Embed Size (px)
Citation preview

Plant Science Letters, 23 (1981) 223--231 223 Elsevier/North-Holland Scientific Publishers Ltd.
INHIBITION OF KAURENE SYNTHETASE FROM CASTOR BEAN SEEDLINGS AND THE GERMINATION OF TOMATO, WHEAT AND CASTOR BEAN SEEDS BY DI-ETHYLENE GLYCOL DISULFIDE
YEDIDYA GAFNI and ISHAIAHU SHECHTER*
Department of Biochemistry, The George S. Wise Faculty for Life Sciences, Tel-Avw Unwersity, Ramat-Avw, Tel-Avw (Israel)
(Received February 9th, 1981) (Revision received June 1st, 1981) (Accepted June 11th, 1981)
SUMMARY
Di-ethylene glycol disulfide (ESSE) inhibits the enzyme kaurene synthe- tase derived from castor bean (Ricinus communis L.) seedlings. The inhibition affects mainly the cyclization of geranylgeranyl pyrophosphate to copalyl pyrophosphate (activity A) and had little or no effect on the further cycliza- tion of copalyl pyrophosphate to polycyclic diterpene hydrocarbons (activity B). The inhibition of activity A of kaurene synthetase has a non- competitive reversible pattern (K i = 10 ~M) with regard to geranylgeranyl pyrophosphate. ESSE inhibits the germination of tomato (Solanum lycopersicum L.) wheat (Triticum aestivum) and castor bean (Ricinus communis L.) seeds. The inhibition is reversible and normal germination was restored by washing the seeds with H20. GAs failed to promote germination of ESSE-treated tomato seeds.
INTRODUCTION
The formation of tetracyclic diterpene ent-kaurene has drawn much attention, particularly with regard to its connection to the biosynthesis of the plant growth regulating substances, the gibberellins [1--3]. ent-Kaurene has been shown to be biosynthesized in cell-free extracts from mevalonate or geranylgeranyl pyrophosphate in preparations from the endosperm of
*To whom correspondence should be sent. Abbreviations: Kaure~e, ent-kaur-16-ene (also referred to as (--)kaur-16-ene); Copalol, trivial name for the trans-isomer of ent4abda-8(16), 13-dien-15-ol; Geranylgeraniol, ali-trans-isomer; MVA, mevalonic acid; ESSE, di-ethyleae glycol disulfide, also known as di-~-hydroxyethyl disulfide.
0304--4211/81/0000--0000/$02.50 © Elsevier/North-Holland Scientific Publishers Ltd.

224
~ OPP
GERAN YLGERANYL-PP
Al
COPALYL-PP
B ~ . ~ ~ • GIBBERELLINS
AC- ent -KAURENE group (*)'SANDARACOPIMAR ADiE NE (+)-STAC H ENE (-I- TRACHYLOBANE
Fig. 1. Scheme for the biosynthesis of diterpene hydrocarbons from geranylgeranyl pyrophosphate.
immature seeds of wild cucumber (Marah macrocarpus) [4,5], in the fungus Fusarium moniliforme [6], in castor bean seedlings (Ricinus cornmunis L.) [7], in pea (Pisum sativum L.) shoot tips [8] and fruits and in cell suspen- sion cultures of tomato (Solanum lycopersicum L.) and tobacco (Nicotiana tabacum L.) [9]. The biosynthesis of ent-kaurene from geranylgeranyl pyrophosphate by kaurene synthetase is known to proceed via a two-step mechanism; (a) the production of bicyclic diterpene copalyl pyrophosphate; (b) further conversion of the latter to kaurene [6]. In castor bean extracts three additional cyclic diterpene hydrocarbons are being produced from copalyl pyrophosphate as shown in Fig. 1 and, together with ent-kaurene, they are known as the C-group [ 7 ].
The reaction of kaurene synthetase has been shown to be the site of inhibition by some plant growth retardants [6,10] and it was suggested to be the site of regulation of the gibberellin biosynthetic pathway on theoretical grounds because of the role of geranylgeranyl pyrophosphate as a branch point metabolite in the biosynthesis of diterpenoids and carotenoids [2,3].
In previous work from our laboratory we reported the production of an inhibitor of kaurene synthetase in castor bean seedlings and cell suspension cultures [11,12]. This inhibitor was shown to be specific for activity A of kaurene synthetase derived from F. moniliforme and seedlings of castor bean. The kinetic pattern of its inhibition was shown to be non-competitive.

225
In a later work we isolated and identified ESSE from castor bean cell suspen- sion cultures [ 13].
The objective of the work described in this report was to ascertain that ESSE is indeed a specific inhibitor of the activity A of kaurene synthetase which would agree with it being the previously described inhibitor [12]. In addition, we investigated the effect of ESSE on the germination of tomato, wheat and castor bean seeds.
MATERIALS AND METHODS
Germination of tomato, wheat and castor bean seeds Seeds of tomato (S. lycopersicum L.) and wheat (T. aestivum) were
imbibed in tap water for 5 h and then were spread in 35 mm Petri dishes containing 4 sheets of Whatman No. 1 filter paper and 1.0 ml of water. Germination of castor bean seeds was done according to Robinson and West [7]. Briefly, the seeds were mechanically freed of their coats and then were spread on 4 sheets of Whatman No. 1 filter paper in 90 mm Petri dishes containing 10 ml of water. The seeds were germinated at 24°C in the dark. Whenever requried, solutions of ESSE or GA3 or both were used for the germination. For the purpose of germination in the presence of ESSE a stock solution of 25 mM was prepared ('Inh.'). A series of dilution of this stock solution was used in each experiment. After 7 days the dishes contain- ing the seeds were photographed, seeds from every dish were divided into two separate dishes and one of each duplicate dishes was rinsed 5 times with 1.0 ml of water. Water (1 ml) was added to every washed dish and all the seeds were allowed to germinate for additional 2 days at 24°C.
Preparation of enzymic extracts Enzymic extracts were prepared from castor bean germinating seeds.
Four-day-old seedlings were harvested and washed thoroughly on filter paper with 0.1 M Tris--HC1 buffer (pH 7.4). The samples were transferred to a precooled mortar and ground in liquid nitrogen to a fine powder. Polyvinylpyrrolidone was added at a ratio of 0.25 g to every gram of germin- ating seeds [14]. After mixing, the paste was allowed to warm to 4°C and 3.0 ml of Tris--HC1 buffer were added to every gram of paste. The mixture was squeezed through four layers of cheesecloth and the effluent was centri- fuged for 30 rain at 35 000 × g. The supernatant was collected and the protein concentration was adjusted t o 1 0 mg/ml by dilution with Tris--HC1 buffer as above. This enzymic extract will be referred to as S3s preparation. Determination of protein concentration was done according to method of Bradford based on binding of the protein to the dye Coomassie brilliant blue G-250 [15].
Enzyme assays The incubation mixture in 1.0 ml, contained 5.0 mM MgC12, 0.5 ml of

226
$3, extract, Tris--HC1 buffer (pH 7.4) and radioactive substrate. The follow- ing amounts of substrate were used in the various incubations: [14C]- geranylgeranyl pyrophosphate, 40 nM (104 cpm) and [ ~4C] copalyl pyrophos. phate, 40 nM (104 cpm). The reactions were started by the addition of the enzyme. The assays were run at 30°C for 20 min. At the end of the incuba- tion period, the reactions were stopped by addition of 1.0 ml of acetone.
Analysis of labeled products by thin-layer chromatography Reaction mixtures were extracted three times with n-hexane. The com-
bined extracts were concentrated under a stream of N2 and applied on 2 × 20 cm chromatography plates. Separation of alcohols from kaurene was done on silica gel plates developed with 10% ethyl acetate in benzene or, in some instances, developed with n-hexane. Separation of hydrocarbons of the C-group was done on silica gel plates impregnated with 10% AgNO3 and developed with n-hexane-benzene (7 : 3) [6].
The radioactivity of materials absorbed on silica gel plates was determined by scanning for radioactivity (Packard Model 7200). For quantitative ana- lysis, zones of the gel were scrapped off and counted for radioactivity (Packard Model Prias). The efficiency of counting of 14C-labeled materials under these conditions was 85%. Non-radioactive reference standards were detected by exposure of the plates to iodine vapour or, in case of AgNO3 plates, by spraying them with ethanolic solution of p-anis-aldehyde and sulfuric acid [16].
Materials [1-14C] R,S-mevalonic acid (MVA) (22.6 Ci/mol) as the N,N'-dibenzyl-
ethylenediamine salt was purchased from the Radiochemical Center, Amersham, U.K. ent-Kaurene standard was obtained through the generosity of Dr. C.A. West, UCLA. Precoated silica gel plates (F-254) in various thick- ness were purchased from Merck.
Preparation of [*4C] geranylgeranyl pyrophosphate and [14C] copalyl pyrophosphate
The above radiolabeled materials were synthesized according to the pro- cedure of Gafni and Shechter [17].
Preparation of ESSE ESSE was synthesized similarly to a previously described procedure [13].
Briefly, to I0 ml of a 2.5 M solution of thioethylene glycol at pH 12, 20 mg of CuSO4 were added and air was bubbled through for 3 h. At the end of 3 h the solution was extracted 5 times with 3.0 ml of ethyl acetate. The combined ethyl acetate extracts were concentrated under a stream of N2 and ESSE was purified on preparative thin-layer chromatography. The purified ESSE was dissolved in H20 and its concentration was determined by measur- ing its absorbance at247 n m [ 13]. Quantitative determination of its disulfide content was done by the Ellman's procedure [18].

227
RESULTS
Inhibition of activity A of haurene synthetase in castor bean Sas enzymic extract
The effect of ESSE on the production of [~4C] diterpene hydrocarbons (the C-group) from [ 14C] geranylgeranyl pyrophosphate and [ 14C ] copalyl pyrophosphate in S3s enzymic extract of castor bean seedlings was investi- gated. ESSE (25 pM) inhibited the synthesis of ~4C-hydrocarbons from [ ~4C] geranylgeranyl pyrophosphate but not from [ ~4C] copalyl pyrophos- phate (Table I). As shown, at a concentration of 25 pM of ESSE about 60% inhibition of activity A was observed whereas activity B was hardly affected, At concentrations of ESSE above 1 mM partial inhibition of activity B was also observed. At these concentrations, activity A was totally inhibited. The inhibition of the production of [14C] kaurene from [~4C]- geranylgeranyl pyrophosphate by ESSE is virtually identical to the inhibition of the production of the whole C-group as shown in Table I. Kinetic analysis of the inhibition indicates that ESSE is a non-competitive inhibitor of the enzyme (Fig. 2). Apparent K i of 10/~M was obtained.
Effect of ESSE on the germination of tomato wheat and castor bean seeds Tomato, wheat and castor bean seeds were germinated in the presence of
increasing concentrations of ESSE, obtained by a series of dilutions of a 25 mM solution of ESSE in water. ESSE inhibited seed germination of the three different plant species to similar extent. We chose to present the inhibition of the germination of tomato seeds by ESSE purely because of the ease at which it can be presented.
Inhibition exerted by ESSE affected both the percentage of tomato seeds that germinated and the degree of growth of the seedlings. In general, at
TABLE I
INHIBITION BY ESSE OF THE PRODUCTION OF CYCLIC DITERPENE HYDROCARBONS FROM [ 14C ] GERANYLGERANYL PYROPHOSPHATE OR [ 14C] COPALYL PYROPHOSPHATE BY CASTOR BEAN EXTRACTS
Additions Total C-group Inhibition ent-Kaurene Inhibition hydrocarbons (%) produced (%) produced (pmol/min/mg prot .) (pmol/rain/rag prot . )
[~4C]GeranyIgeranyl pyrophosphate None 3.07 0 0.89 0 25 #M ESSE 1.25 59.3 0.35 60.7
[14 C] Copaly l pyrophosphate None 7.83 0 2.71 0 25 ~M ESSE 7.53 3.83 2.64 2.6

228
7
"E" ~ $
f
?
T > 2
2 -20 -10
i i w v i i
/ 75~JM
I
k
I0 20 30 40 50 6 0
[GGPP] -I ,uM-I
Fig. 2. Lineweaver-Burk plots. Inhibition of kaurene synthetase in $3~ extract of castor bean seedlings by various concentrations of ESSE. The incubation mixtures contained in 1.0 ml; 0.5 ml S3s enzyme preparation, Tris--HCl buffer (pH 7.4), 5 mM MgCl~ and various amounts of [14C] geranylgeranyl pyrophosphate and of ESSE. Incubation condi- tions and analysis of [ 14C]kaurene was done as described in Materials and Methods. Insert shows variations of V~a x as function of inhibitor concentrations. K i was calcu- lated from this plot.
lower concentrations, ESSE merely delayed the germination of the seeds wi thout any noticeable effect on the percentage of seed germination. How- ever, higher concentrations of ESSE lowered the percentage of seed germina- tion in addition to the delay in germination time. Thus, after 7 days at 0.5 mM ESSE in the germination media (dilution 1 : 50, Fig. 3) only half of the seeds germinated.
When ESSE was washed ou t with water, germination of ESSE-treated tomato seeds is clearly observed after 2 days, even in seeds that were exposed to relatively high concentrations of ESSE (2.5 mM = 1 : 10).
ESSE inhibited seed germination also in the presence of GA3. We could no t detec t any difference in inhibition of seed germination by ESSE, be- tween seeds that were germinated in the presence of 3 mM, 300 ~M and 30 juM of added GA3. The percentage of germination and the size of the seedlings at all three concentrations of GA3 were the same as those of seeds germinated in the absence of GA3.

229
Fig. 3. Effect of ESSE on tomato seed germination. Tomato seeds were spread on wet filter paper in 35 mm Petri dishes containing 1.0 ml of diluted solutions of 25 mM ESSE. The dilution factor is written under each Petri dish. Germination in the presence of undiluted 25 mM ESSE ('Inh.') and water is shown. The seedlings had been germin- ated for 7 days.
DISCUSSION
The inhibition of seed germination (Fig. 3) and the reversal of the inhibi- t ion by rinsing the seeds with water (Fig. 4) indicate that ESSE penetrates the seeds quite easily to reach its target(s). Once there, it can dissociate out again without a permanent damage to the germination process. The revers- ible effect of ESSE is also observed by the kinetic analysis of the inhibition of kaurene synthetase. A reversible non-competitive inhibition is obtained (Fig. 2). Similar to many other growth retardants [6,10] ESSE inhibits at low concentrations (25 pM), specifically the activity A of kaurene synthetase
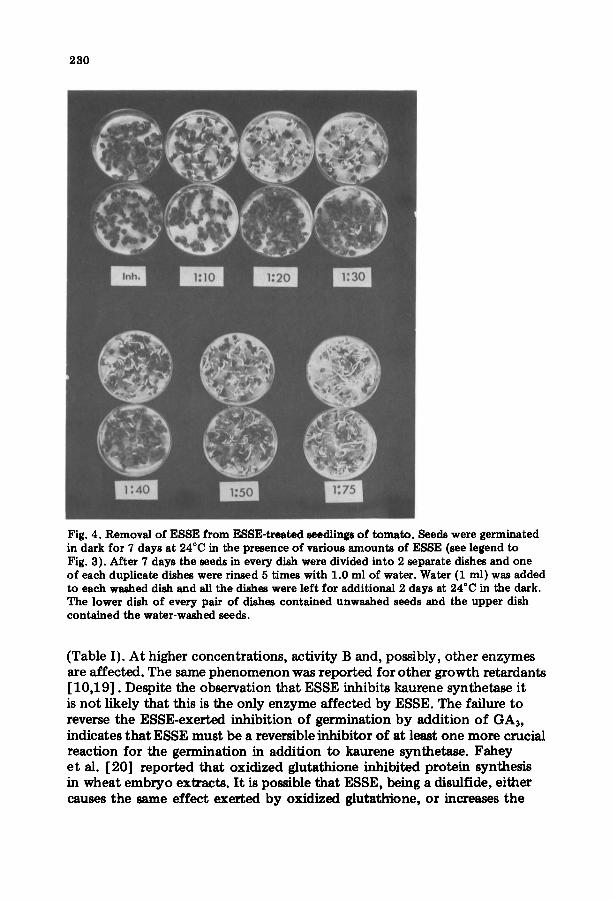
230
Fig. 4. Removal of ESSE from ESSE-treated seedlings of tomato. Seeds were germinated in dark for 7 days at 24°C in the presence of various amounts of ESSE (see legend to Fig. 3). After 7 days the seeds in every dish were divided into 2 separate dishes and one of each duplicate dishes were rinsed 5 times with 1.0 ml of water. Water (1 ml) was added to each washed dish and all the dishes were left for additional 2 days at 24°C in the dark. The lower dish of every pair of dishes contained unwashed seeds and the upper dish contained the water-washed seeds.
(Table I) . A t higher concen t r a t i ons , ac t iv i ty B and, poss ib ly , o the r e n z y m e s are a f fec ted . T h e same p h e n o m e n o n was r e p o r t e d f o r o t h e r g r o w t h r e t a rdan t s [ 1 0 , 1 9 ] . Desp i t e the obse rva t ion t h a t ESSE inhibi ts kau rene syn the t a se i t is n o t l ikely t ha t this is the on ly e n z y m e a f f e c t ed b y ESSE. T h e fai lure t o reverse the ESSE-exe r t ed inh ib i t ion o f g e r m i n a t i o n b y add i t i on o f GA3, indica tes t h a t ESSE m u s t be a revers ib le i nh ib i t o r o f a t least o n e m o r e crucia l r eac t i on fo r the ge rmina t i on in add i t ion to kau rene syn the tase . F a h e y e t ai. [20] r e p o r t e d t h a t ox id ized g lu t a th ione inh ib i t ed p ro t e in synthes is in w h e a t e m b r y o ex t rac t s . I t is poss ib le t h a t ESSE, be ing a disulf ide, e i ther causes t h e same e f f ec t exe r t ed b y ox id ized g lu t a th ione , o r increases t h e

231
c o n c e n t r a t i o n o f the cel lular c o n t e n t o f ox id ized g lu ta th ione by some disulf ide in te rchange mechanism. Whatever the exp lana t ion , the ease at which ESSE pene t ra tes the seeds and diffuses o u t renders i t a useful r eagen t f o r the r e t a rda t ion o f seeds germinat ion .
In a f o r m e r paper [12] we showed t ha t CB-I cell suspension cul tures p r o d u c e an inh ib i to r o f act ivi ty A o f kaurene synthe tase . Later , we isolated and ident i f ied ESSE f rom CB-I cul tures [ 1 3 ] . As r epor t ed , the molecu la r weight of the then u n k n o w n inh ib i to r o f act ivi ty A o f kaurene syn the tase , isolated by an earlier p rocedu re , was 7500 da l tons [ 1 2 ] . Because o f the inh ib i t ion pa t t e rn , we specu la ted tha t the inh ib i to r m a y be b o u n d to a carr ier p ro te in or pep t ide . The p rocedu re used for the isolat ion of ESSE (mol . wt., 154) involved ex t r ac t i on wi th e thy l ace ta te [ 1 3 ] . Such extrac- t ion disrupts any non-cova len t l inkage to a carr ier p ro te in . The re fo re , if a carr ier p ro t e in does exist i ts size should be near ly 7500 dal tons .
REFERENCES
1 P. Hedden, J. MacMillan and B.O. Phinney, Annu. Rev. Plant Physiol., 29 (1978) 149. 2 C.A. West, Biosynthesis of gibberellins, in: B.V. Milborrow (Ed.), Biosynthesis and its
Control in Plants, Academic Press, London, 1972, p. 143. 3 C.A. West, M.W. Dudley and M.T. Dueber, Rec. Adv. Phytochem., 13 (1979) 163. 4 M.O. Oster and C.A. West, Arch. Biochem. Biophys., 127 (1968) 112. 5 C.D. Upper and C.A. West, J. Biol. Chem., 242 (1967) 3285. 6 I. Shechter and C.A. West, J. Biol. Chem., 244 (1969) 3200. 7 D.R. Robinson and C.A. West, Biochemistry, 9 (1970) 80. 8 R.C. Coolbough, T.C. Moore, S.A. Barlow and P.R. Ecldund, Phytochemistry, 12
(1973) 1613. 9 Y. Yafin and I. Schechter, Isr. J. Med. Sci., 15 (1979) 78.
10 R.R. Fall and C.A. West, J. Biol. Chem., 246 (1971) 6913. 11 Y. Gafni and I. Shechter, Isr. J. Med. Sci., 15 (1979) 78. 12 Y. Gafni and I. Shechter, Plant Physiol., 67 (1981) 1169. 13 Y. C-afni and I. Shechter, Phytochemistry, (1981) in press. 14 W.D. Loomis and J. Battaile, Plant Phytochem., 5 (1966) 423. 15 M.M. Bradford, Anal. Biochem., 72 (1976) 248. 16 P.J. Dumphy, J.D. Kerr, J.F. Pennoch and K. Whittle, Chem. Ind., (1966) 1949. 17 Y, Gafni and I. Shechter, Anal. Biochem., 92 (1979) 246. 18 G.L. EUman and H. Lysco, J. Lab. Clin. Med., 70 (1967) 518. 19 T.J. Douglas and L.G. Paleg, Plant Physiol., 49 (1972) 417. 20 R.C. Fahey, D.L. DiStephano, G.P. Meier and R.N. Bryan, Plant Physiol., 65 (1980)
1062.





























