Embed Size (px)
Citation preview

Stephan C. Collins,1,2 Hyun Woong Do,1 Benoit Hastoy,1 Alison Hugill,3 Julie Adam,1
Margarita V. Chibalina,1 Juris Galvanovskis,1,4 Mahdieh Godazgar,1 Sheena Lee,4
Michelle Goldsworthy,3 Albert Salehi,5,6 Andrei I. Tarasov,1,7 Anders H. Rosengren,5
Roger Cox,3 and Patrik Rorsman1,6,7
Increased Expression of the DiabetesGene SOX4 Reduces Insulin Secretionby Impaired Fusion Pore ExpansionDiabetes 2016;65:1952–1961 | DOI: 10.2337/db15-1489
The transcription factor Sox4 has been proposed to underliethe increased type 2 diabetes risk linked to an intronic sin-gle nucleotide polymorphism in CDKAL1. In a mouse modelexpressing a mutant form of Sox4, glucose-induced insulinsecretion is reduced by 40% despite normal intracellularCa2+ signaling and depolarization-evoked exocytosis. Thisparadox is explained by a fourfold increase in kiss-and-runexocytosis (as determined by single-granule exocytosismeasurements) in which the fusion pore connectingthe granule lumen to the exterior expands to a diameterof only 2 nm, which does not allow the exit of insulin.Microarray analysis indicated that this correlated withan increased expression of the exocytosis-regulatingprotein Stxbp6. In a large collection of human isletpreparations (n = 63), STXBP6 expression and glucose-induced insulin secretion correlated positively and nega-tively with SOX4 expression, respectively. Overexpressionof SOX4 in the human insulin–secreting cell EndoC-bH2interfered with granule emptying and inhibited hormonerelease, the latter effect reversed by silencing STXBP6.These data suggest that increased SOX4 expression in-hibits insulin secretion and increased diabetes risk bythe upregulation of STXBP6 and an increase in kiss-and-run exocytosis at the expense of full fusion. Wepropose that pharmacological interventions promot-ing fusion pore expansion may be effective in diabetestherapy.
Reduced glucose-induced insulin secretion (GIIS) is ahallmark of type 2 diabetes (1). This disease results froma complex crosstalk between lifestyle factors (e.g., body weight,age) and genetics (1). Insulin is released by Ca2+-dependentexocytosis of insulin-containing secretory granules. Exo-cytosis involves the fusion of the granular membrane withthe plasma membrane, a process initiated by the establish-ment of a narrow fusion pore that connects the granulelumen with the extracellular space (2). Effective release ofinsulin requires the rapid expansion of the fusion poreso that the secretory granules integrate with the plasmamembrane.
Genome-wide association studies have led to thediscovery of .100 loci associated with increased type 2diabetes risk (3,4), many of which are believed to actthrough reduced b-cell mass or interference with insu-lin secretion (5). For most of these loci, gene annota-tion is only tentative and based on the proximity of thesingle nucleotide polymorphism (SNP) to a certain gene(4). An intronic SNP in the CDKAL1 gene (rs7756992)is associated with a 50% increase in the risk of type 2diabetes (6–9). However, data indicate that CDKAL1 isnot the causative gene (9) and that its effect is insteadmediated by altered expression of a nearby gene encod-ing the transcription factor SOX4 (10).
1Oxford Centre for Diabetes, Endocrinology & Metabolism, Radcliffe Departmentof Medicine, Oxford, U.K.2Université de Bourgogne Franche-Comté, Burgundy, France3Mammalian Genetics Unit, MRC Harwell, Oxfordshire, U.K.4Henry Wellcome Centre for Gene Function, Department of Physiology, Anatomy,and Genetics, University of Oxford, Oxford, U.K.5Lund University Diabetes Centre, Department of Clinical Sciences, Skåne Uni-versity Hospital Malmö, Lund University, Malmö, Sweden6Department of Neuroscience and Physiology, University of Göteborg, Göteborg,Sweden7Oxford National Institute of Health Research, Biomedical Research Centre,Churchill Hospital, Oxford, U.K.
Corresponding author: Stephan C. Collins, [email protected].
Received 29 October 2015 and accepted 8 March 2016.
This article contains Supplementary Data online at http://diabetes.diabetesjournals.org/lookup/suppl/doi:10.2337/db15-1489/-/DC1.
S.C.C., H.W.D., B.H., and A.H. share first authorship.
S.C.C. is currently affiliated with Institut de Génétique et de Biologie Moléculaireet Cellulaire, Illkirch, France.
© 2016 by the American Diabetes Association. Readers may use this article aslong as the work is properly cited, the use is educational and not for profit, andthe work is not altered.
See accompanying article, p. 1791.
1952 Diabetes Volume 65, July 2016
ISLETSTUDIES

We show that increased SOX4 expression is associatedwith reduced GIIS and elevated plasma glucose and thatincreased SOX4 expression impedes the delivery of insulininto the extracellular space through increased expressionof the exocytosis-regulating protein STXBP6 (amisyn).Overexpression of amisyn promotes the stabilization ofthe fusion pore and locks it in a partially expanded state(2–3 nm) (11). This prevents the exit of insulin from thegranule lumen into the extracellular space and thus im-pairs insulin secretion.
RESEARCH DESIGN AND METHODS
Animal HusbandryAnimal work was approved by the local and nationalauthorities. Mice used were described previously (12) butwere heterozygous for the Sox4mutation and were Insr+/+.Mice were killed by cervical dislocation, and islets wereisolated by liberase digestion and handpicking as previ-ously described (13). Static and dynamic measurements ofinsulin secretion were performed as described previously(12). Insulin was measured by mouse insulin ELISA kit(Millipore, Hertfordshire, U.K.).
Mouse Microarray StudiesIslets were isolated from 22-week-old male wild-type andmutant mice (four animals per genotype), and RNA wasextracted with an RNeasy Mini Kit (QIAGEN) and validatedwith an Agilent 2100 Bioanalyzer (Agilent Technologies).Labeled and fragmented complementary RNA (cRNA) washybridized to the Affymetrix 430 2.0 whole-mouse genomemicroarray and processed on an GeneChip Fluidics Station450 and Scanner 3000 (Affymetrix).
Cell Transfections, Small Interfering RNA, andSecretion AssaysThe constructs coding for green fluorescent protein–taggedmouse and human Sox4 and Stxbp6 were purchased fromOriGene Technologies (Rockville, MD). The Y123C mu-tation was introduced into mouse Sox4 by using theQuikChange protocol (Agilent Genomics). INS-1 832/13cells were transfected with Lipofectamine RNAiMAX re-agent (Life Technologies, Paisley, U.K.). The following smallinterfering RNAs (siRNAs) were used: ON-TARGETplussiRNA SMARTpool for Stxbp6 gene and ON-TARGETplusNon-targeting Control siRNAs (Thermo Scientific, HemelHempstead, U.K.). After 24 h, the cells were cotransfectedwith human growth hormone (hGH) and either Discosomaspecies red fluorescent protein (DsRed)–or green fluores-cent protein–tagged mouse Sox4 or Stxbp6 by using Gene-Juice Transfection Reagent (Merck, Nottingham, U.K.).Supernatants and cell pellets were collected, and theamount of growth hormone was measured by using anhGH ELISA kit (Roche Diagnostics, West Sussex, U.K.).EndoC-bH2 cells (14) were transfected with human-specificON-TARGETplus siRNA SMARTpools for STXBP6 andSOX4 used in siRNA knockdown experiments. siRNA andtransfections were performed as described for INS-1 832/13
cells. Quantitative analysis of gene expression was per-formed using QuantiFast SYBR Green PCR kit and gene-specific QuantiTect Primer Assays (QIAGEN). Expressionwas calculated using DCt method, with GAPDH as a ref-erence gene.
Intracellular Calcium MeasurementsIntracellular calcium concentration ([Ca2+]i) was assessedin freshly isolated intact islets by using a dual-wavelengthfluorescence microscopy system (Photon Technology Inter-national, Monmouth Junction, NJ) fitted on an invertedZeiss microscope to allow ratiometric measurements withthe probe Fura-2, AM (Invitrogen, Paisley, U.K.) as describedpreviously (15).
Whole-Cell Measurements of Ca2+ Currents andExocytosisFor patch-clamp measurements, islets were dissociated intosingle b-cells. In whole-cell measurements, insulin-secretingb-cells were identified on the basis of their larger size (.5.5picofarads) and complete inactivation of the Na+ currentat 270 mV (16). Exocytosis and whole-cell Ca2+ currentswere recorded with an Elektronik patch clamp 10 amplifierand Pulse software (HEKA Electronik, Lambrecht [Pfalz],Germany) as described previously (15). Single exocytoticevents and fusion pore expansion were detected in thecell-attached configuration. The standard extracellularsolution described in Zhang et al. (16) supplementedwith 20 mmol/L glucose was used as the pipette fillingmedium. Patchmaster software (HEKA Electronik) and theElektronik patch clamp 10 amplifier together implement aninternal calibration and automatically correct for phaseshifts and frequency-dependent attenuation when a sinu-soidal voltage command of 25 kHz is generated (17). Thescaled apparent capacitance (Im/v) and conductance (Re)are then calculated online by the software.
P2X2 Receptor Expression and Current AnalysisP2X2 currents were recorded in the whole-cell configura-tion in identified b-cells as described previously (19,20).Experiments in EndoC-bH2 cells were performed aftertransfecting cells with both STXBP6 as described previ-ously and P2X2 receptors (as described in Karanauskaiteet al. [20]). To evoke exocytosis, the cells were infusedwith an intracellular medium containing 0–9 mmol/LCaCl2/10 mmol/L EGTA mixture (calculated free [Ca2+]i0 or 2 mmol/L), 3 mmol/L Mg-ATP, and 0.1 mmol/LcAMP. The current spikes reflecting vesicular release ofATP were analyzed using Mini Analysis Program version6.0.3 (Synaptosoft, Decatur, GA) to determine the charge(integrated current [Q]), rise times (t10–90%), and half-widths (HWs) of the individual events (20).
Electron MicroscopyIsolated islets were washed in PBS and fixed in cold 2.5%glutaraldehyde in PBS for 1 h at 4°C and washed twice inPBS. Islets were incubated for 1 h in 1% osmium tetroxideand washed twice in PBS. Islets were dehydrated throughan ethanol series for 5–10 min each and then embedded
diabetes.diabetesjournals.org Collins and Associates 1953
in Agar 100 resin mix (Agar Scientific) pending sectioningand electron microscopy.
Human Islet Insulin Secretion and Microarray StudiesIslets from cadaveric donors of European ancestry wereprovided by the Nordic Islet Transplantation Program(http://www.nordicislets.org). All procedures were approvedby the local ethics committee at Lund University. The isletswere cultured in supplemented CMRL 1066 (MP Biomedi-cals) for 1–9 days before RNA preparation. Total RNA wasisolated with the AllPrep DNA/RNA Mini Kit (QIAGEN).RNA quality and concentration were measured with anAgilent 2100 bioanalyzer and ND-1000 spectrophotom-eter (NanoDrop Technologies). Total RNA was convertedinto biotin-targeted cRNA, and the biotin-labeled cRNAwas fragmented into strands with 35–200 nucleotides.This was hybridized onto a GeneChip Human Gene 1.0 STArray overnight in a GeneChip Hybridization Oven 640(Affymetrix). The arrays were washed and stained in aGeneChip Fluidics Station 450. Scanning was carried outwith a GeneChip Scanner 3000, and image analysis wasdone with GeneChip Operating Software. Data normaliza-tion was performed by using Robust Multiarray Analysis.Insulin secretion in human islets was measured as de-scribed previously (21). Microarray expression data havebeen deposited at Gene Expression Omnibus under acces-sion number GSE50398.
RESULTS
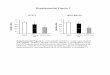
Impaired Insulin Secretion in Sox4mt MiceMice harboring a point mutation (Y123C) in Sox4 (Sox4mtmice) have previously been reported to be glucose intoler-ant and hypoinsulinemic (12), but the precise mechanismby which this mutation results in diabetes is elusive. Asshown in Fig. 1A and B, islets from Sox4mt mice have re-duced (250%) GIIS over a range of glucose concentrations,despite similar insulin content, compared with wild-type(Sox4wt) islets. In dynamic (perifusion) measurements,both Sox4wt and Sox4mt islets responded to an elevationof glucose with a biphasic stimulation of insulin secretion(Fig. 1C), but GIIS was equally reduced at all times, and thesecretion defect could not be corrected by the KATP channelblocker tolbutamide (12).
Glucose-Induced [Ca2+]i Increases, Ca2+ Entry, and
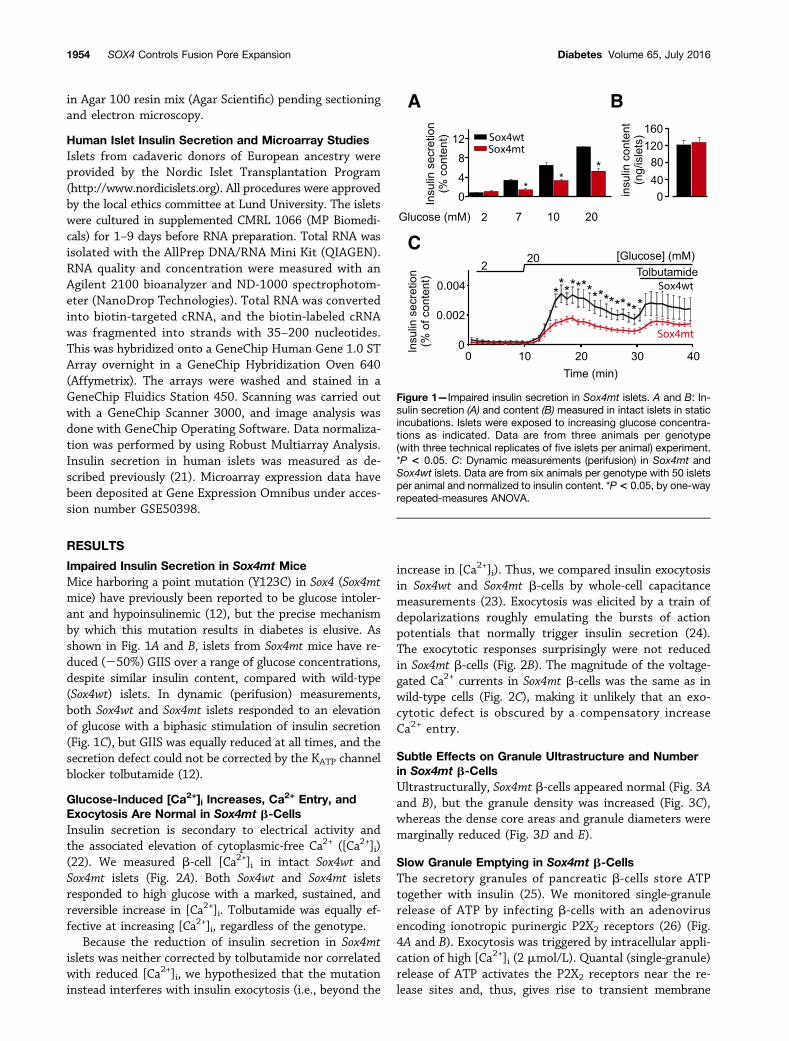
Exocytosis Are Normal in Sox4mt b-CellsInsulin secretion is secondary to electrical activity andthe associated elevation of cytoplasmic-free Ca2+ ([Ca2+]i)(22). We measured b-cell [Ca2+]i in intact Sox4wt andSox4mt islets (Fig. 2A). Both Sox4wt and Sox4mt isletsresponded to high glucose with a marked, sustained, andreversible increase in [Ca2+]i. Tolbutamide was equally ef-fective at increasing [Ca2+]i, regardless of the genotype.
Because the reduction of insulin secretion in Sox4mtislets was neither corrected by tolbutamide nor correlatedwith reduced [Ca2+]i, we hypothesized that the mutationinstead interferes with insulin exocytosis (i.e., beyond the
increase in [Ca2+]i). Thus, we compared insulin exocytosisin Sox4wt and Sox4mt b-cells by whole-cell capacitancemeasurements (23). Exocytosis was elicited by a train ofdepolarizations roughly emulating the bursts of actionpotentials that normally trigger insulin secretion (24).The exocytotic responses surprisingly were not reducedin Sox4mt b-cells (Fig. 2B). The magnitude of the voltage-gated Ca2+ currents in Sox4mt b-cells was the same as inwild-type cells (Fig. 2C), making it unlikely that an exo-cytotic defect is obscured by a compensatory increaseCa2+ entry.
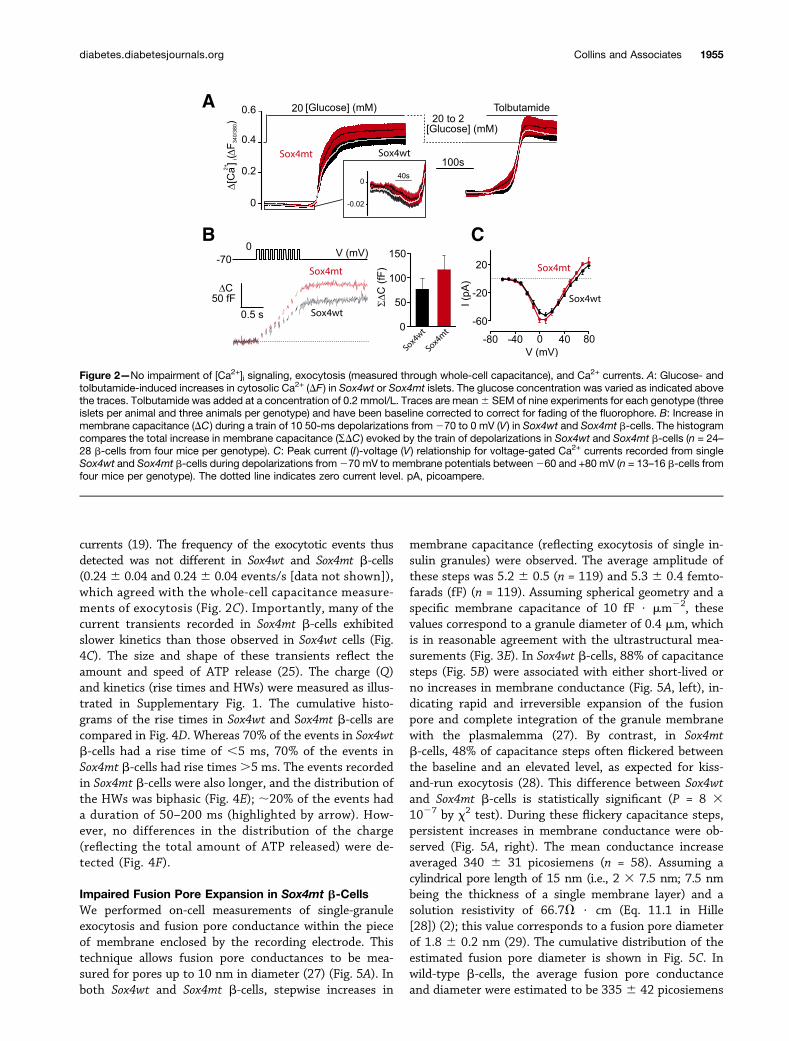
Subtle Effects on Granule Ultrastructure and Numberin Sox4mt b-CellsUltrastructurally, Sox4mt b-cells appeared normal (Fig. 3Aand B), but the granule density was increased (Fig. 3C),whereas the dense core areas and granule diameters weremarginally reduced (Fig. 3D and E).
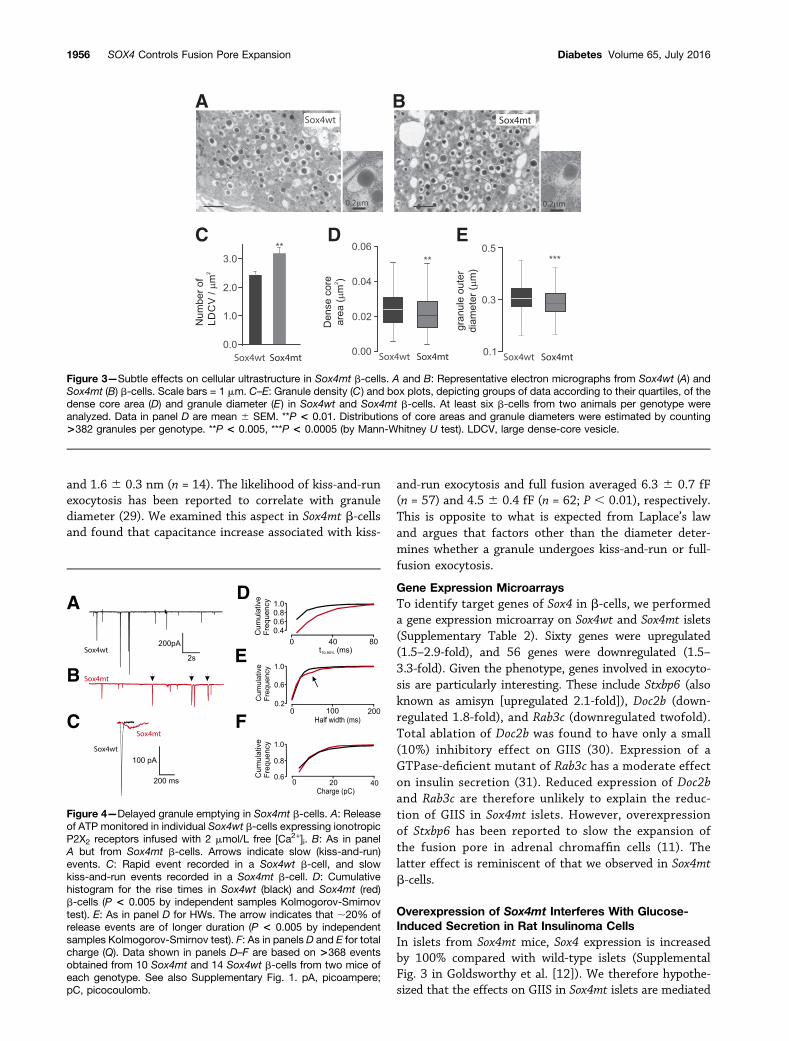
Slow Granule Emptying in Sox4mt b-CellsThe secretory granules of pancreatic b-cells store ATPtogether with insulin (25). We monitored single-granulerelease of ATP by infecting b-cells with an adenovirusencoding ionotropic purinergic P2X2 receptors (26) (Fig.4A and B). Exocytosis was triggered by intracellular appli-cation of high [Ca2+]i (2 mmol/L). Quantal (single-granule)release of ATP activates the P2X2 receptors near the re-lease sites and, thus, gives rise to transient membrane
Figure 1—Impaired insulin secretion in Sox4mt islets. A and B: In-sulin secretion (A) and content (B) measured in intact islets in staticincubations. Islets were exposed to increasing glucose concentra-tions as indicated. Data are from three animals per genotype(with three technical replicates of five islets per animal) experiment.*P < 0.05. C: Dynamic measurements (perifusion) in Sox4mt andSox4wt islets. Data are from six animals per genotype with 50 isletsper animal and normalized to insulin content. *P< 0.05, by one-wayrepeated-measures ANOVA.
1954 SOX4 Controls Fusion Pore Expansion Diabetes Volume 65, July 2016
currents (19). The frequency of the exocytotic events thusdetected was not different in Sox4wt and Sox4mt b-cells(0.246 0.04 and 0.246 0.04 events/s [data not shown]),which agreed with the whole-cell capacitance measure-ments of exocytosis (Fig. 2C). Importantly, many of thecurrent transients recorded in Sox4mt b-cells exhibitedslower kinetics than those observed in Sox4wt cells (Fig.4C). The size and shape of these transients reflect theamount and speed of ATP release (25). The charge (Q)and kinetics (rise times and HWs) were measured as illus-trated in Supplementary Fig. 1. The cumulative histo-grams of the rise times in Sox4wt and Sox4mt b-cells arecompared in Fig. 4D. Whereas 70% of the events in Sox4wtb-cells had a rise time of ,5 ms, 70% of the events inSox4mt b-cells had rise times.5 ms. The events recordedin Sox4mt b-cells were also longer, and the distribution ofthe HWs was biphasic (Fig. 4E); ;20% of the events hada duration of 50–200 ms (highlighted by arrow). How-ever, no differences in the distribution of the charge(reflecting the total amount of ATP released) were de-tected (Fig. 4F).
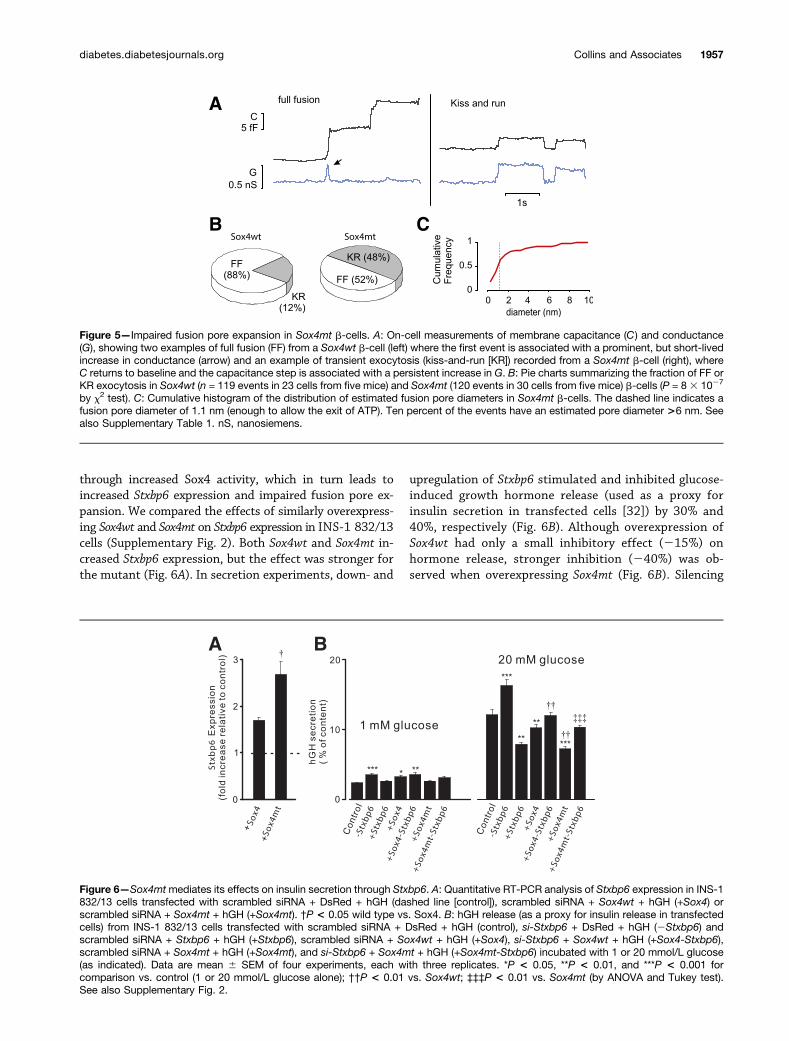
Impaired Fusion Pore Expansion in Sox4mt b-CellsWe performed on-cell measurements of single-granuleexocytosis and fusion pore conductance within the pieceof membrane enclosed by the recording electrode. Thistechnique allows fusion pore conductances to be mea-sured for pores up to 10 nm in diameter (27) (Fig. 5A). Inboth Sox4wt and Sox4mt b-cells, stepwise increases in
membrane capacitance (reflecting exocytosis of single in-sulin granules) were observed. The average amplitude ofthese steps was 5.2 6 0.5 (n = 119) and 5.3 6 0.4 femto-farads (fF) (n = 119). Assuming spherical geometry and aspecific membrane capacitance of 10 fF $ mm22, thesevalues correspond to a granule diameter of 0.4 mm, whichis in reasonable agreement with the ultrastructural mea-surements (Fig. 3E). In Sox4wt b-cells, 88% of capacitancesteps (Fig. 5B) were associated with either short-lived orno increases in membrane conductance (Fig. 5A, left), in-dicating rapid and irreversible expansion of the fusionpore and complete integration of the granule membranewith the plasmalemma (27). By contrast, in Sox4mtb-cells, 48% of capacitance steps often flickered betweenthe baseline and an elevated level, as expected for kiss-and-run exocytosis (28). This difference between Sox4wtand Sox4mt b-cells is statistically significant (P = 8 31027 by x2 test). During these flickery capacitance steps,persistent increases in membrane conductance were ob-served (Fig. 5A, right). The mean conductance increaseaveraged 340 6 31 picosiemens (n = 58). Assuming acylindrical pore length of 15 nm (i.e., 2 3 7.5 nm; 7.5 nmbeing the thickness of a single membrane layer) and asolution resistivity of 66.7V $ cm (Eq. 11.1 in Hille[28]) (2); this value corresponds to a fusion pore diameterof 1.8 6 0.2 nm (29). The cumulative distribution of theestimated fusion pore diameter is shown in Fig. 5C. Inwild-type b-cells, the average fusion pore conductanceand diameter were estimated to be 335 6 42 picosiemens
Figure 2—No impairment of [Ca2+]i signaling, exocytosis (measured through whole-cell capacitance), and Ca2+ currents. A: Glucose- andtolbutamide-induced increases in cytosolic Ca2+ (DF ) in Sox4wt or Sox4mt islets. The glucose concentration was varied as indicated abovethe traces. Tolbutamide was added at a concentration of 0.2 mmol/L. Traces are mean6 SEM of nine experiments for each genotype (threeislets per animal and three animals per genotype) and have been baseline corrected to correct for fading of the fluorophore. B: Increase inmembrane capacitance (DC ) during a train of 10 50-ms depolarizations from270 to 0 mV (V) in Sox4wt and Sox4mt b-cells. The histogramcompares the total increase in membrane capacitance (SDC) evoked by the train of depolarizations in Sox4wt and Sox4mt b-cells (n = 24–28 b-cells from four mice per genotype). C: Peak current (I)-voltage (V ) relationship for voltage-gated Ca2+ currents recorded from singleSox4wt and Sox4mt b-cells during depolarizations from270 mV to membrane potentials between260 and +80 mV (n = 13–16 b-cells fromfour mice per genotype). The dotted line indicates zero current level. pA, picoampere.
diabetes.diabetesjournals.org Collins and Associates 1955
and 1.6 6 0.3 nm (n = 14). The likelihood of kiss-and-runexocytosis has been reported to correlate with granulediameter (29). We examined this aspect in Sox4mt b-cellsand found that capacitance increase associated with kiss-
and-run exocytosis and full fusion averaged 6.3 6 0.7 fF(n = 57) and 4.5 6 0.4 fF (n = 62; P , 0.01), respectively.This is opposite to what is expected from Laplace’s lawand argues that factors other than the diameter deter-mines whether a granule undergoes kiss-and-run or full-fusion exocytosis.
Gene Expression MicroarraysTo identify target genes of Sox4 in b-cells, we performeda gene expression microarray on Sox4wt and Sox4mt islets(Supplementary Table 2). Sixty genes were upregulated(1.5–2.9-fold), and 56 genes were downregulated (1.5–3.3-fold). Given the phenotype, genes involved in exocyto-sis are particularly interesting. These include Stxbp6 (alsoknown as amisyn [upregulated 2.1-fold]), Doc2b (down-regulated 1.8-fold), and Rab3c (downregulated twofold).Total ablation of Doc2b was found to have only a small(10%) inhibitory effect on GIIS (30). Expression of aGTPase-deficient mutant of Rab3c has a moderate effecton insulin secretion (31). Reduced expression of Doc2band Rab3c are therefore unlikely to explain the reduc-tion of GIIS in Sox4mt islets. However, overexpressionof Stxbp6 has been reported to slow the expansion ofthe fusion pore in adrenal chromaffin cells (11). Thelatter effect is reminiscent of that we observed in Sox4mtb-cells.
Overexpression of Sox4mt Interferes With Glucose-Induced Secretion in Rat Insulinoma CellsIn islets from Sox4mt mice, Sox4 expression is increasedby 100% compared with wild-type islets (SupplementalFig. 3 in Goldsworthy et al. [12]). We therefore hypothe-sized that the effects on GIIS in Sox4mt islets are mediated
Figure 3—Subtle effects on cellular ultrastructure in Sox4mt b-cells. A and B: Representative electron micrographs from Sox4wt (A) andSox4mt (B) b-cells. Scale bars = 1 mm. C–E: Granule density (C) and box plots, depicting groups of data according to their quartiles, of thedense core area (D) and granule diameter (E) in Sox4wt and Sox4mt b-cells. At least six b-cells from two animals per genotype wereanalyzed. Data in panel D are mean 6 SEM. **P < 0.01. Distributions of core areas and granule diameters were estimated by counting>382 granules per genotype. **P < 0.005, ***P < 0.0005 (by Mann-Whitney U test). LDCV, large dense-core vesicle.
Figure 4—Delayed granule emptying in Sox4mt b-cells. A: Releaseof ATP monitored in individual Sox4wt b-cells expressing ionotropicP2X2 receptors infused with 2 mmol/L free [Ca2+]i. B: As in panelA but from Sox4mt b-cells. Arrows indicate slow (kiss-and-run)events. C: Rapid event recorded in a Sox4wt b-cell, and slowkiss-and-run events recorded in a Sox4mt b-cell. D: Cumulativehistogram for the rise times in Sox4wt (black) and Sox4mt (red)b-cells (P < 0.005 by independent samples Kolmogorov-Smirnovtest). E: As in panel D for HWs. The arrow indicates that ;20% ofrelease events are of longer duration (P < 0.005 by independentsamples Kolmogorov-Smirnov test). F: As in panels D and E for totalcharge (Q). Data shown in panels D–F are based on >368 eventsobtained from 10 Sox4mt and 14 Sox4wt b-cells from two mice ofeach genotype. See also Supplementary Fig. 1. pA, picoampere;pC, picocoulomb.
1956 SOX4 Controls Fusion Pore Expansion Diabetes Volume 65, July 2016
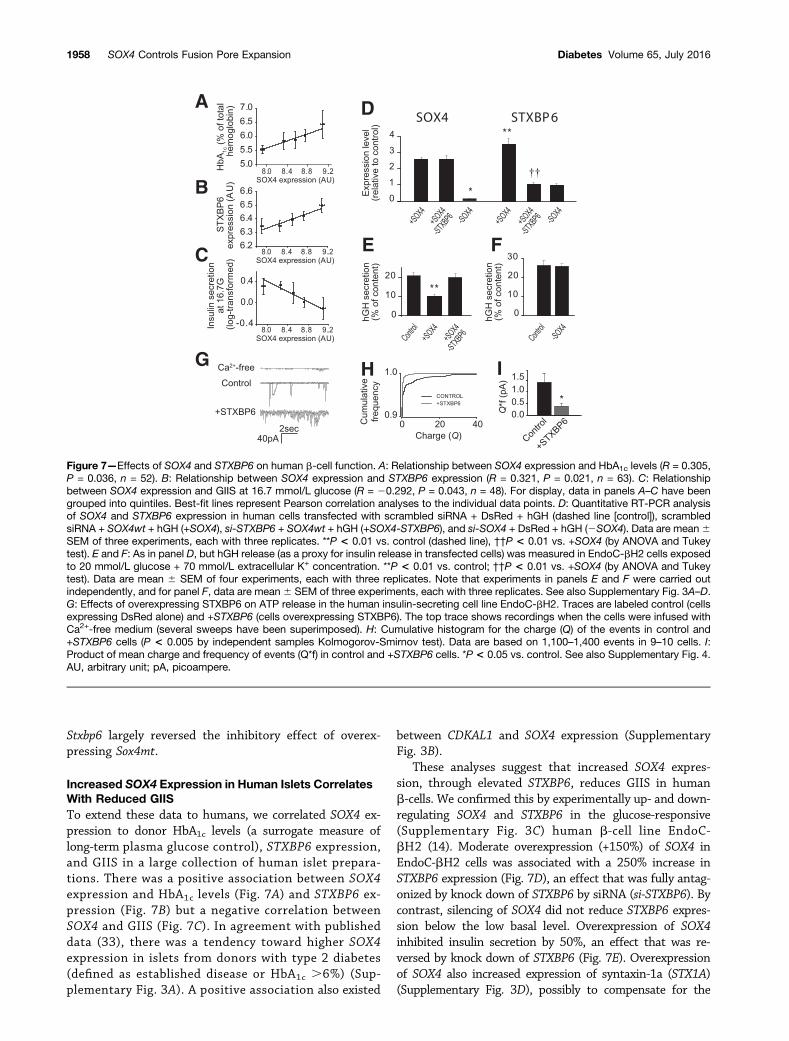
through increased Sox4 activity, which in turn leads toincreased Stxbp6 expression and impaired fusion pore ex-pansion. We compared the effects of similarly overexpress-ing Sox4wt and Sox4mt on Stxbp6 expression in INS-1 832/13cells (Supplementary Fig. 2). Both Sox4wt and Sox4mt in-creased Stxbp6 expression, but the effect was stronger forthe mutant (Fig. 6A). In secretion experiments, down- and
upregulation of Stxbp6 stimulated and inhibited glucose-induced growth hormone release (used as a proxy forinsulin secretion in transfected cells [32]) by 30% and40%, respectively (Fig. 6B). Although overexpression ofSox4wt had only a small inhibitory effect (215%) onhormone release, stronger inhibition (240%) was ob-served when overexpressing Sox4mt (Fig. 6B). Silencing
Figure 5—Impaired fusion pore expansion in Sox4mt b-cells. A: On-cell measurements of membrane capacitance (C) and conductance(G), showing two examples of full fusion (FF) from a Sox4wt b-cell (left) where the first event is associated with a prominent, but short-livedincrease in conductance (arrow) and an example of transient exocytosis (kiss-and-run [KR]) recorded from a Sox4mt b-cell (right), whereC returns to baseline and the capacitance step is associated with a persistent increase in G. B: Pie charts summarizing the fraction of FF orKR exocytosis in Sox4wt (n = 119 events in 23 cells from five mice) and Sox4mt (120 events in 30 cells from five mice) b-cells (P = 8 3 1027
by x2 test). C: Cumulative histogram of the distribution of estimated fusion pore diameters in Sox4mt b-cells. The dashed line indicates afusion pore diameter of 1.1 nm (enough to allow the exit of ATP). Ten percent of the events have an estimated pore diameter >6 nm. Seealso Supplementary Table 1. nS, nanosiemens.
Figure 6—Sox4mtmediates its effects on insulin secretion through Stxbp6. A: Quantitative RT-PCR analysis of Stxbp6 expression in INS-1832/13 cells transfected with scrambled siRNA + DsRed + hGH (dashed line [control]), scrambled siRNA + Sox4wt + hGH (+Sox4) orscrambled siRNA + Sox4mt + hGH (+Sox4mt). †P < 0.05 wild type vs. Sox4. B: hGH release (as a proxy for insulin release in transfectedcells) from INS-1 832/13 cells transfected with scrambled siRNA + DsRed + hGH (control), si-Stxbp6 + DsRed + hGH (2Stxbp6) andscrambled siRNA + Stxbp6 + hGH (+Stxbp6), scrambled siRNA + Sox4wt + hGH (+Sox4), si-Stxbp6 + Sox4wt + hGH (+Sox4-Stxbp6),scrambled siRNA + Sox4mt + hGH (+Sox4mt), and si-Stxbp6 + Sox4mt + hGH (+Sox4mt-Stxbp6) incubated with 1 or 20 mmol/L glucose(as indicated). Data are mean 6 SEM of four experiments, each with three replicates. *P < 0.05, **P < 0.01, and ***P < 0.001 forcomparison vs. control (1 or 20 mmol/L glucose alone); ††P < 0.01 vs. Sox4wt; ‡‡‡P < 0.01 vs. Sox4mt (by ANOVA and Tukey test).See also Supplementary Fig. 2.
diabetes.diabetesjournals.org Collins and Associates 1957
Stxbp6 largely reversed the inhibitory effect of overex-pressing Sox4mt.
Increased SOX4 Expression in Human Islets CorrelatesWith Reduced GIISTo extend these data to humans, we correlated SOX4 ex-pression to donor HbA1c levels (a surrogate measure oflong-term plasma glucose control), STXBP6 expression,and GIIS in a large collection of human islet prepara-tions. There was a positive association between SOX4expression and HbA1c levels (Fig. 7A) and STXBP6 ex-pression (Fig. 7B) but a negative correlation betweenSOX4 and GIIS (Fig. 7C). In agreement with publisheddata (33), there was a tendency toward higher SOX4expression in islets from donors with type 2 diabetes(defined as established disease or HbA1c .6%) (Sup-plementary Fig. 3A). A positive association also existed
between CDKAL1 and SOX4 expression (SupplementaryFig. 3B).
These analyses suggest that increased SOX4 expres-sion, through elevated STXBP6, reduces GIIS in humanb-cells. We confirmed this by experimentally up- and down-regulating SOX4 and STXBP6 in the glucose-responsive(Supplementary Fig. 3C) human b-cell line EndoC-bH2 (14). Moderate overexpression (+150%) of SOX4 inEndoC-bH2 cells was associated with a 250% increase inSTXBP6 expression (Fig. 7D), an effect that was fully antag-onized by knock down of STXBP6 by siRNA (si-STXBP6). Bycontrast, silencing of SOX4 did not reduce STXBP6 expres-sion below the low basal level. Overexpression of SOX4inhibited insulin secretion by 50%, an effect that was re-versed by knock down of STXBP6 (Fig. 7E). Overexpressionof SOX4 also increased expression of syntaxin-1a (STX1A)(Supplementary Fig. 3D), possibly to compensate for the
Figure 7—Effects of SOX4 and STXBP6 on human b-cell function. A: Relationship between SOX4 expression and HbA1c levels (R = 0.305,P = 0.036, n = 52). B: Relationship between SOX4 expression and STXBP6 expression (R = 0.321, P = 0.021, n = 63). C: Relationshipbetween SOX4 expression and GIIS at 16.7 mmol/L glucose (R = 20.292, P = 0.043, n = 48). For display, data in panels A–C have beengrouped into quintiles. Best-fit lines represent Pearson correlation analyses to the individual data points. D: Quantitative RT-PCR analysisof SOX4 and STXBP6 expression in human cells transfected with scrambled siRNA + DsRed + hGH (dashed line [control]), scrambledsiRNA + SOX4wt + hGH (+SOX4), si-STXBP6 + SOX4wt + hGH (+SOX4-STXBP6), and si-SOX4 + DsRed + hGH (2SOX4). Data are mean6SEM of three experiments, each with three replicates. **P < 0.01 vs. control (dashed line), ††P < 0.01 vs. +SOX4 (by ANOVA and Tukeytest). E and F: As in panel D, but hGH release (as a proxy for insulin release in transfected cells) was measured in EndoC-bH2 cells exposedto 20 mmol/L glucose + 70 mmol/L extracellular K+ concentration. **P < 0.01 vs. control; ††P < 0.01 vs. +SOX4 (by ANOVA and Tukeytest). Data are mean 6 SEM of four experiments, each with three replicates. Note that experiments in panels E and F were carried outindependently, and for panel F, data are mean 6 SEM of three experiments, each with three replicates. See also Supplementary Fig. 3A–D.G: Effects of overexpressing STXBP6 on ATP release in the human insulin-secreting cell line EndoC-bH2. Traces are labeled control (cellsexpressing DsRed alone) and +STXBP6 (cells overexpressing STXBP6). The top trace shows recordings when the cells were infused withCa2+-free medium (several sweeps have been superimposed). H: Cumulative histogram for the charge (Q) of the events in control and+STXBP6 cells (P < 0.005 by independent samples Kolmogorov-Smirnov test). Data are based on 1,100–1,400 events in 9–10 cells. I:Product of mean charge and frequency of events (Q*f) in control and +STXBP6 cells. *P < 0.05 vs. control. See also Supplementary Fig. 4.AU, arbitrary unit; pA, picoampere.
1958 SOX4 Controls Fusion Pore Expansion Diabetes Volume 65, July 2016

binding of stxbp6 to syntaxin-1 (34), but this effect is un-likely to explain the suppression of insulin secretion becausea positive correlation between STX1A expression and GIIS hasbeen reported (35). Downregulation of SOX4 affected neitherSTXBP6 expression (Fig. 7D) nor hormone release (Fig. 7F).
To examine the effect of increased STXBP6 expressionon granule emptying in human insulin-secreting cells,we expressed P2X2 receptors in EndoC-bH2 cells andmonitored single-granule ATP release (Fig. 7G). Overex-pression of STXBP6 resulted in a marked decrease in ATPrelease (measured as a decrease in Q) (Fig. 7H). The meancharge of events was reduced by 90%. However, the fre-quency of the events was increased from 1.04 6 0.38 to2.8 6 1.3 Hz (n = 9 or 10; not statistically different).Combining the two effects suggests that there is a 70%reduction in stimulated ATP release following overexpres-sion of STXBP6 (Fig. 7I), which is in reasonable agreementwith the 60% suppression of stimulated hGH release (Fig.6B). Similar results were obtained in INS-1 832/13 cells. NoATP release events were observed when the EndoC-bH2cells were infused with Ca2+-free medium (Fig. 7F, toptrace).
DISCUSSION
Type 2 diabetes is the epitome of a polygenic disorder (1).Genome-wide association studies have led to the identifi-cation of ;120 common gene variants (SNPs) with in-creased type 2 diabetes risk (36). For the majority of thesegene variants, the precise cellular/molecular mechanismsunderlying the increased disease risk remain obscure.The diabetes-associated SNP rs7756992 is commonly re-ferred to CDKAL1, but it has been suggested that thislocus increases disease risk through SOX4 rather thanCDKAL1 (10). Of note, its expression is increased in di-abetic islets (35), but whether increased expression ofSOX4 is causally linked to type 2 diabetes and, if so,the underlying cell biological mechanism has not beenstudied.
In the present study, we show that increased SOX4expression in human pancreatic islets correlates with re-duced GIIS in vitro and increased HbA1c levels in vivo. Wealso found a correlation between CDKAL1 and SOX4 ex-pression in keeping with the idea that the CDKAL1 locusinfluences the expression of both CDKAL1 and SOX4 (10).Furthermore, we observed a tendency toward increasedSOX4 expression in islets from donors with diabetes (Sup-plementary Fig. 3A), which agrees with previous reportsbased on microarray analysis of a b-cell–enriched fractionobtained my laser capture microdissection (35).
To address the mechanisms by which SOX4 may interferewith insulin secretion in humans, we made use of theSox4mt mouse model in which islet Sox4 expression is twicethat of wild-type islet (12). We found that GIIS was reducedby ;50% in Sox4mt islets, a defect that was not correctedby the sulfonylurea tolbutamide. The reduction of insulinsecretion could not be attributed to any impairment of keyfunctional parameters, such as glucose-induced [Ca2+]i, Ca
2+
entry, or exocytosis (determined as depolarization-evokedincreases in whole-cell membrane capacitance).
The delivery of insulin and other granule constituents,like ATP (25), into the extracellular space requires theestablishment and expansion of a fusion pore connectingthe granule lumen and the exterior (27). Normally, theinitial opening of the fusion pore is followed by its rapidexpansion, and following a short delay, the entire granulecollapses into the plasma membrane, ensuring efficientdelivery of the high-molecular-weight cargo into the ex-tracellular space (full fusion). Occasionally, however, fu-sion pore expansion is halted following the initial openingand may eventually close (kiss-and-run exocytosis [27]).
By using on-cell capacitance measurements of individ-ual exocytotic events and the associated fusion poreopenings, we found that full fusion was the predominant(;90%) form of exocytosis in wild-type b-cells, whichagrees with previous reports (29). However, fusion poreexpansion is impaired in Sox4mt b-cells, and the fractionkiss-and-run release events increased to ;50% at theexpense of full fusion. During kiss-and-run events, thefusion pore is locked in a partially expanded state witha diameter of 1–2 nm.
We determined the functional impact of this on theevacuation of the granule lumen by measurements of ATPrelease. The molecular dimensions of ATP are 1.63 1.130.5 nm. With the observed distribution of fusion porediameters, we estimate that ;20% of the total numberof release events in Sox4mt b-cells (i.e., 43% of the 50%showing persistent fusion pore) have fusion pore diame-ters smaller or comparable to the narrowest cross-sectionof ATP (indicated by a dashed line in Fig. 5C). Thus, thefusion pore may function as a molecular sieve that re-stricts the exit of ATP by steric hindrance. This likelyaccounts for the slower rise times of ATP release andthe component of ATP release events with a duration.20 ms (Fig. 4D). Larger molecules like insulin are likelyto face an even greater problem with exiting through thenarrow fusion pore. At concentrations .2 mmol/L, in-sulin exists as a hexamer with a hydrodynamic radiusof 5.6 nm (37). Given that the intragranular insulin con-centration is ;100 mmol/L (22), we conclude that insulinis trapped within the secretory granules undergoing kiss-and-run exocytosis. Indeed, the observed decrease in fullfusion from 88% to 52% is sufficient to account for theobserved 40% reduction of GIIS in Sox4mt islets. It istempting to speculate that the reduction in full fusion ac-counts for the increased granule density observed in Sox4mtb-cells. Although most fusion pores are too small to allowexit of insulin, 10% have a diameter .6 nm. A fusion porediameter as large as this is sufficient to allow the exit ofhexameric insulin, which might explain the slight reductionin dense core area of the insulin granules (Fig. 3D).
Since SOX4 is a transcription factor, it is not surprisingthat the expression of .100 genes were affected inSox4mt islets. However, guided by the functional dataspecifically implicating kiss-and-run exocytosis, we identified
diabetes.diabetesjournals.org Collins and Associates 1959

Stxbp6 (amisyn) as a likely mediator. Overexpression ofStxbp6/STXBP6 mimics the effects of Sox4/SOX4 on secre-tion in rat/human insulinoma cells. Conversely, silencingStxbp6/STXBP6 reverses the inhibitory effect of overexpress-ing Sox4/SOX4. Unexpectedly, and unlike what has previ-ously been observed in rodent insulinoma cells (12),silencing SOX4 in human insulinoma cells did not affecthormone release. The mechanism underlying this dis-crepancy remains unclear.
The present data suggest that increased expression/activity of SOX4, through increased expression of STXBP6and impaired expansion of the fusion pore, plays a rolein insulin secretion and diabetes etiology. This scenarioagrees with the proposal that Stxbp6 forms nonfuso-genic complexes with syntaxin and thereby contributesto the regulation of SNARE complex formation (34). Thisis the first example of a major disease that can be linkedto defective fusion pore expansion. It is possible that themechanisms we have uncovered here may become activatedduring long-term hyperglycemia, a condition reported tobe associated with an increase in kiss-and-run exocytosis (38).It appears that the effect of increased STXBP6 expression isgraded. Whereas a moderate increase (up to +100%) resultsin an increased occurrence of kiss-and-run exocytosis (asseen in the Sox4mt mice), stronger overexpression may re-sult in exocytosis being aborted before granule emptying(Supplementary Fig. 4). In cells overexpressing STXBP6, weobserved small and short events, possibly reflecting repeti-tive openings and closures of the fusion pore, releasing apuff of ATP every time.
If increased SOX4 expression is part of the diseaseetiology and if it acts by interference with fusion poreexpansion, then it may seem paradoxical that type 2 di-abetes is not also associated with defects of neurotrans-mission. There is in fact a link between type 2 diabetesand neuropsychiatric disorders (39). However, slowed orpartial fusion pore expansion will only have a marginaleffect on the release of low-molecular-weight neurotrans-mitters because they are small enough to pass through aconstricted fusion pore.
This study illustrates how detailed cell physiologicalstudies can help us to move from SNPs through theidentification of the actual gene involved to a fullerunderstanding of the causal mechanisms. These findingsraise the interesting possibility that pharmacologicalprocedures promoting full fusion may correct the insulinsecretion defect in type 2 diabetes.
Acknowledgments. The authors thank David Wiggins (Oxford Centre forDiabetes, Endocrinology & Metabolism) for technical assistance and the staff atthe Mary Lyon Centre at MRC Harwell for maintaining the Sox4 mutant mousecolony. The authors also thank Patricia Muller (Leicester University, Leicester, U.K.)for advice on cell transfection and Jochen Lang (Université de Bordeaux, Bordeaux,France) and Raphael Scharfmann (Institut National de la Santé et de la RechercheMédicale [INSERM], Paris, France) for the provision of the INS-1 832/13 andEndoC-bH2 cells, respectively.
Funding. This study was supported by the Medical Research Council (MR/L020149/1 to B.H., R.C., and P.R.), the Wellcome Trust–supported trainingprogramme OXION (to J.G.), the Wellcome Trust (Senior Investigator Award095531/Z/11/Z to P.R.), Swedish Research Council (VR, International Recruitment),and the Knut and Alice Wallenbergs Stiftelse (Wallenberg Scholars Programme).Duality of Interest. No potential conflicts of interest relevant to this articlewere reported.Author Contributions. S.C.C. contributed to the study design, perfor-mance of experiments, data analysis, and drafting and revision of the manuscript.H.W.D., B.H., and A.H. contributed to the performance of experiments, dataanalysis, figure preparation, and drafting of the manuscript. J.A., M.V.C., J.G.,M.God., S.L., M.Gol., A.S., A.I.T., and A.H.R. contributed to the performance ofexperiments and data analysis. R.C. and P.R. contributed to the study design anddrafting and revision of the manuscript. S.C.C. is the guarantor of this work and,as such, had full access to all the data in the study and takes responsibility forthe integrity of the data and the accuracy of the data analysis.
References1. Ashcroft FM, Rorsman P. Diabetes mellitus and the b cell: the last tenyears. Cell 2012;148:1160–11712. MacDonald PE, Braun M, Galvanovskis J, Rorsman P. Release of smalltransmitters through kiss-and-run fusion pores in rat pancreatic beta cells. CellMetab 2006;4:283–2903. McCarthy MI. Genomics, type 2 diabetes, and obesity. N Engl J Med 2010;363:2339–23504. Bonnefond A, Froguel P. Rare and common genetic events in type 2diabetes: what should biologists know? Cell Metab 2015;21:357–3685. Rosengren AH, Braun M, Mahdi T, et al. Reduced insulin exocytosis inhuman pancreatic b-cells with gene variants linked to type 2 diabetes. Diabetes2012;61:1726–17336. Zeggini E, Weedon MN, Lindgren CM, et al.; Wellcome Trust Case ControlConsortium (WTCCC). Replication of genome-wide association signals in UKsamples reveals risk loci for type 2 diabetes. Science 2007;316:1336–13417. Scott LJ, Mohlke KL, Bonnycastle LL, et al. A genome-wide associationstudy of type 2 diabetes in Finns detects multiple susceptibility variants. Science2007;316:1341–13458. Steinthorsdottir V, Thorleifsson G, Reynisdottir I, et al. A variant in CDKAL1influences insulin response and risk of type 2 diabetes. Nat Genet 2007;39:770–7759. Locke JM, Wei FY, Tomizawa K, Weedon MN, Harries LW. A cautionary tale:the non-causal association between type 2 diabetes risk SNP, rs7756992, andlevels of non-coding RNA, CDKAL1-v1. Diabetologia 2015;58:745–74810. Ragvin A, Moro E, Fredman D, et al. Long-range gene regulation linksgenomic type 2 diabetes and obesity risk regions to HHEX, SOX4, and IRX3. ProcNatl Acad Sci U S A 2010;107:775–78011. Constable JR, Graham ME, Morgan A, Burgoyne RD. Amisyn regulatesexocytosis and fusion pore stability by both syntaxin-dependent and syntaxin-independent mechanisms. J Biol Chem 2005;280:31615–3162312. Goldsworthy M, Hugill A, Freeman H, et al. Role of the transcription factorsox4 in insulin secretion and impaired glucose tolerance. Diabetes 2008;57:2234–224413. Shimomura K, Galvanovskis J, Goldsworthy M, et al. Insulin secretion frombeta-cells is affected by deletion of nicotinamide nucleotide transhydrogenase.Methods Enzymol 2009;457:451–48014. Scharfmann R, Pechberty S, Hazhouz Y, et al. Development of a conditionallyimmortalized human pancreatic b cell line. J Clin Invest 2014;124:2087–209815. Collins SC, Hoppa MB, Walker JN, et al. Progression of diet-induced di-abetes in C57BL6J mice involves functional dissociation of Ca2(+) channels fromsecretory vesicles. Diabetes 2010;59:1192–120116. Zhang Q, Chibalina MV, Bengtsson M, et al. Na+ current properties in isleta- and b-cells reflect cell-specific Scn3a and Scn9a expression. J Physiol 2014;592:4677–4696
1960 SOX4 Controls Fusion Pore Expansion Diabetes Volume 65, July 2016

17. Neef A, Heinemann C, Moser T. Measurements of membrane patch capac-itance using a software-based lock-in system. Pflugers Arch 2007;454:335–34418. Braun M, Wendt A, Karanauskaite J, et al. Corelease and differential exit viathe fusion pore of GABA, serotonin, and ATP from LDCV in rat pancreatic betacells. J Gen Physiol 2007;129:221–23119. Obermüller S, Lindqvist A, Karanauskaite J, Galvanovskis J, Rorsman P, BargS. Selective nucleotide-release from dense-core granules in insulin-secretingcells. J Cell Sci 2005;118:4271–428220. Karanauskaite J, Hoppa MB, Braun M, Galvanovskis J, Rorsman P. QuantalATP release in rat beta-cells by exocytosis of insulin-containing LDCVs. PflugersArch 2009;458:389–40121. Rosengren AH, Jokubka R, Tojjar D, et al. Overexpression of alpha2A-adrenergic receptors contributes to type 2 diabetes. Science 2010;327:217–22022. Rorsman P, Renström E. Insulin granule dynamics in pancreatic beta cells.Diabetologia 2003;46:1029–104523. Kanno T, Ma X, Barg S, et al. Large dense-core vesicle exocytosis in pancreaticbeta-cells monitored by capacitance measurements. Methods 2004;33:302–31124. Rorsman P, Eliasson L, Kanno T, Zhang Q, Gopel S. Electrophysiology ofpancreatic b-cells in intact mouse islets of Langerhans. Prog Biophys Mol Biol2011;107:224–23525. Galvanovskis J, Braun M, Rorsman P. Exocytosis from pancreatic b-cells:mathematical modelling of the exit of low-molecular-weight granule content.Interface Focus 2011;1:143–15226. MacDonald PE, Rorsman P. The ins and outs of secretion from pancreaticbeta-cells: control of single-vesicle exo- and endocytosis. Physiology (Bethesda)2007;22:113–12127. Alabi AA, Tsien RW. Perspectives on kiss-and-run: role in exocytosis, en-docytosis, and neurotransmission. Annu Rev Physiol 2013;75:393–42228. Hille B. Ion Channels of Excitable Membranes. Sunderland, MA, Sinauer, 2001
29. MacDonald PE, Obermüller S, Vikman J, Galvanovskis J, Rorsman P, EliassonL. Regulated exocytosis and kiss-and-run of synaptic-like microvesicles in INS-1and primary rat beta-cells. Diabetes 2005;54:736–74330. Li J, Cantley J, Burchfield JG, et al. DOC2 isoforms play dual roles ininsulin secretion and insulin-stimulated glucose uptake. Diabetologia 2014;57:2173–218231. Iezzi M, Escher G, Meda P, et al. Subcellular distribution and function of Rab3A,B, C, and D isoforms in insulin-secreting cells. Mol Endocrinol 1999;13:202–21232. Dubois M, Vacher P, Roger B, et al. Glucotoxicity inhibits late steps of insulinexocytosis. Endocrinology 2007;148:1605–161433. Marselli L, Thorne J, Dahiya S, et al. Gene expression profiles of Beta-cellenriched tissue obtained by laser capture microdissection from subjects with type2 diabetes. PLoS One 2010;5:e1149934. Scales SJ, Hesser BA, Masuda ES, Scheller RH. Amisyn, a novel syntaxin-binding protein that may regulate SNARE complex assembly. J Biol Chem 2002;277:28271–2827935. Andersson SA, Olsson AH, Esguerra JL, et al. Reduced insulin secretioncorrelates with decreased expression of exocytotic genes in pancreatic isletsfrom patients with type 2 diabetes. Mol Cell Endocrinol 2012;364:36–4536. Prasad RB, Groop L. Genetics of type 2 diabetes-pitfalls and possibilities.Genes (Basel) 2015;6:87–12337. Hvidt S. Insulin association in neutral solutions studied by light scattering.Biophys Chem 1991;39:205–21338. Tsuboi T, Ravier MA, Parton LE, Rutter GA. Sustained exposure to highglucose concentrations modifies glucose signaling and the mechanics of se-cretory vesicle fusion in primary rat pancreatic beta-cells. Diabetes 2006;55:1057–106539. Gough SC, O’Donovan MC. Clustering of metabolic comorbidity in schizo-phrenia: a genetic contribution? J Psychopharmacol 2005;19(Suppl.):47–55
diabetes.diabetesjournals.org Collins and Associates 1961