Embed Size (px)
Citation preview

ti»-•••";V:!*y*
[CANCER RESEARCH 45. 4349-4353, September 1985]
Inhibition of Tumor Cell Colony Formation in Culture by a Monoclonal Antibodyto Endogenous Lectins1
Reuben Lotan,2 Dafna Lotan, and Avraham Raz3
Department of Tumor Biology, The University of Texas M. D. Anderson Hospital and Tumor Institute at Houston, Texas 77030 [R. L, D. L], and Department of Cell Biology,The Weizmann Institute of Science, Rehovot 76100, Israel ¡A.R.]
ABSTRACT
The presence of endogenous, galactoside-specific lectin mol
ecules on the surface of various neoplastic cells has beendemonstrated recently using monoclonal antibody (mAb) 5D7[Raz et al., EMBO (Eur. Mol. Biol. Organ.) J., 3: 2979, 1984].The effect of this mAb on the growth of several transformed andtumor cell lines of murine and human origin was investigatedusing in vitro techniques. A dose-dependent reduction (30 to
100%) in colony formation on a solid substrate or in a semisolidmedium was observed when the cells were cultured in thepresence of 15 to 100 ^g of mAb 5D7 per ml of medium. Inhibitionof anchorage-independent growth was more pronounced (2- to3-fold) than inhibition of anchorage-dependent growth for mostof the cells. The growth-inhibitory effects of mAb 5D7 were not
the result of a cytolytic activity, for neither DMA nor proteinsynthesis was suppressed in semiconfluent cell cultures after 3days of exposure to the antibody. Other mAbs that recognizecell surface components, such as chondroitin sulfate or fibronec-
tin, failed to inhibit colony formation. These results suggest thatendogenous tumor cell-surface lectin molecules may be involved
in intercellular interactions or interactions between the cells andexogenous ligands; these interactions are important for growthregulation.
INTRODUCTION
Most of the interactions between cells and their environment,in vivo or in culture, are mediated via cell membrane componentsthat serve as receptors for hormones and growth factors or playa role in recognition and adhesion. Consequently cell surfaceproteins and glycoconjugates have been implicated in the regulation of various fundamental cellular processes including growth,differentiation, and cognitive cellular interactions among adjacentcells or between cells and the extracellular matrix (1).
Malignant transformation is often accompanied by alterationsin cell surface components (2). Some of the modified componentsmay be important for the maintenance of the transformed phe-notype in vitro (e.g., anchorage-independent growth) or for theexpression of tumorigenicity and metastatic properties in vivo(3-5).
Several years ago, we described the presence of endogenouslectins (carbohydrate-binding proteins) in extracts of different
human and murine tumor cells and proposed that these molecules mediated cellular recognition and adhesion (6). This sug-
'This investigation was supported by USPHS Grant 7R01 CA 39422-01awarded by the National Cancer Institute, Department of Health and HumanServices.
2To whom requests for reprints should be addressed.3Incumbent of the Sophie M. T. and Richard S. Richards Career Development
Chair in perpetuity.Received4/2/85; accepted 6/12/85.
gestion was based on our finding that the addition of desialylatedfetuin to tumor cell suspensions promoted cell aggregation following the binding of this glycoprotein to the cell surface (6).Although these aggregation experiments were performed in vitro,they may be indicative of similar aggregation in vivo, inasmuchas a murine melanoma cell variant that we selected for reducedasialofetuin-mediated aggregation exhibited a concomitant de
crease in aggregation in the presence of mouse serum (7). Moreimportant, these variant cells exhibited a marked decrease incolony formation in agarose as well as a reduced colonization oflungs of syngeneic mice following injection into the tail vein (7).
Recently we produced a mAb" directed against a galactose-
specific lectin extracted from B16-F1 melanoma cells (8). This
mAb, designated 5D7, was found to inhibit lectin activity ofextracts from a variety of human and murine tumor cells and tobind to their surface membrane (8). In the present study, weused the mAb 5D7 to examine the possibility that tumor cell-surface galactose-specific lectin molecules are involved in the
regulation of cell growth.
MATERIALS AND METHODS
Cell and Culture Conditions. The following cells were used: mousemelanoma B16-F1 (9); mouse fibrosarcoma UV-2237-IP3 (10); humanmelanoma Hs939 (11); SV40-transformed human fibroblast cell lineSV80; and oncogene-transfected and transformed rat embryo second-ary-fibroblast cell line SPOT. These cells were obtained by transfection
of secondary rat embryo fibroblasts with a mixture of the following DMAs:10 »¿gof pLA-8 containing the adenovirus-2/EIA and part of the EIBregion; 10 ^g of PEJ6.6 containing the activated C-ras"" gene; and 5 ^g
of BALB/c liver DMA. After transfection, cells were cloned in agar, andone of these clones was developed into a transformed cell line designatedSPOT. All the cells were grown in plastic tissue culture dishes in Dulbec-co's modified minimal essential medium containing 10% heat-inactivated
fetal bovine serum, nonessential amino acids, and antibiotics. The cellswere maintained at 37°C in a humidified atmosphere of 7% CO2 and
93% air. Cells were harvested after a brief incubation with 2 mw EDTAin CMF-PBS and washed in CMF-PBS. The cells were resuspended
gently by repeated pipetting, using a Pasteur pipet to dissociate cellclumps into a suspension of single cells. Only cell suspensions exhibiting>90% viability (determined from the proportion of cells that excluded0.1% trypan blue) were used in these studies for assays of colonyformation.
Monoclonal Antibodies. The production of hybridomas secreting mAbto a galactoside-specific endogenous lectin of B16-F1 melanoma cells
has been described recently (8). To obtain the antibody, hybridoma cellswere injected ¡.p.into pristane-primed C57BL/6 x BALB/c Ft mice, and
ascitic fluid was isolated. The ascitic fluid was used either directly or
4The abbreviations used are: mAb, monoclonal antibody; CMF-PBS, Ca2*-andMg2*-free phosphate-buffered salinecontaining, per liter H20, 8.0 g NaCI, 0.2 g
KCI, 1.5 g NazHPO4,and 0.25 g KH2PO«;TCA, trichloroacetic acid; ID.»,dosecausing 50% inhibition of growth; CH, cycloheximide;ara-C, 1-0-D-arabinofurano-sylcytosine; FACS, fluorescence-activatedcell sorter.
CANCER RESEARCH VOL. 45 SEPTEMBER 1985
4349
on April 3, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
TUMOR GROWTH INHIBITION BY ANTILECTIN mAb
after ammonium sulfate precipitation and extensive dialysis. In someexperiments we used mAb to fibronectin, or to chondroitin sulfate, thatwere obtained from Dr. B. Geiger (The Weizmann Institute of Science).The mAb to Ly-1 antigen was obtained from Dr. S. Katzav (The Weiz
mann Institute).Assays for Inhibition of Colony Formation. Two assays for inhibition
were used. One analyzed colony formation on plastic tissue culturedishes by cells that were attached to the solid surface, and the otheranalyzed colony formation by cells suspended in semisolid medium. Forthe first assay, suspensions of single cells were seeded at clonal density(500 to 2000) in 6-cm-diameter dishes, and 24 h later, the medium was
replaced with fresh medium with or without antilectin mAb or controlimmunoglobulin. The cells were grown for 15 days, and then the colonieswere stained with méthylèneblue (0.5% in 70% isopropyl alcohol) andcounted. For the second assay, cells were suspended in medium (8 to16x102/ml) with or without antilectin mAb or control immunoglobulinand incubated for 30 min at 37°C. The cells were then mixed 1:1 (vol/
vol) with a solution of 1% agarose with or without antilectin mAb orcontrol immunoglobulin, and 1-ml aliquots of the mixture were placed ontop of a precast layer of 1% agarose in 3.5-cm-diameter dishes. The
number of colonies that formed after 15 days was determined using aninverted phase microscope. The assays were performed in triplicate in 3independent experiments. Results are expressed as the percentage ofthe number of colonies that formed in cultures grown without anyantibody.
Analysis of DMA and Protein Synthesis. Cells were plated in Costar24-well tissue cultures plates at 5 x 103 cells per well and cultured for
24 h before the addition of medium containing antilectin mAb 507, anti-Ly-1 (both at 50 ^g/ml), or control medium. After an additional 3 days,the cells were labeled with either [3H]leucine (148 Ci/mmol; AmershamCorp., Arlington Heights, IL) or [3H)thymidine (49 Ci/mmol; Nuclear
Research Center, Negev, Israel), both at 1 ¿iCi/mlfor 6 h. Some culturesalso received CH or ara-C, both at 50 ^g/ml, to inhibit protein and DNA
synthesis, respectively. At the end of the radiolabeling incubation, themedium was removed by aspiration, and the cells were rinsed rapidlywith ice-cold PBS and exposed briefly to a solution of 0.25% trypsin: 2
HIM EDTA. When the cells were rounded up, the trypsin was neutralizedwith soybean trypsin inhibitor, and the cells were suspended in 1 ml ofan ice-cold solution of 20% TCA. The precipitates that formed were
collected on GF/C glass fiber filters and washed first with 5% TCA, thenwith ethyl alcohol. The radioactivity remaining on the filters was thencounted in a Packard PLD Tri-Carb scintillation counter using a Lu-
maxixylene scintillation fluid.Analysis of mAb 5D7 Binding to Cells. Binding of mAb 5D7 to the
cell surface was quantitated by indirect immunofluorescence labelingfollowed by FACS analysis.
Cells from subconfluent cultures were detached by incubation with 2rriM EDTA in CMF-PBS, washed in PBS, and suspended at 3 x 107 cells/
ml in PBS containing 0.02% NaN3. Aliquots containing 0.1 ml of thissupension were mixed with 50 to 100 ¿igof mAb 5D7 in 50 ¡Aof PBSand incubated at 4°Cfor 30 min. The cells were then washed twice in
PBS and incubated with a 1:60 dilution of fluorescein-labeled rabbit anti-
mouse immunoglobulin antibodies (Nordic Immunological Laboratory) for30 min at 4°C.The cells were then washed twice in PBS and analyzed
in FACS II (Becton Dickinson). A scatter window was set to eliminatedead cells and cell debris. Ten thousand cells were analyzed for eachcell line using a laser output of 400 mV and fluorescence gain 8. Thefrequency and fluorescence distribution profiles were used to quantitatethe binding of the antibody by comparing the areas under the fluorescence profiles of different cell lines to the area under the profile obtainedwith B16-F1 cells, which was taken as 1.
RESULTS
In previous studies we found that UV-2237-IP3 murine fibro
sarcoma cells expressed relatively high levels of endogenous
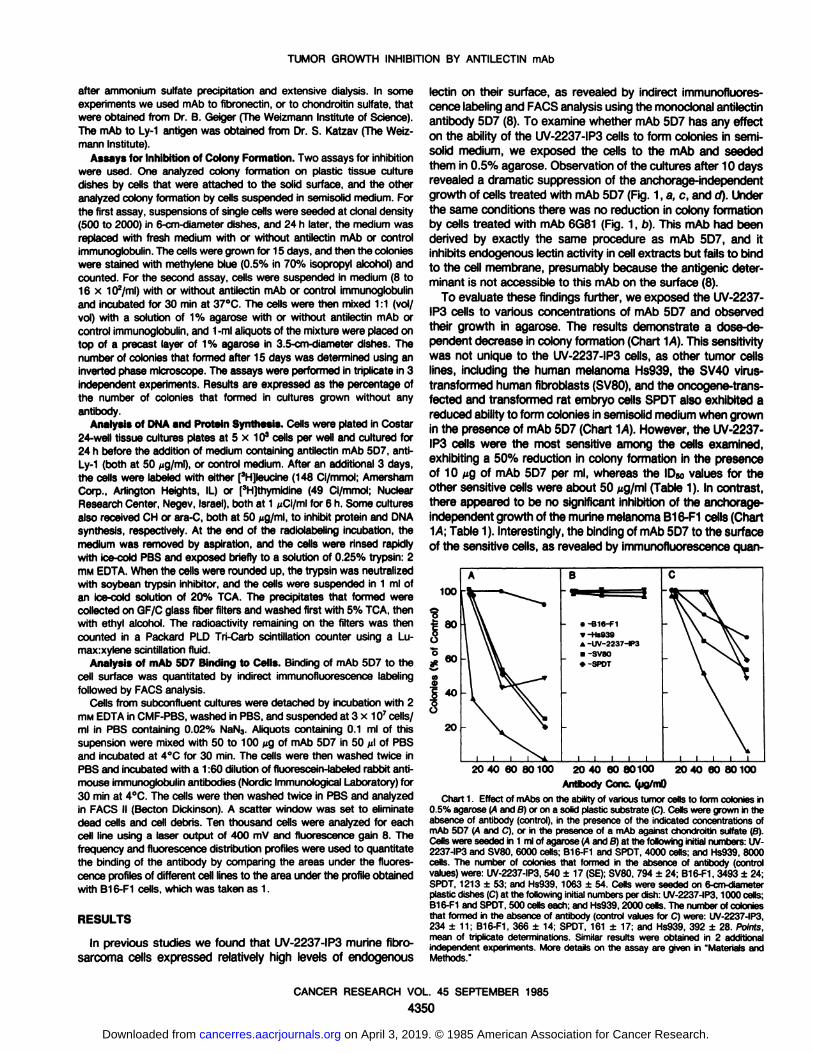
lectin on their surface, as revealed by indirect immunofluorescence labeling and FACS analysis using the monoclonal antilectinantibody 5D7 (8). To examine whether mAb 5D7 has any effecton the ability of the UV-2237-IP3 cells to form colonies in semi-
solid medium, we exposed the cells to the mAb and seededthem in 0.5% agarose. Observation of the cultures after 10 daysrevealed a dramatic suppression of the anchorage-independent
growth of cells treated with mAb 5D7 (Fig. 1, a, c, and d). Underthe same conditions there was no reduction in colony formationby cells treated with mAb 6G81 (Fig. 1, b). This mAb had beenderived by exactly the same procedure as mAb 5D7, and itinhibits endogenous lectin activity in cell extracts but fails to bindto the cell membrane, presumably because the antigenic determinant is not accessible to this mAb on the surface (8).
To evaluate these findings further, we exposed the UV-2237-
IP3 cells to various concentrations of mAb 5D7 and observedtheir growth in agarose. The results demonstrate a dose-dependent decrease in colony formation (Chart "\A). This sensitivity
was not unique to the UV-2237-IP3 cells, as other tumor cellslines, including the human melanoma Hs939, the SV40 virus-transformed human fibroblasts (SV80), and the oncogene-trans-fected and transformed rat embryo cells SPOT also exhibited areduced ability to form colonies in semisolid medium when grownin the presence of mAb 5D7 (Chart 14). However, the UV-2237-
IP3 cells were the most sensitive among the cells examined,exhibiting a 50% reduction in colony formation in the presenceof 10 ^g of mAb 5D7 per ml, whereas the ID50 values for theother sensitive cells were about 50 ^g/ml (Table 1). In contrast,there appeared to be no significant inhibition of the anchorage-independent growth of the murine melanoma B16-F1 cells (Chart~\A;Table 1). Interestingly, the binding of mAb 5D7 to the surface
of the sensitive cells, as revealed by immunofluorescence quan-
100
•-B16-F1
»-HS939A -UV-2237-IP3
•-SV80
•-SPOT
J \ L J 1 1 L_20406080100 20406080100 20406080100
AntibodyCone,(ug/ml)Chart 1. Effect of mAbs on the ability of various tumor cells to form colonies in
0.5% agarose (A and B) or on a solid plastic substrate (C).Cells were grown in theabsence of antibody (control), in the presence of the indicated concentrations ofmAb 5D7 (A and C), or in the presence of a mAb against chondroitin sulfate (B).Cells were seeded in 1 ml of agarose (A and B) at the following initial numbers: UV-2237-IP3 and SV80, 6000 cells; B16-F1 and SPOT, 4000 cells; and Hs939, 8000cells. The number of colonies that formed in the absence of antibody (controlvalues)were: UV-2237-IP3,540 ±17 (SE);SV80, 794 ±24; B16-F1, 3493 ±24;SPOT, 1213 ±53; and Hs939, 1063 ±54. Cells were seeded on 6-cm-diameterplastic dishes (C)at the following initial numbers per dish: UV-2237-IP3,1000 cells;B16-F1 and SPOT,500 cells each; and Hs939, 2000 cells. The number of coloniesthat formed in the absence of antibody (control values for C) were: UV-2237-IP3,234 ±11; B16-F1, 366 ±14; SPOT, 161 + 17; and Hs939, 392 ±28. Points,mean of triplicate determinations. Similar results were obtained in 2 additionalindependent experiments. More details on the assay are given in 'Materials andMethods."
CANCER RESEARCH VOL. 45 SEPTEMBER 1985
4350
on April 3, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
TUMOR GROWTH INHIBITION BY ANTILECTIN mAb
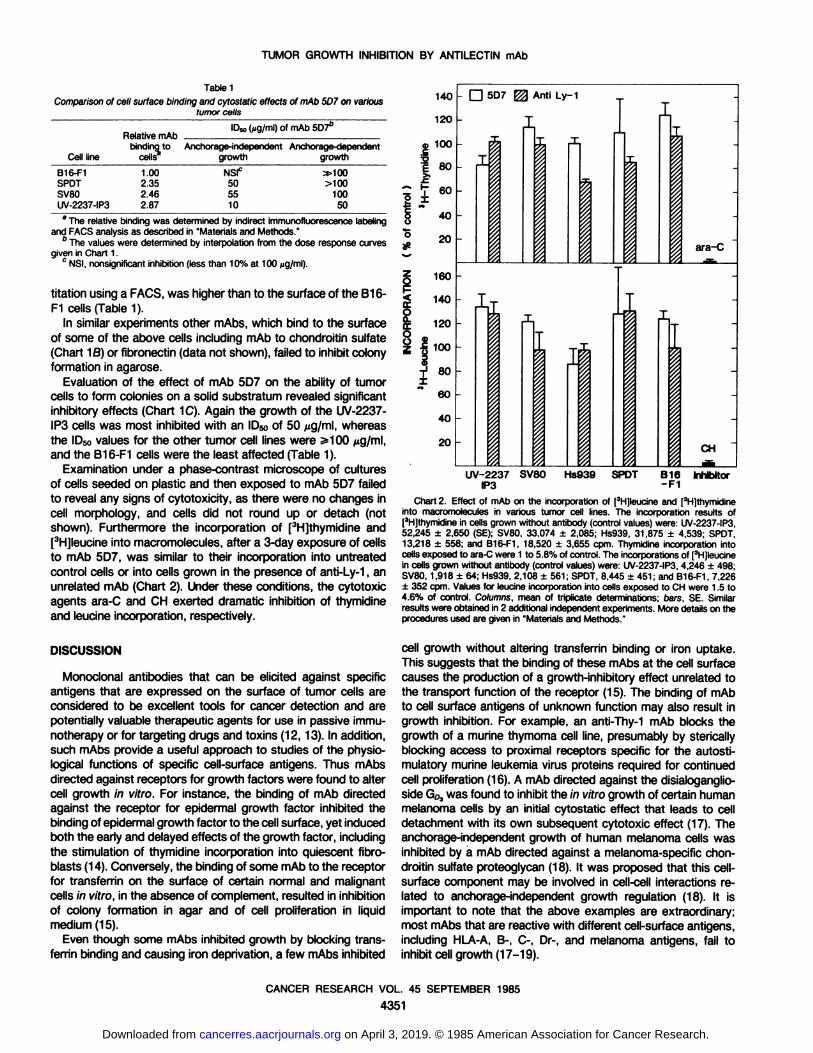
Table 1Comparisonof cell surface binding and cytostatic effects of mAb 5D7 on various
tumor cells
ICell
lineB16-F1
SPOTSV80UV-2237-IP3-¡elative
mAbID«,(iig/ml) of mAb5D76binding
to Anchorage-independentcells^growth1.00
2.352.462.87NSIC
505510Anchorage-dependent
growth»100
>10010050
The relative binding was determined by indirect immunofluorescencelabelingand FACS analysis as described in "Materials and Methods."
6 The values were determined by interpolation from the dose response curves
given in Chart 1.c NSI, nonsignificant inhibition (less than 10% at 100 n9/ml).
titation using a FACS, was higher than to the surface of the B16-
F1 cells (Table 1).In similar experiments other mAbs, which bind to the surface
of some of the above cells including mAb to chondroitin sulfate(Chart 1S) or fibronectin (data not shown), failed to inhibit colonyformation in agarose.
Evaluation of the effect of mAb 5D7 on the ability of tumorcells to form colonies on a solid substratum revealed significantinhibitory effects (Chart 1C). Again the growth of the UV-2237-
IP3 cells was most inhibited with an IDsoof 50 /¿g/ml,whereasthe ID50values for the other tumor cell lines were ^100 ^g/ml,and the B16-F1 cells were the least affected (Table 1).
Examination under a phase-contrast microscope of cultures
of cells seeded on plastic and then exposed to mAb 5D7 failedto reveal any signs of cytotoxicity, as there were no changes incell morphology, and cells did not round up or detach (notshown). Furthermore the incorporation of [3H]thymidine and[3H]leucine into macromolecules, after a 3-day exposure of cells
to mAb 5D7, was similar to their incorporation into untreatedcontrol cells or into cells grown in the presence of anti-Ly-1, an
unrelated mAb (Chart 2). Under these conditions, the cytotoxicagents ara-C and CH exerted dramatic inhibition of thymidine
and leucine incorporation, respectively.
DISCUSSION
Monoclonal antibodies that can be elicited against specificantigens that are expressed on the surface of tumor cells areconsidered to be excellent tools for cancer detection and arepotentially valuable therapeutic agents for use in passive immu-
notherapy or for targeting drugs and toxins (12,13). In addition,such mAbs provide a useful approach to studies of the physiological functions of specific cell-surface antigens. Thus mAbs
directed against receptors for growth factors were found to altercell growth In vitro. For instance, the binding of mAb directedagainst the receptor for epidermal growth factor inhibited thebinding of epidermal growth factor to the cell surface, yet inducedboth the early and delayed effects of the growth factor, includingthe stimulation of thymidine incorporation into quiescent fibro-
blasts (14). Conversely, the binding of some mAb to the receptorfor transferrin on the surface of certain normal and malignantcells in vitro, in the absence of complement, resulted in inhibitionof colony formation in agar and of cell proliferation in liquidmedium (15).
Even though some mAbs inhibited growth by blocking transferrin binding and causing iron deprivation, a few mAbs inhibited
140120a>
100'•5
g80il
60840#
20INCORPORATION
3H-Leucine8e§§88̧-
D 5D7 ^ AntiLy-1I-IIiiijI:LIrrÃŒLIIIrIiara-CCHUV-2237
SV80 Hs939 SPOT B16InhibitorIP3-FI
Chart 2. Effect of mAb on the incorporation of [3H]leucineand [3H]thymidineinto macromolecules in various tumor cell lines. The incorporation results of[3H]thymidinein cells grown without antibody (control values)were: UV-2237-IP3,52,245 ±2,650 (SE); SV80, 33,074 ±2,085; Hs939, 31,875 ±4,539; SPOT.13,218 ±558; and B16-F1, 18,520 ±3,655 cpm. Thymidine incorporation intocells exposed to ara-Cwere 1 to 5.8% of control. The incorporationsof [3H]leucinein cells grown without antibody (control values)were: UV-2237-IP3,4,246 ±498;SV80, 1,918 ±64; Hs939, 2,108 ±561; SPOT, 8,445 ±451; and B16-F1, 7,226±352 cpm. Values for leucine incorporation into cells exposed to CH were 1.5 to4.6% of control. Columns, mean of triplicate determinations; bars, SE. Similarresults were obtained in 2 additional independentexperiments. More details on theprocedures used are given in "Materials and Methods."
cell growth without altering transferrin binding or iron uptake.This suggests that the binding of these mAbs at the cell surfacecauses the production of a growth-inhibitory effect unrelated to
the transport function of the receptor (15). The binding of mAbto cell surface antigens of unknown function may also result ingrowth inhibition. For example, an anti-Thy-1 mAb blocks the
growth of a murine thymoma cell line, presumably by stericallyblocking access to proximal receptors specific for the autosti-mulatory murine leukemia virus proteins required for continuedcell proliferation (16). A mAb directed against the disialoganglio-side Go3was found to inhibit the in vitro growth of certain humanmelanoma cells by an initial cytostatic effect that leads to celldetachment with its own subsequent cytotoxic effect (17). Theanchorage-independent growth of human melanoma cells wasinhibited by a mAb directed against a melanoma-specific chondroitin sulfate proteoglycan (18). It was proposed that this cell-surface component may be involved in cell-cell interactions related to anchorage-independent growth regulation (18). It is
important to note that the above examples are extraordinary;most mAbs that are reactive with different cell-surface antigens,including HLA-A, B-, C-, Dr-, and melanoma antigens, fail toinhibit cell growth (17-19).
CANCER RESEARCH VOL. 45 SEPTEMBER 1985
4351
on April 3, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

TUMOR GROWTH INHIBITION BY ANTILECTIN mAb
In our study, we assessed the effects of a mAb, directedagainst an endogenous tumor cell galactose-specific lectin, on
the growth of several transformed and tumor cell lines. We foundthat this mAb was capable of inhibiting the colony formation ofsome tumor cells under both anchorage-independent and anchorage-dependent growth conditions.
The growth-inhibitory effects of mAb 5D7 appear to be cyto-
static, not cytotoxic, for the following reasons: (a) the effectswere observed in the absence of complement; (b) no cell rounding or detachment was observed; and (c) the incorporation ofthymidine and leucine after 3 days of exposure to the mAb wasnot decreased.
The cytostatic effects of mAb 5D7 were dose dependent, andthe sensitivities of the different cells varied. A preliminary evaluation of this variability suggests a relationship between the levelof mAb 5D7 binding at the cell surface and the extent of thegrowth-inhibitory effect. The inhibition by mAb 5D7 seems to be
specific inasmuch as mAbs for chondroitin sulfate or fibronectin,which bind extensively to the UV-2237-IP3 fibrosarcoma cells
and to a lesser extent to the human and murine melanoma cells,showed no growth-inhibitory effects. Thus the mere binding of a
mAb to the surface is not sufficient for inhibiting growth of thesecells.
The mechanism by which mAb 5D7 inhibits growth is not clear.It is possible that the cell-surface lectin plays a role in growthregulation by functioning as a receptor for some carbohydrate-containing growth-promoting substances. It may mediate spe
cific interactions with extracellular glycoconjugates deposited byculture medium serum on the plastic dishes, or with the galactoseresidues present in the semisolid matrix (agarose). The abrogation of these putative interactions would alter growth regulationand result in growth inhibition. Our results do not, however,exclude the possibility that the binding of mAb 5D7 to the cell-
surface lectin inhibits growth by interfering with the function ofan adjacent membrane constituent, i.e. by sterically hinderingaccess of growth factors to that membrane component.
The ability of cells to grow in semisolid medium is used as acriterion for cell transformation (e.g., after transfection; 20), because this property is usually exhibited only by transformed andtumorigenic cells (21, 22). Inhibition of anchorage-independent
growth of tumor cells is used as a method for screening sensitivity of cancer cells to various chemotherapeutic agents (23). Theability of mAb 5D7 to decrease colony formation in agarose mayresult from suppression of the expression of the transformedphenotype by the antilectin mAb. This suggests that the mAbhas potential for antitumor activity in vivo. Since the metastaticbehavior of the murine fibrosarcoma UV-2237 was correlatedwith the pattern of anchorage-independent growth, with the more
metastatic cells forming larger colonies in agar than cells of lowmetastatic potential (24), it will be of interest to examine the
ability of mAb 5D7 to suppress the expression of the metastaticpotential of these cells in vivo.
REFERENCES
1. Poste, G., and Nicolson, G. L. Cell Surface Reviews, Vols. 1-8. Amsterdam:Elsevier/North-HollandBiomédicalPress, 1976-1982.
2. Makita. A., Tsuiki, S., Fujii, S., and Warren.L. Membranealterations in cancer.GANN Mongr. Cancer Res., 29: 3-220,1983.
3. Smets, L. A. Cell transformation as a modelfor tumor induction and neoplasticgrowth. Biochim. Biophys. Acta, 605; 93-111,1980.
4. Collard, J. G., van Beek, W. P., Janssen, J. W. G., and Schijven, J. F.Transfection by human oncogenes: concomitant induction of tumorigenicityand tumor-associated membrane alterations. Int. J. Cancer, 35: 207-214,1985.
5. Nicolson, G. L., and Poste, G. Tumor cell diversity and host responses incancer metastasis. I. Properties of metastatic cells. In: Current Problems inCancer, Vol. 7, pp. 4-83. Chicago: Year Book Medical Publishers, Inc., 1982.
6. Raz, A., and Lotan, R. Lectin-likeactivitiesassociated with human and murineneoplastic cells. Cancer Res., 41: 3642-3647, 1981.
7. Lotan, R., and Raz, A. Low colony formation in vivo and in culture as exhibitedby metastatic melanoma cells selected for reduced homotypic aggregation.Cancer Res., 43: 2088-2093,1983.
8. Raz, A., Meromsky, L., Carmi, P., Karkash, R., Lotan, D., and Lotan, R.Monoclonal antibodies to endogenous galactose-specific tumor cell lectins.EMBO (Eur. Mol. Biol. Organ.)J., 3. 2979-2983, 1984.
9. Fidler, I. J. Selectionof successive tumor lines for metastasis. Nat. New Biol.,242:148-149, 1973.
10. Kripke, M. L, Gruys, E., and Fidler, I. J. Metastatic heterogeneityof cells froman ultraviolet light induced murine fibrosarcomaof recent origin. Cancer Res.,38: 2962-2967,1978.
11. Lotan, R., and Lotan, D. Stimulation of melanogenesisin a human melanomacell line by retinoids. Cancer Res., 40: 3345-3350,1980.
12. Borowitz, M. J., and Stein, R. B. Diagnostic applications of monoclonalantibodies to human cancer. Arch. Pathol. Lab. Med., 708: 101-105,1984.
13. Ritz, J., and Schlossman, S. F. Utilization of monoclonal antibodies in thetreatment of leukemiaand lymphoma.Blood, 59: 1-11,1982.
14. Schreiber,A. B., Lax, I., Yarden, Y.,Eshhar, Z., and Schtessinger,J. Monoclonal antibodies against receptor for epidermal growth factor induce early anddelayed effects of epidermalgrowth factor. Proc. Nat). Acad. Sci. USA, 78:7535-7539,1981.
15. Taetle, R., Honeysett, J. M., and Trowbridge, I. Effects of anti-transferrinreceptor antibodies on growth of normal and malignant myeloid cells. Int. J.Cancer,32: 343-349, 1983.
16. McGrath, M. S., Pillemer, E.. and Weisman, I. L. Murine teukaemogenesis:monoclonal antibody to T cell determinants arrest T-lymphoma cell proliferation. Nature (Lond.),285: 259-261, 1980.
17. Dippold, W. G., Knuth, A., and Biischenfelde, K. H. M. Inhibition of humanmelanoma cell growth in vitro by monoclonal anti Gra-gangliosideantibody.Cancer Res., 44: 806-810,1984.
18. Harper, J. R., and Reisfeld, R. A. Inhibition of anchorage-independentgrowthof human melanomacells by a monoclonal antibody to a chondroitin sulfateproteoglycan. J. Nati. Cancer Inst., 71: 259-263, 1983.
19. Voflmers,H. P., and Birchmeier,W. Monoclonalantibodies inhibit the adhesionof mouse B16 melanomacells in vitro and block lung metastasis in vivo. Proc.Nati. Acad. Sci. USA,80: 3729-3733,1983.
20. Weinberg, R. A. Use of transfection to analyze genetic information and malignant transformation. Biochim. Biophys. Acta, 657: 25-35,1981.
21. Stocker, M., O'Neill,C., Bern/man,S., and Waxman,V. Anchorageand growthregulation in normal and virus-transformed cells. Int. J. Cancer, 3: 683-693,1968.
22. Freedman,V. H., and Shin, S. Cellulartumorigenicity in nude mice: correlationwith cell growth in semi-solidmedium. Cell, 3: 355-359, 1974.
23. Hamburger, A. W., and Salmon, S. E. Primary bioassayof human tumor stemcells. Science(Wash. DC), 797: 461-463, 1977.
24. Cifone, M. A., and Fidler, I. J. Correlationof patterns of anchorage-independentgrowth with in vivo behavior of cells from a murine fibrosarcoma. Proc. Nati.Acad. Sci. USA, 77: 1039-1043,1980.
CANCER RESEARCH VOL. 45 SEPTEMBER 1985
4352
on April 3, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

TUMOR GROWTH INHIBITION BY ANTILECTIN mAb
Fig. 1. Phase-contrast photomicrographs of UV-2237-IP3 fibrosarcoma cells grown for 10 days in 0.5% agarose without any antibody (a), or with mAb 6G8 at 100^g/ml (D), or with mAb 5D7 at 20 M9/ml (c), or with mAb 507 at 100 ^g/ml (cf). All four, x 160.
CANCER RESEARCH VOL. 45 SEPTEMBER 1985
4353
on April 3, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1985;45:4349-4353. Cancer Res Reuben Lotan, Dafna Lotan and Avraham Raz Monoclonal Antibody to Endogenous LectinsInhibition of Tumor Cell Colony Formation in Culture by a
Updated version
http://cancerres.aacrjournals.org/content/45/9/4349
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/45/9/4349To request permission to re-use all or part of this article, use this link
on April 3, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from






![Growth Inhibition of Human Tumor Cells in Athymic Mice by ...[CANCER RESEARCH 44, 1002-1007, March 1984] Growth Inhibition of Human Tumor Cells in Athymic Mice by Anti-Epidermal Growth](https://img.pdfslide.net/doc/110x75/5e7bcebb508ec15dc92ee12e/growth-inhibition-of-human-tumor-cells-in-athymic-mice-by-cancer-research-44.jpg)






















