Embed Size (px)
Citation preview

INFLUENCE OF CHRONIC CERVICAL VAGOTOMY
ON SALT GLAND SECRETION IN THE GOOSE
JOSEPH P. GILMORE. CATHLEEN GILMOR~. JOHN DIETZ AND
IRVING H. ZUCKER*
Department of Physiology and Biophysics. University of Nebraska College of Medicine Omaha. NB, U.S.A.
(Recrird 16 Seprenlhrr 1976)
Abstract-l. Experiments were carried out to determine the influence of chronic bilateral cervical vago- tomy on salt gland secretion in the goose.
2. In only 3 of 7 animals was salt gland secretion in response to hypertonic sodium chloride per- manently abolished following vagotomy.
3. Survival time following vagotomy ranged from 4. The tachycardia caused by hypertonic sodium
inhibition of a high cardio-vagal tone.
INTRODUCTION
Hanwell et al. (1972) demonstrated clearly that acute bilateral section or blockage of the cervical vagi in the goose abolished salt gland secretion in response to a hypertonic infusion of sodium chloride. Since vagal section below the heart had no effect on salt gland secretion and since the injection of a local anes- thetic into the pericardium abolished secretion. these workers concluded that the heart of the goose con- tains osmoreceptors whose axons traverse the cervical vagi. The present experiments were undertaken to determine if chronic bilateral cervical vagotomy would also prevent salt gland secretion in response to a saline load with the idea that in such a prep- aration compensatory mechanisms for modulating salt gland secretion may be uncovered.
MATERlALS AND METHODS
Domestic geese (Anser. ctnser) maintained on tap water and bird food and ranging in weight from 4 to 5 kg. were employed in all experiments. A catheter was inserted into a wing vein for infusing solutions and withdrawing blood samples and the animal then placed in a plastic restraining box and its head gently immobilized. Salt gland secretion was collected using the technique described by us pre- viously (Gilmore et al.. 1977). A control blood sample was obtained and 50 ml, of a five percent sodium chloride solu- tion was administered intravenously over a 2 min period and the salt gland secretion collected for four 10 min periods, at the end of which time a second blood sample was obtained. After determining this control response to hypertonic sodium chloride the animals were anesthe- tized using intravenous pentobarbital sodium (15 mg/kg). the neck plucked and clipped, both vagi isolated and a loose surgical tie put about each of them. The neck incision was then closed leaving a small opening in the incision
*With the assistance of Robert Huskey and William Tobin. .
Supported by: Nebraska Heart Grant-In-Aid Award and The University of Nebraska Medical Center Research Seed Grant.
6 to 73 days. chloride infusion appears to be the result of the
so that the vagi could be pulled out through the incision for subsequent sectioning. The area over the incision was bandaged and taped and the animals allowed to recover from the anesthetic. On the following day when the ani- mals were fully recovered, the tape and bandage were removed and the area anesthetized using a local anesthetic. The vagi were then gently pulled through the incision and rapidly sectioned. The animals were then forced to con- tinuously move about until they respired spontaneously. This took from 15 to 45 min. If this was not done the animals would often become apneic. In some animals the vagal sectioning was done in such a manner that approxi- mately 1 cm of each nerve was removed. Each animal was then tested on subsequent days as described above to determine the secretory response of the salt gland to an intravenous 59, sodium chloride solution.
The first test after vagotomy was usually done the day following vagotomy. The completeness of vagotomy was verified post mortem. Heart rate was obtained by auscul- tation, sodium by flame photometry. and the hematocrit using capillary tubes and a microcentrifuge. Statistical comparisons were obtained employing the Student’s r test for unpaired data.
RESULTS
Immediately following vagotomy. there were obvious signs of respiratory distress and vomiting usually occurred. One animal died 556 hr after vago- tomy but the cause of death was not determined. If an animal responded after we believed it was vagoto- mized we opened the original incision to verify that the nerves were sectioned. If there was any question concerning the completeness of vagotomy. the pro- cedure for vagotomy (application of surgical ties, etc.) as described in the materials and Methods, was repeated on another section of both vagi and the ani- mal again studied. If the animals developed a tachy- cardia, vagotomy was invariably verified subse- quently. If a section of the nerve was not removed the two ends of the nerve trunk usually rejoined within several days. An interesting observation that we have not pursued was that the birds usually showed a pronounced polydipsia shortly after vagotomy.
119

120 JOSEPH P. GILMORE, et cd.
Table I. Influence of vagotomy on the secretory response of the salt gland to intravenous hypertonic sodium chloride
Bird No.
1
2
3
4
5
6
I
Pre Vagotomy Post Vagotomy wt. Secretion (ml/IO min) exp. * Secretion (ml/l0 min)
(kg) S, Sz s3 s‘s day sz S3
4.1 2.5 3.x 3.6 3.7 3 0 0.5 0.5 10 3.1 0 1.1 I.8
5.0 1.7 2.7 2.x I 0 0.1 0 42 4.4 0 0 0
1.9 3.5 3.2 1 0 0 0 9 0 0 0
4.3 3.4 4.0 3.7 3.1 1 0 0 0 7 3.4 0 0 0.1
4.4 2.4 4.6 4.2 I 0 0 0 41 3.6 0.9 1.X 1.2
4.9 1.8 2.3 2.2 2.2 I 4.4 0.2 3.0 2.6 I3 3.6 0.5 2.0 13
4.9 1.8 2.2 2.0 2.2 3 4.7 0.3 0.4 0.2
s4
0.7 I .8
0
0.2 0
1.2 2.9 0.2
0
* Day following vagotomy that animal was tested. t 0.53 mg atropine administered i.v.
Table 1 shows the salt gland response to hypertonic sodium chloride for the seven animals studied prior and subsequent to bilateral cervical vagotomy. Of the 5 animals studied the day after vagotomy (day 1). 4 showed essentially no response while the fifth ani- mal (No. 6) responded to the same extent as prior to vagotomy and continued to do so through the 13th day following vagotomy. The only apparent effect of bilateral vagotomy was to delay the time that the maximal secretion rate was achieved. After the third collection on the 13th day, 0.53 mg of atropine was administered which almost immediately stopped se- cretion. This animal died later in the day. Bilateral cervical vagotomy was verified post mortem. Both animals that were first studied 3 days post vagotomy (Nos. 1 and 7) showed a greatly attenuated secretory response. One died before it could be tested a second time (No. 7) while the other showed even a greater response (but delayed) when tested on the 10th day post vagotomy. Three animals (Nos. 2,3, and 4) never showed salt gland secretion when tested with S;/ sodium chloride following vagotomy. One of these (No. 2) was studied as late as 42 days after vagotomy and the other two at 7 and 9 days. Animal No. 5 was the most interesting one studied. The salt gland did not secrete the day following vagotomy. However, during 5 subsequent tests done 9941 days after vago- tomy, salt gland secretion occurred when hypertonic saline was administered despite the fact that during this period we bilaterally removed a 1 cm section of both vagi on day 24 while on day 38 did a complete examination of the neck, sectioning all neural struc- tures. On day 56 post vagotomy this animal was given 0.53 mg atropine sulfate before the hypertonic sodium chloride was infused. The salt gland did not secrete in response to the salt load. The animal was again tested on days 66,71 and 73 at which times salt gland secretion did not occur despite the fact that the ani- mal was not given additional atropine. This animal died on day 73 during surgical exploration to deter- mine the nature of an apparent hard mass in the area of the anterior air sac. The “mass” turned out to be a greatly distended esophagus filled with food.
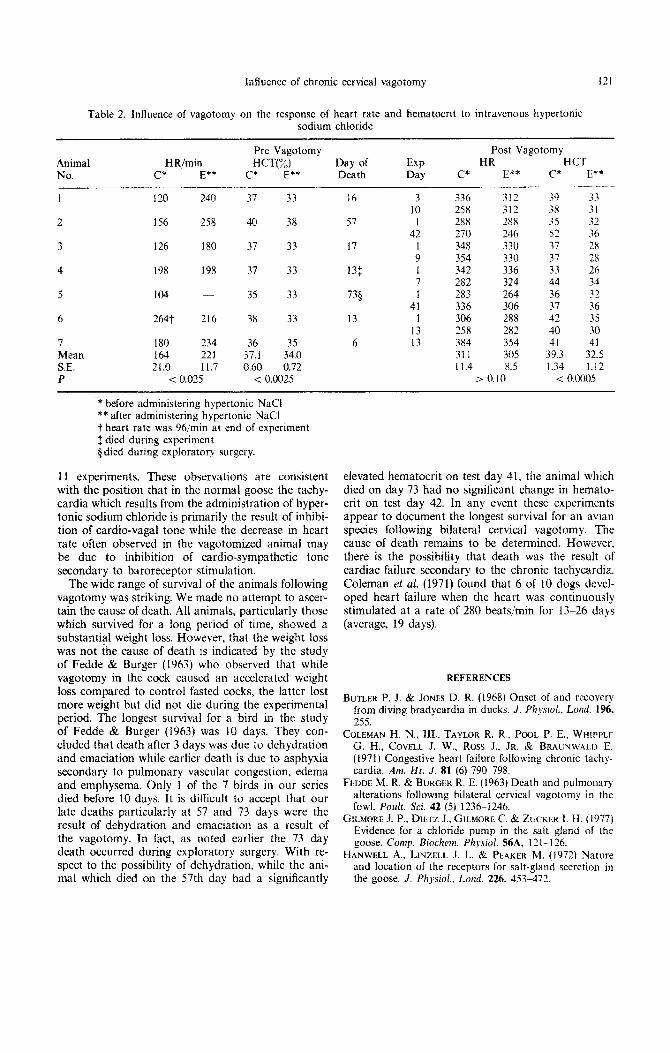
The most consistent consequence of vagotomy was the pronounced sustained tachycardia (Table 2). Prior to vagotomy hypertonic NaCl infusion increased heart rate significantly while following vagotomy the same infusion had no signi~cant effect on heart rate; in fact in individual animals it often decreased. A pro- nounced weight loss (Table 1) and hemo-concen- tration (Table 2) was usually seen in those animals which survived vagotomy for a period of time.
DISCUSSION
The experiments presented above show clearly that intact vagal pathways are not a necessary prerequisite for activating the salt gland in response to the infu- sion of hypertonic sodium chloride. Although in 6 of 7 animals secretion was prevented or greatly attenuated up to 3 days following bilateral cervical vagotomy, 3 of 7 animals showed a significant although somewhat attenuated response after this period. It is important to note further that 2 of these 3 animals showed little or no response through the 3rd day. It would appear, therefore, that in some birds non-vagal pathways are operative in controlling salt gland secretion or, alternatively, are uncovered as a result of vagotomy. The observation that atropine blocks secretion in vagotomized birds which respond to a hypertonic sodium chloride load shows that even in vagotomized responding birds, neural pathways are involved.
Although in the one reported experiment of Han- well et al. (1972) bilateral vagotomy had no influence on heart rate of the conscious goose. our studies clearly document that vagotomy causes a substantial tachycardia, indicating that the vagus exerts a sub- stantial cardio-inhibitory intlucncc in the goose as it does in the duck (Butler & Jones. 1968). Prior to vagotomy. the administration of hypertonic sodium chloride produced a substantial tachycdrdia if resting heart rate was not high. Following vagotomy when resting heart rate was substantially elevated, the ad- ministration of hypertonic sodium chloride had little effect on heart rate or caused a decrease in 7 of the
Influence of chronic cervical vagotomy 121
Table 2. Influence of vagotomy on the response of heart rate and hematocrit to intravenous hypertonic sodium chloride
Animal No.
I
2
3
4
5
6
7 Mean SE. P
HR/min c* E**
120 240
156 258
126 180
198 198
104
2647 216
180 234 164 221 21.0 11,7
< 0.025
Pre Vagotomy Post Vagotomy HCT(%) Day of EXP HR HCT
c* E** Death Day c* E** C* E**
37 33 16 3 336 312 39 33 10 258 312 38 31
40 38 57 I 288 288 35 32 42 270 246 52 36
37 33 17 ; 348 330 37 28 354 330 37 28
37 33 131 I 342 336 33 26 7 282 324 44 34
35 33 735 1 2x3 264 36 32 41 336 306 37 36
38 33 13 1 306 288 42 35 13 258 282 40 30
36 35 6 13 384 354 41 41 37.1 34.0 311 305 39.3 32.5 0.60 0.72 11.4 8.5 1.34 1.12
< 0.0025 > 0.10 < 0.0005
* before administering hypertonic NaCl ** after administering hypertonic NaCl t heart rate was 96/min at end of experiment $ died during experiment p died during exploratory surgery.
11 experiments. These observations are consistent with the position that in the normal goose the tachy- cardia which results from the administration of hyper- tonic sodium chloride is primarily the result of inhibi- tion of cardio-vagal tone while the decrease in heart rate often observed in the vagotomized animal may be due to inhibition of ~rdio-sym~thetic tone secondary to baroreceptor stimulation.
The wide range of survival of the animals following vagotomy was striking. We made no attempt to ascer- tain the cause of death. All animals, particularly those which survived for a long period of time, showed a subs~nti?l weight loss. However, that the weight loss was not the cause of death is indicated by the study of Fedde & Burger (1963) who observed that while vagotomy in the cock caused an accelerated weight loss compared to control fasted cocks, the latter lost more weight but did not die during the experimental period. The longest survival for a bird in the study of Fedde & Burger (1963) was 10 days. They con- cluded that death after 3 days was due to dehydration and emaciation while earlier death is due to asphyxia secondary to pulmonary vascular congestion, edema and emphysema. Only 1 of the 7 birds in our series died before 10 days. It is difficult to accept that our late deaths particularly at 57 and 73 days were the result of dehydration and emaciation as a result of the vagotomy. In fact, as noted earlier the 73 day death occurred during exploratory surgery. With re- spect to the possibility of dehydration, while the ani- mal which died on the 57th day had a significantly
elevated hematocrit on test day 41, the animal which died on day 73 had no significant change in hemato- crit on test day 42. In any event these experiments appear to document the longest survival for an avian species following bilateral cervical vagotomy. The cause of death remains to be determined. However. there is the possibility that death was the result of cardiac failure secondary to the chronic tachycardia. Coleman et al. (1971) found that 6 of 10 dogs devel- oped heart failure when the heart was continuously stimulated at a rate of 280 beats/min for 13-26 days (average, 19 days).
REFERENCES
BUTLER P. J. & JONES D. R. (1968) Onset of and recovery from diving bradycardia in ducks. J. Physiol., Land. 196. 255.
COLEMAN H. N., III.. TAYLOR R. R., POOL P. E., WHIP~LE G. H., COVELL J. W., Ross J., JR. & BRAUNWALD E. (1971) Congestive heart failure following chronic tachy- cardia. Am. Ht. J. 81 (6) 790-798.
FEDDE M. R. & BURGER R. E. (1963) Death and pulmonary alterations following bilateral cervical vagotomy in the fowl. P&t. Sci. 42 (5) 1236-1246.
GILMORE J. P., DIETZ J., GILMOKE C. & ZUCKER I. H. (1977) Evidence for a chloride pump in the salt gland of the goose. Contp. Biochem. Phpsiol. %A, 121- 126.
HANWELL A., LINZELL J. L. & PEAKER M. (1972) Nature and location of the receptors for salt-gland secretion in the goose. J. Physiol., Lond. 226. 453-472.