Embed Size (px)
Citation preview

This article was downloaded by: [Laurentian University]On: 10 October 2014, At: 03:31Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Transactions of the American Fisheries SocietyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/utaf20
Influence of Sex and Reproductive Status on SeasonalMovement of Lake Sturgeon in Namakan Reservoir,Minnesota–OntarioStephanie L. Shaw a f , Steven R. Chipps b , Steve K. Windels c , Molly A. H. Webb d & DarrylT. McLeod ea Department of Natural Resource Management , South Dakota State University , SNP Box2140B, Brookings , South Dakota , 57007 , USAb U.S. Geological Survey, South Dakota Cooperative Fish and Wildlife Research Unit,Department of Natural Resource Management , South Dakota State University , SNP Box2140B, Brookings , South Dakota , 57007 , USAc Voyageurs National Park , 3131 Highway 53, International Falls , Minnesota , 56649 , USAd U.S. Fish and Wildlife Service, Bozeman Fish Technology Center , 4050 Bridger CanyonRoad, Bozeman , Montana , 59715 , USAe Ontario Ministry of Natural Resources, Fort Frances District , 922 Scott Street, FortFrances , Ontario , P9A 1J4 , Canadaf School of Forest Resources and Conservation , University of Florida , 7922 North West 71stStreet, Gainesville , Florida , 32653 , USAPublished online: 04 Dec 2012.
To cite this article: Stephanie L. Shaw , Steven R. Chipps , Steve K. Windels , Molly A. H. Webb & Darryl T. McLeod (2013)Influence of Sex and Reproductive Status on Seasonal Movement of Lake Sturgeon in Namakan Reservoir, Minnesota–Ontario,Transactions of the American Fisheries Society, 142:1, 10-20, DOI: 10.1080/00028487.2012.720625
To link to this article: http://dx.doi.org/10.1080/00028487.2012.720625
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Transactions of the American Fisheries Society 142:10–20, 2013C© American Fisheries Society 2013ISSN: 0002-8487 print / 1548-8659 onlineDOI: 10.1080/00028487.2012.720625
ARTICLE
Influence of Sex and Reproductive Status on SeasonalMovement of Lake Sturgeon in Namakan Reservoir,Minnesota–Ontario
Stephanie L. Shaw*1
Department of Natural Resource Management, South Dakota State University, SNP Box 2140B,Brookings, South Dakota 57007, USA
Steven R. ChippsU.S. Geological Survey, South Dakota Cooperative Fish and Wildlife Research Unit,Department of Natural Resource Management, South Dakota State University, SNP Box 2140B,Brookings, South Dakota 57007, USA
Steve K. WindelsVoyageurs National Park, 3131 Highway 53, International Falls, Minnesota 56649, USA
Molly A. H. WebbU.S. Fish and Wildlife Service, Bozeman Fish Technology Center, 4050 Bridger Canyon Road,Bozeman, Montana 59715, USA
Darryl T. McLeodOntario Ministry of Natural Resources, Fort Frances District, 922 Scott Street, Fort Frances,Ontario P9A 1J4, Canada
AbstractWe evaluated the influence of sex and reproductive condition on seasonal distribution and movement patterns of
Lake Sturgeon Acipenser fulvescens in Namakan Reservoir, Minnesota–Ontario. Blood samples were collected from133 Lake Sturgeon prior to spawning and plasma concentrations of testosterone and estradiol-17ß were analyzed usingradioimmunoassay. Steroid concentrations were used to determine sex and the reproductive stage of each sturgeon.A subset of 60 adults were implanted with acoustic transmitters prior to spawning in 2007 and 2008. Movement wasmonitored using an array of 15 stationary receivers covering U.S. and Canadian waters of Namakan Reservoir and itstributaries. Of the monitored sturgeon, there was no significant difference in the minimum distance traveled betweensexes or among seasons. Site residency did not differ between sexes but differed significantly among seasons, and LakeSturgeon of both sexes had higher residency during winter (mean = 24 d). Five females implanted with transmitterswere characterized as presumed reproductive and 14 as nonreproductive based on plasma steroid concentrations.In general, movement patterns (i.e., migration) of presumed reproductive females corresponded positively withavailability of spawning habitat in tributaries. Moreover, presumed reproductive females traveled greater distancesthan nonreproductive females, particularly during prespawn, spawning, and fall time periods. Distance traveled bypresumed reproductive females was highest in the fall compared with other seasons and may be linked to increasedenergy requirements during late oogenesis before spawning in spring. Combining movement data with information
*Corresponding author: [email protected] address: School of Forest Resources and Conservation, University of Florida, 7922 North West 71st Street, Gainesville, Florida
32653, USA.Received October 27, 2011; accepted August 6, 2012Published online December 4, 2012
10
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
3:31
10
Oct
ober
201
4

SEASONAL MOVEMENT OF LAKE STURGEON 11
on Lake Sturgeon reproductive status and habitat suitability provided a robust approach for understanding theirseasonal migration patterns and identifying spawning locations.
Lake Sturgeon Acipenser fulvescens populations have de-clined across much of their range in North America owing tofactors that include overharvest, impoundment of rivers, and wa-ter pollution (Harkness and Dymond 1961; Auer 1996, 2004).Commercial fishing harvest in the late 1800s and early 1900s de-pleted Lake Sturgeon stocks across North America (Auer 2004).The boom of riverside industry during the same period hinderedpopulation recovery by restricting access to and altering qualityof spawning and nursery habitat (Auer 1999, 2004). Moreover,the life history characteristics of the species, (e.g., late age atreproductive maturity) make Lake Sturgeon vulnerable to over-harvest and slow to recruit to the spawning population (Auer2004). Conservation of Lake Sturgeon populations requires ef-fective management and long-term monitoring. Thus, informa-tion about the abundance, spawning behavior (e.g., seasonalmovement), and reproductive status of Lake Sturgeon providesuseful information for species recovery.
Previous studies have used telemetry to identify habitat useand migration patterns of Lake Sturgeon but often do not ac-count for differences in movement due to sex and reproductivestructure. Research in Ontario, Minnesota, and Michigan usedtelemetry data to define seasonal movement and habitat use byLake Sturgeon (Rusak and Mosindy 1997; McKinley et al. 1998;Auer 1999; Adams et al. 2006); differences in habitat use weregenerally attributed to seasonal changes in water temperatureand behavior related to spawning, foraging, or overwintering.However, there is little information on differences in movementand habitat use related to reproductive strategy. Lake Sturgeonexhibit differing reproductive strategies based on sex. Femalesmature later at 20–25 years of age and have a longer spawn-ing periodicity of 4–9 years, whereas males mature earlier at8–12 years of age and have a shorter spawning periodicity (1–5 years; Craig et al. 2005; Peterson et al. 2007). Thus movementpatterns and habitat use are likely to differ based on sex andreproductive stage. Few studies have combined telemetry datawith reproductive status to examine movement patterns in re-lation to age, sex, and reproductive maturity (McKinley et al.1998).
The reproductive structure of Lake Sturgeon populationsincludes information such as sex ratio, stage of reproductivedevelopment, and proportion of the population capable of re-producing each spawning season and can provide importantinformation to fishery managers (McLeod 1999; Craig et al.2005). McKinley et al. (1998), for example, used information onplasma steroid concentrations to evaluate seasonal reproductivepatterns of male and female Lake Sturgeon from the MattagamiRiver, Ontario. Male and female Lake Sturgeon were shownto have divergent patterns of testicular or ovarian development
after spawning. In male sturgeon, reproductive hormone (i.e.,testosterone) levels and gonadosomatic index (GSI) returnedto prespawning levels quickly (∼1 month), whereas femalesteroid concentrations and GSI remained low after spawningand showed no measureable increase until fall (McKinley et al.1998). Because sex and reproductive status cannot be reliablyidentified externally in sturgeons (Webb and Doroshov 2011),the use of plasma steroid concentrations offers a pragmatic ap-proach to evaluate the reproductive structure of sturgeon species(Webb et al. 2002; Craig et al. 2005; Webb and Erickson 2007;Stahl et al. 2009). Linking reproductive attributes such as sexand stage of maturity with information gathered from teleme-try studies provides a robust approach for verifying spawningareas, evaluating seasonal migration behavior, and quantifyingsex- and maturity-related movement patterns. In this study, wecombined seasonal movement data acquired through acoustictelemetry with information on sex and stage of maturity forLake Sturgeon in Namakan Reservoir, Minnesota–Ontario. Ourobjectives were to evaluate the influence of reproductive at-tributes (sex and stage of maturity) on movement patterns ofLake Sturgeon and identify important spawning areas in theNamakan Reservoir system. In an effort to corroborate thesefindings, we calculated a habitat suitability index (HSI) for po-tential spawning areas and related this to movement patterns ofpresumed reproductive fish.
STUDY SITENamakan Reservoir (26,700 ha) is located along the
Minnesota–Ontario border between Rainy Lake and the Bound-ary Waters Canoe Area Wilderness and Quetico ProvincialPark (Figure 1). It includes five water bodies: Kabetogama,Namakan, Sand Point, Crane, and Little Vermilion lakes, andtheir associated tributaries. The Minnesota–Ontario border tra-verses Namakan, Sand Point, and Little Vermilion lakes. Thereservoir includes six main tributaries: the Vermilion, Loon,Redhorse, Namakan, Ash, and Moose rivers (Figure 1). TheNamakan River is the largest source of inflow to the VoyageursNational Park system and has a mean discharge of 109 m3/s(Kallemeyn et al. 2003). This inflow is located in Ontario at theeastern end of Namakan Lake (Figure 1). The Vermilion River,which empties into Crane Lake, is located at the southwesternend of Namakan Reservoir and has a mean discharge of 9 m3/s(Figure 1; Kallemeyn et al. 2003). The four remaining tribu-taries are smaller with their combined discharge contributingless than 20% of the inflow to the reservoir. Voyageurs NationalPark, established in 1975, encompasses Kabetogama Lake andthe U.S. waters of Namakan and Sand Point lakes. The Superior
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
3:31
10
Oct
ober
201
4

12 SHAW ET AL.
FIGURE 1. The Namakan Reservoir system, Minnesota–Ontario, showing the locations of 15 stationary, Vemco VR2W acoustic receivers to detect LakeSturgeon. Location names are summarized in Table 2.
National Forest borders Crane and Little Vermilion lakes onthe U.S. side of the border. Water levels are regulated by theInternational Joint Commission (IJC) through the InternationalRainy Lake Board of Control (McLeod and Debruyne 2009). In2000, the IJC implemented a hydrologic management regimethat more closely resembles natural flows for Namakan Reser-voir (Kallemeyn et al. 2003).
METHODSLake Sturgeon collection.—Fish were collected from 2007
through 2009 beginning shortly after ice-out and continuinguntil the onset of optimum Lake Sturgeon spawning temper-atures (13◦C, Harkness and Dymond 1961). Biologists fromVoyageurs National Park, the Ontario Ministry of Natural Re-sources, Minnesota Department of Natural Resources, the U.S.Forest Service and the U.S. Geological Survey participated insampling efforts. Lake Sturgeon were collected using multifila-ment gill nets (178-, 203-, 230-, 254-, 305- and 356-mm stretchmeshes) that measured 1.8 m high and varied in length from 30to 100 m. Gill nets were bottom set for approximately 20 h intributaries, embayments, or main-lake locations. Lake Sturgeonwere measured for fork length (FL) and girth (millimeters) andweighed to the nearest gram on a hand scale. A 2- to 3-cm sectionof the pectoral fin spine was obtained for age determination.
Reproductive condition.—The reproductive structure of LakeSturgeon in Namakan Reservoir was recently described by Shaw
et al. (2012). In brief, blood samples were obtained from the cau-dal vasculature of 133 prespawning Lake Sturgeon collected inNamakan Reservoir in 2008 (n = 46) and 2009 (n = 87). Plasmaextracted from the blood samples was analyzed for concentra-tions of estradiol-17β (E2) and testosterone (T) and used todifferentiate sex and classify Lake Sturgeon into reproductivestages as described in Webb et al. (2002). Steroid classifica-tions were validated using a subset of Lake Sturgeon that weresexed and staged by direct visual examination of gonads from asmall incision in the ventral side of the fish anterior to the pelvicgirdle. Of the four females and one male that were classifiedvisually, 100% were classified correctly using steroid concen-trations. Male Lake Sturgeon E2 concentrations did not exceed1.0 ng/mL and T concentrations were greater than 3.0 ng/mL.Males were differentiated into three reproductive stages; mi-totic, meiotic, and spermiating. The spermiating stage (T >
30.0 ng/mL, E2 < 1.0 ng/mL) included reproductive or ripemales capable of reproducing in the season sampled (Webbet al. 2002; Allen et al. 2009; Shaw et al. 2012). Testosteroneis a precursor to E2, and as a result, levels of both steroids varyin females depending on reproductive stage. In nonreproduc-tive females, concentrations of T are less than 3.0 ng/mL andincreasing concentrations of T and E2 are detected upon ini-tiation of vitellogenesis (Webb and Doroshov 2011). Femaleswere classified into three reproductive stages; previtellogenic,vitellogenic, and postvitellogenic. The postvitellogenic stage
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
3:31
10
Oct
ober
201
4

SEASONAL MOVEMENT OF LAKE STURGEON 13
(T > 17.0 ng/mL, E2 > 2.0 ng/mL) was the only reproduc-tive stage that included gravid females that would be capableof spawning in the season sampled (Allen et al. 2009; Shawet al. 2012). Furthermore, Shaw et al. (2012) showed that thereproductive status of female Lake Sturgeon was related to bodysize and girth of postvitellogenic females. Thus, to characterizereproductive status (i.e., presumed reproductive or nonreproduc-tive) of female sturgeon that were implanted with a transmitterbut did not have a blood sample taken, we used weight andgirth as a metric of maturity (Shaw et al. 2012). Presumed re-productive females were defined as those females where weight(kilograms) was equal to or exceeded 2% of FL (millimeters),and girth (millimeters) was equal to or exceeded 40% of FL(Shaw et al. 2012). Nonreproductive females included all thosewith proportionally lower weight and girth in comparison withFL. Because there was no significant difference in size char-acteristics of male Lake Sturgeon of differing maturity stages(Shaw et al. 2012), we were unable to assign a reproductivestatus to male fish and thus excluded them from our analysis.
Acoustic telemetry.—Sixty adult Lake Sturgeon were im-planted with Vemco V16 acoustic transmitters (Vemco–AmirixSystems, Halifax, Nova Scotia) in May 2007 (n = 30) and2008 (n = 30). All Lake Sturgeon implanted with transmit-ters were verified as adults from aging using pectoral spines(range, 17–70 years; Shaw et al. 2012). Of the 60 sturgeon im-planted with acoustic transmitters, 26 (43%) were collected inNamakan Reservoir by a multiagency effort and 34 (57%) werecollected in the Namakan River by the Ontario Ministry of Natu-ral Resources (Table 1). The transmitters were single frequency(69 kHz) coded tags, 68 × 16 mm in size, and weighed 10 gin water (McLeod and Debruyne 2009). Individual transmitterswere uniquely coded and emitted a signal every 60–120 s andhad an operating life of 2,190 d (Welsh and McLeod 2010).
All surgeries performed on fish to implant the V16 trans-mitters were conducted by an experienced biologist (D.T.M.)and followed protocols similar to Hart and Summerfelt (1975)and Adams et al. (2006). Prior to surgery, Lake Sturgeon wereheld for up to 1 h in a 350-L tank filled with approximately300-L of water and 1.5 kg of salt (NaCl). Surgical instrumentsand transmitters were disinfected in a solution of Germiphenegermicidal concentrate (50 mL) and distilled water (4 L). Toimplant an acoustic tag, the Lake Sturgeon was placed ventral
TABLE 1. Number (n), mean fork length (FL), and weight of Lake Sturgeoncollected and implanted with acoustic transmitters in Namakan Reservoir andthe Namakan River in the springs of 2007 and 2008. Values in parenthesesrepresent SE.
Capture 2007 2008 Total Mean FL Mean weightlocation n n n (mm) (kg)
NamakanReservoir
0 26 26 1,218 (31.1) 16.1 (1.2)
NamakanRiver
30 4 34 1,095 (28.2) 11.4 (1.0)
side up in a canvas cradle suspended in the holding tank whilewater was flushed over the gills using a small pump and hose.A sterile, fenestrated polylined towel (Convertors) covered theventral surface of the sturgeon. Using a surgical scalpel, a 3–5 cm incision was made on the ventral surface approximately1 cm off the midline and 3–4 cm anterior to the pelvic girdle.If gonads were readily visible during surgery, the sex and stageof maturity were noted then the transmitter was inserted intothe peritoneal cavity. The peritoneum and muscle tissue wereclosed with a continuous modified Cushings suture technique(3-0 Ethicon PDS II, 12.7-mm CT-2 needle). To close the skin,five simple interrupted sutures were used (2-0 Ethicon Prolene,12.7-mm SH needle). Lake Sturgeon were monitored for ap-proximately 1 h after surgery to assess recovery. Fish handlingand care protocols followed guidelines established by the Amer-ican Fisheries Society (Nickum et al. 2004).
We used a passive telemetry array consisting of 15 stationaryVemco VR2W receivers (Vemco–Amirix Systems) positionedthroughout the main reservoir (Figure 1). We chose receiver lo-cations that would most probably maximize detection efficien-cies based on a manufacturer-recommended maximum range of0.5 km. Thus, locations such as interlake narrows, bays, andtributary mouths were selected throughout the reservoir as de-ployment sites for the receivers. The receivers were attached toa line with a float to keep the receiver suspended approximately1 m off the bottom. Receiver lines were anchored to the bottomusing cement blocks, and anchor lines were attached to a pointon the nearest shoreline. Float lines were shortened dependingon the depth of the water so that floats would remain suspendedapproximately 1 m below the water line, and the anchor lineswere camouflaged on the shore to discourage tampering. Tem-perature loggers (HOBO U22-0001 Water Temperature Logger,Onset Computer Corporation) were attached to the float line ofeach receiver, approximately 1.5 m off the bottom, to recordtemperature once every 4 h. Receivers were range-tested in thesummer of 2008 to determine the detection range using proto-cols established by the manufacturer (Shaw et al. 2012). Rangetesting confirmed that all receivers had a maximum detectionrange of at least 0.5 km. Complete detection efficiency (100%)covered an area from bank to bank in all but two receiver loca-tions, Smuggler’s Point and Larkin Island (Figure 1; Table 2).
Data from the receivers were used to analyze seasonal dis-tribution, site residence, and migration patterns. The degree ofsite residency was defined as the total amount of time (days)an individual Lake Sturgeon spent within each individual re-ceiver’s range (Lowe et al. 2006). We summarized the totalnumber of Lake Sturgeon detected at each receiver. Individualfish detections were exported from the Vemco VUE softwareinto a Microsoft Excel file and summarized by season for eachLake Sturgeon. We used water temperature data from NamakanReservoir to characterize fish movement during spring (pres-pawning and spawning), summer (postspawn), fall, and winter(Figure 2; Rusak and Mosindy 1997). The spring prespawningperiod (April 16–June 4) started as water temperatures increased
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
3:31
10
Oct
ober
201
4

14 SHAW ET AL.
TABLE 2. The total number of Lake Sturgeon detected at each receiver in Namakan Reservoir from May 15, 2008, to October 23, 2009. Also provided are thehabitat characteristics near each receiver location (∼0.5 km radius around each receiver). Receiver locations are shown on Figure 1.
Total number Summer waterLocation Lake Sturgeon Dominant Maximum depth velocity range
Receiver number detected substrate type (m) (m/s)
Loon River 1 3 Sand 2 0.5–1.0Little Vermilion Narrows 2 12 Silt 7 0.5–1.5Vermilion Gorge 3 8 Sand 4 0.75–1.5King William’s Narrows 4 16 Sand 5 1.0–1.5Grassy Bay 5 16 Silt 5 1.0–1.75Redhorse River Bay 6 20 Silt 5 1.0–1.75Namakan Narrows 7 25 Sand 10 1.0–2.0Namakan River 8 35 Silt 8 0.5–2.0Smuggler’s Point 9 25 35 1.0–2.0Blackstone Island 10 20 Bedrock–boulder 3 1.0–1.75Squaw Narrows 11 11 25 1.0–2.0Squirrel Narrows 12 3 Bedrock–boulder 10Moose River 13 0 Silt 2 0.75–1.5Kabetogama Narrows 14 16 Silt 15 1.0–2.0Larkin Island 15 11 15
FIGURE 2. Mean (solid line) and minimum and maximum (dashed lines) reservoir water temperatures recorded from temperature loggers attached to stationaryacoustic receivers. Dates are month/day/year. See text for explanation of seasonal time designations.
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
3:31
10
Oct
ober
201
4

SEASONAL MOVEMENT OF LAKE STURGEON 15
above 2◦C and before the onset of optimum Lake Sturgeonspawning temperatures (13◦C, Harkness and Dymond 1961).The spring spawning season (June 5–June 22) was defined as thetime during which water temperatures were within the range ofreported Lake Sturgeon spawning temperatures (i.e., 13–18◦C,Harkness and Dymond 1961). The summer postspawn periodbegan as temperatures exceeded 18◦C and continued until watertemperatures declined (June 23–September 2). We character-ized fall as the period with decreasing water temperatures fromSeptember 3 to December 11. Winter was characterized as theperiod corresponding to low, stable, water temperatures (0–2◦C;December 12–April 15).
The degree of site residency was summarized overall andby season for individual sturgeon. Upon entering the detectionrange of a receiver, events are logged every 1–3 min. A LakeSturgeon was considered to be within the receiver area untilthe time between detection events exceeded 10 min. Once thetime between detections exceeded 10 min the sturgeon wasconsidered to have left the area of the receiver and the event wasended. For each fish, total time (days) spent within a receiver’sdetection range was summarized by season and for the entirecensus period (May 2008–October 2009).
Individual Lake Sturgeon movements were used to estimateminimum distance traveled (MDT; Chateau and Wantiez 2007)and identify migration patterns (Peterson et al. 2007) using Ar-cMap 9.3 (ArcGIS ESRI, Redlands, California). A least-costdistance map of Namakan Reservoir was created in ArcMap inwhich any water was considered to be no cost to travel and allland was considered to be a barrier to Lake Sturgeon move-ment. The shortest water route between receiver locations wascalculated using the shortest path tool in the Spatial Analyst tool-bar. For each sturgeon, the paths it traversed between receiverswere summarized by season and for the entire time period (May2008–October 2009). Due to the distance between receiver lo-cations we were unable to characterize localized movement pat-terns. Thus, movement was characterized as the MDT aroundthe reservoir (i.e., seasonally or for the entire time series) andwas not an attempt to summarize a continuous movement pat-tern. If a Lake Sturgeon made repeated trips between two of thesame receivers during the time period being summarized, thatpath was only counted once.
A two-way ANOVA was used to examine the effect of sex(i.e., male or female) and season (i.e., spring prespawn, springspawn, summer postspawn, fall, and winter), on MDT and siteresidency (PROC MIXED, SAS 1999). We tested the hypothesisthat mean MDT and mean site residency of Lake Sturgeon wouldnot differ by sex or season. To conduct this analysis, we useddetections that occurred from the beginning of the fall of 2008(September) through the end of the summer of 2009 (August).During this time period, all Lake Sturgeon had been acousticallytagged for at least 4 months, which we assumed was a reasonabletime for fish to resume normal behavior (e.g., movement) aftertransmitter implantation. To evaluate seasonal effects, we used aTukey’s multiple comparison test to compare seasonal means. A
Student’s t-test was used to compare differences between sexes(SAS 1999).
The spawning migration of Lake Sturgeon is an importantcomponent of gonad maturation and can influence spawningsuccess (McKeown 1984; Auer 1996). We used a two-wayANOVA to determine the effect of female maturity level (i.e.,presumed reproductive or nonreproductive) and season on MDTand site residency (PROC MIXED, SAS 1999). We tested thenull hypothesis that MDT and site residency of Lake Sturgeonwould not differ by maturity level or season for female LakeSturgeon. We compared movement data between presumed re-productive and nonreproductive females using telemetry datafrom the fall 2008 through summer 2009, as previously de-scribed. If applicable, post hoc tests (e.g., Tukey’s honestly sig-nificant difference test or t-test) were used to evaluate seasonaldifferences in MDT or site residency.
Habitat suitability.—The presence of reproductively matureLake Sturgeon (i.e., postvitellogenic females and spermiatingmales) in tributaries was used as an indicator of potential spawn-ing areas within Namakan Reservoir. To corroborate these obser-vations, we calculated habitat suitability values for each of thesix main tributaries in the Namakan Reservoir using the habitatsuitability index (HSI) for spawning Lake Sturgeon developedby Threader et al. (1998) and later modified by Daugherty et al.(2008, 2009). Habitat characteristics (i.e., bottom water velocity,depth, and dominant substrate type) were measured in each trib-utary in the spring of 2009 using a systematic sampling design.A 1.5-MHz SonTek RiverCat acoustic doppler profiler (ADP)recorded water velocity to the nearest 0.1 cm/s and depth tothe nearest meter along transects perpendicular to the currentevery 100–150 m. Transects started at the mouth of the tributaryand continued upstream until we reached the first natural bar-rier (i.e., falls, rapids, or impassible low water zones). The ADPrecorded water velocity profiles in 0.5-m cells from the river bot-tom to 1.0 m below the surface. Lake Sturgeon are benthivorous(Scott and Crossman 1973); thus, only the bottom 0.5-m velocitymeasurements were used to estimate water velocity suitability.Substrate samples were collected along every other transect us-ing a petite ponar grab and characterized according to particlesize (Threader et al. 1998). Substrate samples were collected atthree points approximately equidistant across each transect.
Habitat data were imported into ArcGIS 9.3 (ArcGIS ESRI)in georeferenced point format. Spatially explicit maps of eachvariable were interpolated using inverse distance weighting(IDW) with a polyline barrier specific to each river. The IDWwas chosen as it generally provided the lowest root mean squareerror estimates after cross validation compared with otherinterpolation methods (i.e., kriging) indicating that the IDWinterpolation would probably provide more reliable estimatesfor unknown points (Tomczak 1998). Habitat models wereclassified to reflect habitat suitability values for spawning LakeSturgeon (Threader et al. 1998; Daugherty et al. 2008, 2009). Acomposite HSI model for each river was created by calculatingthe geometric mean of the HSI values from each individual
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
3:31
10
Oct
ober
201
4
16 SHAW ET AL.
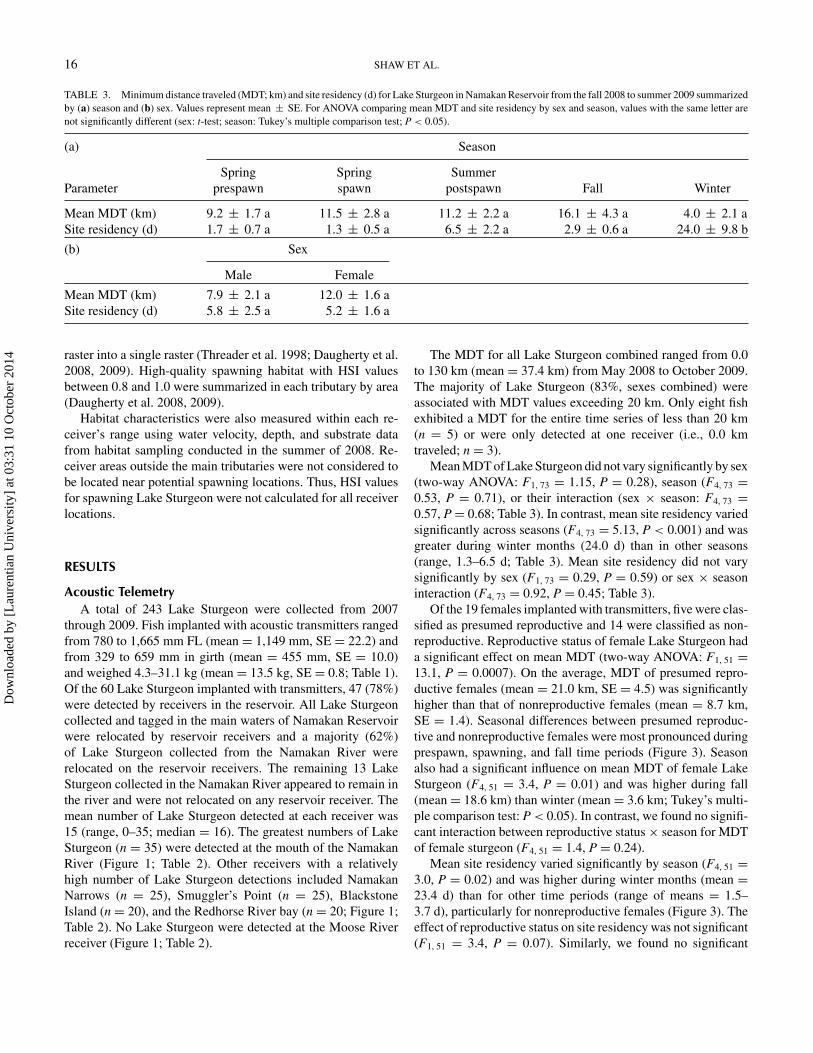
TABLE 3. Minimum distance traveled (MDT; km) and site residency (d) for Lake Sturgeon in Namakan Reservoir from the fall 2008 to summer 2009 summarizedby (a) season and (b) sex. Values represent mean ± SE. For ANOVA comparing mean MDT and site residency by sex and season, values with the same letter arenot significantly different (sex: t-test; season: Tukey’s multiple comparison test; P < 0.05).
(a) Season
Spring Spring SummerParameter prespawn spawn postspawn Fall Winter
Mean MDT (km) 9.2 ± 1.7 a 11.5 ± 2.8 a 11.2 ± 2.2 a 16.1 ± 4.3 a 4.0 ± 2.1 aSite residency (d) 1.7 ± 0.7 a 1.3 ± 0.5 a 6.5 ± 2.2 a 2.9 ± 0.6 a 24.0 ± 9.8 b
(b) Sex
Male Female
Mean MDT (km) 7.9 ± 2.1 a 12.0 ± 1.6 aSite residency (d) 5.8 ± 2.5 a 5.2 ± 1.6 a
raster into a single raster (Threader et al. 1998; Daugherty et al.2008, 2009). High-quality spawning habitat with HSI valuesbetween 0.8 and 1.0 were summarized in each tributary by area(Daugherty et al. 2008, 2009).
Habitat characteristics were also measured within each re-ceiver’s range using water velocity, depth, and substrate datafrom habitat sampling conducted in the summer of 2008. Re-ceiver areas outside the main tributaries were not considered tobe located near potential spawning locations. Thus, HSI valuesfor spawning Lake Sturgeon were not calculated for all receiverlocations.
RESULTS
Acoustic TelemetryA total of 243 Lake Sturgeon were collected from 2007
through 2009. Fish implanted with acoustic transmitters rangedfrom 780 to 1,665 mm FL (mean = 1,149 mm, SE = 22.2) andfrom 329 to 659 mm in girth (mean = 455 mm, SE = 10.0)and weighed 4.3–31.1 kg (mean = 13.5 kg, SE = 0.8; Table 1).Of the 60 Lake Sturgeon implanted with transmitters, 47 (78%)were detected by receivers in the reservoir. All Lake Sturgeoncollected and tagged in the main waters of Namakan Reservoirwere relocated by reservoir receivers and a majority (62%)of Lake Sturgeon collected from the Namakan River wererelocated on the reservoir receivers. The remaining 13 LakeSturgeon collected in the Namakan River appeared to remain inthe river and were not relocated on any reservoir receiver. Themean number of Lake Sturgeon detected at each receiver was15 (range, 0–35; median = 16). The greatest numbers of LakeSturgeon (n = 35) were detected at the mouth of the NamakanRiver (Figure 1; Table 2). Other receivers with a relativelyhigh number of Lake Sturgeon detections included NamakanNarrows (n = 25), Smuggler’s Point (n = 25), BlackstoneIsland (n = 20), and the Redhorse River bay (n = 20; Figure 1;Table 2). No Lake Sturgeon were detected at the Moose Riverreceiver (Figure 1; Table 2).
The MDT for all Lake Sturgeon combined ranged from 0.0to 130 km (mean = 37.4 km) from May 2008 to October 2009.The majority of Lake Sturgeon (83%, sexes combined) wereassociated with MDT values exceeding 20 km. Only eight fishexhibited a MDT for the entire time series of less than 20 km(n = 5) or were only detected at one receiver (i.e., 0.0 kmtraveled; n = 3).
Mean MDT of Lake Sturgeon did not vary significantly by sex(two-way ANOVA: F1, 73 = 1.15, P = 0.28), season (F4, 73 =0.53, P = 0.71), or their interaction (sex × season: F4, 73 =0.57, P = 0.68; Table 3). In contrast, mean site residency variedsignificantly across seasons (F4, 73 = 5.13, P < 0.001) and wasgreater during winter months (24.0 d) than in other seasons(range, 1.3–6.5 d; Table 3). Mean site residency did not varysignificantly by sex (F1, 73 = 0.29, P = 0.59) or sex × seasoninteraction (F4, 73 = 0.92, P = 0.45; Table 3).
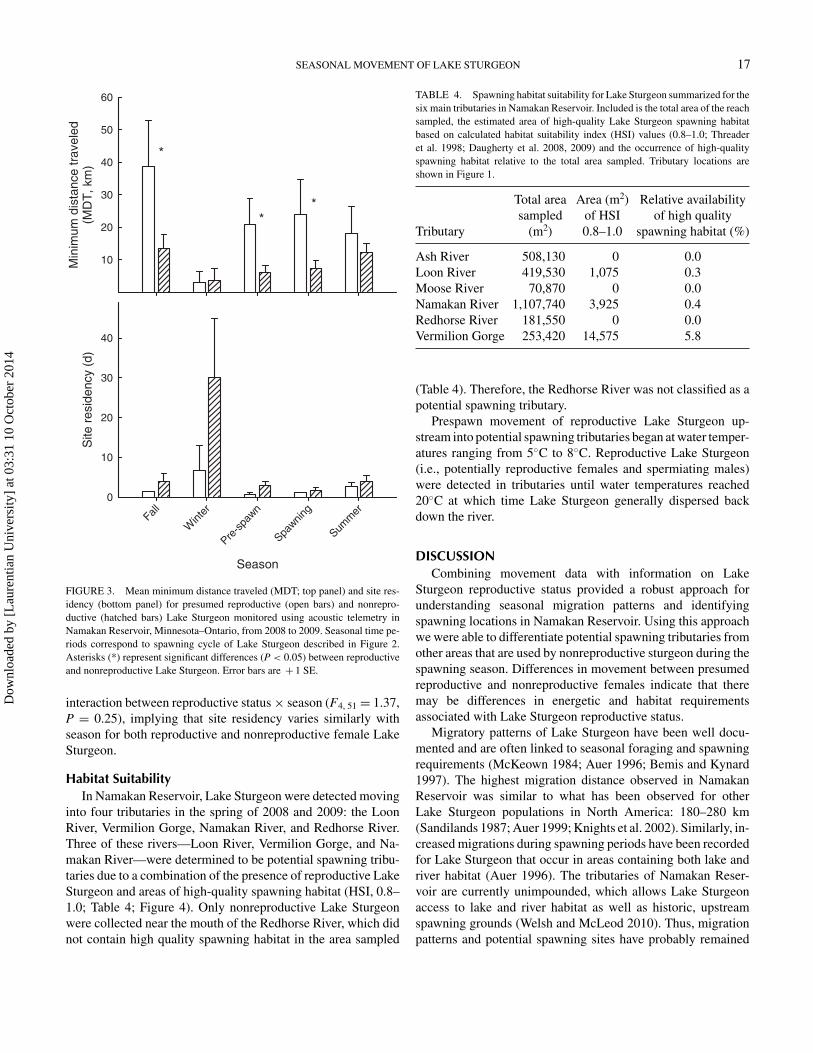
Of the 19 females implanted with transmitters, five were clas-sified as presumed reproductive and 14 were classified as non-reproductive. Reproductive status of female Lake Sturgeon hada significant effect on mean MDT (two-way ANOVA: F1, 51 =13.1, P = 0.0007). On the average, MDT of presumed repro-ductive females (mean = 21.0 km, SE = 4.5) was significantlyhigher than that of nonreproductive females (mean = 8.7 km,SE = 1.4). Seasonal differences between presumed reproduc-tive and nonreproductive females were most pronounced duringprespawn, spawning, and fall time periods (Figure 3). Seasonalso had a significant influence on mean MDT of female LakeSturgeon (F4, 51 = 3.4, P = 0.01) and was higher during fall(mean = 18.6 km) than winter (mean = 3.6 km; Tukey’s multi-ple comparison test: P < 0.05). In contrast, we found no signifi-cant interaction between reproductive status × season for MDTof female sturgeon (F4, 51 = 1.4, P = 0.24).
Mean site residency varied significantly by season (F4, 51 =3.0, P = 0.02) and was higher during winter months (mean =23.4 d) than for other time periods (range of means = 1.5–3.7 d), particularly for nonreproductive females (Figure 3). Theeffect of reproductive status on site residency was not significant(F1, 51 = 3.4, P = 0.07). Similarly, we found no significant
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
3:31
10
Oct
ober
201
4
SEASONAL MOVEMENT OF LAKE STURGEON 17M
inim
um d
ista
nce
trav
eled
(MD
T, k
m)
10
20
30
40
50
60
Season
Fall
Wint
er
Pre-s
pawn
Spawnin
g
Summ
er
Site
res
iden
cy (
d)
0
10
20
30
40
**
*
FIGURE 3. Mean minimum distance traveled (MDT; top panel) and site res-idency (bottom panel) for presumed reproductive (open bars) and nonrepro-ductive (hatched bars) Lake Sturgeon monitored using acoustic telemetry inNamakan Reservoir, Minnesota–Ontario, from 2008 to 2009. Seasonal time pe-riods correspond to spawning cycle of Lake Sturgeon described in Figure 2.Asterisks (*) represent significant differences (P < 0.05) between reproductiveand nonreproductive Lake Sturgeon. Error bars are + 1 SE.
interaction between reproductive status × season (F4, 51 = 1.37,P = 0.25), implying that site residency varies similarly withseason for both reproductive and nonreproductive female LakeSturgeon.
Habitat SuitabilityIn Namakan Reservoir, Lake Sturgeon were detected moving
into four tributaries in the spring of 2008 and 2009: the LoonRiver, Vermilion Gorge, Namakan River, and Redhorse River.Three of these rivers—Loon River, Vermilion Gorge, and Na-makan River—were determined to be potential spawning tribu-taries due to a combination of the presence of reproductive LakeSturgeon and areas of high-quality spawning habitat (HSI, 0.8–1.0; Table 4; Figure 4). Only nonreproductive Lake Sturgeonwere collected near the mouth of the Redhorse River, which didnot contain high quality spawning habitat in the area sampled
TABLE 4. Spawning habitat suitability for Lake Sturgeon summarized for thesix main tributaries in Namakan Reservoir. Included is the total area of the reachsampled, the estimated area of high-quality Lake Sturgeon spawning habitatbased on calculated habitat suitability index (HSI) values (0.8–1.0; Threaderet al. 1998; Daugherty et al. 2008, 2009) and the occurrence of high-qualityspawning habitat relative to the total area sampled. Tributary locations areshown in Figure 1.
Total area Area (m2) Relative availabilitysampled of HSI of high quality
Tributary (m2) 0.8–1.0 spawning habitat (%)
Ash River 508,130 0 0.0Loon River 419,530 1,075 0.3Moose River 70,870 0 0.0Namakan River 1,107,740 3,925 0.4Redhorse River 181,550 0 0.0Vermilion Gorge 253,420 14,575 5.8
(Table 4). Therefore, the Redhorse River was not classified as apotential spawning tributary.
Prespawn movement of reproductive Lake Sturgeon up-stream into potential spawning tributaries began at water temper-atures ranging from 5◦C to 8◦C. Reproductive Lake Sturgeon(i.e., potentially reproductive females and spermiating males)were detected in tributaries until water temperatures reached20◦C at which time Lake Sturgeon generally dispersed backdown the river.
DISCUSSIONCombining movement data with information on Lake
Sturgeon reproductive status provided a robust approach forunderstanding seasonal migration patterns and identifyingspawning locations in Namakan Reservoir. Using this approachwe were able to differentiate potential spawning tributaries fromother areas that are used by nonreproductive sturgeon during thespawning season. Differences in movement between presumedreproductive and nonreproductive females indicate that theremay be differences in energetic and habitat requirementsassociated with Lake Sturgeon reproductive status.
Migratory patterns of Lake Sturgeon have been well docu-mented and are often linked to seasonal foraging and spawningrequirements (McKeown 1984; Auer 1996; Bemis and Kynard1997). The highest migration distance observed in NamakanReservoir was similar to what has been observed for otherLake Sturgeon populations in North America: 180–280 km(Sandilands 1987; Auer 1999; Knights et al. 2002). Similarly, in-creased migrations during spawning periods have been recordedfor Lake Sturgeon that occur in areas containing both lake andriver habitat (Auer 1996). The tributaries of Namakan Reser-voir are currently unimpounded, which allows Lake Sturgeonaccess to lake and river habitat as well as historic, upstreamspawning grounds (Welsh and McLeod 2010). Thus, migrationpatterns and potential spawning sites have probably remained
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
3:31
10
Oct
ober
201
4

18 SHAW ET AL.
FIGURE 4. Relative distribution of (a) reproductive and (b) nonreproductive Lake Sturgeon, sexes combined, collected using gill nets in Namakan Reservoirduring prespawning periods in 2008 and 2009.
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
3:31
10
Oct
ober
201
4

SEASONAL MOVEMENT OF LAKE STURGEON 19
unchanged over time compared with many North American sys-tems that have experienced greater anthropogenic disturbance.
Increased movement by potentially reproductive female LakeSturgeon in the fall contrasts with information reported inother studies that Lake Sturgeon migrate only short distancesto overwintering sites and that movements other than spawn-ing migrations are not extensive (Scott and Crossman 1973;Threader and Brousseau 1986; Rusak and Mosindy 1997). Theincreased movement of presumed reproductive females in Na-makan Reservoir may be an important reproductive strategy foroptimizing forage and spawning success (Auer 2004). Repro-ductive females may travel greater distances in the fall to acquireenergy during the latter stages of oogenesis and (or) to find high-quality overwintering habitat that would allow them to optimizeenergy acquisition and minimize energy expenditure prior tospawning. In a study of Lake Sturgeon in the Mattagami River,Ontario, seasonal patterns in ovarian development were linkedto elevated steroid concentrations and gonadal somatic index(GSI) in the fall (McKinley et al. 1998), supporting the notionof increased energy requirements. Increased movement in thefall may also be associated with a two-step migration pattern,where Lake Sturgeon initiate upstream movement in the fall andoverwinter near the spawning site before resuming a spawningmigration in spring (McLeod and Debruyne 2009). The two-stepmigration pattern is often seen in larger, complex river systemsand may enable sturgeon to use energy reserves for long migra-tions prior to the overwinter season when they will be depleted(Bemis and Kynard 1997; Bruch and Binkowski 2002).
By combining information on female reproductive statuswith acoustic telemetry, we were able to identify three of thesix main tributaries in Namakan Reservoir as containing po-tential spawning sites. Optimum Lake Sturgeon spawning habi-tat includes areas with a combination of fast-flowing water,medium-grain substrate, and depths of 0.5–5 m (Auer 1996,2004). A combination of these characteristics would result inhigh-quality spawning habitat with HSI values between 0.8 and1.0. All three tributaries where reproductive Lake Sturgeon werecollected prior to spawning contained at least one area, in thestretches sampled, with high-quality spawning habitat (e.g., HSIvalues of 1.0). In general, the estimated amount of area withhigh-quality spawning habitat in each tributary should be con-sidered an underestimate. Habitat sampling by boat restrictedus from areas with fast-flowing water (i.e., rapids or falls) orstretches of river upstream from the first natural barrier encoun-tered (i.e., rapids, falls, or low-water zones). However, most ofthese areas do not restrict the upstream movement of Lake Stur-geon. Thus, we were unable to sample upstream reaches thatmay contain high-quality spawning habitat. Each of these trib-utaries differs in its length and distance to potential spawningsites as well as in the amount of high-quality spawning habi-tat available. The Namakan River contains multiple areas withhigh-quality spawning habitat and may therefore have multiplespawning locations, congruent with the relatively high num-bers of Lake Sturgeon documented in this river (McLeod andDebruyne 2009). The Vermilion Gorge and the Loon River are
smaller tributaries compared with the Namakan River that mayhave limited areas with high quality spawning habitat for LakeSturgeon but nonetheless are used by Lake Sturgeon duringspring spawning. It should be noted that the relative impor-tance of any tributary to Lake Sturgeon spawning is difficult toassess based on abundance alone because effort and samplingefficiency varied among tributaries. The Namakan River, for ex-ample, could not be sampled as effectively with gill nets due toits width and relatively higher flows compared with the LoonRiver or the Vermilion Gorge.
A wide range of spawning temperatures (8.8–21.5◦C) hasbeen reported for Lake Sturgeon populations (LaHaye et al.1992; Bruch and Binkowski 2002). In a review of Lake Sturgeonspawning behavior, Bruch and Binkowski (2002) hypothesizedthat spawning activity may be linked more to the rate of watertemperature increase in the spring than to actual water temper-ature. In a 16-year study of Lake Sturgeon spawning behavioron the Wolf River system, Wisconsin, Bruch and Binkowski(2002) noted that the majority of females spawned at watertemperatures of 12–16◦C. The temperature range at which LakeSturgeon in the Namakan Reservoir initiated movement into andout of potential spawning tributaries (5–20◦C) is consistent withother studies and may reflect the realized spawning temperaturerange for Lake Sturgeon in the reservoir.
Reduced movement of Lake Sturgeon during winter monthshas been well documented (Rusak and Mosindy 1997; Auer2004). Rusak and Mosindy (1997) found that Lake Sturgeonin the Rainy River and Lake of the Woods, Ontario, movedinto overwintering sites in the fall as water temperatures rangedbetween 4◦C and 9◦C. Moreover, mean overwinter movementof Lake Sturgeon in Lake of the Woods was relatively low at5–6 km (Rusak and Mosindy 1997), similar to that reportedhere for Lake Sturgeon in Namakan Reservoir. As shown here,seasonal movement data of Lake Sturgeon, combined with infor-mation on their reproductive status, provided important insightabout spawning behavior that may be useful for Lake Sturgeonconservation and restoration efforts.
ACKNOWLEDGMENTSWe thank T. Brezden, T. Burri, J. Butcher, C. Debruyne, J.
Eibler, M. Fincel, B. Flatten, K. Gebhardt, C. Glase, J. Glase,B. Halliday, B. Henning, P. Hogan, D. James, L. Kallemyn, M.Lebron, D. Lilja, B. McLaren, H. Meyer, M. Moseley, W. Nixon,A. Rob, L. Schultz, C. Sundmark, M. Thul, and C. Trembath fortechnical assistance in the field, and E. Cureton at the BozemanFish Technology Center for analysis of sex steroid concentra-tions. G. Adams and S. Mann assisted with age analysis of LakeSturgeon. Funding for this project was provided by the NationalPark Service (CESU Agreement J6820081161). The U.S. Ge-ological Survey, South Dakota Cooperative Fish and WildlifeResearch Unit is jointly supported by the U.S. Geological Sur-vey, South Dakota Department of Game, Fish and Parks, SouthDakota State University, and the Wildlife Management Institute.Any use of trade names is for descriptive purposes only and doesnot imply endorsement by the U.S. Government.
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
3:31
10
Oct
ober
201
4

20 SHAW ET AL.
REFERENCESAdams, W. E., Jr., L. W. Kallemeyn, and D. W. Willis. 2006. Lake Sturgeon,
Acipenser fulvescens, movements in Rainy Lake, Minnesota and Ontario.Canadian Field-Naturalist 120:71–82.
Allen, P. J., M. A. H. Webb, E. Cureton, R. M. Bruch, C. C. Barth, S. J.Peake, and W. G. Anderson. 2009. Calcium regulation in wild populationsof a freshwater cartilaginous fish, the Lake Sturgeon Acipenser fulvescens.Comparative Biochemistry and Physiology 154A:437–450.
Auer, N. A. 1996. Importance of habitat and migration to sturgeons with em-phasis on Lake Sturgeon. Canadian Journal of Fisheries and Aquatic Sciences53(Supplement 1):152–160.
Auer, N. A. 1999. Population characteristics and movements of Lake Sturgeonin the Sturgeon River and Lake Superior. Journal of Great Lakes Research25:282–293.
Auer, N. A. 2004. Conservation. Pages 252–276 in G. T. O. LeBreton,F. W. H. Beamish, and R. S. McKinley, editors. Sturgeons and paddlefishof North America. Kluwer, Dordrecht, The Netherlands.
Bemis, W. E., and B. Kynard. 1997. Sturgeon rivers: an introduction toacipenseriform biogeography and life history. Environmental Biology ofFishes 48:167–183.
Bruch, R. M., and F. P. Binkowski. 2002. Spawning behavior of LakeSturgeon (Acipenser fulvescens). Journal of Applied Ichthyology 18:570–579.
Chateau, O., and L. Wantiez. 2007. Site fidelity and activity patterns of a Hump-head Wrasse, Cheilinus undulatus (Labridae), as determined by acoustictelemetry. Environmental Biology of Fishes 80:503–508.
Craig, J. M., M. V. Thomas, and S. J. Nichols. 2005. Length–weight relation-ship and a relative condition factor equation for Lake Sturgeon (Acipenserfulvescens) from the St. Clair River system (Michigan, USA). Journal ofApplied Ichthyology 21:81–85.
Daugherty, D. J., T. M. Sutton, and R. F. Elliott. 2008. Potential for rein-troduction of Lake Sturgeon in five northern Lake Michigan tributaries: ahabitat suitability perspective. Aquatic Conservation: Marine and FreshwaterEcosystems 18:692–702.
Daugherty, D. J., T. M. Sutton, and R. F. Elliott. 2009. Suitability modeling ofLake Sturgeon habitat in five northern Lake Michigan tributaries: implicationsfor population rehabilitation. Restoration Ecology 17:245–257.
Harkness, W. J. K., and J. R. Dymond. 1961. The Lake Sturgeon. OntarioDepartment of Lands and Forests, Toronto.
Hart, L. G., and R. C. Summerfelt. 1975. Surgical procedures for implanting ul-trasonic transmitters into Flathead Catfish (Pylodictis olivaris). Transactionsof the American Fisheries Society 104:56–59.
Kallemeyn, L. W., K. L. Holmberg, J. A. Perry, and B. Y. Odde. 2003. Aquaticsynthesis for Voyageurs National Park. U.S. Geological Survey, Informationand Technology Report 2003-0001, Washington, D.C.
Knights, B. C., J. M. Vallazza, S. J. Zigler, and M. R. Dewey. 2002. Habitat andmovement of Lake Sturgeon in the upper Mississippi River system, USA.Transactions of the American Fisheries Society 131:507–522.
LaHaye, M., A. Branchaud, M. Gendron, R. Verdon, and R. Fortin. 1992. Re-production, early life history, and characteristics of the spawning grounds ofthe Lake Sturgeon (Acipenser fulvescens) in Des Prairies and L’Assomptionrivers, near Montreal, Quebec. Canadian Journal of Zoology 70:1681–1689.
Lowe, C. G., B. M. Wetherbee, and C. G. Meyer. 2006. Using acoustic telemetrymonitoring techniques to quantify movement patterns and site fidelity ofsharks and giant trevally around French Frigate Shoals and Midway atoll.Atoll Research Bulletin 543:281–303.
McKeown, B. A. 1984. Fish migration. Timber Press, Portland, Oregon.
McKinley, S., G. Van Der Kraak, and G. Power. 1998. Seasonal migrationsand reproductive patterns in the Lake Sturgeon, Acipenser fulvescens, in thevicinity of hydroelectric stations in northern Ontario. Environmental Biologyof Fishes 51:245–256.
McLeod, D. 1999. An assessment of lake sturgeon populations in the lower SeineRiver system, Ontario 1993–1995. Ontario Ministry of Natural Resources,Fort Frances District Report Series 43, Fort Frances.
McLeod, D., and C. Debruyne. 2009. Movement and seasonal distribution ofLake Sturgeon in the Namakan River, Ontario 2007–2008. Ontario Ministryof Natural Resources, Fort Frances District Report Series 82, Fort Frances.
Nickum, J. G., H. L. Bart Jr., P. R. Bowser, I. E. Greer, C. Hubbs, J. A. Jenkins,J. R. MacMillan, J. W. Rachlin, J. D. Rose, P. W. Sorensen, and J. R. Tomasso.2004. Guidelines for the use of fishes in research. American Fisheries Society,Bethesda, Maryland.
Peterson, D. L., P. Vecsei, and C. A. Jennings. 2007. Ecology and biology ofthe Lake Sturgeon: a synthesis of current knowledge of a threatened NorthAmerican Acipenseridae. Reviews in Fish Biology and Fisheries 17:59–76.
Rusak, J. A., and T. Mosindy. 1997. Seasonal movements of Lake Sturgeonin Lake of the Woods and the Rainy River, Ontario. Canadian Journal ofZoology 74:383–395.
Sandilands, A. P. 1987. Biology of the Lake Sturgeon (Acipenser fulvescens) inthe Kenogami River, Ontario. Pages 33–46 in C. H. Olver, editor. Proceedingsof a workshop on the Lake Sturgeon (Acipenser fulvescens). Ontario Ministryof Natural Resources, Fisheries Technical Report Series 23, Toronto.
SAS (Statistical Analysis Systems). 1999. SAS language reference: dictionary,version 8. SAS Institute, Cary, North Carolina.
Scott, W. B., and E. J. Crossman. 1973. Freshwater fishes of Canada. Bulletinof the Fisheries Research Board of Canada 184.
Shaw, S. L., S. R. Chipps, S. K. Windels, M. A. H. Webb, D. T. McLeod, andD. W. Willis. 2012. Lake Sturgeon population attributes and reproductivestructure in the Namakan Reservoir, Minnesota and Ontario. Journal of Ap-plied Ichthyology 28:168–175.
Stahl, M. T., G. W. Whitledge, and A. M. Kelly. 2009. Reproductive biology ofmiddle Mississippi River Shovelnose Sturgeon: insights from seasonal andage variation in plasma sex steroid and calcium concentrations. Journal ofApplied Ichthyology 25(Supplement 2):75–82.
Threader, R. W., and C. S. Brousseau. 1986. Biology and management of theLake Sturgeon in the Moose River, Ontario. North American Journal ofFisheries Management 6:383–390.
Threader, R. W., R. J. Pope, and P. R. H. Schaap. 1998. Development of ahabitat suitability index model for Lake Sturgeon (Acipenser fulvescens).Ontario Hydro, Report H-07015.01-0012, Toronto.
Tomczak, M. 1998. Spatial interpolation and its uncertainty using automatedanisotropic inverse distance weighting (IDW)-cross-validation/jackknife ap-proach. Journal of Geographic Information and Decision Analysis 2:18–30.
Webb, M. A. H., and S. I. Doroshov. 2011. Importance of environmental en-docrinology in fisheries management and aquaculture of sturgeons. Generaland Comparative Endocrinology 170:313–321.
Webb, M. A. H., and D. L. Erickson. 2007. Reproductive structure of theadult Green Sturgeon, Acipenser medirostris, population in the Rogue River,Oregon. Environmental Biology of Fishes 79:305–314.
Webb, M. A. H., G. W. Feist, E. P. Foster, C. B. Schreck, and M. S. Fitzpatrick.2002. Potential classification of sex and stage of gonadal maturity of wildWhite Sturgeon using blood plasma indicators. Transactions of the AmericanFisheries Society 131:132–142.
Welsh, A. B., and D. T. McLeod. 2010. Detection of natural barriers to movementof Lake Sturgeon (Acipenser fulvescens) within the Namakan River, Ontario.Canadian Journal of Zoology 88:390–397.
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
3:31
10
Oct
ober
201
4





























