Embed Size (px)
Citation preview

2369Journal of Cell Science 108, 2369-2379 (1995)Printed in Great Britain © The Company of Biologists Limited 1995
Increased microvascular permeability and endothelial fenestration induced by
vascular endothelial growth factor
W. Gregory Roberts* and George E. Palade
Cellular and Molecular Medicine-0651, University of California-San Diego, 9500 Gilman Drive, La Jolla, CA 92093-0651, USA
*Author for correspondence
The vascular endothelial growth factor (VEGF) was origi-nally described as vascular permeability factor due to itsability to increase microvascular permeability to plasmaproteins. However, the vessel types (arteriolar, venular,and capillary) affected by VEGF and the modification ofendothelial morphology in response to increased perme-ability induced by VEGF in vivo have not been preciselydocumented. By topical application or intradermalinjection of recombinant human VEGF-165 we find thatVEGF increases the permeability of postcapillary venulesas well as muscular venules and capillaries. Surprisingly,we also find that endothelia of small venules and capillar-ies become fenestrated within 10 minutes of VEGF appli-
cation. Fenestrations appeared in vascular beds which donot normally have fenestrated endothelium, namely thecremaster muscle and skin. Histamine, saline, and heat-inactivated VEGF do not cause fenestrations. Increasedpermeability is completely inhibited when VEGF is clearedby immunoprecipitation with anti-VEGF monoclonal anti-bodies. The VEGF effect on permeability is unlike that ofany other mediator described to date since both muscularvenules and capillaries are affected.
Key words: vascular endothelial growth factor, microvascularpermeability, fenestrae
SUMMARY
INTRODUCTION
The vascular permeability factor (VPF) was originallydescribed as a tumor secreted protein which caused substantialvascular leakage (Senger et al., 1983). This same protein waslater independently cloned and named vascular endothelialgrowth factor (VEGF) on account of its mitogenic effect onendothelium in culture (Leung et al., 1989). Alternative splicingof VEGF mRNA accounts for 4 isoforms of 121, 165, 189, and206 amino acids, of which the 165 aa form is most abundant invivo (Tischer et al., 1991). The protein is a homodimer of Mr~46,000 and is produced by many cell types including a varietyof tumors, folliculostellate cells, macrophages and possiblypodocytes or capsular epithelial cells in the renal glomeruliamong others (Senger et al., 1993; Brown et al., 1992a;Monacci et al., 1993; Ferrara et al., 1992). As suggested by itsnames, VEGF/VPF has two known functions: (a) it is involvedand possibly primarily responsible for angiogenesis in woundhealing and a variety of pathologies, including diabeticretinopathy, rheumatoid arthritis, and solid tumor growth (Kochet al., 1994; Berkman et al., 1993; Aiello et al., 1994), and it isalso involved in vasculogenesis, since its expression is spatiallyand temporally regulated at times of endothelial proliferation inthe normal embryo (Breier et al., 1992; Millauer et al., 1993).(b) VEGF/VPF is also a potent enhancer of endothelial perme-ability, being 50,000 times more potent than histamine (Sengeret al., 1990). Due to its persistent expression by epithelial cellsadjacent to fenestrated endothelium it has been hypothesized
that it is involved in the maintenance of fenestrae in endothelia(Breier et al., 1992).
There are two related high affinity receptors known forVEGF, fms-like tyrosine kinase (Flt-1) and fetal liver kinase(Flk-1) (both Mr, 196,000) (De Vries et al., 1992; Quinn et al.,1993). As their names suggest, both are tyrosine kinases andappear to be exclusively expressed on endothelial cells con-sistent with the fact that VEGF does not increase mitogenesisin any other cell type (Yamaguchi et al., 1993; Jakeman et al.,1992). They appear in many tissues but at different levels ofexpression, with proliferating vascular endothelial cells (due toeither pathology or normal development) expressing thehighest levels (Plate et al., 1992, 1993). VEGF expression andthat of its receptor are closely regulated to times of vasculo-genesis and angiogenesis, including wound healing (Peters etal., 1993; Brown et al., 1992b; Miller et al., 1994).
In this paper, we concentrate on the ability of VEGF toincrease the permeability of the microvascular endothelium ofthe cremaster muscle and in skin. We demonstrate that VEGFincreases capillary and venular leakage as a result of openingendothelial intercellular junctions, as well as other drastic mor-phological modifications which occur within 10 minutes oftopical application of VEGF. These modifications include theinduction of fenestrae in venular and capillary endotheliawhich normally are not fenestrated. Increased capillary per-meability and the induction of fenestrae in either capillary orvenular endothelium in situ are novel findings which appear tobe a unique feature associated with VEGF.

2370 W. G. Roberts and G. E. Palade
Table 1. Number of specimens Light microscopy Electron microscopy
VEGF Rats (cremaster) 15 15Mice (skin) 20 7
Heat-inactivated VEGFRats 2 2Mice 4 4
SalineRats 7 5Mice 10 6
HistamineRats 6 6Mice 10 4
MATERIALS AND METHODS
Vascular endothelial growth factor preparationThe cDNA encoding human VEGF-165 was kindly provided by DrJudy Abraham (Scios, Inc., Mountain View, CA). The full length cDNAwas subcloned into the PNUT vector and transfected into baby hamsterkidney cells (BHK) subsequently selected for transfection withmethotrexate (200 µg/ml)(Calbiochem, San Diego, CA). For VEGFisolation, BHK growing in 3-tier culture flasks (Nunc, Naperville, IL)were switched from the normal medium conditions (DMEM high-glucose supplemented with L-glutamine (2 mM), penicillin/strepto-mycin (50,000 U/ml), 10% heat-inactivated FCS, and methotrexate (alltissue culture products except methotrexate from UCSD Cell CoreFacility)) to serum-free, protein-free hybrydoma medium (Sigma, StLouis, MO) without supplementation. Medium was conditioned for 24-48 hours, sterile filtered, and run over a heparin-Sepharose column(Pharmacia, Uppsala, Sweden). VEGF was eluted as previouslydescribed (Ferrara et al., 1991) using step elutions of 0.3 M and 0.9 MNaCl at 4°C. All vascular permeability activity (Miles and Miles, 1952)and in vitro endothelial cell growth activity (Ferrara and Henzel, 1989)were eluted as a single peak after 0.9 M NaCl. This fraction was con-centrated in a Centriprep-30 (Amicon, Beverly, MA), sterile filtered,and re-assayed for its activity on endothelial cell growth and increasedmicrovascular permeability. To assess purity, the active fraction wasrun on a 12% PAGE gel under non-reducing conditions and the gelswere silver stained. The protein was reduced by adding 8 M urea and10% (v/v) β-mercaptoethanol to the loading buffer. VEGF was degly-cosylated by treating with N-glycosidase F for 15 minutes at 37°C(Hayes et al., 1993). Western analysis with anti-VEGF monoclonalantibody (mAb) obtained from Dr W. Cavanee and Pharmingen (seeacknowledgements) confirms the presence of VEGF. There was noendothelial cell growth stimulation with vector alone-transfected BHKconditioned media. VEGF was tested for endotoxin using a commer-cially available Limulus Amebocyte Lysate assay (Biowhittaker, Walk-ersville, MD). Preparations containing more than 0.2 EU/ml werediscarded. VEGF was heat-inactivated by boiling for 30 minutes.
AnimalsMale Fisher rats (200-300 g) or male nude Balb/c mice (25-30g) wereused throughout the study. All procedures were in accordance withNIH standards and were reviewed by an institutional animal carecommittee. Table 1 defines the number of animals used for observa-tions at the light and electron microscopic level.
Experimental methodsAnimals were anesthetized with a mixture of ketamine (75 mg/kgbody weight, Lloyd Laboratories, Shenandoah, IA) and xylazine (25mg/kg body weight, Aveco, Co. Fort Dodge, IA). The cremastermuscle was surgically exposed and either saline (100 µl) or VEGF(20 ng in 100 µl) was topically applied. To detect and localizevascular leakage, the saphenous vein was exposed and used to inject0.5 ml/kg body weight Carbon Black (Faber-Castell Corp., Newark,NJ) or Monastral Blue (Sigma) (used as received from company) or20 nm Au-BSA (prepared as previously described; Hayat, 1989). Todetect anionic sites on fenestral diaphragms, cationized ferritin (70mg/kg body weight) (Sigma) was injected intravenously (i.v.)following VEGF or saline application. After 10 minutes, the cremasterwas fixed in situ for 4 minutes with 1% formaldehyde (freshlyprepared from paraformaldehyde), surgically removed, pinned flat ona piece of dental wax and further fixed overnight in 1% formaldehyde.
For experiments on the skin male nude mice were lightly anes-thetized with methoxyfluorane (Pitman-Moore, Inc., Mundelein, IL),and injected intradermally on the abdomen with either saline (30 µl)or VEGF (20 ng in 30 µl). Carbon Black was injected in the tail veinand the skin injection sites were fixed in situ 10 minutes later andfurther processed as described above for the cremaster. VEGF effects
on the vasculature were surveyed under a dissecting microscope to amagnification of 1,000×.
A Miles permeability assay using Balb/c mice or Fisher rats wasperformed to quantitate the ability of anti-VEGF antibodies to blockthe increase in permeability with VEGF. Animals were anesthetized,shaved, and depilated prior to receiving 0.2 ml (0.75 ml for rats) i.v.injection of Evans Blue dye (0.5%, v/v, in sterile saline). Samples tobe tested were prepared by immunoprecipitating 20 ng of VEGF insterile saline with increasing amounts of monoclonal anti-VEGFantibody. Briefly, VEGF was incubated with gentle agitation for 1hour at room temperature with anti-VEGF mAb; Protein G-Sepharose(Pharmacia) was added to the samples which were further incubatedfor 1 hour to remove antigen/antibody complexes. Samples were cen-trifuged 10 minutes at 14,000 rpm in a microcentrifuge and theresulting supernatants were injected intradermally 2 hours after EvansBlue administration. Controls included isotype-matched irrelevantmAb (Pharmingen, San Diego, CA) and VEGF which was incubatedwith Protein G-Sepharose alone. Extraction and quantitation wereperformed as previously described (Roberts and Hasan, 1993).Briefly, after twenty minutes, animals were euthanized and theinjection site (~1 cm diameter) was removed. Samples were solubi-lized in 1 ml of Soluene 350 (Packard Instruments, Downers Grove,IL) overnight at 37°C and then for 1 hour at 57°C. After the solutioncooled to room temperature, ethyl acetate (2 ml) and 1 N HCl (2 ml)were added and the mixture was vortexed, and centrifuged at 2,500rpm for 15 minutes. Absorption of the upper phase was read at 626nm on a Beckman DU-64 spectrophotometer. Data were calculatedusing the extinction coefficient of Evans Blue at 626 nm (ε=7.81e4)and represent the average ± s.e.m. of 8-12 injection sites obtainedfrom 8 animals. Statistical significance was determined using aStudent’s t-test.
Electron microscopyFollowing overnight formaldehyde fixation, specimens were trimmedinto small pieces selected to allow capillaries to be cross-sectioned.They were then fixed with 1.5% glutaraldehyde in 0.1 M sodiumcacodylate-HCl (pH 7.4) with 5% sucrose for 1 hour. Following 3washes with 0.1 M cacodylate buffer with 7.5% sucrose, specimenswere post-fixed in cacodylate-buffered 1% OsO4 on ice for 1 hour,en-bloc stained with uranyl acetate for 1 hour, dehydrated, embeddedin EPON 812, and cured for 18-24 hours at 60°C. Thin sections werecut, picked up on nickel grids, stained with uranyl acetate and leadcitrate before examination and micrography on a Phillips CM10electron microscope at 80 kV.
RESULTS
VEGF characterization and analysisRecombinant human VEGF-165 was produced by transfected

2371VEGF induces endothelial fenestrae
BHK cells grown in serum-free, protein-free medium andpurified over a heparin-Sepharose column. As determined bySDS-PAGE and silver staining, the purified VEGF was notdetectably contaminated with any other proteins. The proteinmigrated as a doublet of approximately 44 kDa, presumablydue to glycosylation variants (Fig. 1, lane 1). Upon N-degly-cosylation only one band at 38 kDa was observed (lane 2).Often at high VEGF concentrations a 60-66 kDa band recog-nized by the antibody would also appear. Deglycosylation orreduction of this 66 kDa protein results in a single bandmigrating at the appropriate Mr. The anti-VEGF mAb recog-nized the non-reduced glycosylated 44 kDa form (lane 3), thereduced glycosylated form at 22 kDa (lane 4), the non-reduceddeglycosylated form at 38 kDa, and the aggregated 66 kDaform (remnants can be seen in lane 4).
The cremaster muscle model demonstrates the levelof VEGF effect on the microvascular treeAt the light microscope level, topical VEGF application led toextensive tatoo of postcapillary venules and less intense, butclearly detectable, tatoo of capillaries (Fig. 2). Capillaries arerecognizable due to their small size and the fact that they runparallel with muscle fibers (Fig. 2b). It is already known thatvascular tatoo is due to opening of endothelial intercellularjunctions and retention of the Carbon Black tracer by basementmembrane in intramural deposits (Joris et al., 1990). Openingof junctions in venules appears to be extensive, so much so thatintramural deposits outline individual endothelial cells (Fig.2c). The size of the vessels affected by VEGF is greater thanwith other known inflammatory mediators such as histamine,since both small postcapillary venules (arrow) and largermuscular venules (arrowhead) demonstrate extensive leakage(Fig. 2c). Topical administration of VEGF on the cremasteralso causes substantial leakage in capillaries (Fig. 2a,b) leading
kDa
Fig. 1. After elution from heparin-Sepharose affinitychromatography, recombinant human vascular endothelial growthfactor-165 was electrophoresed through a 12% SDS-PAGE gel andsilver-stained. The protein migrates as a doublet, possibly due todifferential glycosylation, at an apparent Mr of 44,000, lane 1. AfterN-glyconase digestion, a single band migrates at Mr 38,000, lane 2.A monoclonal Ab to human VEGF-165 was used forimmunoblotting of VEGF after transfer to PVDF membrane. Thenon-reduced protein is shown in lane 3. Upon reduction, the growthfactor migrates at Mr 22,000; a remnant of an aggregated form (Mr66,000) is also visible (lane 4). The antibody also recognizes thedeglycosylated form of VEGF (lane 5).
to smaller intramural deposits than in venules. All controls(saline or heat-inactivated VEGF) were negative (salineshown, Fig. 2d).
Electron microscopy also reveals that the leakage resultsfrom an extensive opening of endothelial intercellularjunctions (Fig. 3, arrowheads indicate limits of openings).These gaps are large enough to allow the entry of not only thetracer, but circulating cells such as red blood cells (not shown)and platelets (Fig. 3a) into intramural deposits. Many depositsare not in phase with the open junctions because of thetendency of deposits to dissect the vessel wall. Platelet and redblood cell stasis within the lumen in larger vessels, adhesionof platelets to endothelial cells and platelet migration inbetween endothelial cells to reach the basement membranewere common findings. More interesting, however, may be theobservation that platelets and mast cells were not degranulated,suggesting an endothelial cell specific response. Opening of thejunctions and formation of intramural deposits (made upprimarily by the tracer) were also observed in capillaryendothelium (Fig. 3b).
The permeability enhancing activity of VEGF is not limitedto opening endothelial intercellular junctions. Surprisingly, acommon finding on VEGF-treated vessels was the appearanceof endothelial fenestrae. The microvasculature of the cremastermuscle, as with most skeletal muscles, is provided with a con-tinuous endothelium. Endothelial fenestration was found onvenules as shown in Fig. 4a,b as well as capillaries (Fig. 5a,b).These fenestrae generally have a diaphragm and are 60 nm indiameter. In appropriate sections, the characteristic centralknob is observed in essentially all diaphragms (Figs 4b,c, 5a,b,arrowheads). In addition to single diaphragmed fenestrae,transendothelial channels with diaphragms on the luminal andabluminal aspects of the endothelium were found (Fig. 5b,asterisk). No fenestrated endothelium was observed in controltissues. It is known that the luminal aspects of the fenestraldiaphragms in normally fenestrated endothelia are anionic andcan be coated with exogenous cationized ferritin (Simionescuet al., 1981b). As demonstrated in Figs 4c and 5c, the VEGF-induced fenestrae in cremaster vessels are labelled by cation-ized ferritin. The adjacent plasmalemma and stomataldiaphragms on plasmalemmal vesicles are not coated. Coatingby cationized ferritin, however, was variable, since ~70-80%of the fenestrae were labelled (Fig. 4c). Results similar to thosedescribed in the rat cremaster were also observed in the murinecremaster.
Fenestration of skin microvasculature after VEGFadministrationIn an effort to further characterize endothelial morphologicalchanges following VEGF administration, we studied themicrovasculature of normal skin. As with skeletal muscle, theskin microvasculature is provided with a continuous endothe-lium and rarely, if ever, is fenestrated endothelium observed.Similar to the cremaster, skin microvasculature becomestatooed by tracer particles after exposure to VEGF (Fig. 6a).Saline-treated skin vasculature appears normal and does notallow the escape of systemically injected tracer to suben-dothelial spaces (Fig. 6b). The leakage caused by VEGF affectsboth capillaries (Figs 6a, 7b,c) and venules (Fig. 7a). In somevessels, both open junctions and fenestrations are observed(Fig. 7a, venule, c, capillary). Another common finding in

2372 W. G. Roberts and G. E. Palade
Fig. 2. Preparation of a rat cremaster 10minutes after topical application of 20ng of VEGF (a). Vascular leakdemonstrated by the tatooing of vesselswith Carbon Black is evident incapillaries (b) and postcapillary (arrow),as well as, muscular venules(arrowhead) (c). The asterisks in (a)outline the capillary field shown in (b).The opening of junctions caused byVEGF is so extensive that individualendothelial cells are outlined in somevenules (c). Carbon Black leakage is notseen in a saline-treated cremaster (d).Bars: (a) 50 µm; (b,c) 40 µm; (d) 50 µm.
endothelium exposed to VEGF is dilated plasmalemmalvesicles which aggregate to form complexes in which vesiclesare fused to one another and diaphragms provided with centralknobs are found in between the vesicles. (Figs 6a, 7a, asterisk).These complexes were also seen in VEGF-treated cremastermicrovasculature. Interestingly, similar complexes describedas ‘vesiculo-vacuolar organelles’ have been found in tumorvasculature, although in greater numbers (Kohn et al., 1992).
Adjacent to these complexes, diaphragms with their centralknob and radiating fibers can be identified (Fig. 7a, arrow-heads). They may represent either fenestral or stomatal or otherdiaphragms generated between fusing vesicles. Other subcel-lular organelles, such as endoplasmic reticulum, Golgicomplexes, mitochondria, and nucleus, appear normal. As inthe cremaster muscle, VEGF can also induce extensive fenes-tration in skin capillaries (Fig. 7b,c). None of these morpho-
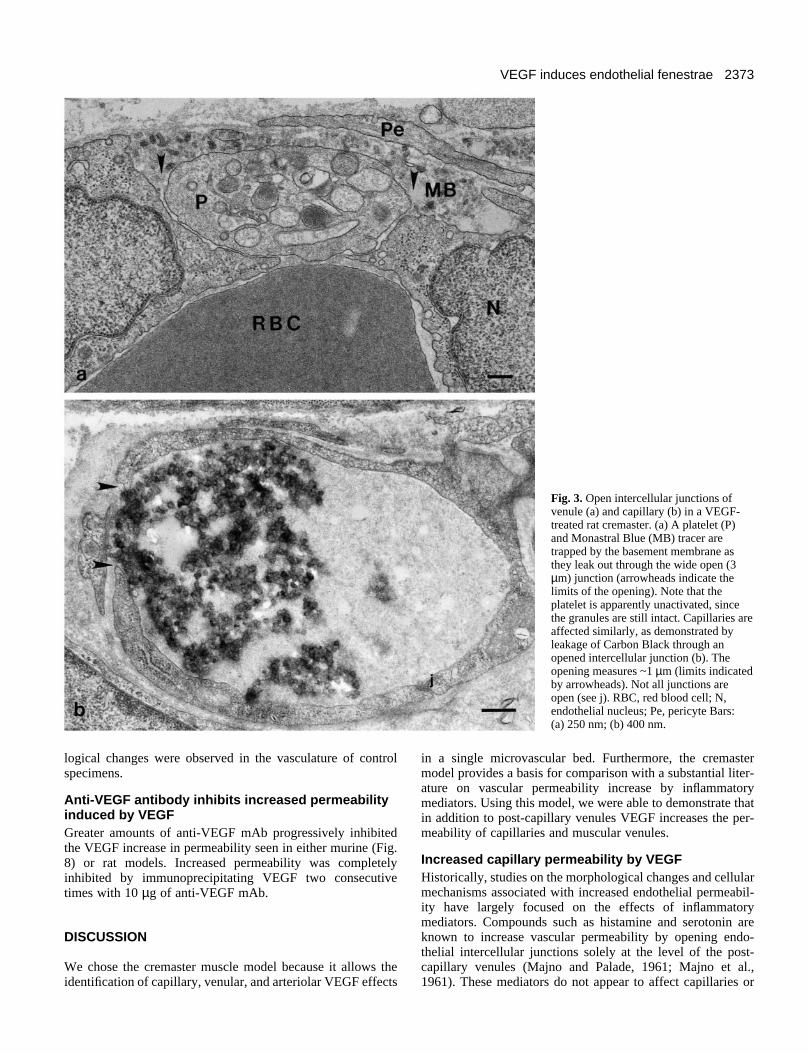
2373VEGF induces endothelial fenestrae
Fig. 3. Open intercellular junctions ofvenule (a) and capillary (b) in a VEGF-treated rat cremaster. (a) A platelet (P)and Monastral Blue (MB) tracer aretrapped by the basement membrane asthey leak out through the wide open (3µm) junction (arrowheads indicate thelimits of the opening). Note that theplatelet is apparently unactivated, sincethe granules are still intact. Capillaries areaffected similarly, as demonstrated byleakage of Carbon Black through anopened intercellular junction (b). Theopening measures ~1 µm (limits indicatedby arrowheads). Not all junctions areopen (see j). RBC, red blood cell; N,endothelial nucleus; Pe, pericyte Bars: (a) 250 nm; (b) 400 nm.
logical changes were observed in the vasculature of controlspecimens.
Anti-VEGF antibody inhibits increased permeabilityinduced by VEGFGreater amounts of anti-VEGF mAb progressively inhibitedthe VEGF increase in permeability seen in either murine (Fig.8) or rat models. Increased permeability was completelyinhibited by immunoprecipitating VEGF two consecutivetimes with 10 µg of anti-VEGF mAb.
DISCUSSION
We chose the cremaster muscle model because it allows theidentification of capillary, venular, and arteriolar VEGF effects
in a single microvascular bed. Furthermore, the cremastermodel provides a basis for comparison with a substantial liter-ature on vascular permeability increase by inflammatorymediators. Using this model, we were able to demonstrate thatin addition to post-capillary venules VEGF increases the per-meability of capillaries and muscular venules.
Increased capillary permeability by VEGFHistorically, studies on the morphological changes and cellularmechanisms associated with increased endothelial permeabil-ity have largely focused on the effects of inflammatorymediators. Compounds such as histamine and serotonin areknown to increase vascular permeability by opening endo-thelial intercellular junctions solely at the level of the post-capillary venules (Majno and Palade, 1961; Majno et al.,1961). These mediators do not appear to affect capillaries or

2374 W. G. Roberts and G. E. Palade
Fig. 4. VEGF induces endothelialfenestrations in venules of the ratcremaster within 10 minutes ofapplication. In an otherwise normalappearing venule, the endotheliumbecomes attenuated and fenestrae appear(a). A closeup of the fenestrae is shownin b (outlined by arrows in a).Cationized ferritin (arrow) was used tolabel the anionic proteoglycans whichconcentrate on fenestral diaphragms (c).The characteristic central fenestral knobcan be seen in many of the fenestrae(arrowheads). Most fenestrae werelabelled with CF, however, within thesame vessel some fenestrae were labeledwhile others were not. Bars: (a) 500 nm;(b,c) 100 nm.
arterioles. In fact, increased capillary permeability in responseto exogenous mediators is rare. These observations, maderepeatedly over many years, have led to the generally acceptedbelief that increased permeability is due to the opening of post-capillary (or pericytic) venular intercellular junctions. The per-meability enhancing effect of VEGF was also believed to occurprimarily on venules in the same manner.
Increased capillary permeability, although rare, is known tooccur in chronic inflammation (De Almeida and Bohm, 1978).It was also observed as a result of in situ resorption of avasculartissue in the cremaster model (Joris et al., 1990). Recently,VEGF has been found to be involved in rheumatoid arthritis(Koch et al., 1994) and to be present in inflammatory effusions(Yeo et al., 1993), although its role in these pathologies is notwell understood. Capillary permeability is also known to occurduring wound healing (Schoefl, 1963; Abell, 1946) and VEGFappears to be involved in wound healing as evidenced by theoverexpression of its receptor on endothelium of regeneratingvessels (Peters et al., 1993; Millauer et al., 1993).
Further evidence of the involvement of VEGF in inflamma-tion reactions is suggested by the adhesion and migrationthrough endothelial intercellular junctions of platelets observedin a number of vessels. VEGF causes the release of von Wille-brand factor from endothelial cells (Brock et al., 1991), storedin Weibel-Palade bodies. P-selectin, also stored in themembranes of these granules, can mediate the adhesion of neu-
trophils and monocytes to endothelium (McEver et al., 1989;Bonafanti et al., 1989). Therefore, VEGF would be expectedto increase the surface expression of P-selectin on endo-thelium, thereby enhancing neutrophil and monocyte adhesion.Interestingly, VEGF has been shown to increase monocytemigration through endothelium in culture (Clauss et al., 1990),possibly due to increased endothelial cell surface P-selectin.The fact that platelets are not degranulated, yet adhere toendothelium in response to VEGF administration suggests thatan endothelial cell-platelet adhesion molecule may also beinvolved, such as CD31 (platelet-endothelial cell adhesionmolecule, PECAM). CD31 is normally found at endothelialintercellular junctions (Muller et al., 1989; Albelda et al.,1991), but its cell surface distribution is modified upon stimu-lation to proliferate (Schimmenti et al., 1992). This redistribu-tion can be mimicked in vitro on HUVEC with VEGF (unpub-lished results).
VEGF induces the formation of endothelialfenestrationsThe finding that topical administration of VEGF couldtransform continuous endothelium into fenestrated enotheliumin both venules and capillaries was completely unexpected.Since VEGF expression in the adult kidney and brain istypically higher in cells adjacent to fenestrated endothelium,the hypothesis that VEGF is involved in inducing and/or main-

2375VEGF induces endothelial fenestrae
Fig. 5. Rat cremaster capillary 10minutes after VEGF administration.Numerous fenestrations appearthroughout the endothelial cell (a).Intracellular organelles such asmitochondria, endoplasmic reticulum,Golgi complex, and endosomes appearnormal. (b) Closeup outlined by arrowsin (a) showing a diaphragmed fenestrawith central knob (arrowhead) and anendothelial channel (asterisk). (c) venulewith the luminal surface of thediaphragm labelled with cationizedferritin (arrow). Bars: (a) 300 nm; (b,c) 100 nm.
Fig. 6. Intradermal injection of VEGF (20 ng) increased vascular permeability of murine skin capillaries, as demonstrated by intramuraldeposits of Carbon Black (a), while there were no deposits after saline injection (b). Bars, 500 nm.
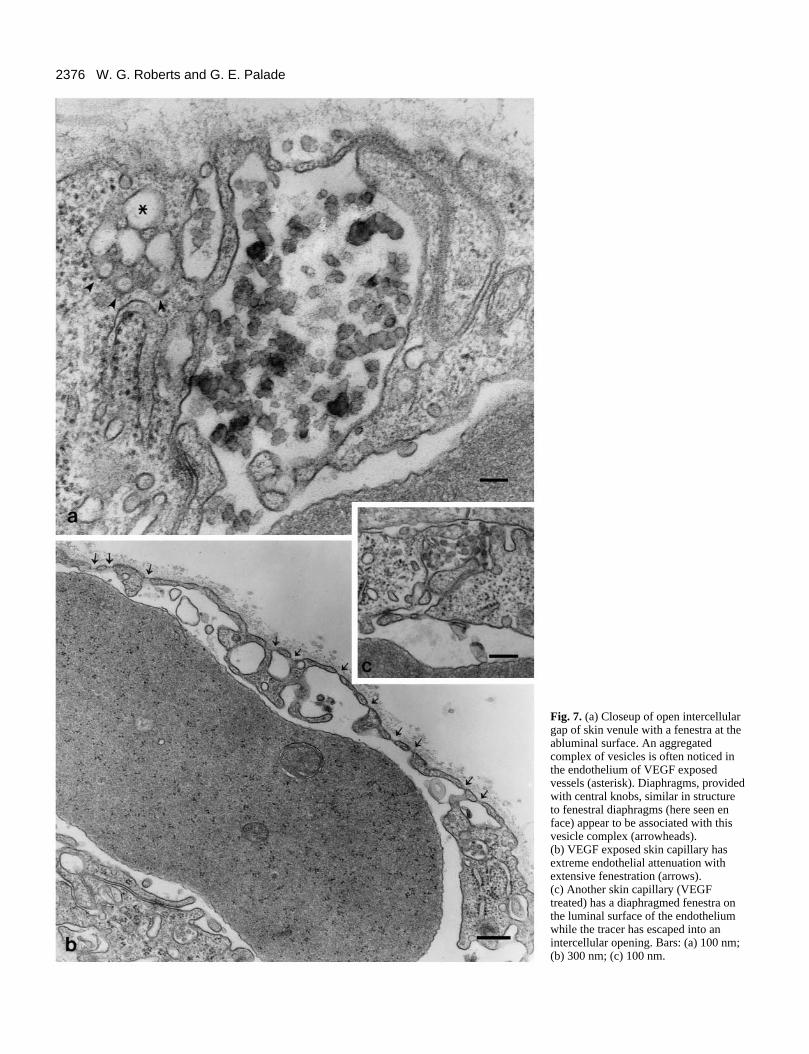
2376 W. G. Roberts and G. E. Palade
Fig. 7. (a) Closeup of open intercellulargap of skin venule with a fenestra at theabluminal surface. An aggregatedcomplex of vesicles is often noticed inthe endothelium of VEGF exposedvessels (asterisk). Diaphragms, providedwith central knobs, similar in structureto fenestral diaphragms (here seen enface) appear to be associated with thisvesicle complex (arrowheads). (b) VEGF exposed skin capillary hasextreme endothelial attenuation withextensive fenestration (arrows). (c) Another skin capillary (VEGFtreated) has a diaphragmed fenestra onthe luminal surface of the endotheliumwhile the tracer has escaped into anintercellular opening. Bars: (a) 100 nm;(b) 300 nm; (c) 100 nm.

2377VEGF induces endothelial fenestrae
P<0.02P<0.01
P<0.005
Fig. 8. Inhibition of increased permeability induced by VEGF inmurine skin with anti-VEGF mAb. A Miles assay was performedusing VEGF (20 ng) which had been reacted with increasingamounts of anti-VEGF mAb. The immunoprecipitate was removedand the cleared supernatant was injected intradermally into micewhich had received a 0.2 ml i.v. injection of 0.5% Evans Blue dye 2hours earlier. Twenty minutes after intradermal injection, theinjection sites were excised and Evans Blue extracted. Controlsconsisted of VEGF immunoprecipitated with isotype matchedirrelevant mAb (C) or with protein G-Sepharose alone (V). Completeinhibition of increased permeability was observed byimmunoprecipitating VEGF two consecutive times with 10 µg anti-VEGF mAb. Data represent the average and s.e.m. of 8-12 injectionsites. Statistical significance versus the control (C) was determinedusing the Student’s t-test.
taining fenestrated endothelium has been put forth (Brown etal., 1992a; Breier et al., 1992). More data suggesting theinvolvement of VEGF with fenestrated endothelium comesfrom studies demonstrating that retinal pigment epithelial cellssecrete VEGF in vitro and in situ (Adamis et al., 1993) andendothelium encapsulated by retinal pigment epitheliumbecomes fenestrated (Burns and Hartz, 1992). Additionally,skeletal muscle capillaries growing into a wound become fen-estrated (McKinney et al., 1977). Lastly, vessels in psoriaticlesions become fenestrated (Braverman and Yen, 1977) andboth VEGF and its receptor are upregulated in psoriatic skin(Detmar et al., 1994). Taken together, these findings implicatedan involvement of VEGF with endothelial fenestration, butthese observations are only correlative and speculative. To ourknowledge, our findings present the first evidence of VEGFdirectly and promptly inducing fenestrae.
Fenestrated endothelium normally occurs in the capillariesof endocrine organs, kidney glomeruli, and visceral mucosae(Farquhar, 1961; Bearer and Orci, 1985; Clementi and Palade,1969). These locations are generally characterized by theirincreased filtration (as in the case of the glomeruli) or increasedtransendothelial transport (as with endocrine organs andintestinal mucosae). Morphologically, fenestrae are regular intheir dimensions, averaging 60 nm in diameter and often tran-sected by a diaphragm (Bearer and Orci, 1985) which consistsof a radial array of fibers that come together in the centerappearing as a knob. Fenestrated capillary endothelium haslong been known to be more permeable to water and smallsolutes than continuous endothelium (Clementi and Palade,1969; Granger et al., 1979). The relative contribution of open
intercellular junctions versus fenestrations in VEGF-inducedpermeability is difficult to assess since the tracers used tomeasure fenestral permeability will also leak through openjunctions. Certainly, protein efflux and escape of cells is dueto opened junctions, while fenestrations will allow theincreased efflux of small solutes. It is likely that bothphenomena are present in VEGF-related pathologies, since ourpreliminary observations demonstrate extensive open intercel-lular junctions and endothelial fenestration in tumor vascula-ture (Roberts and Palade, 1995). However, these results are notin full agreement with those of Ferrara et al. (1993) since theyused VEGF-transfected CHO cells to generate ‘tumors’ andfound that most of the capillaries were of the continuous type.
The fenestrae and fenestral diaphragms detected in ourstudies are similar in morphology to those found in naturallyfenestrated endothelium (Clementi and Palade, 1969; Bearerand Orci, 1985). Furthermore, the unique distribution ofanionic sites, as demonstrated with cationic ferritin labelling,is present on many but not all of the fenestral diaphragmsformed by VEGF administration. This is especially strikingwhen one considers the time frame of these experiments. Thereis a redistribution and concentration of anionic sites within 10minutes. It has been demonstrated that the anionic charge onfenestral diaphragms was removed by heparinase thereforeprovided by heparin and heparan sulfate (Simionescu et al.,1981a). It is worth noting that VEGF (as well as many othergrowth factors, e.g. bFGF) strongly binds heparin and heparansulfate and that heparin enhances the binding of VEGF to itsreceptor (Gitay-Goren et al., 1992). The finding that fusedvesicles have a characteristic fenestral diaphragm betweenthem suggests that all the structural elements necessary to formfenestral diaphragms preexist in the vesicle membranes. Infact, it has been speculated that fenestral and stomataldiaphragms, in general, are the result of the fusion of plas-malemmal vesicle membranes with the plasmlemma proper(Clementi and Palade, 1969).
Our results demonstrate that VEGF can directly increase notonly venular but also capillary permeability. Additionally, theapplication of VEGF can rapidly induce fenestrations inendothelium from microvascular beds where fenestratedendothelium is not normally present. Many questions remainunanswered, such as, the persistance of the fenestrationsfollowing VEGF application and the mechanism of forming thefenestrae. How VEGF can mediate two seemingly differentendothelial effects, namely increased permeability and angio-genesis is unclear. It is possible that these responses are due todifferent VEGF isoforms and/or different receptors, variedlocalization of the receptors, or the result of different signaltransduction pathways modulated by the two receptors. Studiesare presently underway to elucidate the mechanisms involvedin differentiating these two responses.
The authors thank Dr Judy Abraham for the VEGF cDNA, Drs H.-J. S. Huang, W. Arap, W. Cavanee, and C.-H. M. Huang for providingthe G143-264 anti-VEGF monoclonal antibody prior to publication,Dr Ajit Varki and members of his laboratory for supplying theprotocol and reagents of the deglycosylation assay, Dr Dan Predescufor assistance with the cremaster model, and Mike McCaffery andLuann Woodward for helpful suggestions with electron microscopy.In preliminary, abbreviated form (abstract), these findings werepresented at the 1994 American Society for Cell Biology meeting andthe 1995 International Symposium on Endothelium-Derived Factors.

2378 W. G. Roberts and G. E. Palade
REFERENCES
Abell, R. G. (1946). Permeability of blood capillary sprouts and newly formedblood capillaries as compared to that of older blood capillaries. Am. J.Physiol. 147, 237-241.
Adamis, A. P., Shima, D. T., Yeo, K-T., et al. (1993). Synthesis and secretionof vascular permeability factor/vascular endothelial growth factor by humanretinal pigment epithelial cells. Biochem. Biophys. Res. Commun. 193, 631-638.
Aiello, L. P., Avery, R. L., Arrigg, P. G., et al. (1994). Vascular endothelialgrowth factor in ocular fluid of patients with diabetic retinopathy and otherretinal disorders. New Engl. J. Med. 331, 1480-1519.
Albelda, S. M., Muller, W. A., Buck, C. A. and Newman, P. J. (1991).Molecular and cellular properties of PECAM-1 (endoCAM/CD31): A novelvascular cell-cell adhesion molecule. J. Cell Biol. 114, 1059-1068.
Bearer, E. L. and Orci, L. (1985). Endothelial fenestral diaphragms: A quick-freeze, deep-etch study. J. Cell Biol. 100, 418-428.
Berkman, R. A., Merrill, M. J., Reinhold, W. C., et al. (1993). Expression ofthe vascular permeability factor/vascular endothelial growth factor gene incentral nervous system neoplasms. J. Clin. Invest. 91, 153-159.
Bonafanti, R., Furie, B. C., Furie, B. and Wagner, D. D. (1989). PADGEM(GMP140) is a component of Weibel-Palade bodies of human endothelialcells. Blood 73, 1109-1112.
Braverman, I. M. and Yen, A. (1977). Ultrastructure of the capillary loops inthe dermal papillae of psoriasis. J. Invest. Derm. 68, 53-60.
Breier, G., Albrecht, U., Sterrer, S. and Risau, W. (1992). Expression ofvascular endothelial growth factor during embryonic angiogenesis andendothelial cell differentiation. Development 114, 521-532.
Brock, T. A., Dvorak, H. F. and Senger, D. R. (1991). Tumor-secretedvascular permeability factor increases cytosolic Ca++ and von Willebrandfactor release in human endothelial cells. Am. J. Path. 138, 213-221.
Brown, L. F., Berse, B., Tognazzi, K., et al. (1992a). Vascular permeabilityfactor mRNA and protein expression in human kidney. Kidney Int. 42, 1457-1461.
Brown, L. F., Yeo, K.-T., Berse, B., et al. (1992b). Expression of vascularpermeability factor (vascular endothelial growth factor) by epidermalkeratinocytes during wound healing. J. Exp. Med. 176, 1375-1379.
Burns, M. S. and Hartz, M. J. (1992). The retinal pigment epithelium inducesfenestration of endothelial cells in vivo. Curr. Eye Res. 11, 863-873.
Clauss, M., Gerlach, M., Gerlach, H., et al. (1990). Vascular permeabilityfactor: A tumor-derived polypeptide that induces endothelial cell andmonocyte procoagulant activity, and promotes monocyte migration. J. Exp.Med. 172, 1535-1545.
Clementi, F. and Palade, G. E. (1969). Intestinal capillaries. I. Permeability toperoxidase and ferritin. J. Cell Biol. 41, 33-58.
De Almeida, O. P. and Bohm, G. M. (1978). Vascular permeability in the ratgingiva. A model of vessel response in chronic inflammation. Am. J. Path.127, 27-34.
De Vries, C., Escobedo, J. A., Ueno, H., Houck, K., Ferrara, N. andWilliams, L. T. (1992). The fms-like tyrosine kinase, a receptor for vascularendothelial growth factor. Science 255, 989-991.
Detmar, M., Brown, L. F., Claffey, K. P., et al. (1994). Overexpression ofvascular permeability factor/vascular endothelial growth factor and itsreceptors in psoriasis. J. Exp. Med. 180, 1141-1146.
Farquhar, M. G. (1961). Fine structure and function in capillaries of theanterior pituitary gland. Angiology 12, 270-292.
Ferrara, N. and Henzel, W. J. (1989). Pituitary follicular cells secrete a novelheparin-binding growth factor specific for vascular endothelial cells.Biochem. Biophys. Res. Commun. 161, 851-858.
Ferrara, N., Leung, D. W., Cachianes, G., Winer, J. and Henzel, W. J.(1991). Purification and cloning of vascular endothelial growth factorsecreted by pituitary folliculostellate cells. Meth. Enzymol. 198, 391-405.
Ferrara, N., Houck, K., Jakeman, L. and Leung, D. W. (1992). Molecularand biological properties of the vascular endothelial growth factor family ofproteins. Endo. Rev. 13, 18-32.
Ferrara, N., Winer, J., Burton, T., et al. (1993). Expression of vascularendothelial growth factor does not promote transformation but confers agrowth advantage in vivo to chinese hamster ovary cells. J. Clin. Invest. 91,160-170.
Gitay-Goren, H., Soker, S., Vlodavsky, I. and Neufeld, G. (1992). Thebinding of vascular endothelial growth factor to its receptors is dependent oncell surface-associated heparin-like molecules. J. Biol. Chem. 267, 6093-6098.
Granger, D. N., Granger, J. P., Brace, R. A., Parker, R. E. and Taylor, A. E.
(1979). Analysis of the permeability characteristics of cat intestinalcapillaries. Circ. Res. 44, 335-344.
Hayat, M. A. (1989). Colloidal Gold. Principles, Methods and Applications.Vol. 1. Academic Press, San Diego.
Hayes, B. K., Freeze, H. H. and Varki, A. (1993). Biosynthesis ofoligosaccharides in intact Golgi preparations from rat liver. Analysis of N-linked glycans labeled by UDP-[6-3H]N-acetylglucosamine. J. Biol. Chem.268, 16139-16154.
Jakeman, L. B., Winer, J., Bennett, G. L., Altar, C. A. and Ferrara, N.(1992). Binding sites for vascular endothelial growth factor are localized onendothelial cells in adult rat tissues. J. Clin. Invest. 89, 244-253.
Joris, I., Cuenoud, H. F., Doern, G. V., Underwood, J. M. and Majno, G.(1990). Capillary leakage in inflammation. Am. J. Path. 137, 1353-1363.
Koch, A. E., Harlow, L. A., Haines, G. K., et al. (1994). Vascular endothelialgrowth factor. A cytokine modulating endothelial function in rheumatoidarthritis. J. Immunol. 152, 4149-4156.
Kohn, S., Nagy, J. A., Dvorak, H. F. and Dvorak, A. M. (1992). Pathways ofmacromolecular tracer transport across venules and small veins. Lab. Invest.67, 596-607.
Leung, D. W., Cachienes, G., Kuang, W.-J., Goeddel, D. V. and Ferrara, N.(1989). Vascular endothelial growth factor is a secreted angiogenic mitogen.Science 246, 1306-1309.
Majno, G. and Palade, G. E. (1961). Studies on inflammation 1. The effect ofhistamine and serotonin on vascular permeability: An electron microscopicstudy. J. Biochem. Biophys. Cytol. 11, 571-605.
Majno, G., Palade, G. E. and Schoefl, G. I. (1961). Studies on inflammationII. The site of action of histamine and serotonin along the vascular tree: Atopographic study. J. Biophys. Biochem. Cytol. 11, 607-626.
McEver, R. P., Beckstead, J. H., Moore, K. L., Marshall-Carlsen, L. andBainton, D. F. (1989). GMP-140, a platelet α granule membrane protein, isalso synthesized by vascular endothelial cells and is localized in Weibel-Palade bodies. J. Clin. Invest. 84, 92-99.
McKinney, R. V., Singh, B. B. and Brewer, P. D. (1977). Fenestrations inregenerating skeletal muscle capillaries. Am. J. Anat. 150, 213-218.
Miles, A. A. and Miles, E. M. (1952). Vascular reactions to histamine,histamine-liberator and leukotaxine in the skin of guinea-pigs. J. Physiol.118, 228-257.
Millauer, B., Wizigmann-Voos, S., Schnurch, H., et al. (1993). High affinityVEGF binding and developmental expression suggest Flk-1 as a majorregulator of vasculogenesis and angiogenesis. Cell 72, 835-846.
Miller, J. W., Adamis, A. P., Shima, D. T., et al. (1994). Vascular endothelialgrowth factor/Vascular permeability factor is temporally and spatiallycorrelated with ocular angiogenesis in a primate model. Am. J. Path. 145,574-584.
Monacci, W. T., Merrill, M. J. and Oldfield, E. H. (1993). Expression ofvascular permeability factor/vascular endothelial growth factor in normal rattissues. Am. J. Physiol. 264, C995-C1002.
Muller, W. A., Ratti, C. M., McDonnell, S. L. and Cohn, Z. A. (1989). Ahuman endothelial cell-restricted externally disposed plasmalemmal proteinenriched in intercellular junctions. J. Exp. Med. 170, 399-414.
Peters, K. G., De Vries, C. and Williams, L. T. (1993). Vascular endothelialgrowth factor receptor expression during embryogenesis and tissue repairsuggests a role in endothelial differentiation and blood vessel growth. Proc.Nat. Acad. Sci. 90, 8915-8919.
Plate, K. H., Breier, G., Weich, H. A. and Risau, W. (1992). Vascularendothelial growth factor is a potential tumour angiogenesis factor in humangliomas in vivo. Nature 359, 845-848.
Plate, K. H., Breier, G., Millauer, B., Ullrich, A. and Risau, W. (1993). Up-regulation of vascular endothelial growth factor and its cognate receptors in arat glioma model of tumor angiogenesis. Cancer Res. 53, 5822-5827.
Quinn, T. P., Peters, K. G., DeVries, C., Ferrara, N. and Williams, L. T.(1993). Fetal liver kinase 1 is a receptor for vascular endothelial growthfactor and is selectively expressed in vascular endothelium. Proc. Nat. Acad.Sci. 90, 7533-7537.
Roberts, W. G. and Hasan, T. (1993). Tumor-secreted vascular endothelialgrowth factor/vascular permeability factor influences photosensitizer uptake.Cancer Res. 53, 153-157.
Roberts, W. G. and Palade, G. E. (1995). Increased capillary permeability andendothelial fenestration induced by acute administration of vascularendothelial growth factor. Endothelium 2 (supplement), s20.
Schimmenti, L. A., Yan, H.-C., Madri, J. A. and Albelda, S. M. (1992).Platelet endothelial cell adhesion molecule, PECAM-1, modulates cellmigration. J. Cell. Phys. 153, 417-428.

2379VEGF induces endothelial fenestrae
Schoefl, G. I. (1963). Studies on inflammation III. Growing capillaries: Theirstructure and permeability. Virchows Arch. path. Anat. 337, 97-141.
Senger, D. R., Galli, S. J., Dvorak, A. M., Perruzzi, C. A., Harvey, V. S. andDvorak, H. F. (1983). Tumor cells secrete a vascular permeability factor thatpromotes accumulation of ascites fluid. Science 219, 983-985.
Senger, D. R., Connolly, D. T., Van De Water, L., Feder, J. and Dvorak, H.F. (1990). Purification and NH2-terminal amino acid sequence of guinea pigtumor secreted vascular permeability factor. Cancer Res. 50, 1774-1778.
Senger, D. R., Van De Water, L., Brown, L. F., et al. (1993). Vascularpermeability factor (VPF, VEGF) in tumor biology. Cancer Metast. Rev. 12,303-324.
Simionescu, M., Simionescu, N., Silbert, J. E. and Palade, G. E. (1981a).Differentiated microdomains on the luminal surface of the capillaryendothelium II. Partial characterization of their anionic sites. J. Cell Biol. 90,614-621.
Simionescu, N., Siomionescu, M. and Palade, G. E. (1981b). Differentiatedmicrodomains on the luminal surface of the capillary endothelium I.Preferential distribution of anionic sites. J. Cell Biol. 90, 605-613.
Tischer, E., Mitchell, R., Hartman, T., et al. (1991). The human gene forvascular endothelial growth factor. Multiple protein forms are encodedthrough alternative exon splicing. J. Biol. Chem. 266, 11947-11954.
Yamaguchi, T. P., Dumont, D. J., Conlon, R. A., Breitman, M. L. andRossant, J. (1993). flk-1, an flt-related receptor tyrosine kinase is an earlymarker for endothelial cell precursors. Development 118, 489-498.
Yeo, K.-T., Wang, H. H., Nagy, J. A., et al. (1993). Vascular permeabilityfactor (Vascular endothelial growth factor) in guinea pig and human tumorand inflammatory effusions. Cancer Res. 53, 2912-2918.
(Received 30 December 1994 - Accepted 9 March 1995)





























