Embed Size (px)
Citation preview

1
Development of in vitro techniques as supportive
tools for breeding and mass clonal propagation of
Leucocoryne spp. (Amaryllidaceae)
Alejandro Félix Altamira Bravo
2016

2
Pontificia Universidad Católica de Chile Facultad de Agronomía e Ingeniería Forestal
Development of in vitro techniques as supportive tools for breeding and mass clonal propagation of Leucocoryne spp.
(Amaryllidaceae)
Alejandro Félix Altamira Bravo
Thesis to obtain the degree of
Doctor
Ciencias de la Agricultura
Santiago, Chile, December 2016

3
Thesis presented as part of the requirements for the degree of Doctor en Ciencias de la Agricultura, approved by the
Thesis Committee
_____________________ Gloria Montenegro, Advisor
__________________ Dr. Patricio Arce Johnson
___________________ Dr. Levi Mansur
Santiago, December 2016

4
In memory of my grandmother Adela, with whom I spent many days learning to appreciate and propagate plants during my childhood.

5
This work was supported by CONICYT doctoral grant number 21110937
and FIA Project PYT-2012-0079.

6
ACKOWLEDGEMENTS
I would like to thank all those who were members of my committee, Professors
Gloria Montenegro, Patricio Arce, Levi Mansur and Eduardo Olate for all their
valuable contributions in the development of this thesis.
I would also like to thank Professor Marlene Gebauer for her infinite support and
for allowing me to finish this thesis in her laboratory.
I also thank the ex-members of the “Laboratorio de cultivo in vitro y Ornamentales”
with whom I shared during the development of this thesis, especially Nicole Arenas
and Gonzalo Gutierrez for their friendship and support.
Thanks to my parents Sergio and Mónica and my sisters Camila and Josefa for
their love and unconditional support.
And especially I want to thank my partner Mario for all these years of friendship,
company, support and challenges that we have overcome together.

7
CONTENTS
CHAPTER 1.
General Introduction ........................................................................... 8
CHAPTER 2.
An efficient clonal micropropagation protocol for Leucocoryne, a
geophyte genus endemic of Chile .................................................... 26
CHAPTER 3.
An efficient in vitro production system for Leucocoryne spp. plants
initiated from seeds .......................................................................... 68
CHAPTER 4.
Conclusions ...................................................................................... 92

8
CHAPTER 1
GENERAL INTRODUCTION

9
1. Description of Leucocoryne genus.
Leucocoryne genus belongs to the Amaryllidaceae family and is endemic to Chile.
There are 17 described taxons in the genus (15 species and 2 subspecies),
however its taxonomic classification is still confusing due to its high morphological
variability and the occurrence of natural hybrids. Consequently some authors
mention up to 45 species as part of the genus (Muñoz and Moreira 2000;
Riedemannn and Aldunate 2001; Mansur 2002; Zoellner 2002; Mansur and
Cisternas 2005; Zuloaga et al. 2009; Olate and Schiappacasse 2013). The natural
life cycle of Leucocoryne can vary depending on the species but, in general, the
plant takes three years from seed to floral bulb size (Riedemannn and Aldunate
2001; Mansur 2002).
In general, this genus presents high adaptation to dry climates and is widely
distributed along Chile, from lat 20°S in deserted zones until lat 41°S in more rainy
temperate zones, with populations ranging from mild coastal habitats to more
colder mountainous areas where the soil is cover with snow for several months, at
altitudes of 2.800 m.a.s.l. in the Andes mountains. However, most of the
Leucocoryne populations grow in coastal areas in Central Chile (lat 30°-35°S), in
dry climates that show a marked rainy season from May to August and a 70 mm
average annual rainfall (Zoellner 1972; Kim et al. 1998; Mansur et al. 2004; Olate
and Schiappacasse 2013).
Plants of this genus are considered geophytes due the presence of an
underground storage structure, which in the case of Leucocoryne corresponds to a

10
tunicated bulb. These bulbs have a spherical or oval shape, reaching 1.5 to 2.5 cm
in diameter, covered by brown dry membranes and having up to twenty fleshy
scales on the inside. Bulbs have a basal plate from which adventitious roots
develop (Hartmann et al. 1997; Zoellner 2002).
Leaves have a flaccid, glabrous and semi-fleshy appearance. From the bulbs a
single floral scape grow whose height can vary between 30 and 80 cm. Its apical
inflorescence it is an umbel that can have from 3 - 4 to 8 - 12 flowers depending on
the species, although 5 - 15 flowers have also been described. Flowers are perfect,
with the stigma and stamens attached to the floral tube, with 2.5 - 6 cm in diameter,
six tepals 1.4 - 2 cm in length exhibiting colors from white, blue-sky, purple, violet
and others (Zoellner 1972; Mansur 2002; Schiappacasse et al. 2002).
The androecium is composed of three or six fertile stamens and three fleshy
staminodes (infertile stamens) 6 mm in length, with color varying between white,
yellow, greenish or two-colored. The gynoecium has a cylindrical superior ovary,
with a short style and a capitate stigma. Fruit corresponds to a tricarpellary
dehiscent capsule, and each carpel has several seeds. In general, 15 - 40 seeds
per fruit are produced, which are small with sizes of 0.1 - 0.2 cm in diameter
(Muñoz and Moreira 2000; Mansur 2002; Schiappacasse et al. 2002; Zoellner
2002; Verdugo 2013).
Besides the diversity in shape and colors, it has been genetically verified the
existence of diversity in terms of chromosomal number, which can vary from 2n =
10 in Leucocoryne purpurea to 2n = 18 in Leucocoryne coquimbensis, with hybrids
showing 2n = 14, 2n = 20 y 2n = 22 (Mansur 2002; Araneda et al. 2004; Salas and

11
Mansur 2004). Also, it has been reported six Leucocoryne species as self-
incompatible, which would explain the high genetic variability (Mansur 2002;
Mansur et al. 2004).
2. Advances in Leucocoryne domestication and commercial use.
The Leucocoryne genus is known in Chile by the common name "Huilli" and
internationally as "Glory-of-the-Sun". Because the high phenotypic variability of
flowers (colors, shapes and aroma) and also because its long vase life,
Leucocoryne has become a great alternative as an ornamental plant, either as cut
flower, potted or garden plant (Bridgen 2000; Mansur 2002; Olate and
Schiappacasse 2013; De la Cuadra et al. 2016).
The ornamental value of the different species has led to several studies and
domestication efforts and even incipient commercialization of them in countries like
Japan, Netherlands, Israel, New Zealand and USA (Bridgen 2000; Lancaster et al.
2000; Walton et al. 2008; Olate and Schiappacasse 2013; De la Cuadra et al.
2016). This interest is similar to the one occurred with Alstroemeria genus, which
has been used by companies in other countries, for selecting, breeding and
commercialization with high economic success, using Chilean germplasm. These
are examples of importance of identifying, selecting, propagating, breeding and
registration of plant material belonging to Chilean native genetic resources as
Leucocoryne, which could also lead to economic benefits and improve the
conservation of endangered populations (Jorquera et al. 2007).

12
In order to take advantage of the commercial potential and also to conserve
Leucocoryne, which has already been affected by antrophic intervention of their
environment, conservation programs have been developed in Chile. Sustainable
germplasm banks, genetic and agronomic studies for future uses in landscaping,
and also breeding programs for releasing new cut-flower cultivars have been
developed by researchers at the Pontificia Universidad Católica de Valparaíso. To
date, three cut-flower cultivars ('Elena', 'Gabriela' and 'Paulina') have been
patented (Verdugo and Teixeira 2006). Nevertheless, an efficient mass
propagation system is still a pending challenge for successfully commercialization
of these new cultivars (Bridgen 2000; Mansur 2002; Araneda et al. 2004; Olate and
Schiappacasse 2013).
3. Propagation techniques
3.1 Seed propagation
Seed propagation of Leucocoryne naturally occurs when seeds are released from
its dehiscent capsule sometime during the spring. Seeds stay on the soil surface
during all summer and part of the autumn until the next rainy season (autumn and
winter), with temperatures ranging 10 to 15 °C (Mansur 2002; De la Cuadra et al.
2016). Once germination occurs, plants develop a single leaf and after 45 days a
small bulb starts to grow. This bulb can weight 0.06 g in average after 100 days
since germination, and then it enters in dormancy until the next season. During the
next autumn these small bulbs could produce shoots even in absence of water,

13
using for that their reserves getting then ready for the winter rains. During this
second growing season two leaves will appear and the plant will stay active for 100
- 120 days producing more bulb weight increase until it becomes dormant again
until next rainy season. Only during the third or fourth growing season the bulbs will
reach 0.3 g in weight and the flowering process will happen and new seeds will be
produced (Riedemannn and Aldunate 2001; Mansur 2002; De la Cuadra and
Mansur 2004).
Germination studies have been conducted on L. coquimbensis, L. ixioides and L.
purpurea, which have determined that water imbibition for one day and subsequent
cold stratification at 7°C for 7 weeks results in higher germination rates (over 90%)
compared to non-stratified seeds (Salazar 2001; Schiappacasse et al. 2002).
Similar studies have also achieved high germination rates by imbibition of
Leucocoryne seeds for 96 h and subsequent culture at 10 - 15°C, without a
stratification period, higher than germination observed at 20°C and no germination
at 25°C (Jara et al. 2006; De la Cuadra et al. 2016).
Due to the high genetic variability of Leucocoryne and self-incompatibility in some
of its species, seed propagation techniques have great disadvantages to maintain
interesting traits and massive propagation of new registered cultivars. Despite this,
seed propagation is still used as a valuable technique in breeding programs to
obtain new plant segregants (Mansur 2002; Mansur et al. 2004; De la Cuadra et al.
2016).

14
3.2 Vegetative propagation
Vegetative propagation consists in obtaining new plants from other plant organs
like stems, leaves, roots, bulbs, corms and rhizomes. The main advantage related
to this techniques is that new developed plants are genetically identical to the
propagated plant (Schiappacasse et al. 2002; Kumar et al. 2010). In Leucocoryne,
as in other geophyte genus, natural vegetative propagation occurs in the second or
third year of growth, producing 1 - 2 lateral bulblets every one or two years
depending on the genotype. In some species and in particular environmental
conditions the plants can produce even more deep bulbs called "droppers"
(Riedemannn and Aldunate 2001; Mansur 2002; Schiappacasse et al. 2002).
In some tunicate bulb species artificial propagation techniques allow to increase
natural multiplication rates. These techniques are known as bulb-cutting methods
and they consist essentially in to generate a mechanical damage to the basal plate
or to the entire bulb. "Scooping" is one of these techniques that involve the
complete elimination of the basal plate, the apical meristem and the base of the
scales (Fig. 1b). In the "scoring" method, deep cuts to the base of the bulb are
done in order to damage its basal plate and the apical meristem (Fig. 1c). "Coring"
is other of these methods, and consists in the removal of a cylindrical section of the
basal plate, including the apical meristem (Fig. 1d). In other species it is used the
method called "sectioning", where the entire bulb is cut in two or more vertical
sections in order to produce adventitious bulblets (Fig. 1e) (Hartmann et al. 1997;
Schiappacasse et al. 2002; Olate and Bridgen 2005).

15
Fig. 1 Artificial propagation techniques used in some bulb species. a) Intact bulb, b) Scooping, c) Scoring, d) Coring, and e) "Sectioning". Adapted from Kumar et al. (2010).
Studies in Leucocoryne coquimbensis have determined that sectioning a bulb in
equal sections could produce up to 2.2 bulbs of different sizes, while in
Leucocoryne ixiodes up to 5 new bulbs have developed from a singular initial bulb
(Schiappacasse et al. 2002). Nevertheless, in a similar study, L. ixioides
multiplication rates of intact bulbs were compared to bulbs multiplied using
scooping and scoring, founding no significant differences between the natural and
artificial methods (Salazar 2001). These results do not coincide with the successful
results observed in Hyacinthus and Scilla species, where scooping techniques
produce much higher multiplication rates than natural propagation, and it is used
for commercial production (Hartmann et al. 1997).
3.3 In vitro propagation
Micropropagation, or in vitro propagation, corresponds to the culture of explants
under aseptic conditions, onto a nutritive medium, under controlled temperature,
humidity and luminosity conditions, to produce high number of clonal plants in a

16
reduced period of time (Roca and Mroginski 1991; Hartmann et al. 1997; Kyte et al.
2013).
In vitro shoot growth can be achieved using direct organogenesis from axillary or
adventitious buds, or using indirect organogenesis from cells, cell suspensions or
calluses. However, indirect organogenesis has the disadvantage of increasing the
risk of producing genetically non true-to-type plants (somaclonal variation) and thus
affecting the homogeneity of the plants produced (George et al. 2008).
Micropropagation has several stages to obtain new plants. Stage 0 includes the
selection of the genotype and the initiation explant. Stage I corresponds to the
disinfection and establishment of the explant in aseptic culture conditions. Once
the explants are stabilized in vitro conditions, mass plant multiplication could be
performed (Stage II). Stage III of the micropropagation process is where the in vitro
rooting is achieved. Finally, once the plants are fully developed, plants are
transferred to ex vitro conditions to be culture into soil or potting soil in order to
start their acclimatization (Stage IV). Duration and conditions for each of these
stages must be adapted to different plant genotypes (George et al. 2008; Bach and
Sochacki 2012; Kyte et al. 2013).
In contrast with other ornamental geophyte species, only a few studies of in vitro
culture have been done in Leucocoryne sp. One of the main barriers hindering a
wider use of these techniques in geophyte species, is the high contamination
occurring during the in vitro initiation of bulbs, corms, rhizomes and tubers,
because their permanent contact with soil and high level of microorganisms, as
fungi and bacteria (Pedersen and Brandt 1992; Slabbert et al. 1993; Kritzinger et

17
al. 1998; Smith et al. 1999; Chang et al. 2003; Sochacki and Orlikowska 2005;
Paredes et al. 2014). Previous works in Leucocoryne species have reported
contamination rates between 42% and 100% during the in vitro initiation of bulbs
(Fuentevilla 2004). In order to surpass this problem, previous studies involving
Leucocoryne sp. Bulbs have used seeds as initial explants with the consequent
problem of uncertain genotypic homogeneity of the plants used (Ham 2002;
Briones 2003; Escobar et al. 2008). In one of these studies different concentrations
of MS (Murashige and Skoog 1962) were tested as germination media for L.
purpurea seeds (i.e. 12.5%, 25%, 37.5%, 50%, 75% and 100%) resulting in higher
germination rates when seeds were cultured onto 75% MS or lower (Ham 2002).
Escobar et al. (2008) studied indirect organogenesis in L. purpurea by using MS
medium supplemented with 30 g·L-1 sucrose and 0.6% agar, pH 5.7 and a
combination of different plant growth regulators: 2,4-Dichlorophenoxyacetic (2,4-
D), 6-bencyladenine (BA), 1-Phenyl-3-(1-2-3-thiadiazol-5-yl)-urea (TDZ), α-
naphthaleneacetic acid (ANA), 6-y-y-(dimethylallyamino)-purine (2iP) and 4-
amino-3,5,6-trichloropicolinic acid (Picloram). Callus proliferation and shoot
formation was achieved. Then, after seven months of culture, 3 - 4 bulbs/explant
were obtained when subculture was done in absence of plant growth regulators.
Also, in previous studies (Olate and Bridgen 2005; Olate 2006), using MS medium
supplemented with 30 g·L-1 sucrose and 0.7% agar and pH adjusted to 5.7, it was
observed that ligh/darkness and temperature treatments does not affect bulb
production in L. purpurea, L. coquimbensis and L. ixioides. In the same study, in
vitro bulb-cutting methods were also tested. An average of almost 7 bulbs/explants

18
were obtained using a scoring method in L. coquimbensis bulbs, and over 8
bulbs/explant were produced by sectioning the bulbs in four, contrasting with
scarce occurrence of new bulblets on intact bulbs.
4 Hypothesis and objectives.
In terms of commercial potential and native germplasm concerns there is an
obvious need to develop asexual propagation techniques to assist either plant
breeding and conservation programs of Leucocoryne spp. One of the techniques
that provide several advantages over sexual propagation is the clonal
micropropagation. This system could avoid the long life cycle from seed to
flowering bulb and the low vegetative propagation rates that naturally occur in
these species. In this sense, hybrids or interesting clonal lines would benefit from
an in vitro propagation system, using direct organogenesis for example, to
propagate reliable and large populations without altering the genetic and
phenotypic characteristics of the original germplasm. Therefore, future studies
regarding the in vitro multiplication of Leucocoryne spp. should focus on increasing
clonal multiplication rates and on the optimal environmental conditions for the
growth and development of the explants.
In this context, we formulated the following hypothesis:

19
4.1 Hypothesis.
Adjustment and optimization of the culture media, plant growth regulators, explant
type and growing temperature produces a more effective initiation process and
higher multiplication rates during the in vitro propagation of Leucocoryne species,
thus improving breeding and conservation outcomes.
4.2 General objective.
To determine in vitro media culture, propagation techniques and
environmental conditions to increase the growth and propagation rate
of Leucocoryne species to support breeding and conservation programs.
4.3 Specific objectives.
1. To determine the effect of culture media and system on in vitro bulb propagation rates of Leucocoryne sp.
2. To establish the effect of plant growth regulators on in vitro bulb propagation of Leucocoryne sp. bulbs
3. To evaluate the effect of bulb-cutting methods on in vitro bulb propagation rates of Leucocoryne sp.
4. To determine the effect of culture temperature on in vitro seed germination of Leucocoryne sp.

20
References.
Araneda L, Salas P, Mansur L (2004) Chromosome numbers in the Chilean
endemic genus Leucocoryne (Huilli). J Am Soc Hortic Sci 129:77–80.
Bach A, Sochacki D (2012) Propagation of Ornamental Geophytes. In: Ornamental
Geophytes. CRC Press, pp 261–286
Bridgen M (2000) Flowering geophytes of Chile have ornamental potential. In:
Seeds: Trade, Production and Technology. pp 218–230
Briones C (2003) Leucocoryne sp. Pontificia Universidad Católica de Chile
Chang HS, Chakrabarty E, Hahn EJ, Paek KY (2003) Micropropagation of Calla lily
(Zantedeschia albomaculata) via in vitro shoot tip proliferation. Vitr Cell Dev
Biol—Plant 39:129–134. doi: 10.1079/IVP2002362
De la Cuadra C, Mansur L (2004) DESCRIPCIÓN DE LA PRIMERA ETAPA DEL
CICLO DE VIDA DE TRES GENOTIPOS DE Leucocoryne sp.: SEMILLA A
BULBO. Agric Técnica. doi: 10.4067/S0365-28072004000200009
De la Cuadra C, Vidal A, Lefimil S, Mansur L (2016) Temperature Effect on Seed
Germination in the Genus Leucocoryne (Amaryllidaceae ). HortScience
51:412–415.
Escobar L, Jordan M, Olate E, et al (2008) Direct and indirect in vitro
organogenesis of leucocoryne pupurea (Alliaceae) a chilean ornamental
geophyte. Propag Ornam Plants 8:59–64.
Fuentevilla C (2004) Propagación in vitro de algunas especies de Leucocoryne.

21
Pontificia Universidad Católica de Valparaíso
George EF, Hall MA, Klerk G-J De (2008) Plant Propagation by Tissue Culture.
Springer Netherlands, Dordrecht
Ham F (2002) Evaluación de la germinación in vitro de Leucocoryne coquimbensis
F. Phil., geófita enémica con potencial ornamental. Pontificia Universidad
Católica de Chile
Hartmann HT, Kester DE, Davies FT, Geneve RL (1997) Plant Propagation:
Principles and Practices., Sixth edit. Prentice-Hall, Inc., New Jersey, USA
Jara P, Arancio G, Moreno R, Carmona M (2006) Factores abióticos que
influencian la germinación de seis especies herbáceas de la zona árida de
Chile. Rev Chil Hist Nat 79:309–319.
Jorquera C, Moreno R, Arancio G (2007) Domesticación de recursos nativos con
fines comerciales y de conservación: potencial de uso múltiple de una
bullbosa endémica de chile, Leucocoryne spp. Rev Bras Agroecol 2:1512–
1517.
Kim HH, Ohkawa K, Nitta E (1998) EFFECTS OF BULB WEIGHT ON THE
GROWTH AND FLOWERING OF LEUCOCORYNE COQUIMBENSIS F.
PHILL. Acta Hortic 341–346. doi: 10.17660/ActaHortic.1998.454.40
Kritzinger EM, Vuuren RJ Van, Woodward B, et al (1998) Elimimation of external
and internal contaminants in rhizomes of Zantedeschia aethiopica with
commercial fungicides and antibiotics. Plant Cell 61–65. doi:
10.1023/A:1005937016338

22
Kumar GNM, Guse WE, Larsen FE (2010) Propagation of Plants from Specialized
Structures. Pacific Northwest Ext. Publ. 12.
Kyte L, John K, Scoggins H, Bridgen M (2013) Plants from test tubes: An
Introduction to Micro-Propagation, Fourth. Timber press, London
Lancaster JE, Shaw ML, Walton EF (2000) S-Alk(en)yl-l-cysteine sulfoxides,
alliinase and aroma in Leucocoryne. Phytochemistry 55:127–130. doi:
10.1016/S0031-9422(00)00245-4
Mansur L (2002) Ciclo de vida, autoincompatibilidad y plasticidad genética en
diseño y color de flores. In: Leucocoryne un género nativo chileno y su uso
como planta de jardín, 1st edn. Oficina de transferencia tecnológica
Universidad Católica de Valparaíso, Viña del Mar, Chile, pp 9–15
Mansur L, Cisternas M (2005) Leucocoryne talinensis ( Alliaceae ), a New Species
from Chile. Novon 15:324–326.
Mansur L, Gonzalez M, Rojas I, Salas P (2004) Self-incompatibility in the Chilean
Endemic Genus. J Amer Soc Hort Sci 129:836–838.
Muñoz M, Moreira A (2000) Los Géneros Endémicos de Monocotiledóneas de
Chile Continental.
http://www.chlorischile.cl/monocotiledoneas/leucocoryne.htm. Accessed 8 Nov
2016
Murashige T, Skoog F (1962) A Revised Medium for Rapid Growth and Bio Assays
with Tobacco Tissue Cultures. Physiol Plant 15:473–497. doi: 10.1111/j.1399-
3054.1962.tb08052.x

23
Olate E (2006) Chilean Geophytes: Micropropagation and Cut Flower Production.
University of Connecticut
Olate E, Bridgen M (2005) Techniques for the In Vitro Propagation of Rhodophiala
and Leucocoryne spp. Acta Hortic 673:335–342. doi:
10.17660/ActaHortic.2005.673.42
Olate E, Schiappacasse F (2013) Geophyte Research and Production in Chile. In:
Ornamental Geophytes. CRC Press, pp 449–470
Paredes K, Delaveau C, Carrasco P, et al (2014) In vitro bulbing for the
propagation of Traubia modesta (Amaryllidaceae), a threatened plant endemic
to Chile. Cienc e Investig Agrar 41:13–14. doi: 10.4067/S0718-
16202014000200007
Pedersen C, Brandt K (1992) A METHOD FOR DISINFECTION OF
UNDERGROUND RHIZOME TIPS OF ALSTROEMERIA AND HELICONIA.
Acta Hortic 499–504. doi: 10.17660/ActaHortic.1992.325.69
Riedemannn P, Aldunate G (2001) Flora Nativa de valor Ornamental Chile Zona
Centro, Identificación y Propagación, 1st edn. Editorial Andrés Bello, Santiago,
Chile
Roca W, Mroginski L (1991) Cultivo de Tejidos en la Agricultura. Ciat 969.
Salas P, Mansur L (2004) Gene Flow between Parents with Different Ploidy Levels
in a Natural Population of Leucocoryne. J Amer Soc Hort Sci 129:833–835.
Salazar R (2001) Efecto de la estratificación de semillas de Leucocoryne ixioides,

24
L. coquimbensis, L purpurea y de distintas técnicas de Propagación
Vegetativa en Leucocoryne ixioides. Universidad de Talca
Schiappacasse F, Peñailillo P, Yáñez P (2002) Propagación de Bulbosas Chilenas
Ornamentales. Editorial Universidad de Talca, Talca, Chile
Slabbert MM, de Bruyn MH, Ferreira DI, Pretorius J (1993) Regeneration of
bulblets from twin scales of Crinum macowanii in vitro. Plant Cell Tissue
Organ Cult 33:133–141. doi: 10.1007/BF01983226
Smith RH, Burrows J, Kurten K (1999) Challenges associated with
micropropagation of Zephyranthes and Hippesatrum sp. (Amaryllidaceae). Vitr
Cell Dev Biol - Plant 35:281–282. doi: 10.1007/s11627-999-0032-y
Sochacki D, Orlikowska T (2005) The obtaining of narcissus plants free from
potyviruses via adventitious shoot regeneration in vitro from infected bulbs. Sci
Hortic (Amsterdam) 103:219–225. doi: 10.1016/j.scienta.2004.04.018
Verdugo G (2013) FLOWER BREEDING OF NATIVE PLANTS: THE CHILEAN
EXPERIENCE. Acta Hortic 1000:401–405. doi:
10.17660/ActaHortic.2013.1000.55
Verdugo G, Teixeira JA (2006) From Wild to the Table : Leucocoryne and From
Wild to the Table : Leucocoryne and Chloraea. In: Floriculture, Ornamental
and Plant Biotechnology Volume IV. Global Science Books, pp 356–360
Walton EF, Wu RM, Sutherland P (2008) Bulb and inflorescence development in
Leucocoryne ixioides (Hook.) Lind. J Hortic Sci Biotechnol 83:574–580.

25
Zoellner O (2002) Descripción del género Leucocoryne (Alliaceae) su forma de
propagación y distribución. In: Leucocoryne un género nativo chileno y su uso
como planta de jardín, 1st edn. Oficina de transferencia tecnológica
Universidad Católica de Valparaíso, Viña del Mar, Chile, pp 23–27
Zoellner O (1972) El género Leucocoryne. An del Mus Hist Nat Valparaíso 5:9–83.
Zuloaga F, Morrone O, Belgrano M (2009) Catálogo de las plantas vasculares del
cono sur (Argentina, southern Brazil, Chile, Paraguay y Uruguay). In: Inst.
Botánica Darwinion, CONICET.
http://www2.darwin.edu.ar/Proyectos/FloraArgentina/fa.htm. Accessed 8 Nov
2016

26
CHAPTER 2
AN EFFICIENT CLONAL MICROPROPAGATION PROTOCOL FOR
Leucocoryne, A GEOPHYTE GENUS ENDEMIC TO CHILE
Alejandro Altamira1, Eduardo Olate1, Marlene Gebauer1, Levi Mansur2 and Gloria
Montenegro1
1Departamento de Ciencias Vegetales, Facultad de Agronomía e Ingeniería
Forestal, Pontificia Universidad Católica de Chile. Vicuña Mackenna 4860, Macul,
Santiago, Chile.
2Escuela de Agronomía, Pontificia Universidad Católica de Valparaíso, Avda. San
Francisco s/n, La Palma, Quillota, Chile
This chapter was sent to Plant Cell, Tissue and Organ Culture (PCTOC) - Journal of Plant Biotechnology – Springer (December 2016)

27
An efficient clonal micropropagation protocol for Leucocoryne, a geophyte
genus endemic to Chile
Alejandro Altamira1*, Eduardo Olate1, Marlene Gebauer1, Levi Mansur2 and Gloria
Montenegro1.
1 Departamento de Ciencias Vegetales, Facultad de Agronomía e Ingeniería Forestal,
Pontificia Universidad Católica de Chile. Vicuña Mackenna 4860, Macul, Santiago, Chile.
2 Escuela de Agronomía, Pontificia Universidad Católica de Valparaíso, Avda. San
Francisco s/n, La Palma, casilla 4-D, Quillota, Chile
*Corresponding author:
Name: Alejandro Altamira
Address: Departamento de Ciencias Vegetales, Facultad de Agronomía e Ingeniería
Forestal, Pontificia Universidad Católica de Chile. Vicuña Mackenna 4860, Macul,
Santiago, Chile
Telephone: +56 9 90915256
E-Mail: [email protected]
Keywords: Amaryllidaceae, bulb, plant tissue culture, cytokinin, culture media, cut flower.

28
Abstract
Leucocoryne (Amaryllidaceae) is a geophyte genus endemic to Chile with
exceptional characteristics to use it as cut flower, pot or garden plant. The
objective of this work is to develop and improve an in vitro protocol for mass clonal
propagation to assist breeding and conservation efforts. We evaluated different in
vitro culture systems (agar, cotton), nutrient media (MS, LS, B5), BAP
concentrations (0.5, 1.0, 1.5 and 2.0 mgL-1) and different type of explants (intact
bulb, scoring and sectioning) in four Leucocoryne genotypes (Leucocoryne
purpurea, L. vittata, L. ixioides and L. sp. Pichicuy). Using an improved disinfection
process it was possible to initiate bulbs in vitro with a low contamination rate of 7%.
High bulb sprouting percentages were achieved, with values between 80 – 100%.
There was a higher rate of bulb multiplication and higher shoot production when
BAP was added to the growing media. Bulb-cutting methods resulted in the highest
multiplication rates reported to date, obtaining 11-16 bulblets per sectioned bulb,
depending on the genotype used.
Abbreviations: BAP - N6-benzylaminopurine; IAA - indole-3-acetic acid; MS –
Murashige and Skoog medium (1962); LS - Linsmaier and Skoog medium (1965);
B5 – Gamborg et al. medium (1968); Benomyl - Methyl-1-(butylcarbamoyl)-2-
benzimidazole carbamate; Captan - N-(triclorometiltio) ciclohex-4-eno-1,2-
dicarboximida.

29
Introduction
Leucocoryne is a geophyte genus endemic to Chile that belongs to
Amaryllidaceae. It is commonly known as "Huilli" or "Glory-Of-The-Sun" (Mansur
2002; Zoellner 2002; Sassone et al. 2014).
These plants present exceptional characteristics to be used as cut flowers, potted
or landscape plants due to their long life in vase and wide variety of shapes,
designs and colors (Mansur 2002; De la Cuadra et al. 2016).
There are 17 taxa (15 species and 2 subspecies) in the genus. However,
classification of Leucocoryne can be very confusing due to its high variability in
morphology and the occurrence of natural hybrids (Zoellner 1972; Muñoz and
Moreira 2000; Mansur and Cisternas 2005; Olate and Schiappacasse 2013;
Sassone et al. 2014; Jara-Arancio et al. 2014). Plants develop a small tunicate
bulb 1.5-2.5 cm in diameter, thin leaves and a 1-3 floral scape 30-80 cm in height
arranged in a terminal umbel. Its colorful flowers are actinomorphic and trimerous
(6 tepals) varying in quantity from 3-4 to 8-12 depending on the species colors
ranging from white to sky-blue, to purple and violet, forming different patterns and
designs depending on the species. A distinctive characteristic of Leucocoryne
flowers is the presence of three prominent staminodes (sterile stamens) that
protrude from the central part of the corolla (Mansur 2002; Schiappacasse et al.
2002; Zoellner 2002; Olate and Bridgen 2005; Hoffmann et al. 2015).

30
Although Leucocoryne plants have been cultivated in European gardens since the
19th century (Muñoz and Moreira 2000), in the 1990’s the ornamental industry
started to show interest to produce these species commercially, in order to satisfy
the constant demand for new species and cultivars with new shapes and colors.
The latter has led to several research efforts, looking for the domestication and
commercialization of these species, in countries like Japan, Israel, The
Netherlands and New Zealand (Kim et al. 1998; Lancaster et al. 2000; Catley
2003; Walton et al. 2008; Olate and Schiappacasse 2013; Hoffmann et al. 2015).
In the same way, several studies have been conducted in Chile on Leucocoryne
sp., including botanical identification; chromosomal studies; self-incompatibility
processes; life cycle; plant propagation and breeding techniques. These Chilean
studies have been carried out primarily by the Pontificia Universidad Católica de
Valparaíso that has been able to register three new cultivars in addition to the ones
offered from The Netherlands (Bridgen et al. 2002; De la Cuadra and Mansur
2004; Verdugo and Teixeira 2006; Verdugo 2013; Olate and Schiappacasse 2013;
De la Cuadra et al. 2016).
Seeds are usually used for Leucocoryne propagation. However, as in other
geophytes species, such as Narcissus and Tulipa, these species have a very long
life cycle and juvenile period, requiring between 3 and 4 years to form a floral size
bulb from seed (Riedemannn and Aldunate 2001; Mansur 2002; Escobar et al.
2008; Bach and Sochacki 2012). Because it is self-incompatible (Mansur 2002;
Mansur et al. 2004), this genus presents a wide range of genetic variability and
hence, natural populations with individuals showing segregation can be easily

31
found. This fact offers a great potential for breeding purposes but at the same time,
it makes difficult to propagate heterozygous lines having traits of interest by sexual
reproduction (Mansur 2002). Seed propagation, as the most important technique
used for breeding and conservational purposes, has been largely studied in
aspects like germination requirements, and other aspects related to the growth and
development of the plants (De la Cuadra et al. 2002; De la Cuadra and Mansur
2004; De la Cuadra et al. 2016).
Until now vegetative propagation is the best way to maintain over time the genetic
characteristics of Leucocoryne plants, either naturally inherited or produced by
controlled crosses. This techniques allow to produce new individuals with identical
genotype to the original plant from which the propagule was obtained (Mansur
2002; Schiappacasse et al. 2002). It has been reported that natural vegetative
propagation occurs in Leucocoryne species, from either a two or three-year old
plant, which will produce lateral bulblets or droppers (a type of bulblet that grows
deep in the ground) developed from the base of the mother bulb (Riedemannn and
Aldunate 2001; Mansur 2002). Currently, bulblet separation is the propagation
technique commercially used, in spite of its low multiplication rates and its
dependence with the plant genotype (Escobar et al. 2008). On the other hand,
there are artificial techniques that enhance and accelerate the vegetative
propagation capacity in geophytes. These are essentially bulb-cutting methods
consisting in different levels and types of controlled mechanical damages
depending on the genotype to propagate. In the case of the tunicate bulbs,
sectioning, scoring, scooping and coring have been used (Schiappacasse et al.

32
2002). Schiappacasse et al (2002) conducted studies in Leucocoryne
coquimbensis and Leucocoryne ixioides, where the use of bulb sectioning made it
possible to increase the bulb-multiplication rates.
In Leucocoryne, as in other geophytes, natural bulblet multiplication rates are very
low due to the reduced number of axillary meristems existing in modified stems as
bulbs, corms and rhizomes (Bach and Sochacki 2012). In addition to bulb-cutting
methods, there are more sophisticated techniques like in vitro propagation, also
known as micropropagation, by which potentially higher multiplication rates can be
achieved (mass clonal propagation). Micropropagation is composed by several
techniques by which the totipotency of plant cells is used. During micropropagation
small plant pieces or explants are cultured under aseptic conditions onto a media
supplemented with nutrients and plant growth regulators. The explants are then
cultured in an environment with controlled light, temperature and humidity. Using
micropropagation, plant growers are capable to supply stocks for
commercialization by multiplying elite cultivars in large scale without altering the
genotype (Rout et al. 2006; George et al. 2008; Sharma and Agrawal 2012; Bach
and Sochacki 2012). This technique has five stages: Stage 0, which corresponds
to the selection of the genotype and the explant selection and preparation. Stage I
involves the disinfection of the explants and their in vitro establishment. During
Stage II the explants are mass propagated by different means and techniques.
During Stage III the in vitro rooting of the explants is achieved. Finally, in Stage IV
fully developed plants are acclimatized to ex vitro conditions. Duration and

33
conditions for each of these stages must be adapted to each plant genotype
(George et al. 2008; Bach and Sochacki 2012; Kyte et al. 2013).
In vitro culture has been widely used in several ornamental plants as Narcissus,
Lilium, Gladiolus, Freesia, Zantedeshia, Crocus, Hippeastrum, Allium, Tulipa and
Hyacinthus, among others (Bach and Sochacki 2012). In the case of Leucocoryne
only a few studies have been conducted on the in vitro propagation of these
species and specific conditions for a successful in vitro culture are still poorly
understood (Olate and Bridgen 2005; Verdugo and Teixeira 2006; Escobar et al.
2008).
In this context, the objective of this research is to improve the in vitro propagation
techniques for Leucocoryne species and cultivars, to obtain a more efficient
method of mass clonal propagation.
Materials and methods.
Plant material.
Dormant bulbs of Leucocoryne purpurea, L. ixioides, L. vittata and the ecotype
‘Pichicuy’ (reported as L. aff. vittata) were harvested from the Leucocoryne
breeding program of the Pontificia Universidad Católica de Valparaiso (Fig.1a-d).
Bulbs of 1.0 ± 0.2 g in weight of each genotype were selected and stored at room
temperature under dry conditions until their in vitro initiation.

34
Disinfection and in vitro establishment.
In order to establish the plant material under in vitro conditions, the dry protective
tunics of the bulbs were eliminated and bulbs were washed under running tap
water for 5 min. Then, the bulbs were submerged in a fungicide solution of 1gL-1
Benomyl® and 1gL-1 Captan® for 30 min, rinsed with distilled water and washed
with ethanol 95% for 1 min. Subsequently, the explants were disinfected under a
laminar flow chamber using a solution of 50 g·L-1 NaOCl with addition of 2 drops of
Tween® 20, during 30 min and constant agitation. Bulbs were rinsed three times in
sterile distilled water.
After the sterilization all the bulb storage scales and the outer basal plate section
were completely remove from the explant until exposing the vegetative growing
point. Thus, each explant consisted by the apical growing point and a section of the
basal plate (Fig.1e)
The explants were established in culture tubes containing culture media
supplemented with 30 gL-1 sucrose and pH adjusted to 5.7, and previously
sterilized in autoclave at 121°C for 15 min. Macro and micronutrient composition of
the media and the use of a gelling agent varied depending on the experiment.
Explants were cultured under growing chamber conditions at 20±1°C, 16 h
photoperiod and luminous intensity of 125 µmol·m-2·s-1.

35
Experiment 1: Effect of the basal media and BAP on the in vitro growth and
multiplication of Leucocoryne.
To determine a proper culture media for the in vitro propagation of Leucocoryne
purpurea, L. ixioides and L. vittata explants of each of these species were
established in 20 mL of three different basal culture media: MS (Murashige and
Skoog 1962), LS (Linsmaier and Skoog 1965) y B5 (Gamborg et al. 1968). The
media cultures were solidified by adding 0.6% agar. Half of the treatments were
supplemented with BAP 1.0 mgL-1. Each treatment consisted of fifteen individual
explants as repetitions. We evaluated the percentage of bulb sprouting as the
bulbs that developed sprouts, percentage of single shoot as the bulbs that
developed only one shoot, percentage of multiple shoots as the bulbs that
developed two or more shoots, percentage of bulb multiplication as the bulbs that
developed lateral bulblets, bulb production as the number of bulbs obtained per an
initial bulb and bulb fresh weight as the average final weight obtained per explant.
Experiment 2: Effect of the BAP concentration on the in vitro growth and
multiplication of Leucocoryne.
In order to determine an adequate BAP concentration for the in vitro growth and
multiplication of Leucocoryne twenty explants of L. purpurea, L. ixioides and L.
vittata were established onto 20 mL of MS medium solidified with 0,6% agar and
supplemented with 0 (control), 0.5, 1.0, 1.5 and 2.0 mg·L-1 of BAP. Percentage of
bulb sprouting, percentage of single shoot, percentage of multiple shoots,

36
percentage of bulb multiplication, bulb production and bulb fresh weight were
evaluated as described above.
Experiment 3: Effect of bulb-cutting methods, culture system and the addition of
BAP on the in vitro growth and propagation of Leucocoryne vittata.
L. vittata in vitro cultured-bulbs were used as explants and cultured in MS medium.
To evaluate two different in vitro bulb-cutting methods: scoring and sectioning.
Scoring consisted in vertical incisions in the basal plate of the bulbs in order to
damage the main growing point and thus to break the apical dominance (Fig. 1g).
Sectioning consisted in to divide each bulb longitudinally into four isolated identical
parts (Fig. 1h). Cutting treatments were compared to intact bulbs as controls (Fig.
1f). In addition to the bulb-cutting methods two different culture systems were
compared: semi-solid (0.6% agar) and liquid media imbibed in a cotton pad. Bulbs
were cultured onto MS medium, and half of the treatments were supplemented with
1.0 mgL-1 BAP. Five bulbs of 0.8+0.1 g were used in each treatment as replicates.
Percentage of bulb sprouting, percentage of bulb multiplication and bulb production
were evaluated as described above.
Experiment 4: Effect of BAP on the in vitro growth and multiplication of
Leucocoryne sectioned bulbs
In vitro bulbs of L. purpurea, L. ixioides, L. vittata and L. sp. Pichicuy of 0.8+0.1 g
in weight were sectioned in four parts and used as explants (Fig. 1h). Five

37
replicates per treatment were used, each replicate consisted of the four sections
that were obtained from the cut of the same bulb. Each of these four sections were
cultured in the same container onto MS medium solidified with 0.6% agar, and half
of the treatments were supplemented with 1.0 mgL-1 BAP. Percentage of bulb
sprouting, percentage of bulb multiplication, bulb production and bulb fresh weight
were evaluated as described above.
Statistical analysis.
Results of shooting and multiplication were statistically analyzed using Scheffé´s
procedure for multiple comparison of proportions (P≤0.05) (Zar 2010). In the case
of bulb production and bulb fresh weight data were analyzed performing an
analysis of variance (ANOVA) followed by Bonferroni's multiple comparison test
(P≤0.05) using GraphPad Prism 5 software (GraphPad Software Inc., San Diego,
California, USA).

38
Results.
Experiment 1: Effect of the basal media and BAP on the in vitro growth and
multiplication of Leucocoryne.
The methods of disinfection and establishment in vitro resulted in a successful
initiation of the explants, with a contamination level of only 7% after 4 weeks of
culture.
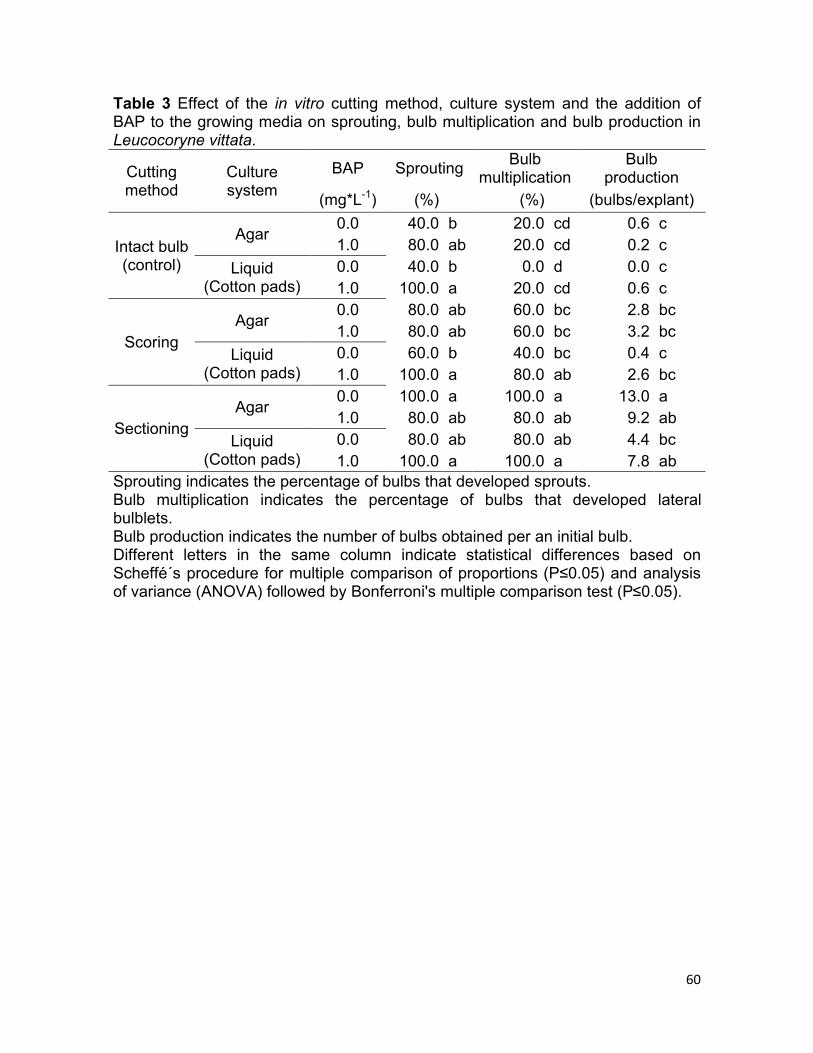
Bulbs with vegetative sprouting were in average 91.9% of the total, varying
between 80 and 100% depending on the treatment. L. purpurea and L. ixioides
explants cultivated onto any of the three culture media assayed presented 100%
bulb sprouting when they were supplemented with 1.0 mgL-1 BAP. In the case of L.
vittata that level of vegetative sprouting was only achieved when MS, B5 and B5 +
1.0 mgL-1 BAP media was used (Table 1). After 13 weeks of culture the addition of
BAP produced the highest bulb sprouting with multiple shoots per bulb. This
difference between the controls and the media supplemented with BAP was even
clearer in L. ixiodes bulbs but not in the other genotypes (Table 1). After 30 weeks
of culture different multiplication rates were observed among the genotypes. In
general, L. ixioides bulbs showed the highest multiplication rate, reaching up to
100% when cultivated onto MS + 1.0 mgL-1 BAP medium (Fig. 2a). In this
treatment it was produced an average of 5.3 bulbs/explant, which was higher than
in MS without BAP, which produced a multiplication rate of 30.8% and only 1.9
bulbs/explant (Fig.2b).

39
In most of the genotype/medium combinations bulb multiplication was produced
except in the case of L. purpurea, where no multiplication of the bulbs cultured onto
B5 + 1.0 mgL-1 BAP medium was observed (Fig. 2a). The latter treatment also
produced the lowest bulb fresh weight. In the case of L. ixioides the highest values
of bulb fresh weight occurred in MS + 1.0 mgL-1 BAP, while in L. vittata no
difference among treatments was observed (Fig. 2c).
Experiment 2: Effect of the BAP concentration on the in vitro growth and
multiplication of Leucocoryne.
In this experiment the in vitro establishment protocol also resulted in a very low
contamination rate (7.7%) after 4 weeks of culture.
A high bulb sprouting rate (98.2% average) was produced, varying between 90 and
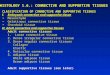
100% depending on the treatment (Table 2). L. purpurea and L. vittata sprouting
rates did not show any significant differences when different BAP concentrations
were added to the culture media. On the other hand, L. ixioides showed the lowest
sprouting rate (90%) when 1.0 mg·L-1 BAP was added to the medium (Table 2).
This genotype also showed a significant higher rate of multiple shoots by the
addition of BAP to the media compared to the control without BAP. However, no
difference was observed between the different BAP concentrations used (Table 2).
In the other two genotypes no clear tendency was observed between treatments in
terms of single or multiple shoot production after 13 weeks of culture (Table 2).

40
Multiplication varied according to the genotype and BAP concentration. The
highest multiplication rates were obtained in L. ixioides with values between 47.9%
(control plants) and 94.4% when bulbs were cultured onto MS + 2.0 mg·L-1 BAP
(Fig. 3a; Fig. 4). In this genotype it was also observed the highest bulb production
per explant, with 6.2 and 5.7 bulbs/explant when cultured onto MS plus 1.0 or 2.0
mg·L-1 BAP respectively. These results were significantly higher than the ones
measured in the control treatment, with only 2.9 bulbs/explant produced. In L.
purpurea and L. vittata no differences were observed between the different
treatments (Fig. 3b).
The highest bulb fresh weight was produced in L. purpurea and L. ixioides when
BAP was added to the culture media. In the case of L. vittata no significant
difference was observed among treatments (Fig. 3c).
Experiment 3: Effect of bulb-cutting methods, culture system and the addition of
BAP on the in vitro growth and propagation of Leucocoryne vittata.
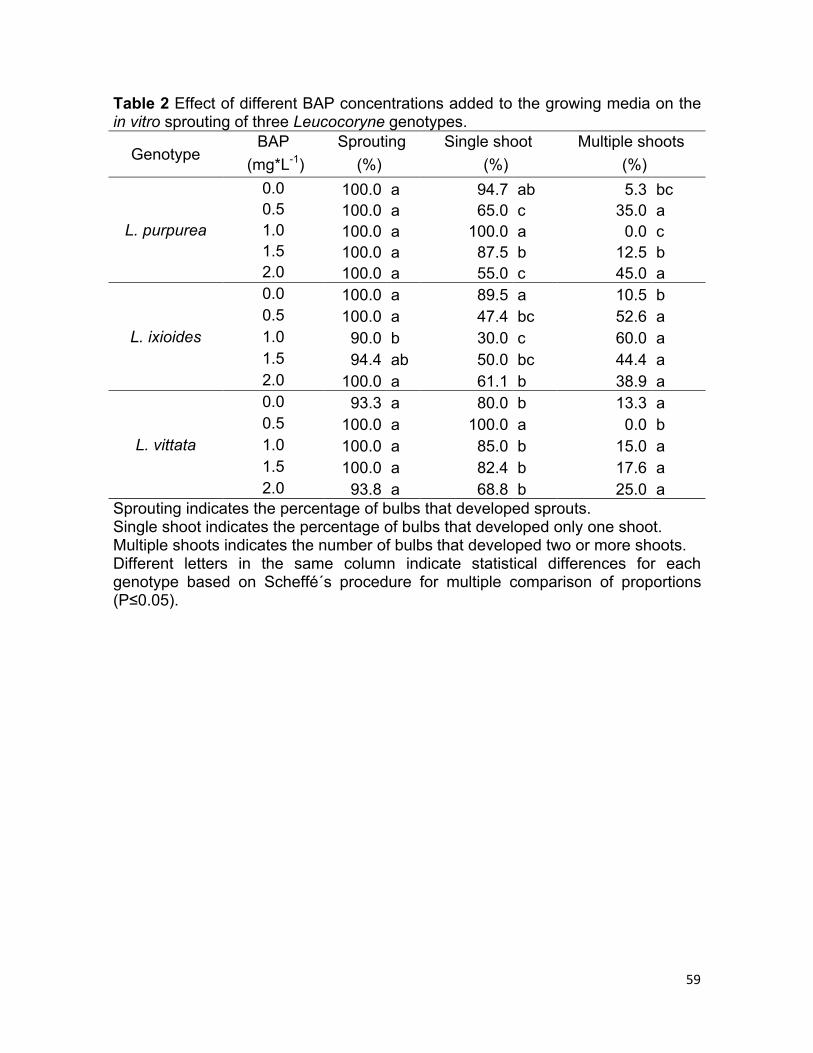
After 8 weeks of culture 100% of sprouting was observed in either intact (control)
or treated bulbs with one of the cutting methods, when they were cultured onto
liquid media imbibed in cotton pads and supplemented with 1.0 mg·L-1 BAP (Table
3). The latter was also observed in sectioned bulbs and cultured onto MS media
solidified with agar and supplemented with 1.0 mg·L-1 BAP. The lowest sprouting
rates (40%) were produced in intact bulbs, when they were cultured onto media
supplemented with BAP, independently of the system used (Table 3).

41
A tendency to increase the multiplication rate was observed when bulb-cutting
treatments were used, especially in those including sectioning (Figure 5). Intact
bulb treatments showed multiplication rates below 20% (Table 3). The highest bulb
production occurred in sectioned bulbs cultured onto media solidified with agar
either supplemented or not with BAP, producing 13 and 9.2 bulbs/explant
respectively. Independently of the culture system used and the addition or not of
BAP, the lowest bulb production was clearly observed in intact bulbs, showing
values between 0 y 0.6 bulbs/explant (Table 3). In terms of culture system,
differences between treatments were observed only when sectioned bulbs were
cultured onto media without BAP. In this case, bulbs cultured on media with agar
produced significantly higher number of bulbs than the ones cultured on cotton
pads imbibed with liquid media, with values of 13 and 4.4 bulbs/explant
respectively (Table 3).
Experiment 4: Effect of BAP on the in vitro growth and multiplication of
Leucocoryne sectioned bulbs.
Considering the results from experiment 3, sectioning was used on four
Leucocoryne genotypes cultured onto growing media either supplemented or not
with BAP. In L. purpurea sprouting rates of 66.6% were observed in control plants
and 50% when BAP was added to the media, but showing no statistical difference
between those groups of plants. On the other hand, L. ixioides and L. vittata plants
showed higher sprouting rates reaching 80% and 100% but there was no influence
of BAP addition to the growing media. In the case of the plants belonging to the L.

42
sp. Pichicuy only 40% of them sprouted in the absence of BA, but 80% of them did
sprout when BAP was added to the media.
In terms of multiplication, after 6 months of culture it was possible to observe
different rates between genotypes, although no statistical difference was founded
when data were analyzed (Fig. 6a). Thus bulb production varied considerably
among genotypes from 2.6 to 16 bulbs/explant (Fig. 7). In L. sp. Pichicuy bulb
production was significantly larger in BAP treatments compared to the controls,
showing values previously mentioned. In L. purpurea and L vittata differences were
also observed but they all remained below the threshold for statistical difference. In
the last genotype, L. ixioides, no differences in bulb production were also observed
among treatments, with values between 12.4 and 14 bulbs/explant for the controls
and BAP treatments, respectively (Fig. 6b).
The addition of BAP to the growing media did not affect the fresh weight of the
bulbs. Values varied between 0.07 and 0.19 g/bulb with no significant differences
between treatments (Fig 6c).
Discussion.
We developed an effective disinfection and in vitro establishment protocol for
Leucocoryne bulbs (Stage 0 and 1). Explant disinfection is one of the most
challenging steps in the process of the in vitro clonal propagation. The protocol to
use will depend on the type and origin of the explant and its sensitivity to

43
disinfectant agents (Smith et al. 1999; Kyte et al. 2013). In addition, the
combination of explant, culture media and controlled environment gives the perfect
conditions for the proliferation of any pathogenic and non-pathogenic
microorganism. Therefore, a proper disinfection system must be implemented to
avoid both explant contamination and extensive tissue damage (Mroginski and
Roca 1991). The disinfection protocol developed in this work resulted in a very
effective and safe protocol for the in vitro establishment of Leucocoryne bulbs,
showing in average only 7% of contamination. This is quite remarkable knowing
the major difficulty that is faced when establishing vegetative tissues that grow on
the ground or even underground in direct contact with soil, such as bulbs, corms,
rhizomes and tubers. In such cases, disinfection protocols should be labor-
intensive and more aggressive for the explant, using combinations of agents and/or
longer disinfections periods (Pedersen and Brandt 1992; Slabbert et al. 1993;
Kritzinger et al. 1998; Smith et al. 1999; Chang et al. 2003; Sochacki and
Orlikowska 2005; Paredes et al. 2014). Studies in other geophytes species have
also reported the difficulty and importance of the disinfection process. In vitro
establishment of Zantedeschia sp. has evidenced low success when using
conventional disinfection techniques (Kritzinger et al. 1998; Chang et al. 2003). In
the case of Narcissus sp., which possess a tunicate bulb similar to Leucocoryne,
problems during the in vitro initiation have been also reported, with high bacterial
and fungal contamination rates, surpassed in some cases only by using hot water
treatments plus use of fungicides (Sochacki and Orlikowska 2005). High
contamination rates have been also reported in other bulb species: 20-40% in
Crinum macowanni (Slabbert et al. 1993), 90% in Eucomis autumnalis (Ault 1995),

44
20-90% in Hippeastrum sp. (Smith et al. 1999) and 50-80% in Traubia modesta,
another endemic species to Chile with ornamental potential (Paredes et al. 2014).
The disinfection protocol developed in this work resulted in low contamination
rates, eventhough it is a more aggressive and laborious procedure than the
conventional ones, survival of the explants was not negatively affected given the
high sprouting rates obtained (80%). In previous studies including Leucocoryne
bulbs, in vitro contamination has been the main limitation for further advance in a
suitable protocol, with values between 40-100% (Fuentevilla, unpublished). Due to
this problem, the few in vitro studies on Leucocoryne micropropagation have
started their plant material from seeds (Escobar et al. 2008).
The different experiments of this work resulted in the development of a suitable
protocol for mass clonal propagation for the Leucocoryne genus. Among the
different media tested, basal MS medium was the most suitable for in vitro growth
and multiplication. It was observed high levels of sprouting and a normal growth
and development of the explants. Previous works on the in vitro propagation of
Leucocoryne have only used MS medium (Escobar et al., 2008; Olate & Bridgen,
2005). The other two media used in this work, LS and B5, also showed to be
suitable for Leucocoryne in vitro culture but in the case of B5 media a lower fresh
weight of the bulbs was obtained. These three media have previously been used
for the micropropagation of other geophyte species. For instance, MS has resulted
in better results than B5 in terms of embryo germination in Alstroemeria (Lu and
Bridgen 1996) and also on callus formation and shoot differentiation in Lilium
longiflorum (Ramsay et al. 2003). Besides, MS medium has been widely used in

45
different ornamental geophytes species, for example, Zhephyra elegans (Vidal et
al. 2012), Narcissus sp. (Sochacki and Orlikowska 2005), Hippeastrum sp. (Smith
et al. 1999), Muscari muscarimi (Ozel et al. 2015) and Zephyranthes sp. (Smith et
al. 1999). B5 medium has been successfully used in other geophytes from the
Allium genus (Dunstan and Short 1977; Shahidul Haque et al. 1997; Barandiaran
et al. 1999; Martınez et al. 2000) in the same way as LS medium have been used
in Allium sp. (Nagakubo et al. 1993; Ayabe and Sumi 1998), Crocus sp. (Mir et al.
2011) and Narcissus sp. (Stone 1973).
The addition of BAP to the culture media contributed to a higher multiplication rate
like the one observed on L. ixioides. However, no significant differences were
observed over the sprouting and multiplication rates when using different BAP
concentrations. Bulb production values of 6.2 and 5.7 bulbs/explants were possible
to obtain when plants were established on MS medium supplemented with 1 or 2
mg·L-1 BAP respectively, which contrast with the much lower values observed in
natural populations. In nature Leucocoryne bulbs are able to produce 1-2 bulblets
per plant every other year but only when they have reached their mature stage
(Riedemannn and Aldunate 2001). A bulb would reach their maturity from seed
only after 2 or 4 years of growth (Riedemannn and Aldunate 2001; Mansur 2002).
We have also concluded that the addition of BAP to the growing media produced a
larger explant fresh weight in L. purpurea and L. ixioides than the controls. In
previous studies Escobar et al. (2008) reported obtaining 3-4 bulbs/explant after
seven months of culture of Leucocoryne plants onto MS medium without any plant
growth regulators. That particular study produced the in vitro bulbs via indirect

46
organogenesis and further induction of the bulbs on MS supplemented with 1.0
mg·L-1 BAP. In Allium cepa micropropagation, it has been reported that 1.0 mg·L-1
BAP added to the growing media resulted in the best treatment, however higher
BAP concentrations resulted in a lower production of new plantlets (Kamstaityte
and Stanys 2004). In Muscari azureum 1.0 mg·L-1 BAP plus 0.25 mg·L-1 IAA
resulted in a larger shoot and bulblet production (Uranbey 2011). Bridgen et al.
(2009) also reported the use of MS medium supplemented with 2.0 mg·L-1 BAP for
an adequate Alstroemeria sp. micropropagation.
Bulb-cutting methods allow accelerating or increasing the efficiency of vegetative
propagation of some geophyte species. These methods include bulb division or
"sectioning", "scoring", "scooping" and "coring". All these methods are focused in a
controlled damage of the main growing point and basal plate and thus favor new
bulblet production (Hartmann et al. 1997; Kumar et al. 2010; Schiappacasse et al.
2002). Hyacinthus is an important plant genus in which these techniques have
been commercially adopted, producing up to 60 bulblets from a single bulb using
scooping, although these bulblets will require four to five years of growth before
flowering (Hartmann et al. 1997). In many other bulb species it has been also
reported successful use of this kind of technique. van Leeuwen and van der
Weijden (1997) using sectioning was able to increase the natural propagation rates
not only in Chionodoxa (from 1.9 to 6.5), but also in Gelanthus (2.7 to 6.5), Muscari
(2.7 to 10.0) and Scilla (1.1 to 8.0). Species with larger bulbs like those belonging
to Hippeastrum, allow a more intensive sectioning obtaining up to 28.5 bulbs when
the initial bulb has been divided in 24 sections (Sandler-Ziv et al. 1997). Scoring

47
has been reported to be a successful technique in Nerine (Mori et al. 1997) and in
Crinum x powellii (Knippels 2012). In the present work we observed a higher
multiplication rate when bulb-cutting methods were applied compared to intact
bulbs, however by using scoring we obtained a lower bulb multiplication and
production rates than sectioning. Similar results were observed by Solgi et al.
(2015) in Fritillaria imperialis where the number of bulbs obtained from an initial
bulb was greater by using sectioning compared to scoring. Schiappacasse et al.
(2002) working with traditional propagation reported the production of 2.2 bulblets
per initial bulb in L. coquimbensis and 5 bulblets in L. ixioides by using sectioning
in two vertical sections. By using in vitro bulb sectioning we obtained up to 11.3
bulbs/explant in L. purpurea and 14 bulbs/ explant in L. vittata and L.ixioides. In the
case of L. sp. Pichicuy, the addition of BAP significantly increased bulb production
from 2.6 to 16 bulbs/explant. Previously Olate and Bridgen (2005) reported on
Leucocoryne coquimbensis 7 bulbs/explant using scoring, 8 bulbs using a 4-
sectioned bulb and almost no bulblets from intact bulbs, but in that case no BAP
was added to the media.
When comparing bulb-cutting methods and culture system in L. vittata, a higher
bulb production was achieved using MS medium solidified with agar than using
liquid MS media imbibed in cotton pads. This result contradicts the potential
advantages of the liquid media in terms of producing a higher multiplication rate
reported by Kim et al. (2003) in Allium sativum.
In conclusion, in this work we have developed a highly efficient mass clonal
micropropagation of Leucocoryne. Our protocol also includes an effective

48
disinfection and explant preparation method for a successful in vitro establishment.
By using different bulb-cutting methods, culture media and growth regulator
addition, we also have been able to culture four different Leucocoryne genotypes,
achieving the highest multiplication rates reported to date. These results represent
an important set of tools to be used by conservational or commercial purposes,
such as breeding programs or nurseries. These techniques may also contribute to
the advance in the knowledge of the propagation of other native and ornamental
geophyte species.

49
References.
Ault JR (1995) In vitro propagation of Eucomis autumnalis , E . comosa , and E.
zambesiaca by twin-scaling. HortScience 30:1441–1442.
Ayabe M, Sumi S (1998) Establishment of a novel tissue culture method, stem-disc
culture, and its practical application to micropropagation of garlic (Allium
sativum L.). Plant Cell Rep 17:773–779. doi: 10.1007/s002990050481
Bach A, Sochacki D (2012) Propagation of Ornamental Geophytes. In: Ornamental
Geophytes. CRC Press, pp 261–286
Barandiaran X, Martín N, Rodríguez-Conde MF, et al (1999) Genetic variability in
callus formation and regeneration of garlic (Allium sativum L.). Plant Cell Rep
18:434–437. doi: 10.1007/s002990050599
Bridgen M, Kollman E, Chunsheng Lu (2009) Interspecific hybridation of
Alstroemeria for the development of new ornamental plants. Acta Hortic
836:73–78. doi: 10.17660/ActaHortic.2009.836.9
Bridgen MP, Olate E, Schiappacasse F (2002) Flowering geophytes from Chile.
Acta Hortic 570:75–80. doi: 10.17660/ActaHortic.2002.570.6
Catley JL (2003) Temperature and Irradiance Effects on Flowering of Two Species
of Leucocoryne. J Am Soc Hortic Sci 128:809–814.
Chang HS, Chakrabarty E, Hahn EJ, Paek KY (2003) Micropropagation of Calla lily
(Zantedeschia albomaculata) via in vitro shoot tip proliferation. Vitr Cell Dev
Biol—Plant 39:129–134. doi: 10.1079/IVP2002362

50
De la Cuadra C, Mansur L (2004) Descripción de la primera etapa del ciclo de vida
de tres genotipos de Leucocoryne sp.: semilla a bulbo. Agric Técnica. doi:
10.4067/S0365-28072004000200009
De la Cuadra C, Mansur L, Verdugo G, Arriagada L (2002) Deterioro de las
semillas de Leucocoryne spp. en función del tiempo de almacenaje. Agric
Técnica 62:3–5. doi: 10.4067/S0365-28072002000100005
De la Cuadra C, Vidal A, Lefimil S, Mansur L (2016) Temperature Effect on Seed
Germination in the Genus Leucocoryne (Amaryllidaceae ). HortScience
51:412–415.
Dunstan DI, Short KC (1977) Improved Growth of Tissue Cultures of the Onion ,
Allium cepa. Physiol Plant 41:70–72. doi: 10.1111/j.1399-3054.1977.tb01525.x
Escobar L, Jordan M, Olate E, et al (2008) Direct and indirect in vitro
organogenesis of Leucocoryne pupurea (Alliaceae) a chilean ornamental
geophyte. Propag Ornam Plants 8:59–64.
Gamborg OL, Miller R a., Ojima K (1968) Nutrient requirements of suspension
cultures of soybean root cells. Exp Cell Res 50:151–158. doi: 10.1016/0014-
4827(68)90403-5
George EF, Hall MA, Klerk G-J De (2008) Plant Propagation by Tissue Culture.
Springer Netherlands, Dordrecht
Hartmann HT, Kester DE, Davies FT, Geneve RL (1997) Plant Propagation:
Principles and Practices., Sixth edit. Prentice-Hall, Inc., New Jersey, USA

51
Hoffmann A, Watson J, Flores AR (2015) Flora Silvestre de Chile, Cuando el
Desierto Florece Vol.I. Monocotiledóneas, 1st edn. Ediciones Fundación
Claudio Gay, Zapallar, Chile
Jara-Arancio P, Arroyo MTK, Guerrero PC, et al (2014) Phylogenetic perspectives
on biome shifts in Leucocoryne (Alliaceae) in relation to climatic niche
evolution in western South America. J Biogeogr 41:328–338. doi:
10.1111/jbi.12186
Kamstaityte D, Stanys V (2004) Micropropagation of onion ( Allium Allium cepa L
.). Acta Univ Latv Biol 676:173–176.
Kim E, Hahn E, Murthy H, Paek K (2003) High frequency of shoot multiplication
and bulblet formation of garlic in liquid cultures. Plant Cell Tissue Organ Cult
73:231–236. doi: 10.1023/A:1023029302462
Kim HH, Ohkawa K, Nitta E (1998) Fall flowering of Leucocoryne coquimbensis F.
Phil. after long-term bulb storage treatments. HortScience 33:18–20.
Knippels PJM (2012) Advanced in Vivo Propagation Techniques for Specialty
Bulbs. Floric Ornam Biotechnol 6:154–157.
Kritzinger EM, Vuuren RJ Van, Woodward B, et al (1998) Elimimation of external
and internal contaminants in rhizomes of Zantedeschia aethiopica with
commercial fungicides and antibiotics. Plant Cell 61–65. doi:
10.1023/A:1005937016338
Kumar GNM, Guse WE, Larsen FE (2010) Propagation of Plants from Specialized
Structures. Pacific Northwest Ext. Publ. 12.

52
Kyte L, John K, Scoggins H, Bridgen M (2013) Plants from test tubes: An
Introduction to Micro-Propagation, Fourth. Timber press, London
Lancaster JE, Shaw ML, Walton EF (2000) S-Alk(en)yl-l-cysteine sulfoxides,
alliinase and aroma in Leucocoryne. Phytochemistry 55:127–130. doi:
10.1016/S0031-9422(00)00245-4
Linsmaier EM, Skoog F (1965) Organic Growth Factor Requirements of Tobacco
Tissue Cultures. Physiol Plant 18:100–127. doi: 10.1111/j.1399-
3054.1965.tb06874.x
Lu C, Bridgen M (1996) Effects of genotype, culture medium and embryo
developmental stage on the in vitro responses from ovule cultures of
interspecific hybrids of Alstroemeria. Plant Sci 116:205–212. doi:
10.1016/0168-9452(96)04385-3
Mansur L (2002) Ciclo de vida, autoincompatibilidad y plasticidad genética en
diseño y color de flores. In: Leucocoryne un género nativo chileno y su uso
como planta de jardín, 1st edn. Oficina de transferencia tecnológica
Universidad Católica de Valparaíso, Viña del Mar, Chile, pp 9–15
Mansur L, Cisternas M (2005) Leucocoryne talinensis ( Alliaceae ), a New Species
from Chile. Novon 15:324–326.
Mansur L, Gonzalez M, Rojas I, Salas P (2004) Self-incompatibility in the Chilean
Endemic Genus Leucocoryne Lindley. J Am Soc Hortic Sci 129:836–838.
Martınez LE, Agüero CB, López ME, Galmarini CR (2000) Improvement of in vitro
gynogenesis induction in onion (Allium cepa L.) using polyamines. Plant Sci

53
156:221–226. doi: 10.1016/S0168-9452(00)00263-6
Mir JI, Ahmed N, Wani SH, et al (2011) In vitro development of microcorms and
stigma like structures in saffron (Crocus sativus L.). Physiol Mol Biol Plants
16:369–373. doi: 10.1007/s12298-010-0044-4
Mori G, Hirai H, Imanishi H (1997) Vegetative propagation of Nerine bulbs by
cross-cutting. Acta Hortic 377–382. doi: 10.17660/ActaHortic.1997.430.58
Mroginski L, Roca WM (1991) Establecimiento de cultivos de tejidos vegetales in
vitro. Cultiv. tejidos en la Agric. Fundam. y Apl. 20–40.
Muñoz M, Moreira A (2000) Los Géneros Endémicos de Monocotiledóneas de
Chile Continental.
http://www.chlorischile.cl/monocotiledoneas/leucocoryne.htm. Accessed 8 Nov
2016
Murashige T, Skoog F (1962) A Revised Medium for Rapid Growth and Bio Assays
with Tobacco Tissue Cultures. Physiol Plant 15:473–497. doi: 10.1111/j.1399-
3054.1962.tb08052.x
Nagakubo T, Nagasawa A, Ohkawa H (1993) Micropropagation of garlic through in
vitro bulblet formation. Plant Cell Tissue Organ Cult 32:175–183. doi:
10.1007/BF00029840
Olate E, Bridgen M (2005) Techniques for the In Vitro Propagation of Rhodophiala
and Leucocoryne spp. Acta Hortic 673:335–342. doi:
10.17660/ActaHortic.2005.673.42

54
Olate E, Schiappacasse F (2013) Geophyte Research and Production in Chile. In:
Ornamental Geophytes. CRC Press, pp 449–470
Ozel CA, Khawar KM, Unal F (2015) Factors affecting efficient in vitro
micropropagation of Muscari muscarimi Medikus using twin bulb scale. Saudi
J Biol Sci 22:132–138. doi: 10.1016/j.sjbs.2014.09.007
Paredes K, Delaveau C, Carrasco P, et al (2014) In vitro bulbing for the
propagation of Traubia modesta (Amaryllidaceae), a threatened plant endemic
to Chile. Cienc e Investig Agrar 41:13–14. doi: 10.4067/S0718-
16202014000200007
Pedersen C, Brandt K (1992) A method for desinfection of underground rhizome
tips of Alstroemeria and Heliconia. Acta Hortic 499–504. doi:
10.17660/ActaHortic.1992.325.69
Ramsay JL, Galitz DS, Lee CW (2003) Basal Medium and Sucrose Concentration
Influence Regeneration of Easter Lily in Ovary Culture. HortScience 38:404–
406.
Riedemannn P, Aldunate G (2001) Flora Nativa de valor Ornamental Chile Zona
Centro, Identificación y Propagación, 1st edn. Editorial Andrés Bello, Santiago,
Chile
Rout GR, Mohapatra A, Jain SM (2006) Tissue culture of ornamental pot plant: A
critical review on present scenario and future prospects. Biotechnol Adv
24:531–560. doi: 10.1016/j.biotechadv.2006.05.001
Sandler-Ziv D, Cohen A, Ion A, et al (1997) Improving Hippeastrum propagation

55
and bulbil yield by cutting and incubation techniques. Acta Hortic 355–360.
doi: 10.17660/ActaHortic.1997.430.55
Sassone AB, Arroyo-Leuenberger SC, Giussani LM (2014) Nueva circunscripción
de la tribu Leucocoryneae (Amaryllidaceae, Allioideae). Darwiniana 2:197–
206. doi: 10.14522/darwiniana.2014.22.584
Schiappacasse F, Peñailillo P, Yáñez P (2002) Propagación de Bulbosas Chilenas
Ornamentales. Editorial Universidad de Talca, Talca, Chile
Shahidul Haque M, Wada T, Hattori K (1997) High frequency shoot regeneration
and plantlet formation fromroot tip of garlic. Plant Cell Tissue Organ Cult
50:83–89. doi: 10.1023/A:1005973929862
Sharma A, Agrawal V (2012) Tissue culture aspects of ornamental plants. CIBTech
J Biotechnol 1:40–48.
Slabbert MM, de Bruyn MH, Ferreira DI, Pretorius J (1993) Regeneration of
bulblets from twin scales of Crinum macowanii in vitro. Plant Cell Tissue
Organ Cult 33:133–141. doi: 10.1007/BF01983226
Smith RH, Burrows J, Kurten K (1999) Challenges associated with
micropropagation of Zephyranthes and Hippesatrum sp. (Amaryllidaceae). Vitr
Cell Dev Biol - Plant 35:281–282. doi: 10.1007/s11627-999-0032-y
Sochacki D, Orlikowska T (2005) The obtaining of narcissus plants free from
potyviruses via adventitious shoot regeneration in vitro from infected bulbs. Sci
Hortic (Amsterdam) 103:219–225. doi: 10.1016/j.scienta.2004.04.018

56
Solgi M, Dastyari K, Hadavi E (2015) The Evaluation Effects of Some Vegetative
Propagation Methods and Plant Growth Regulators on Bulblet Production Rate
in Crown Imperial ( Fritillaria imperialis L .). J Hortic For ans Biotechnol 19:1–
6.
Stone OM (1973) The elimination of viruses from Narcissus tazetta cv. Grand
Soleil d’Or, and rapid multiplication of virus-free clones. Ann Appl Biol 73:45–
52. doi: 10.1111/j.1744-7348.1973.tb01308.x
Uranbey S (2011) In vitro bulblet regeneration from immature embryos of
endangered and endemic Muscari azureum. Arch Biol Sci 63:209–215. doi:
10.2298/ABS1101209U
van Leeuwen PJ, van der Weijden JA (1997) Propagation of specialty bulbs by
chipping. Acta Hortic 351–354. doi: 10.17660/ActaHortic.1997.430.54
Verdugo G (2013) Flower breeding of native plants: the Chilean Experience. Acta
Hortic 1000:401–405. doi: 10.17660/ActaHortic.2013.1000.55
Verdugo G, Teixeira JA (2006) From Wild to the Table : Leucocoryne and From
Wild to the Table : Leucocoryne and Chloraea. In: Floriculture, Ornamental
and Plant Biotechnology Volume IV. Global Science Books, pp 356–360
Vidal AK, Han D, Nakano M, Niimi Y (2012) Decreased time from seed to flowering
corm size in Zephyra elegansvia in vitro cultivation. Cienc e Investig Agrar
39:577–584. doi: 10.4067/S0718-16202012000300017
Walton EF, Wu RM, Sutherland P (2008) Bulb and inflorescence development in
Leucocoryne ixioides (Hook.) Lind. J Hortic Sci Biotechnol 83:574–580.

57
Zar J (2010) Biostatistical Analysis, Fifth. Prentice Hall Inc., New Jersey, USA
Zoellner O (2002) Descripción del género Leucocoryne (Alliaceae) su forma de
propagación y distribución. In: Leucocoryne un género nativo chileno y su uso
como planta de jardín, 1st edn. Oficina de transferencia tecnológica
Universidad Católica de Valparaíso, Viña del Mar, Chile, pp 23–27
Zoellner O (1972) El género Leucocoryne. An del Mus Hist Nat Valparaíso 5:9–83.

58
Table 1 Effect of basal media and BAP addition on the in vitro sprouting of three Leucocoryne genotypes.
Genotype Basal medium
BAP Sprouting Single shoot Multiple shoots (mg*L-1) (%) (%) (%)
L. purpurea
MS 0.0 86.7 b 86.7 b 0.0 b 1.0 100.0 a 71.4 b 28.6 a
LS 0.0 83.3 b 75.0 b 8.3 ab 1.0 100.0 a 92.9 ab 7.1 b
B5 0.0 91.7 ab 91.7 ab 0.0 b 1.0 100.0 a 100.0 a 0.0 b
L. ixioides
MS 0.0 92.3 ab 92.3 a 0.0 b 1.0 100.0 a 53.3 b 46.7 a
LS 0.0 85.7 b 85.7 ab 0.0 b 1.0 100.0 a 64.3 b 35.7 a
B5 0.0 85.7 b 85.7 ab 0.0 b 1.0 100.0 a 53.3 b 46.7 a
L. vittata
MS 0.0 100.0 a 93.3 ab 6.7 bc 1.0 86.7 b 73.3 b 13.3 b
LS 0.0 93.3 ab 93.3 ab 0.0 c 1.0 86.7 b 40.0 c 46.7 a
B5 0.0 100.0 a 100.0 a 0.0 c 1.0 100.0 a 80.0 b 20.0 b
Sprouting indicates the percentage of bulbs that developed sprouts. Single shoot indicates the percentage of bulbs that developed only one shoot. Multiple shoots indicates the number of bulbs that developed two or more shoots. Different letters in the same column indicate statistical differences for each genotype based on Scheffé´s procedure for multiple comparison of proportions (P≤0.05).
59
Table 2 Effect of different BAP concentrations added to the growing media on the in vitro sprouting of three Leucocoryne genotypes.
Genotype BAP Sprouting Single shoot Multiple shoots
(mg*L-1) (%) (%) (%)
L. purpurea
0.0 100.0 a 94.7 ab 5.3 bc 0.5 100.0 a 65.0 c 35.0 a 1.0 100.0 a 100.0 a 0.0 c 1.5 100.0 a 87.5 b 12.5 b 2.0 100.0 a 55.0 c 45.0 a
L. ixioides
0.0 100.0 a 89.5 a 10.5 b 0.5 100.0 a 47.4 bc 52.6 a 1.0 90.0 b 30.0 c 60.0 a 1.5 94.4 ab 50.0 bc 44.4 a 2.0 100.0 a 61.1 b 38.9 a
L. vittata
0.0 93.3 a 80.0 b 13.3 a 0.5 100.0 a 100.0 a 0.0 b 1.0 100.0 a 85.0 b 15.0 a 1.5 100.0 a 82.4 b 17.6 a 2.0 93.8 a 68.8 b 25.0 a
Sprouting indicates the percentage of bulbs that developed sprouts. Single shoot indicates the percentage of bulbs that developed only one shoot. Multiple shoots indicates the number of bulbs that developed two or more shoots. Different letters in the same column indicate statistical differences for each genotype based on Scheffé´s procedure for multiple comparison of proportions (P≤0.05).
60
Table 3 Effect of the in vitro cutting method, culture system and the addition of BAP to the growing media on sprouting, bulb multiplication and bulb production in Leucocoryne vittata.
Cutting method
Culture system
BAP Sprouting Bulb multiplication
Bulb production
(mg*L-1) (%) (%) (bulbs/explant)
Intact bulb (control)
Agar 0.0 40.0 b 20.0 cd 0.6 c 1.0 80.0 ab 20.0 cd 0.2 c
Liquid (Cotton pads)
0.0 40.0 b 0.0 d 0.0 c 1.0 100.0 a 20.0 cd 0.6 c
Scoring Agar 0.0 80.0 ab 60.0 bc 2.8 bc
1.0 80.0 ab 60.0 bc 3.2 bc Liquid
(Cotton pads) 0.0 60.0 b 40.0 bc 0.4 c 1.0 100.0 a 80.0 ab 2.6 bc
Sectioning Agar 0.0 100.0 a 100.0 a 13.0 a
1.0 80.0 ab 80.0 ab 9.2 ab Liquid
(Cotton pads) 0.0 80.0 ab 80.0 ab 4.4 bc 1.0 100.0 a 100.0 a 7.8 ab
Sprouting indicates the percentage of bulbs that developed sprouts. Bulb multiplication indicates the percentage of bulbs that developed lateral bulblets. Bulb production indicates the number of bulbs obtained per an initial bulb. Different letters in the same column indicate statistical differences based on Scheffé´s procedure for multiple comparison of proportions (P≤0.05) and analysis of variance (ANOVA) followed by Bonferroni's multiple comparison test (P≤0.05).

61
Fig. 1 Leucocoryne genotypes and explant types. (a) Leucocoryne purpurea; (b) Leucocoryne ixioides; (c) Leucocoryne vittata; (d) Leucocoryne sp. Pichicuy; (e) Initiation explant composed by the inner basal plate section plus the vegetative growing point; (f) Intact bulb; (g) Scoring and (h) Sectioning done to in vitro bulbs.

62
Fig. 2 Effect of basal media and the addition of BAP on three Leucocoryne genotypes. (a) Percentage of bulbs that developed lateral bulblets; (b) Number of bulbs obtained per an initial bulb; and (c) Average final fresh weight obtained per explant. Different letters indicate statistical differences for each genotype based, in the case of bulb multiplication, on Scheffé´s procedure for multiple comparison of proportions (P≤0.05) and for bulb production and bulb fresh weight, on analysis of variance (ANOVA) followed by Bonferroni's multiple comparison test (P≤0.05).

63
Fig. 3 Effect of BAP concentration on three Leucocoryne genotypes. (a) Percentage of bulbs that developed lateral bulblets; (b) Number of bulbs obtained per an initial bulb; and (c) Average final fresh weight obtained per explant. Different letters indicate statistical differences for each genotype based, in the case of bulb multiplication, on Scheffé´s procedure for multiple comparison of proportions (P≤0.05) and for bulb production and bulb fresh weight, on analysis of variance (ANOVA) followed by Bonferroni's multiple comparison test (P≤0.05).

64
Fig. 4 Effect of BAP concentration on in vitro Leucocoryne ixioides: (a) MS medium (control); (b) MS+0.5mg·L-1 BAP; (c) MS+1.0mg·L-1 BAP d) MS+1.5mg·L-1 BAP and (e) MS+2.0mg·L-1 BAP.

65
Fig. 5 Leucocoryne bulb production in response to different bulb-cutting methods. (a) Intact bulb (control); (b) Scoring; and (c) Sectioning.

66
Fig. 6 Effect of sectioning technique and BAP addition on four Leucocoryne genotypes. (a) Percentage of bulbs that developed lateral bulblets; (b) Number of bulbs obtained per an initial bulb; and (c) Average final fresh weight obtained per explant. Different letters indicate statistical differences for each genotype based, in the case of bulb multiplication, on Scheffé´s procedure for multiple comparison of proportions (P≤0.05) and for bulb production and bulb fresh weight, on analysis of variance (ANOVA) followed by Bonferroni's multiple comparison test (P≤0.05).

67
Fig. 7 Visual appearance of a Leucocoryne bulb and adventitious bulblet produced after in vitro sectioning. (a) Sectioned original explant; (b) Adventitious new bulblets.

68
CHAPTER 3
AN EFFICIENT IN VITRO PRODUCTION SYSTEM FOR
Leucocoryne spp. PLANTS INITIATED FROM SEEDS
Alejandro Altamira1, Eduardo Olate1, Marlene Gebauer1, Levi Mansur2, Carlos De
la Cuadra2 and Gloria Montenegro1*.
1Departamento de Ciencias Vegetales, Facultad de Agronomía e Ingeniería
Forestal, Pontificia Universidad Católica de Chile. Vicuña Mackenna 4860, Macul,
Santiago, Chile.
2Escuela de Agronomía, Pontificia Universidad Católica de Valparaíso, Avda. San
Francisco s/n, La Palma, casilla 4-D, Quillota, Chile
This chapter will be sent to Propagation of Ornamental Plants (POP) –International Journal (March 2017)

69
An efficient in vitro production system for Leucocoryne spp. plants initiated
from seeds
Alejandro Altamira1*, Eduardo Olate1, Marlene Gebauer1, Levi Mansur2, Carlos De la
Cuadra2 and Gloria Montenegro1.
1 Departamento de Ciencias Vegetales, Facultad de Agronomía e Ingeniería Forestal,
Pontificia Universidad Católica de Chile. Vicuña Mackenna 4860, Macul, Santiago, Chile.
2 Escuela de Agronomía, Pontificia Universidad Católica de Valparaíso, Avda. San
Francisco s/n, La Palma, casilla 4-D, Quillota, Chile
*Corresponding author:
Name: Alejandro Altamira
Address: Departamento de Ciencias Vegetales, Facultad de Agronomía e Ingeniería
Forestal, Pontificia Universidad Católica de Chile. Vicuña Mackenna 4860, Macul,
Santiago, Chile
Telephone: +56 9 90915256
E-Mail: [email protected]
Keywords: Amaryllidaceae, in vitro germination, geophyte, sprouting, bulb, optimal
temperature

70
Abstract
Leucocoryne spp. are geophyte plants that belongs to Amaryllidaceae, which have
a great potential in the ornamental plant industry due to its colorful flowers,
distinctive shape and long vase life. The plants are suitable to be used as cut
flowers, potted plants and also as landscape plants. This research focused on
optimizing an in vitro seed germination protocol and the subsequent bulb growth
and development of three Leucocoryne genotypes. Culture temperatures of 15 and
20 °C were used from seed initiation until bulblet formation. Seeds were in vitro
initiated on 12.5% MS medium and subsequently transferred to 50% MS media
optionally supplemented with BAP. Germination rates varied between 85-98% at
15 ºC and 48-83% at 20°C, depending on the genotype. In all the genotypes, the
highest bulb growth and development occurred at 15°C. Use of 50% MS
supplemented with BAP broke down bulb dormancy and stimulated lateral bulblets.
Final fresh weight of the bulbs was higher at 15°C in two of the genotypes and
varied between 0.04 and 0.10 g per bulb.
Abbreviations: BAP - N6-benzylaminopurine; MS – Murashige and Skoog medium
(1962)

71
Introduction.
The ornamental crop industry constantly demands for new cultivars or species with
new shapes and colors (Olate and Bridgen 2005). Leucocoryne is a geophyte
genus endemic to Chile which belongs to Amaryllidaceae and possess exceptional
qualities to be used as cut flower and potted plant, as well as in landscaping, due
to its long vase life and wide flower shape, design and color variety (Bridgen 2000;
Mansur 2002; Sassone et al. 2014; De la Cuadra et al. 2016). There are 15 to 20
species distributed throughout northern and central Chile. Its major diversity center
is located between the Coquimbo and Valparaiso regions (Zoellner 1972; Muñoz
and Moreira 2000; Mansur and Cisternas 2005; Olate and Schiappacasse 2013;
Jara-Arancio et al. 2014). Plants belonging to this genus have a small tunicate bulb
and a single floral scape 30-40 cm in height, which ends in an umbel carrying 3-12
flowers depending on the species. One of the main characteristics are its flower
pattern and color, varying between white, blue and purple (Mansur 2002;
Schiappacasse et al. 2002; Zoellner 2002; Olate and Bridgen 2005; Hoffmann et
al. 2015).
The ornamental value of Leucocoryne has brought special interest in the
knowledge and domestication of the species, with several studies and commercial
efforts done in Japan, The Netherlands, Israel, New Zealand and Chile (Kim et al.
1998b; Bridgen 2000; Lancaster et al. 2000; Schiappacasse et al. 2002; Catley
2003; Walton et al. 2008; Olate and Schiappacasse 2013). This particular interest
and the anthropic and climate threats that natural populations face, have led to the

72
development of a research and breeding program in Chile for the conservation and
commercialization of this genus. This particular program has already registered
three new cultivars in addition to the ones already developed in The Netherlands
(De la Cuadra et al. 2016).
Few studies on the propagation of Leucocoryne have been published to date.
It is therefore necessary to make progress in these techniques in order to help
commercial and conservation advances. has a slow life cycle that takes up to
three or four years from seed to floral bulb (Mansur 2002). It has been reported a
high genetic variation among species and ecotypes, which implies a great potential
to do breeding but, at the same time, it is a sign of the difficulties to maintain
interesting characteristics using sexual propagation (Mansur 2002; Mansur et al.
2004). Leucocoryne, as other geophyte genus, has the natural capacity of
vegetative propagation through the development of one or two lateral bulblets at
the bulb base, or by the production of “droppers”, a type of bulblet that come out as
an additional stem from the center of the bulb but instead grows deeper into the
ground, also from the second or third year of growth (Riedemannn and Aldunate
2001; Mansur 2002; Schiappacasse et al. 2002). Bulb-cutting methods have been
studied in order to increase propagation efficiency (Schiappacasse et al. 2002).
Some of these studies include specific in vitro culture research on Leucocoryne
species (Olate and Bridgen 2005; Escobar et al. 2008).
Sexual propagation of Leucocoryne spp. can be an interesting tool of propagation
for breeding purposes or when genetic stability of the plants is needed.
Nevertheless, seed propagation is still the easiest and inexpensive way to

73
propagate Leucocoryne plants in big quantities, either for conservation purposes or
during the first stages of plant breeding. The in vitro propagation from seeds adds
the extra advantage of a more reliable system under a controlled environment, and
independent from seasonal environmental conditions. It is important to fully
understand the seed propagation process and all the factors that control the
different phases of a particular methodology. In that sense, germination and
growing temperature is one of the main factors to determine. It has been reported
that different Leucocoryne species show different optimal temperatures for seed
germination measured either in natural growing habitats (Jara et al. 2006) or under
artificial conditions (De la Cuadra et al. 2016).
In this context, the objectives of this study are to develop and to optimize the
protocol for the in vitro seed germination of three Leucocoryne genotypes and to
determine the effect of the temperature on the in vitro plant and bulb production.

74
Materials and methods.
Plant material.
Seed belonging to three Leucocoryne species were used as explants: Leucocoryne
purpurea (Fig. 1a), Leucocoryne vittata (Fig. 1b) and ecotype Leucocoryne sp.
Pichicuy (Fig. 1c). Seeds were collected from plants under greenhouse conditions
in the germplasm bank of the Leucocoryne Breeding Program at the Pontificia
Universidad Católica de Valparaíso Experimental Station, located in Quillota (lat.
32°53´ S; long. 71°12’ W). Seeds were stored up to six months in paper bags at
20±2°C in dark conditions, until they were used.
The in vitro studies were conducted in the laboratories of the Plant Science
Department of the Pontificia Universidad Católica de Chile, in Santiago, Chile
(33°29’ S; long. 70° 36’W).
Disinfection and in vitro establishment.
To disinfect the seeds were submerged in a 10 g·L-1 hydric solution of sodium
hypochlorite for 15 min under a laminar flow chamber and rinsed 3 times with
sterile distilled water. Seeds were then imbibed in sterile distilled water for 96 h at
8°C in dark conditions (Ham 2002; De la Cuadra and Mansur 2004; Jara et al.
2006). After imbibition, seeds were sown in 200 mL glass culture vessels
containing 30 mL of 12.5% MS medium supplemented with 30 g·L-1 sucrose, 6 g·L-
1 agar and pH were adjusted to 5.7 (Murashige and Skoog 1962; Ham 2002).

75
In vitro seed germination.
A total of 300 seeds per Leucocoryne genotype were sown in vitro conditions.
Seeds were cultured under two temperature treatments: 15±1°C and 20±1°C, in a
growth chamber with a 16h photoperiod. Treatments consisted in six replicates of
five vessels each, containing five seeds/vessel. After 13 weeks of in vitro culture,
germination rate, bulbing rate, dormancy rate and bulb fresh weight were
evaluated.
Bulb development and multiplication.
After 13 weeks of in vitro culture, bulbs from seeds were transferred onto 25% MS
medium, and after 10 additional weeks they were transferred onto 50% MS
supplemented with 1 mg·L-1 BAP and 30 g·L-1 sucrose, 6 g·L-1 agar and pH
adjusted to 5.7. After 24 weeks, bulbing rate, bulb sprouting, bulblet production,
rooting rate and bulb fresh weight were evaluated.
Statistical analysis.
Bulb fresh weight data were analyzed by analysis of variance (ANOVA) and
Bonferroni's multiple comparison test (P≤0.05) using GraphPad Prism 5 software
(GraphPad Software Inc., San Diego, California, USA). In the case of germination,
bulbing, dormancy, sprouting, bulblet production and rooting data were analyzed
using Sheffé's multiple comparison for proportions test (P≤0.05) (Zar 2010).

76
Results and discussion.
Disinfection and in vitro establishment protocols for Leucocoryne seeds were
highly effective, showing less than 1% contamination and high germination rates in
all the genotypes used. This indicates that disinfection method used did not
damage seeds.
In terms of germination rate after 13 weeks of in vitro culture, the best results were
obtained at 15°C in the three genotypes, with values of 85.3% in L. purpurea,
96.7% in L. vittata and 98.7% in L. sp. Pichicuy, significantly higher than
germination rates at 20°C (Table 1). However, we observed that seed germination
occurred earlier at 20°C than at 15°C (data not showed). Ham (2002) reported 89%
germination in L. coquimbensis using the same culture medium but using 12°C and
darkness conditions, also reporting that 12.5% MS worked as the best MS
concentration for seed germination compared to higher concentrations (25%,
37.5%, 50% and 100%). Our results are also in accordance with reports of De la
Cuadra et al. (2016) that used the same three seed genotypes, but moist
chromatographic paper and darkness as germination conditions. They found that
germination rates were significantly higher at 10ºC and 15°C than at 20°C,
reporting also no seed germination at 25°C. As mentioned by Jara et al. (2006),
these optimal low temperatures for Leucocoryne seed germination and the
inhibitory effect caused by high temperatures under laboratory conditions coincide
with the natural desert and Mediterranean environments in which these species are
found.

77
We also evaluated the occurrence of bulbs (bulbing) from the seedlings produced.
The highest bulbing percentage occurred in seedlings cultured at 15°C (Table 1;
Fig. 2), thus repeating the positive effect of the lower temperature treatment for
seed germination. Bulbs obtained in both treatments were clearly different in size;
therefore bulb fresh weight was also evaluated. In the same way, better results
were obtained at the lowest culture temperature with higher bulb fresh weight at
15°C than at 20°C (Fig. 2; Fig. 3).
After seed germination, during bulb and plant development, we observed the
occurrence of premature senescence of the seedlings. Apparent dormancy started
to show up after 13 weeks of in vitro culture, thus the effect of temperature culture
on dormancy of the bulbs was also evaluated. Bulbs were considered as dormant
when more than 2/3 of the shoot showed senescence before transferring them to
fresh media. In average, L. purpurea showed the lowest dormancy rates between
28% and 53%, while L. vittata and L. sp. Pichicuy showed over 73% of dormant
bulbs (Table 1). Culture temperature affected dormancy rates, occurring lower
dormancy rates at 15°C in all genotypes. De la Cuadra and Mansur (2004)
reported seedling senescence 11 weeks after germination, and completely
senescent plants after 13 weeks of growth.
After transferring the bulbs onto 25% MS medium, we were able confirm that most
of the bulbs were dormant. After 10 weeks, the bulbs achieved a sprouting rate of
only 9.1% (Fig. 4a). During this period, bulb fresh weight did not significantly
change, showing 0.03 g/bulb in L. purpurea in both culture temperatures

78
treatments, and 0.04 and 0.02 g/bulb in L. vittata and L. sp. Pichicuy at 15ºC and
20°C respectively.
Transferring the bulbs onto 50% MS medium supplemented with 1.0 mg*L-1 BAP
triggered dormancy break. The interruption of the bulb dormancy could be verified
with the appearance of vegetative shoots from the explants, regardless of the
culture temperature (Table 2). This would confirm that BAP produces an effect on
bulb dormancy break in Leucocoryne species under in vitro conditions. BAP
addition to in vitro culture media has been used before for stimulating cell division,
shoot formation and growth from axillary buds in other geophyte species (Jha
2005; Maślanka and Bach 2014). We could also observe the appearance of
adventitious bulblets between 4.9% and 12.6% of the explants, independently of
the culture temperature (Table 2; Fig. 4b). Therefore, BAP could also promote and
accelerate adventitious bulblet formation in Leucocoryne species under in vitro
conditions. Although the adventitious bulblet formation observed in this study was
low, it is remarkable that naturally bulb formation in Leucocoryne species does not
occur until the second or third year of growth (Riedemannn and Aldunate 2001;
Mansur 2002). Therefore, the use of in vitro propagation and BAP could delay or
avoid bulbs entering in the natural early dormancy and shorten the time from seed
to adventitious bulblet appearance, thus allowing a faster and more efficient
propagation system. In terms of bulb rooting, statistical differences were observed
between temperature treatments in both L. purpurea and L. sp. Pichicuy, with
higher bulb rooting at 15°C (Table 2).

79
We would also like to highlight the fact that gradual increase of MS concentration
to the growing media (12.5% to 25% and then to 50%) allowed seedlings to
continue growing by gradually adapting to increasing osmotic levels produced by
higher salt contents of the culture media. Previously, Vidal et al. (2012) reported
that Zephyra elegans plants, another geophyte species endemic to Chile with
ornamental potential, were unable to grow in vitro when seedlings germinated on
water agar were directly subcultured onto 100% MS medium.
Final fresh weight of the bulbs varied between 0.04 and 0.1 g depending on both
genotype and culture temperature, but without statistical differences (Fig. 5). It has
been reported that Leucocoryne bulbs must have a fresh weight over 0.3 g in order
to produce quality cut flowers (Kim et al. 1998a) Nevertheless, it has been also
reported that bulbs 0.1-0.2 g in weight are able to increase their weight up to 1 g
after one growing season under greenhouse conditions, producing high quality cut
flowers in the subsequent season (Kim et al. 1998a).
Future works in Leucocoryne sexual propagation under in vitro conditions should
consider increasing the macro and micronutrient concentration of the media
gradually until reaching 100% MS. In that way bulbs will be able to increase and
accelerate their weight constantly. By maintaining this continuous in vitro bulb
growth, and avoiding dormancy, it should be possible to reduce the time needed to
obtain flowering size bulbs.
In conclusion, in this work we determined that 15ºC as culture temperature
produced a very efficient Leucocoryne plant production using in vitro seed
germination. In all the genotypes tested high germination and bulbing rates were

80
achieved at 15°C. Culture temperatures did not produce statistical differences on
bulb fresh weight, but temperatures lower than 15ºC could produce an increase on
this parameter. With the addition of BAP it was possible to break the dormancy of
the bulbs, thus favoring sprouting and the multiplication of the explants. The
protocol developed in this work leads to high in vitro plant production efficiency and
may reduce the time needed to produce flowering size bulbs.

81
References.
Bridgen M (2000) Flowering geophytes of Chile have ornamental potential. In:
Seeds: Trade, Production and Technology. pp 218–230
Catley JL (2003) Temperature and Irradiance Effects on Vegetative Growth of Two
Species of Leucocoryne. J Am Soc Hortic Sci 128:815–820.
De la Cuadra C, Mansur L (2004) Descripción de la primera etapa del ciclo de vida
de tres genotipos de Leucocoryne sp.: semilla a bulbo. Agric Técnica. doi:
10.4067/S0365-28072004000200009
De la Cuadra C, Vidal A, Lefimil S, Mansur L (2016) Temperature Effect on Seed
Germination in the Genus Leucocoryne (Amaryllidaceae ). HortScience
51:412–415.
Escobar L, Jordan M, Olate E, et al (2008) Direct and indirect in vitro
organogenesis of leucocoryne pupurea (Alliaceae) a chilean ornamental
geophyte. Propag Ornam Plants 8:59–64.
Ham F (2002) Evaluación de la germinación in vitro de Leucocoryne coquimbensis
F. Phil., geófita enémica con potencial ornamental. Pontificia Universidad
Católica de Chile
Hoffmann A, Watson J, Flores AR (2015) Flora Silvestre de Chile, Cuando el
Desierto Florece Vol.I. Monocotiledóneas, 1st edn. Ediciones Fundación
Claudio Gay, Zapallar, Chile
Jara-Arancio P, Arroyo MTK, Guerrero PC, et al (2014) Phylogenetic perspectives

82
on biome shifts in Leucocoryne (Alliaceae) in relation to climatic niche
evolution in western South America. J Biogeogr 41:328–338. doi:
10.1111/jbi.12186
Jara P, Arancio G, Moreno R, Carmona M (2006) Factores abióticos que
influencian la germinación de seis especies herbáceas de la zona árida de
Chile. Rev Chil Hist Nat 79:309–319.
Jha TB (2005) Plant Tissue Culture: Basic and Applied. Universities Press
Kim HH, Ohkawa K, Nitta E (1998a) EFFECTS OF BULB WEIGHT ON THE
GROWTH AND FLOWERING OF LEUCOCORYNE COQUIMBENSIS F.
PHILL. Acta Hortic 341–346. doi: 10.17660/ActaHortic.1998.454.40
Kim HH, Ohkawa K, Nitta E (1998b) Fall flowering of Leucocoryne coquimbensis F.
Phil. after long-term bulb storage treatments. HortScience 33:18–20.
Lancaster JE, Shaw ML, Walton EF (2000) S-Alk(en)yl-l-cysteine sulfoxides,
alliinase and aroma in Leucocoryne. Phytochemistry 55:127–130. doi:
10.1016/S0031-9422(00)00245-4
Mansur L (2002) Ciclo de vida, autoincompatibilidad y plasticidad genética en
diseño y color de flores. In: Leucocoryne un género nativo chileno y su uso
como planta de jardín, 1st edn. Oficina de transferencia tecnológica
Universidad Católica de Valparaíso, Viña del Mar, Chile, pp 9–15
Mansur L, Cisternas M (2005) Leucocoryne talinensis ( Alliaceae ), a New Species
from Chile. Novon 15:324–326.

83
Mansur L, Gonzalez M, Rojas I, Salas P (2004) Self-incompatibility in the Chilean
Endemic Genus. J Amer Soc Hort Sci 129:836–838.
Maślanka M, Bach A (2014) Induction of bulb organogenesis in in vitro cultures of
tarda tulip (Tulipa tarda Stapf.) from seed-derived explants. Vitr Cell Dev Biol -
Plant 50:712–721. doi: 10.1007/s11627-014-9641-1
Muñoz M, Moreira A (2000) Los Géneros Endémicos de Monocotiledóneas de
Chile Continental.
http://www.chlorischile.cl/monocotiledoneas/leucocoryne.htm. Accessed 8 Nov
2016
Murashige T, Skoog F (1962) A Revised Medium for Rapid Growth and Bio Assays
with Tobacco Tissue Cultures. Physiol Plant 15:473–497. doi: 10.1111/j.1399-
3054.1962.tb08052.x
Olate E, Bridgen M (2005) Techniques for the In Vitro Propagation of Rhodophiala
and Leucocoryne spp. Acta Hortic 673:335–342. doi:
10.17660/ActaHortic.2005.673.42
Olate E, Schiappacasse F (2013) Geophyte Research and Production in Chile. In:
Ornamental Geophytes. CRC Press, pp 449–470
Riedemannn P, Aldunate G (2001) Flora Nativa de valor Ornamental Chile Zona
Centro, Identificación y Propagación, 1st edn. Editorial Andrés Bello, Santiago,
Chile
Sassone AB, Arroyo-Leuenberger SC, Giussani LM (2014) Nueva circunscripción
de la tribu Leucocoryneae (Amaryllidaceae, Allioideae). Darwiniana 2:197–

84
206. doi: 10.14522/darwiniana.2014.22.584
Schiappacasse F, Peñailillo P, Yáñez P (2002) Propagación de Bulbosas Chilenas
Ornamentales. Editorial Universidad de Talca, Talca, Chile
Vidal AK, Han D, Nakano M, Niimi Y (2012) Decreased time from seed to flowering
corm size in Zephyra elegansvia in vitro cultivation. Cienc e Investig Agrar
39:577–584. doi: 10.4067/S0718-16202012000300017
Walton EF, Wu RM, Sutherland P (2008) Bulb and inflorescence development in
Leucocoryne ixioides (Hook.) Lind. J Hortic Sci Biotechnol 83:574–580.
Zar J (2010) Biostatistical Analysis, Fifth. Prentice Hall Inc., New Jersey, USA
Zoellner O (1972) El género Leucocoryne. An del Mus Hist Nat Valparaíso 5:9–83.
Zoellner O (2002) Descripción del género Leucocoryne (Alliaceae) su forma de
propagación y distribución. In: Leucocoryne un género nativo chileno y su uso
como planta de jardín, 1st edn. Oficina de transferencia tecnológica
Universidad Católica de Valparaíso, Viña del Mar, Chile, pp 23–27
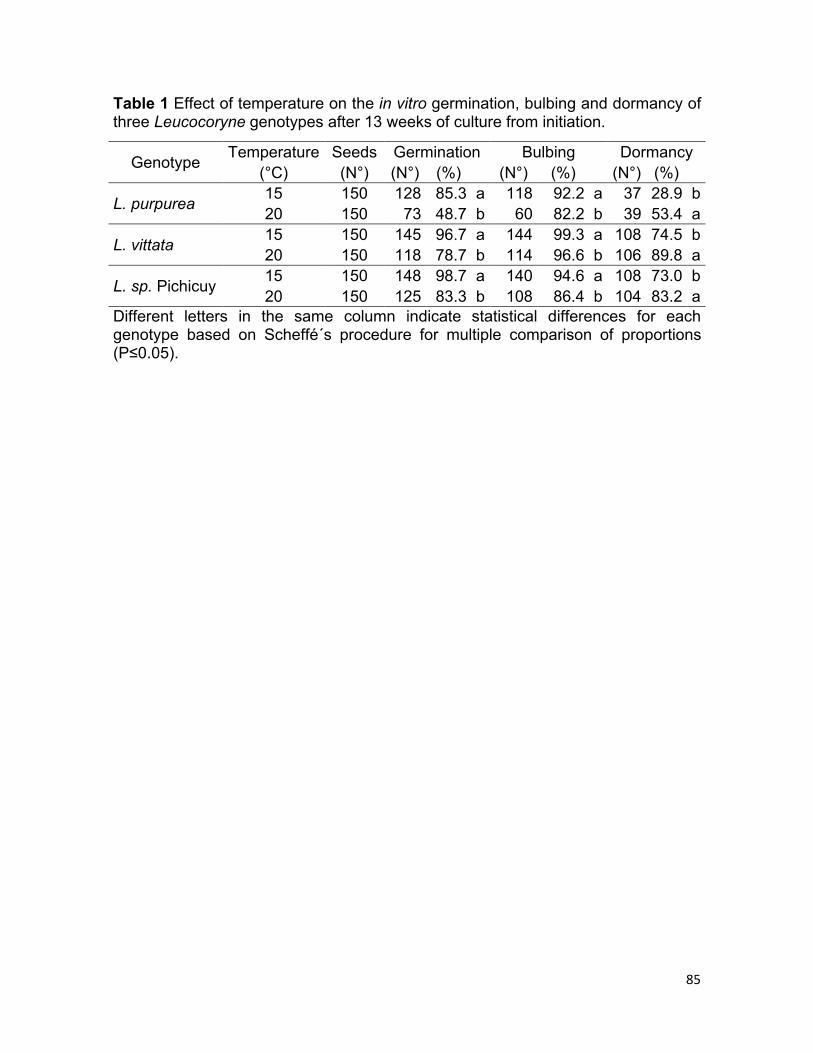
85
Table 1 Effect of temperature on the in vitro germination, bulbing and dormancy of three Leucocoryne genotypes after 13 weeks of culture from initiation.
Genotype Temperature Seeds Germination Bulbing Dormancy (°C) (N°) (N°) (%) (N°) (%) (N°) (%)
L. purpurea 15 150 128 85.3 a 118 92.2 a 37 28.9 b 20 150 73 48.7 b 60 82.2 b 39 53.4 a
L. vittata 15 150 145 96.7 a 144 99.3 a 108 74.5 b 20 150 118 78.7 b 114 96.6 b 106 89.8 a
L. sp. Pichicuy 15 150 148 98.7 a 140 94.6 a 108 73.0 b 20 150 125 83.3 b 108 86.4 b 104 83.2 a
Different letters in the same column indicate statistical differences for each genotype based on Scheffé´s procedure for multiple comparison of proportions (P≤0.05).

86
Table 2 Effect of temperature on bulb occurrence bulbing, bulb sprouting, bulblet production and rooting after 24 weeks in MS 50% + 1.0 mg*L-1 BAP of three Leucocoryne genotypes.
Genotype Temperature Bulbing Bulb sprouting
Bulblet production
Rooting
(°C) (N°) (N°) (%) (N°) (%) (N°) (%)
L. purpurea 15 107 71 66.4 a 7 6.5 a 33 30.8 a 20 53 36 67.9 a 4 7.5 a 11 20.8 b
L. vittata 15 143 94 65.7 a 7 4.9 a 44 30.8 a 20 107 76 71.0 a 7 6.5 a 28 26.2 a
L. sp. Pichicuy 15 135 97 71.9 a 17 12.6 a 66 48.9 a 20 85 63 74.1 a 7 8.2 a 28 32.9 b
Different letters in the same column indicate statistical differences for each genotype based on Scheffé´s procedure for multiple comparison of proportions (P≤0.05).

87
Fig. 1 Leucocoryne genotypes. (a) Leucocoryne purpurea; (b) Leucocoryne vittata; and (c) Leucocoryne sp. Pichicuy.

Fig. 2 genotypes
Fig. 2 Effect of temperature ongenotypes after 13 weeks
Effect of temperature on13 weeks of culture
Effect of temperature on the in vitroof culture from
in vitro bulb production of from seed initiation
bulb production of initiation.
bulb production of three three Leucocoryne
88
Leucocoryne
89
Fig. 3 Effect of temperature on the bulb fresh weight of three Leucocoryne genotypes after 13 weeks from seed in vitro initiation. Different letters indicate statistical differences for each genotype according to analysis of variance (ANOVA) followed by Bonferroni's multiple comparison test (P≤0.05).

Fig. 4Dormant bulbsMS 50% + 1.0 mg*L
Fig. 4 Different vDormant bulbs onMS 50% + 1.0 mg*L
Different visual appearance of on MS 25%
MS 50% + 1.0 mg*L-1 BAP.
isual appearance of MS 25%; (b) Active bulb
BAP.
isual appearance of Leucocoryne(b) Active bulbs with shoot
Leucocoryne bulbs on with shoot and bulblet
bulbs on in vitro conditions: (a) and bulblet producti
conditions: (a) production on
90
conditions: (a) on on

91
Fig. 5 Effect of culture temperature on bulb fresh weight of three Leucocoryne genotypes after 24 weeks of in vitro culture on MS 50% + 1.0 mg*L-1 BAP. Different letters indicate statistical differences for each genotype according to analysis of variance (ANOVA) followed by Bonferroni's multiple comparison test (P≤0.05).

92
CHAPTER 4
CONCLUSIONS

93
In this study we have developed an efficient protocol for the in vitro propagation of
the Leucocoryne genus using different culture techniques.
Different culture media, supplemental addition of BAP, different culture systems
and type of explants were evaluated in order to develop mass clonal bulb
propagation. We are also reporting an effective disinfection method as part of the
in vitro establishment protocol, by which we have surpassed one of the main
barriers for the Leucocoryne in vitro culture.
By studying the effect of culture media and BAP addition, higher bulb production
rates than the naturally occurred were achieved. A higher bulb fresh weight was
also achieved, which allows the use of such larger bulbs in further propagation
techniques.
We have achieved the highest rate of propagation of Leucocoryne bulbs reported
to date, considering both in vitro and ex vitro conditions, when bulbs were
subjected to bulb-cutting methods.
The effects of the culture temperature BAP addition on the in vitro seed
germination and bulb development, was also established. Results regarding both
factors have great potential of use in obtaining active and large plant populations
from sexual reproduction.
Protocols developed in this study will help to overcome the need of a reliable mass
propagation method, which is one of the most important pending challenges for the
Leucocoryne genus. An efficient mass propagation method is an essential and
valuable advance and an important contribution to the domestication and

94
sustainable use of this plant genetic resource. Therefore, the use of these
protocols will assure a high production of plants to be used for conservation,
breeding or commercial purposes. This will greatly increase the value of
Leucocoryne genus in the ornamental crop industry.