MARCH 5, 1960 TREATMENT OF RENAL DISEASES MEDIBA JOURNAL 671
urinary infection is reasonably common in thecommunity, but few
patients in the acute attack enterthe practice of a hospital.
Strict criteria of diagnosiscan be established, using the
facilities available in anyclinical laboratory. Given these
facilities, treatment canbe standardized, and its short-term
success or failurecan be established. Finally, the long-term result
willbecome apparent only after many years, during whichthe patient
will have no symptoms that would makerepeated visits to a hospital
for follow-up seem reason-able; but his family doctor could check
his blood-pressure and examine his urine without any
particulardisturbance.
REFERENCELSFairbrother, R. W. (1959). A Text-book of
Bacteriology, 8th ed.
Heinemann, London.Grollman, E. F., and Grollman, A. (1959). J.
clin. I,zvest., 38,
749.Himsworth, H. P. (1949). Lancet, 1, 465.James, U. (1959).
Lancet, 2, 1001.Merrill, J. P. (1955). The Treatment of Renal
Failure. Grune
and Stratton, New York.Oliver, J. (1950). Amer. J. Med., 9,
88.Parsons, F. M., and McCracken, B. H. (1959). Brit. med. J.,
1,
740.Platt, R. (1952). Ibid., 1, 1313.(1959). Lancet, 1,
159.Rosenheim, M. L., and Spencer, A. G. (1956). Ibid., 2,
313.Sarre, H. (1959). Nierenkrankheiten. Stuttgart.Stanbury, S. W.
(1957). Brit. med. Bull., 13, 57.Stevenson, G. C., Jacobs, R. C.,
Ross, M. W., Collins, W. F.,
and Randt, C. T. (1959). Amer. J. Physiol., 197,
141.Winsbury-White, H. P. (1959). Brit. med. J., 1, 1001.
THE INNERVATION OF SKELETALMUSCLE*
BY
RUTH E. M. BOWDEN, D.Sc., M.B, B.S.Professor of Anatomy, Royal
Free Hospital School of
Medicine, London[WITH SPECTAL PLATE]
Although our knowledge of the pattern of innervationof skeletal
muscle and its functional significance is stillincomplete, there is
much that throws light on theproblem of control of postural and
so-called volitionalmovement. In this brief review attention will
be confinedto the nerve-supply of mammalian striated muscles andto
the connexions that these neurones make in thecentral nervous
system.
The Motor UnitThe smallest functional unit in a muscle is the
motor
unit, which consists of a single anterior horn cell, allits
processes and all the muscle fibres supplied by it.1 2It follows
that the strength of contraction depends, inpart, upon the size of
these motor units as well as onthe number brought into activity.3
Recent estimatesof the average number of muscle fibres in human
motorunits range from 5-9 for extrinsic ocular muscles4
to1,600-1,900 for one of the calf muscles.5 The endingsof the motor
nerves lose their myelin sheaths as theypierce the sarcolemma and
break up into fine brancheswhich rest in the shallow troughs that
form the sub-neural apparatus.6 These troughs are lined by
structureswhich are rich in cholinesterase,' the enzyme
thatdestroys acetylcholine, which is the chemical transmitterof the
nerve impulse across the neuromuscular junction(Special Plate, Figs
1 and 2). The motor end-platesmay be confined to a localized band
or be scatteredthroughout the substance of a muscle. In some
muscles
-for example, the extrinsic ocular and facial muscles-more than
one motor ending can be seen on a singlefibre; and these end-plates
are sometimes supplied bynerve fibres which approach from different
directions.8 9Since motor nerve fibres not only branch in the
mainnerve trunks3 and muscles,10 but also may form intra-muscular
plexuses (Special Plate, Figs. 3 and 4),histological preparations
of healthy muscles de not givereliable evidence about the parent
neurones of thedivergent fibres. For larger muscles in which
thephysical difficulty of examining long fibres throughouttheir
length is increased, there is only scant referenceto histological
evidence of multiple endings on a singlefibre, but
electrophysiological experiments have shownthat large motor nerve
fibres may innervate individualmuscle fibres at two or more
places.'lll 17 Thesemultiple endings may be derived from the
branches ofa single neurone or from several neurones. The
musclefibres of a unit may be in a compact bundle or theymay be
separated widely from each other and give riseto a diffuse motor
unit.8 10 14-17Whatever the spatial arrangement of the motor
units
within the muscle belly, however, there is anatomicaland
physiological evidence that the motor nerve cellssupplying
individual muscles are grouped together inthe brain-stem and spinal
cord, where they formnuclei.'8 26 Sharrard23 reconstructed the
lumbar andsacral segments to scale from serial sections of
threenormal spinal cords and of seven from patients whodied at
intervals ranging from three months to eightyears after the onset
of anterior poliomyelitis. Thereconstructions took no account of
the dendrites andaxons, but gave the relationships between the
bodies ofthe anterior horn cells. By correlating the
histologicaland clinical observations it was possible to
demonstratethe arrangement of the nuclei supplying
individualmuscles. These nuclei varied in size; some
occupiedseveral segments of the cord, while others were confinedto
a single segment, and functionally related nuclei wereanatomically
related to each other. Thus the nucleisupplying the tibialis
anterior and posterior muscleswere contiguous. Both these muscles
invert the foot;the former dorsiflexes the ankle-joint and the
latterplantar-flexes it.The lower motor neurones which activate the
motor
units form the " final common pathway "27 betweenthe central
nervous system and the muscles, and oncethey degenerate there is no
longer any effector pathwaybetween the central nervous system and
the musclefibres. Anterior horn cells have many dendrites whichlie
inside and outside the grey matter of the cord28 29and these
processes account for the greater part of thetotal surface area of
the cells,30 thus increasing the areafor contact with the " end
feet," or houton2s terminatux,of other neurones. It is through
these boiztonis th-tneurones from other levels of the nervous
system exerttheir influence on the lower motor neurone. It hasbeen
estimated that about 80% of the surface of ananterior horn cell is
related to the boutons,"1 at least2,000 of which may rest on the
surface of a single largecell in the cat.9 31 32 This does not
necessarily meanthat there are synaptic connexions with 2,000
otherneurones, for the nerve terminals branch, and many ofthe
boutons might be derived from one parent cell.The boutons vary in
size, but even with electronmicroscopy it has not proved possible
to distinguishA paper read to the Section of Anatomy and Physiology
at the
Annual Meeting of :he British Medical Association,
Birmingham,1958.
INNERVATION OF SKELETAL MUSCLE
between the excitatory and inhibitory endings thatphysiological
evidence has shown to exist 32-34
Conduction of ImpulsesMuscles are not only activated and
influenced by the
central nervous system, but they are also a source ofincoming
sensory stimuli. In a so-called motor nerve,for example, supplying
a limb or trunk muscle, at least30-40% of the fibres are sensory in
function and havetheir cell bodies in the dorsal root ganglia of
thesegmental nerves.1-3 The diameter of these afferentfibres ranges
from Ilt to 20[t,331 and the spectrum offibre sizes has three
peaks-Group I (12-20,t), Group II(4-12,A), and Group III (1-4[t).A5
36 The rate ofconduction of impulses varies from 1 to 125 metresper
second; and the full extent of the range is stressedwhen the rate
of conduction is given as about 2- miles(3.6 km.) per hour for
fibres of the smallest diameterand 280 miles (450 km.) per hour for
those of thelargest diameter.The endings of afferent fibres in
muscle vary enor-
mously in structure. The simplest are very fine,non-myelinated
nerve fibres with no specific endings.'-ome run in perivascular
plexuses (Special Plate, Fig. 5),o:3ers are found in connective
tissue, and it has been6uggested that these are pain fibres.37 3
More complexe:'capsulated Pacinian corpuscles (found also in skina
nd the mesentery) are described. These clearly arenot specific to
muscle; indeed, they are hard to find,37 9but when found they are
related to deep fascia, toaponeuroses, and to intramuscular vessels
and nerves(Special Plate, Fig. 6) ; they are supplied by large,
fast-conducting nerve fibres. In the mesentery they havebeen found
to respond to mechanical stimuli,40 but theirfunction in muscles
has yet to be analysed.The third variety of sensory ending is
presumably
stimulated by contraction of single muscle fibres.Large-,
medium-, and small-diameter nerve fibres formclaw-like endings,
spirals (Special Plate, Figs. 7, 8, 9),and basket-like networks
round individual muscle fibres.These forms of sensory ending have
been found inextrinsic ocular muscles41 and those of the face9 42
andlarynx.s'More is known about the fourth and fifth types of
sensory end-organ, which constitute built-in
automaticdamping-down and alerting mechanisms in striatedmuscles.44
Golgi tendon organs are inhibitory infunction,'546 and are arranged
in series with the musclefibres2 46 and are close to the insertions
of the spindleswhich constitute the alerting mechanism.37 In
tendonorgans the terminal branches of large, fast-conductingnerve
fibres lie at the musculo-tendinous junctions, andthe spray of
endings constitutes either the whole or partof a sensory unit
(Special Plate, Fig. 10). The size ofthese units and their relation
to individual motor unitsare unknown at present. The frequency with
which thetendon organs fire off impulses is a measure of thetension
developed within the muscle either by activecontraction or by
passive stretch.47 Through a reflexpathway in which there is an
interposed connectingneurone these organs exert an inhibitory
effect upon theanterior horn cells which innervate their own
andfunctionally related muscles.45 48
Muscle SpindlesThe muscle spindles are encapsulated, vary in
complexity, and have both motor and sensorynerves.' 37 49 50 In
man they have been found in muscles
of the face, larynx, palate, and infrahyoid region, inextrinsic
ocular muscles, in the tongue, in the musclesof trunk and limbs,
and in the striated upper third ofthe oesophagus." '5 So far they
have not beenfound in muscles of the pharynx or the anal
andurethral sphincters, which are still being investigated.They may
occur singly or in groups,`9 " I and in manare more numerous per
unit volume of muscle in theintrinsic muscles of the hand and in
muscles of the neckthan in the large limb muscles. 2 They are
probablymost plentiful in postural muscles, and nearly asnumerous
in flexors used for delicate movements.56Some indication of the
density of the spindle populationcan be gained from the fact that
47 were found in theinferior rectus and as many as 71 in the
superior obliquemuscle in man.52 In the cat 45 spindles were
foundin the soleus, 57 in the gastrocnemius, and 58 in thetibialis
anterior.39
Spindles are fusiform and may even be visible to thenaked
eye-for example, a single spindle from the vastusmedialis in a
rabbit measured 3.6 mm.,37 and in theintrinsic muscles of the human
hand they ranged from3 to 12 mm. in length. 3 The muscle fibres
inside aspindle are known as the intrafusal fibres and are oftwo
distinct sizes. It is suggested that the larger intra-fusal fibres
are of the fast twitch variety, and that thesmaller ones, like red
muscle fibres, are slow and tonicin their contractions50 (Special
Plate, Fig. 14). Thelargest extend from end to end of the spindle,
and maypass right through one into another, forming
tandemspindles53; the narrowest fibres are inserted into
thecapsule.37 The smallest spindles contain one fibre, butas many
as twelve have been reported.53 Intrafusalfibres differ from the
extrafusal ones in several respects(Special Plate, Figs. 11 and
12): they are usuallynarrower in diameter, stain differently, are
more faintlycross-striated, and have centrally placed nuclei,
which,in the larger intrafusal fibres, become so numeroushalfway
along the length of a spindle that this equatorialregion is also
known as the "nuclear bag" (SpecialPlate, Fig. 13). In the nuclear
bag the sarcoplasm ismuch paler and the cross-striation becomes so
faint that
BRITLSHMEDICAL JOURNAL
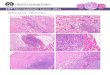
DESCRIPTION OF SPECIAL PLATEFiO. 1.-Motor end-plates in gracilis
of rhesus monkey.(Gold chloride stain.* x250.)FIG. 2.-Subneural
apparatus (marked by arrows) indiaphragm of rat. (Couteaux's
modification of Koellecs
stain. x I 10.)FIG. 3.-Intramuscular plexus formed by trigeminal
andfacial nerves in facial muscle of rhesus monkey. (Gold
chloride stain. x68.)FIG. 4.-Intramuscular plexus in rabbit's
interosseusmuscle. Note scattered motor end-plates. (Gold
chloride
stain. x 68.)FIG. 5.-Perivascular plexus in rabbit's
interosseusmuscle. a=nerve trunk from which a fibre (b) enters
the plexus. (Gold chloride stain. x 68.)FIG. 6.-Pacinian
corpuscles (P.C.) related to neuro-vascular bundle in cat's
interosseus muscle. A =an
arteriole. (Gold chloride stain. x30.)FIO. 7.-Transverse section
of sensory endings (a) inmedial rectus muscle of man. (Romanes's
silver stain.
x 200.)FIGS. 8 and 9.-Longitudinal section of spiral
sensoryendings in medial rectus muscle of man. (Romanes's
silver stain. x 200.)FIG. lO.-Golgi tendon organ in cat's
interosseus muscle.(Gold chloride stain. x 68. Preparation by Miss
B.
Higgs, B.Sc.)*For details of gold chloride stain see J. Anaf.
fLond.),
1956, 90, 217.
672 MARCH 5, 1960
MARCH 5, 1960 INNERVATION OF SKELETAL MUSCLE BRrrUs 673MEDICAL
JOURNAL
in some histological preparations it appears to be lost.In the
narrowest fibres the nuclei form a chain in theequatorial
region.50The capsule may consist of a single layer of
connective
tissue or of several layers (Special Plate, Figs. 14 and 15).It
has been suggested that the more numerous the layersof the capsule
the greater its protective function, but itis possible that this
layering is a means of grading theresponse or of reducing the
sensitivity of the sensoryendings. 5 Since the spindles lie in
parallel with themuscle fibres, they are extended when the
muscleslengthen and are relaxed when they shorten; but asthe
intrafusal fibres have a motor nerve supply theirtension can be
altered by contraction of the intrafusalfibres themselves.
Innervation of Muscle SpindlesThe pattern of innervation of the
spindles is still being
studied, but certain facts are well established. Thereare
several motor end-plates on each pole of an intra-fusal fibre37 53
(Special Plate, Fig. 16), and the nervefibres which supply them are
slow-conducting (20-44metres per second) gamma fibres of 3-8 pt in
diameter.57 58These arise from the small cells of the anterior
horns ofthe cord, leave by the anterior roots of spinal nerves,and
constitute about one-third of the total number ofmotor fibres
destined for skeletal muscle.3 17 The smallmotor fibres branch and
therefore form spindle motorunits (Special Plate, Fig. 17). A
single nerve fibre andits branches may supply each pole of one
intrafusalfibre, and also may innervate other fibres of the sameand
different spindles.37 39 It is also known that thefibres of a
single spindle may be innervated by morethan one spinal segment.39
One pole of a spindle mayreceive additional larger, and therefore
more rapidlyconducting, motor fibres37 47 53 56 (Special Plate,
Fig. 19).
DESCRIPTION OF SPECIAL PLATEFIG. 11.-Part of muscle spindle in
cat's interosseusmuscle. Note the narrower and more darkly
stainingintrafusal muscle fibres. M.F. =the fine motor nervefibre
supplying two motor end-plates on a single musclefibre; S.E. = the
sensory endings supplied by a thick nervefibre. (Gold chloride
stain. x 68. Preparation by Miss
B. Higgs, B.Sc.)FIG. 12.-Longitudinal section of sartorius
muscle ofrhesus monkey, showing the polar region of a spindlewith
narrow, more palely staining muscle fibres.(Romanes's silver stain.
x 100.)FIG. 13.-The nuclear bag or equatorial region (a) of thesame
spindle as Fig. 12. Note b, the thick sensory nervefibre forming
part of the annulo-spiral ending. (x 182.)FIG. 14.-Transverse
section of medial rectus muscle(man). Note a, the single layer of
the sheath of a musclespindle with seven intrafusal fibres of large
and small
diameter. (Romanes's silver stain. x 200.)FIo. 15.-Transverse
section of cat's soleus. Notelayering of capsule (a) of spindle.
(van Gieson's stain.
x152.)FIG. 16.-Multiple motor end-plates (marked by arrows)on
intrafusal muscle fibres in rabbit's interosseus muscle.(Gold
chloride stain. x 178.)FIG. 17.-Gamma motor fibre (3.8 p.) dividing
at a in thespindle shown in Figs. 12 and 13. Four branches couldbe
found by focusing up and down; only three show in
the plane of the photograph. (X200.)FIG. 18. Elaborate
annulo-spiral ending in large spindlein rabbit's tibialis anterior.
(Gold chloride stain. x 173.)FIG. 19.-Spindle in rabbit's
interosseus muscle. Note(a) simple annulo-spiral ending; (b)
medium-sized motor
fibre ending at c. (Gold chloride stain. x 100.)FIG. 20.-Part of
a spindle in sartorius muscle of rhesusmonkey. Note (a) an
intramuscular nerve trunk supply-ing extrafusal motor end-plates
(b) with medium-sizedfibres and (c) the large-diameter fibres
forming theannulo-spiral endings. (Gold chloride stain. x200.)
C
However, this is denied by some workers,58 and thesignificance
of the observation is obscure. Although themotor supply of both
intrafusal and extrafusal fibresmay run in the same intramuscular
nerve trunk (SpecialPlate, Fig. 20), there is some doubt whether
one axonever supplies both types of muscle fibre.The sensory fibres
have their cells of origin in the
dorsal root ganglia of the segmental nerves, and theirperipheral
endings are either at or near the equatorialregion. The most
elaborate annulo-spiral or primaryendings are formed by the largest
myelinated fibres(12-20 IL diameter), which are wound round the
equatorof the intrafusal fibres (Special Plate, Figs. 18 and
20).Other more diffuse flower-spray or secondary endingsare formed
by medium-sized fibres, and there may beone, two, or none in the
spindle. When annulo-spiraland flower-spray endings are found
together the latterlie to the side of the nuclear bag.37 50 Both
types ofending are stimulated by deformation due to a changein the
shape of the contractile intrafusal fibres.A constant stream of
impulses, of frequencies ranging
from 10 to 60 per second, are transmitted by the }ymotor fibres
to the intrafusal fibres. These contract,and, while they add
nothing to the tension of the muscleas a whole, they prime the
sensory endings to fire offwhen a critical level of distortion is
reached. Whenstimulated effectively the large fibres conduct
impulsesback to the spinal cord through the dorsal roots at
ratesranging from 60 to 125 metres per second (216-450 km.or
135-280 miles per hour). The central processes ofthese nerves end
directly on the anterior horn cellssupplying their own and
functionally related muscles.These sensory fibres constitute the
afferent arc of thestretch reflex. There is the minimum synaptic
delay, sincethere is no internuncial or connector neurone in
thisreflex pathway. By means of a branch working throughan
internuncial neurone these same afferent fibresinhibit the
contraction of their antagonistic muscles.33 45 59The activity of
spindles is not only enhanced by activitywithin muscles, but also
by impulses carried to theanterior horn cells from higher levels of
the centralnervous system.Y62 It is damped down or inhibitedby the
cut-out mechanism provided by the Golgi tendonorgans45 as well as
by inhibitory impulses from otherlevels of the nervous system.61
62
Sensory Supply of Trunk and Limb MusclesThe sensory supply of
trunk and limb muscles
provides the afferent arc of a spinal reflex pathway forthe
adjustment of the activity of the lower motorneurones. The precise
afferent pathways for musclessupplied by cranial nerves are not yet
known, but it isclear that these lower motor neurones in the
brain-stemas well as those in the spinal cord are under the
influenceof higher levels of the nervous system. However,
carefulsearch has so far failed to demonstrate any
afferentprojection pathway from the muscles to the cerebralcortex.
This, at first sight, is surprising in the light ofthe many
movements which require delicate gradationof muscular activity. On
the other hand, the capsulesof joints have a rich sensory
innervation, and a projec-tion pathway from joints to cerebral
cortex has beenfound in cats.63-65 Therefore, since the position
ofjoints is signalled to the cortex, and this position, in
anymovement, is the result of muscular activity, there is,in
effect, an indirect cortical representation of themuscles.
Grateful thanks are due to the National Foundation forInfantile
Paralysis Inc.; to my colleague Dr. Charles
674 MARCH 5, 1960 INNERVATION OF SKELETAL MUSCLE
MBDICALJOUwDownman for helpful criticism; to Mr. William
Matthewsfor technical assistance ; to Miss F. Ellis, Miss J.
Cherry,and Messrs. J. M. Crane and V. Willmott for thephotography;
and to the Editor of the Journal of Anatomyfor permission to use
Fig. 20.
REFERENCES1 Sherrington, C. S., J. Physiol. (Lond.) 1894 17,
211.2 Denny-Brown, D., Proc. roy. Soc. B, i929, i04, 252.Eccles, J.
C., and Sherrington, C. S., ibid., 1930, 106, 326.Torre, M.,
Schweiz. Arch. Neurol. Psychiat., 1953, 72, 362.Feinstein, B.,
Lindegard, B., Nyman, E., and Wohlfart, G.,Acta anat. (Basel),
1954, 23, 127.
Couteaux, R., Thesis for Doctorate of University of
Paris.Therien Freres, Montreal, 1947.
Koelle G. B., and Friedenwald, J. S., Proc. Soc. exp.
Biol.(N.Y.), 1949, 70, 617.' Feindel, W., Hinshaw, J. R., and
Weddell, G., J. Anat. (Lond.),
1952, 86, 35.1 Bowden, R. E. M., and Mahran, Z. Y., ibid., 1956,
90, 217.Cooper, S., J. Physiol. (Lond.), 1929, 67, 1.Agduhr, E.,
Anat. Anz., 1919, 52, 273.
2 Jarcho, L. W., Eyzaguirre, C Berman, B., and Lilienthal,J.
L.,jun., Amer. J. Physiol., 1952, 168, 446.
Hunt, C. C., and Kuffler, S. W., J. Physiol. (Lond.), 1954,
126,293.
14 Sherrington C S ibid., 1892, 13, 621.Adrian, E. b., ibid.,
1925, 60, 301.
16 van Harreveld, A., Arch. neerl. Physiol., 1948, 28,
408.Kuffler, S. W., Hunt, C. C., and Quilliam, J. P., J.
Neutro-physiol., 1951, 14, 29.
Romanes, G. J., J. Anat. (Lond.), 1941, 75, 145.- ibid., 1941,
76, 112.20 ibid., 1942, 77, 1.-1 ibid., 1945, 79, 145.22 - J. comp.
Neurol., 1951, 94, 313.
23 Sharrard, W. J. W., J. Bone Jt Surg., 1955, 37B, 540.21
Sprague, J. M., Amer. J. Anat., 1948, 82, 1.25 Walls, E. W.,
Lancet, 1940, 1, 123.24 Warwick, R., J. comp. Neurol., 1953, 98,
449.27 Sherrington, C. S., Report of the 74th Meeting of the
British
Association, Cambridge, 1904, p. 728.28 Hoff, E. C., Proc. roy.
Soc. B, 1932, 111, 175.29 Barr, M. L., J. Anat. (Lond.), 1939, 74,
1.
Aitken, J. T., ibid., 1955, 89, 571.'l Wyckofi, R. W. G., and
Young, J. Z., Proc. roy. Soc. B, 1956,
144, 440.'2Young J. Z., Progress in Neurobiology, ed. J. A.
Kappers,
p. 81. Elsevier Publishing Co., Amsterdam, London, andNew York.
1956.
'3 Lloyd, D. P. C., J. Neurophysiol., 1946, 9, 421.'4 Eccles, J.
C., Harvey Lect., 1955-1956, 51, 1.85 Lloyd, D. P. C., and Chang,
H. T., J. Neurophysiol., 1948, 11,
199.s Rexed, B., and Therman, P. O., ibid., 1948, 11, 133."
Barker, D., Quart. J. micr. Sci., 1948, 89, 143.
Hunt, C. C., J. gen. Physiol., 1954, 38, 117." Hagbarth, K. E.,
and Wohifart, G., Acta anat. (Basel), 1952,
15, 85.Gray, J. A. B., and Malcolm, J. L., Proc. roy. Soc. B,
1950,137, 96.
41 Daniel, P., J. Anat. (Lond.), 1946, 80, 189.42 Kadanoff, D.,
Z. Mikr. anat. Forsch., 1956, 62, 1.4 Lucas Keene, M. F., J. Anal.
(Lond.), 1957, 91, 590; and41 personal communication.Liddell, E. G.
T., Proc. roy. Soc. Med., 1954, 47, 600.'5 Granit, R., J.
Neurophysiol., 1950, 13, 351.46 Fulton, J. F., and Pi-Sufier, J.,
Amer. J. Physiol., 1928, 83, 554." Matthews, B. H. C., J. Physiol.
(Lond.), 1933 78 1.48 Eccles, J. C., Fatt, P., and Landgren, S., J.
Neurophysiol._
1956, 19, 75.Tower, S. S., Brain, 1932, 55, 77.Boyd, r. A., J.
Physiol. (Lond.), 1958, 140, 14 P.
5 Cooper, S., ibid., 1953, 122, 193.52
- and Daniel, P. M., Brain, 1949, 72, 1.513 - - J. Physiol.
(Lond.), 1956, 133, 1 P.54Slawik, F. F., Anat. Anz., 1942, 93,
133.5 Merrillees, N. C. R., Sunderland, S., and Hayhow, W.,
Anal.
Rec., 1950, 108, 23.5 Cooper, S., personal communication.5
Leksell, L., Acta physiol. scand., 1945, 10, Suppl. 31.
Hunt, C. C., and Kuffier, S. W., J. Physiol. (Lond.), 1951,113,
283.
5 Eccles, J. C., Eccles, R. M., and Lundberg, A., ibid., 1957,
137,22.Granit, R., Job, C., and Kaada, B. R., Acta physiol.
scand.,1953, 27, 161.61 Eldred, E., Granit, R., and Merton, P. A.,
J. Physiol. (Lond.),1953, 122, 498.
2Granit, R., and Kaada, B. R., Acta physiol. scand., 1953,
27,130.42 Mountcastle, V. B., J. Neurophysiol., 1957, 20, 408.64-
Davies, P. W., and Berman, A. L., ibid., 1957, 20, 374.
*sBoyd, I. A., J. Physiol. (Lend.), 1954, 124, 476.
METABOLIC AND ENDOCRINE ASPECTSOF DIABETIC NEPHROPATHY
BY
M. NAGY EL MAHALLAWY, M.D.Assistanit Professor of Medicine
MOHAMED SADEK SABOUR, M.D., M.R.C.P.Ed.*Clinical Demonstrator of
Medicine
LAILA MOHAMED OSMAN, D.CI.Sc.Resident in Clinical
Laboratories
AND
SAMIR HANNA SADEK, D.CI.Sc.Clinical Demonstrator of Clinical
Chemistry
Fromn the Departments of General Medicine and RenalClinic, and
Clinical Pathology, A bbassia Faculty of
Medicine, Ein-Shanms University, Cairo, Egypt
In the past few years various metabolic and
endocrinedisturbances have been described in association withthe
specific vascular lesions of diabetes-namely,retinopathy and
nephropathy. It is claimed that manyof these disturbances have a
pathogenic relationshipwith the development of the vascular
complications.
Becker (1952) gave evidence of excessive adrenalcortical
function in diabetics with retinopathy andKimmelstiel-Wilson (K.W.)
lesions. He noted anapparent relationship between the
lipoid-ladenvacuolated cells in the zona fasciculata of the
adrenalcortex and the K.W. lesions in the kidney. A picturesimilar
to early diabetic retinopathy and the K.W. lesioncould be produced
in alloxan-diabetic rabbits by theinjection of corticotrophin or
cortisone. Becker, Allen,et al. (1954), Becker, Maengwyn-Davies et
al. (1954),and Maengwyn-Davies (1956) gave fturther evidence
ofexcessive production of adrenal glucocorticoids inpatients with
diabetic retinopathy.
Becker et al. (1953) also reported a disturbance ofvitamin B, 2
metabolism in patients with diabeticretinopathy. Using the vitamin
B12 excretion test, theyfound that diabetics with retinopathy
cannot retain thevitamin, while those with no retinopathy retain
most ofthe given,dose. However, Field et al. (1957) could notfind a
definite relationship between the development ofdegenerative
complications of diabetes and the excretionpatterns of the B
vitamins.A disturbed carbohydrate metabolism has been found
in patients with clinically evident K.W. syndrome. Thus,Berkman
et al. (1953, 1954), in a study of the serumpolysaccharides in
diabetics and non-diabetics, reportedan increase in the total serum
polysaccharides indiabetics with degenerative vascular
complications, thehighest level being observed in those with
clinicallyevident nephropathy (Adlersberg et al., 1956). It is
notclear whether this increase is due to the renalimpairment and is
thus the result of the diabeticnephropathy, or whether it precedes
the nephropathyand provokes the pathological lesions in the
kidneys.Mann et.al. (1949) have noted hypercholesterolaemia
in association with diabetic glomerulosclerosis. Later,*Part of
a thesis submitted by M. S. Sabour for the degree of
Doctor in Medicine.