (Alexopolilos et al., 2004; Kim et al., 2004). Bacteria
isolated from fish intestines may also have probiotic potential. Fish fed probiotic B.subtilis,
previously isolated from the intestines of a carp fish, were found to have significantly higher
lengths and weights and were also found to have significantly higher survival rates when
challenged with Aeromonas hydrophila compared to that of the fish not fed the probiotic (Ghost
et al., 2008). Since this study did not investigate the antimicrobial or probiotic qualities of our

69
isolates, we cannot conclude that our isolates are antimicrobial or probiotic. Further studies
should be performed to determine if any of our isolates share any of these probiotic properties.
The bacteria isolated from this study were identified using common molecular tools such
as MALDI-TOF MS and genetic sequencing. The MALDI- TOF MS results of this study
indicated all the bacteria are of the Bacillus species. This is consistent with other studies that
isolated bacteria from other traditionally fermented foods such as Dawadawa (Amoa-Awua et
al., 2006), Meju (Cho et al., 2003), Cheonggukjang (Baek et al., 2009), and Douchi (Peng et al.,
2003). These bacteria can survive on legumes because they have the ability to secrete
extracellular enzymes- such as proteases (Cho et al., 2003). MALDI-TOF MS uses bacterial
protein to identify the bacteria and matches the peaks from the mass spectrometry to find its
closest match in the database. However, there are some limitations of the assay. First, in order
for the unknown isolate to have a match, the suspect species would have to be previously
characterized and the profile saved in the database (Hotta et al., 2013). Secondly, the database
must be accurate or else identification will not be reliable (Hotta et al., 2013). Lastly, there
might be some issues with post-translational modification that could alter the peaks which could
incorrectly identify the bacteria (Hotta et al., 2013). It is thus important to confirm the MALDI-
TOF MS identification by other methods, such as 16S rRNA gene sequencing used in this study.
The results of the partial 16S rRNA gene sequencing revealed the Isolate-2, Isolate-4, and F-9
were Bacillus amyloloquefaciens; and Isolate-7, Isolate-8, F-8, and F-11 were Bacillus subtilis.
Although partial 16S RNA gene sequencing can identify many bacterial species, it is more
reliable to sequence the complete 16S rRNA gene, and some other housekeeping gene such as
the gyrB gene. The gyrB gene is responsible for producing the β subunit protein of a DNA

70
gyrase and is one of the genes essential in DNA replication (Wang et al., 2007). This gene is
distributed universally among species and has a faster rate of molecular evolution than the 16S
rRNA gene, thus it has been used to identify different strains of bacteria of the same species
(Wang et al., 2007). Using the gyrB sequence results, we were able to further identify Isolate-2,
Isolate-4, F-9 and F-11 as Bacillus amyloliquefaciens; and Isolate-7, Isolate-8, and F-8 as
Bacillus subtilis. It is interesting to note that the same strain of bacteria were found in both
fermented food and fish intestines, but yet have different enzyme efficiencies. The source from
which bacteria is isolated is very important and can determine which enzymes the bacteria will
secrete to survive. Isolate-2 was one of the eight bacteria we isolated from traditional fermented
food. Given the source, we expected the bacteria to demonstrate high protease activity. This is
important because the bacteria will need to break down the plant’s high molecular weight
proteins in order to better utilize them for their growth. Grass carp is known to be a herbivore
but can utilize both animal and plant sources for nutrients (Zhou et al., 2013). The fish itself
does not produce cellulase but have been known to have microbes in the intestine that can secrete
various enzymes, including cellulase (Li et al., 2009; Zhoa et al.,2013). It was also found that
supplying an exogenous source of cellulase can alter the microbe population in the intestine
which can increase cellulase production ( Zhoa et al.,2013). Given this source we expected the
bacteria we isolated (F-8, F-9, and F-11) to demonstrate the ability to break down cellulose in
order to survive. There has been a study which has successfully isolated cellulase producing
bacteria from the intestine of the grass carp (Li et al., 2009) and thus, we expect NSP
concentrations to also decrease in our fermentation. The change in the NSP concentration of
soybean meal after fermentation with Isolate-2 will be investigated in future experiments.

71
Of the entire Bacillus genus, Bacillus subtilis is the most commonly used in soybean
fermentations. Previous studies have observed an improvement on the total soluble protein
profiles, elimination of allergenic proteins, and a reduction of anti-nutritional factors when
soybean has been fermented with various strains Bacillus subtilis (Wongputtisin et al., 2014;
Ying et al; 2009; Frias et al., 2007). These effects were due the bacteria’s ability to produce
extracellular enzymes which can digest the large proteins into smaller proteins and peptides
(Frias et al., 2007). This species of bacteria has also been found to increase the antioxidant
capacity of soybeans by (Wongputtisin et al., 2007). Many strains of Bacillus subtilis have been
found to have antimicrobial and probiotic effects (Kyriakis et al., 1999; Alexopolilos et al.,
2004; Guo et al., 2006).
Bacillus amyloliquefaciens has been less investigated in fermentation of soybean
compared to Bacillus subtilis. To our knowledge, only one study has investigated Bacillus
amyloliquefaciens in soybean fermentation (Chistyakov et al., 2015). This study was performed
using the spores of the bacteria to ferment the whole soybean as a target dietary supplement for
use in poultry feeds (Chistyakov et al., 2015). The results of the study indicate that the birds fed
the treatment had an 8% higher growth rate and an 8.8% higher feed efficiency when compared
to the control birds (Chistyakov et al., 2015). This species of bacteria has been also found to
have found to have anti-bacteria activity against a strain of E.coli (Chi et al., 2014) and
Vibrionaceae species (Xu et al., 2014) that were both found to be resistant to multiple types of
antibiotics. This anti-microbial activity was attributed to the lipopeptides produced by the
bacteria which destroyed the membrane and whole cells of the pathogens, acting like porins,
causing the cells to rupture (Chi et al., 2014; Xu et al., 2014). The species has also been found

72
to have anti-fungal properties (Song et al., 2013) and produce phytase (Shim & Oh, 2012).
Again, it is important to keep in mind the source of which bacteria is isolated in order to
determine the safety of the bacteria for consumption. Bacteria of the same species can exhibit
different characteristics depending on where it is isolated from, in order to adapt to its
environment. Isolate-2, the bacteria isolated from fermented food and chosen for further
fermentation of soybean meal, has been identified as Bacillus amyloliquefaciens. A strain of
Bacillus amyloliquefaciens isolated from soil was found to have beneficial traits such as being a
probiotic strain (Islam et al., 2011), but other strains isolated from other sources such as the
inside of a moisture damaged building may secrete substances that can be toxic to both humans
and animals (Mikkola et al., 2004).
Preliminary data from the liquid state fermentation shows that the majority of the large
soybean proteins were successfully cleaved to less than 37 kDa, using the newly isolated bacteria
with the exception of Isolate-1. The protein concentration of all the fermented products was
improved compared to the unfermented control. Soy flour fermented by Lactobacillus
plantarum in a liquid state was found to decrease most of its proteins to less than 15 kDa (Frias
et al., 2008). The particle size of the substrate was found to have an influence on these results,
as the same bacteria fermented in a liquid state, using cracked soybean as the substrate, was
found to degrade proteins to less than 40 kDa (Frias et al., 2008). The reduction of particle size
increases the surface area which provides the bacteria with more products to utilize (Frias et al.,
2008). Fermenting soybean in a liquid-state solubilizes the soy proteins which make it easier for
the bacteria to access and thus, hydrolyze using extracellular enzymes (Frias et al., 2008).

73
Although this study and other studies provide evidence that there are benefits to using
liquid-state fermentation, the solid-state application is more desirable to the agricultural industry
setting due to less costly equipment and lower moisture levels (Singhania et al., 2009; Opazo et
al., 2012). Lower moisture content of fermented soybean meal reduces the amount of drying
necessary to eliminate the risk of mould growth (Opazo et al., 2012), and the cost of
transportation. To compensate for fermentations with higher moisture ratios, a previous study
has oven-dried their fermentation product to a moisture level of 10% (Chen et al., 2010). This
extra processing step further increases the cost and the risk of Maillard reactions, negatively
affecting lysine, which effectively decreases the nutrient bioavailability of the feed (Friedman &
Brandon, 2001; Gonzalez-Vega et al., 2011).
Our solid-state fermentation was also successful in improving soybean meal protein
content. Interestingly, the bacteria isolated from fermented soy foods cleaved most of the soy
protein to 27 kDa and under, while fermentation with bacteria isolated from the intestines of the
grass carp, resulted in majority of the soybean protein being degraded to 35 kDa and less. It is
possible that different species and subspecies bacteria may secrete distinct arrays of proteinases
that target different subsets of soybean proteins. The protein distribution patterns are comparable
between 24 and 48 hours of fermentation using Isolate-2 and F-6, respectively. This suggests
that 24 hours is sufficient to achieve maximal protein degradation. Similar peptide profiles have
also been previously observed. Cracked soybeans fermented with A.oryzae for 48 hours resulted
in majority of soybean protein that were 50 kDa or smaller (Frias et al., 2008). In the same
study, cracked soybean fermented with B.subtilis for 48 hours was observed to decrease soybean
proteins to less than 20 kDa, which appears to be more of an improvement than using A.oryzae

74
(Frias et al., 2008). Soybean meal fermented with A.oryzae for 48 hours was found to have
13.4% of its proteins concentrated at 20-60 kDa, while the rest of the proteins were less than 20
kDa (Hong et al., 2004). The reduction of protein sizes is an important aspect to increase the
digestibility of soybean protein as small peptides can be more easily hydrolyzed to free amino
acids that can be absorbed in the intestine (Silk et al., 1979).
The results of the piglet immunoreactivity of the unfermented soybean meal indicated
three distinct soy proteins at: 75 kDa, 40-50 kDa, and 30-40 kDa. These bands would
correspond to the α’ (76 kDa) and β (53 kDa) subunits of β-conglycinin and the acidic (35-40
kDa) subunit of glycinin. The α(72 kDa) subunit of β-conglycinin and the basic (22 kDa)
subunit of glycinin were not visible on the western blot. The western blot as well as the SDS-
PAGE indicated that Isolate-2 was successful at reducing and completely eliminating all subunits
of both allergens. Allergenic proteins found in soybean meal activate the immune system
leading to an inflammation of the intestine, releasing chemical such as histamine, which
increases the incidences of diarrhea (Hao et al., 2009; Zhao et al., 2010). Additionally, they
have been known to impede digestion (Cervantes-Pahm & Stein, 2010). β-conglycinin has also
been found to directly affect piglet intestinal cells in vitro by decreasing cell growth, damaging
the cytoskeleton, and stimulating apoptosis of the intestinal cells (Chen et al., 2010). Thus,
removal of these allergenic soybean proteins is beneficial for both digestion and animal health.
Soybean meal manufactures use a quality control measurement called protein
dispersibility index which should be around 40% (Soy 20/20, 2008). This indicates that at least
40% of the protein found in soybean meal should be recovered when suspended in water. The

75
allergenic proteins focussed in this paper are β-conglycinin and glycinin and each of its subunits
has different degrees of solubility (Zheng et al., 2009). The solubility of the subunits can vary
depending on pH or temperature (Zheng et al., 2009). The subunits of β-conglycinin were found
to have 50% solubility in water at pH equal to or greater than 6 (Zheng et al., 2009). The
subunits of glycinin were found to have a solubility of approximately 100% at pH greater than
6.5 at 20 °C at varying ionic strengths (Lakemond et al., 2000). This thesis only investigated the
changes in the soluble protein when fermented with our isolates and did not measure the
reduction of the insoluble allergenic proteins. Although it was not measured, it cannot be
concluded that the insoluble portion of the allergenic proteins were not also reduced, and thus,
future studies should investigate the changes in the insoluble protein or total protein when
soybean meal is fermented with Isolate-2.
The solid-state fermentation with Isolate-2 significantly increased the crude protein by
5.21 % and 7.84% in 24 hour and 48 hours, respectively. This level of increase is within the
range of crude protein increased using A.oryzae. Soybean meal fermented with A. oryzae has
been reported to have crude protein increased ranging from 1.95 % (Chen et al., 2007) to 10%
(Hong et al., 2004). The increase in crude protein can be a result of two processes. Firstly, the
increase crude protein observed can be due in part to the decrease in carbohydrates (Hong et al.,
2004). Some bacteria can break down cellulose, polysaccharides, and oligosaccharides and
utilize its sugar subunits for their metabolism and respiration processes (Opazo et al., 2012). As
crude protein is a percentage of dry matter, any reduction in dry matter would lead to an increase
in percentage of crude protein. Decrease in dry matter content could also be due to the required
gaseous (carbon dioxide) losses during fermentation. Our finding that Isolate-2 has cellulase
activity is consistent with this possibility. Another aspect of fermentation that would cause an
increase in crude protein, is the production of proteins by the bacteria. Many Bacillus species,

76
including Bacillus amyloliquefaciens, have been found to produce a polymer of glutamic acid
called poly-ɣ-glutamate (PGA) (Feng et al., 2014). Our finding of glutamic acid level increase
in the fermented soybean meal may be a result of PGA production. PGA is biodegradable,
edible, and is safe for humans (Shih & Van, 2001). It can be found in many traditional
fermented foods such as the Japanese fermented food natto (Kada et al., 2012), has high
antioxidant activity, as well as play a role in the immune response (Lee et al., 2014; Lee et al.,
2011). It interacts with toll-like receptors to decrease pathological symptoms caused by airway
asthma (Lee et al., 2011), as well as reduce inflammation caused by angiogenesis and mucosal
inflammation observed in irritated bowel disorder (Sung et al., 2013). It would be interesting to
further investigate if PGA concentration is indeed increased in soybean meal during
fermentation.
The amino acid profile was also improved in the solid-state fermentation with Isolate-2.
Of the 17 amino acids analyzed, levels of valine, phenylalanine, aspartic acid, glutamic acid,
glycine, and proline were significantly higher than the unfermented soybean meal at 24 hours of
fermentation. At 48 hours, levels of histidine, cysteine, and tyrosine increased from a numerical
change (24 hours) to significance; however the levels of valine, aspartic acid, glycine and proline
decline and were no longer statistically significantly different than the unfermented control. The
increase of selective amino acids was also observed in other studies as well. Hong et al. (2004)
observed a significant increase in glycine, glutamic acid, and aspartic acid when soybean meal
was fermented with A. oryzae. Another study using A.oryzae observed similar results with levels
of amino acids threonine and glutamine only significantly increasing when Lb. casei was added
(Chen et al., 2007). On the other hand, using cracked soybean fermented with A.oryzae reported

77
significant increases in all amino acids except arginine and lysine (Frias et al., 2008). Similarly,
cracked soybeans fermented with B.subtilis increased aspartic acid, glutamic acid, serine,
alanine, proline, valine, methionine, cysteine, isoleucine, phenylalanine, tyrosine and lysine
levels after 48 hours (Frias et al., 2008). This increase in amino acids may be in part due to the
protein constituents of bacterial mass and microbial metabolism (Chen et al., 2010). The results
of the current study also showed that levels of arginine decrease at 48 hours of fermentation.
Our result on the increase of total essential amino acid in 24 but not 48 hours (which has an
increase in nonessential amino acid only) of fermentation indicates that bacteria at different
phases of the fermentation may utilize and synthesize a different array of amino acids. This data
again suggests that the 24 hour time point is not only more cost effective, but also desirable for
the fermentation when compared to the 48 hours. The length of the fermentation is an important
factor to consider at the production scale. In an agricultural production facility, like any other
production facility, the manufacturing time of the product may at times be a limiting factor. The
time to manufacture a product may dictate the required fermentation capacity and how quickly
the product can be sold, which influences the profit of the product. The faster a product can be
made the more potential revenue could be had. The length of the fermentation can be greatly
affected by choice of inoculum. The fermentation of soybean into traditional soy sauce can be
greatly reduced from 7 days to 40 hours by using selected strains of A.oryzae which secrete high
amounts of extracellular enzymes (Valyasevi & Rolle, 2002). Other published soybean
fermentation systems have reported fermentation lengths of 40 hours (Chen et al., 2010), 48
hours (Hong et al., 2004, Frias et al., 2008), 72 hours (Lee et al., 2010) or 10 days (Opazo et al
2012). Our fermentation with the newly isolate bacteria has shortened the required length to
only 24 hours in order to obtain the desired protein and amino acid profile.

78
Moisture level is another important factor when designing a fermentation system. Too
little moisture will not allow the bacteria to grow and survive, yet a moisture level too high may
result in undesirable mould growth. Another advantage to our solid-state system is our ratio of
2:1 (weight of liquid: weight of substrate). Previous studies have reported ratios such as
2.4:1(Opazo et al., 2012), 3.5:1 (Refstie et al., 2005), and 10:1 (Rodrigues et al., 2010) which are
all (slightly) higher than our ratio. By optimizing fermentation conditions to have a low moisture
level, we decrease the risk of mould growing and also make it more efficient for the fermented
soybean meal to be inoculated animal feed. Many animal feeds, including swine and poultry
feed, are processed into pellets. The pelleting process itself is very delicate and if moisture level
of the ingredients is too high the pellets will not be strong enough to withstand the transport to
the farm. Our solid-state fermented soybean meal product contains a lower moisture level than
other previous studies, and thus, requires less drying time. Decreasing any additional heating
required for processing also decreases the risk of further heat damage to the proteins.
The results of this study demonstrate that the protein profile, in terms of decreasing
allergenic proteins, decreasing high molecular weight proteins, increasing low molecular weight
proteins, and increasing total amino acids, can be improved by fermenting soybean meal using
our Isolate-2, we were also able to observe a decrease in the oligosaccharides. Our results
indicate that raffinose and stachyose levels decreased to non-detectable levels in soybean meal
after fermentation with Isolate-2. The oligosaccharides in soy are indigestible to piglets and
have been found to cause a decrease in piglet performances (Sohn et al., 1994). Increased levels
of oligosaccharides have been found to negatively affect nutrient and enzyme interaction, as well
as decrease the digestibility of dry matter in the ileum (Smits & Annison, 1996; Van Kempen et
al., 2006). Studies have reported a decrease in soy oligosaccharides, stachyose and raffinose,

79
and NSPs when soybean meal was fermented with bacteria (Refstie et al.,2005; Opazo et al.,
2012) although a much longer term (10 days) fermentation was required. The reduction of these
oligosaccharides could have been due to hydrolysis of the α-1,6-galactose linkage by the enzyme
α-1,6-galactosidase. Some strains of Bacillus have been found to secrete extracellular α-1,6-
galactose (Lee & Cho, 2012) which may explain the reduction of oligosaccharides observed.
This study investigates the fermentation of soybean meal using single bacterium. Other
studies have investigated the benefits of adding more than one microbe to soybean meal
fermentation. It was observed that when a combination of B.subtilis, Lb. fermentum, and
Saccharomyces cerevisae were used to ferment soybean meal with wheat bran, it could reduce
the growth of Enterobacteriaceae in vivo (Ying et al., 2009). The authors attributed this
reduction of pathogenic bacteria to the antimicrobial proteins produced by their strain of
B.subtilis and to the secondary metabolites produced by the lactic acid bacteria which inhibits
spoilage (Ying et al., 2009). Viable Lactobacillus cells and Bacillus spores were found after
fermentation and have led the authors to conclude that this new method of fermentation could be
an alternative to using antibiotics in the animal industry (Ying et al., 2009). Another study
investigated the effects of three newly isolated strains of cellulolytic bacteria isolated from
various sources on soybean meal fermentation (Opazo et al., 2012). The soybean meal was
fermented with 1×109 cells of inoculum (a mixture of bacteria from
Streptomyces, Cohnella and Cellulosimicrobium genuses) /g of substrate for 10 days in 37 °C
under 2.4:1 (weight of water: weight of substrate) with constant agitation. The fermentation
reduced stachyose levels to 0.44 grams/ 100 grams of soybean meal and raffinose levels to 0.48
grams/100 grams of soybean meal compared to that of that of the non-inoculated control groups

80
which were 3.52 grams and 2.11 grams, respectively (Opazo et al., 2012). NSP concentrations
were also reduced to 12.17 grams/ 100 grams of soybean meal compared to the 16.01 grams/100
grams of soybean meal found in the non-inoculated group (Opazo et al., 2012). There have also
been studies exploring the possibility of using a two-step fermentation to observe the dual
benefits of using A.oryzae and Lb.casei in soybean meal fermentation (Chen et al., 2010). The
two-step fermentation, total fermentation time of 40 hours at 30 °C (first-step), and 37 °C
(second-step), successfully decreased soy oligosaccharides to non-detectable levels, increasing
essential amino acids by 9.5%, and decreasing most allergenic proteins, compared to that of the
unfermented control (Chen et al., 2010). Three-step fermentation of soybeans using Lactococcus
lactis, A.orzyae, and B.subtilis was found to have a higher total amino acid concentration than
when fermented with B.subtilis alone (Lee et al. 2007). This improvement in fermentation is due
to the mixture of enzymes secreted by each type bacteria (Lee et al., 2007). Some microbes,
such as A.oryzae, prefer select amino acids, which may result in only certain amino acids being
increased (Hong et al., 2004). Using a combination of bacteria in soybean meal fermentation
may result in the increase in different amino acids, resulting in a larger increase of total amino
acids (Lee et al., 2007). Including a lactic acid bacteria (LAB) in soybean fermentation may add
additional benefits such as increasing amino acid concentration and decreasing the pH of the
fermented product (Lee et al., 2007). In traditional fermentations, many LABs are used as a
starter culture to inhibit the growth of any pathogenic microbes (Leroy& De Vuyst, 2004).
Using LAB in soybean fermentation may also be beneficial to early-weaned pigs. Feeding piglet
metabolites of LAB have demonstrated to reduce incidences of diarrhoea, decreased faecal pH
and pathogenic bacteria, and increase short chain fatty acids (Thu et al., 2011). It would be of
interest to further investigate if combination of other microbes with Isolate-2 can further improve
fermentation of soybean meal.

81
In conclusion, this in vitro solid-state fermentation using newly isolated, food derived
bacteria, resulted in an improved protein profile, an increase in crude protein concentration, an
increase in total amino acid concentration, and a decrease in raffinose and stachyose. This
fermentation system achieved effective fermentation after 24 hours and a relatively low moisture
level. A follow-up study should investigate if further optimization of fermentation conditions
results in further improvements in nutritional value of fermented soybean meal product and to
compare these to commercial or other experimental fermented products. Decreasing the
fermentation length would allow decrease of facilities running cost and product turnover time.
Also, the low moisture level of our system reduces drying needs. The improvement in all of these
aspects could potentially lead to a plant protein source that is less expensive to produce, highly
digestible, and allergenic-free for piglets and other animal species, in order to reduce some of the
negative performance observed during early weaning due to feeding soybean meal. Further
studies should investigate the NSP profiles, as well as the antioxidant profile of the fermented
soybean meal product. Isolate-2 should be further investigated to determine if there are any toxic
effects to mammalian cells and if there is an advantage of combining different a LAB with
Isolate-2 to ferment soybean meal.

82
REFERENCES:
Alexopoulos, C., Georgoulakis, I. E., Tzivara, A., Kyriakis, C. S., Govaris, A., & Kyriakis, S. C. (2004). Field Evaluation of the Effect of a Probiotic‐containing Bacillus licheniformis and Bacillus subtilis Spores on the Health Status, Performance, and Carcass Quality of Grower and Finisher Pigs. Journal of Veterinary Medicine Series A, 51(6), 306-312.
Amadou, I., LE, G. W., SHI, Y. H., Gbadamosi, O. S., Kamara, M. T., & Jin, S. (2011). Optimized Lactobacillus plantarum LP6 solid-state fermentation and proteolytic hydrolysis improve some nutritional attributes of soybean protein meal. Journal of Food Biochemistry, 35(6), 1686-1694.
American Soybean Association (2004) Edited by :Van Eys, J.E., Offner, A.,& Bach, A. Chemical Analysis. Manual of Quality Analysis for Soybean Products in the Feed Industry. http//www.asa-europe. org/ Library/ library_e.htm.
Amoa-Awua, W. K., Terlabie, N. N., & Sakyi-Dawson, E. (2006). Screening of 42 Bacillus isolates for ability to ferment soybeans into dawadawa.International journal of food microbiology, 106(3), 343-347.
Anderson, R. L., & Wolf, W. J. (1995). Compositional changes in trypsin inhibitors, phytic acid, saponins and isoflavones related to soybean processing. The Journal of nutrition, 125(3 Suppl), 581S-588S.
Angkanaporn, K., Choct, M., Bryden, W. L., Annison, E. F., & Annison, G. (1994). Effects of wheat pentosans on endogenous amino acid losses in chickens. Journal of the Science of Food and Agriculture, 66(3), 399-404.
AOAC Official Method 934.01 (1990) Official Methods of Analysis of AOAC International, 15th Ed., AOAC International, Gaithersburg, MD
AOAC Official Method 954.01 (1990) Official Methods of Analysis of AOAC International, 15th Ed., AOAC International, Gaithersburg, MD
AOAC Official Method 994.12 (1990) Official Methods of Analysis of AOAC International, 15th Ed., AOAC International, Gaithersburg, MD
Bach Knudsen, K. E., Hedemann, M. S., & Lærke, H. N. (2012). The role of carbohydrates in intestinal health of pigs. Animal Feed Science and Technology, 173(1-2), 41-53
Baek, J. G., Shim, S. M., Kwon, D. Y., Choi, H. K., Lee, C. H., & Kim, Y. S. (2009). Metabolite profiling of Cheonggukjang, a fermented soybean paste, inoculated with various Bacillus strains during fermentation. Bioscience, biotechnology, and biochemistry, 74(9), 1860-1868.
Baker, K. M., & Stein, H. H. (2009). Amino acid digestibility and concentration of digestible and metabolizable energy in soybean meal produced from conventional, high-protein, or low-oligosaccharide varieties of soybeans and fed to growing pigs. Journal of animal science, 87(7), 2282-2290.

83
Beardslee, T. A., Zeece, M. G., Sarath, G., & Markwell, J. P. (2000). Soybean glycinin G1 acidic chain shares IgE epitopes with peanut allergen Ara h 3.International archives of allergy and immunology, 123(4), 299-307.
Birk, Y. (1985). The Bowman‐Birk inhibitor. Trypsin‐and chymotrypsin‐inhibitor from soybeans. International journal of peptide and protein research, 25(2), 113-131.
Böhme, K., Fernández‐No, I. C., Pazos, M., Gallardo, J. M., Barros‐Velázquez, J., Cañas, B., & Calo‐Mata, P. (2013). Identification and classification of seafood‐borne pathogenic and spoilage bacteria: 16S rRNA sequencing versus MALDI‐TOF MS fingerprinting. Electrophoresis,34(6), 877-887.
Bureau, D. P., Harris, A. M., & Cho, C. Y. (1998). The effects of purified alcohol extracts from soy products on feed intake and growth of chinook salmon (Oncorhynchus tshawytscha) and rainbow trout (Oncorhynchus mykiss). Aquaculture, 161(1), 27-43
Cai, H., Archambault, M., & Prescott, J. F. (2003). 16S Ribosomal RNA Sequence—Based Identification of Veterinary Clinical Bacteria. Journal of veterinary diagnostic investigation, 15(5), 465-469.
Canadian International Grain Institute (2010). Soybean Feed Industry Guide, 1st Ed. Soy20/20 Project. Accessed: http://cigi.ca/wp-content/uploads/2011/12/2010-Soybean-Feed-Industry-Guide.pdf
Carbonnelle, E., Mesquita, C., Bille, E., Day, N., Dauphin, B., Beretti, J. L., & Nassif, X. (2011). MALDI-TOF mass spectrometry tools for bacterial identification in clinical microbiology laboratory. Clinical biochemistry, 44(1), 104-109.
Cervantes-Pahm, S. K., & Stein, H. H. (2010). Ileal digestibility of amino acids in conventional, fermented, and enzyme-treated soybean meal and in soy protein isolate, fish meal, and casein fed to weanling pigs. Journal of animal science, 88(8), 2674-2683.
Chantawannakul, P., Oncharoen, A., Klanbut, K., Chukeatirote, E., & Lumyong, S. (2002). Characterization of proteases of Bacillus subtilis strain 38 isolated from traditionally fermented soybean in Northern Thailand.Science Asia, 28(4), 241-245.
Chattopadhyay, S., Taub, F., Paul, S., Weissman, S. J., & Sokurenko, E. V. (2013). Microbial variome database: Point mutations, adaptive or not, in bacterial core genomes. Molecular Biology and Evolution, 30(6), 1465-1470
Chen, C. C., Chiou, P. W. S., & Yu, B. (2010). Evaluating nutritional quality of single stage-and two stage-fermented soybean meal. Asian-Australasian Journal of Animal Sciences, 23(5), 598
Chen, F., Hao, Y., Piao, X. S., Ma, X., Wu, G. Y., Qiao, S. Y., Li, D.F., & Wang, J. J. (2011). Soybean-derived β-conglycinin affects proteome expression in pig intestinal cells in vivo and in vitro. Journal of animal science, 89(3), 743-753.
Chi, Z., Rong, Y. J., Li, Y., Tang, M. J., & Chi, Z. M. (2014). Biosurfactins production by Bacillus amyloliquefaciens R3 and their antibacterial activity against multi-drug resistant pathogenic E. coli. Bioprocess and biosystems engineering, 1-9.

84
Cho, J. H., Min, B. J., Chen, Y. J., Yoo, J. S., Wang, Q., Kim, J. D., & Kim, I. H. (2007). Evaluation of FSP (fermented soy protein) to replace soybean meal in weaned pigs: growth performance, blood urea nitrogen and total protein concentrations in serum and nutrient digestibility. Asian Australasian Journal of Animal Sciences, 20(12), 1874.
Cho, S. J., Oh, S. H., Pridmore, R. D., Juillerat, M. A., & Lee, C. H. (2003). Purification and characterization of proteases from Bacillus amyloliquefaciens isolated from traditional soybean fermentation starter. Journal of agricultural and food chemistry, 51(26), 7664-7670.
Choct, M., Dersjant-Li, Y., McLeish, J., & Peisker, M. (2010). Soy oligosaccharides and soluble non-starch polysaccharides: a review of digestion, nutritive and anti-nutritive effects in pigs and poultry. Asian-Australasian journal of animal sciences, 23(10), 1386-1398.
Chistyakov, V., Meliknikov, V., Chikindas, M. L., Khutsishvili, M., Chagelishvili, A., Bren, A., & ElisashviliI, V. (2014). Poultry-beneficial Solid-state Bacillus amyloliquefaciens B-1895 Fermented Soybean Formulation. Bioscience of Microbiota, Food and Health.
Clarke, E. J., & Wiseman, J. (2000). Developments in plant breeding for improved nutritional quality of soya beans I. Protein and amino acid content. The Journal of Agricultural Science, 134(02), 111-124.
Cranwell, P. D., Noakes, D. E., & Hill, K. J. (1976). Gastric secretion and fermentation in the suckling pig. British Journal of Nutrition, 36(01), 71-86.
Csaky, I., & Fekete, S. (2004). Soybean: feed quality and safety. Part 1: biologically active components. A review. Acta Veterinaria Hungarica, 52(3), 299-313.
De Angelis, M., Siragusa, S., Berloco, M., Caputo, L., Settanni, L., Alfonsi, G., Amerio, M., Grandi, A., Ragni, A., & Gobbetti, M. (2006). Selection of potential probiotic lactobacilli from pig feces to be used as additives in pelleted feeding. Research in Microbiology,157(8), 792-801
Deacon, J., (2005). Fungal nutrition. In Fungal Biology (Chapter 6). Retrieved from http://archive.bio.ed.ac.uk/jdeacon/FungalBiology/chap6.htm#Fig6.6
Dias, D. R., Vilela, D. M., Silvestre, M. P. C., & Schwan, R. F. (2008). Alkaline protease from bacillus sp. isolated from coffee bean grown on cheese whey. World Journal of Microbiology & Biotechnology, 24(10), 2027-2034.
Dierking, E. C., & Bilyeu, K. D. (2009). Raffinose and stachyose metabolism are not required for efficient soybean seed germination. Journal of Plant Physiology, 166(12), 1329-1335
Doyle, J. J., Schuler, M. A., Godette, W. D., Zenger, V., Beachy, R. N., & Slightom, J. L. (1986). The glycosylated seed storage proteins of Glycine max and Phaseolus vulgaris. Structural homologies of genes and proteins. Journal of Biological Chemistry, 261(20), 9228-9238.
EFSA (European Food Safety Authority)( 2013). Scientific Opinion on the safety and efficacy of of Bacillus amyloliquefaciens (NCIMB 30229) as a silage feed additive for all species.

85
EFSA Journal (11)1:3042 Accessed from: http://www.efsa.europa.eu/en/scdocs/scdoc/543.htm
Fabek, H., Messerschmidt, S., Brulport, V., & Goff, H. D. (2014). The effect of in vitro digestive processes on the viscosity of dietary fibres and their influence on glucose diffusion. Food Hydrocolloids, 35(1), 718-726.
Farina, H. G., Pomies, M., Alonso, D. F., & Gomez, D. E. (2006). Antitumor and antiangiogenic activity of soy isoflavone genistein in mouse models of melanoma and breast cancer. Oncology reports, 16(4), 885-891.
Feng, J., Liu, X., Xu, Z. R., Wang, Y. Z., & Liu, J. X. (2007a). Effects of fermented soybean meal on digestive enzyme activities and intestinal morphology in broilers. Poultry science, 86(6), 1149-1154.
Feng, J., Liu, X., Xu, Z. R., Lu, Y. P., & Liu, Y. Y. (2007b). The effect of Aspergillus oryzae fermented soybean meal on growth performance, digestibility of dietary components and activities of intestinal enzymes in weaned piglets. Animal feed science and technology, 134(3), 295-303.
Feng, J., Gao, W., Gu, Y., Zhang, W., Cao, M., Song, C., & Wang, S. (2014). Functions of poly- gamma-glutamic acid (γ-PGA) degradation genes in γ-PGA synthesis and cell morphology maintenance. Applied microbiology and biotechnology, 1-11.
Florencio, C., Couri, S., & Farinas, C. S. (2012). Correlation between agar plate screening and solid-state fermentation for the prediction of cellulase production by Trichoderma strains. Enzyme research, 2012.
Freire, J. P. B., Guerreiro, A. J. G., Cunha, L. F., & Aumaitre, A. (2000). Effect of dietary fibre source on total tract digestibility, caecum volatile fatty acids and digestive transit time in the weaned piglet. Animal Feed Science and Technology, 87(1), 71-83.
Frias, J., Song, Y. S., Martínez-Villaluenga, C., De Mejia, E. G., & Vidal-Valverde, C. (2007).
Immunoreactivity and amino acid content of fermented soybean products. Journal of agricultural and food chemistry, 56(1), 99-105.
Friedman, M., Brandon, D. L., Bates, A. H., & Hymowitz, T. (1991). Comparison of a commercial soybean cultivar and an isoline lacking the Kunitz trypsin inhibitor: composition, nutritional value, and effects of heating. Journal of Agricultural and Food Chemistry, 39(2), 327-335.
Friedman, M.& Brandon, D. L. (2001). Review: Nutritional and health benefits of soy proteins. J. Agric. Food Chem. 49, 1069–1086
Friesen, K. G., Nelssen, J. L., Goodband, R. D., Behnke, K. C., & Kats, L. J. (1993). The effect of moist extrusion of soy products on growth performance and nutrient utilization in the early-weaned pig. Journal of animal science,71(8), 2099-2109.

86
Fritz, H., Seely, D., Flower, G., Skidmore, B., Fernandes, R., Vadeboncoeur, S., & Fergusson, D. (2013). Soy, Red Clover, and Isoflavones and Breast Cancer: A Systematic Review. PloS one, 8(11).
Gebru, E., Lee, J. S., Son, J. C., Yang, S. Y., Shin, S. A., Kim, B., & Park, S. C. (2010). Effect of
probiotic-, bacteriophage-, or organic acid-supplemented feeds or fermented soybean meal on the growth performance, acute-phase response, and bacterial shedding of grower pigs challenged with Salmonella enterica serotype Typhimurium. Journal of animal science, 88(12), 3880-3886.
Gilbert, E. R., Wong, E. A., & Webb, K. E. (2008). Board-invited review: Peptide absorption
and utilization: Implications for animal nutrition and health. Journal of animal science, 86(9), 2135-2155.
González-Vega, J. C., Kim, B. G., Htoo, J. K., Lemme, A., & Stein, H. H. (2011). Amino acid
digestibility in heated soybean meal fed to growing pigs. Journal of animal science, 89(11), 3617-3625
Ghost, S., Sinha, A., & Sahu, C. (2008). Dietary probiotic supplementation in growth and health of live-bearing ornamental fishes. Aquaculture Nutrition, 14(4), 289-299.
Guo, X., Li, D., Lu, W., Piao, X., & Chen, X. (2006). Screening of Bacillus strains as potential probiotics and subsequent confirmation of the in vivo effectiveness of Bacillus subtilis MA139 in pigs. Antonie Van Leeuwenhoek,90(2), 139-146.
Han, B. Z., Rombouts, F. M., & Nout, M. J. (2001). A Chinese fermented soybean food. International Journal of Food Microbiology, 65(1), 1-10.
Hao, Y., Zhan, Z., Guo, P., Piao, X., & Li, D. (2009). Soybean β-conglycinin-induced gut hypersensitivity reaction in a piglet model. Archives of Animal Nutrition, 63(3), 188-202.
Hedemann, M., & Jensen, B. (2004). Variations in enzyme activity in stomach and pancreatic
tissue and digesta in piglets around weaning. Archives of Animal Nutrition, 58(1), 47-59 Hei, W., Li, Z., Ma, X., & He, P. (2012). Determination of beta-conglycinin in soybean and
soybean products using a sandwich enzyme-linked immunosorbent assay. Analytica Chimica Acta, 734, 62-68.
Held, S., & Mendl, M. (2001). Behaviour of the young weaned pig. Pages 273–297 in The
Weaner Pig—Nutrition and Management. M. A. Varley, and J. Wiseman, ed. CAB Int., Oxon, UK
Helm, R. M., Cockrell, G., Connaughton, C., Sampson, H. A., Bannon, G. A., Beilinson, V., &
Burks, A. W. (2000). A soybean G2 glycinin allergen. 2. Epitope mapping and three-dimensional modeling. International archives of allergy and immunology, 123(3), 213-219.

87
Hong, K. J., Lee, C. H., & Kim, S. W. (2004). Aspergillus oryzae GB-107 fermentation improves nutritional quality of food soybeans and feed soybean meals. Journal of medicinal food, 7(4), 430-435.
Hotta, Y., Sato, J., Sato, H., Hosoda, A., & Tamura, H. (2011). Classification of the genus
Bacillus based on MALDI-TOF MS analysis of ribosomal proteins coded in S10 and spc operons. Journal of agricultural and food chemistry, 59(10), 5222-5230.
Hu, Y., Ge, C., Yuan, W., Zhu, R., Zhang, W., Du, L., & Xue, J. (2010). Characterization of
fermented black soybean natto inoculated with Bacillus natto during fermentation. Journal of the Science of Food and Agriculture,90(7), 1194-1202.
Islam, V. H., Babu, N. P., Pandikumar, P., & Ignacimuthu, S. (2011). Isolation and
characterization of putative probiotic bacterial strain, Bacillus amyloliquefaciens, from North East Himalayan soil based on in vitro and in vivo functional properties. Probiotics and Antimicrobial Proteins, 3(3-4), 175-185.
Jamal, F., Pandey, P. K., Singh, D., & Khan, M. Y. (2013). Serine protease inhibitors in plants:
nature’s arsenal crafted for insect predators.Phytochemistry Reviews, 12(1), 1-34 Jongbloed, A. W., Mroz, Z., & Kemme, P. A. (1992). The effect of supplementary Aspergillus
niger phytase in diets for pigs on concentration and apparent digestibility of dry matter, total phosphorus, and phytic acid in different sections of the alimentary tract. Journal of Animal Science, 70(4), 1159-1168.
Kada, S., Ishikawa, A., Ohshima, Y., & Yoshida, K. I. (2012). Alkaline serine protease AprE
plays an essential role in poly-γ-glutamate production during natto fermentation. Bioscience, biotechnology, and biochemistry, 77(4), 802-809.
Karr-Lilienthal, L. K., Kadzere, C. T., Grieshop, C. M., & Fahey Jr, G. C. (2005). Chemical and
nutritional properties of soybean carbohydrates as related to nonruminants: A review. Livestock Production Science, 97(1), 1-12.
Kelly, D. & King T.P( 2001). Digestive physiology and development in pigs. Pages 179-206 in
The Weaner Pig Nutrition and Management. M. A. Varley and J. Wiseman, ed. CAB International, Wallingford, Oxon, UK
Kemme, P. A., Jongbloed, A. W., Mroz, Z., Kogut, J., & Beynen, A. C. (1999). Digestibility of
nutrients in growing–finishing pigs is affected by Aspergillus niger phytase, phytate and lactic acid levels: 1. Apparent ileal digestibility of amino acids. Livestock Production Science, 58(2), 107-117.
Kerr P.S, & Sebastian S,A. US Patent 6147193. Date Issued:November 14, 2000. Kim, J. C., Hansen, C. F., Mullan, B. P., & Pluske, J. R. (2012). Nutrition and pathology of
weaner pigs: Nutritional strategies to support barrier function in the gastrointestinal tract. Animal Feed Science and Technology,173(1-2), 3-16

88
Kim, S. W., Van Heugten, E., Ji, F., Lee, C. H., & Mateo, R. D. (2010). Fermented soybean meal as a vegetable protein source for nursery pigs: I. Effects on growth performance of nursery pigs. Journal of animal science,88(1), 214-224.
Kim, Y., Cho, J. Y., Kuk, J. H., Moon, J. H., Cho, J. I., Kim, Y. C., & Park, K. H. (2004).
Identification and antimicrobial activity of phenylacetic acid produced by Bacillus licheniformis isolated from fermented soybean, Chungkook-Jang. Current microbiology, 48(4), 312-317.
Klobasa, F., Werhahn, E., & Butler, J. E. (1987). Composition of sow milk during
lactation. Journal of Animal Science, 64(5), 1458-1466. Knudsen, D., Urán, P., Arnous, A., Koppe, W., & Frøkiær, H. (2007). Saponin-containing
subfractions of soybean molasses induce enteritis in the distal intestine of Atlantic salmon. Journal of agricultural and food chemistry, 55(6), 2261-2267.
Kuhn, G., Hennig, U., Kalbe, C., Rehfeldt, C., Ren, M. Q., Moors, S., & Degen, G. H. (2004).
Growth performance, carcass characteristics and bioavailability of isoflavones in pigs fed soy bean based diets. Archives of animal nutrition, 58(4), 265-276.
Kuipers, B. J., Alting, A. C., & Gruppen, H. (2007). Comparison of the aggregation behavior of
soy and bovine whey protein hydrolysates.Biotechnology advances, 25(6), 606-610. Kunst, F., Ogasawara, N., Moszer, I., Albertini, A. M., Alloni, G. O., Azevedo, V., & Haga, K.
(1997). The complete genome sequence of the gram-positive bacterium Bacillus subtilis. Nature, 390(6657), 249-256.
Kyriakis, S. C., Tsiloyiannis, V. K., Vlemmas, J., Sarris, K., Tsinas, A. C., Alexopoulos, C., &
Jansegers, L. (1999). The effect of probiotic LSP 122 on the control of post-weaning diarrhoea syndrome of piglets. Research in veterinary science, 67(3), 223-228.
Lallès, J. P., Bosi, P., Smidt, H., & Stokes, C. R. (2007). Weaning—a challenge to gut
physiologists. Livestock Science, 108(1), 82-93. Lakemond, C. M., de Jongh, H. H., Hessing, M., Gruppen, H., & Voragen, A. G. (2000). Soy
glycinin: influence of pH and ionic strength on solubility and molecular structure at ambient temperatures. Journal of Agricultural and Food Chemistry, 48(6), 1985-1990.
Larsen, N., Thorsen, L., Kpikpi, E., Stuer-Lauridsen, B., Cantor, M., Nielsen, B., . . . Jespersen,
L. (2014). Characterization of bacillus spp. strains for use as probiotic additives in pig feed. Applied Microbiology and Biotechnology, 98(3), 1105-1118
Lee, J. O., Park, M. H., Choi, Y. H., Ha, Y. L., & Ryu, C. H. (2007). New fermentation
technique for complete digestion of soybean protein. Journal of microbiology and biotechnology, 17(11), 1904-1907.

89
Lee, J., Park, I., & Cho, J. (2012). Production and partial characterization of alpha-galactosidase activity from an Antarctic bacterial isolate, Bacillus sp. LX-1. African Journal of Biotechnology, 11(60), 12396-12405.
Lee, K., Kim, S. ‐., Yoon, H. J., Paik, D. J., Kim, J. M., & Youn, J. (2011). Bacillus‐derived
poly‐γ‐glutamic acid attenuates allergic airway inflammation through a Toll‐like receptor‐4‐dependent pathway in a murine model of asthma. Clinical & Experimental Allergy, 41(8), 1143-1156. doi:10.1111/j.1365-2222.2011.03792.x
Lee, N. R., Lee, S. M., Cho, K. S., Jeong, S. Y., Hwang, D. Y., Kim, D. S., & Son, H. J. (2014).
Improved Production of Poly-γ-Glutamic Acid by Bacillus subtilis D7 Isolated from Doenjang, a Korean Traditional Fermented Food, and Its Antioxidant Activity. Applied biochemistry and biotechnology, 1-15.
Lenehan, N. A., DeRouchey, J. M., Goodband, R. D., Tokach, M. D., Dritz, S. S., Nelssen, J. L.,
& Lawrence, K. R. (2007). Evaluation of soy protein concentrates in nursery pig diets. Journal of animal science, 85(11), 3013-3021.
Leroy, F., & De Vuyst, L. (2004). Lactic acid bacteria as functional starter cultures for the food
fermentation industry. Trends in Food Science & Technology, 15(2), 67-78 Li, D. F., Nelssen, J. L., Reddy, P. G., Blecha, F., Hancock, J. D., Allee, G. L., & Klemm, R. D.
(1990). Transient hypersensitivity to soybean meal in the early-weaned pig. Journal of Animal Science, 68(6), 1790-1799.
Li, D. F., Nelssen, J. L., Reddy, P. G., Blecha, F., Klemm, R., & Goodband, R. D. (1991).
Interrelationship between hypersensitivity to soybean proteins and growth performance in early-weaned pigs. Journal of Animal Science,69(10), 4062-4069.
Li, H., Zheng, Z., Cong-xin, X., Bo, H., Chao-yuan, W., & Gang, H. (2009). Isolation of
cellulose—producing microbes from the intestine of grass carp (Ctenopharyngodon idellus). Environ Biol Fish 86, 131–135
Liener, I. E. (1994). Implications of antinutritional components in soybean foods. Critical
Reviews in Food Science & Nutrition, 34(1), 31-67. Liu, S., Xia, Y., Hu, H. Z., Ren, J., Gao, C., & Wood, J. D. (2000). Histamine H3 receptor-
mediated suppression of inhibitory synaptic transmission in the submucous plexus of guinea-pig small intestine.European journal of pharmacology, 397(1), 49-54.
Liying, Z., Li, D., Qiao, S., Johnson, E. W., Li, B., Thacker, P. A., & Han, I. K. (2003). Effects
of stachyose on performance, diarrhoea incidence and intestinal bacteria in weanling pigs. Archives of animal nutrition, 57(1), 1-10.
Lusas, E. W., & Riaz, M. N. (1995). Soy protein products: processing and use. The Journal of
nutrition, 125(3 Suppl), 573S-580S.

90
MacFaddin J.F. 2000. Biochemical tests for identification of medical bacteria, 3rd ed, p 412–
423. Lippincott Williams and Wilkins, Philadelphia, PA. Maenz, D. D., Irish, G. G., & Classen, H. L. (1999). Carbohydrate-binding and agglutinating
lectins in raw and processed soybean meals. Animal feed science and technology, 76(3), 335-343.
Mawadza, C., Hatti-Kaul, R., Zvauya, R., & Mattiasson, B. (2000). Purification and
characterization of cellulases produced by two bacillus strains. Journal of Biotechnology, 83(3), 177-187.
Merlot, E., Meunier-Salaün, M., & Prunier, A. (2004). Behavioural, endocrine and immune
consequences of mixing in weaned piglets. Applied Animal Behaviour Science, 85(3-4), 247-257.
Mikkola, R., Andersson, M. A., Grigoriev, P., Teplova, V. V., Saris, N. E. L., Rainey, F. A., &
Salkinoja-Salonen, M. S. (2004). Bacillus amyloliquefaciens strains isolated from moisture-damaged buildings produced surfactin and a substance toxic to mammalian cells. Archives of microbiology, 181(4), 314-323.
Mitrica, L., & Granum, P. (1979). The amylase-producing microflora of semi-preserved canned
sausages. Zeitschrift Für Lebensmittel-Untersuchung Und Forschung, 169(1), 4-8. Molist, F., de Segura, A. G., Gasa, J., Hermes, R. G., Manzanilla, E. G., Anguita, M., & Pérez, J.
F. (2009). Effects of the insoluble and soluble dietary fibre on the physicochemical properties of digesta and the microbial activity in early weaned piglets. Animal feed science and technology, 149(3), 346-353.
Montagne, L., Pluske, J. R., & Hampson, D. J. (2003). A review of interactions between dietary
fibre and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Animal Feed Science and Technology, 108(1-4), 95-117
Oberleas, D., Muhrer, M. E., & O'dell, B. L. (1962). Effects of phytic acid on zinc availability
and parakeratosis in swine. Journal of Animal Science,21(1), 57-61. Opazo, R., Ortuzar, F., Navarrete, P., Espejo, R., & Romero, J. (2012). Reduction of Soybean
Meal Non-Starch Polysaccharides and α-Galactosides by Solid-State Fermentation Using Cellulolytic Bacteria Obtained from Different Environments. PloS one, 7(9), e44783.
Palacios, M. F., Easter, R. A., Soltwedel, K. T., Parsons, C. M., Douglas, M. W., Hymowitz, T.,
& Pettigrew, J. E. (2004). Effect of soybean variety and processing on growth performance of young chicks and pigs. Journal of animal science, 82(4), 1108-1114.
Pan, B., Li, D., Piao, X., Zhang, L., & Guo, L. (2002). Effect of dietary supplementation with α-
galactosidase preparation and stachyose on growth performance, nutrient digestibility and intestinal bacterial populations of piglets. Archives of Animal Nutrition, 56(5), 327-337

91
Payne, R. L., Bidner, T. D., Southern, L. L., & Geaghan, J. P. (2001). Effects of dietary soy isoflavones on growth, carcass traits, and meat quality in growing-finishing pigs. Journal of animal science, 79(5), 1230-1239.
Peng, Y., Huang, Q., Zhang, R. H., & Zhang, Y. Z. (2003). Purification and characterization of a
fibrinolytic enzyme produced by Bacillus amyloliquefaciens DC-4 screened from douchi, a traditional Chinese soybean food. Comparative biochemistry and physiology part b: biochemistry and molecular biology, 134(1), 45-52.
Pérez, J., Munoz-Dorado, J., de la Rubia, T. D. L. R., & Martinez, J. (2002). Biodegradation and
biological treatments of cellulose, hemicellulose and lignin: an overview. International Microbiology, 5(2), 53-63.
Pluske, J. R., Hampson, D. J., & Williams, I. H. (1997). Factors influencing the structure and
function of the small intestine in the weaned pig: a review. Livestock production science, 51(1), 215-236.
Pyo, Y. H., Lee, T. C., & Lee, Y. C. (2005). Effect of lactic acid fermentation on enrichment of
antioxidant properties and bioactive isoflavones in soybean.Journal of food science, 70(3), S215-S220.
Rackis, J. J., McGee, J. E., Gumbmann, M. R., & Booth, A. N. (1979). Effects of soy proteins
containing trypsin inhibitors in long term feeding studies in rats. Journal of the American Oil Chemists’ Society, 56(3), 162-168.
Refstie, S., Sahlström, S., Bråthen, E., Baeverfjord, G., & Krogedal, P. (2005). Lactic acid
fermentation eliminates indigestible carbohydrates and antinutritional factors in soybean meal for Atlantic salmon (Salmo salar). Aquaculture, 246(1), 331-345.
Rodrigues Brasil, A. P., de Rezende, S. T., do Carmo Gouveia Pelúzio, M., & Guimarães, V. M.
(2010). Removal of oligosaccharides in soybean flour and nutritional effects in rats. Food chemistry, 118(2), 251-255.
Rooke, J. A., Slessor, M., Fraser, H., & Thomson, J. R. (1998). Growth performance and gut
function of piglets weaned at four weeks of age and fed protease-treated soya-bean meal. Animal feed science and technology,70(3), 175-190.
Ruijssenaars, H., & Hartmans, S. (2001). Plate screening methods for the detection of
polysaccharase-producing microorganisms. Applied microbiology and biotechnology, 55(2), 143-149.
Saz, J. M., & Marina, M. L. (2007). High performance liquid chromatography and capillary
electrophoresis in the analysis of soybean proteins and peptides in foodstuffs. Journal of separation science, 30(4), 431-451.
Sazci, A., Erenler, K., & Radford, A. (1986). Detection of cellulolytic fungi by using Congo red
as an indicator: a comparative study with the dinitrosalicyclic acid reagent method. Journal of Applied Microbiology,61(6), 559-562.

92
Shih, I. L., & Van, Y. T. (2001). The production of poly-(γ-glutamic acid) from microorganisms
and its various applications. Bioresource Technology,79(3), 207-225. Shim, J. H., & Oh, B. C. (2012). Characterization and application of calcium-dependent β-
Propeller phytase from Bacillus amyloliquefaciens DS11. Journal of agricultural and food chemistry, 60(30), 7532-7537
Silk, D. B. A., Chung, Y. C., Berger, K. L., Conley, K., Beigler, M., Sleisenger, M. H., & Kim,
Y. S. (1979). Comparison of oral feeding of peptide and amino acid meals to normal human subjects. Gut, 20(4), 291-299.
Singhania, R. R., Patel, A. K., Soccol, C. R., & Pandey, A. (2009). Recent advances in solid-
state fermentation. Biochemical Engineering Journal,44(1), 13-18. Smiricky, M. R., Grieshop, C. M., Albin, D. M., Wubben, J. E., Gabert, V. M., & Fahey, G. C.
(2002). The influence of soy oligosaccharides on apparent and true ileal amino acid digestibilities and fecal consistency in growing pigs.Journal of animal science, 80(9), 2433-2441.
Smits, C. H., & Annison, G. (1996). Non-starch plant polysaccharides in broiler nutrition-
towards a physiologically valid approach to their determination. World's poultry science journal, 52(2), 203-222.
Snoeck, V., Cox, E., Verdonck, F., Joensuu, J. J., & Goddeeris, B. M. (2004). Influence of
porcine intestinal pH and gastric digestion on antigenicity of F4 fimbriae for oral immunisation. Veterinary Microbiology, 98(1), 45-53.
Sohn, K. S., Maxwell, C. V., Buchanan, D. S., & Southern, L. L. (1994). Improved soybean
protein sources for early-weaned pigs: I. Effects on performance and total tract amino acid digestibility. Journal of animal science, 72(3), 622-630.
Song, B., Rong, Y. J., Zhao, M. X., & Chi, Z. M. (2013). Antifungal activity of the lipopeptides
produced by Bacillus amyloliquefaciens anti-CA against Candida albicans isolated from clinic. Applied microbiology and biotechnology,97(16), 7141-7150.
Song, Y. S., Pérez, V. G., Pettigrew, J. E., Martinez-Villaluenga, C., & de Mejia, E. G. (2010).
Fermentation of soybean meal and its inclusion in diets for newly weaned pigs reduced diarrhea and measures of immunoreactivity in the plasma. Animal feed science and technology, 159(1), 41-49.
Sørensen, M., Penn, M., El-Mowafi, A., Storebakken, T., Chunfang, C., Øverland, M., &
Krogdahl, Å. (2011). Effect of stachyose, raffinose and soya-saponins supplementation on nutrient digestibility, digestive enzymes, gut morphology and growth performance in Atlantic salmon (Salmo salar, L). Aquaculture, 314(1), 145-152
Soy Stats, 1998. A reference guide to important soybean facts & figures. American Soybean
Association. Accessed from www.soystats.com.

93
Stalikas, C,D. (2010). Phenolic Acids and Flavonoids: Occurrence and Analytical Methods. In
R.M. Uppu et al. (eds.), Free Radicals and Antioxidant Protocols, Humana Press. 610: 65-90.
Stein, H.H., Berger, L.L., Drackley, J.K., Fahey, G.C., Hernot, D.C., & Parsons., C.M. (2008).
Nutritional properties and feeding values of soybeans and their co-products. Pages 613-660 in Soybeans, Chemistry, Production, Processing, and Utilization. Johnson, L.A., White, P.J., & Galloway, ed AOCS Press, Urbana IL.
Stickler, M. T. 1992. Effect of Feeding the Kunitz Trypsin-Inhibitor Free Soybean on Swine
Growth Performance. M.S. Thesis, Univ. of Illinois, Urbana. Struthers, B. J., MacDonald, J. R., Dahlgren, R. R., & Hopkins, D. T. (1983). Effects on the
monkey, pig and rat pancreas of soy products with varying levels of trypsin inhibitor and comparison with the administration of cholecystokinin. The Journal of nutrition, 113(1), 86.
Sun, P., Li, D., Dong, B., Qiao, S., & Ma, X. (2008a). Effects of soybean glycinin on
performance and immune function in early weaned pigs.Archives of animal nutrition, 62(4), 313-321.
Sun, P., Li, D., Li, Z., Dong, B., & Wang, F. (2008b). Effects of glycinin on IgE-mediated
increase of mast cell numbers and histamine release in the small intestine. The Journal of nutritional biochemistry, 19(9), 627-633.
Sung, M. J., Davaatseren, M., Hwnag, J., Park, J. H., Kim, M. S., & Wang, S. (2013). Poly-
gamma-glutamic acid attenuates angiogenesis and inflammation in experimental colitis. Life Sciences, 93(25-26), e10-e10.
Thomsson, A., Rantzer, D., Weström, B. R., Pierzynowski, S. G., & Svendsen, J. (2007). Effects
of crude red kidney bean lectin (phytohemagglutinin) exposure on performance, health, feeding behavior, and gut maturation of pigs at weaning. Journal of animal science, 85(2), 477-485.
Thu, T., Loh, T., Foo, H., Yaakub, H., & Bejo, M. (2011). Effects of liquid metabolite
combinations produced by lactobacillus plantarum on growth performance, faeces characteristics, intestinal morphology and diarrhoea incidence in postweaning piglets. Tropical Animal Health and Production, 43(1), 69-75
Tonolla M, Benagli C, Rossi V, Fragoso C, Petrini O. 2010. MALDI-TOF MS: a new laboratory
option for the diagnosis of clinical infections. Pipette Swiss Laboratory Medicine 3: 6-10.
Valyasevi, R., & Rolle, R. S. (2002). An overview of small-scale food fermentation technologies
in developing countries with special reference to Thailand: scope for their improvement. International journal of food microbiology, 75(3), 231-239.

94
Van Duursen, M. B. M., Nijmeijer, S. M., de Morree, E. S., de Jong, P. C., & van den Berg, M.
(2011). Genistein induces breast cancer-associated aromatase and stimulates estrogen-dependent tumor cell growth in in vitro breast cancer model. Toxicology, 289(2-3), 67-73.
Van Kempen, T. A. T. G., van Heugten, E., Moeser, A. J., Muley, N. S., & Sewalt, V. J. H.
(2006). Selecting soybean meal characteristics preferred for swine nutrition. Journal of animal science, 84(6), 1387-1395.
Wang, L. T., Lee, F. L., Tai, C. J., & Kasai, H. (2007). Comparison of gyrB gene sequences, 16S
rRNA gene sequences and DNA–DNA hybridization in the Bacillus subtilis group. International Journal of Systematic and Evolutionary Microbiology, 57(8), 1846-1850.
Wang, T., Qin, G., Sun, Z., Zhao, Y., & Zhang, B. (2010). Comparative study on the residual
rate of immunoreactive soybean glycinin (11S) in the digestive tract of pigs of different ages. Food and agricultural immunology,21(3), 201-208.
Weary, D. M., Appleby, M. C., & Fraser, D. (1999). Responses of piglets to early separation
from the sow. Applied Animal Behaviour Science, 63(4), 289-300. Wilson, S., Blaschek, K., & Mejia, E. G. (2005). Antigenic proteins in soybean: processing and
reduction of P34 antigenicity. Nutrition reviews,63(2), 47-58. Wongputtisin, P., Khanongnuch, C., Pongpiachan, P., & Lumyong, S. (2007). Antioxidant
Activity Improvement of Soybean Meal by Microbial Fermentation.Research Journal of Microbiology, 2(7).
Wongputtisin, P., Khanongnuch, C., Kongbuntad, W., Niamsup, P., Lumyong, S., & Sarkar, P.
K. (2014). Use of Bacillus subtilis isolates from Tua‐nao towards nutritional improvement of soya bean hull for monogastric feed application. Letters in applied microbiology, 59(3), 328-333.
Xu, H. M., Rong, Y. J., Zhao, M. X., Song, B., & Chi, Z. M. (2014). Antibacterial activity of the
lipopetides produced by Bacillus amyloliquefaciens M1 against multidrug-resistant Vibrio spp. isolated from diseased marine animals. Applied microbiology and biotechnology, 98(1), 127-136.
Yamanishi, R., Huang, T., Tsuji, H., Bando, N., & Ogawa, T. (1995). Reduction of the Soybean
Antigenicity by the Fermentation with Bacillus natto. Food Science and Technology International, Tokyo, 1(1), 14-17.
Yamamoto, S., & Harayama, S. (1995). PCR amplification and direct sequencing of gyrB genes
with universal primers and their application to the detection and taxonomic analysis of Pseudomonas putida strains. Applied and environmental microbiology, 61(3), 1104-1109

95
Yin, Y. L., Baidoo, S. K., Schulze, H., & Simmins, P. H. (2001). Effects of supplementing diets containing hulless barley varieties having different levels of non-starch polysaccharides with β-glucanase and xylanase on the physiological status of the gastrointestinal tract and nutrient digestibility of weaned pigs. Livestock Production Science, 71(2), 97-107.
Ying, W., Zhu, R., Lu, W., & Gong, L. (2009). A new strategy to apply Bacillus subtilis MA139
for the production of solid‐state fermentation feed. Letters in applied microbiology, 49(2), 229-234.
Yoon, M. Y., & Hwang, H. J. (2008). Reduction of soybean oligosaccharides and properties of
α-d-galactosidase from Lactobacillus curvatus R08 and Leuconostoc mesenteriodes JK55. Food microbiology, 25(6), 815-823.
Yuan, F. J., Zhao, H. J., Ren, X. L., Zhu, S. L., Fu, X. J., & Shu, Q. Y. (2007). Generation and
characterization of two novel low phytate mutations in soybean (Glycine max L. Merr.). Theoretical and Applied Genetics, 115(7), 945-957
Zhang, J. H., Tatsumi, E., Fan, J. F., & Li, L. T. (2007). Chemical components of Aspergillus‐
type Douchi, a Chinese traditional fermented soybean product, change during the fermentation process. International journal of food science & technology, 42(3), 263-268.
Zhang, Y., C. M. Parsons, K. E. Weingartner, and W. B. Wijeratne. (1993). Effects of extrusion
and expelling on the nutritional quality of conventional and Kunitz trypsin inhibitor-free soybeans. Poult. Sci.
Zheng, H. G., Yang, X. Q., Ahmad, I., Min, W., Zhu, J. H., & Yuan, D. B. (2009). Soybean β-
conglycinin constituent subunits: Isolation, solubility and amino acid composition. Food research international, 42(8), 998-1003.
Zhao, Y., Qin, G., Sun, Z., Zhang, X., Bao, N., Wang, T., & Sun, L. (2008). Disappearance of
immunoreactive glycinin and β-conglycinin in the digestive tract of piglets. Archives of animal nutrition, 62(4), 322-330.
Zhao, Y., Qin, G. X., Sun, Z. W., Zhang, B., & Wang, T. (2010). Effects of glycinin and β-
conglycinin on enterocyte apoptosis, proliferation and migration of piglets. Food and agricultural immunology, 21(3), 209-218.
Zhou, Y., Yuan, X., Liang, X. F., Fang, L., Li, J., Guo, X., Bai, X., & He, S. (2013).
Enhancement of growth and intestinal flora in grass carp: The effect of exogenous cellulase. Aquaculture, 416, 1-7
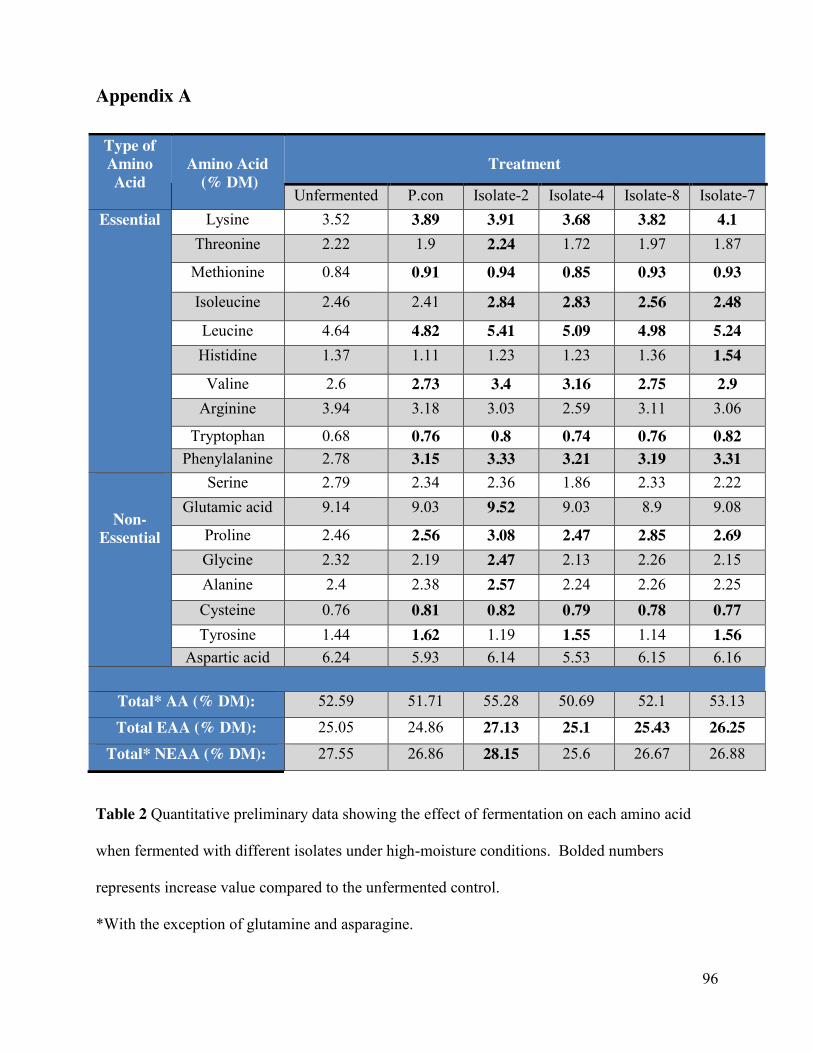
96
Appendix A
Table 2 Quantitative preliminary data showing the effect of fermentation on each amino acid
when fermented with different isolates under high-moisture conditions. Bolded numbers
represents increase value compared to the unfermented control.
*With the exception of glutamine and asparagine.
Type of Amino Acid
Amino Acid
(% DM)
Treatment
Unfermented P.con Isolate-2 Isolate-4 Isolate-8 Isolate-7 Essential Lysine 3.52 3.89 3.91 3.68 3.82 4.1
Threonine 2.22 1.9 2.24 1.72 1.97 1.87
Methionine 0.84 0.91 0.94 0.85 0.93 0.93
Isoleucine 2.46 2.41 2.84 2.83 2.56 2.48
Leucine 4.64 4.82 5.41 5.09 4.98 5.24 Histidine 1.37 1.11 1.23 1.23 1.36 1.54
Valine 2.6 2.73 3.4 3.16 2.75 2.9 Arginine 3.94 3.18 3.03 2.59 3.11 3.06
Tryptophan 0.68 0.76 0.8 0.74 0.76 0.82 Phenylalanine 2.78 3.15 3.33 3.21 3.19 3.31
Non-Essential
Serine 2.79 2.34 2.36 1.86 2.33 2.22 Glutamic acid 9.14 9.03 9.52 9.03 8.9 9.08
Proline 2.46 2.56 3.08 2.47 2.85 2.69 Glycine 2.32 2.19 2.47 2.13 2.26 2.15 Alanine 2.4 2.38 2.57 2.24 2.26 2.25 Cysteine 0.76 0.81 0.82 0.79 0.78 0.77 Tyrosine 1.44 1.62 1.19 1.55 1.14 1.56
Aspartic acid 6.24 5.93 6.14 5.53 6.15 6.16
Total* AA (% DM): 52.59 51.71 55.28 50.69 52.1 53.13
Total EAA (% DM): 25.05 24.86 27.13 25.1 25.43 26.25
Total* NEAA (% DM): 27.55 26.86 28.15 25.6 26.67 26.88





























