Embed Size (px)
Citation preview

Im
YMa
b
c
d
e
a
ARRAA
KPPANTP
0d
Journal of Ethnopharmacology 139 (2012) 616– 625
Contents lists available at SciVerse ScienceDirect
Journal of Ethnopharmacology
jo ur nal homep age : www.elsev ier .com/ locate / je thpharm
n vitro and in vivo anti-inflammatory activities of Polygonum hydropiperethanol extract
anyan Yanga, Tao Yua, Hyun-Jae Jangb, Se Eun Byeona, Song-Yi Songc, Byoung-Hee Leec,an Hee Rheed, Tae Woong Kimb, Jaehwi Leee,∗∗, Sungyoul Honga, Jae Youl Choa,∗
Department of Genetic Engineering, Sungkyunkwan University, Suwon 440-746, Republic of KoreaDepartment of Biochemistry, Kangwon National University, Chuncheon 200-701, Republic of KoreaDivision of Biological Resources Coordination, National Institute of Biological Resources, Incheon 404-708, Republic of KoreaCollege of Veterinary Medicine, Kyungpook National University, Daegu 702-701, Republic of KoreaCollege of Pharmacy, Chung-Ang University, Seoul 156-756, Republic of Korea
r t i c l e i n f o
rticle history:eceived 12 July 2011eceived in revised form 29 October 2011ccepted 2 December 2011vailable online 13 December 2011
eywords:olygonum hydropiper L.olygonaceaenti-inflammatory effectsitric oxideumour necrosis factor-�rostaglandin E2
a b s t r a c t
Ethnopharmacological relevance: Polygonum hydropiper L. (Polygonaceae) has been traditionally used totreat various inflammatory diseases such as rheumatoid arthritis. However, no systematic studies on theanti-inflammatory actions of Polygonum hydropiper and its inhibitory mechanisms have been reported.This study is therefore aimed at exploring the anti-inflammatory effects of 99% methanol extracts (Ph-ME)of this plant.Materials and methods: The effects of Ph-ME on the production of inflammatory mediators in RAW264.7cells and peritoneal macrophages were investigated. Molecular mechanisms underlying the effects, espe-cially inhibitory effects, were elucidated by analyzing the activation of transcription factors and theirupstream signalling, and by evaluating the kinase activities of target enzymes. Additionally, a dextransulphate sodium (DSS)-induced colitis model was employed to see whether this extract can be used asan orally available drug.Results: Ph-ME dose-dependently suppressed the release of nitric oxide (NO), tumour necrosis fac-tor (TNF)-�, and prostaglandin (PG)E2, in RAW264.7 cells and peritoneal macrophages stimulated bylipopolysaccharide (LPS). Ph-ME inhibited mRNA expression of pro-inflammatory genes such as inducibleNO synthase (iNOS), cyclooxygenase (COX)-2, and TNF-� by suppressing the activation of nuclear factor(NF)-�B, activator protein (AP-1), and cAMP responsive element binding protein (CREB), and simultane-ously inhibited its upstream inflammatory signalling cascades, including cascades involving Syk, Src, and
IRAK1. Consistent with these findings, the extract strongly suppressed the kinase activities of Src andSyk. Based on HPLC analysis, quercetin, which inhibits NO and PGE2 activities, was found as one of theactive ingredients in Ph-ME.Conclusion: Ph-ME exerts strong anti-inflammatory activity by suppressing Src/Syk/NF-�B and IRAK/AP-1/CREB pathways, which contribute to its major ethno-pharmacological role as an anti-gastritisglandin E2; NO, nitric oxide; COX, cyclooxygenase; iNOS, inducible NO synthase; (TNF)-�,eceptors (TLR); MAPK, mitogen activated protein kinase; NF-�B, nuclear factor-�B; AP-1,ctivating transcription factor 2; CREB, cAMP response element-binding; IKK, I�B� kinase;D88, myeloid differentiation primary-response protein-88; TRAF6, tumor necrosis factor-oinositide-dependent kinase-1; Syk, spleen tyrosine kinase; TRIF, TIR-domain-containing
remedy.
Abbreviations: Ph-ME, methanol extract of Polygonum hydropiper L.; PGE2, prostatumour necrosis factor; ERK, extracellular signal-related kinase; TLR, Toll-like ractivator protein-1; JNK, c-Jun N-terminal kinase; Akt, protein kinase B; ATF2, aIRAK1, interleukin-1 receptor-associated kinase 1; MKK, MAP Kinase Kinase; Myreceptor-associated factor-6; TAK1, TGF-Beta-Activated Kinase-1; PDK1, Phosph
adapter-inducing interferon-�; EIA, enzyme immunoassay; ELISA, enzyme linked imbromide, a tetrazole; PI3K, phosphoinositide 3-kinases; LPS, lipopolysaccharide; RT-PC∗ Corresponding author. Tel.: +82 2 820 5606; fax: +82 2 816 7338.∗∗ Corresponding author. Tel.: +82 31 290 7868; fax: +82 31 290 7870.
E-mail addresses: [email protected] (J. Lee), [email protected], [email protected] (
378-8741/$ – see front matter © 2011 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.jep.2011.12.003
© 2011 Elsevier Ireland Ltd. All rights reserved.
munosorbent assay; MTT, (3-4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumR, reverse transcriptase-polymerase chain reaction; DSS, dextran sulphate sodium.
J.Y. Cho).

pharmacology 139 (2012) 616– 625 617
1
ivtat2s
FLfwpa
Table 1Sequences of primers used in real-time PCR analysis.
Gene Primer sequences
TNF-� F 5′-TTGACCTCAGCGCTGAGTTG-3′
R 5′-CCTGTAGCCCACGTCGTAGC-3′
iNOS F 5′-CCCTTCCGAAGTTTCTGGCAGCAGC-3′′ ′
Y. Yang et al. / Journal of Ethno
. Introduction
Inflammation is one of the body’s self defence systems thats classified as part of our innate immunity. Thus, bacterial oriral infections trigger numerous immunobiological events such ashe production of cytokines, chemokines, and inflammatory medi-
tors [including nitric oxide (NO), prostaglandin E2 (PGE2) andumour necrosis factor (TNF)-� (Kinne et al., 2000; Owens et al.,005)], migration and infiltration of leukocytes, increased expres-ion of surface molecules such as MHC molecules, complementRAW264.7Peritoneal macrophage
120
140
160 HEK293
80
100
20
40
60
00 25 50 100 200
Cel
l via
bilit
y (%
of c
ontr
ol)
Ph-ME (μg/ml)
Peritoneal macrophages
80
100
120
40
602pr
oduc
tion
*
0
20NOPGEN
O a
nd P
GE
(% o
f con
trol
)
**
**
**
2
0 25 50 100 200Ph-ME ( μg/ml)
140 RAW264.7 cells
100
120α
60
80
2an
d TN
F-
**
20
40 NOPGE2
NO
, PG
Epr
oduc
tion
(% o
f con
trol
)
**
*
0TNF-α
0 25 50 100 200Ph-ME (
**
μg/ml )
A
B
C
ig. 1. Effect of Ph-ME on the production of inflammatory mediators. (A and B)evels of NO, PGE2, and TNF-� were determined by Griess assay, EIA, and ELISArom culture supernatants of RAW264.7 cells or peritoneal macrophages treatedith Ph-ME and LPS (1 �g/ml) for 6 or 24 h. (C) Cell viability of RAW264.7 cells,eritoneal macrophages, and HEK293 cells was determined by MTT assay. *P < 0.05nd **P < 0.01 compared to control.
R 5 -GGCTGTCAGAGCCTCGTGGCTTTGG-3COX-2 F 5′-CACTACATCCTGACCCACTT-3′
R 5′-ATGCTCCTGCTTGAGTATGT-3′
IL-1� F 5′-CAGGATGAGGACATGAGCACC-3′
R 5′-CTCTGCAGACTCAAACTCCAC-3′
IL-6 F 5′-GTACTCCAGAAGACCAGAGG-3′
R 5′-TGCTGGTGACAACCACGGCC-3′
GAPDH F 5′-CACTCACGGCAAATTCAACGGCAC-3′
R 5′-GACTCCACGACATACTCAGCAC-3′
Table 2The half maximal effective concentration (EC50) of Ph-ME in the production of NOand PGE2 in RAW264.7 cells and peritoneal macrophages.
Cells EC50 value (�g/ml)
NO PGE2 TNF-�
RAW264.7 cells 107.3 118.9 252.7Peritoneal macrophages 252.7 95.4 >300
receptors, and release of hydrolytic enzymes, to clear out suchinfections (Deban et al., 2009). Unfortunately, acute and chronic
inflammation can cause lead to acute or chronic inflammatorydiseases such as septic shock, rheumatoid arthritis, gastritis, andatherosclerosis (Michaelsson et al., 1995; Stuhlmuller et al., 2000).120
80
100IL-1βIL-6
60β an
d IL
-6
20
40IL-1
expr
essi
on (%
of c
ontr
ol)
00 50 100 200
Ph-ME ( μg/ml)
120
80
100
αan
d C
OX-
2
60
iNO
S, T
NF-
expr
essi
on (%
of c
ontr
ol)
20
40iNOSTNF-αCOX-2
**
* **
*
*
00 50 100 200
**
Ph-ME ( μg/ml)
A
B
Fig. 2. Effect of Ph-ME on the mRNA expression of pro-inflammatory genes. (A andB) The mRNA levels of iNOS, COX-2, TNF-�, IL-1�, and IL-6 were determined byreal-time PCR. *P < 0.05 and **P < 0.01 compared to control.

618 Y. Yang et al. / Journal of Ethnopharmacology 139 (2012) 616– 625
Fig. 3. Effect of Ph-ME on the activation of NF-�B. (A–C) HEK293 cells co-transfected with plasmid constructs NF-�B-Luc, adaptor molecules (MyD88 and TRIF) (each 1 �g/ml)and �-gal (as a transfection control) were treated with Ph-ME in the presence or absence of PMA (100 nM). Luciferase activity was measured by luminometer. (D) Levels ofNF-�B (p65/p50) in nuclear fractions were determined by immunoblotting analysis with antibodies against the total protein. *P < 0.05 and **P < 0.01 compared to control.
Fig. 4. Effect of Ph-ME on the activation of AP-1. (A–C) HEK293 cells co-transfected with plasmid constructs AP-1-Luc, adaptor molecules (MyD88 and TRIF) (each 1 �g/ml)and �-gal (as a transfection control) were treated with Ph-ME in the presence or absence of PMA (100 nM). Luciferase activity was measured by luminometer. (D) Levels ofmembers of the AP-1 family (ATF-2, phospho (p)-ATF-2, c-Jun and c-Fos) in nuclear fractions were determined by immunoblotting analysis with antibodies against the totalprotein. *P < 0.05 and **P < 0.01 compared to control.
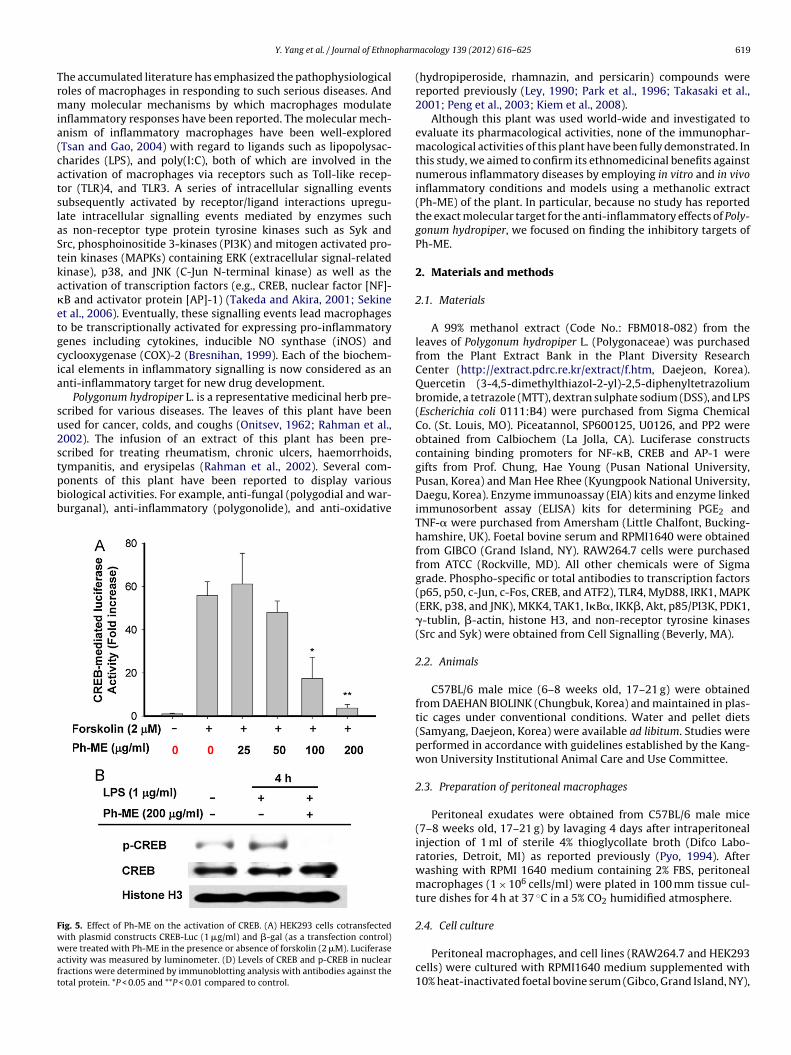
pharm
Trmia(catslaStka�etgcia
su2stpbb
Fwwaft
Y. Yang et al. / Journal of Ethno
he accumulated literature has emphasized the pathophysiologicaloles of macrophages in responding to such serious diseases. Andany molecular mechanisms by which macrophages modulate
nflammatory responses have been reported. The molecular mech-nism of inflammatory macrophages have been well-exploredTsan and Gao, 2004) with regard to ligands such as lipopolysac-harides (LPS), and poly(I:C), both of which are involved in thectivation of macrophages via receptors such as Toll-like recep-or (TLR)4, and TLR3. A series of intracellular signalling eventsubsequently activated by receptor/ligand interactions upregu-ate intracellular signalling events mediated by enzymes suchs non-receptor type protein tyrosine kinases such as Syk andrc, phosphoinositide 3-kinases (PI3K) and mitogen activated pro-ein kinases (MAPKs) containing ERK (extracellular signal-relatedinase), p38, and JNK (C-Jun N-terminal kinase) as well as thectivation of transcription factors (e.g., CREB, nuclear factor [NF]-B and activator protein [AP]-1) (Takeda and Akira, 2001; Sekinet al., 2006). Eventually, these signalling events lead macrophageso be transcriptionally activated for expressing pro-inflammatoryenes including cytokines, inducible NO synthase (iNOS) andyclooxygenase (COX)-2 (Bresnihan, 1999). Each of the biochem-cal elements in inflammatory signalling is now considered as annti-inflammatory target for new drug development.
Polygonum hydropiper L. is a representative medicinal herb pre-cribed for various diseases. The leaves of this plant have beensed for cancer, colds, and coughs (Onitsev, 1962; Rahman et al.,002). The infusion of an extract of this plant has been pre-cribed for treating rheumatism, chronic ulcers, haemorrhoids,
ympanitis, and erysipelas (Rahman et al., 2002). Several com-onents of this plant have been reported to display variousiological activities. For example, anti-fungal (polygodial and war-urganal), anti-inflammatory (polygonolide), and anti-oxidativeig. 5. Effect of Ph-ME on the activation of CREB. (A) HEK293 cells cotransfectedith plasmid constructs CREB-Luc (1 �g/ml) and �-gal (as a transfection control)ere treated with Ph-ME in the presence or absence of forskolin (2 �M). Luciferase
ctivity was measured by luminometer. (D) Levels of CREB and p-CREB in nuclearractions were determined by immunoblotting analysis with antibodies against theotal protein. *P < 0.05 and **P < 0.01 compared to control.
acology 139 (2012) 616– 625 619
(hydropiperoside, rhamnazin, and persicarin) compounds werereported previously (Ley, 1990; Park et al., 1996; Takasaki et al.,2001; Peng et al., 2003; Kiem et al., 2008).
Although this plant was used world-wide and investigated toevaluate its pharmacological activities, none of the immunophar-macological activities of this plant have been fully demonstrated. Inthis study, we aimed to confirm its ethnomedicinal benefits againstnumerous inflammatory diseases by employing in vitro and in vivoinflammatory conditions and models using a methanolic extract(Ph-ME) of the plant. In particular, because no study has reportedthe exact molecular target for the anti-inflammatory effects of Poly-gonum hydropiper, we focused on finding the inhibitory targets ofPh-ME.
2. Materials and methods
2.1. Materials
A 99% methanol extract (Code No.: FBM018-082) from theleaves of Polygonum hydropiper L. (Polygonaceae) was purchasedfrom the Plant Extract Bank in the Plant Diversity ResearchCenter (http://extract.pdrc.re.kr/extract/f.htm, Daejeon, Korea).Quercetin (3-4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide, a tetrazole (MTT), dextran sulphate sodium (DSS), and LPS(Escherichia coli 0111:B4) were purchased from Sigma ChemicalCo. (St. Louis, MO). Piceatannol, SP600125, U0126, and PP2 wereobtained from Calbiochem (La Jolla, CA). Luciferase constructscontaining binding promoters for NF-�B, CREB and AP-1 weregifts from Prof. Chung, Hae Young (Pusan National University,Pusan, Korea) and Man Hee Rhee (Kyungpook National University,Daegu, Korea). Enzyme immunoassay (EIA) kits and enzyme linkedimmunosorbent assay (ELISA) kits for determining PGE2 andTNF-� were purchased from Amersham (Little Chalfont, Bucking-hamshire, UK). Foetal bovine serum and RPMI1640 were obtainedfrom GIBCO (Grand Island, NY). RAW264.7 cells were purchasedfrom ATCC (Rockville, MD). All other chemicals were of Sigmagrade. Phospho-specific or total antibodies to transcription factors(p65, p50, c-Jun, c-Fos, CREB, and ATF2), TLR4, MyD88, IRK1, MAPK(ERK, p38, and JNK), MKK4, TAK1, I�B�, IKK�, Akt, p85/PI3K, PDK1,�-tublin, �-actin, histone H3, and non-receptor tyrosine kinases(Src and Syk) were obtained from Cell Signalling (Beverly, MA).
2.2. Animals
C57BL/6 male mice (6–8 weeks old, 17–21 g) were obtainedfrom DAEHAN BIOLINK (Chungbuk, Korea) and maintained in plas-tic cages under conventional conditions. Water and pellet diets(Samyang, Daejeon, Korea) were available ad libitum. Studies wereperformed in accordance with guidelines established by the Kang-won University Institutional Animal Care and Use Committee.
2.3. Preparation of peritoneal macrophages
Peritoneal exudates were obtained from C57BL/6 male mice(7–8 weeks old, 17–21 g) by lavaging 4 days after intraperitonealinjection of 1 ml of sterile 4% thioglycollate broth (Difco Labo-ratories, Detroit, MI) as reported previously (Pyo, 1994). Afterwashing with RPMI 1640 medium containing 2% FBS, peritonealmacrophages (1 × 106 cells/ml) were plated in 100 mm tissue cul-ture dishes for 4 h at 37 ◦C in a 5% CO2 humidified atmosphere.
2.4. Cell culture
Peritoneal macrophages, and cell lines (RAW264.7 and HEK293cells) were cultured with RPMI1640 medium supplemented with10% heat-inactivated foetal bovine serum (Gibco, Grand Island, NY),

6 pharm
guctb
2
mwwPPie
2
tfa1(cmdeu
FSkfc
20 Y. Yang et al. / Journal of Ethno
lutamine and antibiotics (penicillin and streptomycin) at 37 ◦Cnder 5% CO2. For each experiment, cells were detached with aell scraper. Under our experimental cell density (2 × 106 cells/ml),he proportion of dead cells was less than 1%, according to Trypanlue dye exclusion tests.
.5. NO, PGE2 and TNF- ̨ production
After preincubation of RAW264.7 cells or peritonealacrophages (1 × 106 cells/ml) for 18 h, cells were pre-treatedith Ph-ME (0–400 �g/ml) for 30 min and were further incubatedith LPS (1 �g/ml) for 24 h. The inhibitory effect of Ph-ME on NO,
GE2, and TNF-� production was determined by analyzing NO,GE2, and TNF-� levels with the Griess reagent and enzyme linkedmmunosorbent assay (ELISA) kits, as described previously (Chot al., 2000).
.6. Cell viability test
After preincubation of RAW264.7 cells (1 × 106 cells/ml) for 18 h,he Ph-ME (0–100 �g/ml) was added to the cells and incubatedor 24 h. The cytotoxic effect of the Ph-ME was then evaluated by
conventional MTT assay, as reported previously (Pauwels et al.,988). At 3 h prior to culture termination, 10 �l of an MTT solution10 mg/ml in phosphate buffered-saline, pH 7.4) was added and theells were continuously cultured until termination of the experi-
ent. The incubation was halted by the addition of 15% sodiumodecyl sulphate into each well, solubilising the formazan (Kimt al., 2008). The absorbance at 570 nm (OD570–630) was measuredsing a Spectramax 250 microplate reader.
ig. 6. Effect of Ph-ME on the activation of upstream signalling enzymes for NF-�B translyk, p85/PI3K, and �-actin from cell lysates were determined by phospho-specific or totalinase assay using purified enzymes. Control was set as 100% with each enzyme (Src or Syrom LPS-treated RAW264.7 cells pre-treated with standard Src and Syk inhibitors (PP2ompared to control.
acology 139 (2012) 616– 625
2.7. mRNA analysis by quantitative real-time polymerase chainreactions
To determine cytokine mRNA expression levels, total RNA wasisolated from LPS-treated RAW264.7 cells with TRIzol Reagent(Gibco BRL), according to the manufacturer’s instructions. TotalRNA was stored at −70 ◦C until use. Quantification of mRNAwas also performed using real-time RT-PCR with manufacturer’sinstructions of SYBR Premix Ex Taq (Takara, Japan) using real-timethermal cycler (Bio-Rad, USA) as reported previously (Sun et al.,2010; Kim and Lee, 2011). The results were expressed as the ratioof optimal density to GAPDH. The primers (Bioneer, Daejeon, Korea)used are indicated in Table 1.
2.8. Luciferase reporter gene activity assay
HEK293 cells (1 × 106 cells/ml) were transfected with 1 �g ofplasmids containing NF-�B-Luc, CREB-Luc, or AP-1-Luc as wellas �-galactosidase using the calcium phosphate method in a 12-well plate according to the manufacturer’s protocol (Jin et al.,2009). The cells were used for experiments 48 h after transfec-tion. Luciferase assays were performed using the Luciferase AssaySystem (Promega) as reported previously (Jung et al., 2006).
2.9. Preparation of total lysates and nuclear fraction,immunoblotting, and immunoprecipitation
RAW264.7 cells (5 × 106 cells/ml) were washed 3 timesin cold PBS with 1 mM sodium orthovanadate and lysedin lysis buffer (20 mM Tris–HCl, pH 7.4, 2 mM EDTA, 2 mM
ocation. (A and B) Phospho-protein or total protein levels of I�B�, IKK�/�, Akt, Src, protein antibodies. (C) Kinase activities of Syk and Src were determined by a directk) activity only obtained with vehicle treatment. (D) Culture supernatants prepared
and piceatannol) were assayed for NO, TNF-�, and PGE2. *P < 0.05 and **P < 0.01

pharm
esgaca
epofntnKTtn
otws5
FJ(tfh
Y. Yang et al. / Journal of Ethno
thyleneglycotetraacetic acid, 50 mM �-glycerophosphate, 1 mModium orthovanadate, 1 mM dithiothreitol, 1% Triton X-100, 10%lycerol, 10 �g/ml aprotinin, 10 �g/ml pepstatin, 1 mM benzimide,nd 2 mM PMSF) for 30 min with rotation, at 4 ◦C. The lysates werelarified by centrifugation at 16,000 × g for 10 min at 4 ◦C and storedt −20 ◦C until needed.
Nuclear lysates were prepared in a three-step procedure (Byeont al., 2009). After treatment, cells were collected with a rubberoliceman, washed with 1× PBS, and lysed in 500 �l of lysis buffern ice for 4 min. Cell lysates were then centrifuged at 19,326 × gor 1 min in a microcentrifuge. In the second step, the pellet (theuclear fraction) was washed once in washing buffer, which washe same as the lysis buffer without Nonidet P-40. In the final step,uclei were treated with an extraction buffer containing 500 mMCl, 10% glycerol, and several other reagents as in the lysis buffer.he nuclei/extraction buffer mixture was frozen at −80 ◦C, and thenhawed on ice and centrifuged at 19,326 × g for 5 min. The super-atant was collected as a nuclear extract.
For immunoprecipitation, cell lysates containing equal amountsf protein (500 �g) from RAW264.7 cells (1 × 107 cells ml−1)
reated with or without LPS (1 �g/ml) for 2.5 min were pre-clearedith 10 �l of protein A-coupled Sepharose beads (50%, v/v) (Amer-ham, UK) for 1 h at 4 ◦C. Pre-cleared samples were incubated with �l of anti-TLR4 antibody overnight at 4 ◦C. Immune complexes
ig. 7. Effect of Ph-ME on the activation of upstream signalling enzymes for AP-1 tranNK), their upstream kinases (MKK, TAK1, and IRAK1), and �-actin from cell lysates wer5 × 106 cells/ml) were incubated with Ph-ME in the presence or absence of LPS (1 �g/ml)
o TLR4 were identified by immunoprecipitation with anti-TLR4 antibody and immunoblorom LPS-treated RAW264.7 cells pre-treated with standard p38 and JNK inhibitors (SB2eavy chain. *P < 0.05 compared to control.
acology 139 (2012) 616– 625 621
were mixed with 10 �l of protein A-coupled Sepharose beads (50%,vv) and rotated for 3 h at 4 ◦C.
Soluble cell lysates or boiled immunoprecipitated beads wereimmunoblotted and phosphorylated or total levels of transcriptionfactors (p65, p50, c-Jun, c-Fos, CREB, and ATF2), TLR4, MyD88, IRK1,MAPK (ERK, p38, and JNK), MKK4, TAK1, I�B�, IKK�, Akt, p85/PI3K,PDK1, �-actin, histone H3, and non-receptor tyrosine kinases (Srcand Syk) were visualized as previously reported (Chung et al., 2010;Rhee et al., 2010).
2.10. Syk and Src kinase assays
To evaluate the ability of extracts to inhibit Syk and Src kinaseactivity using purified enzymes, a kinase profiler service from Mil-lipore was used. In a final reaction volume of 25 �l, Src or Syk(human) (1–5 mU) was incubated with the reaction buffer. Thereaction was initiated by the addition of MgATP. After incuba-tion for 40 min at room temperature, the reaction was stoppedby the addition of 5 ml of a 3% phosphoric acid solution. Ten
microliters of the reaction product was then spotted onto a P30filtermat and washed three times for 5 min in 75 mM phospho-ric acid and once in methanol prior to drying and scintillationcounting.slocation. (A–C) Phospho-protein or total protein levels of MAPK (ERK, p38, ande determined by phospho-specific or total protein antibodies. (D) RAW264.7 cellsfor 0, 2, and 5 min. After preparing total lysates, binding levels of MyD88 and IRAK1tting with antibodies to MyD88, IRAK1, and TLR4. (E) Culture supernatants prepared03580 and SP600125) were assayed for NO, TNF-�, and PGE2 determination. H.C:
6 pharmacology 139 (2012) 616– 625
2
Dcwmam
2
pcs1aTgp
2
cpiwaao
3
emchppwectecs2vc
mutttu29biA
e
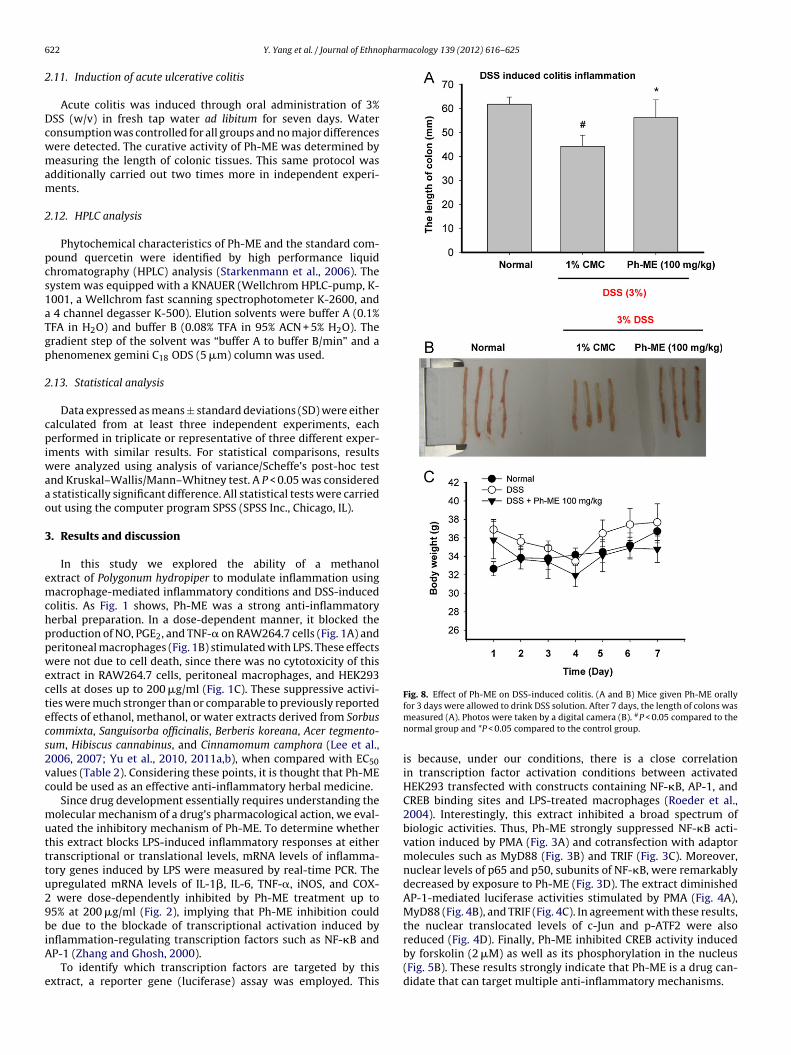
Fig. 8. Effect of Ph-ME on DSS-induced colitis. (A and B) Mice given Ph-ME orally
22 Y. Yang et al. / Journal of Ethno
.11. Induction of acute ulcerative colitis
Acute colitis was induced through oral administration of 3%SS (w/v) in fresh tap water ad libitum for seven days. Wateronsumption was controlled for all groups and no major differencesere detected. The curative activity of Ph-ME was determined byeasuring the length of colonic tissues. This same protocol was
dditionally carried out two times more in independent experi-ents.
.12. HPLC analysis
Phytochemical characteristics of Ph-ME and the standard com-ound quercetin were identified by high performance liquidhromatography (HPLC) analysis (Starkenmann et al., 2006). Theystem was equipped with a KNAUER (Wellchrom HPLC-pump, K-001, a Wellchrom fast scanning spectrophotometer K-2600, and
4 channel degasser K-500). Elution solvents were buffer A (0.1%FA in H2O) and buffer B (0.08% TFA in 95% ACN + 5% H2O). Theradient step of the solvent was “buffer A to buffer B/min” and ahenomenex gemini C18 ODS (5 �m) column was used.
.13. Statistical analysis
Data expressed as means ± standard deviations (SD) were eitheralculated from at least three independent experiments, eacherformed in triplicate or representative of three different exper-
ments with similar results. For statistical comparisons, resultsere analyzed using analysis of variance/Scheffe’s post-hoc test
nd Kruskal–Wallis/Mann–Whitney test. A P < 0.05 was considered statistically significant difference. All statistical tests were carriedut using the computer program SPSS (SPSS Inc., Chicago, IL).
. Results and discussion
In this study we explored the ability of a methanolxtract of Polygonum hydropiper to modulate inflammation usingacrophage-mediated inflammatory conditions and DSS-induced
olitis. As Fig. 1 shows, Ph-ME was a strong anti-inflammatoryerbal preparation. In a dose-dependent manner, it blocked theroduction of NO, PGE2, and TNF-� on RAW264.7 cells (Fig. 1A) anderitoneal macrophages (Fig. 1B) stimulated with LPS. These effectsere not due to cell death, since there was no cytotoxicity of this
xtract in RAW264.7 cells, peritoneal macrophages, and HEK293ells at doses up to 200 �g/ml (Fig. 1C). These suppressive activi-ies were much stronger than or comparable to previously reportedffects of ethanol, methanol, or water extracts derived from Sorbusommixta, Sanguisorba officinalis, Berberis koreana, Acer tegmento-um, Hibiscus cannabinus, and Cinnamomum camphora (Lee et al.,006, 2007; Yu et al., 2010, 2011a,b), when compared with EC50alues (Table 2). Considering these points, it is thought that Ph-MEould be used as an effective anti-inflammatory herbal medicine.
Since drug development essentially requires understanding theolecular mechanism of a drug’s pharmacological action, we eval-
ated the inhibitory mechanism of Ph-ME. To determine whetherhis extract blocks LPS-induced inflammatory responses at eitherranscriptional or translational levels, mRNA levels of inflamma-ory genes induced by LPS were measured by real-time PCR. Thepregulated mRNA levels of IL-1�, IL-6, TNF-�, iNOS, and COX-
were dose-dependently inhibited by Ph-ME treatment up to5% at 200 �g/ml (Fig. 2), implying that Ph-ME inhibition coulde due to the blockade of transcriptional activation induced by
nflammation-regulating transcription factors such as NF-�B andP-1 (Zhang and Ghosh, 2000).
To identify which transcription factors are targeted by thisxtract, a reporter gene (luciferase) assay was employed. This
for 3 days were allowed to drink DSS solution. After 7 days, the length of colons wasmeasured (A). Photos were taken by a digital camera (B). #P < 0.05 compared to thenormal group and *P < 0.05 compared to the control group.
is because, under our conditions, there is a close correlationin transcription factor activation conditions between activatedHEK293 transfected with constructs containing NF-�B, AP-1, andCREB binding sites and LPS-treated macrophages (Roeder et al.,2004). Interestingly, this extract inhibited a broad spectrum ofbiologic activities. Thus, Ph-ME strongly suppressed NF-�B acti-vation induced by PMA (Fig. 3A) and cotransfection with adaptormolecules such as MyD88 (Fig. 3B) and TRIF (Fig. 3C). Moreover,nuclear levels of p65 and p50, subunits of NF-�B, were remarkablydecreased by exposure to Ph-ME (Fig. 3D). The extract diminishedAP-1-mediated luciferase activities stimulated by PMA (Fig. 4A),MyD88 (Fig. 4B), and TRIF (Fig. 4C). In agreement with these results,the nuclear translocated levels of c-Jun and p-ATF2 were also
reduced (Fig. 4D). Finally, Ph-ME inhibited CREB activity inducedby forskolin (2 �M) as well as its phosphorylation in the nucleus(Fig. 5B). These results strongly indicate that Ph-ME is a drug can-didate that can target multiple anti-inflammatory mechanisms.
pharm
PsIofmci((r(tfiSaii
Fcp
Y. Yang et al. / Journal of Ethno
Further analysis of targeting was done by determining whetherh-ME modulates upstream events in the activation of tran-cription factors NF-�B, AP-1, and CREB in LPS/TLR4 signalling.nterestingly, this extract strongly suppressed the phosphorylationf I�B� (from 5 to 30 min, Fig. 6A), an important biochemical eventor the nuclear translocation of NF-�B (Wang et al., 2002). Further-
ore, Ph-ME strongly suppressed a series of upstream kinases thatontribute to the activation of I�B�. Indeed, this extract clearlynhibited the phosphorylation of IKK and Akt from 5 to 30 minFig. 6A). The phosphorylation of TLR-associated tyrosine kinasesSrc and Syk) and their downstream substrate p85/PI3K were alsoemarkably suppressed by Ph-ME at early time points (2 and 3 min)Fig. 6B), suggesting that the activation of these enzymes could beargets of Ph-ME. In fact, direct kinase assay performed with puri-ed enzymes demonstrated this possibility. Namely, the activity of
yk and Src, which play a critical role in regulation of TLR signallingnd inflammatory responses (Hu et al., 2007; Kim et al., 2010), wasnhibited by this extract up to 95% at 200 �g/ml (Fig. 6C), imply-ng that the inhibition of Syk and Src by Ph-ME could explain itsig. 9. Analysis of the phytochemical profile of Ph-ME and the inhibitory activity of quercharacteristics of Ph-ME were analyzed by high performance liquid chromatography (HProduction in LPS-treated RAW264.7 cells was examined by Griess assay and EIA. *P < 0.0
acology 139 (2012) 616– 625 623
NF-�B suppressing activity. Indeed, piceatannol, a Syk inhibitor,and PP2, a Src inhibitor, strongly suppressed NF-�B-dependent pro-duction of inflammatory mediators such as NO, PGE2, and TNF-�(Fig. 6D). Additionally, considering our report that early phase (at5 min) phosphorylation of I�B� is regulated by Syk activation andintermediate phase phosphorylation (30 and 60 min) is mediatedby Src (Lee et al., 2009), the inhibition of I�B� phosphorylation at5–30 min by Ph-ME strongly supports its simultaneous inhibitionof Syk and Src.
In addition, to explore molecular mechanisms of the inhibi-tion of AP-1 and CREB by Ph-ME, phosphorylation patterns of theirupstream proteins were evaluated. At an early time point (5 min),the phosphorylation of p38 was suppressed by Ph-ME (Fig. 7A).JNK phosphorylation at 15 and 30 min was also reduced by thisextract (Fig. 7A). Moreover, Ph-ME blocked ERK phosphorylation
at 30 min (Fig. 7A), indicating that MAPK can be time-dependentlysuppressed by this extract. The activation of upstream enzymes(MKK and TAK1) for MAPK signalling was also negatively modu-lated by Ph-ME, so that phosphorylation of MKK and TAK1 wasetin on NO and PGE2 production in LPS-treated RAW264.7 cells. (A) PhytochemicalLC), equipped with KNAUER. (B) The inhibitory effect of quercetin on NO and PGE2
5 and **P < 0.01 compared to the control group.

6 pharm
svdsd(gstctiiENTtf
osiDtcb
c3go2qdMd
24 Y. Yang et al. / Journal of Ethno
uppressed by Ph-ME at 2–5 min (Fig. 7B). Finally, since the acti-ation of MKK and TAK1 is regulated by IRAK1 activation, weetermined whether Ph-ME suppresses IRAK1 activity by mea-uring its protein level. As Fig. 7C shows, LPS treatment inducedegradation of IRAK1, an essential pathway to activate this enzymeHuang et al., 2004) while Ph-ME blocked its degradation, sug-esting that Ph-ME-mediated inhibition of MAPK could be due touppression of IRAK1 activation. However, it has been found thathis extract did not suppress an initial signalling complex formationomposed of TLR4 and MyD88, according to immunoprecipita-ion (IP) and immunoblotting (IB) analysis. To confirm whethernhibition of these pathways contributes to Ph-ME-mediated anti-nflammatory actions, specific inhibitors of JNK (SP600125) andRK (U0126) were tested under the same conditions. Interestingly,O release was not blocked by these inhibitors, while both PGE2 andNF-� production was remarkably reduced, suggesting that Ph-MEargeted inhibition of JNK and ERK pathways could be importantor PGE2 and TNF-� production.
To show that in vitro anti-inflammatory activity of Ph-ME wasbserved in vivo, a DSS-induced colitis model was employed. Aseen in Fig. 8A and B, this extract strongly ameliorated the DSS-nduced decrease in colon length that had been triggered bySS-induced colon inflammation (Nishitani et al., 2009). Seven-day
reatment with Ph-ME (100 mg/kg) did not elicit a body weighthange (Fig. 8C), suggesting that there was no metabolic distur-ance by this extract.
Many anti-oxidative flavonoids, sesquiterpenoids, andoumarin compounds, including 7,4′-dimethylquercetin,′-methylquercetin, quercetin, isoquercitrin, polygodial, warbur-anal, hydropiperoside, rhamnazin, and persicarin are constituentsf Polygonum hydropiper (Horhammer and Rao, 1954; Kiem et al.,008; Yang et al., 2011; Sultana et al., 2011). Of these compounds,
uercetin has already been reported as a strong anti-inflammatoryrug with Syk and Src inhibitory activities (Lee et al., 2008).oreover, MAPK activation and subsequent translocation of AP-1uring UV and PMA treatment were reduced by quercetin (Lim and
Fig. 10. Putative inhibitory pathway of LPS-activated
acology 139 (2012) 616– 625
Kim, 2007). The activation of MAPK and AP-1 by insulin-inducedsignalling in hepatocytes was blocked by quercetin (Sidhu et al.,2001). These reports strongly suggest that quercetin is a majoractive anti-inflammatory ingredient in Ph-ME. In fact, HPLC analy-sis of Ph-ME strongly supported the idea that this compound is oneof the major ingredients. There was a clear peak around 19 min inthe chromatographic analysis of the methanol extract, and a peakwith a similar retention time was observed by HPLC analysis ofstandard quercetin. In a spiking experiment of combined sampleswith Ph-ME and quercetin (1:1), the area of the quercetin peakwas proportionally enhanced, suggesting that the peak similarlyseen for standard quercetin is clearly this compound. Indeed, ithas been reported that quercetin was included 1.3% in methanolextract of this plant leaves (Peng et al., 2003). Furthermore, it iswell known that quercetin is a representative anti-inflammatoryflavonoid with NF-�B and AP-1 suppressing activities by simulta-neously suppressing Syk, Src, ERK, p38, and JNK (Hou et al., 2007),although its molecular inhibitory mechanisms remain unknown.Indeed, under our conditions, quercetin significantly suppressedNO and PGE2 production in LPS-treated macrophages (Fig. 9B) aswell as luciferase activity mediated by NF-�B and AP-1 activation(Fig. 9C). Therefore, these data and reports strongly suggest thatPh-ME-mediated anti-inflammatory activity could be regulatedby the action of quercetin as an active principle. Nonetheless,since there are several peaks with similar retention times in HPLCprofile, additional works will be focused on the identification andisolation of flavonoid derivatives with multiple anti-inflammatoryactions.
In summary, we have demonstrated that Ph-ME can stronglyblock macrophage-mediated inflammatory responses such as theproduction of NO, TNF-�, and PGE2, during the activation of TLR4.In particular, this extract selectively suppresses the activation of
NF-�B, AP-1 and CREB by inhibition of their upstream signallingenzymes such as Src, Syk, TAK1, ERK, JNK, and IRAK1 as summarizedin Fig. 10. In vivo curative activity of Ph-ME also strongly impliesa great potential of this extract as an anti-inflammatory remedy.inflammatory signalling responses by Ph-ME.

pharm
Tnwddp
A
gf0
R
B
B
C
C
D
H
H
H
H
J
J
K
K
K
K
K
L
L
L
L
L
Y. Yang et al. / Journal of Ethno
herefore, it is suggested that Ph-ME should be developed as aovel anti-inflammatory remedy. To investigate this possibility, weill do additional in vivo efficacy tests using models of acute disor-ers (septic shock and carageenan-induced arthritis) and of chronicisorders (collagen- or adjuvant-induced arthritis) in a follow-upre-clinical study.
cknowledgements
This research was supported by the Basic Science Research Pro-ram through the National Research Foundation of Korea (NRF)unded by the Ministry of Education, Science and Technology (no.004975).
eferences
resnihan, B., 1999. Pathogenesis of joint damage in rheumatoid arthritis. Journalof Rheumatology 26, 717–719.
yeon, S.E., Lee, J., Lee, E., Lee, S.Y., Hong, E.K., Kim, Y.E., Cho, J.Y., 2009. Functionalactivation of macrophages, monocytes and splenic lymphocytes by polysaccha-ride fraction from Tricholoma matsutake. Archives of Pharmacal Research 32,1565–1572.
hung, S.H., Yuan, H.D., Kim, S.J., Quan, H.Y., Huang, B., 2010. Ginseng leaf extract pre-vents high fat diet-induced hyperglycemia and hyperlipidemia through AMPKactivation. Journal of Ginseng Research 34, 369–375.
ho, J.Y., Baik, K.U., Jung, J.H., Park, M.H., 2000. In vitro anti-inflammatory effects ofcynaropicrin, a sesquiterpene lactone, from Saussurea lappa. European Journalof Pharmacology 398, 399–407.
eban, L., Bottazzi, B., Garlanda, C., de la Torre, Y.M., Mantovani, A., 2009. Pentraxins:multifunctional proteins at the interface of innate immunity and inflammation.Biofactors 35, 138–145.
orhammer, L., Rao, S.B., 1954. Isolation of quercitrin from Polygonum hydropiperL. Archiv der Pharmazie und Berichte der Deutschen PharmazeutischenGesellschaft 287, 34–38.
ou, D.X., Masuzaki, S., Hashimoto, F., Uto, T., Tanigawa, S., Fujii, M., Sakata, Y.,2007. Green tea proanthocyanidins inhibit cyclooxygenase-2 expression in LPS-activated mouse macrophages: molecular mechanisms and structure–activityrelationship. Archives of Biochemistry and Biophysics 460, 67–74.
u, X., Chen, J., Wang, L., Ivashkiv, L.B., 2007. Crosstalk among Jak-STAT, Toll-likereceptor, and ITAM-dependent pathways in macrophage activation. Journal ofLeukocyte Biology 82, 237–243.
uang, Y., Li, T., Sane, D.C., Li, L., 2004. IRAK1 serves as a novel regulator essen-tial for lipopolysaccharide-induced interleukin-10 gene expression. Journal ofBiological Chemistry 279, 51697–51703.
in, M., Park, S., Pyo, M.Y., 2009. Suppressive effects of T-412, a flavone on interleukin-4 production in T cells. Biological & Pharmaceutical Bulletin 32, 1875–1879.
ung, K.K., Lee, H.S., Cho, J.Y., Shin, W.C., Rhee, M.H., Kim, T.G., Kang, J.H., Kim, S.H.,Hong, S., Kang, S.Y., 2006. Inhibitory effect of curcumin on nitric oxide pro-duction from lipopolysaccharide-activated primary microglia. Life Sciences 79,2022–2031.
iem, P.V., Nhiem, N.X., Cuong, N.X., Hoa, T.Q., Huong, H.T., Huong Le, M., Minh,C.V., Kim, Y.H., 2008. New phenylpropanoid esters of sucrose from Polygonumhydropiper and their antioxidant activity. Archives of Pharmacal Research 31,1477–1482.
im, J.R., Oh, D.R., Cha, M.H., Pyo, B.S., Rhee, J.H., Choy, H.E., Oh, W.K., Kim, Y.R.,2008. Protective effect of Polygoni cuspidati radix and emodin on Vibrio vulnificuscytotoxicity and infection. Journal of Microbiology 46, 737–743.
im, J.Y., Lee, Y.G., Kim, M.Y., Byeon, S.E., Rhee, M.H., Park, J., Katz, D.R., Chain, B.M.,Cho, J.Y., 2010. Src-mediated regulation of inflammatory responses by actinpolymerization. Biochemical Pharmacology 79, 431–443.
im, Y.O., Lee, S.W., 2011. Microarray analysis of gene expression by ginseng waterextracts in a mouse adrenal cortex after immobilization stress. Journal of Gin-seng Research 35, 111–123.
inne, R.W., Brauer, R., Stuhlmuller, B., Palombo-Kinne, E., Burmester, G.R., 2000.Macrophages in rheumatoid arthritis. Arthritis Research 2, 189–202.
ee, H.J., Hyun, E.A., Yoon, W.J., Kim, B.H., Rhee, M.H., Kang, H.K., Cho, J.Y., Yoo,E.S., 2006. In vitro anti-inflammatory and anti-oxidative effects of Cinnamomumcamphora extracts. Journal of Ethnopharmacology 103, 208–216.
ee, J.H., Kim, J.W., Ko, N.Y., Mun, S.H., Kim, D.K., Kim, J.D., Kim, H.S., Lee, K.R.,Kim, Y.K., Radinger, M., Her, E., Choi, W.S., 2008. Camellia japonica suppressesimmunoglobulin E-mediated allergic response by the inhibition of Syk kinaseactivation in mast cells. Clinical and Experimental Immunology 38, 794–804.
ee, Y.G., Byeon, S.E., Kim, J.Y., Lee, J.Y., Rhee, M.H., Hong, S., Wu, J.C., Lee, H.S., Kim,M.J., Cho, D.H., Cho, J.Y., 2007. Immunomodulatory effect of Hibiscus cannabinusextract on macrophage functions. Journal of Ethnopharmacology 113, 62–71.
ee, Y.G., Chain, B.M., Cho, J.Y., 2009. Distinct role of spleen tyrosine kinase in
the early phosphorylation of inhibitor of kappaB alpha via activation of thephosphoinositide-3-kinase and Akt pathways. International Journal of Biochem-istry and Cell Biology 41, 811–821.ey, S.V., 1990. Synthesis of antifeedants for insects: novel behaviour-modifyingchemicals from plants. Ciba Foundation Symposium 154, 80–87.
acology 139 (2012) 616– 625 625
Lim, H., Kim, H.P., 2007. Inhibition of mammalian collagenase, matrixmetalloproteinase-1, by naturally-occurring flavonoids. Planta Medica 73,1267–1274.
Michaelsson, E., Holmdahl, M., Engstrom, A., Burkhardt, H., Scheynius, A., Holm-dahl, R., 1995. Macrophages, but not dendritic cells, present collagen to T cells.European Journal of Immunology 25, 2234–2241.
Nishitani, Y., Tanoue, T., Yamada, K., Ishida, T., Yoshida, M., Azuma, T., Mizuno,M., 2009. Lactococcus lactis subsp. cremoris FC alleviates symptoms of colitisinduced by dextran sulfate sodium in mice. International Immunopharmacology9, 1444–1451.
Onitsev, P.I., 1962. Biological evaluation of preparations from Polygonum hydropiper.Aptechnoe Delo 11, 49–52.
Owens, T., Babcock, A.A., Millward, J.M., Toft-Hansen, H., 2005. Cytokine andchemokine inter-regulation in the inflamed or injured CNS. Brain Research BrainResearch Review 48, 178–184.
Park, J.C., Yu, Y.B., Lee, J.H., Hattori, M., Lee, C.K., Choi, J.W., 1996. Protective effect ofOenanthe javanica on the hepatic lipid peroxidation in bromobenzene-treatedrats and its bioactive component. Planta Medica 62, 488–490.
Pauwels, R., Balzarini, J., Baba, M., Snoeck, R., Schols, D., Herdewijn, P., Desmyter, J.,De Clercq, E., 1988. Rapid and automated tetrazolium-based colorimetric assayfor the detection of anti-HIV compounds. Journal of Virological Methods 20,309–321.
Peng, Z.F., Strack, D., Baumert, A., Subramaniam, R., Goh, N.K., Chia, T.F., Tan, S.N.,Chia, L.S., 2003. Antioxidant flavonoids from leaves of Polygonum hydropiper L.Phytochemistry 62, 219–228.
Pyo, S., 1994. The mechanism of poly I:C-induced antiviral activity in peritonealmacrophage. Archives of Pharmacal Research 17, 93–99.
Rahman, E., Goni, S.A., Rahman, M.T., Ahmed, M., 2002. Antinociceptive activity ofPolygonum hydropiper. Fitoterapia 73, 704–706.
Rhee, D.K., Kim, E.H., Lee, M.J., Kim, I.H., Pyo, S., Choi, K.T., 2010. Anti-apoptotic effectsof red ginseng on oxidative stress induced by hydrogen peroxide in SK-N-SHcells. Journal of Ginseng Research 34, 138–144.
Roeder, A., Kirschning, C.J., Schaller, M., Weindl, G., Wagner, H., Korting, H.C., Rupec,R.A., 2004. Induction of nuclear factor-kappa B and c-Jun/activator protein-1via toll-like receptor 2 in macrophages by antimycotic-treated Candida albicans.Journal of Infectious Diseases 190, 1318–1326.
Sekine, Y., Yumioka, T., Yamamoto, T., Muromoto, R., Imoto, S., Sugiyma, K., Oritani,K., Shimoda, K., Minoguchi, M., Akira, S., Yoshimura, A., Matsuda, T., 2006. Mod-ulation of TLR4 signaling by a novel adaptor protein signal-transducing adaptorprotein-2 in macrophages. Journal of Immunology 176, 380–389.
Sidhu, J.S., Liu, F., Boyle, S.M., Omiecinski, C.J., 2001. PI3K inhibitors reverse thesuppressive actions of insulin on CYP2E1 expression by activating stress-response pathways in primary rat hepatocytes. Molecular Pharmacology 59,1138–1146.
Starkenmann, C., Luca, L., Niclass, Y., Praz, E., Roguet, D., 2006. Comparison of volatileconstituents of Persicaria odorata (Lour.) Sojak (Polygonum odoratum Lour.) andPersicaria hydropiper L. Spach (Polygonum hydropiper L.). Journal of AgriculturalFood Chemistry 54, 3067–3071.
Stuhlmuller, B., Ungethum, U., Scholze, S., Martinez, L., Backhaus, M., Kraetsch, H.G.,Kinne, R.W., Burmester, G.R., 2000. Identification of known and novel genesin activated monocytes from patients with rheumatoid arthritis. Arthritis andRheumatism 43, 775–790.
Sultana, R., Hossain, R., Adhikari, A., Ali, Z., Yousuf, S., Choudhary, M.I., Ali, M.Y.,Zaman, M.S., 2011. Drimane-type sesquiterpenes from Polygonum hydropiper.Planta Medica 77, 1848–1851.
Sun, H., Lee, O.R., Kim, Y.J., Jeong, S.K., In, J.G., Kwon, W.S., Kim, S.Y., Yang, D.C., 2010.Identification of ‘Chunpoong’ among Panax ginseng cultivars using real time PCRand SNP marker. Journal of Ginseng Research 34, 47–50.
Takasaki, M., Kuroki, S., Kozuka, M., Konoshima, T., 2001. New phenylpropanoidesters of sucrose from Polygonum lapathifolium. Journal of Natural Product 64,1305–1308.
Takeda, K., Akira, S., 2001. Roles of Toll-like receptors in innate immune responses.Genes to Cells 6, 733–742.
Tsan, M.F., Gao, B., 2004. Endogenous ligands of Toll-like receptors. Journal of Leuko-cyte Biology 76, 514–519.
Wang, T., Zhang, X., Li, J.J., 2002. The role of NF-kappaB in the regulation of cell stressresponses. International Immunopharmacology 2, 1509–1520.
Yang, X., Wang, B.C., Zhang, X., Yang, S.P., Li, W., Tang, Q., Singh, G.K., 2011. Simultane-ous determination of nine flavonoids in Polygonum hydropiper L. samples usingnanomagnetic powder three-phase hollow fibre-based liquid-phase microex-traction combined with ultrahigh performance liquid chromatography-massspectrometry. Journal of Pharmaceutical and Biomedical Analysis 54, 311–316.
Yu, T., Lee, J., Lee, Y.G., Byeon, S.E., Kim, M.H., Sohn, E.H., Lee, Y.J., Lee, S.G., Cho, J.Y.,2010. In vitro and in vivo anti-inflammatory effects of ethanol extract from Acertegmentosum. Journal of Ethnopharmacology 128, 139–147.
Yu, T., Lee, Y.J., Jang, H.J., Kim, A.R., Hong, S., Kim, T.W., Kim, M.Y., Lee, J.,Cho, J.Y., 2011a. Anti-inflammatory activity of Sorbus commixta water extractand its molecular inhibitory mechanism. Journal of Ethnopharmacology 134,493–500.
Yu, T., Lee, Y.J., Yang, H.M., Han, S., Kim, J.H., Lee, Y., Kim, C., Han, M.H., Kim, M.Y.,Lee, J., Cho, J.Y., 2011b. Inhibitory effect of Sanguisorba officinalis ethanol extract
on NO and PGE2 production is mediated by suppression of NF-kappaB and AP-1activation signaling cascade. Journal of Ethnopharmacology 134, 11–17.Zhang, G., Ghosh, S., 2000. Molecular mechanisms of NF-kappaB activation inducedby bacterial lipopolysaccharide through Toll-like receptors. Journal of EndotoxinResearch 6, 453–457.





























