Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 76, No. 8, pp. 3880-3884, August 1979Botany
Induction of cytokinin autonomy by N,N'-diphenylurea in tissuecultures of Phaseolus lunatus L.
(plant hormones/genetic variation/bean callus)
MACHTELD C. MOK*, SANG-GU KIMt, DONALD J. ARMSTRONGt, AND DAVID W. S. MOK**Department of Horticulture and Genetics Program, Oregon State University, Corvallis, Oregon 97331; and tDepartment of Botany and Plant Pathology, OregonState University, Corvallis, Oregon 97331
Communicated by Folke Skoog, June 4,1979
ABSTRACT The ability of NN-diphenylurea (Ph2urea) tosubstitute for cytokinin-active adenine derivatives in promotingcallus growth of Phaseolus lunatus has been examined. Ingeneral, Ph2urea stimulated callus growth at high concentra-tions, although the growth of most callus tissues was irregular.Variability in the sensitivity and uniformity of the growth re-sponse to Phsurea was found among different genotypes of P.lunatus. Most importantly, tissues cultured on .Ph2urea-con-taining medium for one passage had acquired the ability toproliferate in subsequent passages in the absence of eitherPh2urea or cytokinin-active adenine derivatives. Correspondingtissues maintained on kinetin-containing medium remainedcytokinin-dependent. It appears that the effect of Ph2urea inpromoting the growth of P. lunatus callus tissue resides in itsability to induce cytokinin autonomy. This result suggests thatthe cytokinin activity of Ph2urea may be due to promotion ofendogenous cytokinin biosynthesis in the bioassay systems inwhich it is active.
The ability of N,N'-diphenylurea (Ph2urea) to stimulate celldivision in carrot tissue cultures was reported by Shantz andSteward (1) in 1955. Ph2urea and a number of related com-pounds have subsequently been shown to exhibit activity similarto that of N6-substituted adenine derivatives in various cyto-kinin bioassay systems. Ph2urea exhibits cytokinin activity inthe tobacco callus bioassay (2-5) as well as in cytokinin bioassaysbased on chlorophyll retention, bud development, and seedgermination (6). The activity of Ph2urea was, however, weakand variable in some of these tests (2, 4, 5).The similarity in the biological activity of two such struc-
turally unrelated classes of compounds (N6-substituted adeninederivatives and substituted ureas) has posed one of the moreinteresting problems in cytokinin structure-activity relation-ships. The ability of benzylurea derivatives to antagonize theeffects of both types of compounds led Kefford et al. (7) toargue that there must be a common site of action. Miller (8)suggested that Ph2urea might serve as a precursor for the N6-side chain of cytokinin-active adenine derivatives. In supportof this hypothesis, certain synthetic ureidopurines are knownto possess cytokinin activity (9), and the activity of Ph2urea inpromoting the growth of callus tissues has been reported to becorrelated with the ability of the tissues to metabolize Ph2urea(10). However, the only Ph2urea metabolite to be isolated andidentified from plant tissues has been a simple glucosyl deriv-ative (1 1).We have examined the effect of Ph2urea on the growth of
cytokinin-dependent tissue cultures of Phaseolus lunatus. Theresults reported here indicate that the activity of Ph2urea inpromoting the growth of P. lunatus callus tissue is associatedwith an induction of cytokinin autonomy (i.e., transformation
to cytokinin independence as evidenced by the ability of thetissue to grow on cytokinin-free medium). The possible sig-nificance of this result in interpreting the mechanism of actionof Ph2urea is discussed.
MATERIALS AND METHODSChemicals. Ph2urea and kinetin (N6-furfurylaminopurine)
were obtained from Sigma. Picloram (4-amino-3,5,6-trichlo-ropicolinic acid) was a gift from Dow Chemical.
Plant Materials. Seeds of P. lunatus L. cv. Kingston werelocally obtained. Seeds of plant introduction lines (P.I. lines)of P. lunatus were provided by the Regional Plant IntroductionStation, Washington State University (Pullman, WA).
Tissue Culture Medium. The tissue culture medium con-sisted of the mineral nutrients as described by Murashige andSkoog (12) with the following organic substances added: sucrose(30 g/liter), myo-inositol (100 mg/liter), thiamine-HCl (1mg/liter), nicotinic acid (5 mg/liter), pyridoxine-HCl (0.5mg/liter), and picloram (2.5 tiM). (The latter compound is usedto supply the auxin requirement of Phaseolus tissue cultures.)Kinetin (5 ,uM) was included in medium used for callus initia-tion and stock cultures. The pH of the medium was adjustedto 5.7 and Difco Bacto-agar (10 g/liter) was added. The me-dium was dispensed into 125-ml erlenmeyer flasks (50 ml perflask) and autoclaved at 120'C for 15 min. In tests of the effectof Ph2urea on callus growth, appropriate amounts of Ph2ureawere dissolved in dimethyl sulfoxide (13) and added to theautoclaved tissue culture flasks prior to solidification of themedium. (The final amount of dimethyl sulfoxide in the tissueculture medium was 0.025 ml per flask.)Growth and Harvest of Phaseolus Callus Cultures.
Phaseolus callus cultures were established from the hypocotylsof 5-day-old seedlings as described (14) or from the petioles ofmature plants. The callus tissue that formed on the initial ex-plants was transferred once (first passage) on medium con-taining 5 ,uM kinetin. Tests for Ph2urea response were per-formed in the second passage of the callus tissue by using 4-week-old first-passage cultures as stock tissue. Three pieces ofcallus weighing approximately 25 mg each were planted perflask. Tissues were harvested and weighed after 35 days ofgrowth at 280C in the dark. All tests were repeated at least onceby using newly established cultures.
RESULTSThe activity of Ph2urea in promoting P. lunatus callus growthwas first tested in cultures derived from hypocotyl tissues of P.lunatus cv. Kingston. The results of this initial test are illustratedin Fig. 1. Response to Ph2urea was detectable at concentrationsof 32 and 100 AM Ph2urea. Although the amount of growth was
Abbreviations: Ph2urea, N,N'-diphenylurea; P.I., plant introduction.
3880
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be hereby marked "ad-vertisement" in accordance with 18 U. S. C. §1734 solely to indicatethis fact.
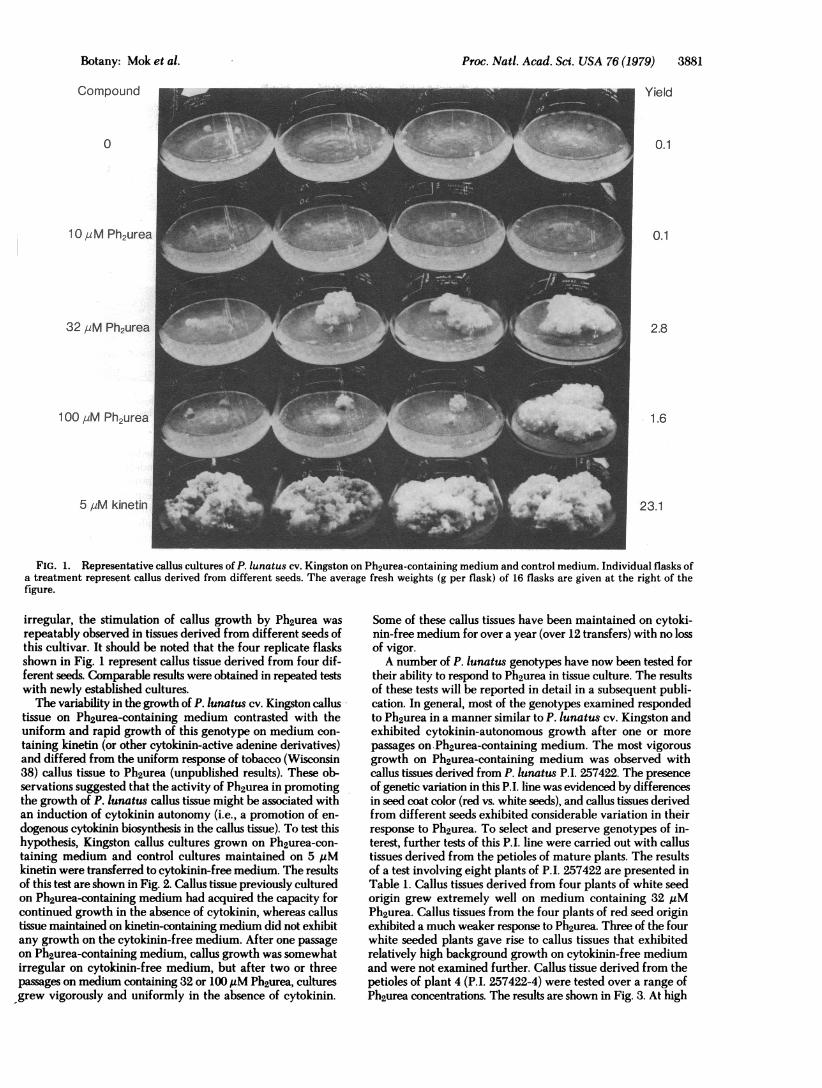
Proc. Natl. Acad. Sci. USA 76 (1979) 3881
Compound
0
10PM Ph2urea
32 yM Ph2urea
100 MM Ph2urea
5 jiM kinetin
Yield
0.1
0.1
2.8
1.6
23.1
FIG. 1. Representative callus cultures of P. lunatus cv. Kingston on Ph2urea-containing medium and control medium. Individual flasks ofa treatment represent callus derived from different seeds. The average fresh weights (g per flask) of 16 flasks are given at the right of thefigure.
irregular, the stimulation of callus growth by Ph2urea wasrepeatably observed in tissues derived from different seeds ofthis cultivar. It should be noted that the four replicate flasksshown in Fig. 1 represent callus tissue derived from four dif-ferent seeds. Comparable results were obtained in repeated testswith newly established cultures.The variability in the growth of P. lunatus cv. Kingston callus
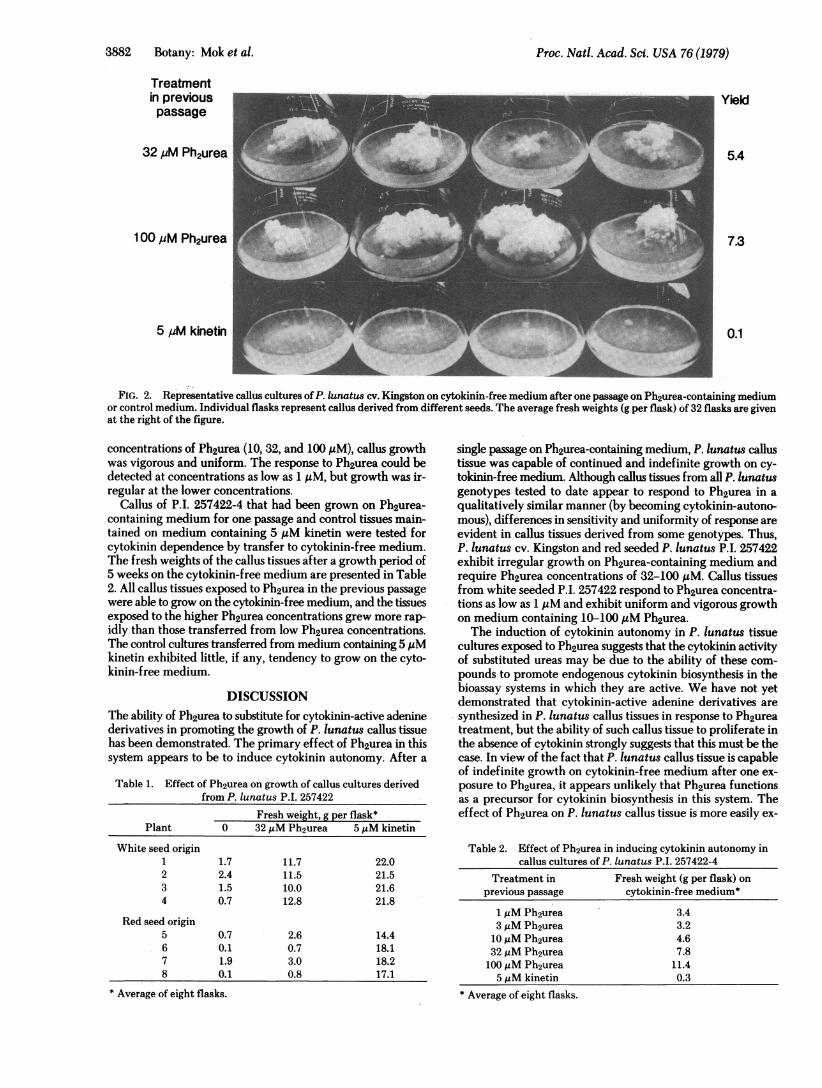
tissue on Ph2urea-containing medium contrasted with theuniform and rapid growth of this genotype on medium con-taining kinetin (or other cytokinin-active adenine derivatives)and differed from the uniform response of tobacco (Wisconsin38) callus tissue to Ph2urea (unpublished results). These ob-servations suggested that the activity of Ph2urea in promotingthe growth of P. lunatus callus tissue might be associated withan induction of cytokinin autonomy (i.e., a promotion of en-dogenous cytokinin biosynthesis in the callus tissue). To test thishypothesis, Kingston callus cultures grown on Ph2urea-con-taining medium and control cultures maintained on 5 ,uMkinetin were transferred to cytokinin-free medium. The resultsof this test are shown in Fig. 2. Callus tissue previously culturedon Ph2urea-containing medium had acquired the capacity forcontinued growth in the absence of cytokinin, whereas callustissue maintained on kinetin-containing medium did not exhibitany growth on the cytokinin-free medium. After one passageon Ph2urea-containing medium, callus growth was somewhatirregular on cytokinin-free medium, but after two or threepassages on medium containing 32 or 100 ,uM Ph2urea, culturesgrew vigorously and uniformly in the absence of cytokinin.
Some of these callus tissues have been maintained on cytoki-nin-free medium for over a year (over 12 transfers) with no lossof vigor.A number of P. lunatus genotypes have now been tested for
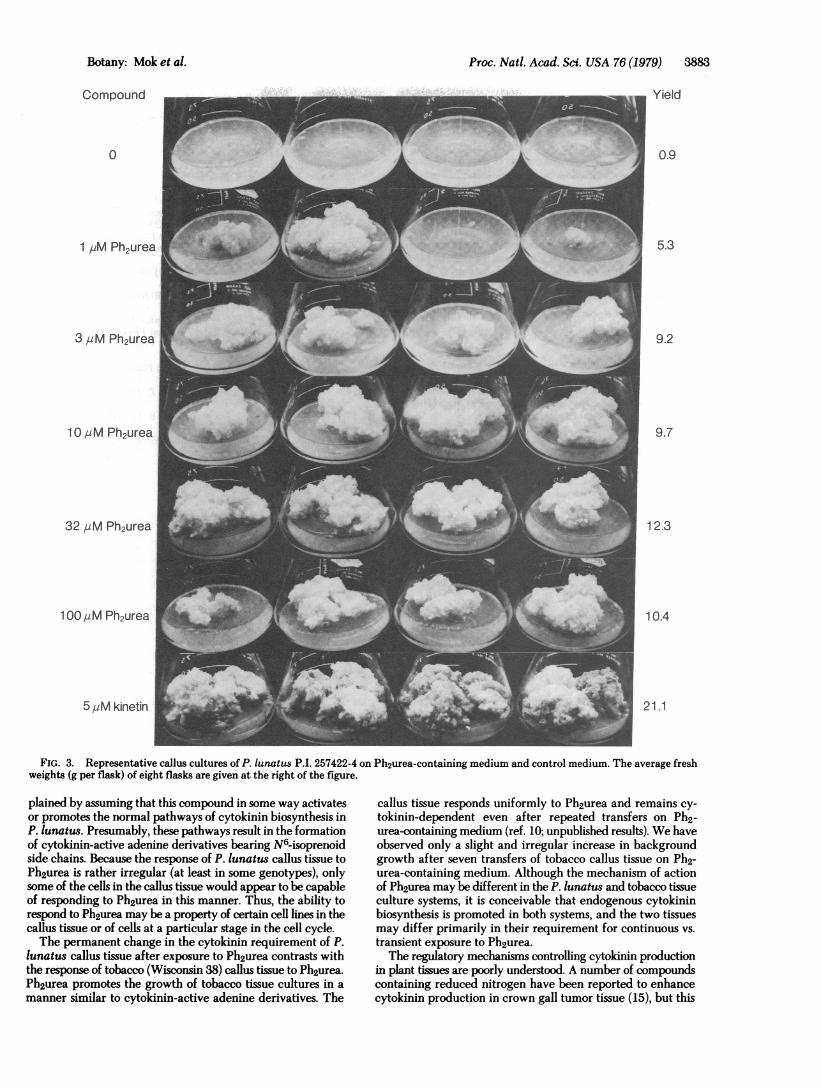
their ability to respond to Ph2urea in tissue culture. The resultsof these tests will be reported in detail in a subsequent publi-cation. In general, most of the genotypes examined respondedto Ph2urea in a manner similar to P. lunatus cv. Kingston andexhibited cytokinin-autonomous growth after one or morepassages on Ph2urea-containing medium. The most vigorousgrowth on Ph2urea-containing medium was observed withcallus tissues derived from P. lunatus P.I. 257422. The presenceof genetic variation in this P.1. line was evidenced by differencesin seed coat color (red vs. white seeds), and callus tissues derivedfrom different seeds exhibited considerable variation in theirresponse to Ph2urea. To select and preserve genotypes of in-terest, further tests of this P.I. line were carried out with callustissues derived from the petioles of mature plants. The resultsof a test involving eight plants of P.I. 257422 are presented inTable 1. Callus tissues derived from four plants of white seedorigin grew extremely well on medium containing 32 ,uMPh2urea. Callus tissues from the four plants of red seed originexhibited a much weaker response to Ph2urea. Three of the fourwhite seeded plants gave rise to callus tissues that exhibitedrelatively high background growth on cytokinin-free mediumand were not examined further. Callus tissue derived from thepetioles of plant 4 (P.1. 257422-4) were tested over a range ofPh2urea concentrations. The results are shown in Fig. 3. At high
B.-.Aany: Mok et al.
Proc. Natl. Acad. Sci. USA 76 (1979)
Treatmentin previouspassage
32 ,uM Ph2urea
100pM Ph2urea
5 pM kinetin
Yield
5.4
7.3
0.1L~~~~~~~~~~~~~~~~~~~~~~~~~~.*.-.. ... .S.. ;;m
FIG. 2. Representative callus cultures of P. lunatus cv. Kingston on cytokinin-free medium after one passage on Ph2urea-containing mediumor control medium. Individual flasks represent callus derived from different seeds. The average fresh weights (g per flask) of 32 flasks are givenat the right of the figure.
concentrations of Ph2urea (10, 32, and 100 ,uM), callus growthwas vigorous and uniform. The response to Ph2urea could bedetected at concentrations as low as 1 ,uM, but growth was ir-
regular at the lower concentrations.Callus of P.I. 257422-4 that had been grown on Ph2urea-
containing medium for one passage and control tissues main-tained on medium containing 5 ,uM kinetin were tested forcytokinin dependence by transfer to cytokinin-free medium.The fresh weights of the callus tissues after a growth period of5 weeks on the cytokinin-free medium are presented in Table2. All callus tissues exposed to Ph2urea in the previous passagewere able to grow on the cytokinin-free medium, and the tissuesexposed to the higher Ph2urea concentrations grew more rap-idly than those transferred from low Ph2urea concentrations.The control cultures transferred from medium containing 5 JIMkinetin exhibited little, if any, tendency to grow on the cyto-kinin-free medium.
DISCUSSIONThe ability of Ph2urea to substitute for cytokinin-active adeninederivatives in promoting the growth of P. lunatus callus tissuehas been demonstrated. The primary effect of Ph2urea in thissystem appears to be to induce cytokinin autonomy. After a
Table 1. Effect of Ph2urea on growth of callus cultures derivedfrom P. lunatus P.I. 257422
Fresh weight, g per flask*Plant 0 32,uM Ph2urea 5 AM kinetin
White seed origin1 1.7 11.7 22.02 2.4 11.5 21.53 1.5 10.0 21.64 0.7 12.8 21.8
Red seed origin5 0.7 2.6 14.46 0.1 0.7 18.17 1.9 3.0 18.28 0.1 0.8 17.1
* Average of eight flasks.
single passage on Ph2urea-containing medium, P. lunatus callustissue was capable of continued and indefinite growth on cy-tokinin-free medium. Although callus tissues from all P. lunatusgenotypes tested to date appear to respond to Ph2urea in aqualitatively similar manner (by becoming cytokinin-autono-mous), differences in sensitivity and uniformity of response areevident in callus tissues derived from some genotypes. Thus,P. lunatus cv. Kingston and red seeded P. lunatus P.I. 257422exhibit irregular growth on Ph2urea-containing medium andrequire Ph2urea concentrations of 32-100 ,gM. Callus tissuesfrom white seeded P.1. 257422 respond to Ph2urea concentra-tions as low as 1 ,uM and exhibit uniform and vigorous growthon medium containing 10-100,uM Ph2urea.The induction of cytokinin autonomy in P. lunatus tissue
cultures exposed to Ph2urea suggests that the cytokinin activityof substituted ureas may be due to the ability of these com-pounds to promote endogenous cytokinin biosynthesis in thebioassay systems in which they are active. We have not yetdemonstrated that cytokinin-active adenine derivatives aresynthesized in P. lunatus callus tissues in response to Ph2ureatreatment, but the ability of such callus tissue to proliferate inthe absence of cytokinin strongly suggests that this must be thecase. In view of the fact that P. lunatus callus tissue is capableof indefinite growth on cytokinin-free medium after one ex-
posure to Ph2urea, it appears unlikely that Ph2urea functionsas a precursor for cytokinin biosynthesis in this system. Theeffect of Ph2urea on P. lunatus callus tissue is more easily ex-
Table 2. Effect of Ph2urea in inducing cytokinin autonomy incallus cultures of P. lunatus P.I. 257422-4
Treatment in Fresh weight (g per flask) onprevious passage cytokinin-free medium*
1 MM Ph2urea 3.43AM Ph2urea 3.210M Ph2urea 4.632 MM Ph2urea 7.8
100 AM Ph2urea 11.45MM kinetin 0.3
* Average of eight flasks.
3882 Botany: Mok et al.
Proc. Natl. Acad. Sci. USA 76 (1979) 3883
Compound
0
1 pM Ph2urea
3pM Ph2urea
10pM Ph2urea
32 pM Ph2urea
100pM Ph2urea
5 PM kinetin
Yield
0.9
5.3
9.2
9.7
12.3
10.4
21.1
FIG. 3. Representative callus cultures of P. lunatus P.1. 257422-4 on Ph2urea-containing medium and control medium. The average freshweights (g per flask) of eight flasks are given at the right of the figure.
plained by assuming that this compound in some way activatesor promotes the normal pathways of cytokinin biosynthesis inP. lunatus. Presumably, these pathways result in the formationof cytokinin-active adenine derivatives bearing N6-isoprenoidside chains. Because the response of P. lunatus callus tissue toPh2urea is rather irregular (at least in some genotypes), onlysome of the cells in the callus tissue would appear to be capableof responding to Ph2urea in this manner. Thus, the ability torespond to Ph2urea may be a property of certain cell lines in thecallus tissue or of cells at a particular stage in the cell cycle.The permanent change in the cytokinin requirement of P.
lunatus callus tissue after exposure to Ph2urea contrasts withthe response of tobacco (Wisconsin 38) callus tissue to Ph2urea.Ph2urea promotes the growth of tobacco tissue cultures in amanner similar to cytokinin-active adenine derivatives. The
callus tissue responds uniformly to Ph2urea and remains cy-tokinin-dependent even after repeated transfers on Ph2-urea-containing medium (ref. 10; unpublished results). We haveobserved only a slight and irregular increase in backgroundgrowth after seven transfers of tobacco callus tissue on Ph2-urea-containing medium. Although the mechanism of actionof Ph2urea may be different in the P. lunatus and tobacco tissueculture systems, it is conceivable that endogenous cytokininbiosynthesis is promoted in both systems, and the two tissuesmay differ primarily in their requirement for continuous vs.transient exposure to Ph2urea.The regulatory mechanisms controlling cytokinin production
in plant tissues are poorly understood. A number of compoundscontaining reduced nitrogen have been reported to enhancecytokinin production in crown gall tumor tissue (15), but this
JPL,-.tany: Mok et al.

Proc. Natl. Acad. Sci. USA 76 (1979)
observation does not appear to be directly related to the re-sponse of P. lunatus callus tissue to Ph2urea. The Phaseolustissue culture medium already contains high concentrations ofreduced nitrogen in the form of ammonium ions, and urea itselfdoes not promote the growth of P. lunatus callus cultures. Toour knowledge, the only previous reports of chemical inductionof autonomy in cytokinin-dependent plant tissue cultures haveinvolved exposure of tobacco tissue cultures to inhibitory con-centrations of streptomycin (16) or to treatments with mor-phactins or chemically related compounds (aminofluorenes)(17, 18). The latter compounds are carcinogenic in animalsystems, and it is doubtful that their mode of action in inducingcytokinin autonomy in tobacco callus tissue is equivalent to thatof Ph2urea in P. lunatus callus cultures. The results reportedhere do not permit any conclusions concerning the mechanismby which Ph2urea induces cytokinin autonomy in P. lunatuscallus tissue, but the possibility that Ph2urea may promote en-dogenous cytokinin production in other plant systems meritsinvestigation.
This is technical paper 5146 of the Oregon Agricultural ExperimentStation. Research was supported by the Science and Education Ad-ministration of the U.S. Department of Agriculture under Grant5901-0410-8-0028-0 from the Competitive Research Grants office andby the Oregon Agricultural Experiment Station.
1. Shantz, E. M. & Steward, F. C. (1955) J. Am. Chem. Soc. 77,6351-6353.
2. Bottomly, W., Kefford, N. P., Zwar, J. A. & Goldacre, P. L. (1963)Aust. J. Biol. Sci. 16,395-409.
3. Bruce, M. I. & Zwar, J. A. (1966) Proc. R. Soc. London Ser. B 165,245-265.
4. Miller, C. 0. (1960) Plant Physiol. 35, xxvi.5. Strong, F. M. (1958) in Topics in Microbial Chemistry (Wiley,
New York), pp. 98-157.6. Bruce, M. I., Zwar, J. A. & Kefford, N. P. (1965) Life Sci. 4,
461-466.7. Kefford, N. P., Zwar, J. A. & Bruce, M. I. (1968) in Biochemistry
and Physiology of Plant Growth Substances, eds. Wightman,F. & Setterfield, G. (Runge, Ottawa, ON, Canada), pp. 61-69.
8. Miller, C. 0. (1961) Annu. Rev. Plant Physiol. 12,395-408.9. McDonald, J. J., Leonard, N. J., Schmitz, R. Y. & Skoog, F. (1971)
Phytochemistry 10, 1429-1439.10. Dyson, W. H., Fox, J. E. & McChesney, J. D. (1972) Plant Physiol.
49,506-513.11. Burrows, W. J. & Leworthy, D. P. (1976) Biochem. Biophys. Res.
Commun. 70, 1109-1114.12. Murashige, T. & Skoog, F. (1962) Physiol. Plant. 15,473-497.13. Schmitz, R. Y. & Skoog, F. (1970) Plant Physiol. 45,537-538.14. Mok, M. C. & Mok, D. W. S. (1977) Physiol. Plant 40, 261-
264.15. Peterson, J. B. & Miller, C. 0. (1976) Plant Physiol. 57, 393-
399.16. Kaminek, M. & Lustinec, J. (1974) Z. Pflanzenphysiol. 73,
74-81.17. Bednar, T. W. & Linsmaier-Bednar, E. M. (1971) Proc. Natl.
Acad. Sci. USA 68, 1178-1179.18. Bednar, T. W. & Linsmaier-Bednar, E. M. (1971) Chem.-Biol.
Interact. 4, 233-238.
3884 Botany: Mok et al.





























