Embed Size (px)
Citation preview

Informe final* del Proyecto G007 Genética poblacional y filogeografía de las tortugas marinas golfina (Lepidochelys
olivacea) y laúd (Dermochelys coriacea) en el Pacífico mexicano Responsable: Dr. Federico Alberto Abreu Grobois Institución: Universidad Nacional Autónoma de México
Instituto de Ciencias del Mar y Limnología Estación Mazatlán Laboratorio de Genética
Dirección: Explanada de la Azada y Cerro del Crestón s/n, Mazatlán, Sin, 82240 , México
Correo electrónico: [email protected]
Teléfono/Fax: N/D Fecha de inicio: Febrero 29, 1996 Fecha de término: Septiembre 13, 2000 Principales resultados: Base de datos, Informe final, Hojas de cálculo
Forma de citar** el informe final y otros resultados:
Abreu Grobois F. A., 1999. Genética poblacional y filogeografía de las tortugas marinas golfina (Lepidochelys olivacea) y laúd (Dermochelys coriacea) en el Pacífico mexicano. Universidad Nacional Autónoma de México. Instituto de Ciencias del Mar y Limnología. Informe final SNIB-CONABIO proyecto No. G007. México D. F.
Forma de citar las hojas de cálculo
Abreu Grobois F. A., 1999. Genética poblacional y filogeografía de las tortugas marinas golfina (Lepidochelys olivacea) y laúd (Dermochelys coriacea) en el Pacífico mexicano. Universidad Nacional Autónoma de México. Instituto de Ciencias del Mar y Limnología. Hojas de cálculo SNIB-CONABIO proyecto No. G007. México D. F.
Resumen: A pesar de los intensos esfuerzos de conservación dirigidos hacia las tortugas marinas a nivel internacional, aún están catalogadas como especies en peligro de extinción, situación en gran manera resultado de su singular ciclo de vida que incluye: (1) un tiempo de generación prolongado y virtualmente desconocido, (2) el escaso o nulo conocimiento sobre el período pelágico de las crías y juveniles y (3) la ocurrencia de extensas migraciones como adultos entre sus hábitats de alimentación y de reproducción. Las estrategias de manejo para la recuperación y conservación de cualquier especie amenazada de extinción tienen como meta fundamental el mantener o incrementar el tamaño y la diversidad de las poblaciones. Para ello, es esencial definir las unidades reproductoras básicas que componen las especies (stocks o "unidades de manejo"-UM). En tortugas marinas el análisis del ADN ha permitido determinar las diferencias genéticas entre UM con precisión, rapidez y a costos relativamente bajos y, con ello, clarificar el grado de aislamiento y los niveles de migración entre poblaciones. El presente proyecto constituye un primer esfuerzo en el país por aplicar la genética molecular al estudio de las tortugas marinas y representa un esfuerzo de colaboración en el que participan diversas instituciones nacionales (Instituto de Ciencias del Mar y Limnología y la Facultad de Ciencias, UNAM) y extranjeras (University of Florida; National Marine Fishery Service, La Jolla). Está dirigido a las colonias más importantes de dos de las especies de tortugas marinas que anidan en las costas del Pacífico mexicano: la tortuga golfina (Lepidochelys olivacea) y la tortuga laúd (Dermochelys coriacea). Se analiza la variación en las secuencias de la región control del ADN mitocondrial (tortuga golfina) así como en loci de microsatélites en el ADN nuclear (ambas especies). Los resultados nos permitirán: (1) cuantificar el grado de estructuración y aislamiento reproductivo entre colonias anidadoras así como (2) estimar los niveles de variabilidad genética y (3) el grado de intercambio genético entre poblaciones. Como consecuencia, se podrá delimitar las UM en estas especies, conocimiento requerido para fundamentar los programas de conservación tanto en el ámbito local (México) como regional (Pacífico oriental). Al correlacionar estos resultados con información derivada de estudios previos sobre patrones de migración, tamaño poblacional, distribución geográfica y diferenciación genética a nivel mundial, nos permitirá también reconstruir la biogeografía e historia evolutiva reciente de las especies dentro de la región de estudio.

_______________________________________________________________________________________________
• * El presente documento no necesariamente contiene los principales resultados del proyecto correspondiente o la descripción de los mismos. Los proyectos apoyados por la CONABIO así como información adicional sobre ellos, pueden consultarse en www.conabio.gob.mx
• ** El usuario tiene la obligación, de conformidad con el artículo 57 de la LFDA, de citar a los autores de obras individuales, así como a los compiladores. De manera que deberán citarse todos los responsables de los proyectos, que proveyeron datos, así como a la CONABIO como depositaria, compiladora y proveedora de la información. En su caso, el usuario deberá obtener del proveedor la información complementaria sobre la autoría específica de los datos.

GENETICA POBLACIONAL Y FILOGEOGRAFIA DE LAS TORTUGAS MARINAS GOLFINA (Lepidochelys olivacea) Y LAUD (Dermochelys coriacea) EN EL PACIFICO MEXICANO
(proyecto G-007) INFORME FINAL
Febrero, 2000
Responsable:
Dr. F Alberto Abreu Grobois Laboratorio de Genética y
Banco de Información sobre Tortugas Marinas Unidad Académica Mazatlán
Instituto de Ciencias del Mar y Limnología UNAM
Co-investigadores:
M. en C. Raquel Briseño Dueñas Cand. M. en C. Ana Barragán Rocha Laboratorio de Genética y Laboratorio de Tortugas Marinas Banco de Información sobre Tortugas Marinas Facultad de Ciencias Unidad Académica Mazatlán UNAM Instituto de Ciencias del Mar y Limnología UNAM

INFORME FINAL - Proyecto G007
TITULO. GENETICA POBLACIONAL Y FILOGEOGRAFIA DE LAS TORTUGAS MARINAS GOLFINA (Lepidochelys olivacea) Y LAUD (Dermochelys coriacea) EN EL PACIFICO MEXICANO INSTITUCIÓN. Instituto de Ciencias del Mar y Limnología, UNAM. Laboratorio de Genética. RESPONSABLE DEL PROYECTO. Dr. F. Alberto Abreu Grobois AREA. Genética de la conservación. GRUPO. Zoológico. AMBIENTE. Marino. REGIONES PRIORITARIAS: El área de muestreo y estudio comprende (además de otras) la región costera de 11 áreas prioritarias (según CONABIO/USAID/WWF/FMCN/PACKARD): #4 Bahía Magdalena, #20 Piaxtla-Urías, # 22 Bahía de Banderas, # 25 Mismaloya-Pta. Soledad, #28 Cuyutlán-Chupadero, #29 Maruata-Colola, #30 Mexiquillo-Delta del Balsas, #31 Tlalcoyunque, #34 Chacahua-Escobilla, #35 Puerto Angel-Mazunte, #39 Punta Arista. ESPECIES. Tortugas golfina (Lepidochelys olivacea) y laúd (Dermochelys coriacea), ambas considerada en peligro de extinción en la norma oficial vigente (NOM-059-ECOL/1994). PALABRAS CLAVE. Especies en peligro, mtDNA, microsatélites, genética, conservación, marcadores moleculares.

INDICE 1. INTRODUCCION ......................................................................................................................................................5
Biología de las especies ................................................................................................................................................. 6 Tortuga golfina, Lepidochelys olivacea ................................................................................................................... 6 Tortuga laúd, Dermochelys coriacea ........................................................................................................................ 7
Problemática de las especies ......................................................................................................................................... 9 ¿Por qué son susceptibles las tortugas marinas a la extinción? ................................................................................. 9
Factores de mortalidad ............................................................................................................................................... 10 Hábitos migratorios ................................................................................................................................................ 11 Fidelidad a los sitios de anidación........................................................................................................................... 11
3. ANTECEDENTES .................................................................................................................................................. 12 Aplicaciones de la genética molecular en las tortugas marinas ................................................................................ 12 Conocimiento actual sobre la genética de las especies objeto de este estudio .......................................................... 13
4. OBJETIVOS DEL PROYECTO ........................................................................................................................ 15 Objetivo general .......................................................................................................................................................... 15
Objetivos particulares ............................................................................................................................................ 15
5. MATERIALES Y METODOS ............................................................................................................................ 16 Arcas de Estudio ......................................................................................................................................................... 16 Obtención y preservación de muestras ...................................................................................................................... 18
Muestras de tejido sólido ....................................................................................................................................... 18 Muestras de sangre ................................................................................................................................................ 18
Análisis bioquímico ..................................................................................................................................................... 19 Extracción de DNA ............................................................................................................................................... 19 Cebadores para la amplificación del DNA ............................................................................................................. 19 Reacción en cadena de la polimerasa (PCR) .......................................................................................................... 19
Región control en tortugas golfina ................................................................................................................19 Microsatélites del DNAnuclear en tortugas golfina ...................................................................................... 20 Microsatélites del DNAnuclear en tortugas laúd .......................................................................................... 21
Análisis estadístico de la información genética ........................................................................................................... 22 Región control ...........................................................................................................................................................
Variabilidad genética .................................................................................................................................... 22 Diferenciación genética ................................................................................................................................. 22 Estructura genética de la población inferida por el Análisis de Varianza Molecular (AMOVA) ................. 23 Las comparaciones de los valores de F st’s entre pares de colonias, ............................................................ 23 Estimación del flujo génico entre poblaciones (Nm)...................................................................................... 23 Compraración de frecuencias haplotípicas y sus intervalos de confianza .................................................... 23 Microsatélites ................................................................................................................................................24
Personal participante .................................................................................................................................................. 24 Estudio de tortuga golfina- .................................................................................................................................... 24 Estudio de tortuga laúd- ......................................................................................................................................... 24
6. RESULTADOS Y DISCUSION ......................................................................................................................... 25 Tortuga golfina ........................................................................................................................................................... 25
Variación en la región control del DNA mitocondrial (por Raquel Briseño Dueñas y F. Alberto Abreu Grobois) .............................................................................. 25 Secuencias ..................................................................................................................................................... 25 Análisis estadístico de los resultados ............................................................................................................ 25
Niveles de variabilidad genética y su distribución ................................................................................... 25 Diversidad genética ................................................................................................................................. 29 Frecuencias haplotipicas y sus intervalos de confianza ............................................................................ 29 Estructura de la población ....................................................................................................................... 30
Pruebas de diferenciación interpoblacional .................................................................................................. 30 Distancias genéticas ................................................................................................................................ 30

Estimaciones de las tasas de migración .................................................................................................. 30 Variación en loci de microsatélites del DNA nuclear en tortuga golfina
(por F. Alberto Abreu Grobois y Felipe López Chávez) ................................................................................ 32 Frecuencias alélicas, genotipos y niveles de variación ................................................................................. 32Estructura poblacional evaluada por análisis de microsatélites ...................................................................... 32
Información complementaria de Lepidochelys olivacea en el Pacifico oriental (por Raquel Briseño Dueñas y F. Alberto Abreu Grobois) ............................................................................ 38 Flujo génico entre colonias ........................................................................................................................... 38
Discusión ............................................................................................................................................................... 40 Diversidad genética ....................................................................................................................................... 40Estructura y unidades de manejo ................................................................................................................... 41
Tortuga laúd ............................................................................................................................................................... 43 Variación genética en loci de microsatélites del DNA nuclear
(por Ana Barragán Rocha) ............................................................................................................................. 43 Discusión ............................................................................................................................................................... 46
7. CONCLUSIONES Y RECOMENDACIONES ...................................................................................................................................................48
8. AGRADECIMIENTOS .................................................................................................................50 9. PERMISOS DE COLECTA ....................................................................................................................................................................................................5010. LITERATURA CITADA ...................................................................................................................................... 51

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 5
1. INTRODUCCION Las tortugas marinas modernas se agrupan en dos familia que integran a siete especies distribuidas en
los mares tropicales y subtropicales del mundo. En la familia Cheloniidae se encuentran las tortugas de caparazón constituido por placas óseas o concha dura, con seis especies. Estas son, la tortuga verde (Chelonia mydas), la caguama (Caretta caretta), la kikila (Natator depressus), la carey (Eretmochelys imbricata), la golfina (Lepidochelys olivacea) y la lora (L. kempii). Algunos especialistas consideran a la tortuga verde del Pacífico oriental, también denominada Chelonia agassizii, como una octava especie. En la familia Dermochelyidae se encuentra una sola especie, la laúd (Dermochelys coriacea), y son tortugas que tienen un caparazón parecido a un cuero grueso y flexible carente de placas óseas.
Estas especies permanecen casi toda su vida en el mar y han desarrollado modificaciones anatómicas para su adaptación a la vida acuática. Sin embargo, como vestigios de su origen terrestre incursionan sobre playas arenosas tropicales para incubar sus huevos. Las especies y aún poblaciones de una misma especie, difieren en sus ciclos estacionales, la amplitud de su distribución geográfica y el comportamiento. Por ejemplo, algunas especies realizan extensas migraciones entre los sitios de alimentación y de anidación. Cuando esto sucede, el desgaste de las travesías y los requerimientos energéticos para la producción de huevos, condicionan fisiológicamente la periodicidad reproductoras de las hembras a uno, dos, tres o cuatro años. Los machos y las hembras migran a las áreas de reproducción y se aparean en aguas cercanas a los sitios de anidación. Unicamente las hembras salen a las playas durante la fase del desove. El número de huevos en cada nidada es de 50 a 160 y depositan de 1 a 7 nidadas por temporada, según la especie. Los altos niveles de fecundidad contrarrestan la elevada tasa de mortalidad de las tortugas marinas durante el proceso de crías a la fase adulta. Algunos autores estiman que en condiciones naturales, la proporción de sobrevivencia vs. mortalidad es de 1:1000 y otros de 1:10,000.
Los huevos permanecen en los nidos a una profundidad de 40 a 70 cm, durante un período de 45-60 días dependiendo de la especie y de la temperatura de incubación (que también determina el sexo de las crías). Al concluir el periodo de desarrollo, los neonatos ascienden a la superficie y se dirigen inmediatamente al mar, habiendo recibido durante su estancia en la fase terrestre una «impronta» (no se sabe el tipo de información adquirida, sin embargo se ha especulado que «guardan» una memoria sobre las características fisicoquímicas de su playa de origen, que permitirá el regreso a su sitio de nacimiento una vez que alcancen la edad de primera reproducción.
En el mar, las crías se integran a la comunidad pelágica durante períodos de unos cuantos meses a varios años, según la especie. Ya como juveniles, cambian sus hábitats hacia zonas someras protegidas, como bahías y estuarios. El desarrollo a la madurez sexual se prolonga durante varias décadas antes de regresar como adultos para reproducirse a las playas de su nacimiento.
Estas son características compartidas por todas las especies. Sin embargo, existen diferencias entre las especies que están relacionadas con los hábitos alimenticios (especialmente de los juveniles y adultos), distribución geográfica y rutas migratorias que las distingue y que explica muchas de las diferencias entre las especies en el grado de afectación por las actividades del hombre. Las características más conspicuas para cada una de las dos especies investigadas son descritas en los párrafos subsiguientes. No se pretende hacer una revisión exhaustivos con los datos aquí presentados, están basados en documentos que incluyen, entre otros, los Planes de Recuperación de las agencias de pesquerías y vida silvestre de EUA, la revisión de Márquez (1990) así como en trabajos realizados principalmente en México.
En general, las tortugas marinas representan uno de los grupos marinos más sujetos al uso y veneración tradicional de las comunidades costeras en las zonas tropicales del mundo. También tienen un largo antecedente histórico de explotación comercial, al principio artesanal y más recientemente (en los últimos siglos), que llegó a alcanzar niveles vastos e intensivos. Esta práctica contribuyó de manera significativa al colapso y extinción de muchas de las poblaciones de tortugas marinas a lo largo del mundo. Varias de estas eran poblaciones gigantezcas que ahora han desaparecido. Las evidencias de poblaciones reducidas, principalmente por actividades antropogénicas se encuentran documentadas a lo largo de su distribución global. Para las poblaciones de las tortugas golfina y laúd, las tendencias poblacionales en el Pacífico hacia la baja ha sido documentada en la última década (Eckert, 1993) particularmente para la segunda especie que ha sufrido desplomes en sus tamaños

6 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacifico mexicano
aún más recientemente (Eckert y Sarti, 1997). Los principales factores causales de las disminuciones en las tortugas marinas han sido la
extracción directa en niveles insostenibles de adultos o de huevos, la captura incidental (por diversas artes de pesca enfocada a otras especies pero que ocurren en sitios de alimentación o migración de las tortugas marinas) y, más recientemente, la modificación o degradación de hábitats críticos (p. ej., los de reproducción)
La disminución drástica ha provocado que todas las especies estén catalogadas en las diversas listas como organismos amenazados o en peligro de extinción- en la Lista Roja de la UICN (Unión Mundial para la Naturaleza); en el Apéndice I de la CITES (Convención Internacional sobre el Comercio de las Especies Amenazadas o en Peligro) y de la CMS (Convención para Especies Migratorias, o Convención de Bonn), entre otras. En México desde 1994 todas las especies de este grupo que se encuentran en aguas territoriales, están catalogadas por la Norma Oficial Mexicana NOM-0059-94, como en peligro.
Por lo anterior, resulta obvio que una conservación efectiva para estas especies requiere del concierto entre el mejor conocimiento de sus hábitos, de su distribución geográfica, la estructura genética de sus poblaciones para derivar las mejores prácticas de manejo que por un lado fomenten el crecimiento de las tortugas marinas y contrarresten los impactos nocivos del hombre. Biología de las especies
Tortuga golfina, Lepidochelys olivacea Nombres comunes: golfina, pinta, frijolilla y caguama (lora en algunos países de Centroamérica). Taxonomía: En 1829, Eschscholtz la describió bajo el nombre de Chelonia olivacea.
Posteriormente fue ubicada en el género Lepidochelys por Fitzinger (1843) Descripción: Las tortugas del género Lepidochelys son las más pequeñas de las tortugas marinas que
xisten actualmente. Los adultos pesan entre 30 y 45 kg., el largo del carapacho en línea recta es de 55 a 75 cm.
A diferencia de otra especie de este género, L. kempii, las golfinas tienen mayor altura corporal (carapacho más elevado y aplanado en la porción dorsal). El carapacho es redondeado, con escudos marginales convexos y dirigidos ligeramente hacia arriba. Carapacho sin fontanelas laterales, de color
olivo amarillento, tiene cinco escudos centrales, 5, 6 o más pares de escudos laterales. El plastrón presenta un poro en cada uno de lo escudos inframarginales. El diámetro de los huevos es 40 mm aprox. y el peso varía entre 30 y-37 g. Las crías miden de 3.1-4.4 cm del largo del carapacho y pesan entre 14-21 g.
Alimentación : La tortuga golfina es predominantemente carnívora, capaz de comer un solo tipo de alimento por largos períodos. Por ejemplo, las poblaciones que se localizan en las costa occidental de Baja California, se alimentan exclusivamente de la langostilla Pleuroncodes planipes. Sin embargo, existe una lista documentada de la amplia variedad de alimento que consumen tanto de la comunidad del bentos como del necton: camarón, medusas. cangrejos, caracoles, tunicados y, en algunas áreas, también de algas (Márquez, 1990)
Crecimiento: La información sobre el crecimiento de las tortugas golfinas en el medio natural no está bien definida. Sin embargo, se tienen evidencias de que los adultos continúan creciendo aún después de alcanzar la edad de primera
reproducción, estimada por Márquez (com.pers.) entre los 10-15 años. Reproducción El apareamiento puede ocurrir durante la migración a las áreas de reproducción, pero
generalmente se ha observado frente a las playas de anidación, antes y durante la temporada. El período de anidación generalmente es durante el verano y otoño. En algunas playas es posible observar hembras anidando durante todos los meses del año. La fecundidad promedio para esta especie se ha estimado en 285 huevos por estación y 95 huevos por desove.
Distribución de las poblaciones: Por mucho, esta es la especie más abundante en las costas del Pacífico oriental. Su área de distribución es esencialmente tropical. Observaciones de la Comisión Interamericana del Atún Tropical (CIAT), muestran una distribución estacional para la tortuga golfina. Los valores del índice de frecuencia relativa de ocurrencia (FRO) son generalmente altos cerca de las costas de Centro y Sudamérica
Lepidochelys olivacea (ilustración tomada de WATS,
1983)

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas. Ana R. Barragán Rocha 7 durante los meses pico de la anidación (julio a diciembre, Pitman?) . Sin embargo el FRO para México siempre es bajos, a pesar de la alta densidad de anidación de la población reproductora de Oaxaca.
Además de la distribución estacional, también se ha encontrado una distribución diferencial por sexo y talla. Juveniles de esta especie se han localizado en las áreas de alimentación de Ecuador y Colombia. También se les ha observado en el Pacífico mexicano en zonas caracterizadas por su alta productividad: la región del Istmo de Tehuantepec, en el Golfo de California y en el margen occidental de la península de Baja California. Informes de la CIAT indican que la distribución de esta especie abarca desde los 30° N a los 15° S. Los registros de la CIAT clarifican que las dos áreas más importantes para la tortuga golfina en el Pacífico oriental son las costas de Centro América y las áreas de alimentación y desarrollo de Colombia y Ecuador. Las anidaciones aparecen desde el sur de Sonora y Baja California Sur (México) hasta Colombia. También se encuentran individuos en zonas de alimentación en aguas del sudoeste de EUA y, en menor densidad, en aguas de Perú y Chile. En el Océano Atlántico, las golfinas ocurren ampliamente pero en menor abundancia en la costa oriental de la Guyana Francesa y la noroccidental de Guyana. Existen también anidaciones escasas de esta especie en la costa de Brasil. Es la única especie que, en algunas colonias, manifiesta el fenómeno de "arribazones" (anidaciones masivas) en las que decenas a centenas de miles de hembras anidan a lo largo de las 1-3 noches que dura la anidación.
Hábitat: Fuera de sus áreas de anidación, los adultos de esta especie se les localiza en hábitats neríticos, desplazándose o descansando, manteniéndose a la deriva por periodos prolongados. También se ha observado que realizan inmersiones hasta los 200 ni de profundidad.
Migraciones: En el Pacífico oriental, las tortugas golfinas efectúan sus migraciones principalmente a lo largo del margen continental, se desplazan en forma gregaria formando "flotillas" de varias decenas, cientos o miles. No se le conocen migraciones transoceánicas regulares.
Situación actual: Según los criterios de la UICN y de la Norma Oficial Mexicana NOM-059-ECOL-1994, la tortuga golfina se encuentra catalogada como especie EN PELIGRO. Los criterios aplicados se han fundamentado fuertemente en los desplomes significativos observados para colonias reproductoras con amplios antecedentes de conformar arribazones (por ejemplo, las que llegaban a las playas de Piedra de Tlalcoyunque, Guerrero y el Playón de Mismaloya, Jalisco- ambas en México) ocurridos después de la sobrepesca en la década de los 60's y la sensible disminución en la densidad y número de arribazones en Escobilla, Oaxaca (México) en el período de 1977 a 1988 y en Nancite, Costa Rica en los últimos años (R. Valverde, pers. com.) .
No obstante, la demografía mostrada en las ultimas seis temporadas indican incrementos en el número de anidaciones estimadas, por lo menos a lo largo de la costa de México. Esta tendencia, sugiere que las poblaciones de esa región debe dársele un seguimiento más cuidadoso para analizar su condición real a nivel regional.
Principales amenazas: En México las principales amenazas para la golfina son la extracción de huevos y tortugas en playa. También son afectadas por la pesca ilegal y la captura incidental por artes de pesca destinadas a otros recursos marinos. La degradación del hábitat principalmente por desmonte y cambio de uso del suelo del hábitat adyacente a la zona de anidación, la contaminación de la playa y de la zona costera, son factores que están incidiendo en el menoscabo de las poblaciones. Los niveles de impacto real no se han evaluado con precisión en la mayor parte de sus áreas de distribución.
Tortuga laúd, Dermochelys coriacea Nombres comunes: tortuga laúd, garapacho, siete filos, tinglado, machincuepo, tortuga de canal.
Taxonomía: El nombre genérico Dermochelys fue introducido por Blainville (1816). El nombre específico coriacea fue usado por Vandelli (1761) y adoptado por Linneaus en 1766
Descripción: Esta es la más grande de las tortugas marinas vivientes y es tan diferente del resto que se ha clasificado en una familia distinta: Dermochelyidae. Mientras que las otras tortugas tienen placas óseas cubiertas por escamas de naturaleza córnea en el carapacho, el de ésta tiene una piel gruesa y continua, interrumpida por siete quillas longitudinales que se unen en la porción distal del carapacho. Las aletas anteriores son proporcionalmente más largas que en otras tortugas marinas, alcanzando unos 270 cm en el adulto. El largo curvo del carapacho en hembras adultas varía entre 137 y 183 cm. En los adultos, la epidermis en la parte dorsal

8 Genética poblacional y frlogeografia de las tortugas marinas golfina y laúd en el Pacifico mexicano
es predominante negra con un número variable de motas blancas. El área ventral es de color gris rosado con manchas blancas.
Alimentación: Se alimentan de medusas, tunicados, sifonóforos y salpas. La alimentación de estos organismos ocurre a profundidades que frecuentemente rebasan los 50 m y en ocasiones alcanzan 1,300 m o más.
Crecimiento: No se tiene información sobre la tasa de crecimiento de juveniles en su medio natural. Una de las causas principales es el hecho de que la distribución de fase de los juveniles es prácticamente desconocida. En cautiverio el problema se agudiza debido a la baja tasa de sobrevivencia (un promedio de dos años) bajo estas condiciones.
Reproducción: Existen evidencias indirectas de que los apareamientos ocurren antes o durante la migración a la playa de anidación (Eckert y Eckert, 1988). El comportamiento de las hembras durante el proceso de la anidación es similar al de las otras tortugas marinas. La laúd tiene un ciclo de anidación de entre 2 y 3 años, oviposita de cuatro a cinco veces por temporada. En cada nido depositan entre 61 y 126 huevos de dos tipos. Una proporción (40-85) son huevos viables de color blanco y forma esférica, de 4.5 a 6 cm de diámetro. El resto (20-50) no son viables, de color blanco, varían en forma y tamaño, son más pequeños y carentes de yema o vitelo. Un problema común en la sobrevivencia de la crías es el hecho de que las hembras prefieren playas de alta energía, expuestas a modificaciones bruscas por ello las nidadas se pierden frecuentemente debido a la erosión. Las principales áreas de concentración para la anidación en el
continente americano se localizan en el Pacifico oriental, principalmente en los estados de Guerrero, Michoacán y Oaxaca.
Distribución de las poblaciones: La laúd es la especie más oceánica y migratoria de las tortugas marinas además que, gracias a singulares adaptaciones fisiológicas que le permite sobrevivir en ambientes fríos, alrededor del continente americano se le encuentra en el Pacífico desde Canadá hasta Chile y desde Labrador (Canadá) hasta Mar del Plata (Argentina) en el Atlántico.
Hábitat: a pesar de que se le considera la más pelágica de todas las tortugas, son escasos los registros de su presencia en mar abierto (Márquez, 1990 y Pitman, 1993). Los requerimientos de hábitat de los juveniles y neonatos son prácticamente desconocidos
Migraciones: Las laúd realizan migraciones mas extensas y se acercan a latitudes de aguas más frías que cualquier otro reptil marino (Pritchard, 1976). Abarcan aguas boreales, templadas y tropicales probablemente optimizando oportunidades tanto de anidación como de alimentación.
Situación actual: Se le considera EN PELIGRO de extinción dentro de toda su distribución global por la UICN, en la Norma Oficial Mexicana (NOM-059-EOL-1994), en el Acta de Especies en Peligro de los EUA y está incluida en el Apéndice 1 del CITES.
En el Pacífico, las poblaciones de laúd se encuentran en severa declinación. Una evaluación reciente (Sarti el al.,1996) realizada en la región del Pacífico mexicano, considerada por diversos autores como el sostén de la mitad de toda la población global, ha mostrado un descenso alarmante en sus registros durante los últimos 10 años. La últimas cifras estiman que en la región del Pacífico mexicano durante la temporada 1995-1996 anidaron menos de 1,000 hembras. Se les captura incidentalmente en las pesquerías pelágicas de palangre en el Pacífico Norte en cantidades que, considerando estas declinaciones, muy probablemente representen un impacto significativo. Las tendencias observadas hace que los planes de recuperación de esta especie deban considerarse como de alta prioridad (Bolten et al., 1996).
Principales fuentes de impacto: Las principales fuentes de impacto antropogénico las constituyen la extracción de las nidadas en la playas de anidación, la captura incidental en palangres y redes de deriva para la pesca comerciales en zonas pelágicas y en redes agalleras en otras zonas. Debido a sus hábitos alimenticios, las laúd también ingieren una amplia gama de desechos que flotan en el mar: bolsas y pedacería de diferentes plásticos, aglomeraciones de chapopote, etc. con consecuencias que podrían ser nocivas para las tortugas.
Dermochelys coriacea (ilustración tomada de WATS. 1983)

Informe Final - F. Alberto Abreu Grobois. Raquel Briseño Dueñas, Ana R. Barragán Rocha 9
Desgraciadamente aún no se conocen los niveles de impacto por estos hábitos. Se le captura en ocasiones incidentalmente por flotas de pesca comercial de camarón.
Problemática de las especies
¿Por qué son susceptibles las tortugas marinas a la extinción? La preocupación por la extinción de las tortugas marinas está basada en las evidencias ampliamente
documentadas de la extirpación completa de muchas poblaciones y dramáticos colapsos en otras. En la actualidad, con excepción de la kikila que se considera como especie vulnerable a la extinción, las demás tortugas están catalogadas como especies en peligro crítico o en peligro por los nuevos criterios de la UICN (UICN, 1994) que incrementan la objetividad al aplicar criterios de clasificación que toman en cuenta, entre otros elementos, factores que definen las tendencias poblacionales y un marco de tiempo relativo al tiempo de generación de la especie. En la actualidad solamente se tienen clasificadas las tortugas marinas como especies, aplicando los procedimientos a la información disponible para su distribución total, a nivel mundial. Se espera que en futuro, en la medida que se tenga la capacidad para identificar las poblaciones o stocks individuales y se cuente con estimaciones de las tendencias demográficas en series de tiempo con las escalas necesarias, se puedan aplicar los procedimientos a este nivel también.
Mientras que la persistencia de las tortugas marinas a lo largo de millones de años refleja un éxito evolutivo, muchos de los rasgos observados en los ciclos vitales de las tortugas marinas hace que,bajo condiciones de explotación, sean muy vulnerables a la extinción. En particular porque son organismos que maduran sexualmente muy lentamente y sus hábitos altamente migratorios hacen que el desenvolvimiento del ciclo de vida utilice una variedad de hábitats, dispersos en áreas muy extensas (abarcando, inclusive, aguas territoriales de varios países) y durante períodos prolongados. Estos rasgos dificultan su estudio y complican su conservación y proteccion ya que para lograrla de manera efectiva se requiere un conocimiento de los diferentes hábitats utilizados, así como los requerimientos de las diferentes fases del ciclo vital y la naturaleza y magnitud de las presiones que les afectan.
La determinación del estado de una especie se basa en una evaluación de tendencias demográficas con el tiempo, y del balance entre animales jóvenes y veteranos. Debido a que es extremadamente difícil realizar censos de las tortugas en el mar abierto, normalmente se obtienen estimaciones de las tendencias poblacionales por medio de muestreos representativos de alguna fase de desarrollo. En el caso de estas especies, comúnmente se usa el número de hembras anidando en playa y sus tendencias de las fluctuaciones demográficas observadas a largo plazo proporciona estimadores del estado de la población. Disminuciones en los tamaños de las mismas pueden incrementar la susceptibilidad a la
extinción ya que poblaciones pequeñas o reducidas son más vulnerables a los impactos de eventos catastróficos de ocurrencia azarosa, o provocar problemas por consanguinidad y las enfermedades asociadas.
Si bien es cierto que, en general, la sobrevivencia de una especie está determinada por el balance entre las tasas de reproducción y de mortalidad, y que éstas, a su vez, son moduladas por varios fenómenos (algunos naturales y otros antropogénicos), el factor de mayor repercusión sobre la velocidad de incremento poblacional, es el tiempo que tardan los organismos en alcanzar la maduración sexual. Cuando este proceso es rápido, la aparición de nuevas generaciones
Figura 2.1 Efectos de una cosecha total de hembras anidadoras. Ejemplo simplificado e hipotético en el que se sacrifica el 100% de las hembras reproductoras antes de que puedan anidar ( indicado por las "X") a partir del año de 1974. Las crías que se producen antes de la explotación realizan su maduración en un lapso de varias décadas antes de regresar para la reproducción (en la ilustración se usa un tiempo 25 años, aunque este valor varía entre especies) que ocurre aún bajo el régimen de explotación. Nótese que el efecto de esta captura excesiva no se observa sino hasta después del año 2000, para cuando es demasiado tarde (adaptado de Mortimer. 1995).

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 11 por alto la normatividad vigente para cada país. En México y otros países, por ejemplo, donde se tiene desde 1990 una veda permanente sobre la captura o comercialización de las tortugas marinas o sus productos, aún persisten prácticas de captura directa con trasmallos en algunas regiones que ponen en riesgo los programas de conservación afectando la fase de reproductores y pre-adultos en corredores migratorios.
Entre las amenazas antropogénicas más importantes que afectan a la sobrevivencia de las tortugas marinas se tiene la mortalidad por captura incidental en diversas artes de pesca comercial diseñadas para otras especies. Incluyen, además de otras, las redes de arrastre para la captura del camarón que, como caso especial en el contexto del presente trabajo se verá con mayor detalle en la siguiente sección.
La destrucción, degradación o modificación de hábitats (particularmente áreas de reproducción) también disminuyen la capacidad reproductiva de las tortugas marinas. En este último rubro se incluyen los desarrollos turísticos en playa que, como varía regionalmente, afecta a las especies en distinto grado dependiendo de su distribución (ver sección anterior).
Hábitos migratorios. La extensión geográfica del ciclo completo de las tortugas marinas es extremadamente grande,
abarcando circuitos de varios cientos o miles de kilómetros y muchas veces extendiéndose a través de jurisdicciones territoriales de varias naciones. Esta característica acarrea importantes implicaciones para el manejo. Significa que cualquier menoscabo que sufran las poblaciones en una porción de su distribución, ocasionara repercusiones observables en el resto. Además, el comportamiento de las tortugas marinas también causa que en algunos sitios de alimentación y en corredores migratorios, converjan tortugas originarias de varias colonias. Donde ocurre esta mezcla de poblaciones se complica notablemente la conservación de las especies ya que, además de requerir la competencia de diversos países es necesario poder identificar las colonias individuales y la proporción de su ocurrencia en las mezclas.
Fidelidad a los sitios de anidación. No obstante su hábitos migratorios, las tortugas marinas son muy específicas en el lugar donde
anidan, regresando a la misma playa en sucesivas migraciones para la reproducción. Pero lo que es más impresionante es que para su primera anidación, las hembras tienden a regresar a las playas donde nacieron ("filopatría"). Este hecho tiene importantes implicaciones para el manejo. Primero, cada colonia anidadora tiende a comportarse como unidad poblacional con demografía independiente. Segundo, este nivel de estructuración significa que las colonias que sufran merma o colapsos por cualquier causa (natural o antropogénica) no podrán recuperarse a través un reclutamiento natural proveniente de poblaciones externas, por lo menos dentro de las escalas de tiempo relevantes para fines de manejo.

12 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacífico mexicano
3. ANTECEDENTES En la última década se ha fortelecido la llamada Biología de la Conservación que se aboca a tres
objetivos principales: l) el mantenimiento de la viabilidad de las poblaciones y especies; 2) el mantenimiento del potencial evolutivo de las poblaciones, por medio de la diversidad genética inherente; y 3) el mantenimiento de la capacidad para la especiación. Estos objetivos se han abordado de diversos ángulos, a partir de diferentes disciplinas por lo que la Biología de la Conservación requiere escencialmente un enfoque multidisciplinario, entre ellas la Genética.
Dentro de las contribuciones que ejerce el enfoque molecular de la genética a este nuevo campo está la caracterización a nivel genética de las poblaciones para evaluar los niveles de variabilidad que contienen, tanto dentro de las poblaciones como entre ellas. El conocimiento de los niveles de variabilidad genética son importantes ya que se considera que reducciones en este parámetro reduce la capacidad para adaptación de los organismos y se incremente el riesgo de extinción (Frankham, 1995).
La capacidad para identificar las características genéticas de las poblaciones es también util para otro propósito que impacta sobre la capacidad de conservar los recursos: la demarcación de las unidades demográficas en las poblaciones naturales. La identificación de estas unidades, también conocidas como "stocks" en pesquerías, ha sido complicado anteriormente. Por ejemplo, algunos métodos usados para este propósito en especies de importancia pesquera, se han basados en la aplicación de marcas físicas, diferencias en aspectos ecológicos entre poblaciones, distribución diferencial de parásitos, características merísticas y morfométricas, carcteres fisiológicos y de comportamiento, estructuras calcificdas, citogenéticas, inmunogenéticas, proteínas y finalmente, en las últimas décadas, marcas moleculares (Carvalho y Hauser, 1995). Estos últimos se han desarrollado rápidamente, transformándose en herrarmientas con una excepcional capacidad de resolución y eficiencia, contando con unas mismas técnicas y variables aplicables a diferentes especies y a diferentes niveles de diferenciación genética.
En el ámbito del manejo de recursos naturales, la identificación de las unidades demográficas básicas es necesario para modular y enfocar los esfuerzos hacia las diferentes unidades que, al comportarse como unidades reproductoras independientes, tendrán también dinámicas de crecimiento y niveles de condición independientes. De este procedimiento no solo delimitan la distribución geográfica de las poblaciones, sino también se pueden inferir niveles de flujo génico y las relaciones evolutivas entre ellas.
No obstante, ya que las herramientas de la genética molecular permiten identificar unidades poblacionales dentro de una gama amplia de diferenciación genética- desde poblaciones gemelas hasta diferentes especies- es necesario definir y demarcar los límtes de trabajo de estas diferencias detectables, además de su significado y aplicabilidad para los fines de manejo o conservación, de acuerdo a los objetivos y la escala de tiempo de los mismos. De esta manera, el nivel óptimo de resolución del análisis genético podrá ser utilizado. En este sentido, Moritz (1994), distinguió dos distintos niveles de resolución de la demarcación genética de las poblaciones: 1) conjuntos de poblaciones que se han mantenido aislados evolutivamente durante un período prolongado y han acumulado diferencias genéticas substanciales, los cuales denominó Unidades de Significancia Evolutiva (ESU, por sus siglas en inglés) y 2) las poblaciones cuyo nivel de diferenciación no es tan marcado pero que sin embargo, al comportarse como unidades independientes reproductivamente, han acumulado discontinuidades significativas en las frecuenciasindividuales de conjuntos de alelos compartidos. Estas últimas, sinónimos de los "stocks", son denominadas unidades de manejo (UM) por su utilidad para los propósitos de la conservación y manejo de las poblaciones arriba mencionados. En muchos casos, la utilización de los marcadores genéticos para la identificación de las UM representan la única opción expedita para estos propósitos y se han constituido como puntos de apoyo para la toma de decisiones en programas de conservación y manejo (Avise, 1966), en conjunto con otras consideraciones dentro de otros ámbitos; políticos, económicos y sociales.
Aplicaciones de la genética molecular en las tortugas marinas Los trabajos precursores de genética molecular en tortugas marinas se remontan a Bonhome et al.
(1987) quienes utilizaron alozimas para estudiar la tortuga verde C. mydas. La escasa capacidad de resolución de esta técnica imposibilitó la detección de diferencias encontradas más adelante. Sin embargo, se pudo distinguir entre poblaciones del Atlántico y del Indo-Pacífico, más no entre muestras del Pacífico e Indico. La implementación de metodologías para analizar la variación genética directamente en el DNA inició con estudios de polimorfismos en las longitudes de fragmentos de restricción (RFLPs, por sus siglas en inglés) aplicado por Bowen a poblaciones

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 13 de la tortuga verde (Bowen et al., 1992). Incrementos importantes en la precisión del análisis surgieron a partir de la posibilidad de secuenciar regiones específicas del DNA, primordialmente dentro del DNA mitocondrial, gracias al desarrollo de la reacción en cadena de la polimerasa (PCR, por sus siglas en inglés). Esta técnica facilitó y abarató el estudio de la constitución genética de poblaciones de organismos. En tortugas marinas se determinó la estructura genética de la tortuga verde en el Atlántico occidental (Encalada el al., 1996), la tortuga carey en el Caribe (Bass et al., 1996), la laúd (Dutton el al., 1999) y la golfina a nivel global (Bowen el al., 1998)
Las relaciones sistemáticas y evolutivas entre poblaciones de tortugas marinas ha estado sujeto a polémicas de difícil resolución debido a que los organismos presentan rasgos morfológicos extremadamente conservados y que no se prestan facilmente a un análisis filogenético. A niveles altos de la clasificación de los quelonios sobrevivientes, las herramientas moleculares han resuelto dudas sobre la singularidad específica de la tortuga lora (Lepidochelys kempii) con respecto a su especie hermana, la tortuga golfina (L. olivacea; Bowen y Karl, 1996); la mayor cercanía evolutiva de la tortuga carey (Eretmochelys imbricata), con hábitos espongívoros, y la subfamilia Carettini (que incluye el género Caretta) de hábitos carnívoros (Bowen el al., 1993); la confirmación de hibridización entre especies (inclusive entre diferentes géneros) de tortugas marinas- Caretta con Chelonia; Caretta con Eretmochelys y Lepidochelys con Caretta (Bowen y Karl, 1996).
La identificación de caracteres genéticos que identifica poblaciones tiene varias aplicaciones que sirven, por ejemplo, para resolver algunas de las problemáticas presentadas anteriormente. Una aplicación de las técnicas moleculares ha sido en el análisis de la composición de las poblaciones (por lugar de origen) en áreas de convergencia corno sitios de alimentación o rutas migratorias (Broderick et al., 1994; Bowen el al., 1995; Bowen et al., 1996). Por ejemplo, se pudo demostrar que de las 1,000 caguamas al año capturadas incidentalmente por los palangres y redes de deriva que varios países emplean en el Pacífico del Norte, un 95% nace en colonias japonesas y el resto en playas australianas (Bowen et al., 1995). Todas convergen para su desarrollo juvenil en la zona de alimentación localizada en aguas costeras de Baja California, México. Así, la jurisdicción sobre su conservación es compleja, ya que compete a los varios países que comparten la población en diferentes fases de su ciclo vital. Mientras tanto, los impactos causados deben analizarse en función de la frecuencia relativa de las UM's en la captura y de la condición de cada una de las colonias fuente. Sobre este punto se ha estimado una estabilidad actual relativa en la población anidadora de la especie en Japón mientras que las anidaciones en Australia tienden hacia la baja. Sin embargo, para mantener niveles poblacionales saludables en ambas UM's es indispensable controlar la mortalidad de los juveniles en las pesquerías aludidas (Bolten et al., 1996).
Los niveles de flujo genético entre poblaciones también son amenos a evaluación por medio de marcadores moleculares y, en casos donde se comparan los resultados de diferenciación en loci nucleares (de herencia biparental) con los de loci mitocondriales (de herencia materna), es posible disinguir el flujo génico mediado por las hembras de. aquel mediado por los machos. Así, Karl et al. (1992) encontraron que mientras que las líneas maternas de las colonias reproductoras de la Isla Ascensión y de Surinam son disinguibles genéticamente, existe un reconocible flujo génico que afecta la composición de loci nucleares, sugiriendo un intercambio genético propiciado por el comportamiento promiscuo de machos, indistinamente de su colonia origen, sobre hembras fieles a sus sitios de anidación.
La tendencia de las hembras hacia la filopatría es otro de los aspectos que han sido elucidados con marcadores moleculares. En términos genéticos, este fenómeno es reconocible por medio de diferencias genéticas entre colonias que anidan en diferentes playas. Estudios de genética en la tortuga verde (Chelonia mydas; Encalada et al., 1992), en la carey (Eretmochelys imbricata; Bass et al., 1996) y en la caguama (Caretta caretta; Bowen et al., 1994) han demostrado que todas estas especies exhiben esta subdivisión genética entre colonias y confirman una marcada tendencia hacia el comportamiento de filopatría. Conocimiento actual sobre la genética de las especies objeto de este estudio
A pesar de que la tortuga golfina es una especie pantropical y la más abundante de todas las tortugas marinas, es también sobre la que menos se ha investigado. Esto podría obedecer al hecho de que no abunda en regiones del mundo en donde se han dirigido la mayor parte de los recursos para la investigación de estas especies. Además, una buena parte de la información que si se recoge sobre esta especie ocurre en países en vías de desarrollo donde una buena parte de la información permanece en la literatura "gris" como informes técnicos,

14 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacifico mexicano
tesis o artículos de difusión con escasa circulación. Aun así, Pritchard (1969) planteó un escenario sobre el proceso de colonización de esta especie a través de las cuencas oceánicas que ahora ocupa, y Márquez (1976) sobre la posible diferenciación de la especie dentro del Pacífico oriental.
Bowen et al. (1993), usando secuencias del citocromo b, compararon las relaciones evolutivas entre las especies de tortugas marinas, incluyendo Dermochelys y Lepidochelys. En el trabajo más completo sobre las filogenia de este grupo, Dutton et al. (1996) utilizaron secuencias del ND4-tRNA leucina y de laregión control.
Ambas especies han sido objeto de estudios recientes sobre su filogeografía global. En tortuga golfina, Bowen et al. (1998) utilizaron secuencias de la región control del DNA mitocondrial, para abordar las predicciones de Pritchard sobre los patrones de colonización de la especie en las cuencas oceánicas y demostrar una diferenciación significativa entre las grandes regiones. En laúd, Dutton et al. (1999), también usaron secuencias de la región control. En este caso, la divergencia evolutiva encontrada fué mucho más somera que en el resto de las especies estudiadas aunque existen fuertes subdivisiones a nivel global y dentro de cuencas a pesar de la extensa capacidad migratoria de la especie.
El presente es un estudio pionero para las tortugas golfina y laúd en la región. Extiende y profundiza los trabajos de genética sobre estas especies, enfocando sobre las colonias anidadoras de mayor importancia en la costa del Pacífico mexicano, que constituyen para ambas especies, de las más abundantes en el mundo.

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 15
4. OBJETIVOS DEL PROYECTO
Objetivo general Analizar las relaciones genéticas entre las principales colonias reproductoras de las tortugas golfina
(Lepidochelys olivacea) y laúd (Dermochelys coriacea) en el litoral del Pacífico mexicano.
Objetivos particulares • Instalar y validar metodologías de aislamiento de DNA a partir de muestras de tejidos de tortugas
marinas • Amplificación de regiones específicas del DNA por medio de la reacción en cadena de la
polimerasa (técnica de la PCR por sus siglas en inglés) y el subsecuente análisis comparativo entre las poblaciones estudiadas. En la tortuga golfina se enfocó sobre la región control del DNA mitocondrial además de loci de microsatélites en el DNA nuclear, en la laúd se estudiaron loci de microsatélites.
• Contrastar estos resultados, con los obtenidos en otras regiones con poblaciones de las mismas especies al igual que con el resto de las especies de tortugas marinas.
• Analizar la composición genética de las especies, en el área de estudio para validar la hipótesis de la existencia de una estructuración en colonias individuales ("stocks", unidades "demográficas" o "de manejo") con tendencias hacia la independencia reproductiva, como ha sido demostrado para otras especies de tortugas marinas y en otras regiones.
• Cuantificar los niveles de variabilidad genética y su distribución entre y dentro de las colonias reproductoras para determinar si existe algún indicio de reducción en los valores, como posible consecuencia de las drásticas declinaciones documentadas para las especies bajo estudio
• Analizar las implicaciones de la información aquí generada, en programas de conservación de tortugas marinas a nivel nacional y regional
• Generar una tesis de maestría sobre genética poblacional de la tortuga laúd (Ana Barragán Rocha) y dos sobre la tortuga golfina, una empleando secuencias de la región control del DNA mitocondrial (Raquel Briseño Dueñas) y la segunda microsatélites del DNA nuclear (Felipe López)

16 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacífico mexicano
5. MATERIALES Y METODOS
Areas de Estudio De acuerdo a lo sugerido por Norman et al. (1994), el área de estudio abarcó colonias de anidación
representativas de la distribución de la especie a diferentes escalas geográficas de la cuenca del Pacífico. Se intentó muestrear por lo menos una colonia anidadora de golfina en cada uno de los estados en la
costa del Pacífico mexicano, pero particularmente para el estudio con la región control del DNAmit, se tomaronen cuenta los siguientes criterios- a) los extremos en la distribución de su anidación en la costa de México (#1y #13, ver Figura 5.1 y Tabla 5.1), b) una playa de anidaciones masivas (La Escobilla, Oaxaca # 10), c) unaplaya con antecedentes de arribazones, con una condición actual de reducción poblacional severa (Piedra deTlalcoyunque, Guerrero #7) seleccionada por haber reunido arribazones sustanciales todavía a finales de los
Figura 5.1 Distribución de los sitios de muestreo (ver Tabla 5.1 para la identificación de las playas de anidación).
60's, d) la siguiente escala geográfica, comprendiendo otros sitios del Pacífico americano (la coloniaanidadora de golfinas en Playa Nancite, Costa Rica #I4), de gran importancia regional por ser una de lascolonias de arribazones de mayor tamaño en el ámbito mundial, e) para tener una cobertura de cuenca, seutilizaron muestras del Pacífico occidental (la colonia anidadora de Arnhem Land, Northern Territory,Australia #17, en la región sur-occidental del Pacífico) (Tabla 5.1 y Fig. 5.1). Finalmente, para realizar unacomparación de los niveles de diferenciación genética entre especies del mismo género, se incorporarontambién muestras de L. kempii proporcionadas por el Dr. Brian Bowen.
Para la obtención de muestras de tejidos de tortuga golfina, también se obtuvo la colaboración debiólogos de campo, residentes en los campamentos de conservación durante las temporadas de anidación, en lossitios de muestreo. Sus nombres aparecen en la sección de agradecimientos.
Para la tortuga laúd, se muestrearon las cuatro principales colonias anidadoras del país: � Playón de Mexiquillo, Mich. (#6)

16 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacífico mexicano
5. MATERIALES Y METODOS
Areas de Estudio De acuerdo a lo sugerido por Norman et al. (1994), el área de estudio abarcó colonias de anidación
representativas de la distribución de la especie a diferentes escalas geográficas de la cuenca del Pacífico. Se intentó muestrear por lo menos una colonia anidadora de golfina en cada uno de los estados en la
costa del Pacífico mexicano, pero particularmente para el estudio con la región control del DNAmit, se tomaronen cuenta los siguientes criterios- a) los extremos en la distribución de su anidación en la costa de México (#1 y #13, ver Figura 5.1 y Tabla 5.1), b) una playa de anidaciones masivas (La Escobilla, Oaxaca # 10), c) una playa con antecedentes de arribazones, con una condición actual de reducción poblacional severa (Piedra de Tlalcoyunque, Guerrero #7) seleccionada por haber reunido arribazones sustanciales todavía a finales de los
Figura 5.1 Distribución de los sitios de muestreo (ver Tabla 5.1 para la identificación de las playas de anidación).
60's, d) la siguiente escala geográfica, comprendiendo otros sitios del Pacífico americano (la colonia anidadora de golfinas en Playa Nancite, Costa Rica #I4), de gran importancia regional por ser una de las colonias de arribazones de mayor tamaño en el ámbito mundial, e) para tener una cobertura de cuenca, se utilizaron muestras del Pacífico occidental (la colonia anidadora de Arnhem Land, Northern Territory, Australia #17, en la región sur-occidental del Pacífico) (Tabla 5.1 y Fig. 5.1). Finalmente, para realizar una comparación de los niveles de diferenciación genética entre especies del mismo género, se incorporaron también muestras de L. kempii proporcionadas por el Dr. Brian Bowen.
Para la obtención de muestras de tejidos de tortuga golfina, también se obtuvo la colaboración de biólogos de campo, residentes en los campamentos de conservación durante las temporadas de anidación, en los sitios de muestreo. Sus nombres aparecen en la sección de agradecimientos.
Para la tortuga laúd, se muestrearon las cuatro principales colonias anidadoras del país: • Playón de Mexiquillo, Mich. (#6)

Informe Final F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 17
Tabla 5.1 Colonias reproductoras de L. olivacea y D. coriacea que fueron muestreadas en la cuenca del Pacífico
#en mapa
clave BITMAR
País Estado Playa Especie Colección donde se guardaron
tejidos
Temporada Núm muestras
1
4030 México Sinaloa El Verde ohvacea BITMAR, ICMyL UNAM
94, 95 56
2 5010 México Nayarit Platanitos ohvacea BITMAR, ICMyL UNAM
96, 97 34
3 6030 México Jalisco La Gloria ohvacea BITMAR, ICMyL UNAM
94, 95, 96 112
4 7070 México Colima Chupadero ohvacea BITMAR, ICMyL UNAM
96 73
5 8030 México Michoacán Maruata ohvacea BITMAR, ICMyL UNAM
97 10
6 8100 México Mexiquillo coriacea Laboratorio de Tortugas marinas, FC UNAM
94, 95 41
ohvacea BITMAR, ICMyL UNAM
95, 96 22
7 9030 México Guerrero Piedra de Tlalcoyunque
ohvacea BITMAR, ICMyL UNAM
95 41
8 9110 México Tierra Colorada coriacea Laboratorio de Tortugas marinas, FC UNAM
95 55
ohvacea BITMAR, ICMyL UNAM
96 11
9 10020 México Oaxaca San Juan Chacahua coriacea Laboratorio de Tortugas marinas, FC UNAM
95
33
ohvacea BITMAR, ICMyL UNAM
96 31
10 10030 México Oaxaca La Escobilla ohvacea BITMAR, ICMyL UNAM
93 22
11 10040 México Oaxaca Barra de la Cruz coriacea Laboratorio de Tortugas marinas, FC UNAM
95 36
12 10050 México Oaxaca Morro Ayuta ohvacea BITMAR, ICMyL UNAM
96 35
13 11020 México Chiapas Puerto Arista ohvacea BITMAR, ICMyL UNAM
95 30
14 50001 Costa Rica ND Nancite ohvacea BITMAR, ICMyL UNAM
93 18
15 50003 Costa Rica Guanacaste Playa Naranjo ohvacea BITMAR, ICMyL UNAM
88 5
16 52001 Nicaragua ND Playa La Flor ohvacea BITMAR, ICMyL UNAM
95 27
17 90000-90029
Océano pacifico
ND zona de alimentación Pacífico Oriental*
ohvacea BITMAR, ICMyL UNAM
93, 95 48
18 100001 Australia Northern Territory
Fog Bay ohvacea Laboratorio de Tortugas marinas, FC UNAM
ND 11
Total 782 ND= no determinado
*= aunque también se obtuvieron muestras de esta área de alimentación y los tejidos se sumaron a la colección de material genético, no se analizaron para los fines de este proyecto

18 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacífico mexicano
Tabla 5.2 Ubicación geográfica de los sitios de muestreo en México
Clave BITMAR
Estado Municipio Código INEGI
playa Latitud N (grados)
Latitud N (mins)
Longitud W (grados)
Longitud W (mins)
4030 Sinaloa Mazatlán 012 El Verde 23 26 106 34 5010 Nayarit Compostela 004 Platanitos 21 20 105 14 6030 Jalisco Tomatlán 100 La Gloria 19 56 105 277070 Colima Tecomán 009 Chupadero 18 41 103 47 8020 Michoacán Aquila 008 Colola 18 18 103 26 8030 Michoacán Aquila 008 Maruata 18 15 103 21 8100 Michoacán Aquila 008 Mexiquillo 18 5 102 48 9030 Guerrero Tecpan de
Galeana 057 Piedra de
Tlalcoyunque17 16 101 3
9110 Guerrero Cuajinicuilapa 023 Tierra Colorada
16 3 98 43
10020 Oaxaca San Pedro Tututepec
334 San Juan Chacahua
15 58 97 47
10030 Oaxaca Sta. María Tonameca
439 La Escobilla 15 43 96 45
10040 Oaxaca Santiago Astata 453 Barra de la Cruz
15 50 95 57
10050 Oaxaca Santiago Astata 453 Morro Ayuta 15 53 95 46 11020 Chiapas Tonalá 097 Puerto Arista 15 55 93 48
NA= no aplicable
• Tierra Colorada, Gro. (#8) • Chacahua (#9) y Barra de la Cruz (#11 ), Oax. Los muestreos de tejidos para esta especie fueron realizados por Ana Barragán con el apoyo logístico
y de personal estudiantil del Laboratorio de Tortugas Marinas (Facultad de Ciencias, UNAM).
Obtención y preservación de muestras Debido a la situación de ambas especies, catalogadas como especies en peligro de extinción, se tuvo
especial cuidado de impactar al mínimo las poblaciones. De tal manera, se aprovecharon métodos de obtención de tejido que no representara el sacrificar ningún animal- sangre extraída de hembras anidadoras o tejido (músculo, corazón o hígado) de crías muertas por causas naturales en el nido. En todos momentos se aseguró que las muestras de crías provenían de madres distintas.
Muestras de tejido sólido Las muestras de tejido sólido (hígado, corazón o músculo), se colectaron de crías que morían antes de
la eclosión y que son recuperadas durante la revisión rutinaria de los nidos. De esta manera se elimina la necesidad de lastimar o sacrificar organismos durante la colecta. El material fue preservado de acuerdo al protocolo de Proebstel et al. (1993), modificado por Bowen (com. pers.). El fijador utilizado es conocido como SED, (por las siglas en ingles de sus componentes), y consiste en una solución saturada de NaCl, con EDTA y DMSO al 20%, amortiguado a pH 7.5. Las muestras mantenidas en esta solución, se pueden conservar por varios meses a temperatura ambiente o en refrigeración hasta por un año
Muestras de sangre Las muestras de sangre se extrajeron del seno cervical de hembras anidadoras, usando la técnica
descrita por Owens y Ruiz (1980), y se preservaron en un buffer lítico de Sulfato Dodecil de Sodio (SDS) conteniendo 100 mM Tris.HCl, 100 mM EDTA, 10 mM Na CI, 0.5% SDS a un pH de 8.0. La proporción sangre-amortiguador, se mantuvo en una relación de 1:10, de acuerdo a lo recomendado por White y Densmore (1992). Las muestras para el análisis genético, se conservan en este buffer lítico y a temperatura ambiente por períodos >1 año, sin

Informe Final - F. Alberto Abreu Grobois. Raquel Briseño Dueñas, Ana R. Barragán Rocha 19 una degradación sustancial del DNA.
Análisis bioquímico
Extracción de DNA En este trabajo, el DNA genómico total (suma del mitocondrial y nuclear) fue aislado por el método
de extracción secuencial con fenol/ cloroformo/ alcohol isoamílico, seguido por precipitación en frío con alcohol etílico según la técnica descrita por Hillis y Moritz (1990). Los productos del DNA, se almacenaron siempre en una solución de Tris-EDTA amortiguada a un pH = 8.0 y una temperatura de -20°C.
Las muestras de tortuga golfina fueron extraídas por Raquel Briseño, en el Laboratorio de Genética, de la Unidad Académica"Mazatlán" del ICMyL-UNAM y algunas en el Laboratorio de Análisis Genético del Dr. Brian Bowen, del Instituto de Desarrollo Biotecnológico de la Universidad de Florida en Gainesville durante una estancia de trabajo. Las de laúd fueron extraídas por Ana Barragán en la Facultad de Ciencias y en el laboratorio de Genética y Evolución del Instituto de Ecología, ambos de la UNAM.
Cebadores para la amplificación del DNA A partir del DNA total se procedió a amplificar los loci específicos que habían sido seleccionados
debido a su polimorfismo en las especies, usando cebadores ("primers") desarrollados a partir de tortugas marinas en su mayoría (con la excepción de N32 que se derivó de tortugas terrestres, Tabla 5.3). La identificación de loci polimórficos en la tortuga golfina fue lograda por medio de una estancia de trabajo del responsable con el Dr. Peter Dutton en el Soutwest Fisheries Science Center, NMFS-NOAA. En la tortuga laúd se conocía de antemano el grado de variabilidad de estos loci (Dutton, 1995).
Reacción en cadena de la polimerasa (PCR)
Región control en tortugas golfina Las condiciones de reacción para la amplificación por PCR están descritas en las Tablas 5.4 y 5.5.
Los productos de la amplificación para los distintos loci fueron primero analizados en geles de 10 x 10 cm de agarosa al 3% para verificación de existencia de producto y de su tamaño con respecto a estándares de tamaño molecular (medido en pares de bases, bp) el cual generalmente fue ΦX174/HaellI que presenta varios fragmentos entre 72 y 1353 bp's (Figura 5.2)
Al final de las amplificaciones, los productos fueron limpiados con filtros Millipore 30,000 MV® Ultra-Free-MC y enviados a secuenciar en una estación robotizada Applied Biosystems 800 y 373A en el DNA Sequencing Core de la Universidad de Florida, Las secuencias obtenidas fueron alineadas inicialmente por el mismo secuenciador usando el algoritmo de Sequencher y posteriormente con el programa Clustal X (Thompson
Figura 5.2 Electroforesis en agarosa de los productos de PCR para la región control del DNAmt de tortuga golfina

20 Genética poblactonal y filogeografia de las tortugas marinas golfina y laúd en el Pacífico mexicano
Tabla 5.3 Cebadores utilizados para ambas especies y sus secuencias
Epecie en la que se aplico
Desig. cebador
Locus Secuencia Ref.
Lepidochelys olivacea
LTCM1 Región control del DNA mitocondrial (hebra pesada) 5’CCCAAAACCGGAATCCTAT 3’ (F)
Allard et al. (1994)
Lepidochelys olivacea HDCM1
Región control del DNA mitocondrial (hebra pesada)
5’ ACTACCGTATGCCAGGTTA 3’ (R) Allard et al. (1994)
Lepidochelys olivacea Cm72 Microsatélite en DNA nuclear
5’ CTATAAGGAGAAAGCGTTAAGACA 3’(F) 5’ CCAAATTAGGATTACACAGCCAAC 3’ (R)
FitzSimmons et al. (1995)
Lepidochelys olivacea Cm84 Microsatélite en DNA nuclear
5’ TGTTTTGACATTAGTCCAGGATTG 3’ (F) 5’ATTGTTATAGCCTATTGTTCAGGA 3’ (R)
FitzSimmons et al. (1995)
Lepidochelys olivacea Cc141 Microsatélite en DNA nuclear
5’ TAGTACGTCTGGCCTGACTTTC 3’ (F) 5’ CAGCAGGCTGCTAGTTCTCCA 3’ (R)
FitzSimmons et al. (1995)
Dermochelys coriacea
DC99 Microsatélite en DNA nuclear 5’ CACCCATTTTTTCCCATTG 3’ (F) 5’ ATTTGAGCATAAGTTTTCGTGG 3’ (R) Dutton (1995)
Dermochelys coriacea
Nigra32 Microsatélite en DNA nuclear
5’ CGTGTGTTTGGACAGAAGATGAAC 3’ (F) 5’ AGGCAAAGCACCTGCAAATC 3’ (R)
Louis com. Pers. Citado en dutton
(F)= para amplificación de la hebra pesada ("Forward") (R)= para amplificación de la hebra ligera ("reverse")
Tabla 5.4 Protocolo para el termociclaje de la amplificación de la región control del DNAmit en tortuga golfina
Paso Segmento Temp. (°C)
Tiempo (min)
Total Ciclos
1 95.0 3:00 1 2 1 95.0 01:00 35
2 50.0 01:00 3 72.0 01:00 3 72.0 03:00 FIN
et al., 1994). La amplificación y secuenciación de las muestras de golfina se llevaron a cabo durante una estancia en el laboratorio del Dr. Bowen, en la Universidad de Florida.
Microsatélites del DNAnuclear en tortugas golfina Para la amplificación de los loci de microsatélites en tortugas golfinas se utilizó el mismo
protocolo y condiciones del termociclador que para la región control. La amplificación de las muestras de El Verde, Sinaloa y de Escobilla, Oaxaca fueron realizadas en el Laboratorio del Dr. Dutton (SW Fisheries Science Center del Servicio de Pesquerías Marinas de EUA, SWFSC-NMFS-NOAA), mientras que el resto fueron procesadas en el Laboratorio de Genética de la Unidad Académica Mazatlán del ICMyL.
Mientras que los productos de la amplificación de las muestras procesadas en el SWFSC fueron analizadas siguieron el protocolo descrito para el material de tortuga laúd, el resto fue corrido en el LGUAM en geles de poliacrilamida de 8% y visualizadas con unción de plata de acuerdo a la técnica descrita por Bassam y Caetano-Anolles (1993, ver figura 5.3). Una vez obtenida la resolución de los electroferotipos se procedió a identificar los alelos, catalogarlos y interpretar los genotipos, con base a patrones de estándares de peso molecular. Todas las designaciones se realizaron con base en el número correspondiente de pares de bases (bp por sus siglas en inglés) y así se catalogaron los genotipos de cada individuo analizado en la Base de Datos. Todos los análisis de microsatélites de tortugas golfina fueron llevados a cabo por el Cand. MC Felipe López.

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas. Ana R. Barragán Rocha 21
Tabla 5.5 Protocolo para el cocktail de reacción para la PCR, basado en el protocolo recomendado en Moritz et al (1997)
Reactivos Conc Stock
Concentraciónfinal en reacc
C a n t i d a d ,por tubo
(µl)
Agua dest EsteriI* - - - - 9.6*dNTPs 2 mM 0.2 mM 2.5 MgCl2 10 mM 3.5 mM 5Cebador 1 10 µM 0.48 µM 1.2 Cebador 2 10 µM 0.48 µM 1.2 10Xbuffer 10 X 1 X 2.5 Polimerasa Taq 1 U 1 U 1
TEMPLETE DNA 2 Volúmen total (µl) 25
Microsatélites del DNAnuclear en tortugas laúd
Las amplificaciones por medio de la PCR de microsatélites en tortuga laúd (DC99 y N32), se realizó en el el laboratorio del Dr. Dutton por Ana Barragán para las muestras de laúd del Pacífico mexicano. Se utlizó una variante denominada " Touchdown PCR", la cual consiste en cuatro fases:
1) Desnaturalización del DNA a 95° C durante 2 minutos. 2) 30 ciclos de desnaturalización a 95° C durante 45 segundos, fusión del primer ("annealing")
durante 2 min. y extensión a 72° C durante 2 min. Entre cada ciclo la temperatura de alineación disminuye
Figura 5.3 Gel de poliacrylamida al 8% resolviendo los electroferotipos de Cm84 (carriles 1-11, 13-15 y 17) y Cm72 (16 y 18). El carril 17 lleva una mezcla de ambos loci.

22 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacífico mexicano
0.5° C, por lo que la temperatura inicial de alineamiento es de 60° C y la final es de 45° C. 3) 15 ciclos con temperatura de alineación constante de 55° C (sin disminución) (95°C 45 seg/
55°C 2 min/ 72°C 2 min). 4) Extensión final a 72° C durante 5 min. La técnica del Touchdown permite optimizar los fragmentos generados en las temperaturas de
fusión ("annealing") más altas que son las más apegadas al sitio objetivo de la reacción. La PCR fue realizada con un termociclador Perkin Elmer GeneAmp 9600 y cebadores marcados con tinciones fluorescentes para una detección diferencial en el secuenciador. Los productos amplificados se corrieron en geles de poliacrilamida 6% de 0.4 mm de espesor, dentro de un sistema secuenciador de DNA Applied Biosystems 370A, el cual separa los fragmentos de DNA por tamaños, con una resolución de 1 par de bases.
Los alelos de loci de microsatélites se distinguen por sus diferentes tamaños, el análisis de los alelos por locus para las muestras amplificadas se realizó usando el programa GENESCAN 672, el cual identifica las bandas en el gel de poliacrilamida por su fluorescencia y las compara con un marcador de tamaño estándar para la asignación de los diferentes alelos.
Análisis estadístico de la información genética
Región control El análisis estadístico de la información genética de este locus, dentro y entre las colonias anidadoras,
fue inferido por la comparación de las secuencias aplicando el programa Arlequín versión 1.1 (Schneider et al., 1997).
Variabilidad genética Indice de diversidad haplotípica (h). Equivalente a la heterocigosidad esperada para datos
diploides, estima la variabilidad genética en términos del número de haplotipos presentes en la muestra y sus respectivas frecuencias. Este índice, es definido como la probabilidad de que dos haplotipos de la muestra, elegidos al azar, sean diferentes y se expresa por la fórmula
∑=
−−
=k
ipi
nnH
1
2 )1(1
donde n es el tamaño de la muestra, k es el número de haplotipos y Pi , es la frecuencia del haplotipo imo de la muestra (Nei, 1987).
Indice de diversidad nucleotídica (π). Determina la heterocigosidad o diversidad genética al nivel
de nucleótidos, por medio de las diferencias entre secuencias de haplotipos, estimando la probabilidad de que dos nucleótidos homólogos escogidos al azar sean diferentes (Nei y Li, 1979, Nei y Tajima, 1981). Se obtiene valorando el número promedio de nucleótidos diferentes por sitio entre dos secuencias con la siguiente relación:
L
dpp ijjiij
k
in
∑∑<== 1π
donde las p's son las frecuencias de cada uno de los nucleótidos i, y j que son comparados, des la distancia genética entre nucleótidos, y L es el número de loci.
La determinación del valor promedio se derivó del promedio de los valores de π para cada una de las colonias, mientras que el valor global se obtuvo agrupando solamente las secuencias.
Diferenciación genética Distancias genéticas. Es medida por el número de substituciones de nucleótidos entre un par de
secuencias como el número de substituciones observadas entre dos secuencias. También se puede utilizar la proporción de

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas. Ana R. Barragán Rocha 23
diferencias entre un par de secuencias, dividiéndo por L, el número de loci.
Número promedio de diferencias de bases entre las secuencias. Es el número promedio de diferenciaspor sitio. Es decir, el número promedio de nucleótidos diferentes por sitio entre dos haplotipos (p). y se determina por la fórmula:
∧
<=
∧
∑∑= ijjiij
k
idpp
1π
donde d es un estimado del número de mutaciones que ha ocurrido desde la divergencia de los
haplotipos i y j, k es el número de haplotipos y p es la frecuencia del haplotipo i. Un análisis adicional aplicado, fue la corrección a los valores de distancia genética según el modelo
de dos parámetros de Kimura (Kimura, 1980). Este estadístico toma en cuenta que no todas las substituciones tienen la misma probabilidad de ocurrir y además corrige por las diferencias entre las tasas de substitución transicionales y transversionales. Estructura genética de la población inferida por el Análisis de Varianza Molecular (AMOVA)
La determinación de la distribución de la diversidad genética distribuida dentro y entre las colonias anidadoras, fue estimada con un análisis de varianza molecular implementado en el programa ARLEQUIN (versión 1.1, de acuerdo a Excoffier et al., 1992). Este mismo algoritmo, calcula los niveles de por permutaciónno paramétrica.
Las comparaciones de los valores de Φ st’s entre pares de colonias, También se puede utilizar el valor obtenido del AMOV A a manera de medir distancias genéticas entre
la poblaciones, aplicando una ligera transformación al tiempo de divergencia de la población, para linealizar la distancias (Reynolds et al 1993, Slatkin.1995). Los valores de la comparaciones por pares de poblaciones de la Φst's son expresados en una matriz. La distribución nula de estos valores bajo la hipótesis de no-diferencia, entre las poblaciones fue obtenida por permutaciones (bootstrapping) de los haplotipos entre todas las poblaciones. El valor P de la prueba es la proporción de permutaciones que dan a un valor mas grande o igual al observado.
Estimación del flujo génico entre poblaciones (Nm). Los resultados del AMOVA también fueron utilizados para cuantificar el flujo génico por vía
materna (aproximación de la tasa de migrantes por generación), por la comparación entre pares de las colonias anidadoras. Para ello, se utilizó la relación: )1/1(2
1 −Φ= STemN
donde N es el tamaño efectivo de la población y ni es la tasa de de migración Para estae estadística se uso el parámetro ΦST del programa AMOVA. incluido en el paquete
estadístico Arlequín.
Compraración de frecuencias haplotípicas y sus intervalos de confianza Utilizando los tamaños de muestra de cada colonia y las frecuencias haplotípicas observadas, se
determinaron los errores std. al 95% por medio de una prueba de remuestreo numérico con reemplazo o, "bootstrapping", como se le denomina en inglés (Weir, 1996). Con este procedimiento se generan nuevos estimados con los que se obtiene una aproximación de la distribución teórica de los datos. El procedimiento se aplicó a todos los datos (no solamente a los haplotiopos de la región control, usando 1000 remuestreos aleatorios a partir de la matriz original. Este método de análisis estadístico es uno de los mas empleados en estudios de genética poblacional, debido a la naturaleza de estos trabajos en los que difícilmente se puede analizar un número muy

24 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacífico mexicano grande de muestras (por los altos costos del estudio molecular y de que en muchos casos no se puede obtener una mayor cantidad de muestras).
Microsatélites Se obtuvieron las frecuencias de los alelos observados asi como de los genotipos con base a los
alelomorfos detectados por medio de la separación electroforética de las muestras analizadas. A partir de estos datos de derivó también la estimación de diversidad génica, medida como heterocigosidad observada (ho) y esperada (he). Mientras que la primera medida representa la proporción de heterocigotos observados, la segunda se deriva a partir de las frecuencias alélicas, bajo el supuesto de que la población esté en equilibrio Hardy-Weinberg.
)1( 2
1pih
k
ie ∑
=
−=
Los valores de heterocigosidad por locus fueron también empleados para calcular valores
promedio, entre todas las poblaciones para un locus (en tortugas golfina) y para cada población para todos los loci analizados (ambas especies)
Personal participante • Dr. F. Alberto Abreu Grobois (Laboratorio de Genética y BITMAR, Unidad Académica
Mazatlán, ICMyL.-UNAM) responsable global del proyecto
Estudio de tortuga golfina - M. en C. Raquel Briseño Dueñas (Laboratorio de Genética y BITMAR, Unidad Académica
Mazatlán, ICMyL-UNAM)- responsable directa del estudio poblacional de la tortuga golfina. • Cand. M. en C. Felipe Javier López Chávez (Centro de Ecología Costera, Universidad de
Guadalajara en Melaque, Jalisco) estudiante responsable de la porción del estudio con microsatélites de DNA de la tortuga golfina. Se incorporó al proyecto en 1998.
• Dr. Brian Bowen (BEECS Genetic Analysis Core, Universidad de Florida) colaborador y contraparte internacional, también suministró el uso de laboratorio de genética y de técnicos adscritos a su cargo para algunos de los análisis genéticos de tortuga golfina
• Dr. Peter Dutton (SW Fisheries Science Center, NMFS-NOAA) colaborador y contraparte internacional, facilitó el uso del laboratorio de genética y de personal técnico a su cargo para algunos de los análisis genéticos de tortuga laúd. Asimismo participa en el desarrollo de metodologías de microsatélites para el análisis molecular de muestras de tortuga golfina.
Estudio de tortuga laúd -
• Cand. M. en C. Ana Rebeca Barragán Rocha. (Laboratorio de Tortugas Marinas, Fac. Ciencias, UNAM) responsable directa del estudio genético de la tortuga laúd, también participó en colectas de muestras de tortuga golfina
• Cand. M. en C. Adriana Laura Sarti Martínez (Laboratorio de Tortugas Marinas, Fac. Ciencias, UNAM)- especialista en tortuga laúd, apoyo logístico y coordinación de estudiantes-colaboradores para el muestreo de tortuga laúd
• Dr. Peter Dutton (SW Fisheries Science Center, NMFS-NOAA) colaborador y contraparte internacional, facilitó el uso del laboratorio de genética y de personal técnico a su cargo para algunos de los análisis genéticos de tortuga laúd. Asimismo participa en el desarrollo de metodologías de microsatélites para el análisis molecular de muestras de tortuga golfina.

nforme Final - F. Alberto Abreu Grobois. Raquel Briseño Dueñas, Ana R. Barragán Rocha 25
6. RESULTADOS Y DISCUSION
Tortuga golfina
Variación en la región control del DNA mitocondrial (por Raquel Briseño Dueñas y F. Alberto Abreu Grobois)
(nota: cualquier mención a resultados en esta sección deberá hacer referencia a R. Briseño Dueñas (1998)Variación genética en la región control del ADN mitocondrial de poblaciones de la tortuga golfina,Lepidochelys olivacea, en el Pacífico oriental y las implicaciones para su conservación. Tesis deMaestría. Universidad Autónoma de Sinaloa)
Secuencias Se obtuvieron secuencias de las muestras provenientes de 7 playas de anidación de tortuga golfina,
distribuidas en una gran área de la cuenca del Pacífico, incluyendo la de la población en el norte deAustralia para fines comparativos. También se incorporaron al análisis secuencias de la tortuga lora (L. kempii) para estimar dimensiones de las diferencias interespecíficas usando este locus. Dentro de esteconjunto de secuencias, se identificaron 14 haplotipos distintos en L. olivacea, mismos que se presentan alineados en la Tabla 6.1. Para identificar a cada haplotipo, se designó un código que incluye las siglas dela especie (p. ej., LO para L. olivacea) y una letra. Los haplotipos asignados a las playas de anidación delPacífico oriental (México y Costa Rica) fueron catalogados con las letras B, C, E. E, G, I, K, M, N, O, P y Z mientras que los haplotipos del Pacífico occidental (Australia) con las letras A y D.
Análisis estadístico de los resultados
Niveles de variabilidad genética y su distribución La lista de la distribución geográfica de los haplotipos y las frecuencias a las que se encontraron en
cada colonia reproductora, se compilaron en la Tabla 6.1. El haplotipo "K" fue el dominante en todas laspoblaciones del Pacífico oriental. La distribución de las frecuencias de este haplotipo en al área de estudio, distinguió a dos

26 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacifico mexicano Tabla 6.1 Secuencias de los haplotipos encontrados en poblaciones de tortuga golfina (LO) en la cuenca del Pacífico, además de una representativa de la tortuga lora (LK) tabuladas por el programa GeneDoc (Karl y Karl. 1997). Los números en la parte superior de cada bloque indican el número de la posición, mientras que la secuencia representa la «secuencia de consenso». Los sitios con letra minúscula indican loci polimórficos, los espacios en blanco representan sitios abajo del % mínimo de consenso. Los sitios o loci sin variación se presentan en fondo negro con letras blancas, los sitios con fondo gris y letras blancas señalan sitios con variación mínima y los sitios de fondo gris con letras negras representan sitios de alta variabilidad. Esta representación simplifica la identificación de los sitios variables dentro de secuencias muy conservadas como las de tortugas marinas. Los guiones representan "gaps" de alineamiento. Nota: en la base de datos el número de gaps al inicio de las secuencias es mayor debido a que se incluyó el número necesario para alinear las secuencias con el inicio de la región control del DNA mitocondrial de las tortugas.
5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 CTTaATTCTCTGCCGTGCACAACAGAACAATAGCCATAcTATCTATCTATGTATTATTGTACATCTACTT LO-A : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70
LO-D : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70LO-B : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70LO-C : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70LO-E : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70LO-F : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70LO-G : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70LO-I : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70LO-K : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70LO-M : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70LO-N : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70LO-O : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70LO-P : --------- CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 70LO-Z : --------C CTTAATTCTCTGCCGTGCACAACAGAACAATAGCCATACTATCTATCTATGTATTATTGTACATCTACTT : 71LK-LK3B : --------- CTTTATTCTCTGCCGTGCACAACAGAACAATAGCCATATTATCTATCTATGTATTATTGTACATCTACTT : 70
85 90 95 100 105 110 115 120 125 130 135 140 145 150 155 160
ATTTACCgtTAGCATATGATCAGTAATaTTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAACT LO-A : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAACT :150LO-D : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAACT :150LO-B : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAACT :150LO-C : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAAAT :150LO-E : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAACT :150LO-F : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAAAT :150LO-G : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAAAT :150LO-I : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAACT :150LO-K : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAACT :150LO-M : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAACT :150LO-N : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAACT :150LO-O : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAACT :150LO-P : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAACT :150LO-Z : ATTTACCGTTAGCATATGATCAGTAATATTGTCGATTAATTTGGCTTTAAACATAAAAATTCATTAATTTTACATAAACT :151LK-LK3B : ATTTACCACTAGCATATGATCAGTAATGTTGTCGATTAATTTGACCTTAAACATAAA-ATTCAATAATTTTACATAAACT :149
165 170 175 180 185 190 195 200 205 210 215 220 225 230 235 240 gTTTTtACCAAAtGAATATTATAAaGaTgATAATAGTGAAATGATATAGGACATAAAATTAAAC ATTATTCTCgACCAT
LO-A : GTTTTAACCAAATGAATATTATAAAGATGATAATAGTGAAATGATATAGGACATAAAATTAAACCATTATTCTCAACCAT :230LO-D : GTTTTAACCAAATGAATATTATAAGGATGATAATAGTGAAATGATATAGGACATAAAATTAAACCATTATTCTCAACCAT :230LO-B : GTTTTTACCAAATGAATATTATAAAGATGATAATAGTGAAATGATATAGGACATAAAATTAAACTATTATTCTCGACCAT :230LO-C : GTTTTTACCAAATGAATATTATAAAGATGATAATAGTGAAATGATATAGGACATAAAATTAAACTATTATTCTCGACCAT :230LO-E : GTTTTTACCAAATGAATATTATAAAGATGATAATAGTGAAATGATATAGGACATAAAATTAAACTATTATTCTCGACCAT :230LO-F : GTTTTTACCAAATGAATATTATAAAGATGATAATAGTGAAATGATATAGGACATAAAATTAAACTATTATTCTCGACCAT :230LO-G : GTTTTTACCAAATGAATATTATAAAGATGATAATAGTGAAATGATATAGGACATAAAATTAAACTATTATTCTCGACCAT :230LO-I : GTTTTTACCAAATGAATATTATAAAGATGATAATAGTGAAATGATATAGGACATAAAATTAAACCATTATTCTCGACCAT :230LO-K : GTTTTTACCAAATGAATATTATAAAGATGATAATAGTGAAATGATATAGGACATAAAATTAAACCATTATTCTCGACCAT :230LO-M : GTTTTTACCAAATGAATATTATAAAGATGATAATAGTGAAATGATATAGGACATAAAATTAAACCATTATTCTCGACCAT :230LO-N : GTTTTTACCAAATGAATATTATAAAGATGATAATAGTGAAATGATATAGGACATAAAATTAAACCATTATTCTCGACCAT :230LO-O : GTTTTTACCAAATGAATATTATAAAGGTGATAATAGTGAAATGATATAGGACATAAAATTAAACCATTATTCTCGACCAT :230LO-P : GTTTTTACCAAATGAATATTATAAAGATGATAATAGTGAAATGATATAGGACATAAAATTAAACCATTATTCTCGACCAT :230LO-Z : GTTTTTACCAAATGAATATTATAAAGATGATAATAGTGAAATGATATAGGACATAAAATTAAACCATTATTCTCGACCAT :231LK-LK3B : ATTTTAACCAAACGAATATTATAAAGGTAATAATAGTGAAATGATATAGGACATAAAATTAAACCATTATTCTCAACCAT :229

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 27
Tabla 6.1 (cont.)
325 330 335 340 345 350 355 360 365 370 375 380 385 390 395 400
405 410 415 420 425 430 435 440 445 450 455 460 465 470 475 480
GAATATCGTCaCAGTAATTGGTtATTTCTTAGTTCAaCTCATCACGAGAAATAAGCAAtCCTTGTTAACAAGATACaACA LO-A : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :310LO-D : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :310LO-B : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :310LO-C : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :310LO-E : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :310LO-F : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :310LO-G : GAATATCGTC-CAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :309LO-I : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :310LO-K : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :310LO-M : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :310LO-N : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :310LO-O : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :310LO-P : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :310LO-Z : GAATATCGTCACAGTAATTGGTTATTTCTTAGTTCAACTCATCACGAGAAATAAGCAATCCTTGTTAACAAGATACAACA :311LK-LK3B : GAATATCGTCACAGTAATTGGTCATTTCTTAGTTCAGCTCATCACGAGAAATAAGCAACCCTTGTTAACAAGATACGACA :309
TTACCAGTTTCAgGCcCATTAAAcTAcgaCGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAgGCACATTAA LO-A : TTACCAGTTTCAGGCCCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :390LO-D : TTACCAGTTTCAGGCCCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :390LO-B : TTACCAGTTTCAGGCCCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :390LO-C : TTACCAGTTTCAGGCCCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :390LO-E : TTACCAGTTTCAGGCCCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :390LO-F : TTACCAGTTTCAGGCCCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :390LO-G : TTACCAGTTTCAGGCTCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :389LO-I : TTACCAGTTTCAGGCTCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :390LO-K : TTACCAGTTTCAGGCCCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :390LO-M : TTACCAGTTTCAGGCCCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :390LO-N : TTACCAGTTTCAGGCCCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :390LO-O : TTACCAGTTTCAGGCCCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :390LO-P : TTACCAGTTTCAGGCCCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :390LO-Z : TTACCAGTTTCAGGCCCATTAAACTACGACGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAGGCACATTAA :391LK-LK3B : TTACCAGTTTCAAGCCCATTAAATTATACCGTACATAACTGATCTATTCTGGCCTCTGGTTGTTTTTTCAAGCACATTAA :389
gGtA TGAAGTTCATTCGTTCCTCTTTAAAAGGCCTtTGGTTcgacgataAATGAGTTCTATACATTgAATTTATAACC LO-A : GGTAA-TGAAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :469LO-D : GGTAG-TGAAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGA-AAATGAGTTCTATACATTGAATTTATAACC :468LO-B : GGTAA-TGAAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :469LO-C : GGTAA-TGAAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :469LO-E : GGTAG-TGAAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :469LO-F : GGTAG-TGAAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :469LO-G : GGTAG-TGAAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :468LO-I : GGTAA-TGAAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :469LO-K : GGTAA-TGAAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :469LO-M : AGCAA-TGAAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :469LO-N : GGTAAATGAAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :470LO-O : GGTAAATGAAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :470LO-P : GGTAAATGCAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :470LO-Z : AGTAA-TGCAGTTCATTCGTTCCTCTTTAAAAGGCCTTTGGTTCGACGATAAATGAGTTCTATACATTGAATTTATAACC :470LK-LK3B : AGTAA-TGAAGTTCATTCGTTCCTCTTTAAAAGGCCTCTGGTTA-------AATGAGTTCTATACATTAAATTTATAACC :461
245 250 255 260 265 270 275 280 285 290 295 300 305 310 315 320

28 Genética poblacional y Filogeografía de las tortugas marinas golfina y laúd en el Pacifico mexicano
Tabla 6.2 Resumen de las características genéticas de las colonias reproductoras de L olivacea bajo estudio
El Verde, SIN
La Gloria JAL.
P. Tlalcoy. GRO
Escobilla, OAX
Pto Arista, CHIS
Ostional, COSTA RICA
Arnhem L., AUSTRALIA
Promedios (global)
No de hembras por colonia.
300-500 300-500 300-700 250.000 100-500 50-100,00 100-500 Tamaño Muestra
15 12 18 21 20 15 8 16.17
Núm Haplotipos
4 6 4 7 4 4 2 4.17 (13)
Núm. de loci analizados
487 487 487 487 487 487 487 487
Núm. de sitios polimórficos 5 11 6 8 7 3 3 5.33
(14)
Diversidad Haplotipica
0.62 ± 0.1196
0.68 ± 0.148
0.59± 0.1086
0.61±- 0.1164
0.49± 0.1167
0.73± 0.0669
0.54± 0.1232
0.60± 0.1086 (0.67)
Núm. de transiciones 4
5 4 6 3 2 2 8
Núm. de Transversiones
0 1 1 1 2 0 0 3
Prop. trans:transv n.d. 5 4 6 1.5 n.d. n.d. 8 3 = 2.67
Núm. de indels 1 5 1 1 2 1 1 1.2 (3)
Composición nucleotidica- %C %T %A %G
17.4 35.1 33.3 14.0
17.5 35.1 33.4 14.0
17.5 35.1 33.3 13.9
17.4 35.1 33.3 14.0
17.4 3 51 33 3 14.0
17.4 35.2 33.3 14.0
17.5 34.8 33.5 13.9
17.4 35.1 33.3 14.0
Diversidad nucleotídica
0.0023 0.0046 0.0030 0.0022 0.0020 00.0025 0 0033 0.0026 (0.0025)
grupos: i) las colonias de México con una frecuencia ≥ de 0.60 y ii) la colonia de Costa Rica con una frecuencia de 0.40. Por otro lado, el haplotipo "N". que también se encontró presente en todas las colonias del Pacífico oriental, marca dos grupos. Solo que en el haplotipo "N", la mayor frecuencia (0.33) se encontró en la colonia deCosta Rica y las colonias de México presentaron frecuencias < 0.20. (ver fig. 6.1).
Se destaca también, que las frecuencias de los haplotipos K y N en la población de Costa Rica se mantienen en proporciones similares. Analizando solamente estos dos haplotipos y tomando en cuenta los límitesde confianza (Fig.6.2) la diferencia entre las colonias mexicanas y la de Costa Rica es significativa.
Los haplotipos I y C, se catalogaron como privados o endémicos para la colonia de Oaxaca. De manera
tabla 6.1 (cont. ) 485 490 495 TGGCATACGGTAGtttt LO-A : TGGCATACGGTAGTTTT : 469 LO-D : TGGCATACGGTAGTTTT : 468 LO-B : TGGCATACGGTAGTTTT : 469 LO-C : TGGCATACGGTAGTTTT : 469 LO-E : TGGCATACGGTAGTTTT : 469 LO-F : TGGCATACGGTAGTTTT : 469 LO-G : TGGCATACGGTAGTTTT : 468 LO-I : TGGCATACGGTAGTTTT : 469 LO-K : TGGCATACGGTAGTTTT : 469 LO-M : TGGCATACGGTAGTTTT : 469 LO-N : TGGCATACGGTAGTTTT : 470 LO-O : TGGCATACGGTAGTTTT : 470 LO-P : TGGCATACGGTAGTTTT : 470 LO-Z : TGGCATACGGTAG---- : 470 LK-LK3B : TGGCATACGGTAGTTTT : 461

Informe Final - F. Alberto Abreu Grobois. Raquel Briseño Dueñas. Ana R. Barragán Rocha 29 análoga, los haplotipos G y P para la colonia de Chiapas. el haplotipo F para la colonia de Guerrero, y el haplotipo Z para la de Jalisco. En la colonia de Sinaloa, no se encontraron haplotipos privados. En total, ocho de los trece haplotipos identificados se encontraron corno privados para algunas colonias individuales. En las muestras de la colonia australiana de L. olivacea, los dos haplotipos identificados (A y D) no se comparten con ninguna colonia del PO y la frecuencia dominante fue la del haplotipo "A".
Diversidad genética La caracterización genética de las colonias analizadas, se enlista en la Tabla 6.2. Las estimaciones
de la diversidad de haplotipos (h) fluctuaron entre 0.49 y 0.73, con los valores más altos en las colonias de Costa Rica, El Verde y Oaxaca. Las estimaciones de la diversidad nucleotídica presentaron un intervalo π=0.0033-0.0022, correspondiendo estos valores a las poblaciones de Australia y Oaxaca respectivamente.
La diversidad nucleotídica, π, es la diversidad genética a nivel de nucleótidos, tomando en cuenta el grado de divergencia estructural entre ellos. Dependiendo de los loci y especies estudiadas la diversidad nucleotídica en una población típica se encuentra en el intervalo de 0.0005-0.020 (Stephan y Langley, 1992 citado en Avise, 1994). El valor global, tomando en cuenta solo las 13 secuencias fue de 0.006 ± 0.003, mientras que el promedio de los valores para cada población fue de 0.003. Los valores de diversidad haplotípica observados en este trabajo, se encuentran dentro del intervalo global estimado por Bowen et al. (1998) para L. olivacea (h=0.0-0.76).
Tabla 6.3 Distribución de la variación genética entre las pobaciones de México y Costa Rica Fuente de la variación g. l.
Suma de Cuadrados
Componentes de La varianza
Porcentaje de la variación
Entre Grupos
1 0.842 0.01612 2.82
Entre Poblaciones Dentro de Grupos
4 1.716 -0.00062 0.77
Dentro de Poblaciones
95 41.753 0.43951 96.41
Total 100 44.310 0.45500 ΦST = 0.036
Frecuencias haplotípicas y sus intervalos de confianza El procedimiento de re-muestreo iterativo o "bootstrapping" deriva las inferencias sobre la
distribución de las frecuencias de los haplotipos en la población, basándose en las frecuencias absolutas de los haplotipos encontrados en la muestra. Los resultados muestran la evaluación de los errores std de las frecuencias, representados en la Fig. 6.1. Solo se incluyó la distribución de las frecuencias de las poblaciones de México y de Costa Rica. No se consideraron los datos de Australia ya que simplemente no comparten haplotipos con las poblaciones del margen oriental del Pacífico.
Tabla 6.4 Distribución de la variación genética entre las pobaciones del Pacífico oriental (agrupando poblaciones de México y Costa Rica en un solo grupo)
Fuente de La
i i
g.l. Suma de Cuadrados
Componentes de la varianza
porcentaje de la variación
Entre Poblaciones 5 3.584 0.00419 0.64
Dentro de Poblaciones
95 61.445 0.64679 99.36
Total 100 65.030 0.65099 ΦST = 0.006

30 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacífico mexicano
Estructura de la población El análisis de varianza molecular puede ser aplicado a varias escalas geográficas de la distribución
espacial de las poblaciones para identificar las "fronteras" de unidades que demuestran homogeneidad genética. En el presente caso se aplicó a las poblaciones muestreadas dentro de toda la cuenca del Pacífico pero particularmente en la porción del Pacífico oriental, abarcando las poblaciones de golfina en México y Costa Rica.
En primer lugar se analizó la distribución de la variación al agrupar las poblaciones de México por separado de la de Costa Rica. El resultado del AMOVA (Tabla 6.3) demuestra que la mayoría de la variación en la región control del DNAmit, se localiza dentro de las poblaciones (>96°/o; o sea ΦST= 0.04) y no entre ellas (entre poblaaciones dentro de grupos) o aún entre las poblaciones de México y la de Costa Rica (entre grupos). Cuando se consideraron todas las poblaciones del Pacífico oriental como un solo grupo, la estimación, del componente de la varianza permanece dentro de las poblaciones (> 98%) y solamente el 0.64% de la variación ocurre entre las poblaciones (Tabla 6.4). Eso es, el valor de ΦST
., = 0.006 lo cual evidencia una escasa diferenciación entre colonias de esta especie en la región del Pacífico oriental. Pruebas de diferenciación interpoblacional
Distancias genéticas La prueba de las diferencias de los valores de las Fst’S por pares de poblaciones, basada en
distancias entre haplotipos empleando el método de Kimura de 2 parámetors derivó los datos de la Tabla 6.5. En este análisis se demostró que solamente existen diferencias significativas en las comparaciones de las poblaciones de Australia contra las poblaciones del Pacífico oriental.
Esta estimación de distancias genéticas entre las colonias de L. olivacea en la región del Pacífico oriental,
Tabla 6.5 Distancias genéticas derivadas de las FST’S para comparaciones entre pares de poblaciones y sus valores de P
colonia Sinaloa Guerrero Oaxaca Chiapas COSTA RICA
Guerrero -0.005
Oaxaca -0.037 0.019
Chiapas -0.009 0.069 -0.032
COSTA RICA -0.026 0.066 0.027 0.063
AUSTRALIA 0.713 0.636 0.725 0.771 0.714
P < < . 0 1
< .20
< .50
n.s.
confirmó la carencia de una diferenciación genética en esta escala geográfica. Por otro lado, al comparar las distancias genéticas entre la colonia de Australia y las colonias del margen oriental, se encontró consistentemente un valor alrededor de 0.70. Este estimado, permite una aproximación de la separación geográfica necesaria para que se establezcan diferencias genéticas significativas en este locus para esta especie.
Estimaciones de las tasas de migración En la comparación entre todas las poblaciones del Pacifico oriental realizado para estimar la tasa
de migración (m) basándose en el estadístico FST (Tabla 6.5), los niveles de flujo génico fueron muy variables, presentando valores desde 6.8 a >50. Dentro de este intervalo, Guerrero presentó una tasa de migración mayor con Sinaloa, que con Oaxaca, Chiapas y Costa Rica.
Las tasas de migración estimadas entre la colonia de Costa Rica y las colonias de Guerrero. Oaxaca y Chiapas fueron muy similares (» 7) y relativamente reducidas, si se comparan con la alta tasa de migración estimada para con la colonia de Sinaloa.
Es necesario señalar que se debe tener cautela en la interpretación de las estimaciones de la tasas de

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas. Ana R. Barragán Rocha 31 migración a partir de los valores de FST, ya que este calculo por sí solo no permite distinguir entre las consecuencias de eventos de colonización reciente y el resultado de intercambio genético contemporáneo entre la colonias.
Según lo informado en otros trabajos (Bass et al., 1996; Bowen et al., 1998), tasas de migración mayores de 1 a 4, son consideradas como evidencia de que el flujo de genes es suficiente para mantener un pool génico
Tabla 6.6 Tasa de migración entre colonias de L. olivacea (Nm) derivadas del estadístico Fst
colonia Sinaloa Jalisco Guerrero Oaxaca Chiapas COSTARICA
Jalisco infinito
Guerrero infinito infinito
Oaxaca infinito infinito 25.20
Chiapas infinito 21.24 6.79 infinito
COSTA RICA infinito 11.44 7.10 18.00 7.41
AUSTRALIA 0.20 027 0.29 0.19 0.15 0.20
relativamente homogéneo (Slatkin, 1987). Los valores estimados para este trabajo, además de mostrar un alto grado de intercambio genético, también evidencian que éste no es totalmente homogéneo entre todas las colonias del Pacífico oriental.
Los valores entre la colonia de Australia y las del Pacífico oriental (< < 0 . 5 ) son usados como punto de comparación. Estos valores de Nm, confirman el grado de aislamiento genético entre las colonias del Pacífico oriental y el Pacífico occidental.
Nm << 1
1-10
10-30
> 50

32 Genética poblacional y Filogeografía de las tortugas marinas golfina y laúd en el Pacifico mexicano
Variación en loci de microsatélites del DNA nuclear en tortuga golfina (por F. Alberto Abreu Grobois y Felipe López Chávez)
(nota: cualquier mención a resultados en esta sección deberá hacer referencia a F. López Chávez (2000) Determinación de la estructura genética de las poblaciones de tortuga golfina en el Pacífico central mexicano con microsatélites del DNA nuclear. Tesis de Maestría. Universidad de Colima)
Frecuencias alélicas, genotipos y niveles de variación Se seleccionaron tres de los loci de microsatélites identificados por FitzSimmons et al. (1995) en
tortugas marinas que demostraron ser polimórficos en tortuga golfina: Cm72, Cc 141 y Cm84. El locus Cm84 resultó ser el más polimórfico, presentando un total de 13 alelos, tres alelos fueron encontrados para el locus Cm72 y dos para Cc141. En la Tablas 6.6- 6.8 se enlistan los alelos de cada uno de los loci, identificados por su tamaño medido en pares de bases (bp). Con estos loci se analizaron un total de 213 organismos provenientes de siete de las colonias anidadoras de tortuga marina golfina, localizadas a lo largo de la distribución de la especie en el Pacífico mexicano y centroamericano. Cinco de estas poblaciones provienen del litoral mexicano, una de Costa Rica y la otra de Nicaragua. Se utilizaron las mismas poblaciones a las que se les analizó la región control del DNA mit para realizar una comparación de resultados y empleando diferentes loci. Sin embargo se añadió la población de Platanitos (Nayarit) en México y la de La Flor en Nicaragua para extender la cobertura del análisis, siguiendo la lógica planteada para la estrategia de muestreo (ver Materiales y Métodos).
Los niveles de variación observados varían entre estos loci, abarcando valores de heterocigosidad esperada promediada para las poblaciones (He prom) entre 0.14 el menos polimórfico (Cm72) hasta 0.82 el más variable (Cm84).
Partiendo de la frecuencia de alelos observada, se realizaron corridas de "remuestreo" ("bootstrapping")para determinar los errores std. de la frecuencia alélica reportada. Los resultados pueden observarse en las Figuras 6.6b, 6.8b y 6.10b para Cm l 41, Cm72 y Cm84, respectivamente, graficadas contra la distribución de las frecuencias alélicas de cada población. De ésto resulta evidente que los niveles del error std. son pequeños en los dos primeros loci, mientras que para Cm84 los errores std son grandes. Esto es de esperarse debido a que, al aumentar el número de alelos en el locus, la cantidad de representantes de cada alelo en cada muestra llega a ser muy pequeño y disminuye la precisión de la medición.
Aunque los niveles de variación para un mismo locus varía entre especies de tortugas marinas analizadas, los valores obtenidos en este estudio se encuentran dentro de la gama observada por FitzSimmons et al.(1995)
Los genotipos observados para los tres loci pueden verificarse en las Tablas 6.7, 6.9 y 6.1 1, junto con las frecuencias en las que se encontraron. Debido a la baja frecuencia de algunos de los alelos, no se observaron algunos de los posibles genotipos. Por ejemplo, para Cc141 no se encontraron homocigotos del alelo menos común (178/178), tampoco para el homocigoto 238/238 en Cm72. El caso para Cm84 es todavía más extremo debido al gran número de alelos y en la tabla 6.11 solo se muestran los genotipos encontrados con sus correspondientes frecuencias.
Los valores observados y esperados de heterocigosidad para el conjunto de los tres loci (Ho y He, resp.) es muy semejante (Tabla 6.12). No obstante, en unos de los loci para algunas poblaciones se encontraron desviaciones de los valores esperados de acuerdo a la ley de Hardy-Weinberg. Esto particularmente en Cm84 y para las muestras de Platanitos, Nay. y La Gloria, Jal. En estas dos poblaciones este resultado anomalo probablemente fué causado por un muestreo realizado en dos años distintos y con variación en las frecuencias alélicas. Esto se puede inferir ya que al separar los muestreos por año (Tabla 6.14) los valores de P no son significativos para esta prueba. Este resultado es objeto de investigaciones.
Estructura poblacional evaluada por análisis de microsatélites La prueba AMOVA aplicada a la variación genética observada en los loci de microsatélites permite
determinar la proporción de esta variación que se encuentra distribuida entre poblaciones y dentro depoblaciones. Los resultados (Tabla 6.15), indican, una vez más, que la mayoría de la variación se encuentra dentro de las poblaciones, arrojando un valor de ΦST=0.12. Este resultado corrobora y complementa aquel obtenido con esta misma prueba a partir de los datos con la región control y refleja la existencia de un intercambio genético entre las poblaciones, por lo menos en escalas de tiempo genético (varias generaciones).

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 33
Tabla 6.6a Frecuencias alélicas para el locus Cc141en colonias de la tortuga golfina Colonia
Alelo
El Verde Sin. N=9
La Gloria Jal.
N=29
TlacoyunqueGro. N=18
Escobilla Oax. N=10
Pto Arista Chis. N=17
La Flor NIC N=18
Nancite C.R. N=16
178 0.11 0.09 0.13 0.05 0.12 0.06 0.03 188 0.89 0.91 0.88 0.95 0.88 0.94 0.97
Tabla 6.6b Distribución de las frecuencias alélicas en el locus Cc141 y sus errores std. determinados por «bootstrapping» de los valores observados.
Colonia Genotipo
Escobilla Oax
El Verde Sin
La Gloria Jal
TlacoyunqueGro
Pto Arista Chis
La Flor Nic
Nancite C.R.
178/178 0.00 0.00 0.00 0.06 0.00 0.00 0.00 178/188 0.10 0.20 0.17 0.13 0.24 0.1 1 0.06 188/188 0.90 0.80 0.83 0.81 0.76 0.89 0.94
ho 0.10 0.20 0.17 0.13 0.24 0.11 0.06 he 0.10 0.20 0.10 0.22 0.21 0.10 0.06
Ho prom 0.14 He prom 0.14

Tabla 6.7 Frecuencias genotípicas y heterocigosidad para el locus Cc141, ho= heterocigosidad observada, he
= heterocigosidad esperada. Hoprom y Heprom heterocigosidad observada y esperada promediada para todas las poblaciones a partir de ho y he� resp.
34 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacifico
mexicano Tabla 6.8a Frecuencias alélicas para el locus Cm72 en colonias pie la tortuga golfina
Colonia Alelo
El verde Sin. N=20
Platanitos Nay. N=32
La Gloria Jal. N=35
Tlacoyunque Gro N=15
Escobilla Oax. N=20
Pto Arista Chis. N=16
La Flor NIC N=23
Nancite C.R. N=17
234 0.73 0.76 0.77 0.73 0.62 0.72 0.75 0.65
236 0.18 0.21 0.21 0.27 0.25 0.22 0.22 0.32
238 0.10 0.03 0.01 0.00 0.10 0.06 0.03 0.03
Tabla 6.8b Distribución de las frecuencias alélicas en el locus Cm72 y sus errores std. determinados por «bootstrapping» de los valores observados.
Cólonia Genotipo
El Verde Sin
Platanitos Nay
La Gloria Jal
Tlacoyunque Gro
Escobilla Oax
Pto Arista Chis
La Flor NIC
Nancite C.R.
234/234 0.55 0.58 0.60 0.47 0.30 0.50 0.50 0.41 236/236 0.05 0.03 0.06 0.00 0.00 0.06 0.00 0.12 238/238 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 234/236 0.20 0.35 0.31 0.53 0.50 0.31 0.44 0.41 2341238 0.15 0.00 0.03 0.00 0.20 0.06 0.06 0.06 236/238 0.05 0.03 0.00 0.00 0.00 0.06 0.00 0.00
ho 0.40 0.39 0.34 0.53 0.70 0.44 0.50 0.47
he 0.42 0.38 0.36 0.39 0.51 0.43 0.39 0.48 Ho prom 0.47 He prom 0.42

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 35 Tabla 6.10a Frecuencias alélicas para el locus Cm84 en colonias de la tortuga golfina
Colonia Alelo
El Verde Sin.
N=20
Platanitos Nay
N=34
La Gloria Jal.
N=25
Tlacoyunque Gro. N=18
Escobilla Oax. N=20
Pto Arista Chis. N=18
La Flor NIC N=6
Nancite C.R. N=18
320 0.00 0.02 0.01 0.000 0.00 0.00 0.00 0.00
322 0.03 0.12 0.16 0.03 0.10 0.00 0.17 0.25
324 0.13 0.26 0.33 0.03 0.00 0.03 0.08 0.22
326 0.23 0.12 0.15 0.28 0.28 0.42 0.00 0.17
328 0.18 0.17 0.08 0.22 0.18 0.19 0.25 0.11
330 0.03 0.18 0.17 0.08 0.03 0.22 0.08 0.08 332 0.05 0.06 0.09 0.03 0.10 0.03 0.08 0.03 334 0.03 0.08 0.01 0.28 0.10 0.06 0.08 0.06 336 0.15 0.00 0.00 0.0b 0.00 0.03 0.17 0.08 338 0.08 0.00 0.00 0.00 0.05 0.03 0.08 0.00 340 0.03 0.00 0.00 0.00 0.05 0.00 0.00 0.00 342 0.10 0.00 0.00 0.00 0.05 0.00 0.00 0.00 344 0.00 0.00 0.00 0.00 0.05 0.00 0.00 0.00 346 0.00 0.00 0.00 0.00 0.03 0.00 0.00 0.00
Tabla 6.10b Distribución de las frecuencias alélicas en el locus Cm84 y sus errores std. determinados por <bootstrapping» de los valores observados.

36 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacífico mexicano
Tabla 6.11 Frecuencias genotipicas y heterocigosidad para el locus Cm84, ho = heterocigosidad observada, he.= heterocigosidad
esperada. H0 prom y He prom= heterocigosidad observada y esperada promediada para todas las poblaciones a partir de ho y he’ resp Nota: Para mayor claridad, se eliminaron los genotipos no observados y los valores de 0 fueron substituidos por « - »
Colonia El Verde Sin
Platanitos Nay
La Gloria Jal
Tlalcoyunque Gro
Escobilla Oax
Pto Arista Chis
La Flor Nic
Nancite C.R
Genotipo 320/330 --- 0.03 --- --- --- --- -- --- 320/334 --- --- 0.02 --- --- --- --- --- 322/322 --- 0.03 0.02 --- --- --- --- --- 322/324 --- 0.06 0.02 --- --- --- --- --- 322/326 --- 0.03 0.02 --- 0.15 --- --- 0.17 322/328 --- --- 0.02 --- --- --- --- --- 322/330 --- 0.06 0.16 --- --- --- --- 0.06 322/332 --- --- 0.05 --- 0.05 --- --- --- 322/334 --- 0.03 --- --- --- --- --- 0.06 322/336 0.05 --- --- --- --- --- 0.33 0.11 322/338 --- --- --- --- --- --- --- 0.11 324/324 0.05 0.09 0.20 --- --- --- --- 0.11 324/326 --- 0.03 0.07 --- --- 0.06 --- 0.06 324/328 0.10 --- 0.07 --- --- --- 0.17 --- 324/330 --- 0.12 0.05 --- --- --- --- --- 324/332 --- 0.06 0.05 --- --- --- --- --- 324/334 --- 0.06 --- 0.05 --- --- --- 0.06 324/336 --- --- --- --- --- --- --- 0.11 324/340 0.05 --- --- --- --- --- --- --- 326/326 -- 0.06 --- 0.05 0.15 0.22 --- 0.06 326/328 0.05 0.03 0.07 0.10 --- --- --- --- 326/330 -- 0.03 0.09 --- --- 0.17 --- 0.11 326/332 --- --- 0.05 --- 0.10 --- --- --- 326/334 --- --- --- 0.25 --- 0.06 --- --- 326/336 0.15 --- --- 0.05 --- 0.06 --- --- 326/338 0.15 --- --- --- --- 0.06 --- --- 326/342 0.10 --- --- --- --- --- --- --- 328/328 0.05 0.09 --- 0.05 0.05 0.17 --- --- 328/330 0.05 0.06 --- --- --- 0.06 0.17 --- 328/332 0.05 0.06 --- 0.10 0.05 --- --- --- 328/334 --- --- --- 0.10 0.05 --- 0.33 --- 328/338 --- --- --- --- 0.05 --- --- --- 328/340 --- --- --- --- 0.10 --- --- --- 330/330 --- 0.03 0.02 0.05 --- 0.06 --- --- 330/332 --- --- --- --- --- 0.06 --- --- 330/334 --- --- --- 0.05 --- 0.06 --- --- 330/342 --- --- --- --- 0.05 --- --- --- 332/332 --- --- 0.02 --- --- --- --- --- 332/342 0.05 --- --- --- --- --- --- --- 334/334 --- 0.03 --- --- --- --- --- --- 334/336 0.05 --- --- 0.05 --- --- --- --- 334/338 --- --- --- --- 0.05 --- --- --- 334/342 --- --- --- --- 0.05 --- --- --- 334/346 --- --- --- --- 0.05 --- --- --- 336/342 0.05 --- --- --- --- --- --- --- 344/344 --- --- --- --- 0.05 --- --- --- Total l.00 1.00 1.00 1.00 1.00 1.00 1.00 l.00 ho 0.90 0.67 0.68 0.75 0.75 0.56 1.00 0.83 he 0.86 0.83 0.80 0.78 0.85 0.73 0.85 0.83 Ho prom 0.77 He prom 0.82

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 37
Tabla 6.12 Heterocigosidad observada y esperada para cada población promediada para los tres loci (Ho y He).
El Verde Sin
Platanitos Nay
La GloriaJal
Tlalcoyunque Gro
Escobilla Oax
Pto Arista Chis
La Flor Nic
Nancite C.R.
Ho 0.50 0.41 0.38 0.46 0.56 0.35 0.83 0.47 He 0.49 0.44 0.46 0.42 0.52 0.41 0.74 0.47
Tabla 6.13 Probabilidad estimada de que las colonias se encuentren en equilibrio Hardy-Weinberg
Colonia
Locus El Verde
Sin
Platanitos Nay
(dos años)
La GloriaJal
(dos años) Tlalcoyunque
Gro. Escobilla
Oax. Pto Arista
Chis. La Flor
Nic Nancite
C.R. Cm72 0.48 0.05 0.72 0.51 0.01 0.33 0.32 1.00 Cm84 0.14 0.05 0.00 0.46 0.03 0.11 0.00 0.16Cc 141 l.00 0.06 1.00 0.29 1.00 1.00 1.00 1.00
Tabla 6.14 Valores de P de que las colonias estén en equilibrio H-W para La Gloria y Platanitos, separando los resultados por año.
Colonia
Locus Platanitos dos años
Platanitos I996
Platanitos 1997
La Gloria dos años
La Gloria I994
La Gloria 1996
Cm72 0.05 0.51 0.06 0.72 1.00 0.66 Cm84 0.05 0.43 0.13 0.003 0.09 0.07Cc141 0.06 0.41 0.14 1.00 1.00 1.00
Tabla 6.15 AMOVA para los datos de microsatelites en tortuga golfina
Fuente de variación g.l. Suma de
cuadradosComponentesde variación
Porcentaje de variación
Entre poblaciones 7 22.816 0.10719 Va 12.21
Dentro de poblaciones 180 138.663 0.77035 Vb 87.79
Total 187 161.479 0.87754
ΦST=0.12

38 Genética poblacional y filogeografia de las tortugas marinas golfina y laúd en el Pacífico mexicano
Información complementaria de Lepidochelys olivacea en el Pacífico oriental (por Raquel Briseño Dueñas y F. Alberto Abreu Grobois)
(nota: cualquier mención a información en esta sección deberá hacer referencia a R. Briseño Dueñas (1998) Variación genética en la región control del ADN mitocondrial de poblaciones de la tortuga golfina, Lepidochelys olivacea, en el Pacífico oriental y las implicaciones para su conservación. Tesis de Maestría. Universidad Autónoma de Sinaloa)
Flujo génico entre colonias Con el propósito de validar o contrastar los resultados obtenidos a partir del análilsis genético, se analizó
una fuente de información que también permitiera evaluar los niveles de flujo génico entre colonias, pero de manera independiente. Se analizaron los datos de marca-recaptura aplicado a hembras reproductoras de L. olivacea provenientes de los programas de marcado en México y Costa y Rica. El dato de relevancia en este contexto sería sobre la existencia y frecuencia de hembras que anidan en más de una playa (Tabla 6.6) en contraposición con un comportamiento de estricta fidelidad a una sola playa. Si fuera el segundo caso, se esperaría (como ocurre en otras especies de tortugas marinas) ausencia de evidencias de traslocación de sus sitios de anidación a hembras de la especie junto con una diferenciación genética significativa entre colonias (ver Bowen, 1996)
De este conjunto de datos, se observa, por ejemplo, que tortugas que anidaron en Oaxaca, tienen la capacidad de desplazarse subsecuentemente para anidar a otras playas en regiones tan distantes como algunas ubicadas tanto al norte, en México (Jalisco) como al sur y fuera del país (Colombia). Por otro lado, tortugas marcadas durante su anidación en la playa "El Verde", Sinaloa se han recapturado anidando en playas de una región que abarca prácticamente la extensión total del litoral mexicano (ver por ejemplo Ríos-Olmeda, 1997).
Esta información corrobora y extiende las observaciones de los resultados genéticos e implican la ocurrencia de cantidades suficientes de emigrantes por generación entre colonias del Pacífico oriental como para afectar la distribución de la variación genética en escalas de tiempo evolutivo.
En la tabla 6.16, se observa igualmente que las tortugas golfinas que anidan en las playas de Nancite y Ostional (Costa Rica) tienen una alta frecuencia en la alternancia de uso de estas dos playas para la anidación. Independencia demográfica entre colonias
No obstante que los resultados de genética y de los programas de marcaje sugiere un flujo genético
Tabla 6.16 Datos de hembras de Lepidochelys olivacea marcadas durante la anidación en una playa y recapturadas anidando en playas diferentes.
frac sitios de
anidación alternativos
Países que comparten
anidaciones
dist. aprox.(km)
ref.
1 Sinaloa – Nayarit México 300 Ríos com. pers
1 Sinaloa – Oaxaca México 1,487 Ríos com. pers.
1 Sinaloa – Cbiapas México 2,050 Ríos com. pers
1 Colima – Guerrero México 280 Márquez, com. pers.
1 Oaxaca – Jalisco México 1,171 Márquez, com. pers
1 Oaxaca – Michoacán México 700 Márquez, com. pers.
2
Oaxaca – Guerrero
México 230 Márquez, com. pers.
1 Oaxaca – Guatemala
México – Guatemala 7 Márquez, com. pers.
2 Oaxaca – El Salvador
México - El Salvador 900 Márquez, com. pers.
1 Oaxaca – Colombia
México – Colombia 2,600 Márquez, com. pers.
50 Nancite- Ostional Rica Costa Rica 50 Cháves com. pers.
10 Nancite Chacocente Costa Rica – Nicaragua 150 Cháves com. pers.

informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 39 importante en escalas de tiempo de significancia evolutiva, existen otras evidencias que sugieren que, enescalas de tiempo de relevancia para el manejo, la demografía de las colonias de esta especie se comportande manera independiente! Una visión de este enfoque puede observarse en la Figura 6.11 en donde secompendia la información histórica de las tendencias poblacionales. en tres colonias reproductoras concomportamiento de arribazones en México en los últimos 40 años: El Playón de Mismaloya, Jal.(de donde seobtuvo la muestra de La Gloria para este trabajo): Piedra de Tlalcoyunque, Gro. y Escobilla, Oax. Lasfrancas diferencias en el comportamiento del número de nidadas registradas, entre la Escobilla y las otrasdos playas, se encuentran directamente relacionadas con eventos antropogénicos (captura comercialintensiva) que provocaron un abatimiento de estas últimas observables en años posteriores. Esto es, lascolonias de Jalisco y Guerrero estuvieron sometidas durante la década de los 60's a una captura comercial sinprecedentes, rebasando una extracción 1.5 y 2 millones de tortugas en México, principalmente golfinas, deacuerdo a datos extraoficiales, en las dos décadas entre 1965 y 1975
Figura 6.11 Anidaciones de L olivacea en tres playas de arribada y registros oficiales de captura comercial en el periodo 1961-1994 (Márquez et al., 1996b; Silva-Bátiz, 1994; Márquez et al., 1976; Nava, 1994)
No existen registros oficiales de los niveles de extracción real posterior a ese período, porque lacaptura legal fue limitada a franquicias especiales y en 1990, se declaró la veda total.
La información que es particularmente relevante para analizar posibles interconexiones oindependencia demográfica entre las colonias es la dinámica de crecimiento poblacional en las mismasdurante los últimos 20 años. Mientras que las colonias de Jalisco y Guerrero que fueron seriamentedisminuidas por la pesca no muestran síntomas de una recuperación significativa, y pesar de los esfuerzos deconservación la colonia de Escobilla ha mostrado una recuperación sostenida a partir de 1990.
El mismo comportamiento hacia incrementos poblacionales se ha observado también en otras playasdel litoral mexicano. Inclusive, en los inventarios de las anidaciones de golfina en el Pacifico mexicano, setienen registros colonización de sitios en los que no se habían registrado anidaciones en temporadasprevias.

40 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacífico mexicano
Discusión
Diversidad genética Mientras que estudios anteriores sobre la diversidad genética de las tortugas marinas a partir del
análisis del DNA mitocondrial habían descrito a la tortuga golfina como un taxón de baja diversidad y una filogenia del DNA mitocondrial "poco profunda" (Bowen y Karl, 1996) o, en la investigación más reciente con un nivel de diversidad haplotípica entre "moderado" y "bajo" (Bowen et al., 1998), los resultados aquí obtenidos para la diversidad haplotípica (h) indican que dentro de una escala regional (Pacífico oriental), los valores son muy similares y caen dentro de la gama de los observados para otras especies (Tabla 6.17)
Por otro lado, los valores de diversidad nucleotídica (π) contrastan marcadamente con los valores en otros grupos de tortugas marinas, siendo menores y más homogéneos. Esta homogeneidad podría ser ocasionado por recientes colonizaciones regionales de acuerdo a los comentarios de Bowen et al. (1998) y sería consistente con la pequeña diferenciación estructural entre los haplotipos. Bajo esta explicación, el poco tiempo transcurrido no habría permitido alcanzar niveles de diferenciación que son encontrados más comúnmente en las otras espcecies.
Los resultados del análisis en el DNA nuclear por medio de microsatélites tampoco encontró evidencias de disminución en los niveles de variabilidad genética. Los valores de heterocigosidad promedio y del números de alelos se encuentran dentro de la gama identificada para otras especies de tortugas marinas (Tabla 6.18) que investigaron con los mismos loci.
Los análisis de la composición genética podrían también abordar la pregunta sobre la posible erosión de la variabilidad genética, posible producto de los cuellos de botella a la que han sido sujetas las poblaciones en las últimas décadas y que resultaría de una subsecuente endogamia. Desgraciadamente, no se cuenta con un valor umbral universal que permitiría objetivamente determinar si un cierto valor de diversidad reflejan una endogamia o no, y menos para tortugas marinas. Como alternativa viable, podemos comparar entre especies usando el mismo segmento del DNA, con la ventaja de poder comparar y contrastar dentro de un mismo grupo con ancestrías comunes. Bajo este contexto, los resultados de diversidad genética para las colonias de tortuga golfina en el Pacífico oriental exhibe niveles que caen dentro de la gama observada en otras especies de tortugas marinas y no reflejan disminuciones observables.
Tabla 6.17 Niveles de diversidad genética en la región control del DNA mitocondrial de tortugas marinas
Esp
ecie
Áre
a G
eogr
áfic
a
Col
onia
s
Hap
lotip
os Diversidad
Haplotipica (h)
rango (prom)
Diversidad Nucleotidica
(π) rango
(global) Ref
eren
cia
E. imbricata Caribe 7 17 0.12 - 0.78 (0.85)
0 - 0.006 (0.0119)
Basa et al. (1996)
C. mydas Atlántico y Medlterrá-
neo 9 18 0 - 0.82
(0.83) 0 - 0.006 (0.0050)
Encalada et al. (1996)
D coriacea Global 8 0.54 0.002 (0.0051)
Dutton (1996)
L. kempi Golfo de México 1 4 0.69 0.003 Bowen
et al. (1996)
L. olivacea Global 7 12 0-0.76 (0.81)
0-0.21 (0.0077)
Bowen et al.(1998)
L. olivacea Pacifico Oriental 6 13 0.49 - 0.73
(0.67)
0.0020- 0.0033
(0.0025) este trabajo

Informe Final - F. Alberto Abreu Grobois. Raquel Briseño Dueñas. Ana R. Barragán Rocha 41
Tabla 6.18 Variabilidad en loci de microsatélites del DNA nuclear en varias especies de tortugas marinas.
Especie Cm72
h (núm de alelos)
Cm84 h
(núm de alelos)
Cc 141 h
(núm de alelos) Referencias
Eretmochelys imbricata 0.615 (7) 0.706 (15) - FitzSimmons
et al. (1995) Chelonia mydas 0.900 (25) 0.707 (19) - FitzSimmons
et al. (1995) Natator
depressus 0.286 (4) 0.391 (3) - FitzSimmons et al. (1995)
Caretta caretta 0.705 (5) 0.929 (11) - FitzSimmons
et al. (1995) Dermochelys
coriacea 0,000 (1) 0.667 (4) - FitzSimmons et al. (1995)
Lepidochelys olivacea 0.900 (2) 0.444 (6) - FitzSimmons
et al. (1995)
L. kempii 0.521(3) 0.746 (18) 0.660 (6) Kichler (1996)
L. olivacea 0.491(3) 0.912 (16) 0.135 (2) Kichler (1996)
L. olivacea 0.47 (3) 0.77 (13) 0.14 (2) Este trabajo
Estructura y unidades de manejo
El análisis de la estructura de las poblaciones por medio de AMOVA o procedimientos similares ayuda aidentificar la presencia de discontinuidades dentro del conjunto estudiado que reflejaría la presencia de unidadesdemográficamente independientes o unidades de manejo discretas. En varias especies, por ejemplo en la tortugaverde (Encalada et al., 1996) y en carey (Bass et al., 1996) este comportamiento prevalece y así, a partir de resultados con secuencias de la región control del DNA mitocondrial, se encuentran valores altos del estadísticoque mide el grado de diferenciación interpoblacional (Tabla 6.19). En ambas de estas dos especies, su comportamiento típico incluye una marcada tendencia hacia la filopatría y, por consiguiente se obtiene una plenaidentificación de unidades de manejo. En el presente estudio con la región control del DNAmt en las tortugas golfinas, se obtuvieron valores muy bajos de Φst (1.8 - 3.6%), junto con altos valores de Nm (flujo génico). Estonos conduce a determinar que las líneas maternas de las tortugas golfinas del Pacífico oriental son escencialmentepanmícticas, por lo menos dentro de esta escala geográfica y dentro de una escala de tiempo que abarca varias generaciones. Los resultados de microsatélites también arrojan una conclusión similar ya que, aunque los valoresde Φst son más elevados (12%) , son marcadamente menores de el rango de 20-60% encontrado para especies
Tabla 6.19 Estimadores de estructura poblacional y tasas de migración en diferentes especies de tortugas marinas usando la región control del DNA mitocondrial
Especie Área Geográfica
Grado de Diferenciación
Interpoblacional (FST. GST o 4'sT)
Nm (flujo
génico) Referencias
E. imbricata Caribe 64% 0 – 2.0 Bass et al. (1996)
C. mydas Atlántico y Mediterráneo 20 – 97% 0.02 - 2.5 Encalada et at.
(1996)
D. coriacea Atlántico 8 - 23% 0.4 - 1.4 Dutton et al. (1995)
L. olivacea Global 59% 0.2 - 4.5 Bowen et al. (1998)
L. olivacea Pacífico 33% 0.14-0.28 Este trabajo
L. olivacea Pacífico Oriental 1.8-3.6% 4 - >50 Este trabajo

42 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacifico mexicano
tortugas marinas donde se ha demostrado la presencia de unidades de manejo bien definidas (carey y verde, Tabla 6.19). Los mayores valores obtenidos con microsatélites refleja la mayor resolución obtenible a partir del análisis de este tipo de loci y que aparentemente puede detectar diferencias más sutiles entre las poblaciones que la que se obtiene con la región control del DNAmt.
Una alternativa a la panmixis, para explicar los resultados, implicaría un evento de colonización reciente (en escala evolutiva, dentro de los últimos 300,000 años según estimaciones de Bowen et al., 1998 con base en los datos moleculares), más que un flujo contemporáneo de genes.
Sin embargo, las evidencias independientes obtenidas de los resultados de marca-recaptura de tortugas golfinas (ver sección Información complementaria) también apoya una conclusión de panmixia en esta región, por lo menos en escalas de tiempo evolutivo.

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 43
Tortuga laúd
Variación genética en loci de microsatélites del DNA nuclear (por Ana Barragán Rocha)
(nota: cualquier mención a resultados en esta sección deberá hacer referencia a A. Barragán Rocha (2000) Genética Poblacional de la tortuga laúd (Dermochelys coriacea) en el Pacifico mexicano. Tesis de Maestría. Universidad Nacional Autónoma de México)
Se analizaron un total de 83 muestras de tortuga laúd de las cuatro principales playas de anidación
del Pacífico, para dos loci, DC99 y Nigra32 (N32). Se obtuvieron las frecuencias alélicas y las frecuencias genotípicas. En la tabla 6.17 se muestran los alelos encontrados para los loci DC99 y Nigra32, clasificados según la clasificació de Dutton (1995) (Nota: en la base de datos enviada a la CONABIO se identifica a los genotipos observados tanto con la asignación de letras, como por medio del tamaño del alelo, expresado en pares de bases). Se encontraron 7 alelos para el locus DC99 y 5 para N32. Es importante resaltar que el alelo denominado "Z", encontrado para nuestras muestras de laúd en el locus DC99, no se había descrito antes y por tanto corresponde a un alelo nuevo.
Las frecuencias genotípicas relativas encontradas para cada loci por colonia anidadora se muestran en la tablas 6.18 y 6.19 para los loci DC99 y N32, respectivamente.
Los loci empleados en el análisis de las muestras presentaron 5 o más alelos cada uno; un grado de variabilidad que se considera adecuada para describir las características genéticas a nivel poblacional.
Los niveles de heterocigosis observados en las colonias anidadoras de tortuga laúd del Pacífico mexicano (mayores a 0.5) corresponden a una población que no manifiesta efectos de endogamia ni de fijación de alelos que pudieran resultar de deriva génica a consecuencia de tamaños pequeños de poblacionales mantenidos durante varias generaciones. Este resultado podría indicar que la reducción de las anidaciones observadas en los últimos años para las principales playas de anidación del Pacífico, en caso de que refleje una reducción real en el número de hembras anidadoras de esta especie en la región, ha ocurrido demasiado rápido para causar efectos en la composición genética, tomando en cuenta que dichos efectos sólo son perceptibles después de varias generaciones. Por consiguiente, la tortuga laúd del Pacífico mexicano no está en riesgo inmediato de presentar síntomas de depresión por endogamia relacionados con bajos niveles de heterocigosis. Sin embargo, al desconocerse el tamaño efectivo de las poblaciones (el cual considera también el número de machos adultos) no es posible predecir en este momento si el alto nivel de heterocigosis observado en este estudio se puede mantener en el futuro.
Tabla 6.17 Frecuencias alélicas para dos loci en muestras de tortuga laúd de las cuatro principales colonias anidadoras del Pacífico mexicano
DC99 Nigra32
Alelo Mexiquillo,
Mich.
(N=23)
Tierra Col., Gro
(N=NI7)
Chacahua,
Oax
(N=22)
Barra dela Cruz,
Oax
(N=22)
Mexiquillo,Mich
(N=20)
Tierra col., Gro
(N=11)
Chacahua,
Oax
(N=23)
Barra de la Cruz,
Oax
(N=22)A 0.00 0.03 0.07 0.09 0.10 0.09 0.11 0.11
B 0.30 0.21 0.34 0.30 0.68 0.50 0.59 0.48
C 0.16 0.09 0.11 0.14 0,08 0.21 0.20 0.23
D 0.14 0.15 0.11 0.11 0 0.00 0.00 0.00
E 0.30 0.32 0.27 0.27 0.13 0.21 0.11 0.16
F 0.09 0.12 0.07 0.05 0.03 0.00 0.00 0.02
Z 0.02 0.09 0.02 0.05 - - - -

44 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacifico mexicano
Frecuencia alélica del locus DC99 en poblaciones de tortuga laúd
Frecuencia alélica del locus N32 en poblaciones de tortuga laúd

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 45
Tabla 6.18 Frecuencias genotípicas observadas en muestras de tortuga laúd de las cuatro principales colonias anidadoras del Pacífico mexicano para el locus DC99.
Tabla 6.19 Frecuencias genotípicas observadas en muestras de tortuga laúd de las cuatro principales colonias anidadoras del Pacífico mexicano para el locus Nigra32.
Alelo Mexiquillo,
Mich.
(N=22)
Tierra Col., Gro
(N=17)
Chacahua,
Oax
(N=21)
Barra de la Cruz,
Oax
(N=22)
AB - - 0.10 0.14
AC - 0.06 0.05 -
AE - - - 0.05
BB 0.18 - 0.10 0.14
BC 0.09 - 0.10 0.05
BD 0.05 0.06 0.14 0.09
BE 0.05 0.18 0.10 0.05
BF 0.05 0.18 0.05 -
CD 0.09 - 0.05 -
CE 0.14 0.12 0.05 0.18
CF - - - 0.05
EE 0.14 0.12 0.14 0.09
EF 0.14 - 0.05 0.05
DD 0.05 0.06 - 0.05
DF - 0.06 0.05 -
ZD 0.05 0.06 - 0.05
ZE - 0.12 0.05 0.05
he 0.42 0.80 0.78 0.80
ho 0.64 0.82 0.76 0.73
Alelo Mexiquillo,
Mich.
(N=20)
Tierra Col., Gro
(N=17)
Chacahua,
Oax
(N=22)
Barra de la Cruz,
Oax
(N=22)
AA 0.050 - 0.045 -
A B 0.100 0.176 0.091 0.136
A E - - 0.045 0.091
BB 0.450 0.235 0.318 0.273
BC 0.100 0.235 0.273 0.227
BE 0.200 0.118 0.182 0.045
BF 0.050 - - -
CC - 0.059 0.045 -
CE 0.050 0.059 - 0.182
CF - - - 0.045
EE - 0.118 - -
he 0.52 0.66 0.62 0.57
ho 0.50 0.59 0.59 0.73

46 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacífico mexicano
Tabla 6.20 AMOVA con base en el estadístico Fst para los datos de microsatélites en tortuga laúdrealizado mediante el programa ARLEQUIN 1.1 (Schneider et al., 1997)
Fuente de variación
g.I. Suma de cuadrados
Componentes de variación
Porcentaje de
variación
Entre poblaciones 3 0.923 -0.00839 Va -1.28
Dentro de poblaciones 166 1 10.124 0.66340 Vb 101.28
Total 169 111.047 0.65500
ΦST = 0.13
El análisis de frecuencias alélicas permite identificar la presencia de alelos raros. Conocer la existencia de alelos raros es importante porque generalmente éstos caracterizan las poblaciones que los contienen. Las frecuencias alélicas de todas las colonias anidadoras del Pacífico mexicano consideradas en conjunto son en general similares a las encontradas para Costa Rica por Dutton (1995), a excepción de la presencia de un alelo raro en el locus DC 99 (nombrado Z en este estudio) que no se reporta en la población mencionada. Aunque todavía es prematuro concluir que este alelo es característico de las tortugas del Pacífico mexicano, ya que el tamaño de muestra es muy diferente (84 en México contra 24 en Costa Rica), puede ser un indicio de que las tortugas que anidan en México pueden distinguirse de las que anidan en Costa Rica.
Las frecuencias alélicas consideradas por colonia (labia 6.17) muestran que el alelo Z no se encuentra en las muestras de Mexiquillo, aunque si se observa en el resto de las playas y en muy baja frecuencia.
Discusión Los valores de heterocigosis en las colonias anidadoras mexicanas analizadas fué alto, tanto por
locus (Tablas 6.18 y 6.19) . Es interesante notar que los valores de H para el Pacífico mexicano son muy similares al valor de diversidad haplotípica encontrada para México por Dutton y colaboradores (1999), usando la región
Pop. MEX TC CHA
TC 0.014 ± 0.017 -
CHA 0.001 ± 0.007
0.004 ± 0.003 -
BC
0.018 ±0.024
-0.003 ±0.006
-0.003 ± 0.006
MEX: Mexiquillo; TC: Tierra Colorada; CHA: Chacahua; BC: Barra de la Cruz
control del DNAmt (h = 0.712 ± 0.073). Esto es un indicio de que la alta heterocigosis encontrada en el presenteestudio no es producto del tipo de marcador molecular utilizado (siendo los microsatélites marcadores hipervariables),sino reflejo de una verdadera variabilidad genética elevada en la población. Estos resultados sugieren que, a pesar de la declinación en el número de hembras anidadoras observadas en las playas del Pacífico mexicano enlos últimos años, esta ha ocurrido demasiado rápido para tener un efecto cuantificable sobre la variación genéticaa nivel poblacional.
Los resultados del análisis de varianza molecular (AMOVA), realizado a las colonias anidadoras del Pacífico mexicano como un solo grupo, se muestran en la tabla 6.20. El análisis se realizó con base en el númerode alelos diferentes (Fst; Wright, 195 1). También se calcularon los índices Fst para las cuatro colonias anidadoras mexicanas, en comparaciones por pares de colonias (Tabla 6.21). Podemos observar que los valores para ambos
Tabla 6.21 Valores de Fst para comparaciones por pares de colonias anidadoras de tortuga laúd Dermochelys coriacea de México. Los valores de error estándar fueron obtenidos mediante bootstrapping exhaustivo con el programa MICROSAT (Minch, 1997).

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 47 índices se encuentran muy cercanos a cero, por lo que no existe evidencia de subdivisión poblacional en el conjunto de muestras.
Una de las más importantes aplicaciones de los datos genéticos se encontrará en el análisis de la composición de capturas incidentales en la pesca comercial que opera en aguas internacionales del Océano Pacífico. En una reunión reciente (Bolten et al., 1996), se analizó la condición actual de las poblaciones de tortugas marinas en esa región y el impacto de las pesquerías de palangre sobre ellas. Tomando en cuenta la delicada condición en las poblaciones de tortuga laúd evidenciada por los datos de anidaciones de las colonias más importantes y mejor estudiadas (Chan y Liew, 1996; Sarti et al., 1996) se considera prioritario el contar con medios para identificar los orígenes de organismos capturados incidentalmente con los cuales derivar estimaciones del impacto real sobre las colonias fuente. El presente trabajo representa un avance significativo en este objetivo ya que complementan el estudio de Dutton (1995) con poblaciones de Malasia, Sudáfrica y Costa Rica. Un análisis de la composición alélicas en los loci de microsatélites estudiados sugiere que existen diferencias importantes en las frecuencias con las cuales se podrían distinguir en poblaciones mezcladas:
• individuos provenientes de colonias mexicanas de aquellos provenientes de colonias costarricenses • individuos provenientes de colonias americanas de aquellos provenientes de colonias
malasias y sudafricanas.

48 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacífico mexicano
7. CONCLUSIONES Y RECOMENDACIONES En este apartado se resumen las conclusiones de relevancia para los objetivos de este proyecto,
obtenidas a partir de los resultados. Cualquier referencia a éstos debería citar las tesis respectivas identificadas en las secciones de resultados respectivas para cada técnica y especie investigada.
(1) Instalar y validar metodologías de aislamiento de DNA a partir de muestras de tejidos de tortugas marinas • Con ligeras modificaciones, los protocolos de reacción y cebadores que fueron diseñados a partir
del DNA de Chelonia mydas (Allard et al.. 1994) resultaron efectivos para la amplificación por medio de la PCR y la secuenciación de la región control del DNA mitocondrial de L. olivacea.
• Los cebadores para microsatélites de DNA nuclear de tortugas marinas, derivados por FitzSimmons et al. (1995) a partir de material de Chelonia mydas (Cm72 y Cm84) y de Caretta caretta (Cc l41) resultaron efectivos para amplificar en DNA de L. olivacea. En la tortuga laúd, un mismo éxito fuéobtenido pero con cebadores derivados de la misma especie (DC99) y de una especie de tortuga terrestre (N32) por los trabajos de Dutton (1995).
• Sería recomendable el desarrollar cebadores adicionales para amplificar microsatélites en L. olivacea con el propósito de contar con niveles intermedios de diversidad (en términos de número de alelos) que complementen los extremos en este parámetro de los aquí usados. Dado que la tendencia es para que los loci estudiados son más variables en la especie fuente (FitzSimmons, 1995), es recomendable que los nuevos cebadores provengan de L. olivacea.
(2) Cuantificar los niveles de variabilidad genética y su distribución entre y dentro de las colonias
reproductoras para determinar si existe algún indicio de reducción en los valores, como posible consecuencia de las drásticas declinaciones documentadas para las especies bajo estudio. • Dado que los niveles de variabilidad genética determinados a partir de la región control como
diversidad haplotípica (h, prom.= 0.60) y nucleotídica (π, prom. = 0.0026) en colonias de la región del Pacífico oriental (México y Centro América) para L. olivacea son comparables en magnitud a los de otras especies no se apoya la noción de erosión genética en estas poblaciones.
• Las evaluaciones de heterocigosidad en DNA nuclear en L. olivacea de la misma región, utilizando los tres loci de microsatélites, tampoco evidencian erosión de la variabilidad y son muy similares a los observados en otras especies de tortugas marinas.
• En D. coriacea, a pesar de la dramática reducción en las poblaciones del Pacífico oriental observada en la última década (Sarti et al., 1996), los valores de H en las colonias más importantes en el litoral mexicano no indican disminuciones en los niveles de variabilidad genética.
• Los niveles de variabilidad genética detectados son similares, y altos, en todas las poblaciones del conjunto estudiado para ambas especies.
(3) Analizar la composición genética de las especies, en el área de estudio para validar la hipótesis de
la existencia de una estructuración en colonias individuales ("stocks", unidades "demográficas" o "de manejo") con tendencias hacia la independencia reproductiva, como ha sido demostrado para otras especies de tortugas marinas y en otras regiones. • Con base en el análisis AMOVA para resultados de la región control del DNA mitocondrial en L. olivacea, los límites de las unidades de manejo se localizan fuera del Pacífico oriental, solamente
siendo discernibles entre esta región y el Pacífico occidental (Australia). • La totalidad de los análisis moleculares de la distribución genética (AMOVA, Fst por pares de
poblacioes, flujo génico medido por Nm) en secuencias de la región control del DNAmit en L. olivacea demuestran que no existe suficiente diferenciación interpoblacional que demuestre una tendencia hacia la independencia reproductiva entre las poblaciones del Pacífico oriental (México hasta Centro América).

Informe Final - F. Alberto Abreu Grobois, Raquel Briseño Dueñas, Ana R. Barragán Rocha 49
• Los resultados de la caracterización genética matrilineal de las poblaciones de L. olivacea en el Pacífico oriental, sugieren un comportamiento panmíctico de la especie en escalas de tiempo evolutivo (varias generaciones).
• Esta misma conclusión es apoyada y extendida por los análisis de otra región del genoma en L. olivacea, sujeto a mayores tasas de mutación (tres loci de microsatélites- Cm72, Cm84 y Cc 141).
• La hipótesis de panmixia se fortalece con datos de anidamientos en más de una playa de anidación (separadas por distancias de hasta miles de kilómetros translocaciones entre México y Centro-Sudamérica) proporcionados por los programas de marca-recaptura de L. olivacea en el Pacífico oriental.
• En D. coriacea, los resultados del análisis de la distribución de la variación genética entre las colonias más importantes del Pacífico mexicano, en dos microsatélites en el DNA nuclear (DC99 y Nigra32) demuestran también una nula tendencia hacia la diferenciación interpoblacional.
• La identificación de una tendencia hacia la panmixia en ambas especies estudiadas contrasta marcadamente con los resultados genéticos para otras tortugas marinas (p. ej., la verde y la carey) que típicamente mantienen una independencia reproductiva entre poblaciones que ocurren dentro de regiones de dimensiones similares a las que se estudió para este proyecto. Esta tendencia hacia la panmixia sugiere que el hábito filopátrico no es tan rígido en estas especies como en las otras y es congruente con el comportamiento oportunista y dinámico de la tortuga golfina y de adaptación a utilzar ambientes dinámicos de anidación en la tortuga laúd.
• Los caracteres genéticos resueltos con la metodología empleada (los haplotipos y sus frecuencias de la región control del DNAmit de L. olivacea, los alelos de microsatélites y sus frecuencias en L. olivacea y D. coriacea) serán elementos de gran utilidad para distinguir la contribución de las poblaciones de estas especies en sitios de mezcla poblacional (p. ej., rutas migratorias) sujeta a impactos por pesca incidental en el Pacífico central y otros (ver Bolten et al., 1996).
(4) Analizar las implicaciones de la información aquí generada, en programas de conservación de
tortugas marinas a nivel nacional y regional. • Para determinar las implicaciones de la información genética a los programas de conservación y
manejo, es necesario considerar las escalas de tiempo de los fenómenos bajo estudio. Si bien en escalas de tiempo evolutivo (varias generaciones) tanto la tortuga golfina como la laúd tienden hacia la panmixia en la región del Pacífico oriental (posible con apenas 4 eventos de imigración o más por generación) para escalas de tiempo relevantes para manejo (10-20 años) es necesario considerar fenómenos que ocurren con velocidades mayores.
• Una observación de relevancia que permite conocer la dinámica y evengadura de eventos propiciados por el hombre que ocurren en lapsos de pocas décadas se tiene la marcada diferencia en el comportamiento demográfico de las golfinas en las colonias de Jalisco y Guerrero (que luego del colapso por sobrepesca en los 60s no se han recuperado) y Oaxaca (que se ha incrementado excepcionalmente en los últimos cinco años). Esto sugiere un nivel de independencia entre poblaciones que limita la difusión de tortugas de una colonia a otra, dentro de escalas de tiempo más pequeñas que aquellas definibles por el análisis a partir de secuencias de la región control del DNAmit..
• El análisis de la información genética en conjunto con evidencias proporcionadas por otras líneas de investigación que muestran tendencias diferentes entre distintas colonias, sugiere que el modelo de panmixia para la tortuga golfina y la laúd en el Pacífico oriental es aplicable dentro de escalas de tiempo evolutivo (varias generaciones) pero que dentro de períodos de tiempo menores (décadas)- de relevancia para programas de manejo y conservación- la dinámica de intercambio entre colonias es demasiada lenta para la repoblación entre colonias.
• La aplicación del principio precautorio, en conjunto con el análisis integral de la información proveniente de varias líneas de investigación, permite el planteamiento de que las colonias de L. olivacea y de D. coriacea en el Pacífico oriental deben ser consideradas como unidades demográficas de manejo ("stocks") para fines de su conservación..

50 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacifico mexicano
8. AGRADECIMIENTOS Se extiende un especial reconocimiento a los Dres. Brian Bowen (Universidad de Florida) y Peter
Dutton (Southwest Fishery Science Center, NMFS) por la capacitación de los participantes en este proyecto en las técnicas de genética molecular, así como la utilización de las facilidades de sus laboratorios para la realilzación de parte de las investigaciones. A los diversos biólogos y doctores, que generosamente contribuyeron con su tiempo y esfuerzo en la colecta de material en el campo, nuestro mayor agradecimiento: Daniel Ríos, Eréndida Alvarez López, Jesús Romero Villarroel, Francisco Silva, Felipe López, Noemí Barajas Campusano, Ramiro Sánchez Pérez, Ana Rebeca Barragán, Ninel García, Luis Enrique Nava, Eduardo Ramírez, Héctor Aguilar, Mario Provencio, Miguel Hernández, Dennys Ramos, Walter Vaughan, R. M. Ball, Robert Pitman, Craig Moritz, estudiantes de la Biología de Campo de la Facultad de Ciencias UNAM
9. PERMISOS DE COLECTA La obtención de material biológico para este estudio se realizó bajo los siguientes permisos de colecta:
• Permiso No. 190396-213-03, oficio 0745, de Pesca de Fomento para la toma de muestras de tortugas marinas golfina y laúd, emitido por la Dirección General de Administración de Pesquerías de la SEMARNAP el 20 de marzo de 1996. Otorgado a Alberto Abreu y colab.
• Oficio No. A00-700-(2) 3075, Permiso de colecta para la toma de muestras de tortugas marinas golfina y laúd, emitido por la Dirección General de Aprovechamiento Ecológico de los RecursosNaturales, INE-SEMARNAP, de fecha 23 agosto 1995. Otorgado a Alberto Abreu y colab.
• Oficio No. A00-700-(2) 3843, Permiso de colecta para la toma de muestras de tortuga marina laúd, emitido por la Dirección General de Aprovechamiento Ecológico de los Recursos Naturales, INESEMARNAP, de fecha 18 de octubre de 1995 a nombre de Ana Barragán..

Informe Final - F. Alberto Abreu Grobois. Raquel Briseño Dueñas, Ana R. Barragán Rocha 51
10. LITERATURA CITADA
Allard, M.W., M.M. Miyamoto, K.A . Bjorndal, A.B. Bolten y B,W.Bowen. 1994. Support for natal homing in
green turtles from mitochondrial DNA sequences. Copeia. 1994. (1) 34-41. Avise, J.C. 1994. Molecular Markers, Natural History and Evolution. Chapman & Hall. New York, N.Y. 511
PP. Bass, A., D.A., Good, K.A. Bjorndal, J.I. Richardson, Z.M. Hillis, J.A. Horrocks y B.W. Bowen. 1996. Testing
models of female reproductive migratory behaviour and populations structure in teh Caribbean hawksbillturtle, Eretmochelys imbricata, with mtDNA sequences. Molecular Ecology 5: 321-328.
Bass, A., D.A., Good, K.A. Bjorndal, J.I. Richardson, Z.M. Hillis, J.A. Horrocks y B.W. Bowen. 1996. Testingmodels of female reproductive migratory behaviour and populations structure in teh Caribbean hawksbillturtle, Eretmochelys imbricala, with mtDNA sequences. Molecular Ecology 5: 321-328.
Bassam, B.J. y G. Caetano-Anolles. 1993. Silver staining of DNA in polyacrilamide gels. Appl. Biochem. Biotech.42: 181-188
Baverstock, P.R. y C. Moritz. 1996. Project Design. Pp. 17-28. In: Hillis, DM, C. Moritz, and BK Mable (eds)Molecuar Systematics. Sinauer Assoc., lnc.
Bolten, A.B., J.A. Wetherall, G.H. Balazs, y S.G. Pooley (comps.). 1996. Status of marine turtles in the Pacific Ocean relevant to incidental take in the Hawaii-based pelagic longline fishery. U.S. Dept. Commer., NOAA Tech. Memo., NOAA-TM-NMFS-SWFSC-230.
Bonhomme, F., S. Salvido, A. LeBeau, y G. Pasteur. 1987. Comparison genetique de tortues vertes (Chelonia mydas) des Oceans Atlantique. Indien et Pacifique: une ilutratin apparente de la theorie mullerienne classique de ka structure genetique des populations? Genetica 74:89-94.
Bowen, B. W. 1996. Literature on marine turtle population structure, molecular evolution, conservation genetics,and related topics. En: Proceedings of the International Symposium on Sea Turtles Conservation Genetics (Bowen, B. y W.N. Witzell, Eds) NOAA Tech. Memo, NMFS-SEFSC.396, National Technical information Service, Springfield, VA. 205 pp.
Bowen, B.W. y J.C. Avise. 1995. Conservation genetics of marine turtles. En: Avise, J.C. y J.L. Hamrick (eds) Conservation Genetics: case histories from nature. Chapman and Hall, New York.. pp. 190-237. Bowen
B. y S. Karl. 1996. Populations genetics, phylogeography, and molecular evolution, 29-50. En: The Biology of Sea Turtles (Lutz, P. y J. Musick, eds.) CRC Press, Boca Raton Fla. 420 pp
Bowen, B.W., A.B. Meylan, y J.C. Avise. 1989. An odyssey of the green turtle: Ascension island resisisted. Proc. Natl. Acad.Sci., USA. 86:573-576.
Bowen, B.W., A.B. Meylan, J.P.Ross, C.J.Limpus, G.H. Balazzs, y J.C. Avise. 1992. Global Population estructure and natural history of the green turtle (Chelonia mydas) in terms of matriarchal phylogeny.Evolution 46:865-881.
Bowen, B.W., W.S. Nelson y J.C. Avise. 1993a. A molecular phylogeny for marine turtles: trait mapping, rate assessment and conservation relevance. Proc. Natl. Acad. Sci. USA 90: 5574-5577.
Bowen, B.W., J.I. Richardson, A.B. Meylan, D. Margaritoulis, S. Hopkins-Murphy y J.C. Avise. 1993b. Population structure of the loggerhead turtle (Caretta caretta) in the northwest Atlantic Ocean and Mediterranean Sea. Conservation Biology 7: 834-844.
Bowen, B.W., N. Kamezaki, C.J. Limpus, G.R. Hughes, A.B. Meylan y J.C. Avise. 1994. Global phylogeographyof the loggerhead turtle (Caretta caretta) as indicated by mitochondrial DNA genotypes. Evolution 48(6): 1829-1828.
Bowen, B.W., F.A. Abreu-Grobois, G.H. Balazs, N. Kamezaki, C.J. Limpus y R.J. Ferl. 1995. Trans-Pacific migrations of the loggerhead turtle demonstrated with mitochondrial DNA markers. Proc. Natl. Adad.

52 Genética poblacional y filogeografía de las tortugas marinas golfina y laúd en el Pacifico mexicano
Sci. USA 92: 3731-3734. Bowen, B.W., A.L. Bass, A. García-Rodríguez, C.E. Diez. R. van Dam, A. Bolten, K.A. Bjorndal, M.M.
Miyamoto y R.J. Ferl. 1996. Origin of hawssbill turtles (Eretmochelys imbricata) in a Caribbean feedingarea as indicated by mitochondrial DNA sequence analysis. Ecol. Appl. (6) 566.
Briseño-Dueñas, R. y F. A Abreu-Grobois. 1994. Las tortugas Marinas y sus playas de anidación en México. Informe Final Proyecto UNAM-CONABIO FB025-P063 / 93.
Broderick, D., C. Moritz, J.D. Miller, M. Guinea, R.J. Prince and C.J. Limpus. 1994. Genetic studies of the hawskbill turtle (Eretmochelys imbricata): evidence for multiple stocks in Australian waters. Pacific Conservation Biol. 1: 123-131.
Carvalho G.R. y L. Hauser.1995. Molecular genetics and the stock concept in fisheries. 55-79. En: Molecular Genetics in Fisheries (Carvahlo, G. y T. Pitcher, Eds.) Chapman&Hall, London. 120 pp.
Chan, E.-H. y Liew, H.-C. 1996. Decline ofthe leatherback population in terengannu, Malaysia 1956-1995. Chel. Conserv. Biol. 2: 196-203
Dutton, P. 1995. Molecular Evolution of Sea Turtles with Special Reference to the Leatherback, Dermochelys coriacea . Tesis Doctoral. Texas A&M University. 137 pp.
Dutton, P. 1996. Use of molecular makers for stock identification, fingerprinting, and the study of mating behaviorin leatherback turtles. In.- Bowen, B. W. and W.N. Witzell.(Editors). Proceedings of the lnternational Syimposium on Sea Turtle Conservation Genetics. NOAA Technical Memorandum NMF-.SEFSC-396.173 pp.
Dutton, P. H. y G.H. Balazs. 1995. Simple biopsy technique for sampling skin for DNA analysus of sea turtles. Marine Turtle Newsletter 69:9-10.
Dutton, P. H., Davis, S. K.,. Guerra, T., y Owens D. 1996. Molecular phylogeny for marine turtles based on sequences of the ND4-leucina t-RNA and control region of mitochondrial DNA Mol. Phyl. Eyol. 5: 511.
Dutton, P.H., Bowen, B.W., Owens, D.W., Barragán, A.R. y Davis, S.K, 1999. Global phylogeography of the leatherback turtle (Dermochelys coriacea). J. Zool. Lond. 248: 397-409.
Eckert y Eckert, 1988 Eckert, K.L. 1993. The biology and population status of marine turtles in the North Pacific Ocean, NOAA Tech.
Memor. NMFSNOAA-TM-NMFS-SWFSC-186. Eckert, S. A. y L. Sarti. 1997. Distant fisheries implicated in the loss of the world's largest leatherback nesting
population. Marine Turtle Newsletter 78. Pp. 2-7. Eckert. K. L. 1993. The biology and status of marine turtles in the north Pacific Ocean. U.S. Dept. Commer.,
NOAA/TM/NMFS/SWFSC-186. 156 p. Encalada, S.E., P.N. Lahanas, K.A. Bjorndal, A.B. Bolten, M.M. Miyamoto y B.W. Bowen. 1996.
Phylogeography and population structure of the Atlantic and Mediterranean green turtle Chelonia mydas: a mitochondrial DNA control region sequence assessment. Molecular Ecology 5: 473-483.
Excoffier, L., P. Smouse y J. Quattro. 1992. Analysis of molecular variante inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 131:479-491.
FitzSimmons, N.N., C. Moritz y S.S. Moore. 1995. Conservation and dynamics of microsatellite loci over 300 million years of marine turtle evolution. Mol. Biol. Evol. 12: 432-440.
Frankham, R. 1995. Conservation Genetics. Ann. Rev. Genetics. 29: 305-327. Hillis, D. M., A. Larson, S. K. Davis y E. A. Zimmer. 1990. Nucleic Acids 111: Sequencing. En: Hillis, D. M. y
C. Moritz (eds). Molecular Systematics. Sinauer Associates Inc. Sunderland, MA. USA. 318-370 Hillis, DM, C. Moritz, y BK Mable. 1996. (eds) Molecuar Systematics (2nd Ed.). Sinauer Assoc., Inc. Hudson, R. R. 1990. Gene genealogies and the coalescent process. pp. 1-44 in Oxford Surveys in Evolutionary

Informe Final - F. Alberto Abreu Grobois. Raquel Briseño Dueñas, Ana R. Barragán Rocha 53
Biology, edited by Futuyama, and J. D. Antonovics. Oxford University Press, New York. Karl, S., B.W. Bowen y J.C. Avise. 1992. Global population structure and male-mediated gene flow in the
green turtle (C'helonia mvdas): RFLP analyses of anonymous nuclear DNA regions. Genetics 131:163-173.
Kichler, K. 1996. Microsatellites and conservation genetics: genetic variability and mating behaviour of the Kemp's ridley sea turtle, Lepidochelys kempii. Tesis M.C. Texas A&M University. College Station, TX
Kimura, M. 1980 A simple method for estimating evolutionary rate of base substitution through comparative studies of nucleotide sequences. J. Mol. Evol. 16:1 11-120.
Lahanas, P.N., M.M. Miyamoto, K.A. Bjorndal y A. B. Bolten. 1994. Molecular evolution and population genetics ofGreater Caribbean green turtles (Chelonia mydas) as inferred from mitochondrial DNA control region sequences. Genetica 94: 57-67.
Márquez, M.R. 1990. FAO species catalogue. Vol. 11: Sea turtles of the world. An annotated and illustrated catalogue of sea turtle species known to date. FAO Fisheries Synopsis. No. 125, Vol. 11. Rome, FAO. 81 p.
Márquez, R., A. Villanueva y C. Peñaflores. 1976. Sinopsis de datos biológicos sobre la tortuga golfina Lepidochelys olivacea (Eschscholtz, 1829) en México. INP Sinopsis sobre la Pesca (2): 61 p. Márquez, R., C. Peñaflores y J. Vasconcelos. 1996. Olive ridley turtles (Lepidochelys olivacea) show signs
of recovery at La Escobilla, Oaxaca. Marine Turtle Newsletter, 1996, No. 73: 5-7. Moritz,C. 1994. Defining "Evolutionary Significants Units" for conservation. Trends Ecology & Evolution.
9:373.375 Mortimer, J. 1995. Enseñanza de conceptos críticos para la conservación de las tortugas marinas. Noticiero de
Tortugas Marinas. No. 71: 1-4. Nei, M. 1987. Molecular Evolutionary Genetics. Columbia University Press, New York, NY, USA. 456 pp
Nicholas, K.B. y H.B. Nicholas, Jr. 1997. GeneDoc: a tool for editing and annotating multiple sequence alignments. Distributed by the authors.
Norman, J, C. Moritz, C. Limpus y R. Prince. 1994. Population genetics as a tool for managing marine turtle populations. Methodological Reviews. 101-117.
Ottenwalder, J.A. y J.P. Ross. 1992. The Cuban sea turtle fishery: description and needs for management. En: (M. Salmon y J. Wyneken, comps) Proc.11th. Workshop on Sea Turtle Biol. Conserv. NOAA Tech. Memo. NMFS-SEFC-302. pp. 90-92
Owens, D. W., and G. W. Ruiz. 1980. New methods of obtaining blood and cerebrospinal fluid from marine turtles. Herpetologica 36:17-20.
Parsons, J. 1962. The Green Turtle and Man. Univ. de Florida Press, Gainesville. 301 pp. Pritchard, P.C.H. 1969. Studies on the svtenzatics and reproductive cycles on the genus Lepidochelys. Tesis
Ph.D., University of Florida, Gainisville. 202 pp. Pritchard, P.C.H. 1979. Encyclopedia of Turtles. Ed. T.F.H. Publications, Inc. Ltd. 895 p. Proebstel, D.S., R.P. Evans, D.K. Shiozawa y R.N. Williams. 1993. Preservation of nonfrozen tissue samples
from a salmonine fish Brachymystax lenok (Pallas) for DNA analysis. Journal of ichthyology 9: 9-17. R. Valverde, pers. com.
Ríos-Olmeda, D., H. Parra, S. Robles y E. Varela. 1996. Informe final de actividades del programa «Investigación y Conservación de tortugas marinas en la playa El Verde, Sinaloa- Temporada 22va. INP-CRIP Mazatlán. 45pp.
Sarti, L., S.A. Eckert, N. García y A.R. Barragán. 1996. Decline of the world's largest nesting assemblage of leatherback turtles. Marine Turtle Newsletter 74: 2-5.
Schneider, S., J-M Kueffer, D. Roessli y L. Excoffier. 1997. Arlequin: A software for population genetic data analysis. Genetics and Biometry Laboratory, University of Geneva, Suiza. http://anthropologie.unige.ch/ arlequin

54 Genética poblacional y filogeografia de las tortugas marinas golfrna y laúd en el Pacífico mexicano
Slatkin, M. 1987. Gene flow and the geographic structure of natural populations. Science 236: 787-792. Slatkin, M. 1995 A measure of population subdivision based on microsatellite allele frequencies. Genetics 139:
457-462. Smith, M. H., H. O. Hillestad, M.N. Manlove, D.O. Straney y J.M. Dean. 1978. Management implications of
genetic variability in loggerhead and green sea turt le. Pp. 302-312. In Proceedings of the 1th International Congress of Game Biologists. 308pp
Tajima, F. 1993. Measurement of DNA polymorphism. In: Mechan isms of Molecular Evolutior. Introduction to Molecular Paleopopulation Biology, edited by Takahata, N. y Clark, A.G., Tokyo, Sunderland, MA:Japan Scientific Societies Press, Sinauer Associates, Inc., p. 37-59.
Tajima, F. y M. Nei. 1984. Estimation of evolutionary distance between nucleotide sequences. Mol. Biol. Evol. 1:269-285.
Takahata, N. y S.R. Palumbi. 1985. Extranuclear differentiation and gene flow in the finite island model. Genetics 109: 441-457.
Thompson, J.D., Higgins, D.G. y Gibson, T.J. 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucleic Acids Research 22:4673-4680.
UICN. 1994. Categorías de las Listas Rojas de la UICN. UICN, Gland. Weir, B. S. 1996. Genetic Data Analysis II: Methods for Discrete Population Genetic Data. Sinauer Assoc., Inc.
Sunderland, MA, USA. Weir, B.S. y C.C. Cockerham. 1984. Estimating F-statistics for the analysis of population structure.
Evolution 38:1358-1370. White, P. S. y L. D. Densmore. 1992. Mitochondrial DNA isolation . En: A. R. Hoezel (ed.) Molecular
Genetics Analysis of Populations: a practical approach. IRL Press. Oxford Univ. Press, N.Y. USA. 29-58.













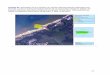



![[GEOLOGIA]O HOMEM E AS ZONAS COSTEIRAS 2015 - Artigo... · 2015. 10. 19. · espécies: Dermochelys coriacea (tartaruga-de-couro), Chelonia mydas (tartaruga-verde), Erectmochelys](https://img.pdfslide.net/doc/110x75/6119e0c28794b2637d09f459/geologiao-homem-e-as-zonas-costeiras-2015-artigo-2015-10-19-espcies.jpg)









