Embed Size (px)
Citation preview

Increased Thermostability of Thylakoid Membranesin Isoprene-Emitting Leaves Probed with ThreeBiophysical Techniques1[W][OA]
Violeta Velikova, Zsuzsanna Varkonyi, Milan Szabo, Liliana Maslenkova, Isabel Nogues, Laszlo Kovacs,Violeta Peeva, Mira Busheva, Gyozo Garab, Thomas D. Sharkey, and Francesco Loreto*
Institute of Plant Physiology and Genetics (V.V., L.M., V.P.) and Institute of Biophysics and BiomedicalEngineering (M.B.), Bulgarian Academy of Sciences, 1113 Sofia, Bulgaria; Institute of Plant Biology, BiologicalResearch Center, Hungarian Academy of Sciences, 6726 Szeged, Hungary (Z.V., M.S., L.K., G.G.); Institute ofAgroenvironmental and Forest Biology, National Research Council, 00015 Monterotondo, Rome, Italy (I.N.);Department of Biochemistry and Molecular Biology, Michigan State University, East Lansing, Michigan 48824(T.D.S.); and Institute for Plant Protection, National Research Council, 50019 Sesto Fiorentino, Florence, Italy(F.L.)
Three biophysical approaches were used to get insight into increased thermostability of thylakoid membranes in isopre-ne-emittingplants.Arabidopsis (Arabidopsis thaliana) plants genetically modified to make isoprene and Platanus orientalis leaves,in which isoprene emission was chemically inhibited, were used. First, in the circular dichroism spectrum the transitiontemperature of the main band at 694 nm was higher in the presence of isoprene, indicating that the heat stability of chiralmacrodomains of chloroplast membranes, and specifically the stability of ordered arrays of light-harvesting complex II-photosystem II in the stacked region of the thylakoid grana, was improved in the presence of isoprene. Second, the decay ofelectrochromic absorbance changes resulting from the electric field component of the proton motive force (DA515) wasevaluated following single-turnover saturating flashes. The decay of DA515 was faster in the absence of isoprene when leaves ofArabidopsis and Platanus were exposed to high temperature, indicating that isoprene protects the thylakoid membranesagainst leakiness at elevated temperature. Finally, thermoluminescence measurements revealed that S2QB
2 charge recombi-nation was shifted to higher temperature in Arabidopsis and Platanus plants in the presence of isoprene, indicating higheractivation energy for S2QB
2 redox pair, which enables isoprene-emitting plants to perform efficient primary photochemistry ofphotosystem II even at higher temperatures. The data provide biophysical evidence that isoprene improves the integrity andfunctionality of the thylakoid membranes at high temperature. These results contribute to our understanding of isoprenemechanism of action in plant protection against environmental stresses.
Vegetation is a source of large quantities of biogenicvolatile organic compounds emitted into the atmo-sphere, of which isoprene is the most abundant(Guenther et al., 2006). Owing to the high reactivityand fast oxidation by hydroxyl radicals in the atmo-sphere, isoprene can increase the formation of atmo-
spheric ozone (Monson and Holland, 2001; Kleindienstet al., 2007), organic nitrates (O’Brien et al., 1995), andperoxyacylnitrates (Sun and Huang, 1995). Isoprene isalso a significant factor in secondary aerosol forma-tion, with direct and indirect effects on the global ra-diation balance of the atmosphere (Claeys et al., 2004;Matsunaga et al., 2005; Kroll et al., 2006; Ervens et al.,2008; Paulot et al., 2009).
Besides having significant influences on atmo-spheric chemistry, many biogenic volatile organiccompounds play an important role in plant biology.Experimental evidence shows that isoprene protectsphotosynthesis under thermal and oxidative stressconditions (for review, see Vickers et al., 2009a; Loretoand Schnitzler, 2010). It was demonstrated that leavesin which isoprene biosynthesis was blocked by themethyl erythritol pathway inhibitor fosmidomycin,were more sensitive to high temperature and ozoneexposure, and developed stronger oxidative damage,compared to isoprene-emitting leaves (Loreto andVelikova, 2001; Sharkey et al., 2001; Velikova andLoreto, 2005). Plants fumigated with isoprene suffer
1 This work was supported by bilateral projects in the frameworkof Bulgarian Academy of Sciences, Italian National Research Coun-cil, and Hungarian Academy of Sciences agreements, by the Euro-pean Science Foundation-Eurocores project EuroVOL-MOMEVIP(to F.L.), by the grant OTKA/NKTH-CNK (grant no. 80345 to G.G.),and by the National Science Foundation (grant no. IOS–0950574 toT.D.S.).
* Corresponding author; e-mail [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Francesco Loreto ([email protected]).
[W] The online version of this article contains Web-only data.[OA] Open Access articles can be viewed online without a sub-
scription.www.plantphysiol.org/cgi/doi/10.1104/pp.111.182519
Plant Physiology�, October 2011, Vol. 157, pp. 905–916, www.plantphysiol.org � 2011 American Society of Plant Biologists. All Rights Reserved. 905 www.plantphysiol.orgon May 19, 2020 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.

less damage when exposed to oxidative stresses (Loretoet al., 2001) and recover more rapidly from heat stressthan untreated controls (Singsaas et al., 1997; Sharkeyet al., 2001; Velikova et al., 2006). Genetic approaches todevelop isoprene-emitting species from nonemittingwild types (Sharkey et al., 2005; Loivamaki et al.,2007; Sasaki et al., 2007; Vickers et al., 2009b) or tosuppress isoprene synthesis in strong emitters (Behnkeet al., 2007) have also been used to clarify the role ofisoprene in plant protection. Studies with geneticallymodified plants mostly confirmed improved toleranceassociated with the capacity to form and emit iso-prene. Recently important compensatory responses,such as the activation of alternative defensive bio-chemical pathways, e.g. phenolics biosynthesis, to copewith stressful conditions, were also highlighted (Fareset al., 2010), especially when isoprene biosynthesis isknocked out (Behnke et al., 2009).
Several hypotheses have been put forward toexplain the physiological mechanism(s) by which iso-prene protects the photosynthetic apparatus (Loretoand Schnitzler, 2010). The oldest and most widelyaccepted idea is that isoprene stabilizes chloroplastmembranes (Sharkey and Singsaas, 1995). Thylakoidmembranes become leaky at high temperature (Pastenesand Horton, 1996; Bukhov et al., 1999; Schrader et al.,2004; Zhang et al., 2009). It was suggested that thepositive effect of isoprene might be due to the hydro-phobic nature of the molecule, the localization ofisoprene synthase enzyme near the thylakoid mem-branes (Wildermuth and Fall, 1998; Schnitzler et al.,2005), and the high octanol/water partitioning coeffi-cient (Copolovici and Niinemets, 2005). Lipophilicisoprene partitioned into membranes might preventthe formation of water channels responsible for themembrane leakiness at high temperature, or may pre-vent the formation of nonlamellar aggregates, or helpstabilize the photosynthetic complexes embedded inthylakoid membranes (Singsaas et al., 1997; Sharkeyet al., 2001). Isoprene could also enhance hydrophobicinteractions within thylakoids and thereby stabilizeinteractions between lipids and/or membrane proteinsduring episodes of heat shock or high-temperaturestress conditions (Sharkey and Yeh, 2001). Based onmolecular dynamics simulations of phospholipid bi-layers with and without isoprene, Siwko et al. (2007)suggested that isoprene enhances the packing of lipidtails. The authors suggested that the role of isopreneas a membrane stabilizer can be related to the factthat it fits well into the available pockets of the freevolume inside the membrane, and adds cohesiveness,amplifying membrane packing, while not affectingthe dynamics of phospholipid bilayers (Siwko et al.,2007). However, Logan and Monson (1999) workingwith reconstituted liposomes were not able to provethat isoprene improves the thermal stability of mem-branes.
Another hypothesis to explain the generally positiverole of isoprene in plant metabolism is based on thehigh reactivity of volatile isoprenoids with radicals
and other reactive compounds. Loreto et al. (2001) firstsuggested that isoprene might operate as a volatilemolecule, scavenging reactive oxygen species in theintercellular spaces of the leaf mesophyll. More re-cently, it was proven that isoprene also removes reac-tive nitrogen species from the mesophyll (Velikovaet al., 2008). Vickers et al. (2009a) reviewed the possiblemechanisms of isoprene function and suggested thatthe molecule may have a general antioxidant role.
A demonstration of effects of isoprene on biophys-ical measurements of thylakoid function at high tem-perature could help in working toward a resolution ofthe primary mechanism of isoprene action. Here threewell-known techniques in biophysical studies of thy-lakoid membrane function were tested for the effect ofisoprene at high temperature. Two plant systems wereused. Arabidopsis (Arabidopsis thaliana), which doesnot normally make isoprene, was engineered with anisoprene synthase gene from kudzu (Pueraria lobata) sothat wild-type plants (nonemitting) could be com-pared to the transformed plants that do make iso-prene. Leaves of Platanus (plane tree) normally domake isoprene but this was inhibited by fosmido-mycin so that emitting (water-fed) and nonemitting(fosmidomycin-fed) leaves could be compared. Thethermal stability of the thylakoid membranes was char-acterized with biophysical approaches not previouslyused in isoprene studies, namely circular dichroism(CD) spectrosocopy, electrochromic absorbance tran-sients (DA515), and thermoluminescence (TL). Thesemeasurements revealed that in the presence of iso-prene, the macroorganization of the pigment-proteincomplexes in the membranes were more stable toelevated temperature, the membranes were better ableto maintain a light-induced transmembrane electricfield at elevated temperatures, and the recombinationof the PSII donor and acceptor side charges occurred upto higher temperature.
RESULTS
Suitability of Plant Material
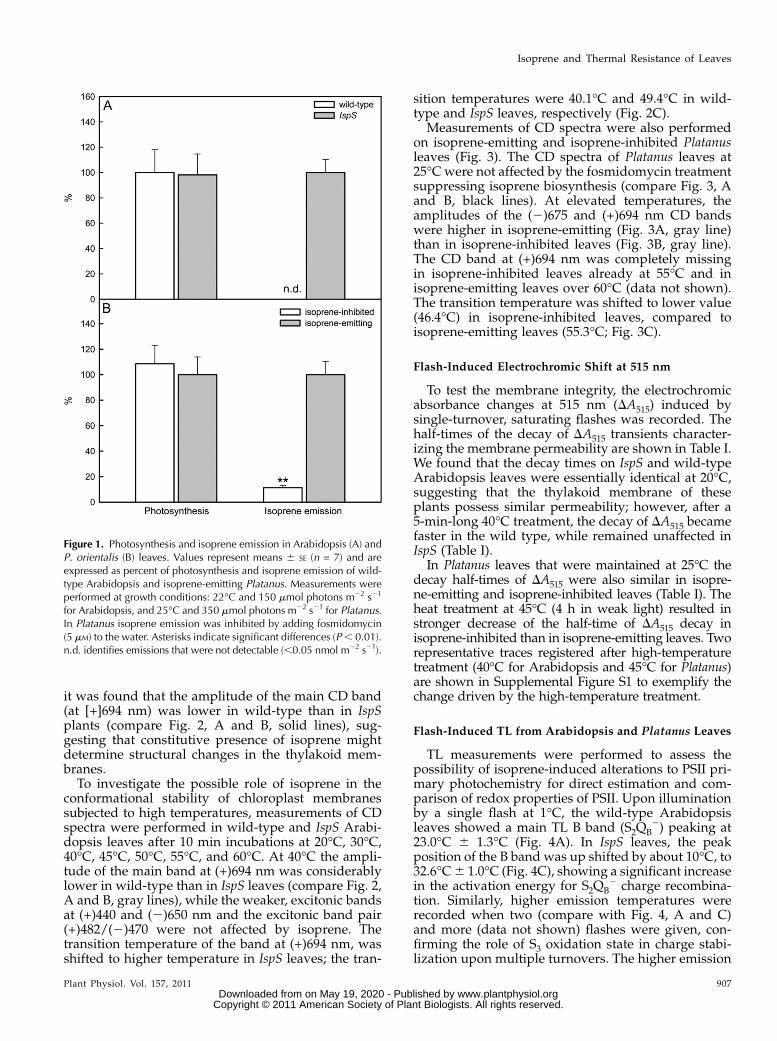
Genetic manipulation of Arabidopsis plants didnot affect their photosynthetic performance. IspS andwild-type plants were characterized by similar photo-synthesis (Fig. 1; absolute values 7.0 6 0.3 mmolm22 s21, n = 14). Inhibition of isoprene emission inPlatanus leaves by the chemical inhibitor also did notinfluence photosynthesis. Absolute values of photo-synthesis in Platanus leaves were 5.3 6 0.4 mmolm22 s21, n = 14. Isoprene emission was not detect-able in wild-type plants of Arabidopsis, whereas infosmidomycin-fed Platanus leaves isoprene emissionwas inhibited to about 10% of the original level (Fig. 1).
CD Spectroscopy
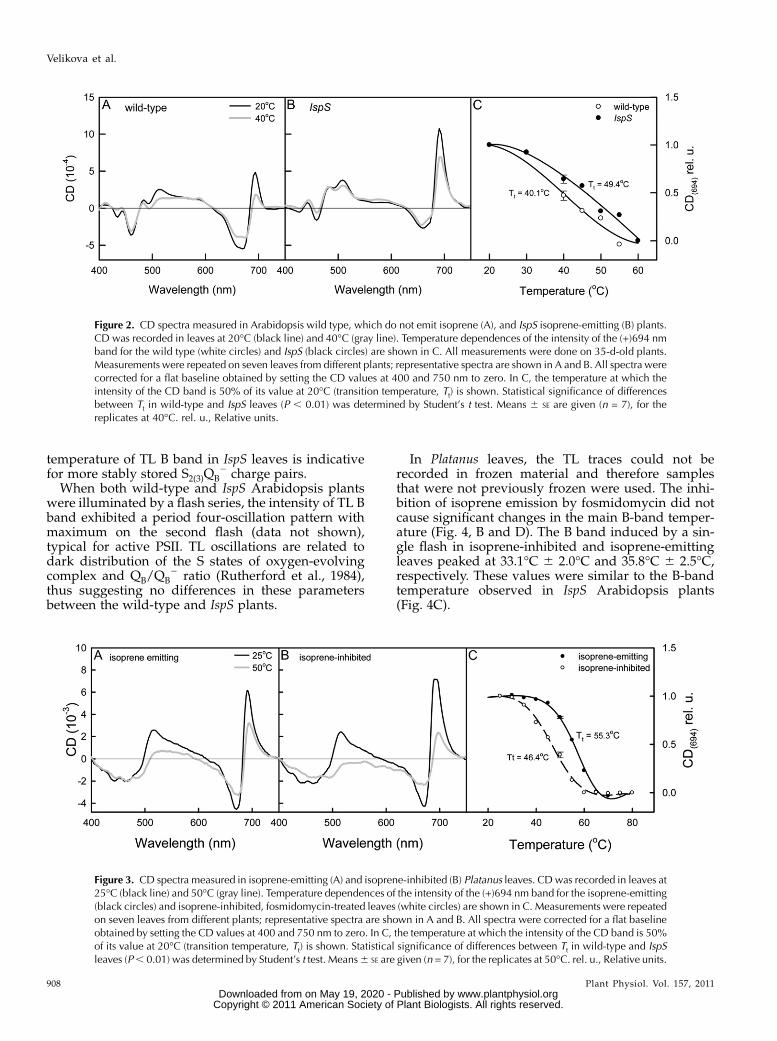
CD spectra in wild-type and IspSArabidopsis plantsshowed considerable differences at 20�C. In particular,
Velikova et al.
906 Plant Physiol. Vol. 157, 2011 www.plantphysiol.orgon May 19, 2020 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.
it was found that the amplitude of the main CD band(at [+]694 nm) was lower in wild-type than in IspSplants (compare Fig. 2, A and B, solid lines), sug-gesting that constitutive presence of isoprene mightdetermine structural changes in the thylakoid mem-branes.To investigate the possible role of isoprene in the
conformational stability of chloroplast membranessubjected to high temperatures, measurements of CDspectra were performed in wild-type and IspS Arabi-dopsis leaves after 10 min incubations at 20�C, 30�C,40�C, 45�C, 50�C, 55�C, and 60�C. At 40�C the ampli-tude of the main band at (+)694 nm was considerablylower in wild-type than in IspS leaves (compare Fig. 2,A and B, gray lines), while the weaker, excitonic bandsat (+)440 and (2)650 nm and the excitonic band pair(+)482/(2)470 were not affected by isoprene. Thetransition temperature of the band at (+)694 nm, wasshifted to higher temperature in IspS leaves; the tran-
sition temperatures were 40.1�C and 49.4�C in wild-type and IspS leaves, respectively (Fig. 2C).
Measurements of CD spectra were also performedon isoprene-emitting and isoprene-inhibited Platanusleaves (Fig. 3). The CD spectra of Platanus leaves at25�Cwere not affected by the fosmidomycin treatmentsuppressing isoprene biosynthesis (compare Fig. 3, Aand B, black lines). At elevated temperatures, theamplitudes of the (2)675 and (+)694 nm CD bandswere higher in isoprene-emitting (Fig. 3A, gray line)than in isoprene-inhibited leaves (Fig. 3B, gray line).The CD band at (+)694 nm was completely missingin isoprene-inhibited leaves already at 55�C and inisoprene-emitting leaves over 60�C (data not shown).The transition temperature was shifted to lower value(46.4�C) in isoprene-inhibited leaves, compared toisoprene-emitting leaves (55.3�C; Fig. 3C).
Flash-Induced Electrochromic Shift at 515 nm
To test the membrane integrity, the electrochromicabsorbance changes at 515 nm (DA515) induced bysingle-turnover, saturating flashes was recorded. Thehalf-times of the decay of DA515 transients character-izing the membrane permeability are shown in Table I.We found that the decay times on IspS and wild-typeArabidopsis leaves were essentially identical at 20�C,suggesting that the thylakoid membrane of theseplants possess similar permeability; however, after a5-min-long 40�C treatment, the decay of DA515 becamefaster in the wild type, while remained unaffected inIspS (Table I).
In Platanus leaves that were maintained at 25�C thedecay half-times of DA515 were also similar in isopre-ne-emitting and isoprene-inhibited leaves (Table I). Theheat treatment at 45�C (4 h in weak light) resulted instronger decrease of the half-time of DA515 decay inisoprene-inhibited than in isoprene-emitting leaves. Tworepresentative traces registered after high-temperaturetreatment (40�C for Arabidopsis and 45�C for Platanus)are shown in Supplemental Figure S1 to exemplify thechange driven by the high-temperature treatment.
Flash-Induced TL from Arabidopsis and Platanus Leaves
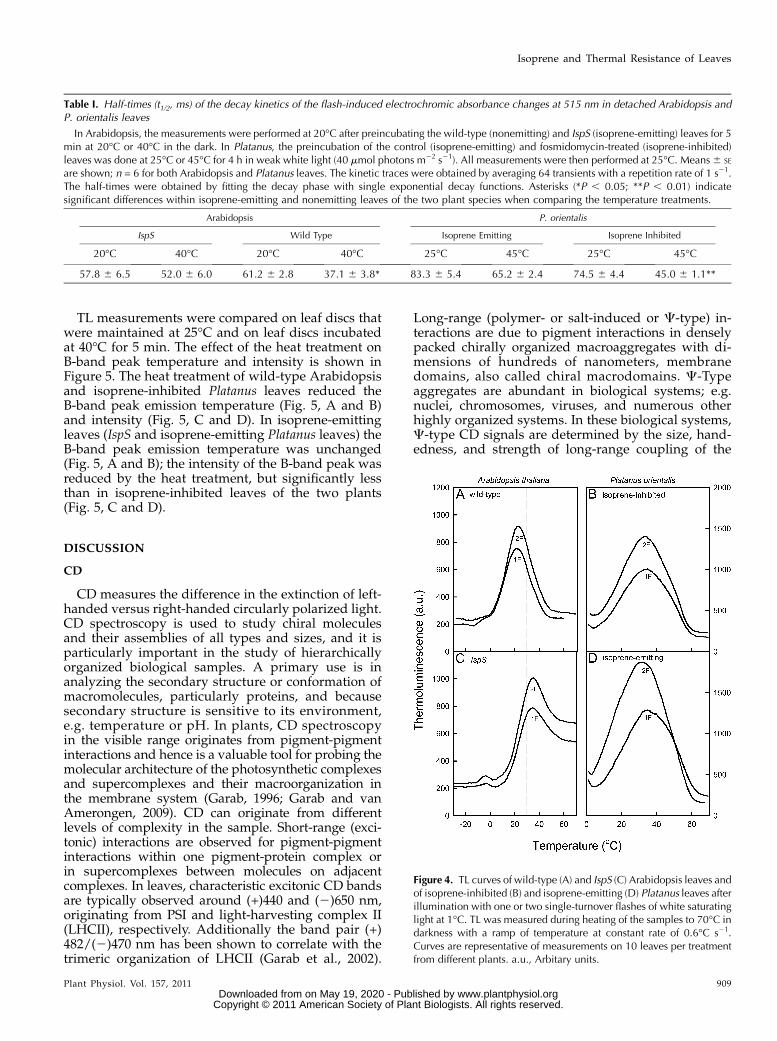
TL measurements were performed to assess thepossibility of isoprene-induced alterations to PSII pri-mary photochemistry for direct estimation and com-parison of redox properties of PSII. Upon illuminationby a single flash at 1�C, the wild-type Arabidopsisleaves showed a main TL B band (S2QB
2) peaking at23.0�C 6 1.3�C (Fig. 4A). In IspS leaves, the peakposition of the B band was up shifted by about 10�C, to32.6�C6 1.0�C (Fig. 4C), showing a significant increasein the activation energy for S2QB
2 charge recombina-tion. Similarly, higher emission temperatures wererecorded when two (compare with Fig. 4, A and C)and more (data not shown) flashes were given, con-firming the role of S3 oxidation state in charge stabi-lization upon multiple turnovers. The higher emission
Figure 1. Photosynthesis and isoprene emission in Arabidopsis (A) andP. orientalis (B) leaves. Values represent means 6 SE (n = 7) and areexpressed as percent of photosynthesis and isoprene emission of wild-type Arabidopsis and isoprene-emitting Platanus. Measurements wereperformed at growth conditions: 22�C and 150 mmol photons m22 s21
for Arabidopsis, and 25�C and 350 mmol photons m22 s21 for Platanus.In Platanus isoprene emission was inhibited by adding fosmidomycin(5 mM) to the water. Asterisks indicate significant differences (P, 0.01).n.d. identifies emissions that were not detectable (,0.05 nmol m22 s21).
Isoprene and Thermal Resistance of Leaves
Plant Physiol. Vol. 157, 2011 907 www.plantphysiol.orgon May 19, 2020 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.
temperature of TL B band in IspS leaves is indicativefor more stably stored S2(3)QB
2 charge pairs.When both wild-type and IspS Arabidopsis plants
were illuminated by a flash series, the intensity of TL Bband exhibited a period four-oscillation pattern withmaximum on the second flash (data not shown),typical for active PSII. TL oscillations are related todark distribution of the S states of oxygen-evolvingcomplex and QB/QB
2 ratio (Rutherford et al., 1984),thus suggesting no differences in these parametersbetween the wild-type and IspS plants.
In Platanus leaves, the TL traces could not berecorded in frozen material and therefore samplesthat were not previously frozen were used. The inhi-bition of isoprene emission by fosmidomycin did notcause significant changes in the main B-band temper-ature (Fig. 4, B and D). The B band induced by a sin-gle flash in isoprene-inhibited and isoprene-emittingleaves peaked at 33.1�C 6 2.0�C and 35.8�C 6 2.5�C,respectively. These values were similar to the B-bandtemperature observed in IspS Arabidopsis plants(Fig. 4C).
Figure 2. CD spectra measured in Arabidopsis wild type, which do not emit isoprene (A), and IspS isoprene-emitting (B) plants.CD was recorded in leaves at 20�C (black line) and 40�C (gray line). Temperature dependences of the intensity of the (+)694 nmband for the wild type (white circles) and IspS (black circles) are shown in C. All measurements were done on 35-d-old plants.Measurements were repeated on seven leaves from different plants; representative spectra are shown in A and B. All spectra werecorrected for a flat baseline obtained by setting the CD values at 400 and 750 nm to zero. In C, the temperature at which theintensity of the CD band is 50% of its value at 20�C (transition temperature, Tt) is shown. Statistical significance of differencesbetween Tt in wild-type and IspS leaves (P , 0.01) was determined by Student’s t test. Means 6 SE are given (n = 7), for thereplicates at 40�C. rel. u., Relative units.
Figure 3. CD spectra measured in isoprene-emitting (A) and isoprene-inhibited (B) Platanus leaves. CDwas recorded in leaves at25�C (black line) and 50�C (gray line). Temperature dependences of the intensity of the (+)694 nm band for the isoprene-emitting(black circles) and isoprene-inhibited, fosmidomycin-treated leaves (white circles) are shown in C. Measurements were repeatedon seven leaves from different plants; representative spectra are shown in A and B. All spectra were corrected for a flat baselineobtained by setting the CD values at 400 and 750 nm to zero. In C, the temperature at which the intensity of the CD band is 50%of its value at 20�C (transition temperature, Tt) is shown. Statistical significance of differences between Tt in wild-type and IspSleaves (P, 0.01) was determined by Student’s t test. Means6 SE are given (n = 7), for the replicates at 50�C. rel. u., Relative units.
Velikova et al.
908 Plant Physiol. Vol. 157, 2011 www.plantphysiol.orgon May 19, 2020 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.
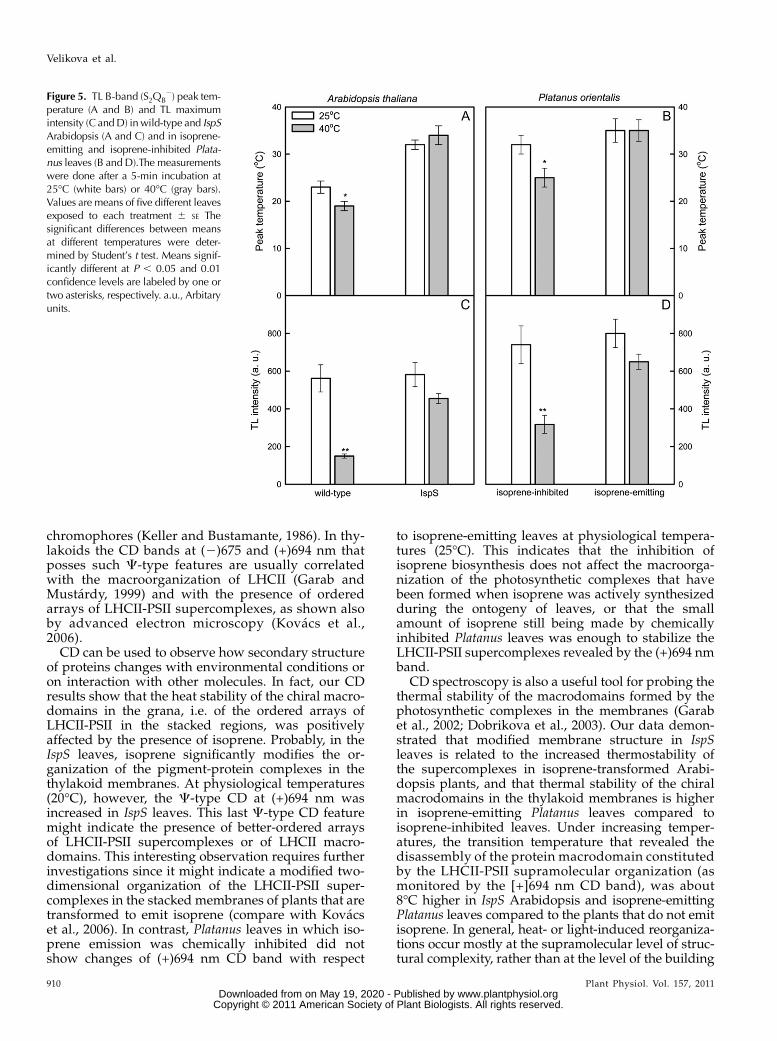
TL measurements were compared on leaf discs thatwere maintained at 25�C and on leaf discs incubatedat 40�C for 5 min. The effect of the heat treatment onB-band peak temperature and intensity is shown inFigure 5. The heat treatment of wild-type Arabidopsisand isoprene-inhibited Platanus leaves reduced theB-band peak emission temperature (Fig. 5, A and B)and intensity (Fig. 5, C and D). In isoprene-emittingleaves (IspS and isoprene-emitting Platanus leaves) theB-band peak emission temperature was unchanged(Fig. 5, A and B); the intensity of the B-band peak wasreduced by the heat treatment, but significantly lessthan in isoprene-inhibited leaves of the two plants(Fig. 5, C and D).
DISCUSSION
CD
CD measures the difference in the extinction of left-handed versus right-handed circularly polarized light.CD spectroscopy is used to study chiral moleculesand their assemblies of all types and sizes, and it isparticularly important in the study of hierarchicallyorganized biological samples. A primary use is inanalyzing the secondary structure or conformation ofmacromolecules, particularly proteins, and becausesecondary structure is sensitive to its environment,e.g. temperature or pH. In plants, CD spectroscopyin the visible range originates from pigment-pigmentinteractions and hence is a valuable tool for probing themolecular architecture of the photosynthetic complexesand supercomplexes and their macroorganization inthe membrane system (Garab, 1996; Garab and vanAmerongen, 2009). CD can originate from differentlevels of complexity in the sample. Short-range (exci-tonic) interactions are observed for pigment-pigmentinteractions within one pigment-protein complex orin supercomplexes between molecules on adjacentcomplexes. In leaves, characteristic excitonic CD bandsare typically observed around (+)440 and (2)650 nm,originating from PSI and light-harvesting complex II(LHCII), respectively. Additionally the band pair (+)482/(2)470 nm has been shown to correlate with thetrimeric organization of LHCII (Garab et al., 2002).
Long-range (polymer- or salt-induced or C-type) in-teractions are due to pigment interactions in denselypacked chirally organized macroaggregates with di-mensions of hundreds of nanometers, membranedomains, also called chiral macrodomains. C-Typeaggregates are abundant in biological systems; e.g.nuclei, chromosomes, viruses, and numerous otherhighly organized systems. In these biological systems,C-type CD signals are determined by the size, hand-edness, and strength of long-range coupling of the
Table I. Half-times (t1/2, ms) of the decay kinetics of the flash-induced electrochromic absorbance changes at 515 nm in detached Arabidopsis andP. orientalis leaves
In Arabidopsis, the measurements were performed at 20�C after preincubating the wild-type (nonemitting) and IspS (isoprene-emitting) leaves for 5min at 20�C or 40�C in the dark. In Platanus, the preincubation of the control (isoprene-emitting) and fosmidomycin-treated (isoprene-inhibited)leaves was done at 25�C or 45�C for 4 h in weak white light (40 mmol photons m22 s21). All measurements were then performed at 25�C. Means6 SE
are shown; n = 6 for both Arabidopsis and Platanus leaves. The kinetic traces were obtained by averaging 64 transients with a repetition rate of 1 s21.The half-times were obtained by fitting the decay phase with single exponential decay functions. Asterisks (*P , 0.05; **P , 0.01) indicatesignificant differences within isoprene-emitting and nonemitting leaves of the two plant species when comparing the temperature treatments.
Arabidopsis P. orientalis
IspS Wild Type Isoprene Emitting Isoprene Inhibited
20�C 40�C 20�C 40�C 25�C 45�C 25�C 45�C
57.8 6 6.5 52.0 6 6.0 61.2 6 2.8 37.1 6 3.8* 83.3 6 5.4 65.2 6 2.4 74.5 6 4.4 45.0 6 1.1**
Figure 4. TL curves of wild-type (A) and IspS (C) Arabidopsis leaves andof isoprene-inhibited (B) and isoprene-emitting (D) Platanus leaves afterillumination with one or two single-turnover flashes of white saturatinglight at 1�C. TL was measured during heating of the samples to 70�C indarkness with a ramp of temperature at constant rate of 0.6�C s21.Curves are representative of measurements on 10 leaves per treatmentfrom different plants. a.u., Arbitary units.
Isoprene and Thermal Resistance of Leaves
Plant Physiol. Vol. 157, 2011 909 www.plantphysiol.orgon May 19, 2020 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.
chromophores (Keller and Bustamante, 1986). In thy-lakoids the CD bands at (2)675 and (+)694 nm thatposses such C-type features are usually correlatedwith the macroorganization of LHCII (Garab andMustardy, 1999) and with the presence of orderedarrays of LHCII-PSII supercomplexes, as shown alsoby advanced electron microscopy (Kovacs et al.,2006).
CD can be used to observe how secondary structureof proteins changes with environmental conditions oron interaction with other molecules. In fact, our CDresults show that the heat stability of the chiral macro-domains in the grana, i.e. of the ordered arrays ofLHCII-PSII in the stacked regions, was positivelyaffected by the presence of isoprene. Probably, in theIspS leaves, isoprene significantly modifies the or-ganization of the pigment-protein complexes in thethylakoid membranes. At physiological temperatures(20�C), however, the C-type CD at (+)694 nm wasincreased in IspS leaves. This last C-type CD featuremight indicate the presence of better-ordered arraysof LHCII-PSII supercomplexes or of LHCII macro-domains. This interesting observation requires furtherinvestigations since it might indicate a modified two-dimensional organization of the LHCII-PSII super-complexes in the stacked membranes of plants that aretransformed to emit isoprene (compare with Kovacset al., 2006). In contrast, Platanus leaves in which iso-prene emission was chemically inhibited did notshow changes of (+)694 nm CD band with respect
to isoprene-emitting leaves at physiological tempera-tures (25�C). This indicates that the inhibition ofisoprene biosynthesis does not affect the macroorga-nization of the photosynthetic complexes that havebeen formed when isoprene was actively synthesizedduring the ontogeny of leaves, or that the smallamount of isoprene still being made by chemicallyinhibited Platanus leaves was enough to stabilize theLHCII-PSII supercomplexes revealed by the (+)694 nmband.
CD spectroscopy is also a useful tool for probing thethermal stability of the macrodomains formed by thephotosynthetic complexes in the membranes (Garabet al., 2002; Dobrikova et al., 2003). Our data demon-strated that modified membrane structure in IspSleaves is related to the increased thermostability ofthe supercomplexes in isoprene-transformed Arabi-dopsis plants, and that thermal stability of the chiralmacrodomains in the thylakoid membranes is higherin isoprene-emitting Platanus leaves compared toisoprene-inhibited leaves. Under increasing temper-atures, the transition temperature that revealed thedisassembly of the protein macrodomain constitutedby the LHCII-PSII supramolecular organization (asmonitored by the [+]694 nm CD band), was about8�C higher in IspS Arabidopsis and isoprene-emittingPlatanus leaves compared to the plants that do not emitisoprene. In general, heat- or light-induced reorganiza-tions occur mostly at the supramolecular level of struc-tural complexity, rather than at the level of the building
Figure 5. TL B-band (S2QB2) peak tem-
perature (A and B) and TL maximumintensity (C andD) inwild-type and IspSArabidopsis (A and C) and in isoprene-emitting and isoprene-inhibited Plata-nus leaves (B and D).The measurementswere done after a 5-min incubation at25�C (white bars) or 40�C (gray bars).Values are means of five different leavesexposed to each treatment 6 SE Thesignificant differences between meansat different temperatures were deter-mined by Student’s t test. Means signif-icantly different at P , 0.05 and 0.01confidence levels are labeled by one ortwo asterisks, respectively. a.u., Arbitaryunits.
Velikova et al.
910 Plant Physiol. Vol. 157, 2011 www.plantphysiol.orgon May 19, 2020 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.

blocks of macroassemblies. For example, the stability ofthe single constituents of pigment-protein complexeshas been shown to be considerably higher than thestability of their macroassemblies (Cseh et al., 2000,2005; Dobrikova et al., 2003). The experiment using CDspectroscopy therefore confirms that a higher thermalstability is conferred by isoprene to thylakoid mem-branes, and suggests that this might be due to a sus-tained stability of the ordered arrays of supercomplexesin the thylakoid membranes, rather than to a specifi-cally induced resistance in a single photosystem com-ponent.It was shown that the structural flexibility of LHCII
macrodomains depends largely on the lipid environ-ment of the complexes (Simidjiev et al., 1998). Thepositive stabilizing effect of isoprene on membranescould therefore be related to changes in the lipid phaseand/or alterations induced by isoprene in the phasebehavior of lipids. The mixed lipid phases can havesignificant effects on the organization of thylakoidmembranes (Williams and Quinn, 1987). Lateral or-ganization of membranes into domains can dependcritically on temperature and small molecules havebeen found to significantly affect large-scale phaseseparations. Molecules that affect the miscibility of dif-ferent phases will significantly alter the stability of dif-ferent membrane domains under heat stress (Veatch,2007). As under physiological conditions the CD spec-trum at (+)694 nm is similar in isoprene-emitting andisoprene-inhibited Platanus leaves, we infer that thechemical inhibition of isoprene does not alter the mac-romolecular structure of the membranes. If isoprenesimply alters the lipid phase of membranes, then theprotective effect may be more rapidly lost after iso-prene inhibition. This would explain why the protec-tive effect of isoprene is seen on both Arabidopsis IspSplants and in isoprene-emitting Platanus under heatstress, independently of the stability of the LHCII-PSIIsupramolecular organization and of the way isopreneemission is manipulated. Recent data have shown thatthe dgd1 Arabidopsis mutant, which is deficient in thebilayer lipid digalatosyldiacyl glycerol and enriched inthe nonbilayer monogalactosyldiacyl glycerol, displayconsiderably lower thermal stability of the chiralmacrodomains, measured with the temperature de-pendence of the main C-type CD bands (Krumovaet al., 2010).Inwhole leaves and intact granal chloroplasts, usually
the C-type bands are dominating the spectra and themuch weaker excitonic bands become visible only upondisintegration of the macrodomains, e.g. at hypotoniclow-salt medium (Garab et al., 1991) or at elevatedtemperatures (Cseh et al., 2000). Interestingly, the ca-pacity to emit isoprene does not seem to affect signifi-cantly the excitonic (non-C-type) bands arising fromsupercomplexes as also monitored by CD spectroscopy.Namely, the trimer-to-monomer transition temperaturesof LHCII, between 55�C and 60�C, as tested with theexcitonic band pair (+)482/(2)470 (Garab et al., 2002)was unchanged (data not shown), indicating that the
trimeric LHCII is resistant to heat independently ofisoprene presence, in both tested species.
Electrochromic Shift and Membrane Conductance
Electrochromic absorbance changes at 515 nm(DA515) are used to monitor ion permeability of mem-branes (Junge, 1977; Witt, 1979; Peters et al., 1984)because of the effect on the electrical potential acrossthe thylakoid membranes. In particular, the decaykinetics of the absorbance changes are proportionalto the ion flux across the thylakoid membranes and aresensitive indicators of thylakoid membrane intactness(Peters et al., 1984), i.e. of the ability of membranes tomaintain the light-induced transmembrane electricfield. In dark-adapted leaves the ATP synthase isturned off and so membrane effects are more prom-inent than in light-adapted leaves. The decay is slowerin dark-adapted leaves than light-adapted leaves butis variable among species. Therefore, the absolute de-cay rate cannot be compared among species but thedecay for one species can be compared under differ-ent conditions such as moderate or high temperatureand with or without isoprene present.
Increased permeability has been found to occurparticularly at high temperatures, concurrently withdenaturation of membrane lipids (Bukhov et al., 1999;Schrader et al., 2004; Zhang and Sharkey, 2009; Zhanget al., 2009). The increased membrane permeability ofheat-treated samples can be due to conformationalchanges of membrane lipids, opening of ion channels,or changes in lipid-lipid interactions (Santarius, 1980).At moderate (20�C–25�C) temperature, the thylakoidmembranes exhibit similar electrochromic absorbancechanges at 515 nm (DA515), in spite of the presence (IspSArabidopsis, and control water-fed Platanus leaves)or absence (wild-type Arabidopsis, and fosmidomy-cin-fed Platanus leaves) of isoprene. Thus, the mem-brane permeability to ions is low and is not affectedby isoprene presence at growth temperature. How-ever, at high temperature (40�C–45�C) wild-type Arab-idopsis membranes, and isoprene-inhibited Platanusmembranes became more permeable to ions, whereasisoprene-emitting Platanus membranes were lessaffected, and IspS membranes remained unaffected.Thus, isoprene presence is associatedwith better mem-brane integrity under heat-stress conditions, as shownby lower ion leakage at high temperatures.
TL
TL measurements also support the thermotolerancehypothesis, demonstrating the protective role of iso-prene in increasing the membrane thermostability.TL is a sensitive and reliable tool for monitoring thefunctionality of PSII donor and acceptor side compo-nents (Sane and Rutherford, 1986; Vass and Inoue,1992; Ducruet, 2003; Sane, 2004). Illumination of dark-adapted samples with single-turnover flashes gener-ates charge pairs within PSII reaction centers that are
Isoprene and Thermal Resistance of Leaves
Plant Physiol. Vol. 157, 2011 911 www.plantphysiol.orgon May 19, 2020 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.

energetically stabilized on the primary and secondaryquinone acceptors of the chloroplast’s electron trans-port chain. The samples are rapidly cooled down totrap separated charges that, on heating, yield a TLlight emission at characteristic temperatures (for de-tails, see Sane, 2004). Two major TL bands, B and Q,appear in TL curves as a result of the recombination ofthe trapped electrons and stabilized positive holes onthe reduced quinone acceptors (QA or QB) and on theS2 (or S3) oxidation state of the water-splitting enzyme.The B band is generated by the recombination of QB
2
with S2 state of the water-splitting complex. Evensmall changes in the redox properties of the radicalpairs affect the intensity and the peak position of TLbands. This complexity of information of TL emissioncurves is widely used for detection of structuralchanges in both the donor and acceptor side of PSII.
We tested with TL measurements the impact ofisoprene presence in leaves that were exposed tophysiological or high temperatures before runningthe assay. In our experiment with Arabidopsis plantsexposed to physiological temperatures, the peak posi-tion of the B band was up shifted by about 10�C in IspSleaves, compared to wild-type leaves. Alterations inthe peak positions of a TL band indicates changes inthe stability, redox potential, or activation energy ofthe recombining charge pairs, while the peak ampli-tude is proportional to the concentration of the corre-sponding charge pairs (Rutherford et al., 1984). Theobserved stabilization of the charge separation statebetween the redox components on PSII donor andacceptor side in IspS Arabidopsis clearly indicates anisoprene-induced modification that may enable plantsto perform efficient primary photochemistry of PSII athigher temperatures. In Platanus leaves exposed tophysiological temperatures, however, the B band wasnot shifted to lower temperatures when isoprene waschemically inhibited, as we would have expected. Asin the case of CD spectroscopy, we attribute this resultto the possibility that the chemical inhibition of iso-prene does not interfere with the molecular structureof membrane architecture that was assembled whenisoprene was still available. Specifically, our resultsmight indicate that the isoprene-related membranemodification affecting PSII reaction centers, and gen-erating the shift in the energy levels of the S2(3)QB
2, is along-term, structural effect that cannot be altered byfast removal of isoprene. Alternatively, however, thelow rate of isoprene synthesis in the fosmidomycin-fedPlatanus leaves might be sufficient to stabilize chargeseparation. In any case, a change in TL peak position isnot directly associated with the presence of isopreneunder physiological conditions.
When TL measurements were repeated in samplespreviously exposed to heat stress, the B band clearlypeaked at a lower temperature in plants that do notemit isoprene, whereas the B-band peak position wasnot affected in isoprene-emitting leaves of both Arab-idopsis and Platanus. Absence of isoprene caused astrong reduction of the intensity of the B band in both
plant species that were subjected to the heat treat-ment. It has been shown that the peak temperature ofB band increases in stress-tolerant photosyntheticorganisms; for example in thermophilic cyanobacte-ria or desiccation-tolerant species (Govindjee et al.,1985; Sass et al., 1996; Maslenkova and Homann,2000; Peeva and Maslenkova, 2004). It has also beenshown that stresses do not affect the energetic levelsof the stabilized S2QB
2 charge pairs in stress-tolerantplants (Sass et al., 1996; Georgieva et al., 2005).
It is of interest that the heat stress also affected TLparameters in isoprene-inhibited Platanus leaves thatdid not show TL changes at physiological tempera-tures. However, the TL intensity of the B band inisoprene-inhibited Platanus leaves was less affectedafter heat treatment, than in wild-type Arabidopsisthat constitutively do not emit isoprene. We interpretthis result as showing that a continuous biosynthesisof isoprene is needed to maintain membrane stabilityunder heat stress, whereas the residual isoprene thatis likely stabilizing membranes under physiologicaltemperatures in isoprene-inhibited Platanus (see above)may only partially fulfill this role under elevated tem-peratures.
Since the production of isoprene is strongly stimu-lated at elevated temperatures in the range of 30�C to40�C (Monson et al., 1992; Singsaas and Sharkey, 2000),we suggest that isoprene emission represents an adap-tive mechanism of membrane stabilization under hightemperatures. Isoprene is highly lipophilic so that itmay partition into chloroplast membranes and there-fore it could act within the thylakoid lipid phase byrendering membranes more viscous and changing thefeatures of secondary electron acceptor QB. It wasshown that this plastoquinone molecule is the mostsensitive to the lipid environment (Gombos et al.,2002). However, this does not rule out other volatileisoprenoids (e.g. monoterpenes, sesquiterpenes), zea-xanthin synthesis, accumulation of soluble thermo-protectants such as sugars or free amino acids, orchanges in membrane lipid composition involvementin membrane stabilization (Ducruet et al., 2007).
CONCLUSION
There currently is a controversy about the mecha-nism of action of isoprene (Loreto and Schnitzler, 2010).Vickers et al. (2009b) suggests that the mechanismcould be interaction/quenching of reactive oxygenspecies that allows plants to tolerate heat and oxidativestress. Sharkey et al. (2008) suggest that isoprene mayaffect membrane organization and function and thatthis allows plants to tolerate heat and reduces theformation of reactive oxygen. The molecular dynamicsmodeling of Siwko et al. (2007) and the octanol/waterpartitioning (Copolovici and Niinemets, 2005) showthat isoprene will accumulate inside membranes butuntil now there were no data showing that this couldaffect biophysical signals of membrane function.
Velikova et al.
912 Plant Physiol. Vol. 157, 2011 www.plantphysiol.orgon May 19, 2020 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.

Three biophysical techniques, which are often usedto study thylakoid membrane function, show thatisoprene can extend thylakoid membrane functionalityto higher temperature while having little or no effect atmoderate temperature. While the three techniquesspecifically investigated different aspects of the pho-tochemical apparatus, they converge in indicating thatisoprene improves the integrity of the photochemistryof photosynthesis under heat-stress conditions. TheCD spectra in particular show that isoprene can affectlarge-scale thylakoid membrane organization while notaffecting the short-range interactions that arise frompigment-protein complexes and their supercomplexes.As more is learned about how heat affects photo-
synthetic function and what mechanisms improvethermotolerance it will be possible to interpret thesebiophysical measures of isoprene effects in terms ofimproved thermotolerance and ultimately in terms ofthe evolutionary pressure that favors isoprene emis-sion from plants in some conditions. These experi-ments show that there are biophysical mechanisms bywhich the integrity and functionality of the thylakoidmembranes are better preserved at high temperaturein the presence of isoprene. This strongly supports theidea that isoprene improves thermotolerance of plants(Sharkey and Singsaas, 1995; Loreto and Schnitzler,2010) and favors effects on membrane function as thereason. This suggests that isoprene-emitting plantswill cope better with temperature extremes that maybecomemore frequent in the near future due to currentclimate change (e.g. De Boeck et al., 2011).
MATERIALS AND METHODS
Plant Material and Growth Conditions
Wild-type Arabidopsis (Arabidopsis thaliana; ecotype Wassilewskija), which
does not naturally emit isoprene (wild type), and genetically modified
Arabidopsis plants (IspS) producing isoprene as a natural metabolite were
used in this study. Arabidopsis IspS plants were transformed inserting the
kudzu (Pueraria lobata) ISPS genomic sequence under the control of a consti-
tutive promoter (Sharkey et al., 2005). The T4 generation was used for the
experiments. Plants were grown in a climate chamber (Percival Scientific)
under a photoperiod of 8 h light at 22�C and 16 h dark at 20�C. Light intensitywas 150 mmol photons m22 s21 and relative humidity was 50% to 60%. Fully
grown rosettes were used for the experiments.
Three-year-old Platanus (Platanus orientalis) plants were grown in 5-L pots
with sand and peat-based commercial soil (1:1) in a greenhouse under con-
trolled environmental factors. In particular the greenhouse was thermostated
with air conditioning modules and the light intensity was supplemented,
if needed, with artificial lamps characterized by a solar spectrum (Osram
PowerStar HQI-TS 150W). Growth temperature was 25�C/22�C (day/night),
daily light intensity was 350 mmol photons m22 s21, with a 12-h photoperiod,
and relative humidity was maintained around 65%. The experiments were
performed on fully expanded leaves.
Plants were regularly watered to pot water capacity and fertilized once a
week with full-strength Hoagland solution.
Fosmidomycin and High-Temperature Treatments of
Platanus Leaves
To inhibit isoprene emission from Platanus leaves, fosmidomycin, a specific
inhibitor of 2-deoxyxylulose 5-phosphate reductoisomerase (Zeidler et al.,
1998), was used. Platanus leaves were cut under water and maintained with
the petiole in a glass vial with 15 mL of distilled water for 1 h. Fosmidomycin
(5 mM) was then added to the water and taken up by the leaf through
transpiration stream. After 1 h of incubation isoprene emission was inhibited
to approximately 10% of the original level, as monitored by gas exchange and
gas chromatography (Loreto and Velikova, 2001). Cut leaves with the petiole
immersed in a vial with distilled water only were used as a control (isoprene-
emitting leaves). Isoprene-emitting and isoprene-inhibited leaves were used
for all measurements.
Measurements
Gas-Exchange and Isoprene Emission Measurements
CO2 and water exchange were measured by using a LI-7000 infrared gas
analyzer (LI-COR). Two entire Arabidopsis plants were placed in a 1.7-L glass
cuvette. Plants were exposed to synthetic air made by mixing O2, N2, and CO2
from cylinders deprived of contaminants. The absence of contamination by
trace gases that could interfere with isoprene measurements was tested every
time a cylinder was changed, using the spectrometric analysis outlined below.
Atmospheric concentrations of the three gases (20%, 80%, and 390 mmol
mol21, respectively) were set with mass flow controllers. The air flow was set
at 2 L min21. During gas-exchange measurements leaf temperature, light
intensity, and relative humidity were maintained at constant levels close to
growth conditions (22�C, 150 mmol m22 s21, and 56%, respectively).
Steady-state photosynthesis, stomatal conductance, and transpiration in
Platanus leaves were measured by a portable gas-exchange system (LI-6400)
equipped with a 6-cm2 cuvette. Measurements were made in ambient CO2
concentration (390 mmol mol21) on individual leaves enclosed into a leaf
cuvette under a rate of 0.44 L min21 air flow, relative humidity within the
cuvette at 50% to 55%, a leaf temperature of 25�C, and 500 mmol photons m22
s21 light intensity.
Isoprene emission was measured on both Arabidopsis and Platanus plants
online, by diverting the air at the exit of the gas-exchange cuvettes into a
proton transfer reaction-mass spectrometer (Ionicon). Details on isoprene
analysis by proton transfer reaction-mass spectrometer can be found in Tholl
et al. (2006).
Data represent means6 SE of measurements on seven different plants. The
significant differences between means were determined by Student’s t test.
Means significantly different at P, 0.05 and 0.01 confidence levels are labeled
by one or two asterisks, respectively.
CD Spectroscopy
CD was measured in a JobinYvon CD6 dichrograph (JobinYvon ISA)
equipped with a thermostated sample holder on Arabidopsis leaves, and with
Jasco 815 dichrograph equippedwith Peltier sample holder on Platanus leaves.
The spectra were recorded between 400 and 750 nm in 1-nm steps with an
integration time of 0.3 s and a band pass of 2 nm. For the measurements of
thermal stability, Arabidopsis or Platanus leaves were placed in the sample
holder and were dark adapted for 10 min at 20�C. Then the temperature was
gradually increased up to 80�C (60�C in Arabidopsis due to higher thermal
sensitivity of this plant) in 5�C steps. The leaves were incubated for 10 min
before measurements at each temperature. The transition temperature (Tt) is
the temperature at which the intensity of the CD band is decreased by 50% of
its value at 25�C (Cseh et al., 2000). The transition temperatures for the main
C-type CD band at around (+)694 nm were calculated from the inflection
point of the fitted curve of the temperature dependence of the CD ampli-
tude. The experiment was repeated on seven leaves of different plants per
treatment, on each one of the two species. The mean transition temper-
atures of isoprene-emitting and nonemitting leaves were separated by
Student’s t test and positively tested against a P , 0.01.The spectra were
corrected for a flat baseline obtained by setting the CD values at 400 and
750 nm to zero.
Electrochromic Absorbance Changes (DA515)
Electrochromic absorbance changes (DA515; also named electrochromic
shift) induced by saturating single-turnover flashes were measured at 515 nm
on detached leaves in a setup described earlier (Horvath et al., 1979; Barabas
et al., 1985; Buchel and Garab, 1995). Briefly, the single-beam kinetic spectro-
photometer consists of a tungsten light source, a grating monochromator, and
Isoprene and Thermal Resistance of Leaves
Plant Physiol. Vol. 157, 2011 913 www.plantphysiol.orgon May 19, 2020 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.

a photomultiplier with an optical filter for the blue spectral region, and is
equipped with two xenon lamps (Stroboslave, General Electric) providing red
flashes of 3 ms half-duration. For recording the signal and data acquisition, a
differential amplifier and a digital averaging storage scope (Tektronix AM502
and 2224, respectively) are used in combination with a home-built timer unit
and a personal computer. The Arabidopsis plants used for these measure-
ments were dark adapted at 20�C for 20 min. Then, detached wild-type and
IspS leaves were incubated for 5 min either at 20�C or 40�C in the dark, and
thereafter measured at 20�C. In Platanus, control (isoprene-emitting) and
fosmidomycin-treated (isoprene-inhibited) leaves were exposed to 25�C or
45�C for 4 h in weak white light (40 mmol photons m22 s21). The treatment
with Platanus leaves was much longer than in the case of Arabidopsis and was
carried out in dim light to allow complete infiltration of fosmidomycin in the
leaves. In the last 20 min of the heat treatment, the light was turned off to
ensure the dark-adapted state of the leaves. Kinetic traces (n = 64) were
collected with a repetition rate of 1 s21, and averaged; the time constant of the
measurements was adjusted to 100 ms. The measurements were repeated at
least six times on different leaves. The half-times of the decay were obtained
by fitting the curves with single exponential decay functions.
TL
TL was detected with a photomultiplier tube (Hamamatsu R943-02,
Hamamatsu Photonics) linked to an amplifier in a home-built apparatus. A
detailed description of the block diagram of the TL equipment, handling,
temperature regulation, personal computer-based TL data acquisition, and
graphical simulation is in Zeinalov and Maslenkova (1996). After a 30-min
dark incubation at room temperature leaf segments were placed on a sample
holder and covered with a thin plastic plate. TL was excited by one or two
saturating (4J) single-turnover flashes (approximately 10 ms half-band with
1-Hz frequency) given at 1�C. After the flash exposure, the sample from
Arabidopsis leaves was quickly cooled to 250�C with the aid of a metal block
cooled by liquid nitrogen before being warmed to 70�C with 0.6�C/s heatingrate. TL emission from Platanus leaves was recorded similarly, but samples
were not frozen before warming from 0�C to 70�C. This way the distortion of
the TL signal during freezing that was observed in Platanus as well as in some
other species (Homann, 1999), was avoided. The nomenclature of Vass and
Govindjee (1996) was used for characterization of the main TL curve peaks. TL
measurements were repeated on 10 leaves from different plants per treatment,
on each plant species. TL curves shown in Figure 4 are representative of 10
measurements on leaves from different plants.
In another experiment, Arabidopsis and Platanus leaves were incubated for
5 min at 25�C or 40�C before TL measurements. TL emission was then
recorded following the same procedure as outlined above. Means and SEs for
TL B-peak parameters (temperature positions and amplitude; Fig. 5) were
calculated from five replicates of each nontreated and high-temperature-
treated leaf samples from different plants. The significant differences between
means at different temperatures were determined by Student’s t test. Means
significantly different at P, 0.05 and 0.01 confidence levels are labeled by one
or two asterisks, respectively. Analysis of the TL curves was carried out using
a Microcal Origin v.6.0 software package (Microcal Software).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. Representative traces of electrochromic absor-
bance changes measured at 515 nm in isoprene-emitting and nonemit-
ting leaves.
Received June 27, 2011; accepted July 28, 2011; published August 1, 2011.
LITERATURE CITED
Barabas K, Zimanyi L, Garab G (1985) Kinetics of the flash-induced
electrochromic absorbency changes in the presence of background
illumination—turnover rate of the electron transport. 1. Isolated intact
chloroplasts. J Bioenerg Biomembr 17: 349–364
Behnke K, Ehlting B, Teuber M, Bauerfeind M, Louis S, Hansch R, Polle
A, Bohlmann J, Schnitzler JP (2007) Transgenic, non-isoprene emitting
poplars don’t like it hot. Plant J 51: 485–499
Behnke K, Kleist E, Uerlings R, Wildt J, Rennenberg H, Schnitzler JP
(2009) RNAi-mediated suppression of isoprene biosynthesis in hybrid
poplar impacts ozone tolerance. Tree Physiol 29: 725–736
Buchel C, Garab G (1995) Electrochromic absorbance changes in the
chlorophyll-c-containing alga Pleurocloris meiringensis (Xanthophyceae).
Photosynth Res 43: 49–56
Bukhov HG, Wiese C, Neimanis S, Heber U (1999) Heat sensitivity of
chloroplasts and leaves: leakage of protons from thylakoids and revers-
ible activation of cyclic electron transport. Photosynth Res 59: 81–93
Claeys M, Wang W, Ion AC, Kourtchev I, Gelencserb A, Maenhaut W
(2004) Formation of secondary organic aerosols from isoprene and its
gas-phase oxidation products through reaction with hydrogen peroxide.
Atmos Environ 38: 4093–4098
Copolovici LO, Niinemets U (2005) Temperature dependencies of Henry’s
law constants and octanol/water partition coefficients for key plant
volatile monoterpenoids. Chemosphere 61: 1390–1400
Cseh Z, Rajagopal S, Tsonev T, Busheva M, Papp E, Garab G (2000)
Thermooptic effect in chloroplast thylakoid membranes: thermal and
light stability of pigment arrays with different levels of structural com-
plexity. Biochemistry 39: 15250–15257
Cseh Z, Vianelli A, Rajagopal S, Krumova S, Kovacs L, Papp E, Barzda V,
Jennings R, Garab G (2005) Thermo-optically induced reorganizations
in the main light harvesting antenna of plants. I. Non-Arrhenius type of
temperature dependence and linear light-intensity dependencies. Pho-
tosynth Res 86: 263–273
De Boeck HJ, Dreesen FE, Janssens IA, Nijs I (2011) Whole-system
responses of experimental plant communities to climate extremes im-
posed in different seasons. New Phytol 189: 806–817
Dobrikova AG, Varkonyi Z, Krumova SB, Kovacs L, Kostov GK,
Todinova SJ, Busheva MC, Taneva SG, Garab G (2003) Structural
rearrangements in chloroplast thylakoid membranes revealed by dif-
ferential scanning calorimetry and circular dichroism spectroscopy:
thermo-optic effect. Biochemistry 42: 11272–11280
Ducruet JM (2003) Chlorophyll thermoluminescence of leaf discs: simple
instruments and progress in signal interpretation open the way to new
ecophysiological indicators. J Exp Bot 54: 2419–2430
Ducruet JM, Peeva V, Havaux M (2007) Chlorophyll thermofluorescence
and thermoluminescence as complementary tools for the study of
temperature stress in plants. Photosynth Res 93: 159–171
Ervens B, Carlton AG, Turpin BJ, Altieri KE, Kreidenweis SM, Feingold
G (2008) Secondary organic aerosol yields from cloud-processing of
isoprene oxidation products. Geophys Res Lett 35: L02816
Fares S, Oksanen E, Lannenpaa M, Julkunen-Tiitto R, Loreto F (2010)
Volatile emissions and phenolic compound concentrations along a
vertical profile of Populus nigra leaves exposed to realistic ozone con-
centrations. Photosynth Res 104: 61–74
Garab G (1996) Linear and circular dichroism. In J Amesz, AJ Hoff, eds,
Biophysical Techniques in Photosynthesis. Kluwer, Dordrecht, The
Netherlands, pp 11–40
Garab G, Cseh Z, Kovacs L, Rajagopal S, Varkonyi Z, Wentworth M,
Mustardy L, Der A, Ruban A, Papp E, et al. (2002) Light-induced trimer
to monomer transition in the main light-harvesting antenna complex of
plants: thermo-optic mechanism. Biochemistry 41: 15121–15129
Garab G, Kieleczawa J, Sutherland JC, Bustamante C, Hind G (1991)
Organization of pigment-protein complexes into macrodomains in the
thylakoid membranes of wild-type and chlorophyll b-less mutant of barley
as revealed by circular dichroism. Photochem Photobiol 54: 273–281
Garab G, Mustardy L (1999) Role of LHCII-containing macrodomains in the
structure, function and dynamics of grana. Aust J Plant Physiol 26: 649–658
Garab G, van Amerongen H (2009) Linear dichroism and circular dichro-
ism in photosynthesis research. Photosynth Res 101: 135–146
Georgieva K, Maslenkova L, Peeva V, Markovska Y, Stefanov D, Tuba Z
(2005) Comparative study on the changes in photosynthetic activity of
the homoiochlorophyllous desiccation-tolerant Haberlea rhodopensis and
spinach leaves during desiccation and rehydration. Photosynth Res 85:
191–203
Gombos Z, Varkonyi Z, Hagio M, Iwaki M, Kovacs L, Masamoto K, Itoh
S, Wada H (2002) Phosphatidylglycerol requirement for the function of
electron acceptor plastoquinone Q(B) in the photosystem II reaction
center. Biochemistry 41: 3796–3802
Govindjee, Kambara T, Coleman W (1985) The electron donor side of
photosystem II: the oxygen evolving complex. Photochem Photobiol 42:
187–210
Velikova et al.
914 Plant Physiol. Vol. 157, 2011 www.plantphysiol.orgon May 19, 2020 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.

Guenther A, Karl T, Harley P, Wiedinmyer C, Palmer PI, Geron C (2006)
Estimates of global terrestrial isoprene emissions using MEGAN (Model
of Emissions of Gases and Aerosols from Nature). Atmos Chem Phys
Discuss 6: 107–173
Homann PH (1999) Reliability of photosystem II thermoluminescence
measurements after sample freezing: few artifacts with photosystem II
membranes but gross distortions with certain leaves. Photosynth Res 62:
219–229
Horvath G, Niemi HA, Droppa M, Faludi-Daniel A (1979) Characteristics
of the flash-induced 515 nm change of intact isolated chloroplasts. Plant
Physiol 63: 778–782
Junge W (1977) Membrane potentials in photosynthesis. Annu Rev Plant
Physiol 28: 503–536
Keller D, Bustamante C (1986) Theory of the interaction of light with large
inhomogeneous molecular aggregates. II. Psi-type circular dichroism.
J Chem Phys 84: 2972–2979
Kleindienst TE, Lewandowski M, Offenberg JH, Jaoui M, Endey EO
(2007) Ozone-isoprene reaction: re-examination of the formation of
secondary organic aerosols. Geophys Res Lett 34: L01805
Kovacs L, Damkjaer J, Kereıche S, Ilioaia C, Ruban AV, Boekema EJ,
Jansson S, Horton P (2006) Lack of the light-harvesting complex CP24
affects the structure and function of the grana membranes of higher
plant chloroplasts. Plant Cell 18: 3106–3120
Kroll JH, Ng NL, Murphy SM, Flagan RC, Seinfeld JH (2006) Secondary
organic aerosol formation from isoprene photooxidation. Environ Sci
Technol 40: 1869–1877
Krumova SB, Laptenok SP, Kovacs L, Toth T, van Hoek A, Garab G, van
Amerongen H (2010) Digalactosyl-diacylglycerol-deficiency lowers the
thermal stability of thylakoid membranes. Photosynth Res 105: 229–242
Logan BA, Monson RK (1999) Thermotolerance of leaf discs from four
isoprene-emitting species is not enhanced by exposure to exogenous
isoprene. Plant Physiol 120: 821–826
Loivamaki M, Gilmer F, Fischbach RJ, Sorgel C, Bachl A, Walter A,
Schnitzler J-P (2007) Arabidopsis, a model to study biological functions
of isoprene emission? Plant Physiol 144: 1066–1078
Loreto F, Mannozzi M, Maris C, Nascetti P, Ferranti F, Pasqualini S (2001)
Ozone quenching properties of isoprene and its antioxidant role in
leaves. Plant Physiol 126: 993–1000
Loreto F, Schnitzler J-P (2010) Abiotic stresses and induced BVOCs. Trends
Plant Sci 15: 154–166
Loreto F, Velikova V (2001) Isoprene produced by leaves protects the
photosynthetic apparatus against ozone damage, quenches ozone prod-
ucts, and reduces lipid peroxidation of cellular membranes. Plant
Physiol 127: 1781–1787
Maslenkova L, Homann P (2000) Stabilized S2 state in leaves of the
desiccation tolerant resurrection fern Polipodium polipodioides. Compt
Rend Bulg Acad Sci 53: 99–102
Matsunaga SN, Wiedinmyer C, Guenther AB, Orlando JJ, Karl T, Toohey
DW, Greenberg JP, Kajii Y (2005) Isoprene oxidation products are a
significant atmospheric aerosol component. Atmos Chem Phys Discuss
5: 11143–11156
Monson RK, Holland E (2001) Biospheric trace gas fluxes and their control
over tropospheric chemistry. Annu Rev Ecol Evol Syst 32: 547–576
Monson RK, Jaeger CH, Adams WW, Driggers EM, Silver GM, Fall R
(1992) Relationships among isoprene emission rate, photosynthesis, and
isoprene synthase activity as influenced by temperature. Plant Physiol
98: 1175–1180
O’Brien JM, Shepson PB, Muthuramu K, Hao C, Niki H, Hastie DR,
Taylor R, Roussel PB (1995) Measurements of alkyl and multifunctional
organic nitrates at a rural site in Ontario. J Geophys Res 100: 22795–
22804
Pastenes C, Horton P (1996) Effect of high temperature on photosynthesis
in beans. (I. Oxygen evolution and chlorophyll fluorescence). Plant
Physiol 112: 1245–1251
Paulot F, Crounse JD, Kjaergaard HG, Kurten A, St Clair JM, Seinfeld JH,
Wennberg PO (2009) Unexpected epoxide formation in the gas-phase
photooxidation of isoprene. Science 325: 730–733
Peeva V, Maslenkova L (2004) Thermoluminescence study of photosystem
II activity in Haberlea rhodopensis and spinach leaves during desiccation.
Plant Biol (Stuttg) 6: 319–324
Peters RLA, van Kooten O, Vredenberg WJ (1984) The kinetics of P515 in
relation to the lipid composition of the thylakoid membrane. J Bioenerg
Biomembr 16: 283–294
Rutherford AW, Govindjee, Inoue Y (1984) Charge accumulation and
photochemistry in leaves studied by thermoluminescence and delayed
light emission. Proc Natl Acad Sci USA 81: 1107–1111
Sane PV (2004) Thermoluminescence: a technique for probing photosystem
II. In R Carpentier, ed, Photosynthesis Research Protocols. Humana
Press, Totowa, NJ, pp 229–248
Sane PV, Rutherford AW (1986) Thermoluminescence from photosynthetic
membranes. In Govindjee, J Amesz, DC Fork, eds, Light Emission by
Plants and Bacteria. Academic Press, Orlando, FL, pp 329–360
Santarius KA (1980) Membrane lipids in heat injury of spinach chloro-
plasts. Physiol Plant 49: 1–6
Sasaki K, Saito T, Lamsa M, Oksman-Caldentey KM, Suzuki M, Ohyama
K, Muranaka T, Ohara K, Yazaki K (2007) Plants utilize isoprene emis-
sion as a thermotolerance mechanism. Plant Cell Physiol 48: 1254–1262
Sass L, Csintalan Z, Tuba Z, Vass I (1996) Thermoluminescence studies on
the function of photosystem II in the desiccation tolerant lichen Cladonia
convoluta. Photosynth Res 48: 205–212
Schnitzler J-P, Zimmer I, Bachl A, Arend M, Fromm J, Fischbach RJ (2005)
Biochemical properties of isoprene synthase in poplar (Populus x
canescens). Planta 222: 777–786
Schrader SM, Wise RR, Wacholtz WF, Ort DR, Sharkey TD (2004)
Thylakoid membrane responses to moderately high leaf temperature
in pima cotton. Plant Cell Environ 27: 725–735
Sharkey TD, Chen X, Yeh S (2001) Isoprene increases thermotolerance of
fosmidomycin-fed leaves. Plant Physiol 125: 2001–2006
Sharkey TD, Singsaas EL (1995) Why plants emit isoprene. Nature 374: 769
Sharkey TD, Wiberley AE, Donohue AR (2008) Isoprene emission from
plants: why and how. Ann Bot (Lond) 101: 5–18
Sharkey TD, Yeh S (2001) Isoprene emission from plants. Annu Rev Plant
Physiol Plant Mol Biol 52: 407–436
Sharkey TD, Yeh S, Wiberley AE, Falbel TG, Gong D, Fernandez DE
(2005) Evolution of the isoprene biosynthetic pathway in kudzu. Plant
Physiol 137: 700–712
Simidjiev I, Barzda V, Mustardy L, Garab G (1998) Role of thylakoid lipids
in the structural flexibility of lamellar aggregates of the isolated light-
harvesting chlorophyll a/b complex of photosystem II. Biochemistry 37:
4169–4173
Singsaas EL, Lerdau M, Winter K, Sharkey TD (1997) Isoprene increases
thermotolerance of isoprene-emitting species. Plant Physiol 115:
1413–1420
Singsaas EL, Sharkey TD (2000) The effects of high temperature on
isoprene synthesis in oak leaves. Plant Cell Environ 23: 751–757
SiwkoME,Marrink SJ, de Vries AH, Kozubek A, Schoot Uiterkamp AJM,
Mark AE (2007) Does isoprene protect plant membranes from thermal
shock? A molecular dynamics study. Biochim Biophys Acta 1768:
198–206
Sun EJ, Huang MH (1995) Detection of peroxyacetyl nitrate at phytotoxic
level and its effects on vegetation in Taiwan. Atmos Environ 29:
2899–2904
Tholl D, Boland W, Hansel A, Loreto F, Rose US, Schnitzler J-P (2006)
Practical approaches to plant volatile analysis. Plant J 45: 540–560
Vass I, Govindjee (1996) Thermoluminescence from the photosynthetic
apparatus. Photosynth Res 48: 117–126
Vass I, Inoue Y (1992) Thermoluminescence in the study of photosystem II.
In J Barber, ed, The Photosystems: Structure, Function and Molecular
Biology. Elsevier, Amsterdam, pp 259–294
Veatch SL (2007) From small fluctuations to large-scale phase separation:
lateral organization in model membranes containing cholesterol. Semin
Cell Dev Biol 18: 573–582
Velikova V, Fares S, Loreto F (2008) Isoprene and nitric oxide reduce
damages in leaves exposed to oxidative stress. Plant Cell Environ 31:
1882–1894
Velikova V, Loreto F (2005) On the relationship between isoprene emission
and thermotolerance in Phragmites australis leaves exposed to high
temperature and during the recovery from a heat stress. Plant Cell
Environ 28: 318–327
Velikova V, Loreto F, Tsonev T, Brilli F, Edreva A (2006) Isoprene prevents
the negative consequences of high temperature stress in Platanus
orientalis leaves. Funct Plant Biol 33: 931–940
Vickers CE, Gershenzon J, Lerdau MT, Loreto F (2009a) A unified mech-
anism of action for volatile isoprenoids in plant abiotic stress. Nat Chem
Biol 5: 283–291
Vickers CE, Possell M, Cojocariu CI, Velikova VB, Laothawornkitkul J,
Isoprene and Thermal Resistance of Leaves
Plant Physiol. Vol. 157, 2011 915 www.plantphysiol.orgon May 19, 2020 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.

Ryan A, Mullineaux PM, Nicholas Hewitt C (2009b) Isoprene synthesis
protects transgenic tobacco plants from oxidative stress. Plant Cell
Environ 32: 520–531
WildermuthMC, Fall R (1998) Biochemical characterization of stromal and
thylakoid-bound isoforms of isoprene synthase in willow leaves. Plant
Physiol 116: 1111–1123
Williams WP, Quinn PJ (1987) The phase behavior of lipids in photosyn-
thetic membranes. J Bioenerg Biomembr 19: 605–624
Witt HT (1979) Energy conversion in the functional membrane of photo-
synthesis: analysis by light pulse and electric pulse methods: the central
role of the electric field. Biochim Biophys Acta 505: 355–427
Zeidler J, Schwender J, Muller C, Wiesner J, Weidemeyer C, Beck E,
Jomaa H, Lichtenthaler HK (1998) Inhibition of the non-mevalonate
1-deoxy-D-xylulose-5-phosphate pathway of plant isoprenoid biosyn-
thesis by fosmidomycin. Z Naturforsch 53c: 980–986
Zeinalov Y, Maslenkova L (1996) Mechanisms of photosynthetic oxygen
evolution. In M Pessarakli, ed, Handbook of Photosynthesis, Marcel
Dekker, New York, pp 129–150
Zhang R, Cruz JA, Kramer DM, Magallanes-Lundback ME, Dellapenna
D, Sharkey TD (2009) Moderate heat stress reduces the pH component
of the transthylakoid proton motive force in light-adapted, intact
tobacco leaves. Plant Cell Environ 32: 1538–1547
Zhang R, Sharkey TD (2009) Photosynthetic electron transport and proton
flux under moderate heat stress. Photosynth Res 100: 29–43
Velikova et al.
916 Plant Physiol. Vol. 157, 2011 www.plantphysiol.orgon May 19, 2020 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.





























